AMBER AND RUSSET - LATE COLOUR CHANGE GENES Copyright 2014, Sarah HartwellThe
ancestors of the
domestic cat were nondescript
black /
brown striped
tabbies. Over the
centuries , mutation produced a
wide array of
colours based on 2
different pigments. Eumelanin gives the blacks,
browns and
blues while phaeomelanin gives the reds, fawns and creams.
A few
other genes give
further variations on those colours
such silvers, colourpoints and solids/selfs. Mutations
continue to
occur and unexpected colours also
turn up due to inbreeding where recessive
genes,
hidden for generations, start showing up.
AMBER
AND LIGHT AMBERDuring the
1990s , some purebred
Norwegian Forest Cats in
Sweden produced
chocolate /lilac and
cinnamon /fawn offspring.
However , those colours
are not
found in the purebred Norwegian Forest Cat gene pool. Had the
gene pool become polluted by
someone ,
perhaps generations ago,
breeding their Norwegian Forest Cat to
another breed? Was it a
spontaneous mutation?
Crossing of those cats with
known chocolate and
cinnamon colour cats of other breeds ruled out chocolate/lilac and
cinnamon/fawn genes.
These cats were a totally new colour, peculiar
to the Norwegian Forest Cat gene pool and dubbed the "X
Colours". They are now called Amber and Light Amber. The Amber
effect is due to the extension gene (also called red
factor ) which
controls the
production of red and black
pigment . The
dominant version of the gene produces normal black pigment in the
coat while
the recessive version produces red pigment. The name
comes from the
effect of black or brown pigment not being
extended throughout the
whole coat, but being restricted to the
skin of the extremities and
to the
eyes (for example in bay
horses ).
This
Norwegian Forest Cat was bred by Yve Hamilton Bruce from a
silver mackerel tabby
female (imported from Denmark) and a
classic red tabby
and white
male . The
result was 1 silver tabbies and 2 silver tabbies
with white. At just over 3 months old, this silver and white tabby
male
developed a large
patch of
bright red
hair on his
back which
continued to
spread . Eventually the whole fur will become amber. The
effect of amber during the colour-change stage depends on the
original colour -
solid black or
blue , bicolour or tabby. The cat
pictured is not a
typical amber as it has the silver gene so the
amber effect is overlaid on silver.
A
non-agouti amber Norwegian forest Cat resembles a silver tabby, but
has a distinctive black
nose and black paw-pads instead of the
pink /reddish nose with black outlining found on conventional silver
tabbies. The
photo below shows a non-agouti amber. According to Rui
Pacheco,
although this cat is
clearly non-agouti because of the black
nose, he has the
phenotype of a silver shaded. Rui's theory is that
the amber gene invalidates the
action on the non-agouti gene;
wide-banding will further lighten the
base colour
giving a very
pale cat with black nose and paws. Amber cats are basically black cats
with a colour modifier that affects the deposition of pigment on the
hair shaft, but not the skin colour. That is why in non-agouti cats
the nose remains black and in agouti cats it remains pink.
PLATINUMThe
platinum effect is found in some lines of Silver Persian/Chinchilla
Longhair. In 1986/7, Cheryl Bennett
reported in The Silver and
Golden Persian Newsletter (ACFA) that one of her shaded silver females
(Kelley
Lane Contessa of WeANDE) had changed from
pure silver to pale
golden. Contessa’s
parents were a shaded golden and a shaded
silver; these were
full siblings. At 10 – 12 months of age,
Contessa began to “tarnish” i.e. show
cream /reddish patches and
by 3
years of age she was entirely pale golden. She produced a number
of Silver Persians
without tarnish, but she failed to produce any
offspring when mated to a golden
sire . One of Contessa’s male
offspring turned from silver to golden as an
adult . An adult Silver
Persian from
England (Lynchard Silver Shadow) was exported to
Australia and also turned golden. Shadow had a few golden hairs on
his paw, but did not turn golden
until he was a
year old when his
coat turned to pale beige. By 3 years old he was entirely pale
golden. Shadow was bred to a genetically golden female, but the
pairing only produced silver offspring. However, at
least one of his
silver offspring
later turned pale golden. Other descendents of
Contessa also
went through the late colour change. During the 1980s,
several other breeders of Shaded Silver and Chinchilla Persians
came forward to
report that their cats had developed reddish, brownish or
golden-coloured fur
along their spines as they
aged . Many of the cats
had no golden in their ancestry. At
first it was dismissed as an
unavoidable genetic
fault where silver was incompletely dominant and
did not hide the recessive golden colour.
A
common ancestor of all the colour-
changing Silver Persians was a
stud cat called Kelly Lane Andromeda (in the UK)
whose descendents were
exported to the USA in the 1950s and
1960s . Some of those descendents
were influential stud cats and would have spread the mutant gene far
and wide. Controlled inbreeding (linebreeding) helped establish the
gene, which would later
double up to produce colour-changing Silver
Persians. The effect of this gene on Golden Persians, if indeed it
has any effect, is not known.
Except
for red-silvers and cream-silvers, silver cats should entirely
lack phaeomelanin (red or cream pigment) and only have eumelanin (black,
brown, blue etc)
present . Perhaps some other gene
causes the
eumelanin structure to change so that it is
perceived as a golden
colour. Chemical
analysis proved that the late colour change Silver
Persians did not have phaeomelanin pigment present.
AMBER-TYPE
EFFECT IN MANXAn
amber-type effect has also been
seen in a Manx cat. Janet McArthur
bred a male Manx (a longy i.e. with
tail ) that was born an ordinary
black/brown mackerel tabby without much rufousing, as was his
sister .
In
addition , the male had white ticking at the
tips of his hair. As
the male grew, his stripes
faded out and
became spots while the black
markings also changed from black to a burnt red-brown colour and his
white ticking became smaller. The change from
stripe to spots was
probably an optical effect caused by the change from black to
red-brown. His dorsal line became a burnt golden brown, and the
background colour was
similar to a washed out sorrel colour (sorrel
is a colour found in Abyssinians).
There are a few oddities in his ancestry. His
mother ,
Pearl , was a black
Manx and his sire was a normal black/brown tabby from the
same black
Manx mother. The inbreeding was
accidental , and may have
brought out
a recessive gene. Pearl was born to a black/red tortie (carrying
dilute and colourpoint) and a red van-pattern sire; this pairing
should not produce solid black females so, barring an anomaly in her
sire's germ line, Pearl may be a tortie where the red has not been
expressed in the coat. Pearl has also produced a
kitten that changed
from black with white ticking at the hair-tips, to black
smoke and
then to solid black without ticking.
BIMETALLIC
/ SUNSHINEThe
informal
term "bimetallic" (referring to a mix of silver
and golden or silver-gilt) describes some Siberian silver tabbies
that turn golden in a
rather patchy fashion. Unlike the amber gene,
the colour change does not start on the back and
work downwards, but
seems distributed throughout the coat. The
formal name "sunshine"
has been proposed for this
emerging gene.
RUSSETSimilar
to amber is russet, which turned up in a line of seal (brown)
European-style Burmese in New
Zealand in 2007. It has subsequently
occurred found in the
related Mandalay (similar to the
Asian in
Europe ) and
appears to be a mutation of the extension gene. The first
known russet was a pure-bred Burmese called “Molly” in 2007.
There is now an experimental
programme in NZ to breed Russet Burmese
and to investigate dilute russet, russet tabby and solid russet (as
opposed to the Burmese sepia form of russet).
The
first russet kitten “Molly” was born an "odd-coloured lilac
(lavender)" which gradually lightened as she grew, progressing
through lilac-caramel and chocolate
ticked tabby and then
dramatically changing to red. Chocolate ticked tabby and red were
both impossible from her pedigree, and in any
case , reds are not born
as chocolate tabbies! Several more unusually coloured kittens were
born in different litters and all went through the same
colour/pattern
changes . The ancestors of these kittens were seal
Burmese and had no silver, tabby or Mandalay (Asian)
blood . The
pedigree had both dilute and chocolate, there were no reds, creams or
torties until 4 generations back. DNA
testing showed some kittens to
be genetically seal and other genetically chocolate.
Hence the new
colour is due to a different mutation currently known as russet.
Russet appears to be due to a recessive mutation that causes black
pigment (eumelanin) to gradually
fade to a
minimal amount while
leaving red pigment (phaeomelanin)unaffected.
Russet
kittens resemble tabbies at
birth , but have pink noses and paw-pads,
pale fur
around the pads and genitalia and a pale tail-tip - all of
which would be
dark in tabbies. The muzzle and fur around the eye is
ivory. The back is solidly dark rather
than ticked, becoming pale
ivory halfway down the flanks. The back becomes more ticked
appearance,
almost a saddle, as kittens undergo the colour change and
the
face becomes reddish. By age two, they may resemble a red
Burmese. It differs from amber as ambers have dark noses and
paw-pads. Non-agouti (solid) amber kittens are very dark with a dark
face that is last to go red while russet kittens have off-white faces
(possible due to Burmese sepia gene in the mix), which are the first
part to go red (rather than the last as in ambers), and pale
undersides. The russet colour change appears to be slower than the
amber colour change. Russet kittens to
date have been larger at birth
than their siblings and somewhat on the large side as adults.
RECESSIVE
BROWN - THE ENIGMATIC BARRINGTON BROWN GENECopyright
2010 Sarah HartwellThis
page pulls together what is known about Don
Shaw 's Barrington Brown
gene, a form of recessive brown/colour dilution only reliably
recorded in a
colony of
laboratory cats,
none of which are believed
to have
left the laboratory. To make this comprehensible to the
non-genetics
expert I have
referred to "copies of genes" or
"versions" of genes although the
correct terminology is
"alleles". There is also a brief guide to Shaw's
terminology at the end as Shaw's writing pre-dated modern "standard"
symbols and terminology.
Don
Shaw was an
early feline geneticist in the USA. During the 1950s and
1960s, there was no standard form of genetic coding and Shaw used his
own system of genetic coding which can be difficult to read
today . He
also referred to chocolate, which is a mutation of the black gene, as
"chocolate dilution". What modern fanciers
call dilution,
Shaw called "maltesing" (Maltese i.e. blue cats were a
genuine dilution of black). Shaw viewed dilution as being due to the
reducing amount of melanin present in the hair and not by the way
pigment
forms clumps in the hair. Chocolate had less melanin than
black,
therefore Shaw called this dilution. Blue has a the same
amount of melanin, but arranged differently, so Shaw did not
consider this dilution.
In
simple terms , the black (eumelanin) colours in cats is due to a
series of genes (alleles to be precise) with the
following order of
dominance/recessive (most dominant at the top, most recessive at the
bottom ). The
chart also shows how these combine with dilution genes
and dilution modifier genes. A cat inherits 2 copies of the black
gene and only the more dominant version will show up while the
recessive gene will be hidden. The caramel form only shows up in cats
that
already have the dilute colour and it lightens and adds a
brownish
cast to the dilute colour.
ORIGINAL COLOURDILUTE VERSIONCARAMEL VERSION OF THE DILUTE VERSION (DILUTE MODIFIER, DOUBLE DILUTION)Black
Blue
Caramel (UK: Blue-based caramel)
Chocolate
Lavender (Lilac)
Taupe (UK: Lilac-based caramel)
Cinnamon
Fawn
UK: Fawn-based Caramel
These
colours will naturally
look a
little different on Burmese (due to the
sepia gene), Tonkinese (
mink gene) and cats with the silver Inhibitor
gene. For clarity, this article will
ignore those additional genes
and just look at the eumelanin (black) series of colours.
An
additional type of recessively carried brown colour has been reported
in laboratory cats, but not in the
outside world. Termed "Barrington
Brown", a cat with two copies of this gene had black pigment
diluted as shown below. Because it hasn't (yet) been seen in the cat
fancy , this gene is enigmatic to many fanciers.
Some
have suggested it is the same as caramel (dilute modifier), however
some reports indicate cats with 2 copies of the Barrington Brown gene
were different in colour from cats with only one
copy of the gene
(which seems odd
since the gene is recessive and cats with only one
copy should have dilution at all). When 2 copies were present,
Barrington Brown had an additive effect on black and chocolate, and
presumably on cinnamon (which may have been unfamiliar to Shaw).
ORIGINAL COLOURBARRINGTON BROWN DILUTIONBlack
Deep mahogany brown
Chocolate
Light brown
Cinnamon
Cafe -au-
lait (
milk coffee colour)
The
dilution only happens if 2 copies of Barrington Brown are present,
although some
sources suggest the genes had additive effect on each
other to create the pale milk coffee coloured cats when there were 2
Barrington Brown genes. Since Barrington Brown is a recessive gene.
If only one copy was present it should be masked by the dominant
non-Barrington copy. This is part of the problem when interpreting
reports that use an old, non-standard genetics notation!
ba - Barrington Brown Locus (unverified)Genotype Description PhenotypeBa/Ba
Homozygous - Non Barrington Brown
Cat unaffected - i.e. Black/Brown/Chocolate etc
Ba/ba
Heterozygous - Non Barrington Brown - carrying Barrington Brown
Cat unaffected - i.e. Black/Brown/Chocolate etc
ba/ba
Homozygous - Barrington Brown -
liberty of renaming
Mahogany Brown/Light Brown/milk coffee in colour (depending on whether cat is black/chocolate/cinnamon)
In
the 1960s, chocolate was
described as a dilution of black and the
third possible version of the gene, cinnamon, was apparently
unfamiliar to Shaw (or not
reflected in his terminology). Shaw’s
Barrington Brown article was first written in the 1960’s in the
Journal of Cat Genetics, and reprinted in the early 1970’s in Cats
Magazine and has been
subject to much reinterpretation, especially
when a dilute modifier emerged in the cat fancy.
According
to Shaw's breeding data, Barrington Brown (mahogany brown) dilution
was inherited in much the same way as “standard chocolate dilution"
(black/chocolate alleles) by reducing the amount of pigment in the
hair and by producing elliptical pigment granules instead of
round granules. Elliptical pigment granules refract light differently and
give a reddish-brown colour instead of black/sepia. In Shaw's
terminology which can confuse modern readers, chocolate is a dilute
of black (while blue is "maltesing" of black).
It
became apparent that Barrington dilution gene was in a different
location to the ordinary black/chocolate genes and was inherited
independently of black/chocolate. That
means it wasn't the modern
cinnamon. Genes in different locations can
affect different enzymes
involved in production of the same protein, in this case the
production of eumelanin pigment. Shaw referred to the “standard
chocolate dilution" as affecting
enzyme D while the Barrington
system
affected enzyme B. He identified Barrington Brown as
having 2
alleles; the dominant
wild type and the recessive Barrington Brown
dilution.
B+
= Wild type gene. Apparently
responsible for normal Enzyme B
production, giving full intensity of melanin.
b = Barrington Brown
recessive gene. When 2 copies are present there is less Enzyme B
produced which means less melanin produced; the pigment granules are
elliptical (reddish-brown) instead of round (black).
He
gave it the name Barrington Dilution or Barrington Brown because it
was
discovered at the Quaker Oats Nutritional Laboratories in
Barrington, Illinois. The first cat known to be homozygous for
Barrington Brown was born at the Barrington Laboratories. This was a
strangely colored light brown (
rich caramel or perhaps cafe-au-lait)
male kitten homozygous for Barrington Brown as well as having
“standard chocolate dilution". The 2 gene systems were
additive in
nature and each expresses its effect as if the other were
not present i.e. the end result is the sum of both Barrington Brown
and "standard chocolate dilution" being expressed. Shaw's
description of "rich caramel" (light tan) misled 1970s
breeders to believe the emerging dilute modifier was the same as
Barrington Brown.
Don
Shaw and Wayne Durdle investigated this gene in depth. Cats carrying
the Barrington gene were donated by the Quaker Oats Nutritional
Laboratory to the Feline Research Laboratory at Tuskegee Institute,
Alabama (maintained there from
summer 1966 to mid-1969). No
Barrington Brown carriers were believed to
exist outside of the
colonies and Barrington and Tuskegee.
Studies established that the
light tan or cafe-au-lait colouration was due to the combined
effects of "standard chocolate dilution" and their Barrington Brown
gene.
Shaw
had left the
project in July 1968 and Durdle reportedly terminated
the colony in
June 1969 (this suggests the cats were destroyed,
neutered or used in other research). To their
knowledge - and to the
ongoing chagrin of many breeders - the Barrington Brown cats had no
direct descendents in the cat fancy and the gene was
lost . However,
Shaw noted that some "doubly diluted" kittens were
appearing in the cat fancy. These had no apparent connection to the
Barrington or Tuskegee cats. He also mentioned unconfirmed reports of
a dilution system similar to the “standard chocolate dilution"
but not traceable to any known chocolate ancestry.
Shaw
offered the following possibilities:
- a leak of the Barrington Brown gene from one or other of the laboratory colonies into the cat fancy population; perhaps some cats or kittens had been taken home as pets and bred.
- a new mutation in a completely different gene location
- a new mutation at the same location as his Barrington Brown gene
- an independent recurrence of the Barrington Brown mutation.
To
identify the
relationship , if any
between the Barrington Brown cats
and those with "double dilution" would
require test-matings
between the new "doubly diluted" cats and some known
Barrington Brown cats. Since there were no known Barrington Brown
cats outside of the laboratory and the lab colony had been
terminated, this wasn't possible. In the 1970s, Pat
Turner stated ,
without any supporting
evidence , that caramel (the double dilution
mentioned by Shaw) was the same as Barrington Brown. The dilute
modifier (caramel) colours are not the same as the colours described
by Shaw, though some websites continue to repeat Turner's
claim that
Shaw called the caramel colour Barrington Brown. Turner had simply
jumped to conclusions. Barrington Brown
cannot be the same as caramel
because Barrington Brown affected black and chocolate, but caramel
only affects blue, lilac and cream.
Shaw
noted that recessive
traits emerge in pedigree cats because of
line-breeding and inbreeding which matches together genetically
similar individuals. This means a better
chance of cats inheriting 2
recessive versions of a gene and new traits showing up as result -
just as they did in the Barrington colony of cats.
From
a breeder viewpoint, it is sad that "recessive brown" has
been lost. Shaw's descriptions indicate that it wasn't the same as
caramel. Its effects on the wider palette of feline colours -
torties, ticked and patterned tabbies, Burmese sepias, minks and
colourpoints - can only be hypothesised. From time to time there are
reports of odd colours in cats,
including a tantalising "palomino"
described as "the colour of a brown
paper grocery bag" from
the USA that might just have been the light tan noted by Shaw.
SHAW'S
TERMINOLOGYModern
feline geneticists use b/b for chocolate, but Shaw defined chocolate
as d/d, because to him it was a dilution of black.
Reading Shaw's
work on black, chocolate and Barrington Brown without knowing his
terminology is therefore confusing. In Shaw's defence, the standard
gene symbols had not been defined in the 1950s and 1960s.
Light
tan/cafe-au-lait in Shaw's terminology is d/d, b/b. To a modern
reader, d/d, b/b indicates lilac. This, as much as any
visual similarity, helps
explain why his Barrington Brown is so often
confused with the dilute modifier.
Shaw’s
Chocolate alleles
D+-
= Normal colour (black/sepia)
d/d = chocolate (in Shaw’ s
words ,
Chocolate Dilution)
Shaw’s
Maltese alleles
M+
=
Dense colours e.g. black
m/m = Maltese (or in our terms
dilution) e.g. blue is the maltese of black
Shaw's
Barrington Brown
allele B+
= Normal coat
b/b = Barrington Brown
Lilac
(in Shaw's terminology) would be d/d, m/m = Dilution allele
(chocolate) + Maltese allele = lilac.
Lilac in modern
terminology is b/b, d/d = Chocolate allele + dilution allele = lilac.
Although
there is no
official modern
symbol for the lost Barrington Brown, Ba
is often used for convenience in modern discussion (since it isn't
allocated to
anything else ).
Brown
Locus (modern symbols)
B/-
= Black/Brown (Shaw's black/sepia)
b/b = chocolate (Shaw's
"standard chocolate dilution")
bI/bl = cinnamon
(probably not recognised by Shaw)
Barrington
Brown Locus (modern symbols)
Ba/Ba
- no Barrington Brown dilution
ba/ba - Barrington Brown
dilution/recessive brown
THE BASIC SELF (SOLID) COLOURS OF CATSCopyright
2002 - 2013
Sarah
HartwellGenetically
speaking, there are
four basic self (or solid) colours of cats:
black, chocolate, cinnamon and red. All other self colours are
modifications of these. Although covered
here as a self colour, red
is a form of tabby and it is impossible to completely eliminate the
tabby markings. Why are there not
five basic colours? White is
counted as an absence of colour rather than a colour.
Different
countries, registries and breeds have different
names for some of the
same basic colours.
Even where the same name is used, there may be
different
views on what is an acceptable or
ideal version of that
colour. Colours which appear identical to the human eye are caused by
different genetic interactions.
The
same colours are called by different names in different breeds. Even
in the same breed, the colour may have different names depending on
which
country the cat comes from and which registry it is registered
with. American registries like to add "mink" after the
Tonkinese colours whereas
British registries use the same name for
that colour as is used in the
equivalent Siamese or Burmese colour.
Confused? Don't worry - there are some
cross -
reference tables later
on!
The
jet-black colour you known as "black" is called "
ebony "
and "ebony tabby" in Orientals, "black" in solid
coloured domestic shorthairs, "brown" when it refers to
brown tabby domestic shorthairs, "bronze" in Egyptian Maus,
"tawny" in Ocicats and "ruddy" in Abyssinians. In
colour-pointed cats, "black" is called "seal". In
Burmese it is "
sable " or "seal sepia" and in
American Tonkinese it is "cinnamon" or "natural mink".
In the Asian breed (self Burmese cats) it has a breed name to itself
"Bombay". Shaded silvers, black smokes and chinchilla cats
may look various shades of
grey or silver, but they are black cats
with silver
roots to their fur. Add dilution and it becomes "blue".
Modify the dilution and it becomes "caramel". Yet it is
still basically a black cat.
WHY
RED SELF CATS ARE STILL RED TABBIESStrange as it may
seem , all red cats are actually red tabby because the
non-agouti gene (the gene that turns a tabby into a self/solid
colour) does not affect the way red pigment is deposited. A variety
of other genes, called polygenes or modifiers,
control the intensity
of colour and
contrast between markings and background colour.
Selective breeding has reduced the tabby markings to produce a cat
that
looks solid red by breeding from those cats with the least red
markings (cats with "low contrast" between markings and
background colour). Because the non-agouti gene does not work on the
red pigment, red tabby
ghost markings can
never be completely
eliminated and may be seen on the tail,
legs and
forehead and as a
darker
region along the
spine . Even though red cats are registered as
"red self" they are still red tabbies, albeit red tabbies
with very reduced markings. Because they lack the polygenes for high
contrast between markings and background colour, their offspring also
appear to be red self. More information can be found in
Robinson 's
"Genetics for Cat Breeders".
DILUTION
AND MODIFIED DILUTION - MALTESING AND CARAMELISINGThe
simplest modification of the 4 basic colours is dilution. As the name
suggests, this "washes out" the original colour to
something paler. This is also called maltesing since the it was first
identified in black cats and blue cats - Maltese is term for
blue-grey and some of the eary blue cats imported into
Britain were
known as Maltese cats.
A
secondary type of dilution is called the dilute modifier. It only
affects already diluted colours. Since it gave
rise to the colour
"caramel". I have referred to it here as "caramelising"
purely to
avoid confusing the layperson. Textbooks always
refer to it
as the dilute modifier.
ORIGINAL COLOURDILUTE VERSIONCARAMELISED VERSION OF THE DILUTE VERSION (DILUTE MODIFIER, DOUBLE DILUTION)White
N/A
N/A
Black
Blue
Caramel (UK: Blue-based caramel)
Chocolate
Lavender (Lilac)
Taupe (UK: Lilac-based caramel)
Cinnamon
Fawn
UK: Fawn-based Caramel
Red
Cream
Apricot Amber
Light Amber
Black,
chocolate and cinnamon are all versions of the same gene. With the
exception of "red" which is a
special case, genes are
inherited in pairs - one from each
parent . Some genes are dominant
over
others and only the dominant one will be expressed (show up).
The other gene (the recessive) will still be lurking in the
background and can be
passed on to offspring. Depending on which
pairing a cat inherits, it will be one of those basic colours. The
fact that at it may look different depends on many other genes which
alter the way in which these three basic colours are expressed.
If
it inherits black + black OR black + chocolate OR black + cinnamon it
will be black.
If it inherits the dilution gene it will be
blue.
If it inherits the dilution gene AND the caramelising gene
it will be caramel.
If
it inherits chocolate + chocolate OR chocolate + cinnamon it will be
chocolate.
If it inherits the dilution gene it will be
lavender.
If it inherits the dilution gene AND the caramelising
gene it will be taupe.
If
it inherits cinnamon + cinnamon it will be cinnamon in colour.
If
it inherits the dilution gene it will be fawn.
If it inherits the
dilution gene AND the caramelising gene it will be a pink-brown..
Many
early caramels were probably registered as fawns or lilacs, probably
as
poor quality individuals. The colour
differences are subtle enough
that it may be necessary to check what is in a cat's pedigree to work
out the exact colour. To the
rest of us, such cats are simply
"biscuit colour" and are no less attractive for it!
Red
is a different gene to black, chocolate and cinnamon. It is a
sex-
linked gene which means a female must inherit 2 copies of the red
gene in order to show up as a red cat. A male only
needs one copy of
the red gene to be a red cat. This is why
ginger tomcats are more
common than ginger females (though contrary to
popular belief , ginger
females are neither
rare nor infertile!). If a female only inherits
one red gene, she is a tortoiseshell.
If
a female inherits red + red she will be red (ginger).
If she
inherits 1 red gene she will be tortoiseshell (red/black,
red/chocolate or red/cinnamon depending on the other genes
present)
If she inherits red + red AND the dilution gene she will
be cream.
If she inherits 1 red gene AND dilution gene she will be
dilute tortoiseshell (blue/cream, lilac/cream or fawn/cream)
If
she inherits red + red AND the dilution AND the caramelising gene she
will be apricot.
If she inherits 1 red gene AND dilution gene AND
the caramelising gene she will be dilute tortoiseshell in the
caramelised range of colours (though this may not show up very well
and may be impossible to identify.)
If
a male inherits 1 red gene he will be red.
If he inherits 1 red
gene AND dilution gene he will be cream.
If he inherits 1 red gene
AND dilution gene AND caramelising gene he will be apricot.
In
reds, there are genes for rufism i.e. for the depth of the red
colour. This is why show-quality reds are a rich, deep red colour
while alley cats are more often marmalade or ginger. Early reds
(1880s) were known as yellows; the depth of colour was
improved over
many generations of selective breeding.
In
1924, a series of breeding experiments between a Siamese female and a
tabby male resulted in black offspring that themselves produced tabby
offspring. This suggests a gene for black that is dominant to tabby,
the opposite of the known behaviour of tabby and black! It is
possible that the Siamese female, one of a
pair imported from
Bangkok , had a mutation for black colour that was dominant instead of
recessive. There have been no further reports of dominant black in
the cat population. Dominant black is genetically different to the
black colour described
above , but
unless it occurs
again its
interaction with other genes will
remain unknown.
Note :
The usual explanation of black (B), brown/chocolate (b) and cinnamon
(bl) is that there are 3 alleles of the black locus gene. Chocolate
carrying cinnamon can appear as light chocolate in some breeds, but
basically there are 3 phenotypes: black, brown/chocolate and
cinnamon. An alternative explanation suggests 2 gene loci, each with
two alleles. One locus is black/brown. The other locus is a modifer
that, when homozygous (2 copies are present), changes black to
cinnamon and brown to pale cinnamon. This gives 4 phenotypes: black,
brown/chocolate, dark cinnamon/light chocolate and light cinnamon.An
additional type of recessively carried brown colour was reported in
laboratory cats in the USA in the 1960s, but not in the outside
world. Termed "Barrington Brown", a cat with two copies of
this gene has black diluted to deep mahogany brown, chocolate dilutes
to light brown and cinnamon dilutes to a pale cafe-au-lait. There are
suggestions it may have been the caramel gene, but the descriptions
of the colours
quite different. The confusion seems to stem from the
discovering describing the light tan of the Barrington Brown cats as
"caramel coloured". Barrington Brown cannot be the same as
caramel because Barrington Brown affected black and chocolate, but
caramel only affects blue, lilac and cream.
CARAMELThe
Dilute Modifier gene was posited by Patricia Turner who erroneously
believed it to be the same as the Barrington Brown gene reported in a
colony of laboratory cats. In essence, caramel lightens and gives a
brownish cast to the underlying colour. How can
adding brown make
something lighter? Brown (in terms of colour, not genetics) comes in
a variety of shades ranging from fawn to chocolate and adding a
brownish hue is not the same as mixing
paint . With the dilute
modifier, blue becomes caramel, lilac becomes taupe (brownish grey)
and red becomes apricot. In
practice , these colours may only be
visually distinguishable from each other by knowing the cat's
genetics. Some
find caramel cats resemble golden series cats and
suggest that a theoretical hypostatic (hidden) silver/golden gene is
showing through, especially in the lighter colours where there is
nothing to
mask a hidden silver/golden colour.
In
the past, the Dilute Modifier has also been called “double
dilution” and “caramelising”. Some people are still not
convinced about caramel because almost identical colours can result
from blue + caramel and from lavender/lilac + caramel. With a
modifier gene, it is possible to get the same end colour (visually)
from 2 different base colours (genetically). It depends on what the
Dm gene is modifying (the expression of a protein) which is different
from mixing paint. Two different original proteins (relating to
melanin production or deposition)
could be modified by such a gene to
produce almost identical expression.
EXTENSION
GENES (BLACK MODIFIER) - AMBER/LIGHT AMBER AND RUSSETThe
Extension gene (formerly the Black Modifier gene) brightens black
areas of the coat. At birth, kittens appear to be black or blue (it
has not been found in combination with other genes recessive to
black), but become brighter as they grow. The Black Modifier has so
far only been
observed in the Norwegian Forest Cat. Recognition of
the colour by registries will
allow the cats to be correctly
registered and distinguish
them from chocolate, lilac, cinnamon and
fawn (not found in Norwegian Forest Cats).
COLOURWITH EXTENSION GENE/BLACK MODIFIERBlack
Amber
Blue
Light Amber
Chocolate
Lilac
Cinnamon
Fawn
During
the 1990s, some purebred Norwegian Forest Cats in
Sweden produced
chocolate/lilac and cinnamon/fawn offspring. Because these colours
are not recognised (they indicate outcrossing) they were called
"x-colours". Crossing a "cinnamon spotted"
x-colour with a fawn
Somali produced blacks and blues, not fawns.
Some x-colour cats were registered as "golden", but the
x-colour can be found in combination with silver (x-colour silver
tabbies), ruling out golden: a cat can be either silver or golden,
but not both. The offspring of an x-colour "cinnamon spotted"
and a chocolate point Birman were black and blue tabbies, ruling out
the recessive chocolate, lilac, cinnamon or fawn and ruling out a
recessive masking factor (i.e. that found in colourpoint cats). To
produce black and blue offspring, the x-colour must be genetically
black (dominant to chocolate). Further test-matings upheld these
conclusions.
Self
(solid) x-colour cats were born as poorly coloured black-silver or
blue-silver tabbies. Their tabby ghost-markings faded as they matured
and the colour became bright apricot to cinnamon colour with dark
brown paw pads and nose
leather with no black rim (a black rim is
characteristic of silvers). Mating a self x-colour cat to a
black-and-white and mating two self x-coloured cats together showed
that the colours were not agouti (not ticked), but were new colours,
now called Amber and Light Amber. Kittens are born dark and undergo a
period of
extreme brightening of the black/blue areas as they mature.
Their original birth colour is often seen only on the back and tail,
allowing amber and light amber to be distinguished from one another.
Amber
is apricot-to-cinnamon colour with brown paw pads, nose leather and
eye rims. Kittens are born dark or black, with ghost markings, and
brighten as they mature. Light Amber is a pale beige colour. Kittens
are born blue and brighten with age, becoming pink-beige to fawn at
maturity. The nose leather, eye rims and paw pads are dark blue grey.
(To
complete the description of amber/light amber: Amber Tabbies are
born apricot with black markings; the markings brighten to
reddish-brown/cinnamon at maturity. The nose is pink and the paw pads
and eye rims are brown. Light Amber Tabbies are born beige with blue
tabby markings; the markings brighten to pink-beige/fawn at maturity.
The nose is pink and the eye rims and paw pads are blue-grey.
Amber/light amber replaces black/blue in torties. Amber also occurs
with silver and in bicolours/tabbies -and-white.)
The
provisionally
named Russet Burmese is an experimental Burmese colour
in New Zealand. In 2007, Nicki and Bob Mackenzie's line of seal
(brown) Burmese (carrying dilute and chocolate) produced
"odd-coloured lilac" kittens which gradually lightened as
they grew, progressing through chocolate ticked tabby to red. The
kittens traced to the same
father and two related females (mother and
daughter ) and all three shared a common ancestress only a few
generations back. DNA testing showed the russet kittens to have the
standard seal or standard chocolate genes,
meaning there is an a
different gene causing the colour change. Russet kittens also tended
to be larger than their littermates, both at birth and as adults.
Russet appears to be a recessive gene from a spontaneous mutation. It
causes the black pigment (eumelanin) to gradually fade to almost
nothing while leaving the red pigment (phaeomelanin) unaffected. Like
the amber colour change, it might be an extension gene mutation.
Russet
kittens have pink noses and paw-pads and a pale tail-tip. The muzzle
and fur aroundthe eye is ivory. The back is solidly dark rather than
ticked, becoming pale ivory halfway down the flanks. The back becomes
more ticked appearance as kittens undergo the colour change and the
face becomes reddish. By age two, they may resemble a red Burmese. It
differes from amber (in Norwegian forest Cats) as ambers have dark
noses and paw-pads. Non-agouti (solid) amber kittens are very dark
with a dark face that is last to go red while russet kittens have
off-white faces (possible due to Burmese sepia gene in the mix),
which are the first part to go red, and pale undersides. The russet
colour change appears to be slower than the amber colour change.
PINK- EYED DILUTIONThe
type of dilution seen in cats is blue dilution (it dilutes black to
blue). A second type of dilution seen in many
mammals is "pink-eyed
dilution". Pink-eyed dilution is characterised by a pink or
ruby glimmer to the eye (depigmentation). The coat colour is often diluted
to bluish-fawn and the pink-eyed dilution factor is generally
inherited as a recessive gene. A possible pink-eyed dilute female cat
was reported in 1961. She was described as pink-eyed with a light tan
coat. She was mated to a chocolate point Siamese and produced three
tabby kittens 10
days premature. Sadly none of the kittens survived.
The colour of the kittens implies that pink-eyed dilution in cats is
inherited as a recessive
trait and is independent of the colourpoint
genes. The pigment granules in the hairs of pink-eyed dilute cats
were very small and yellowish brown,instead of the normal dark-brown
or black. (
Todd NB: A pink-eyed dilution in the cat. JHered 52:202,
1961.27.)
ALBINO,
DOMINANT WHITE, WHITE SPOTTING Albino
is generally
thought of as pure white, but the situation in cats is
more
complex . There are five known alleles for
albinism : blue-eyed
albino, pink-eyed albino, Burmese pattern, Siamese pattern and full
colour (non-albino). Full colour is dominant to all of the other four
alleles. Burmese pattern is incompletely dominant to Siamese pattern;
cats that inherit one of each of those genes will be
intermediate in
pattern and is known as Tonkinese. A quirk of the Siamese form of
albinism is that it is temperature
dependent with
warm areas of the
body being paler than cooler areas. For this
reason , it is often
described as "colour restriction" rather than albinism.
Pink-eyed albino appears to be recessive to all of the other albino
mutations.
The
albino cat reported in Europe and the USA seems to be intermediate
between pink-eyed albino and blue-eyed albino. Although it has the
white coat of a true albino, its eyes have ruby red pupils and pale
blue irises. A true pink-eyed albino was reported in
1931 and again
in 1980s in the USA. Albino kittens have turned up more recently in
the
Bengal breed, unsurprising since albinism is found in the Asian
Leopard Cat (the wild parent of the Bengal).
Dominant
white is the colour associated with deafness in cats. Dominant white
masks all other colours and cats may have blue, orange or odd eyes.
Those with blue eyes have a high chance of deafness. Those with one
blue eye have a high chance of deafness on the blue-eyed side. Those
with orange eyes are far less likely to be
deaf . Some dominant white
kittens are born with smudges of coloured fur on top of the head,
this smudge of colour
usually disappears by adulthood, but kittens
with colour smudges are more likely to have normal hearing.
The
gene for white spotting can also create the
impression of a self
white cat. This gene is semi-dominant and is variable in the way it
is expressed - a cat may have no
visible white spots or may be wholly
white and all stages in between those two extremes. Unlike dominant
white, white spotting is not linked to deafness.
COLOUR
RESTRICTION, TICKING AND TIPPINGAnother
form of modification is colour restriction. This is seen in
colourpointed cats where the colour is restricted to the head, tail
and legs (
plus scrotum in
males ). This is a form of partial albinism.
As well as restricting where the colour will be exhibited, it tends
to "bleach out" the colour to a
greater or lesser
degree depending on the type of colour restriction. It may seem odd to think
of these as solid colour cats, but genetically they are; they have
just had a "special effect" overlaid on them.
There
are three colour-restriction
patterns :
In
the Siamese (Himalayan) pattern, there is maximum contrast between
the
points and the body colour.
In the Burmese, there is the least
contrast but just enough to be
able to see that the cat is not a
single solid colour.
In the "Mink" Tonkinese, the effect
is moderate.
The
self colours may have different names in these cats. Alternatively a
colour which appears identical to one of the solid colours may be
genetically different because of the
mild to moderate bleaching
effect. A seal-point Siamese may appear to be brown, but genetically
it is black!
In
Abyssinian and Somali cats, the agouti gene causes the colour to be
distributed in bands along each hair. This creates a ticked effect.
The most confusing aspect is "red". The bright red colour
in sorrel Abyssinians is
really cinnamon. The dark red of the Usual
Abyssinian is genetically black. Where registries recognise the
sex-linked red and sex-linked cream colours the colour name is
prefixed by the words "sex-linked". In the USA, the
sex-linked colours are apparently not recognised to avoid confusion
with the sorrel and fawn colours. To the
naked eye (or without the
benefit of a pedigree chart), the colours are practically
indistinguishable.
It
has been thought there were 2 different shades of sorrel red: a
bright coppery red and a dark red characterised by an influential
American Abyssinian sire called
Champion Dhmahl's Diablo. Some pale
kittens darken to red, while others become deep solid-
looking red
with minimum ticking. The dark red is more likely to be chocolate.
Chocolate and lilac, along with the silver series, are
considered alien to many
North American breeders, but are familiar in Europe.
Full ExpressionSepia (Burmese)Mink (Tonkinese)Pointed (Siamese)Abyssinian/Somali Australian MistBlack/Brown (in tabbies)/Ebony
Brown/Sable/Seal Sepia
Sable/Natural Mink
Sable/Seal
Usual/Tawny/Ruddy/Brown
Brown
Blue
Blue/Blue Sepia
Blue
Blue
Blue
Blue
Chocolate/ Chestnut/Brown
Chocolate/ Champagne Sepia
Chocolate/ Champagne Mink
Chocolate
Chocolate
Chocolate
Lilac/Lavender/
Frost Lilac/Platinum Sepia
Lilac/Platinum Mink
Lilac/Platinum
Lavender
Lilac
Cinnamon
Cinnamon/Cinnamon Sepia
Cinnamon/
Honey Mink
Cinnamon
Red/Sorrel
Gold Fawn/Light Lilac
Fawn/Fawn Sepia
Fawn
Fawn
Fawn/Beige/Dilute Sorrel (non sex-linked)
Peach
Red
Red/Red Sepia
Red
Red
Sex-linked red
Red
Cream
Cream/Cream Sepia
Cream
Cream
Sex-linked cream
Cream
The
colour can also be restricted to the ends of each hair. This is
caused by a gene for silver. Chinchilla (
shell ) is the lightest form
of tipping or shading - hair tip is coloured and hair shaft is
silver, giving a sparkling appearance. Shaded is the next degree -
the colour extends further along the hair shaft, darkest on the back
(where fur is longer) to create a mantle of shading. Smoke is the
heaviest tipping - the undercoat colour is reduced to a small
band near the hair
root so that the cat appears to be solid colour with a
pale ruff until the coat is parted and you can see the silver roots.
In goldens, the effect is similar except that the hair shaft is gold
rather than white. These cats are still self colours, but the colour
distribution has been modified!
There
is another effect known as silver-tipping; this is the sparkling
silver tips to the otherwise black fur of Chausies. It is an effect
inherited from
Jungle Cats (F chaus) and is seen in purebred
melanistic Jungle Cats. Affected cats are self coloured, but with a
modifier which is new to domestic cat genetics.
A
WIDER PALETTE OF COLOURSThe
previous sections dealt with solid colours and how they are affected
by two
types of dilution, three types of colour restriction, agouti
ticking and silver/golden shading.
There
are genes which control how evenly the hair is coloured i.e. the
bands of colour on each hair. In agouti cats, it is
easy to see
different bands of colour using a magnifying
glass . In cats with
apparently solid-coloured fur, you would need a microscope. Even
though the effect on an
individual hair can't be detected, the
overall effect on an area of fur is make the colour more or less
dense i.e. darker or lighter.
In
general the self colours (with
plain English descriptions) are:
Colour name Plain English DescriptionAlbino
White
Amber
Born black, brightens to bright apricot to cinnamon with age.
Apricot
Pink-brown or hot cream, with a metallic sheen,
Beige
Abyssinian/Somali: Non sex-linked cream, Fawn
Black
Jet-black
Blue
Blue-grey
Brown
Chocolate. Genetically black cats with tabby pattern are known as brown tabbies. (Burmese "Brown" is equivalent to black.)
Caramel
Caramelised blue, cafe-au-lait colour (biscuit colour), cool toned bluish fawn, metallic sheen
Champagne
Burmese/Tonkinese: equivalent to chocolate
Chestnut
Oriental:
Medium -dark brown, equivalent to chocolate
Chocolate
Medium-dark brown
Cinnamon
Milk-chocolate (reddish) colour
Cream
Buff, dilute of sex-linked red
Ebony
Foreign : equivalent to black
Fawn
Abyssinian/Somali : hot cream (non sex-linked cream), equivalent to Light Lilac
Light Amber
Born blue, brightens to pink-beige or fawn with age.
Frost
Lilac/lavender
Gold
Australian Mist: equivalent to cinnamon
Honey
Burmese/Tonkinese : Equivalent to chocolate/chestnut
Indigo Lavender
Pinkish grey (dove grey)
Lilac
Lavender
Light Brown
Equivalent to cinnamon
Light chocolate
Burmilla: Milk chocolate
Light Lilac
Equivalent to fawn
Natural
Tonkinese: equivalent to sable/seal
Peach
Australian Mist: Pink-brown, equivalent to light lilac/fawn. Peach is also seen as a dilute of
Russian Blues and may be caramel.
Platinum
Burmese/Tonkinese: Equivalent to lilac/lavender
Red
Rich ginger red (poor reds are yellowish due to other genes)
Red
Abyssinian/Somali: Equivalent to cinnamon
Ruddy
Abyssinian/Somali: Equivalent to black/brown
Sable
Burmese: Dark brown (genetically black)
Seal
Siamese: Dark brown (genetically black)
Sorrel
Abyssinian/Somali: Equivalent to cinnamon, honey mink of Tonkinese
Taupe
Caramel dilution of lilac/lavender
Tawny
Abyssinian/Somali: Equivalent to black
Usual
Abyssinian/Somali: Equivalent to black
White
Non-albino white, this is the absence of colour.
In
all likelihood, there are many other genes which subtly alter the
colour e.g. by modifying the hair structure
slightly to change the
way it absorbs or reflects light or by affecting the distribution of
the pigment granules in the hair shaft. The appearance of
silver-tipped black cats suggests that hybridisation is
going to
introduce new colours and effects into domestic cats.
Some
colours which have already
appeared are still disputed. For example
"caramel" is a subtle form of dilution. It does not affect
non-dilute colours (black, chocolate, cinnamon), but it changes the
appearance of already dilute colours (blue, lilac, fawn). Caramels
which are genetically blue cats have a brownish cast while those
which are genetically lilac are lighter in
tone . So should they have
separate colour names e.g. blue-caramel (caramel), lilac-caramel
(taupe) and fawn-caramel or should all the slightly varying colours
be lumped under the name "caramel". They jury is still out
on this one, but it demonstrates the importance of knowing what
colours the cat has in its pedigree!
Another
new colour, indigo, is described as a richer, darker version of blue.
This suggests some sort of intensifying effect acting on diluted
colours. However since blues can be variable in hue, there is no
confirmation that indigo is due to a specific gene or that it is
genetically heritable. The only example I have seen depicted a
tortoiseshell cat with blue-black and hot cream patches i.e. to
richly coloured to be blue-cream tortoiseshell, but not a red-black
tortoiseshell. This could
mean a gene with the opposite effect to
caramel i.e. it acts on dilute colours only, but it intensifies them.
If so, it is probably invisible to the naked eye and only noticed in
individuals with the most extreme effects of that gene. The same gene
would have no discernible effect on non-dilute colours since there
are already at their most
intense .
Note:
since other breeders describe indigo as a darker version of blue, the
"blue-black" may have been exaggerated by lighting.If there
is a colour intensifier
turning blue into indigo, it could be
expected to work on the other dilute colours as well e.g. turning
lavender into deep lavender, fawn into rich fawn and cream into rich
cream. The differences would probably be too slight to be visible -
it is already
hard enough to detect the caramels!
According
to Inina Sadovnikova, "I believe every breeder has had cats of
one and the same colour, but of a different
shade . Take a black
(brown) tabby, for instance. They range in shades from a light grey
with a black pattern to a rich brown with a black pattern. Reds are
also very different, from a ginger to an
intensive orange. The same
concerns blue, I have seen blues so dark that one doubts if they are
really blue and not black. From my
experience , I
tend to think that
there is a modifier, though I don't
know of any studies of this
factor. The litters of my female Konkordia always had at least one
kitten coloured intensively, either a very dark black tabby or a very
dark red tabby. She has a light shade of colour, but her grandmother
is darker and also had several dark kittens in her litters. This
looks like a recessive, but one needs a lot more of statistics to
prove if and how it is inherited. A dark shade is desirable for red,
but at present undesirable for blue. So I don't know if "indigo"
has any future."
ODDITIESStrange
as it may seem, in some
animals the mother's
diet can affect coat
colour.
Recent work at Duke
University in Durham, North Carolina, has
shown that in a certain
strain of
mice , the "agouti" coat
colour gene can be turned off through methylation. The extent of
methylation was dependent on the mother's vitamin intake during
pregnancy. (Methylation turns off genes by chemically modifying
them). The effect has not been observed in cats, but in theory it
could turn genetically agouti (ticked) cats into solid colours.
Some
cat shelter workers have noticed apparently self red kittens turning
into black adults. Solid black is known to be
prone to rustiness
through damp or sunlight and evidently, in some
cases , to
conditions in the womb. Self black kittens are often "rusty" at birth,
though few cases are as striking as that of a ginger kitten called
"Marmalade" who was entirely black at 6 months old
(personal correspondence).
A
better documented oddity is that of apparently self red or self black
female cats that are genetically tortoiseshell cats. In rare cases,
one colour predominates to such an extent that the other is
represented by a few isolated hairs.
BEAUTIFUL
BICOLOURS - TUXEDO AND MAGPIE CATS
Copyright
2002 - 2016 , Sarah HartwellBicolour
cats go by various names and
come in many patterns. They range from
almost solid colour cats with a white
throat locket or white
tail-tip, through to almost solid white cats with black smudges on
the nose or between the
ears . You may have heard of them as tuxedo
cats (white mitts, white
belly and white
chin with an optional white
tail-tip) or patched, pied, particoloured, harlequin or magpie cats
(usually white with coloured splashes on the back and top of the
head). The term covering all bicolour cats is "piebald" or
"white spotted" with some variants of the pattern being
called the Seychelles pattern.
DIFFERENT
TYPES OF PIEBALD The
diagram above shows a typical progression from solid colour through
to solid white. The number by each diagram is the "
Grade "
of spotting from Grade 0 (no spotting) through to Grade 10 (white
spotting has obscured all of the base colour). The "solid
colour" can be one of the true solid colours e.g. black or grey,
or a tabby colour or tortoiseshell (the bicolour is then known as a
calico and is actually a tricolour). Non-pedigree cats have a diverse
range of combinations not recognised in the pedigree world - the
"solid" area can be Abyssinian ticked (Aby-and-white),
smoke, shaded or tipped. Black-smoke-and white is an attractive
combination if not actually encouraged!
The
Van pattern seen in pedigree Turkish Van cats and in the Seychellois
(a Van-patterned Oriental) is Grade 8 - Grade 9 Piebald and
represents the most extreme of the 'Seychelles' patterns. In the
Turkish Van, the colour is restricted to auburn (red) or cream
patches on the head at the base of each ear; the tail is the same
colour as patches, often with darker
rings because red and cream are
not true solid colours. Although auburn has long been the
traditional colour of Turkish Van markings, other colours are being developed
within different cat fancies, but in its
native Turkey , the Turkish
Van actually refers to an all-white cat (Van Kedi).
(Almost) Van Pattern (Grade 8/9 Piebald)
Grade 7 Piebald Littermates
Grade 7 Piebald
The
Van pattern is also found in Persian Longhairs in the form of Van
Bi-Colours & Harlequins. These often have additional markings on
legs, and one or two small splashes of colour on the body. The
Seychellois is an Oriental type cat, occurring in both longhair and
shorthair forms, exhibiting varying degrees of the Van pattern:-
Seychellois
Neuvieme is a white cat with coloured tail and head
splashes
Seychellois Huitieme is white with coloured tail
and head splashes plus additional splashes of colour on the
legs
Seychellois Septieme has splashes of colour on the legs and
body in addition to those on the head and the coloured tail.
British
cat fancier Pat Turner defined the different levels of white spotting
as: Grade 2 - "White
Trim "; Grade 3 - "Mitted";
Grade 4 - "
Irish "; Grade 5 - "Saddle"; Grade 6 -
"Pied"; Grade 7 - "
Chinese "; Grade 8 -
"Harlequin"; Grade 9 - "Van". In
exhibition -quality bicolour cats, symmetrical markings were
preferred .
The
placement of the colour patches is variable and related to how the
embryo develops and expresses the genes it has inherited. As a
result, even more extreme colour restrictions can turn up, for
example Wanda below (
photos provided by Peggy Alden-Clapper) is an
eyecatching
variation on Grade 9 Piebald - all white apart from a
black tail, but no splashes of colour on the head or body.
Variation Grade 9 (Extreme Grade 9)
"Moorish-headed" Cat (Variation on Grade 9)
In
1896, Jean Bungartz wrote in his "Illustrated Book of Cats"
(in
German only): The black-headed or Mohrenkopf (Moorish-headed) cat
must be
clean white, with contrasting colour on the head and tail.
Consistent specimens of this variety are extremely rare and it can
probably be regarded as one of the most peculiar colour patterns of
the domestic cat. The colour of the head and tail can be either
black, grey, blue or
yellow with no white hairs except those
regularly showing up on the head. As previously mentioned, cats with
good and correct markings are
highly valued.
THE
TYPICAL RANGE OF BICOLOUR PATTERNSPiebald
cats are extremely common and varied in appearance. The white
spotting can occur with any colour/pattern and varies from minimal
white through to almost all white. The smallest amount of white may
be no more than a white locket and a small white belly patch, each no
more than a few white hairs. Though this pattern doesn't have a
descriptive namem the term "locket cat" has been suggested.
The
image at the top of this page and the description below is a
typical progression through the different grades of piebald. Due to
developmental effects at the embryo stage, there are further
variations on this general
theme !
Tuxedo Pattern
White
facial markings
At
one end of the
scale is the white locket, followed by the familiar
tuxedo pattern where the belly is largely white. As the degree of
white spotting
increases , the white spreads up the neck and onto the
chin (as shown in the three-
quarter Persian above). The cheeks may be
white or have white spots and there may be a white blaze (an
inverted V) from the muzzle up between the eyes or a white "teardrop"
on the nose (as shown in the cat above right). If the neck and chin
are white, the
front paws are also usually white.
Mask-and-mantle pattern
Saddle pattern
Grade 7 Piebald (with white
ringed tail)
As
the degree of white spotting increases further, the white extends up
the
sides of the cat, up the forelegs (stockings) and onto the hind
paws. The next step is all white hind legs, white fully around the
neck and the white blaze may
extend right between the ears. Some cats
also have white ear tips. This pattern is sometimes known as
"mantled" because the cat appears to be wearing a coloured
cape with a hood or mask. The mask-and-mantle is a common bicolour
pattern as shown above.
As
the amount of white increases further, it simply shrinks the mantle
into a saddle. The mask may also shrink back to a "cap"
giving the cap-and-saddle pattern (as with the cat in the above
centre photo). Further white spotting breaks the saddle up into
smaller patches as shown in the black-and-white Grade 7 piebald cat.
Sometimes one or more white rings encircle the tail - for some reason
this ringed effect seems more common in tabbies.
With
any degree of spotting, there may be coloured smudges on the cheeks,
chin or nose. Some cats also have black
toes on otherwise white paws.
Jean
Bungartz described two bicolour patterns in his 1896 book "Die
Hauskatze, ihre Rassen und Varietäten" (Housecats, Their
Races and
Varieties ) in " Illustriertes Katzenbuch" (An
Illustrated Book of Cats). His description of the "masked cat"
is the tuxedo pattern, which he describes as "sometimes the
tail-tip is white also. The eyes of this variety are bright yellow
with black rims.
Regular and sharply defined white patches create the
most beautiful Mask Cats which have many admirers."
Bungartz
describes a less familiar bicolour he calls the black-headed or
Moor -headed cat (Mohrenkopf) which was clean white, except for the
head and tail, which were black, grey, blue or yellow with no white
interspersed "except those showing up regularly on the head".
Consistent specimens of this variety were extremely rare and
valuable . His illustration depicts a cat with wholly black head, but
it is possible he was describing the Van pattern.
It
is possible to combine piebald with bicolour. Bicolour Siamese have
been bred in Europe to a mixed reception. A few Himalayan breeders
breed bicolour Himalayans, using bicolour Persians to introduce the
white spotting pattern. Paul Beall, Richmond,
Texas is one such
breeder.
Paula 's photo (below) shows a bicolour (piebald) Himalayan,
but
unfortunately this cat has not produced piebald-point kittens and
Paula has decided not to reintroduce the gene from Persians.
GENETICS
OF PIEBALD CATSWhen
the amount of white is small (under 40%), it is called low-grade
spotting and comprises the white belly patch, mitts, locket and
blaze. In medium grade spotting, the amount of white varies from 40%
to 60% of the coat and is the typical pattern of a show quality
bicolour (a mask and mantle effect). When it is extensive (more than
60%), it is called high-grade spotting - the Seychellois or Van
pattern.
Piebald
spotting is a semi-dominant gene with very variable expression. Low
grade spotted cats and some medium grade, spotted cats are
heterozygotes i.e. they have only one copy of the white spotting
gene. Other medium grade spotted cats and all high grade spotted cats
are homozygotes i.e. they have 2 copies of the gene. Where cats are
in the medium spotted range it is generally impossible to know
whether they are heterozygotes or homozygotes unless they are bred to
a cat known to be either homozygous or heterozygous. To further
complicate matters for breeders, some apparently non-piebald cats are
really piebald cats whose white bits are so minimal they might be no
more than a few hairs in the groin or at the tip of the tail!
In
January 2016 , it was announced that piebald patches form when cells
fail to
develop in the womb. The gene for white spotting affects the
embryo cells (melanoblasts) which will become pigment-producing skin
cells (melanocytes) which make the pigment for hair. Pigment cells
move and multiply as an embryo
grows and there aren't enough cells to
cover all the skin, so the
animal gets a white belly. The findings
were published in the journal Nature Communications. The pigment
cells 'fail to follow instructions' during early
development . They
move and multiply randomly as an embryo grows, without complex
cell-to-cell
communication sending them in one direction as
once thought. The University of
Edinburgh 's Dr
Richard Mort said that the
cells move and multiply at
random which is not what was expected. The
University of Bath's Dr Christian Yates said: that piebald patterns
could be caused by a faulty version of a gene called kit. What the
researchers found was
counter -intuitive. Previously it was widely
thought that the defective kit gene slowed cells down, but instead
they found that it reduced the
rate at which the pigment cells
multiply. This did not
surprise cat fanciers who had noticed that
pigmented patches could fit together like a jigsaw and likened the
process to “plate tectonics” where pigmented patches
drift across the embryo’s skin leaving unpigmented areas inbetween the patches.
Researchers at the Universities of Bath and Edinburgh said that in
addition to kit, there are many other genes that can create piebald
patterns, but their the mathematical model can explain piebald
patterns regardless of the genes involved.
This
wasn't entirely surprising to many cat fanciers with an
interest in
genetics or embryology! Independently of
scientific researchers, and
based on observation, fanciers had some ideas of their own, which
have turned out to be
close to the mark.
Many
cats have patches which look as though they could be fitted together
like jigsaw pieces; for example a coloured
spot on a leg might
correspond to an inlet on a coloured patch on the flank, exactly as
though a
piece of coloured area has
broken off and migrated
elsewhere. Spots could end up almost anywhere depending on the timing
of cracking, the
size of the coloured domains, and the trajectories
they take over the embryonic surface as it grows. Terada and Watanabe
found it possible to fit all the coloured areas of cat coat together.
The seams can then be projected onto the surface of a sphere (the
embryo). As the sphere expands, the coloured area cracks apart. Think
of a
balloon covered in solidified chocolate: the balloon can expand,
but the chocolate can't. The cracked solid surfaces then gave
patterns just like those found on bicolour cats. The swirled patterns
in
particular fit this hypothesis.
Part
of this theory was that the patterning of black and white was the
result of the skin surface "cracking" during early embryo
development. The skin of such bicolour cats at an early embryo stage
would be basically pigmented, but the dominant S (white spotting)
gene, causes the pigmented surface to
crack into domains or
islands .
These islands drift apart over the embryonic surface as the embryo
grows, but the pigment cells don't multiply fast enough to fill the
spaces between the
island . That leaves white areas in the
regions between the islands of pigment. It may help to think of it as similar
to the
movement of the earth's continents with areas of sea
inbetween. It is just as though the white areas were scar tissue
produced by the cracking. There are not enough melanocytes
available to fill the white areas as their surface expands. This means that the
black domains can end up pushed together to form a single tuxedo
style black area (albeit there may be some small white markings in
this single black area). The white belly area might be a ventral
(belly) seam from a ventral crack earlier on - it is suspected that
the belly area expands
greatly during embryo
growth . Megacolon in
cats and exposed gut conditions in some rabbits upholds this theory.
Black
feet could indicate a black domain that has been pushed to the
foot extremity by the expansion of the ventral region at the same
time that the
limbs are being
formed .
Where
the white spotting occurs over the eyes, it may affect the eye
colour. Thus a few bicolour cats have blue eyes. Another
interesting effect of white spotting is in tortoiseshell cats. Tortie cats with
little or no white tend to have brindled coats with intermingled
black and orange hairs. However, the more white there is, the more
the black and white will also be separated out into patches instead
of being intermingled. This is also explained by the mechanism
detailed in the 2016 report findings. The black pigmented cells that
have moved across the embryo surface multiply to form black islands
(clonal patches), while the red pigmented cells that moved across the
surface multiply to form red patches.
OLDER
/ ALTERNATIVE THEORIES FOR BICOLOUR PATTERNSAbove:
"Olga", an attractive brown bicolour
owned by Sarah Richie
It
used to be thought that when the melanoblasts
arose from the "neural
crest" - the area along the back of the embryo -they migrate all
over the body during
formation of the skin. Where cells failed to
reach their allotted positions
before the skin was fully formed,
those areas lacked pigment. This
seemed to explain why the white is
most often found on the paws, belly and
chest - those areas are the
most remote from the neural crest and take
longest to reach. The
slower the
migration of cells, the less colour there will be. It also
explained why the back and the tail may be coloured in an otherwise
all white cat - those areas are closest to the neural crest and the
pigment cells didn't have to migrate very far.
Another
theory involved either of two cellular mechanisms in white spotting
that "turned off" pigment. One mechanism was apoptosis
(programmed cell
death ) reducing the melanoblast (pigment granule)
population. In other words, the colour cells migrate over the whole
surface of the embryo, but then selectively die out. The other
cellular mechanism involved intracellular chemical communication
whereby colour cells are biochemically turned off in certain areas.
Both of these suggest a chemical
gradient so that cells at the
extremities tend to be first affected and stop producing colour.
Another
hypothesis for the distribution of white was originally presented in
papers published by T Terada and T Watanabe in a
1930s Japanese journal. This hypothesis is now being investigated using computer
simulation. There are several "unknowns" with the
current hypotheses: the relative frequencies of the 10 grades of white
spotting
given that white spotted cats may be either SS or Ss; the
relationship, if any, between birth order and patterning; and whether
the "swirled" pattern is related to any birth defects or
any difficulties during pregnancy. The melanocyte migration
hypothesis may not
contain the whole story, especially where swirled
patterns, black feet on white socks and
skunk stripes are concerned.
Some
cat breeders believed there were flaws in the conventional melanocyte
migration theory. For example a breeder
working with American
Shorthairs has been crossing Van pattern American Shorthairs to
bicolour and spotted Van pattern cats since the mid 1990s. She found
that the melanocyte migration theory could not
account for some of
the spotted patterns these breedings produced. Alternative theories
from other white spotting genes and mechanisms in other
species (
dogs ,
horse , pigs and goats) also could not account for what was
appearing in her cats,
particularly with 60%, 70%, 80% and 90% white
on a cat. The terminology used differs from the Grades 1 - 9 of the
cat fancy; 90% white with a
lot of
random spots is referred to as Level 15. Selective breeding has
isolated most of the white on the cats to a single
layer of white.
She can now produce solid coloured cats from the mating of two
bi-colours/pied/vanish cats. In addition, all of her
best breeding
cats have black feet. Breeding experiments involved mating a level 15
stud to a solid black, two Level 7 piebalds, and a Level 9 cat. A cat
with a large degree of white is most probably homozygous for the
white spotting gene and mating it to other white-marked cats which
also
carry at least one white spotting gene should not, in theory,
produce solid-coloured cats. An alternative hypothesis is that the
cats have the dominant white gene (the one related to deafness) and
that a second gene is causing this to break down so that spotting
appears. It is possible to produce coloured cats from mating two
all-white cats since each parent needs only one copy of the dominant
white gene to make them all white and can carry masked genes for
other colours.
OTHER
PIEBALD CATSThere
are quite possibly several genes which
influence white spotting. For
example, the patterns above might not be due to a single gene - there
may be several other genes which modify its effects. There are also
other piebald patterns which appear to be due to different genes or
to cats being homozygous for a modifier gene since the effect is
relatively
uniform and predictable.
The
gloves (mitts) on Birman and Snowshoe cats appear to be due to an
incompletely dominant gene. The gloves may
vary in
length but they
are restricted to feet and/or legs. The white markings on the
forehead and chest of breeds such as the Snowshoe may also be due to
a gene which
limits the amount of white. Lockets - those small spots
on the throat, chest, stomach and groin of otherwise solid coloured
cats also seem to be due to a different gene - one which is normally
hidden by the more extensive white spotting of piebald cats. There is
also the
phenomenon of white toes occurring on otherwise coloured
cats.
In
the
York Chocolate breed, there is a particular white displacement in
bicoloured individuals. The placement of white maintains the same
configuration in all successive generations. This has been termed
"Spotting Particolour" and may be due to an allele
(variant) of the White Spotting gene. According to this theory, the
White Spotting gene appears to have 4 variants: non-spotted, spotted,
particolor, and Birman mitted. Spotted is the dominant form and is
variable expressed. Non-spotted is the recessive wild-type and
produces a coat without white. The hypothetical Birman allele (or
Birman mitted) is also variable, but confines the white spotting to
the legs and feet. The hypothetical Particolour allele produces an
inverted white "V" with the apex in the centre of the
forehead and passing through the centres of the eyes plus a white
chin, chest, belly, legs and feet are white. Particolour is also
variable and its least expression may be a white locket or white spot
on the forehead. The existence of breeds such as the York Chocolate
(consistent particolour pattern) and Birman and Snowshoe (consistent
mitted pattern) appear to support the existence of a white
particolour gene and a white mitted gene.
A
pattern which has only rarely been reported in cats is the
belt ,
blanket or sheet marking. This is common in pigs and
cattle and in
Dutch rabbits where a band of white encircles the animal's body like
a belt.
Another
cause of white spotting is a somatic mutation i.e. some skin cells
have a chance mutation which prevents them from producing pigment.
This is not hereditary.
Leukoderma
("white skin"), leukotrichia ("white hair") or
vitiligo, is a
cosmetic condition that produces a "cobweb"
or "snowflake" effect and is most easily seen on black
cats. White spots appear on the coat; these become more extensive
with age until the cat has a white lace pattern on the black fur.
Ultimately the cat may go completely white or be left with
diminishing isolated patches of colour. This condition has been seen
in black leopards ("cobweb panthers"),
humans , dogs and
other animals. This must not be confused with the normal sprinkling
of isolated white hairs which appear in a cat's fur during its
lifetime . "Leukotrichia" is a generic term. Acommon term is
"piebaldism" because it causes white patches in the skin
and fur. It is an "aquired depigmentation" that occurs
during the cat's lifetime, is usually progressive and may be
triggered by
illness or environmental factors. Ultimately, a cat with
leukoderma may become almost entirely white. Antibodies are formed
against the pigment-producing melanocytes. The melanocytes are
destroyed
leading to the white areas. A type of leukoderma has been
identified in some Persian cats and these are used as laboratory
subjects in the
study of depigmentation conditions. Periocular
leukotrichia, causes the fur around the cat's eyes to become pale -
as though the cat is wearing spectacles.
Vitiligo in a big cat - the "cobweb panthers" that turned progressively white.Charva with feline vitiligo (Jason Reeves). "Charva"
(Jason Reeves) has vitiligo; she was a black cat with patches of
slightly longer/softer black hair. In 2009 she turned 4 and developed
a patch of white hair. Since then the patches of white have turned
into stripes, spots, and patches. Her skin is turning pink as well
due to the loss of pigmentation.
In
2011, Kimberly Sexton of New York City
sent these photos of her cat
Frankie . Kimberly's 14 year old cat
Miko (white with grey markings on
his head and tail) had recently died and in his last 3 years had
formed a close bond with her younger cat, Frankie, a mackerel tabby.
The two cats often slept intertwined. Two years before Miko passed
away , Frankie developed a white spot of fur under his neck, while his
chin and nose became paler. As time passed, Frankie's white areas
spread and new
ones developed around his neck and
behind his
shoulder blades. Kimberly joked to friends that Miko's white colour was
rubbing off on Frankie as they slept! Frankie's white areas are
likely to continue spreading, but the effect appears entirely
cosmetic.
On
coloured cats, hair regrowth around scars is often white. Lizzie
Ellis (The Feline Rescue
Association Inc, Maryland, USA) provided
these photos of a grey male cat with white patches due to scarring.
Where the skin was injured or burned, the fur has grown back white,
not grey. The cat had suffered frostbite injuries. Some dark coloured
cats show white hairs in their coat as they grow older - some
follicles stop producing pigment, just as humans get "grey"
hairs. Unlike the cobweb effect (where pigment loss occurs in
patches, like snowflakes or strands of gossamer), age-related white
hairs are evenly sprinkled across the body giving a salt-and-pepper
effect; there may also be greying around the muzzle. It is a normal
part of ageing although cats seem far less prone to
getting grey
hairs than do dogs. It should not be confused with the white patches
found in vitiligo.
SWIRLED
PATTERNS AND SKUNK STRIPESWhile
the piebald pattern is generally fairly symmetrical until the white
exceeds 60% of the body surface, some cats
exhibit a variety of
swirled patterns. This may simply be due to the way the embryo
developed or it may be the interaction between different genes which
affect white spotting. The two cats below are unrelated, but both
have swirled black and markings and black markings on the face. Both
also have black toes. Coloured toes sometimes occur on bicolours -
these can be individual toes or multiple toes. It may even look as
though a white-footed cat has paddled through coloured paint, in
which case the colour usually extends up the back of the leg.
Coloured toes may possibly be another gene (modifier) interacting
with the white spotting gene.
Note:
The cat shown top left has a mild form of radial hypoplasia which has
caused her deformed forelegs; she also has a slightly deformed skull .Swirled Patterns
More
unusual is the "skunk marking" - a white dorsal stripe.
Because pigment producing cells migrate away from the neural crest,
the dorsal area itself usually remains coloured in low to medium
grade spotting. The appearance of a white dorsal stripe is unusual
and appears to be hereditary.
Photos courtesy of
Magnificent MunchkinsBRINDLED
BICOLOURS AND "TWEED" MUTATIONPandora (owned by
Bill B, Granby, MA, USA) is a brindled cat with what
appears to be an unusual mutation. She has the brindled pattern
normally seen on tortoiseshell cats, but the patches which should be
red are white! The brindling and facial pattern is typical of
tortoiseshell cats, but it seems that 15 year old Pandora has a
mutation that prevents her producing red pigment. An alternative
explanation is that Pandora is chimera formed when a black embryo and
a white embryo fused in the womb; however chimeras tend to have a
patched appearance rather than being thoroughly brindled, this
makes chimerism a less likely explanation. A third possibility is that
Pandora has additional X chromosomes (XXX or XXXX instead of the
normal XX female), however this genetic anomaly is associated with
mental retardation and
physical anomalies/deformities (this is not
Klinefelter
syndrome , Klinefelter syndrome is
only found
in males e.g. XXY or XXXY). These two conditions can be identified
using tissue samples, but this isn't recommended in an older cat as
it involves sedation or anaesthesia. It seems likely that the gene
that should produce red pigment is faulty or that some other gene is
masking the red pigment out. Since Pandora was spayed before Bill
obtained her, it isn't possible to breed her to see if it can be
inherited. I have seen the opposite mutation - where a tortoiseshell
cat produced red pigment, but not black, resulting in a red, cream
and white brindled cat - but this is the first time I have seen a
black-and-white brindle where the red is absent.
More
brindled bicolours are explained in the separate page on
Roan,
Tweed and "Salt and Pepper" ColoursTHE
CHINESE HARLEQUINDuring
the 1980s, there were
attempts to breed shorthaired cats with a black
tail, black patches on the head and small black (or other solid
colour) patches or spots on the body. Named Chinese Harlequin, it was
to resemble cats found in
ancient Chinese art. Although still listed
by some registries, it appears to now be
extinct due to the
difficulty of breeding bicolours with consistent spotted markings. A
similar "strikingly spotted" cat, the Gao Taem, is seen in
ancient
Thai art and is described as having, black marked forepaws,
black and white ears, black shoulders, two black spots on the back
and black shoulders. In
total there should be nine horse-like black
spots on an all white background. If it existed as a breed, the
mutation creating these
features has been lost. In all
probability it
was a form of Seychelles pattern. Some bicolours have black toes or
paws and and some breeders have attempted to fix this trait to create
a black-footed Van-type bicolour.
Also
depicted in Thai art (1676) is the Vichiens Mas which is shown as
white with dark ears, nose, paws and whiskers. Although depicted as
black and white, it seems to be a stylised or idealised depiction of
the seal-point Siamese.
"BELTED"
OR "SHEETED" PATTERN"Belted"
means a solid coloured animal with a belt of white around its
middle ,
such as is seen in Dutch Rabbits. This pattern has
occasionally been
seen in
Spanish feral cats. "Sheeted" means a wider the
band of white i.e. from shoulders to haunches.
THE
"DALMATIAN" OR "APPALOOSA" CATThe
concept of a white cat with coloured spots, akin to a Dalmatian dog
or Appaloosa horse, has intrigued numerous people. In some feral
colonies, white cats with numerous small black splashes have been
found alongside bicolour cats with the conventional Grades 1 - 9 of
white spotting. This suggests the presence of modifier genes which
affect the distribution of colour and white. Inbreeding causes genes
to double up so that recessive genes and traits controlled by
multiple genes become visible. It would be possible to have genes
which interact with the semi-dominant white spotting gene such that
small patches of colour break through the white or which cause a
breakdown in white spotting.
Some
breeders report a
consistently high
incidence of numerous small splashes of colour on the body in certain
breeding lines which suggests to them that some other gene(s) is
being inherited alongside the white spotting gene. This may see a
repetition of the Chinese Harlequin programme.
"BIMETALLIC"
SIBERIANS & PLATINUM PERSIANS
2013-2014,
Sarah Hartwell
Many
thanks to Silvia Perego and Lesley Morgan for allowing the use of
their photographs on this page.Note
on Terminology: At present, terminology isn't standardised.
"Bi-metallic" describes a visual effect, while "sunshine"
is the (current) proposed genetic name. In Siberian Cats,
sunshine-silver produces the bi-metallic pattern.For
some years, strangely patterned Siberian cats have been turning up.
Some are covered in A Torbie Dynasty:
A
Torbie Dynasty.
Caroline Sharp in
Germany had several generations of apparently
tortoiseshell-tabby male cats. This should be impossible as tortie
tomcats are genetically abnormal in some way and even when
fertile they should not produce further generations of tortie tomcats. They
couldn’t all be chimeras, so there had to be some unidentified
mechanism at work.
Other
Siberian cats had both silver and golden areas of fur and were dubbed
“bimetallic.” This X-colour (mystery colour) looked similar to
the partway stages of the amber colour change seen in Norwegian
Forest Cats. Siberian breeders had referred to their cats as "golden"
but this term clashed with the wide band colour found in Persians,
Exotics and British Shorthairs. The official term "sunshine"
has been proposed to distinguish Siberian golden from "wide band
golden".
At
birth, Siberian "Sunshine" resembles the early stages of
the amber (ee) colour change in Norwegian Forest Cats caused by a
non-extension gene. However, the Siberian cats tested negative for
the amber gene. There were no genetic tests for Wide-band or for
silver inhibitor, let alone for Siberian sunshine, but it can be
distinguished visually and by studying pedigrees. Bimetallic
colouration has also been seen in the Kurilian Bobtail and in
random-bred cats in the
Ukraine .
Judge Lesley Morgan come across a number of strangely coloured cats that
had been nicknamed “bimetallic” as they displayed a mix of silver
tabby and golden tabby in their coats. The effect was
beyond the
rufousing or breakthrough colouring sometimes seen in silver tabbies
and in some cats the golden areas increased as the kittens grew into
adulthood. The only
link she found was that every bimetallic Siberian
she saw had close Polish ancestry. Other Siberians had been
registered as black silver tortie tabby, but were more likely to be
bimetallics (silver tabby + sunshine).
3
views of "bimetallic" including a close-up of the fur.
In
2013, Silvia Perego of Veselka Siberian Cattery in
Italy described
more such cats. Her stud male, Yankee, came from Poland and is a
black silver tabby with white. When he
arrived at her cattery at 4
months old, she noticed that he had a "strange"
golden-brown spot on his neck and some little other spots on the
back. At first this appeared to be rufism. His first
litter included a silver tabby daughter,
Lady , who had the same golden-brown spot on
her neck. Another litter included a daughter, Lizzy, with the same
spot. Then Lady, from the first litter, had her own litter with
another silver-and-golden kitten like them. In summer 2013, Yankee
had two kittens with the golden-brown patch. Although a judge
suggested chimerism, there appeared to be a newly emerging gene at
work.
The
classic golden colour found in Persian cats is due to the dominant
wide-band (Wb) gene. This widens the pale brown area at the base of
each agouti (banded) hair and confines the darker colour to the ends
of the hairs. The paw-pads are either white (i.e. pink) or match the
colour at the hair tip. Combined with the silver inhibitors, this
resulted in chinchilla and shaded silver Persians. A golden Persian
has wide-band and the recessive form of the inhibitor gene.
Research
by Eleonora Ruggiero into the pedigrees of Siberian Cats brought out
some intriguing
details . For example in “Onix Gloria” lines there
are golden tabby Siberians born of non-golden parents. In the
pedigrees of “Siberian golden” (provisionally denoted sg), golden
offspring are born from two non-golden parents. This means the gene
was recessive and only expressed when two copies are inherited. It is
not the same as Persian golden (Wb with recessive inhibitor).
Bimetallic females were often registered as tortoiseshell, but didn't
breed as a tortoiseshell as they lacked the red (O) gene (evident
when the paw-pads are examined and are never reddish). Their nose
leather was dark pink, unlike the nose leather of tabby or genetic
tortie cats.
When
golden cats
started turning up in silver Siberians, it resulted in
cats expressing both colours in the coat; something impossible were
it the same as Persian golden. The degree and intensity of golden
colour (probably influenced by polygenes) rules out rufism or
tarnish. These cats were initially referred to as “bi-
metal ” and
later as "sunshine". Siberian sunshine was established in a
breeding programme that used much inbreeding to bring out this
recessive trait. Early sunshine cats were dismissed as silvers with
high degrees of rufism and not bred from. In 2013, all known sunshine
Siberians are agouti (tabby markings) and the colour hadn’t been
seen in non-agouti (solid) cats. The sunshine gene affects the agouti
(ticked) hairs only".
Sunshine
silver looks very different from a silver with rufism; the nose
leather is pinkish unlike that of a tabby cat. The red colour of a
sunshine red tabbies and sunshine torties is brighter and the paws
are lighter. The absence of pigment in the sunshine tabbies extends
beyond the edges of the nose leather so they have no nose-liner and
have whitish fur at the bottom of the nose.
There
is a link to one of the Snowknight cats, Joschka, around 7
generations back, so his torbie dynasty continues. The common
ancestor from Poland is Snowknight Destiny. Either Destiny or her
brother Snowknight Derringer had a very colourful torbie male
offspring, but the breeder had him neutered rather than test-mating
him. This was carried over from Joschka, who had an
extra Chromosome.
Caroline Sharp’s torbie male Bogomir Tschudovich of Misinza aka
Meli is black-and-white, with a big flame of red over and around his
ears and no sex-linked red parentage. Because those cats don't have
the inhibitor (silver) gene, they looked even more like
tortie-tabbies.
Michelangelo,
a colourful torbie male (without silver) from Caroline Sharp's
breeding line.
While
silver-and-gold has turned up in Siberians, a silver-TO-gold effect
has turned up in some lines of Silver Persian/Chinchilla Longhair
where it has been dubbed "platinum". In 1986/7, Cheryl
Bennett reported in The Silver and Golden Persian Newsletter (ACFA)
that one of her shaded silver females (Kelley Lane Contessa of
WeANDE) had changed from pure silver to pale golden. Contessa’s
parents were a shaded golden and a shaded silver; these were full
siblings. At 10 – 12 months of age, Contessa began to “tarnish”
i.e. show cream/reddish patches and by 3 years of age she was
entirely pale golden. She produced a number of Silver Persians
without tarnish, but she failed to produce any offspring when mated
to a golden sire. One of Contessa’s male offspring turned from
silver to golden as an adult. An adult Silver Persian from England
(Lynchard Silver Shadow) was exported to Australia and also turned
golden. Shadow had a few golden hairs on his paw, but did not turn
golden until he was a year old when his coat turned to pale beige. By
3 years old he was entirely pale golden. Shadow was bred to a
genetically golden female, but the pairing only produced silver
offspring. However, at least one of his silver offspring later turned
pale golden. Other descendents of Contessa also went through the late
colour change. During the 1980s, several other breeders of Shaded
Silver and Chinchilla Persians came forward to report that their cats
had developed reddish, brownish or golden-coloured fur along their
spines as they aged. Many of the cats had no golden in their
ancestry. At first it was dismissed as an unavoidable genetic fault
where silver was incompletely dominant and did not hide the recessive
golden colour.
A
common ancestor of all the colour-changing Silver Persians was a stud
cat called Kelly Lane Andromeda (in the UK) whose descendents were
exported to the USA in the 1950s and 1960s. Some of those descendents
were influential stud cats and would have spread the mutant gene far
and wide. Controlled inbreeding (linebreeding) helped establish the
gene, which would later double up to produce colour-changing Silver
Persians. The effect of this gene on Golden Persians, if indeed it
has any effect, is not known. Except for red-silvers and
cream-silvers, silver cats should entirely lack phaeomelanin (red or
cream pigment) and only have eumelanin (black, brown, blue etc)
present. Perhaps some other gene causes the eumelanin structure to
change so that it is perceived as a golden colour. Chemical analysis
proved that the late colour change Silver Persians did not have
phaeomelanin pigment present.
BIMETALLIC
ORIENTAL SHORTHAIRThis
is an Oriental Shorthair, genetically a chestnut silver spotted tabby
and white, but with with non-silver patches resulting in a "merle"
type effect. This looks similar to the bimetallic effect seen in the
Siberians. Kotchera
Trail Blazer of San-Toi was bred by Wendy Heidt,
a Siberian breeder and is being investigated by Pat Decano
(CasaDecano Orientals, Siamese and Colorpoint Shorthairs). Pat has
also considered chimerism as a possible explanation.
COLOURPOINTED
AND MASKED CATS
Copyright
2002-2015, Sarah HartwellThe
colourpoint pattern is a form of albinism and is known as the
"Himalayan pattern" although it's more often called
"Siamese" pattern by cat lovers. It is found in a number of
mammals including rabbits and goats from the Himalayas region. The
exact effect of the gene depends on the temperature since its effect
is inhibit colour in warm areas of the body and allow colour to
develop on the cooler areas such as the ears, face, legs, tail and
(in males) scrotum. In warmer climes, there is less contrast between
the body colour and the point colour. Colourpoint kittens are born
with almost pure white coats on a pink skin because of the uniform
warmth in the womb; only after birth does the point colour develop.
In cooler regions, the contrast may be very pronounced. The
characteristic blue eyes reflects reddish at night because albinism
affects the tapetum (reflective layer) of the eye; non-albino cats'
eyes reflect greenish.
The
most familiar colourpoint breeds are the Siamese and Himalayan
(Colourpoint Longhair/Colourpoint Persian), but there are a number of
others. Most, if not all, colourpoint cats have Siamese somewhere in
the history, or at least have Thai, Asian or Japanese cats in the
family tree since this pattern occurs naturally throughout
Asia and
into
Russia . The Burmese has a "low contrast" form of the
colourpoint gene andthe Tonkinese is halfway between Burmese and
Siamese. There are a series of gene variants and combinations
controlling the contrast between body colour and point colour; these
are shown in the charts below.
SIAMESE/COLORPOINT
SHORTHAIRModern
Siamese come in different varieties - one is the "Modern"
Siamese with its exaggerated wedge-shaped head, bat ears, a muzzle
sometimes referred to as a "banana nose" and an often
excessively tubular body. Another is the "Classic" Siamese
that resembles the 1950s-1960s Siamese and intermediate between the
more
robust "Old-Style" that resemble early Siamese imports
and the extreme modern Siamese. There is a continuum between the
old-style and the modern Siamese, with most people considering there
to be two types: old and modern. Personally speaking, I find many of
the modern showbench Siamese extremely
ugly creatures and many
owners, as opposed to breeders,
prefer a less extreme style of cat.
For this reason, there are also traditional (old) style Siamese (Thai
Siamese) being bred, recreating the chunkier
shape familiar in the
late 1800s and early 1900s. Both originate from the same cats.
The
naturally-occurring colourpoints I have seen in
Malaysia and Thailand
are closer in type to those being bred to the older (non-
ultra -typed)
Siamese standard. Of the conformation of Siamese cat,
Phyllis Lauder
wrote in "The British, European and American Shorthair Cat"
(1981): "Cats of
eastern provenance have not shown long heads:
in the 1930s and 40s there was, at the Natural History
Museum in
London, a stuffed Siamese cat, and this animal’s head was ‘as
round as an
apple ’ to
quote one of England’s
prominent experimental breeders, the late B. A. Stirling-
Webb . The
taxidermist’s work showed a large cat of strictly ‘domestic’
type. "
There
are many theories about the
origin of the Siamese. The Seal Point
Siamese was known as the
Royal Cat of Siam because it was found in
palaces. It seems to have been familiar in the ancient Siam city of
Ayudha which was
founded in 1350 and was Siam's capital until
destroyed by fire during the Burmese invasion of 1767. In Bangkok's
National Library there are manuscripts from Ayudha which describe
native fauna. "The Cat Book of
Poems " depicts pale coated
Seal Point Siamese, describing them as having black
tails , feet and
ears with white hair and blue eyes. No one really knows what the cats
of ancient Siam looked like or whether they were bred for body type
or purely for colour and pattern with little regard for shape.
Another problem is that artists often stylize their illustrations.
The
extinct Annamese from Vietnam is also considered by some to be the
ancestor of modern Siamese and that the Siamese cat of Thailand arose
through interbreeding Birman and Annamese cats. An early description
of the Seal Point Siamese in 1676 calls it "Vichien Mas"
(meaning "diamond mouth") and drawings depict extreme
expression of the colourpoint pattern - dark ears, nose, paws and
whiskers. A cat with Siamese markings appeared on an old
engraving discovered by the naturalist Pallas on his journey into Southern
Russia between 1793 and 1794. It was the opinion of Sir
Russell Gordon (who closely studied these cats) and
Auguste Pavie (
French explorer and one time resident at Bangkok), that the Siamese cat
derived from a cross between the Burmese
Temple Cat (i.e. Burmese,
not the Birman which was known as the "
Sacred Burmese Temple
Cat") and the Annamite Cat. The Annamite cat was described as a
slender, small and gracefully
built variety of cat with a short tail.
Like so many varieties it was said to have disappeared due to
interbreeding with introduced domestic cats.
Although
apparently favoured as
palace cats, there is no
clear record of an
distinct Siamese breed until the 1800s. The British became interested
in Siamese cats and imported them from Siam. The earliest documented
imports were during the 1870s, but these were apparently not bred. In
1884, the departing British Consul-General
Gould was given a Siamese
cat by the Siamese
king . He brought the cat to England and its
progeny were exhibited at
Crystal Palace in 1885. The early Siamese
cats were round-headed, solid and muscular, but even so, their
appearance was so extraordinary that they were described as an
"
unnatural
nightmare of a cat".
In Ceylon, the Siamese cat was, for a while, known as "Gould's
Cat", having been introduced there by Mr Gould. The Burmese
Sacred Cat was known to early British cat fanciers as the "Gold
Cat". A wild cat of the region was known as the "Golden
Cat" (Temminck's Golden Cat) or "Bay Cat". HC Brooke
believed these similarities of name to be the reason that Temminck's
Golden Cat was
claimed to be an ancestor of the Siamese. The first
champion Siamese, "Wankee," was born in Hong
Kong in
1895 and exhibited in 1898. He was relatively large and round-headed by
modern standards, but had a more distinct muzzle and longer body than
modern appleheaded (as in the rounded headed "applehead"
dolls of the USA) Siamese - more of an intermediate type.
Frances Simpson ,
editor of "The Book of the Cat" (1903) included
contributions from several early breeders of Siamese cats. While
acknowledging the existence of blues, blacks, whites and tabbies in
Siam, she stated that only the "Royal Siamese" and
"Chocolate Siamese" were recognised in England at that
time. These were sometimes bred to each other although opinions on
the quality of the offspring were contradictory. The royal Siamese
was sometimes bred to white short-hairs because the English type was
preferred over the foreign type by judges. Because white is dominant
and masks other colours, "
sports " with "any other
colour" points occurred a generation or two later.
Most
early breeders considered the Siamese cat to be more delicate than
the English cat, having delicate lungs and being prone to
disease and
other upsets. Many did not risk sending their precious cats to shows.
One early breeder noted a rarity of female kittens in a litter, the
average being 5 males to 2 females. The kittens were said to be
difficult to
rear , as they suffered from worms and teething, and it
was common to foster Siamese kittens on English cats to make them
more robust and healthier. Males were described as extremely
powerful, great fighters, had terrible voices and would kill strange
cats and
fight dogs.
"The
males are, however, antagonistic to others of their sex, and fight
with a terrible persistency. I have heard of a stalwart fellow who,
being allowed his liberty, cleared the neighbourhood of all other
wandering toms. When made neuter, Siamese become most charming home
pets."Miss Forestier-
Walker and her sister, Mrs Vyvyan, had
received a pair of
Siamese cats from the Siamese Palace in 1884-5; Miss Forestier-Walker
wrote
"Siamese
cats were first introduced into England about twenty -five years ago,
but were not often seen until a few years later. Since then they have
become fairly common. There are two distinct varieties in the present
day. (1) The royal cat of Siam" by which she meant the seal
point Siamese, "(2) The chocolate cats are deep brown in colour
showing hardly any markings, and have blue eyes. The tails are
sometimes straight , which is not a fault; but a knot or kink in the
tail is a peculiarity of the breed, and therefore desirable. In
England it has been asserted that this is a defect , but in Siam it is
highly prized […] In the East a cat with a kinked tail fetches a higher price .""There
is a legend that the light-coloured cats with blue eyes represent silver; the dark cats with yellow eyes, gold; and that the possessor
of both will have plenty. This rather gives the idea that originally
the eyes of the pure chocolate cat were yellow, and that the present
variety has been crossed with the royal cat. It is a great mistake to
mix the varieties, as the result after they become adult is a
blurring of the markings and a patchy coat." What
Miss Forestier-Walker was describing was the range of colours from
the brown Burmese (the "Burmese cat" depicted in the 1903
book was an Oriental ticked tabby), through the "blurred"
or less distinct markings of the mink range (Tonkinese) to the
sharply defined colourpoint of the Siamese.
Champion
Wankee was bred in Hong Kong in 1895, the offspring of a female
kitten stolen from the Palace in Siam, and imported to England aged 6
months. His
owner , Mrs Robinson, wrote
"One
of the most beautiful of the short-haired cats is undoubtedly the
royal cat of Siam, and the breed is increasing in popularity ; but is
never likely to be common, as the cats are delicate in this country.
[…] The [standard of] points of the chocolate Siamese are the same
as the royal, with the exception of body colour, which is a dark rich
brown all over, thus making the markings less noticeable. All Siamese
darken with age, and when they get dark there is a tendency to call
them chocolates. I know of only one real chocolate - Mr C Cooke's
'Zetland Wanzies' - so consider them more likely to be a freak than a
distinct variety."She
also described the different body types: "
Of
the royals there seem to be two types in England: the one - rather a
small, long-headed cat, with glossy , close lying coat and deep blue
eyes, and with a decided tendency to darken with age - is generally
the imported cat or having imported parents; the other is a larger
cat, with a rounder head, a much thicker, longer and less close-lying
coat, and the eyes a paler blue (these cats do not darken as much or
as soon as the other type, and have generally been bred for several
generations in England)."According
to another early breeder, Mrs Parker Brough,
"There
are distinct varieties of Siamese known to fanciers - the palace or
royal cat, the temple cat (chocolate), and there is likewise the
common cat of the country, which is also found within the palace. The
points of the chocolate cat are identical for shows with those of the
royal except body colour, but the imported chocolate is often dark
chocolate, with blue eyes, stumpy tail with a marked kink, short
legs, and heavy , thick body. There are not many chocolates exhibited,
owing to the preference given to the royal variety. It must be
understood that there is no definite royal breed as such, but the
palace breed seems to have originated by selection ." Lady
Marcus Beresford wrote that Siamese imported from a Bangkok temple
"
differed
from the royal Siamese, being darker and having a more pointed head
and face, and their eyes were larger and fuller."According
to Miss Forestier Walker
"This
breed is said to be kept very carefully in the palace in Bangkok -
hence the title 'royal' - and is by no means the common cat of Siam.
One gentleman (a missionary), who had lived there fifteen years, had
during that time seen only three. A few years ago there was a pair of
these cats in the Zoological Gardens at Bangkok, but they were very
poor specimens. […] The first specimens were brought to England
about twenty-five or thirty years ago, and Mr Harrison Weir says that among those who possessed them were Lady Dorothy Nevill, whose cats
were 'imported and presented by Sir R Herbert of the Colonial Office.
The late Duke of Wellington imported the breed, also Mr Scott of
Rotherfield.' "Mrs
Parker Brough
wrote
"Until recently the Siamese was but little known in Europe, but
occasionally was to be found in the various zoological gardens. At
present there is a fine female specimen to be seen at the Zoo at
Frankfort-on-the-Main, having been purchased from the King of
Roumania. One or two are to be seen at the Hague. London has the
first one it has had for six years, but it is not shown owing to its
want of condition. […] A point on which the Siamese fancy is divided is where the ideal cat should have a kink in the tail or
not.[…] There is a peculiarity in breeding the Siamese - i.e. the
rarity of female kittens in a litter, the average seeming to be five
males to two females. […] They have naturally rather delicate
lungs."Some
of those early (1890-
1902 ) Siamese were evidently large, robust
creatures, for example a neutered male called Attache was described
as very large and powerful, with
massive limbs, and an unconquerable
antipathy to all other cats of any description. Frances Simpson
summed up in 1903 by saying
"I
do not believe that Siamese will ever become common in England, for
many reasons. These cats are expensive to purchase , difficult to
rear, and fancier are afraid to risk them in the show pen; but in
spite of these drawbacks, I think as time goes on and the Siamese Club extends its labours, we shall see and hear more of these really
curious creatures, for what we call the royal Siamese bears no
resemblance to any other cat, and the distinguishing difference ,
being so great, tend to make the breed one of our best show cats and
a clear class to itself, for the Siamese of the purest blood should
not be crossed with other cats. We have heard of 'any other colour'
Siamese, but these cats of varied hue claiming to be Siamese are but
the offspring of a cross. We have been told of black and blue and
tabby Siamese; but the fanciers of Siamese look askance at these
freaks, and feel that it is worse than useless to attempt to produce
any other variety than that which we have learned by custom to
designate the Royal cat of Siam."A
great deal has been written about the origins of the Siamese breed.
Sydney W
France , editor of "Cats and Kittens" magazine,
wrote the book "Siamese Cats" in 1949. This was the first
major publication on Siamese cats since 1936.
"The
history of the Siamese in this country is a very short one, and it is
true to say that they have only been here within living memory, and
that the first ones actually were from The Royal Palace of Siam. Even
on this point there is much controversy and it is interesting to note
that the first Siamese of which there is any record were said to have
been brought to England in 1884 by Mr. Gould who was then Consul
General in Bangkok at that time. In 1886 a pair of cats and two
kittens were brought to England by Mrs. Vyvyan, these had actually
been procured from the King’s Palace in Bangkok. In the same year,
Mrs. Walker, the General’s daughter, brought over one male and
three females. There is no doubt that at that time the true Siamese
were kept in the Royal Palaces and Temples , and that few of them ever
found their way from there except as gifts, which were then
considered as of great worth."In
direct contradiction, Ida M. Mellen, well-known American
authority on
cats, in her "
Practical Cat Book" (1939) writes
"Although
this cat generally is referred to as the Royal, and even as the
Sacred Siamese, it is the common cat of Siam, just as the Manx,
equally an aristocrat, is the common cat of the Isle of Man."France
also reproduces a
letter Mellen received from Dr. Hugh M. Smith,
Adviser in Fisheries to His Siamese Majesty’s
Government between
1923 and 1934. In addition to the Siamese cat, he mentions a "mauve"
cat which is no doubt the Si-Sawat (Korat) and kinked-
tailed cats
which were, and still are, common throughout Thailand, Malaysia and
Singapore (and in the modern Japanese Bobtail breed).
"I
was well Acquainted with cats in Siam, but made no special study of
them. There appear to be two races peculiar to the country: the
common form with pale fawn colour, black or dark brown feet, tips of
ears, tail and muzzle, and blue eyes, well-known to cat fanciers all
over the world, and a form of uniform mauve or Maltese colour with
yellow eyes. There are no "palace" cats in Siam. There are
no "royal" cats, although the strikingly marked creatures
would be the natural ones to be kept in palaces. Any person can have
a Siamese cat, and as a matter of fact there are many people outside
the palaces and many foreigners who keep such cats as household pets.
There are no "temple" cats. The Buddhist priests, who do
not live in the temples but in special buildings in the temple
grounds, may keep cats, as they do dogs. A Siamese prince whom I know
very well was visiting in London and was interviewed by one of the
thousands of Siamese cat fanciers there. He told her there were more
Siamese cats in London than in all Siam.You
probably know about the cat not peculiar to Siam but found over much
of South Eastern Asia, which always has a sharp kink near the tip of
its tail. It is of various colours but never of the special Siamese
cat colour, and is of no interest except for its tail."There
is no doubt that Siamese cats came from Siam (modern day Thailand),
but some disagreement in the 1940s as to their "royal"
origins. France disagreed about Siamese not being Royal cats. As well
as mentioning the various royal legends relating to the tail kink and
squint (faults in show cats), he stated:
"This
does not bear out my own information on this point, because whilst
living in Jersey I had the good fortune to meet and become friendly
with Major Walton of Verona House, Grouville, who was until recently,
in Siam in connection with the Rice purchasing Commission, when he
and his wife , both cat lovers, became friendly with the Prince Regent of Siam. Major Walton told the Prince Regent that he and his wife wanted to bring back some Siamese to England, but had not been able
to find any at all in the country that were for sale. Before leaving
Siam, Mrs. Walton was presented with a pair, male and female by the
Prince Regent, and I actually went and saw them in quarantine in
Jersey where they had a litter of five kittens, of which, later on, I
bought two; a male and female.These
Siamese cats had coats of extremely fine quality texture, and colour,
and very good head and body shape, splendid long whip-like tails, but
with eyes which definitely failed in colour according to our
standards here in England. They appear to be hazel , whereas there is
no doubt that we have enormously improved the eye colour and have
cats with beautiful deep blue eyes."Major
Walton’s remarks to France about the Siamese cats of the Royal
Palaces were also mentioned in an
issue of "Cats and Kittens"
magazine, of which France was editor. This prompted the following
letter from Mr. A. N. M. Garry of Minehead, Somerset:
"My
wife and I are ardent cat lovers, and having spent most of my working
life in Borneo, I feel I have some justification for writing to you
about Siam and its cats. When I was in Siam in 1930, I was told that
there were two distinct types of Siamese cats - apart from the Malay
cat and crosses with it. The first is the one we see in England, but
I think its points are a shade different, [chocolate] brown instead
of seal. The second, which was said to be peculiar to the Royal
family and palaces, had the body colour of the first - but not the
points; and hazel eyes.Having
been a contemporary at Eton with the then King, I got a special permit to see the Bangkok Palace more thoroughly than the usual tourist does, and I saw one or two of these "Royal" cats,
whose appearance was (to the best of my recollection after so long)
as I have described. At that time, the export of the first type,
except neuter ones, was absolutely forbidden, owing to the fear that
they might become extinct in Siam, because so many had been exported.The
second type was absolutely unobtainable, far less exportable, for it
was not to be seen outside the royal palaces. But owing to the war,
and the various constitutional changes that Siam has undergone, the
rules may well have been relaxed. It certainly looks as though Major
Walton has been lucky enough to get hold of a pair of the scarcer
"Royal type." Incidentally, I hope he is aware of the
tendency of Siam born Siamese to chest troubles ~ damp or cool weather . I know of several people even out there, who have lost their
pets from this cause.I
am afraid I am quite unable to agree with Mrs. Adney’s friend that
a Siamese must have a kinked tail to be considered pure bred out
East. In my experience, the .connoisseur out there, just as at home,
demands the straight tail; but the fact remains that the majority of
so-called Siamese cats in Malaya and Borneo have kinked tails, owing
to one of their parents or forefathers having been a Malay cat. They
still make lovely pets, and have the characteristics of pure-bred
Siamese, such as the deep voice and the love of following their
owners like dogs.The
Malay cat, like the British domestic cat, is of varied colours,
ginger, black, black and white, tabby and tortoiseshell. Some having
Persian forbears, are long haired. Practically all have kinks, and I
had one, a beloved ginger, called Peter, who was considered a cat of
particularly good omen , as he had two kinks quite close together.
They were very tender, and he hated them being touched. He lived to
the age of seventeen - very old indeed for an animal in the tropics.
He died a few months before the Japanese invaded Borneo."The
first Siamese fanciers club was founded in Britain in 1902 at which
time they were apparently variable in type. Possibly the conformation
depended on which cats the early Siamese had been out-crossed to,
there being few pure Siamese in the country at the time, or to the
variability of imported cats and inbreeding from a limited gene pool.
Early
photos show differences between the "compact" cats and the
"lithe" cats, but the difference is nowhere near as extreme
as that between modern "classic" and "old-style"
Siamese. Early Siamese are more robust and have more rounded heads
than modern classic Siamese, however, they were longer-bodied (less
cobby) and more wedge-headed than the British Shorthairs and Persians
of the time. The 1892 Siamese breed standard (Harrison Weir)
described them as a
marten -faced, Oriental type of cat distinctly
different from the cobby, round British cats. Weir described the
Siamese wedge as beginning at eye level, at the muzzle (in modern
cats it begins from the ears downwards).
Based
on only a few imported Siamese, early breeders believed that there to
be 2 types of Siamese. Judge and Siamese-fancier, Mrs. Carew Cox,
said (reported 1903) "
There
appear to be two distinct types - the compactly built, short in body,
short on legs, and round in head; and the long-bodied, long-faced,
lithe, sinuous, and peculiarly foreign-looking variety."
An
early Siamese breeder, Mrs. Robinson said, "
Of
the royals there seem to be two types in England: the one - rather a
small, long-headed cat, with glossy, close-lying coat and deep blue
eyes, and with a decided tendency to darken with age - is generally
the imported cat or having imported parents; the other is a larger
cat, with a rounder head, a much thicker, longer, and less
close-lying coat, and the eyes a paler blue (these cats do not darken
as much or as soon as the other type, and have generally been bred
for several generations in England)."The
Siamese Cat Society of America was founded in 1909 although the date
of their arrival in the United
States is not precisely known. Many
early Siamese had kinked tails and cross-eyes or a squint; these
faults have largely been bred out of modern Siamese.
In
1949, Kit Wilson (Vice-Chairman of GCCF) wrote in the Sydney W
France's book "Siamese Cats"
"A
few years before World War 2, interest was beginning to be shown in
Blue Pointed Siamese. These had caused some considerable controversy
among breeders, many of whom were of the opinion that they were
"sports" and therefore could not be bred true, but a few,
whose convictions based on research refuted these opinions, and "The
Blue Pointed Siamese Cat Club" was formed.Lately
particular interest is being taken by some breeders in Chocolate
Pointed Siamese. The greatest living authority on this fascinating
variety is Miss Wentworth Fitz-William of Slingsby Yorks., one of the
most valued breeders and judges of many years standing who has always
owned some, and from whose stock most of those in existence to-day
have sprung. [...] It is an interesting fact however that there are
considerable numbers of chocolate points in France, but their Owners
have no idea as to their origin, the strain however must have been
good, as they are without exception the only Siamese worth showing
over there (or at least they were at the January 1948 show, when I
was judging. [...] For those who have never seen a Chocolate they are
often smaller in build than the seal or blue point, and their points
are of a rich milk chocolate colour.Blue
points are very popular in the U.S.A. many of them winning high
awards at shows over there, but, although I am open to correction, I
have not heard of any chocolates. I only had one of these in my classes at the Danish (Darak) show in 1946, and this was a very poor
specimen and hardly worthy to carry on the breed, and was, in all
probability a " sport ." It had been bought at quite a high
price when a kitten as a seal point, and having met the vendor as
well as the owner I am of the opinion that neither of them had the
slightest idea that it was anything other than the usual seal.While
writing this an interesting fact has come to light with regard to
chocolate points. Mr. Brian Stirling-Webb, the Hon. Treasurer of the
Siamese Cat Club has been making a close study of this variety, and
he states that practically every chocolate in the country can trace their pedigree back to an imported cat brought into this country in
1896, this may also account for those on the continent . Who knows,
perhaps another from the litter was taken over there and that those
now being shown are direct descendants."In
that book, Wilson wrote in disapproving tones of cross-breeding of
Siamese. Luckily the experiments in breeding Siamese with other
breeds (not species!) did continue in spite of early disapproval,
leading to a wide variety of attractive Oriental and Foreign
varieties.
"From
America we have heard of the Black Siamese with orange eyes but to my
knowledge no specimen has ever been seen in this country. Then there
have been long haired specimens, described as Burmese, they have the
same colouring as the seal point, and long fur, which although in no
way comparable with the Persian yet is definitely more long than
short. Experiments however have proved that in breeding Siamese to
other species - varied forms can be made - this practise is not to be
encouraged, as it may lead to definite malpractices as have occurred
in other livestock."Her
comments show some confusion between Burmese and Birman. The "Black
Siamese" is probably the Foreign Black; essentially a Seal Point
Siamese without the colourpointing gene.
While
they became more popular in the US and Europe, the purebred Siamese
was dwindling in its native Thailand. In the 1950s, a breeder
obtained 3 Siamese kittens of the robust type from Thailand. While
the debate over "2 types" of early Siamese continues to
this day, what is certain is that early Siamese cats were far more
moderate in type than either of their modern counterparts. During the
early 1960s, the "robust" Siamese lost out to the longer
thinner type. While owners often preferred a moderate cat, exhibitors
preferred an extremely slender cat with a very long, triangular head,
almond-shaped eyes, and flaring ears. This look caught on with
show-oriented Siamese breeders and judges. A cat called "Fan Tee
Cee" was an early example of the increasingly extreme
conformation that was impressing the judges. Fan Tee Cee appears many
times in some pedigrees, demonstrating his contribution to the
new-style Siamese. The
growing number of new-style Siamese put
pressure on cat fancies to change the breed standard to reflect the
changing shape, putting old-style cats at a disadvantage.
The
original Siamese cats were Seal Points ("black" points)
although Blue Points and Chocolate Points also appeared in Siamese
litters and were for many years considered "poorly marked"
Seal Points. In the USA at the end of the 1960s, breeders in Britain
and the USA were working with different colours of Siamese-type cats
e.g. ebonies, reds, chestnuts and lavender.
Lynx -points (tabby
points) were being bred in Britain. Because blue-pointed Siamese
appeared spontaneously in seal-point litters, they were known as
"sports" or "freaks". In 1896 Louis Wain, father
of the modern cat show, refused to judge a Siamese on grounds that it
was blue rather than seal. When the first book on Siamese was written
by Phil Wade she wrote, ‘"Even the best blue pointed cannot, I
think, equal the
beauty of our seal pointed cats and I can see no
real object in trying to breed them. Their
value at the moment is
their scarcity, but I cannot believe there will ever be a great
demand for them."
Tabby
Point Siamese had been mentioned as early as 1902 in Britain. Between
1944 and 1949, they were bred in
Scotland and known as Silverpoint
Siamese. They were introduced to the cat fancy at a London cat show
in the 1960s and in 1966, the Tabby Point Siamese was granted
recognition by the Governing
Council of the Cat Fancy (GCCF).
By
1986 the old-style Siamese had vanished from the showbench and many
people assumed that Siamese cats had always had a long body,
wedge-shaped head and disproportionately large ears. Siamese cats had
become extreme parodies of the original imports. In Australia, some
Siamese appear even more extreme than their American cousins - with
larger ears and even more fragile bodies. In Britain they have also
become more extreme - and ugly - in type, especially facially. It
really is up to judges to do a sanity check and
prevent the cat from
being taken to ever greater extremes of type.
In
the USA, the old-style cat was still being bred in a small way even
though it could no longer
compete against the modern style cats. It
was popular with the general public. There is a separate Siamese
breed known as the traditional and recognised as distinct from
classic Siamese by some registries in the USA. The term "Old-Style
Siamese" is often used.
In
Britain, interest from the public wanting to buy old-style Siamese
cats has led to a revival of the Traditional style Siamese during the
late 1990s. Those would be owners expressed dismay at the
wedge-headed Siamese seen on the showbench. On mainland Europe the
Thai Pointed is a naturally occurring colourpointed (Blue-point,
Lilac-point) Korat variant. The Thai Siamese is a European term for
the traditional (round-headed) style of Siamese derived purely from
non-ultra-typed Siamese cats. The term Thai Siamese has sometimes
been used for colourpointed Korat variants.
In
the USA, the Siamese is recognised in seal point, blue point,
chocolate point and lilac point. Other colours (red point, cream
point, tabby point, tortie point etc) are known as Colorpoint
Shorthair (Colorpoint Oriental Shorthair). Elsewhere, these other
colours are classed as Siamese. The additional colours were
introduced through out-crossing, however they have been so
extensively back-crossed with Siamese cats that only the most ardent
purist would claim that they are not Siamese! The additional colours
are not permitted in the traditional old-style of Siamese which seeks
to breed only the traditional colours as well as the more moderate
conformation.
Lynx point (UK) 1990.
Lynx point (UK) 1990.
Breakthrough
Tabby Pattern in Colourpoints
Cats
born of Siamese parents sometimes have a tabby/spotted pattern
breaking through so strongly that the cat appears to be a
tabby/spotted Oriental or a tabby/spotted sepia (Burmese) pattern.
Viewed closely, the pattern colour is heavily ticked on a paler
ticked background. This breakthrough pattern is evident from
kittenhood and not the same as age-related darkening in Colourpoint
cats. It is inherited, possibly due to modifier genes that prevent
the colourpoint gene from properly inhibiting the colour/pattern on
the normally pale areas of the body.
A
stongly marked seal-tabby Siamese with a breakthrough spotted pattern
(looked like a Spotted Tabby Oriental with blue eyes) mated to a
Siamese produced a litter containing one strongly marked classic
tabby that looked more like a Classic Tabby Oriental with blue eyes.
The breakthrough pattern has become a problem in some Australian
lines. Another example was a lynx-point Siamese female (old-style
conformation) in England that looked like a slightly washed out tabby
with blue eyes. Her colour raised
questions as to whether she was
genuinely colourpoint or was genetically mink or sepia pattern from
Tonkinese lines.
BALINESE/JAVANESE
AND SINGHALESEAs
with the Siamese, there are 2 types of Balinese - Balinese and the
traditional style of Balinese (Apple- or round-head). In both cases,
they are semi-longhaired versions of the 2 different types of Siamese
Semi-longhaired
variants have appeared now and again in purebred Siamese litters.
There have been various explanations. The recessive nature of the
longhair gene means it can be carried for many generations without
coming to light - in which case the gene was present in one of the
early imports of Siamese. Some say that at least one Chinese tapestry
depicts pointed longhairs. Longhair can also occur as a simple
mutation; again the gene may be carried hidden for many generations.
An unlikely explanation is that it came from the domestication of the
longhaired Pallas cat (F manul) of
western China although genetic
evidence is lacking. More feasibly early british breeders may have
outcrossed early Siamese imports to longhaired cats or to shorthaired
cats which had a recessive gene for longhair. Possibly they were
crossed with Turkish Angoras, another breed popular at the time.
After
many years of hiding their existence, a "Longhair Siamese"
was registered with the Cat Fanciers Federation in 1928, however they
were not bred in earnest until 1955 when Marion Dorsey began breeding
and showing the longer-haired variety in the USA. Since long hair is
a recessive trait, these long-haired Siamese bred true. To keep it
separate from the conventional shorthaired Siamese it
required a new
name. "Balinese" was chosen to reflect its
grace , like that
of Balinese dancers, and also because
Bali is close to Thailand and
this reflects the breed's
links with its Siamese ancestors.
In
1961 it was recognized in the USA and accepted for
registration in
the same point colours as Siamese: seal, chocolate, blue and lilac.
Other colours were introduced from outcrossing: red, cream, tabby,
tortie, cinnamon, fawn, smoke, silver and more. These were registed
in the USA in 1979 as Javanese, but are considered Balinese in other
countries since they have been so extensively back-crossed to
Balinese that they can no longer be considered
hybrids .
The
development of the Balinese paralleled that of the Siamese. In the
1950s most Siamese and Balinese cats were less extreme than we are
used to today. The more extreme look gained in popularity for the
Siamese and its longhaired counterpart followed suit since Balinese
were bred back to Siamese to
improve the type of the Balinese. In
addition, much of the Balinese cat's long fur was lost, leaving it as
a more-or-less shorthaired cat with a plumy tail, a side-effect of
breeding back to the Siamese. As with the Siamese, a few breeders
preserved the older style and it is now becoming more popular in its
own right.
In
the 1960s, a cat very similar to the Balinese was bred in the USA and
Canada . These resulted from crossing Siamese cats to red/red-tabby
Angora (presumably the Turkish Angora) resulting in semi-longhairs
with Siamese conformation and red or tortie points. The fur was
shorter, but fluffier than the Balinese and the tail much fluffier.
The
temperament was also calmer than the Siamese due to the Angora
influence. Essentially these were the red-series equivalents of the
Balinese which US registries recognised only in seal, blue, lilac anc
chocolate. The Singhalese was allowed to be bred to either Balinese
or Himalayan and the progeny could be registered as Singhalese. It
was judged to the Balinese standard with allowances made for its
different coat type. Over time, it appears to have lost out to the
Balinese.
BIRMANThe
Birman has the colourpoint pattern with the addition of white
bootees/gauntlets. The Birman arrived in Europe around 1920 and has
also been known as the Tibetan Temple Cat and the Sacred Cat of
Burma . Modern Birmans are descended from a pair brought to Europe by
Mr. Vanderbilt. It is also reported that 2 Englishmen, Major
Gordon-Russell and August Pavie, then living in France, received a
pair of Birmans from the Kittah people in 1919 as gratitude for their
part in saving the temple from being overrun by invaders. The
remainder of the
tale is the same: the male unfortunately died during
the voyage, but the female was pregnant. Of her kittens, only a
female survived. This meant two female Birmans and no male. To
re-establish the type, these cats were crossed to Siamese,
Colourpoints and White Longhairs.
In
1927, judge Mrs Basnett reported on the
Paris Cat Show
held on
14th and
15th of
January by the Cat Club de France and wrote "The Sacred Burmese
Temple Cats interested me very much, with their long fur on the tail
and coat resembling that of a poorly bred Persian; their colouring is
exactly like that of the Siamese, but their feet sometimes have white
toes. I was given to understand that they are very difficult to rear,
only about one in ten
survive . I do not think they possess the same
quick movements as the Siamese, life to them seems much more dreamy
and
slow , but they are very loving and intelligent." This
clearly referred to the Birman; confusingly the name Burmese Temple
Cat was also used at that time for the gold-eyed brown Thai cats
analogous to modern Burmese or brown Orientals.
As
a result of its early near-loss and re-creation, some say the Birman
comes from temples in Burma, while others claim it is a breed
entirely manufactured in France using Siamese andblack-and-white
longhairs. French breeders had to re-create the Birman not once, but
twice since the Second World War caused pedigreed cats in Europe a
great set back. Only two purebred Birmans survived and their
offspring formed the new post-war foundation of the Birman breed in
France. They were bred with other longhairs out of necessity, but by
1955, the Birman was once more recognised as pure bred and there was
no more outcrossing. In the early 1960's Mrs Elsie Fisher and Mrs M.
Richards imported the first Birmans into Britain from France.
While
some claim the cats were created in France, it is interesting that in
1960, a pair of "Tibetan Temple kittens" was given to a
North American cat
lover and were identical to the Birman cats being
bred in Europe. In recent years new point colours have been
introduced, including chocolate, lilac point, red, cream, tabby and
tortie point. Not all societies recognise the newer colours. It's
worth noting that the older style of Birmans tended to develop
"hood-type" head colouring.
Early
in the breeding of Birmans in the USA, Gertrude Griswold attempted to
keep the Birman breed under her control. It was possible to adopt a
female Birman from her for breeding, but to actually breed the cat,
there had to be a notarized agreement (a contract with the Griswolds)
that the sire would be of Gertrude Griswold's choosing and the
adoption of the resulting kittens into new homes would be under her
jurisdiction. Later on, Ann
Baker attempted a similar degree of
control over Ragdolls, another colourpointed breed.
Birman
variants are known e.g. the "
Khmer " being a bootee-less
Birman-type cat and the Tibetan being a Birman in solids colours and
tabbies. Some of the early Birmans lacked the gauntlets.
TEMPLECAT
(BIRMAN SHORTHAIR)The
Templecat is the shorthaired equivalent of the Birman. It is a Birman
in every
respect apart from the slightly springy short hair. It was
developed in New Zealand by outcrossing Birmans to a cinnamon spotted
tabby Oriental.
The
Templecat began development in 1995 and was originally called the
Birman Shorthair. It later became the Tsuncat (after a temple priest
in
mythology surrounding the Birman cat) and became the Templecat in
2001.
The
early colours were seal point, blue point, chocolate point, lilac
point, cinnamon point and fawn point It is now also in red point,
cream point and in tabby and toritie points.
HIMALAYANThe
Himalayan is a Persian in a colourpoint coat and with blue eyes. In
the UK it is known as the Colorpoint Longhair (or Colourpoint
Persian). In the USA, Himalayans with tabby or tortie points are
known as Colorpoint Longhair in some registries.
Experimental
breeding of Siamese and Persians
dates back as far as the
1920s . The
first recorded deliberate cross between Siamese and longhaired cats
was made by
Swedish geneticist T Tjebbes in 1924 using white
longhairs. However it was not his
intention to create a new breed for
the cat fancy. In 1931 in the USA,
Virginia Cobb and Dr.
Clyde Keeler
began an experimental breeding programme with the
purpose of learning
the inheritance involved in producing a Colourpoint Longhair. In
1936, they produced the first Himalayan kitten. They published an
article about colourpoint inheritance in the American Journal of
Heredity. Having accomplished their aim, the breeding programme was
abandoned.
Sydney
W France's book "Siamese Cats" (1949) also mentioned the
crossing of Siamese cats with Persians, a practice which eventually
led to the now familiar Colourpoint Persians (Himalayans). In 1949,
such breedings were regarded with a mix of
curiosity and disapproval.
"Some
curious experiments have been made from time to time in an effort to
cross Siamese cats with Persians and even the tabby cat, and at least
two treatises have been written on the subject. "Siamese-Persian
Cats" by Clyde E. Keeler and Virginia Cobb, "Journal of
Heredity" v. 27. No. 9. Sept. 1936, and "Crosses with
Siamese Cats" by K. Tjebbes, Journal of Genetics, V. 14. p. 335,
1924. From this we find that Swedish Dr. K. Tjebbes in about 1924
crossed a white Persian female with a Siamese and the colour of the
Persian dominated to the extent of seven white kittens and three
coloured ones. Back crosses all gave 50 per cent . white. Like Siamese
colouring, short hair dominated.It took Mrs. Virginia R. Cobb five years of experiment working in
conjunction with Dr. Clyde E. Keeler of the Harvard Medical School to
produce the first successful experiment and breed long-haired Siamese
kittens, using black Persians instead of white, as used by Dr.
Tjebbes in his experiments. Only pedigree cats were used, and from
each litter only the most perfect kittens were selected, to be mated
up in due course to carry on the experiments. A black Persian male
was mated to a Siamese female, and a Siamese male was mated to a
black Persian female. The kittens in every case were black and
short-haired. After a time a female of one of these litters was mated
to a male of the other, and produced among her kittens a long-haired
black female! this female being bred back to her short-haired black
father. She subsequently had a litter containing two Siamese-Persian
kittens which had the long hair of the Persian and the markings of
the Siamese. This mating was repeated and of the eleven kittens three
were long-haired Siamese, the other eight being black. These three
long-haired Siamese kittens had the blue eyes of the Siamese, and the
same voice, which they used just as often as the true Siamese do.In
1939 when Ida M. Mellen reported these facts in her practical cat
book the experiments were still proceeding."In
1931, Virginia Cobb (
Newton Cattery) and Dr. Clyde Keeler (Harvard
Medical School) conducted an experimental breeding programme to
produce a Colourpoint Longhair. They wanted to study the inheritance
of longhair and colourpoints to produce a "longhaired Siamese".
They first crossed Siamese with Black Persians. This resulted in
black shorthair kittens. They mated the offspring together and
achieved the desired longhaired Siamese. In 1936, Keeler and Cobb
produced the first Colourpoint Longhair (Himalayan) type kitten,
"Newton's Debutante". They published an article and the
"
formula " for producing her in the American Journal of
Heredity and then abandoned the breeding programme. They had
accomplished the goal of studying how the factors were inherited
(longhair and colourpoint are both recessive), but did not intend to
create a new breed.
In
Britain, at the Siamese Cat Club Show in 1947, Brian Stirling-Webb
was approached by a cat owner, Mrs Barton-Wright, who wanted to mate
her "long-haired Siamese
queen " to one of his Siamese stud
cats. She lived close to his Briarry cattery and was anxious to breed
more long-haired Siamese, believing they would make a great addition
to the Cat Fancy. Stirling-Webb disagreed and told her that she would
get only short-haired kittens from mating her cat with a Siamese.
Stirling-Webb had seen the
results of Keeler and Cobb's experiments
and described their longhairs as having Siamese type (far less
extreme in type than modern Siamese). He had
felt that Keeler and
Cobb's experiments were the worst possible
thing that could happen to
the Siamese - the Siamese cats' beauty was due to their svelte
outlines, which Stirling-Webb felt would be obscured by a long coat.
Stirling-Webb therefore recommended either having the queen spayed or
mating her to a Black Persian and then mating the offspring together.
The owner was "averse to either course" and
asked to take
the queen to Stirling-Webb for him to see her for
himself .
Stirling-Webb
later wrote:
"When
I saw this queen, I was astonished at her beauty. Apart from her
colouring, she possessed practically no Siamese characteristics, and
was reasonably Persian in type. I was also amazed that nothing was
known of her origin due to the fact that she had been a stray living
in a country churchyard for about six weeks before being adopted by
this lady. The police had been informed of her adoption, and advertisements were placed in the local papers in an attempt to
contact the original owner, but without success." The
stray queen later became known as "Bubastis Georgina".
Unfortunately her
habits meant she was "quite unfit to live in a
civilised house" and in spite of persevering with her, her
adopter offered her to Stirling-Webb as a cattery cat. By then,
Stirling-Webb was enthusiastic about producing colourpoint Persians
(but emphatically against longhaired Siamese!). He was joined by Mrs
SM Harding (Mingchiu cattery) and began to develop the variety
further. He repeated Cobb and Keeler's "formula" and also
acquired a Black Longhair male who had one Siamese grandparent. This
male was mated to the colourpoint queen and they produced kittens
close to his ideal.
Getting
GCCF recognition for Colourpoint Longhairs (Persians) was harder as
he needed to produce 3 generations of Colourpoint Longhairs for it to
gain breed
status and his stock was becoming inbred. In 1955, the
cats achieved breed recognition and to continue outcrossing his
breeding stock. He recommended the use of "bad" Siamese
(round-headed) in Colourpoint Longhair breeding programmes. From the
outset, the variety was considered a colour variety of Longhair
(Persian): Breed Number 13B. Side-effects of his efforts were the
Self Chocolate Longhair (Briarry
Bruno ) and Self Lilac Longhair
(Minghiu Lilac).
In
the 1940s and 1950s, Ben and Ann Borrett bred cattle at their
Chestermere ranch in western Canada. They were also lovers of Siamese
cats and became aware of Stirling-Webb's experimental breeding
programme in England. The Borretts travelled to England to visit
Stirling-Webb and to purchase several of his cats to start their own
breeding
program . Their experience as cattle breeders meant that the
Borretts understood the amount of work required to establish the new
breed and the need for multiple bloodlines. They built up a large
cattery (Chestermere cattery) and developed several different and
totally separate bloodlines. Only the very best cats from each
generation were bred. In 1957, they exhibited two of their imported
cats at an ACFA show in Calgary, Alberta, Canada and were asked to
create the "Himalayan" breed standard (Himalayan is the
name given to the colourpoint pattern in rabbits and goats from the
Himalayan region).
Around
the same time,
California artist and cat breeder Marguerita Goforth
(Goforth cattery) agreed to look after a friend's cat. The cat was a
seal-point Longhair called "Princess Himalayan Hope" and
had come from the San
Diego Humane Society. Goforth gained permission
to breed the cat to create a Persian-type cat with Siamese colouring.
When seeking acceptance for the breed in the USA, she used the
Borretts' breed standard. In the 1960s, Goforth wrote that she
regretted the decision to recognise the Himalayan as a distinct
breed: "I have felt for many years that the Himalayan cat should
be recognized as a colour of Longhair". While the Himalayans
were kept separate from Persians they began to diverge in type,
becoming a long-nosed, colorpoint longhair.
In
the 1950s, the Himalayan was recognised in the same 4 colours as the
Siamese of the time: seal point, chocolate point, blue point, and
lilac point. Red (flame) points and tortie points are recognised in
the USA in 1964. Over the next several years these were joined by
blue-cream point, cream point, lynx (tabby) points and tortie points.
In 1984, the CFA made the Himalayan a division of the Persian breed.
Left: Flame-point (red-point) Himalayan
Right: Seal Point Himalayan
Bottom: Chocolate-point and Seal-point Himalayans
Only
a few Himalayan breeders will ever breed bicolour Himalayans, using
bicolour Persians to introduce the white spotting pattern. Paul
Beall, Richmond, Texas is one such breeder. Paula's photo (left)
shows a bicolour (piebald) Himalayan, but unfortunately this cat has
not produced piebald-point kittens and Paula has decided not to
reintroduce the gene from Persians. At present, Paula is working with
the cinnamon gene in Himalayans; though the gene should have been
present in Siamese cats used to develop the Himalayan, it does not
seem to hve shown up in the Himalayn breed and breeders may have to
introduce it using out-crosses to Persians.
For
a while in the 1950s there was a French breed known as the Khmer. It
resembled a semi-longhair colourpoint and was apparently overtaken by
the Himalayan. "La Vie a Campagne", 1935 dated Khmers back
to the 1920s and the photographs resembled early Colourpoint
Persians. The story goes that a pair was taken to Paris and abandoned
there by a returned serviceman from Indochina. By 1935, this strain
of Khmer had evidently almost petered out. The French Khmer is also
been described as a bootee-less Birman-type cat. The Khmer is
described in an aside by Fernand Mery in 1966 when he describes the
"Colourpoint Longhair" as being "remarkably like a
breed recognised in France, the Khmer, which has its own standard in
that country, though it is not recognised by La Federation
Internationale Feline d'Europe". The term "Khmer" had
actually been dropped in 1955.
COLOURPOINT EXOTIC SHORTHAIRThe
Exotic Shorthair is a shorthaired version of the Persian. It was
developed during the 1950s and recognised in the mid 1960s. The
Colourpoint Exotic Shorthair is an Exotic Shorthair with colourpoint
pattern and blue eyes.
RAGDOLL
AND RAGAMUFFINRagdolls
are medium to large boned, fairly long cats and possess the Siamese
colourpoint gene and the white spotting gene. The have
broad modified
wedge shaped head - best described as a large piece of pie, with the
corners rounded. Their ears are wide set and slightly tilted forward
and the eyes are very blue, but not round in shape.
The
Ragdoll comes in seal point, chocolate point, blue point and lilac
point although red points, cream points and tabby points are now
being bred. The "colourpoints" are exactly that. The
"mitted" versions have the colour points but have white
mittens, white boots and a white blaze on the nose. The "bicoloured"
has additional white on the stomach, chest and ruff. Genetics means
that there are three other patterns: "High Mitted" where
the mitts extend up legs; "Mid-High White" which is a
Bi-colour with additional white in the "saddle" area; and
"High White" which is a Bi-colour with even greater degree
of white, the "saddle" may be absent.
The
colourpoint and white spotting genes interact to produce the
different patterns. The three recognised patterns are colourpoint,
mitted and bicolour. All Ragdolls are colourpointed and some also
have white. Due to the way genes interact, some cats Ragdolls which
look identical actually have different genotypes (different genetic
make-up). For example, High Mitted, Mid High White and "true"
Bicolour can all appear identical, though the exact amount of white
is variable (almost infinitely so!) and some High Mitteds display
more white than a show quality Bicolour while some Mid High Whites
may have so much white on them that they appear to be High Whites! At
times like this, only a knowledge of what genes the cat has inherited
allows breeders to know what colour their cat really is!
High
White is also known as Van pattern or Harlequin in some associations
and is a recognised colour in CFA (America).
Ragdolls
also produce pet quality variants which are not shown (except as
household pets), but which have the same excellent temperament. The
variants
include white toes on colourpoint Ragdolls, dark toes on
mitted Ragdolls, odd white
marks appearing in dark areas and dark
marks appearing in light areas! The white spotting is hard to get
perfectly right!
The
Ragdoll originated in California in the early 1960s and is surrounded
by myth thanks to their somewhat eccentric breeder, Ann Baker. Ann
Baker bred Persian cats.
Josephine was a white Persian-Angora cat
belonging to Baker's
neighbour ; she was a semi-feral who produced
kittens as wild as
herself . After Josephine was hit by a car she had
to stay indoors while recovering and she became somewhat tamer.
While
Josephine's earlier litters had been
half wild like their mother, the
litters born after the
road accident were very relaxed, docile and
social. The exact reason why is not known and later became the
subject of wild claims. Since kittens inherit much of their
personality from their sire, the most likely explanation is that she
was breeding with different males, ones carrying genes for more
sociable offspring. Judging by the kittens' appearance, Josephine's
suitors were most likely Birman and Burmese. In addition, Josephine
was less wild and perhaps did not
train her kittens to be quite so
fearful of humans.
Baker
decided to create a new breed, one which retained the very sociable,
relaxed traits, non-matting fur, large size and the property of going
limp when handled. She acquired a black Burmese-looking female called
Buckwheat and a bicolour female called Fugianna. Fugianna's sire was
another of Josephine's kittens, a seal-point Birman-type cat called
Daddy Warbucks. There were to be no more
founding cats since
Josephine and her next litter were destroyed by their owner. Based on
the appearance of the two founding females, Baker split their progeny
into two groups: Fugianna's kittens were termed "the Light
Side"; Buckwheat's kittens were termed "the Dark Side".
Ann
Baker formulated a strict breeding policy to preserve the purity of
the Ragdoll and charged other breeders (franchisees) a royalty fee
for every kitten they
sold . She also made wild claims about the cats:
their docility was due to changes in Josephine after she was hit by a
car (a long-discredited Lamarckian theory), that while Josephine was
being treated for injuries she was infused experimentally with skunk
and/or racoon and/or human genes (even if
done , these genes would not
have entered the egg cells in the ovaries), that Ragdolls were immune
to pain (they have normal pain thresholds but are remarkably tolerant
cats) and even that they are a link between us and
space aliens! A
more plausible
suggestion was that Josephine had a mutation in her
ovaries and this mutation was present in the egg cells (a germ-line
mutation) and was passed on to her offspring.
Her
claims and methods alienated other breeders, some of whom broke away
to develop the Ragdoll into a registrable breed using
sound breeding
practices. Depending on which history you read, the Ragdoll was
recognised as a purebred cat in 1965, 1966 or 1967. Also according to
many sources, Denny Dayton achieved US recognition for the Ragdoll in
1967, however he didn't buy his first Ragdolls until November 1969.
In 1971 Baker created the International Ragdoll Cat Association
(
IRCA ) and claimed that non-IRCA cats to be fakes, frauds,
look-alike, half-bred, not authentic etc while promoting her cats as
the only legitimate Ragdolls. In 1975 Baker patented the Ragdoll
name; she died in 1997 but her
patent remained valid until 2005 and
allowed only IRCA breeders to use the name "Ragdoll".
Earlier breakaway breeders felt that the restriction did not apply to
them because they had purchased and bred their cats
prior to the time
of the patent. In 1981, Ragdolls were exported overseas for the first
time.
The
stories of the two types of Ragdoll ran
parallel for many years.
Baker's trademarked umbrella term for all of her IRCA varieties was
Cherubim Cat meaning "Angels non-fighting cat". In 1985 and
1987, one UK breeder also advertised "Cherubinis" which
appear to have been non-colourpointed offspring from outcrossing
Ragdolls to other breeds. "Cherubini" was a legally safe
equivalent of IRCA's trademarked "Cherubim" name. The IRCA
Ragdolls were not recognised by any other registry and
upon Baker's
death, a number of breeders re-registered their
Miracle Ragdolls,
Honeybears and other IRCA varieties as foundation stock for the new
RagaMuffin breed which is now recognised by many registries around
the world. The RagaMuffin is not merely a non-pointed Ragdoll, it
also has a different conformation from the Ragdoll. After Baker's
death, some breeders tried to sell their IRCA Ragdolls as "rare
mink-pointed Ragdolls" or "rare solid colour Ragdolls"
to unwary owners/breeders who did not know that there had been two
conflicting Ragdoll breeds. These "rare" colour cats,
though attractive, were not registrable as Ragdolls in any other cat
association.
Throughout
her life, the eccentric Baker defended her breeds and claimed that
only IRCA Ragdolls were the real article and guaranteed free from
genetic defect, while other Ragdolls were half-bred or cross-bred
inferior lookalikes trading fraudulently on the Ragdoll name.
Quarter-page advertisements to this effect appeared in an American
cat magazine in 1992. Periodically, she would send threatening mail
to breeders non-IRCA Ragdolls, including a mailshot that contained
photos of dozens of dead cats
laid out side-by-side along with a
claim that someone had broken in and
killed her cats (some believe
Baker killed her own cats and might not have known what she was doing
due to a claimed history of head
trauma ).
The
RagaMuffin is another descendent of the original Ragdolls, but in a
more extended colour range. Their similarity to the accepted Ragdoll
meant that many registries would not
accept them, though the
conformation is not the same as the TICA Ragdoll (note: the NZCF
accepts "Solid Ragdolls instead). In the days when Ann Baker and
IRCA were
active , a there were a number of other "breeds"
or named breeding lines. IRCA's other named breeds/lines were the
Honeybear/Honey Bear (resembled old style Persians), Ragdoll Hobby
Cat, Miracle Ragdoll ("a highly upgraded Ragdoll" formerly
known as the "Ragdoll Tu", "experimental Persian"
or simply "Miracle"), Baby Dolls/
Doll Babies, Angels, Shu
Schoo, Symoneese/Symonees, Manxees and Fuzz. The named IRCA
Ragdoll/Miracle breeding lines included Maximillions (silver
tabbies), Minks, Bears (thicker fur, shorter ears, rounder eyes,
flatter face, cobbier body) and Catenoids (blue-eyed solids
surrounded by claims they could be bred to any cat to produce an
IRCA-type Ragdoll!). After the demise of Baker and of IRCA, many of
these were absorbed into the RagaMuffin and some were apparently
re-registered as TICA Ragdolls.
SNOWSHOE,
SNOWSHOE SIAMESE AND SNOW -TOESOriginally,
the Snowshoe (or Silver Laces Cat) resulted from crossing Siamese, or
Oriental Shorthairs, with American Shorthairs. The white mitts occur
natually in some lines of Siamese where they are considered a fault.
Today Snowshoes generally result from Snowshoe to Snowshoe breedings
although they may still be outcrossed to one of the founding breeds
in order to expand their gene pool, introduce new colours, or improve
type or colour/pattern. Officially the breed claims no Birman
ancestry despite the superficial resemblance in pattern.
Snowshoes
were developed in the USA by Dorothy Hinds in the late 1960s when her
two Siamese cats produced kittens each with four white feet. White
spotting sometimes showed up in early Siamese cats (white toes are
occasionally still found). She dubbed them Snowshoes. The variant did
not recur so she crossed a Siamese with a bicolour American
Shorthair. In addition to the white mitts, this eventually produced
the inverted white 'V' on the face.
The
Snowshoe was recognised by various American registries in 1974,
however in 1977 there was only one registered breeder of Snowshoes.
Luckily, interest in the breed picked up and the Snowshoe has become
a popular breed in the USA.
It
has Siamese style points and blue eyes combined with white paws and
white spotting and comes in blue, seal, lilac and chocolate points.
The Snowshoe pattern is divided into two categories: Bicolour
Snowshoes where up to two-thirds of the cat can be white and Mitted
Snowshoes where the white
ranges from just the toes to halfway up the
leg. Snowshoes have a white inverted 'V' extending from between the
eyes down over the nose.
Early
on the breed was erroneously thought to be a Birman Shorthair, based
purely on its appearance. The Birman Shorthair (Templecat) only came
into existence in 1995 in New Zealand.
The
Snowshoe conformation is intermediate between the parent breeds and
is sometimes described as close to the older (or traditional) and
more moderate style of Siamese cat although it is sometimes described
as resembling the American Shorthair. The Snowshoe inherits the
muscular body from the American Shorthair and the body length from
the Siamese/Oriental. The conformation is, to some extent, affected
by the outcross breeds permitted in countries where the American
Shorthair is not found.
In
the 21st
Century , an experimental breed called the Snowshoe Siamese
has also been announced. Its body type is more extreme and is closer
to the modern Siamese, while the original Snowshoe maintains the more
moderate body type. Some Siamese cats have appeared with white toes
and this was considered a
serious fault in the breed.
The
Snow-Toes was bred in the late 1960s by crossing Birmans to
Himalayans, seemingly with the aim of a mitted Himalayan. Whether
this will see a revival alongside the Snowshoe Siamese has yet to be
seen!
NEVA
MASQUERADEIn
1990, the first Siberians were imported into the USA. However the
colourpointed variety with blue eyes was not imported and colourpoint
Siberians were not recognised by registries on in American breed
standards. However Russian breeders have recognised the colourpointed
variety since they began to keep records (in around 1987). According
to anecdotes, colourpointed feral cats bred with feral Siberian cats
along the banks of the Neva river at St Petersburg (then Stalingrad).
The name Neva Masquerade reflects the origins of this masked cat.
The
first Siberian cats were imported into the USA in 1990 by
Elizabeth Terrell of Louisiana, in exchange for Himalayan cats sent to
establish that breed in Russia. The first colourpoint Siberians were
imported into the USA in 1997 and registries are have recently
accepted it following early controversy.
TONKINESEThe
Tonkinese is intermediate in type between Burmese and Siamese. Its
pointed pattern is less pronounced than that of the Siamese and its
shape is less extreme. Long known as a variety in Thailand, it has
existed in the
West for at least a
hundred years. Towards the end of
the 1800s, a cat described as a chocolate variety of Royal Siamese
was exported from Singapore to England. It was a rich chocolate
colour with a darker face, ears and tail; and its legs were darker
than the body. It was not, however, recognised as anything other than
a brown Siamese and interest in them had waned because it had been
hard to compete against the striking coat and eye colour of the
Siamese. These cats were either Burmese or Tonkinese, or most likely
a mix of both types since some, when bred to the royal Siamese,
produced offspring with "poor" markings (i.e. "mink")
while others evidently produced offspring had royal Siamese markings.
In 1903, the "Burmese Cat" (shown here) would be, in modern
terms, an Oriental Ticked Tabby!
The
Tonkinese is considered a
hybrid of Siamese and Burmese and this has
hindered its acceptance in some countries e.g. Australia because "as
a hybrid it cannot breed true". The alternative standpoint is
that it is that Tonkinese is the naturally occurring intermediate
form of a colour range where the two extremes are represented by
Siamese and Burmese. The two extreme forms have been selectively
bred, but the intermediate form was ignored until relatively
recently. Tonkinese type cats have been described for many years: the
Chocolate Siamese (Britain, 1880s; generally considered to be
Burmese), Golden Siamese (USA, 1950s, Siamese/Burmese hybrids),Golden
Chechongs, Si-Burms and Zibelines (the French name for Burmese).
These all had tan to brown bodies with seal or almost black masks and
points.
The
self-coloured Chocolate Siamese (as opposed to the Chocolate Point
Siamese) was described by Harrison Weir in 1889 and by Frances
Simpson in1903, however the "Royal Siamese" with its
greater contrast between body and points was the form that
predominated in Britain and the Chocolate Siamese was lost. The
Chocolate Siamese is variously claimed to be an early Burmese or an
early Havana Brown type cat. The founding female of the Burmese
breed, Wong Mau, was a walnut-brown Tonkinese type cat imported into
the USA from Rangoon in 1930. She was described as brown with darker
points on her face, legs, feet and tail. When bred to a seal-point
Siamese, she produced kittens identical to herself and some with a
Siamese coat pattern. Over several generations and through mating her
to her own
sons , three varieties of offspring arose: those with a
Siamese pattern, those which were dark brown all over (Burmese) and
those with a dark brown body but identifiable darkening at the points
much like Wong Mau herself. Wong Mau is considered to be the founding
mother of the Burmese breed but was herself a Tonkinese and breeders
chose to develop her Burmese offspring.
In
the early 1940s, a Hawaiian breeder, Lelia Volk, bred a dark variety
of Siamese: its colour was lighter than the Burmese, but darker than
the Siamese. In a report in the Journal of Heredity, the colour was
said to be close enough to the Siamese that it was unlikely to be
perpetuated. Between the 1950s and early 1960s, Milan Greer in New
York City bred Golden Siamese. These were Siamese/Burmese hybrids
which had a rich mahogany body and dark points. Greer had been told
that such cats were normally considered defective and destroyed
because they did not breed true. Using Seal Point Siamese and Sable
Burmese and cross-breeding their offspring, he produced chocolate
brown cats with darker points and claimed to have bred pure "Golden
Siamese" for 5 generations. Having
proven his point, he stopped
breeding, though his cats were popular with pet buyers.
In
the 1960s and 1970s, the Tonkinese was revived in Canada and the USA
by crossing Siamese with Burmese in spite of opposition from breeders
of both Siamese and Burmese who viewed them as embodying everything
they were trying to breed out of those two breeds. In the mid 1960s,
Jane Barletta of New Jersey
switched from the increasingly extreme
Siamese to breeding the more moderate Tonkinese. At about the same
time,
Margaret Conroy in Canada bred a female Burmese to a Seal point
Siamese as she was unable to find a suitable Burmese stud in her
locality. Her tan-bodies cats bred true for 5 generations and were
dubbed Tonkanese (with an "a"). The original cats were
Sable (US: Natural Mink), but crosses between Blue Burmese and Blue
Point Siamese introduced Blue Tonkinese. Later on, the other
Siamese/Burmese colours were also bred together to expand the
Tonkinese colour range: red, lilac (US: platinum), chocolate (US:
champagne) and cinnamon (US: honey).
Though
the Tonkinese breed in the USA has been recreated by crossing the
Siamese and the Burmese, the variety has occurred naturally in south
east Asia for hundreds of years. Rather than being a hybrid, it is a
naturally occurring breed and the Burmese, Tonkinese and Siamese
represent the two extremes and the mid-point of an overall genetic
pattern caused by 2 mutant genes. Recognition in various cat fancying
countries came from the 1980s onward. In the UK, it was recognised in
1990. In Australia it was grudgingly accepted in the 1990s despite
being condemned as a hybrid and unable to breed true.
The
colours of Tonkinese are (the term "mink" is not used by
some registries) blue mink, champagne mink (light chocolate), honey
mink (chocolate/chestnut), natural mink (seal), platinum mink
(lavender/lilac), red mink, cinnamon mink, fawn mink, cream mink. In
some registries Burmese are accepted in tortie patterns and this will
be reflected in the Tonkinese. Its genetic make-up means that
Tonkinese-to-Tonkinese breedings will always produce a mix of
patterns: self (Burmese-type) variants, Tonkinese and colourpointed
variants. The colourpointed Tonkinese find favour with those who
seek a less extreme version of the Siamese. Although most registries don't
accept the variants for exhibition, the variants are an
important part of the breeding program; bred together they produce
Tonkinese-pattern cats.
COLOURPOINT
SHORTHAIR (COLOURPOINT BRITISH SHORTHAIR) The
Colourpoint British Shorthair is exactly what the name suggests - a
British Shorthair in a colourpoint coat with blue eyes. It was
developed in the 1980s by crossing the British with the Himalayan.
OPAL
(COLORPOINT AMERICAN SHORTHAIR)In
the 1990s, breeders of appleheaded (roundheaded) Siamese were invited
to
register their cats as Opals. Little has been heard of the Opal
since then although the traditional style of Siamese goes from
strength to strength (despite the registry's rather bizarre refusal
to allow its actual breed name to be used in
articles or
reviews in
case it erodes, rather than promotes, the breed). The Opal appears to
be a Colourpoint American Shorthair.
THAI
POINTED AND THAI SIAMESEThe
Thai Pointed is a naturally occurring colorpointed (Blue-point,
Lilac-point) Korat variant. Korats are related to Siamese and have
interbred naturally in their native Thailand.
The
Thai Siamese is a European term for the traditional style of Siamese
derived purely from non-ultra-typed Siamese cats.
The
term Thai Siamese has also been used for colorpointed cats derived
from naturally occurring colorpointed variants in Korat litters, but
this usage is not encouraged.
Andrea
and Beata Brunetti, in Italy, have a Thai (the European name for the
less extreme, traditional or old style of Siamese cat) called Urban
Hunter with a visible blotched tabby pattern and an interesting
dorsal stripe. Urban Hunter has been mated to a red-point Thai called
Kasia Ruda, resulting in one male and four female kittens. All of
these have dorsal stripes similar to Hunter.
In
some lines of old type Siamese, there is less contrast between body
and points. The Siamese pattern is caused by an albinism gene, but is
partly dependent on temperature, with a darker colour
developing in
cooler regions. This is unlikely as Urban Hunter lives in a climate
where the temperature ranges between 25 and 30 celsius.
Polygenes
can also help to allow or suppress the colour. Variable contrast
between body and points is seen more often in the older style
Siamese, probably due to greater genetic diversity compared to the
modern extreme-type cats. It is also possible that a novel mutation
has occurred as Urban Hunter has
aqua , rather than Siamese blue,
eyes. Because of the aqua-blue eyes, I initially
wondered if Urban
Hunter had Tonkinese in his ancestry (Tonkinese is intermediate
between Siamese and Burmese in colourpoint). Genetic testing and
test-mating would be required to determine whether Hunter’s
colouring is due to a mutation of the colourpointing gene.
AUSTRALIAN CLOUD The Cloud is an Australian shorthair/semi-longhair bred in mink colors. This does not appear to be a recognised breed, but is a cross-breed from Birmans, Ragdolls and British Shorthairs.
Some photos depict a Ragdoll-like cat, others depict a cat similar to the traditional style of Siamese or a colourpointed British Shorthair (as seen here). The accompanying photo is provided by an owner of a cat bought as a "Cloud".
KUCING
MALAYSIA/PIAWAIAN KUCINGKucing is Malaysian for "cat". The Kucing Malaysia is similar in type to the Tonkinese with a colourpointed pattern similar to Ragdoll and blue/blue-
green eyes. A White blaze on face and muzzle is desirable.
FRENCH
KHMERA French breed known as the Khmer was described by Mery in 1966. He noted the similarity of emerging colourpoint Persians (Himalayans) to the existing French Khmer, although the photographs of the Khmer suggested a semi-longhair or even a a
fluffy , cobby colourpoint shorthair. Although Mery described it in 1966 and noted that it had a breed standard albeit one not recognised by FIFe, the term Khmer had been dropped back in 1955 when Colourpoint Longhairs and Himalayans were being developed in America/Canada and Britain respectively. There are earlier descriptions of the Khmer dating from 1935 (by which time it was close to being lost) and photos from that time depict a longhaired cat similar to an early colourpoint Persian. "La Vie a Campagne" in the late 1920s showed a longer haired Khmer, so possibly the coat length was seasonal. A pair of Khmer cats were supposedly taken to Paris from Indochina by a French serviceman - a tale very similar to that of the Birman. Two Birman were reputedly obtained by Major Gordon Russell from Burmese priests whom he had helped escape to Tibet; when he went to live in France in 1919 he took his Birman cats with him. This suggests that the French Khmer was a Birman that lacked the characteristic white markings of the Birman. This corresponds to the use of the term Khmer to describe bootee-less Birmans - and at one time there were indeed unsuccessful attempts to create"Himalayans" from Birman stock.
MASKED
SILVERA
variety described by Mery and others in the 1960s, this is not
related to Siamese cats but was considered a form of shaded silver
(agouti + silver) longhair with a black mask on its face. It is a
poorly marked black smoke. Similar cats appear in black smoke
(non-agouti + silver) individuals of
curly -haired breeds due to the
black colour being more intense on the straighter fur of the face and
legs.
Martine
Sansoucy of Butterpaws LaPerms has seen a number of masked silvers
and
notes that they appear to be smokes rather than shaded silvers.
Butterpaws BC The Crow, known as "Cairo" is a black smoke
that meets the general description of a masked silver. As can be seen
from the photo, the different textures of fur on the face and the
body give the impression of a masked cat.
Historical Masked Silver.
Masked Silver (black smoke) LaPerm.
Historical Masked Silver.
Some
authors have written that the masked silver dates back to 1900, but
may have been referring to shaded silvers, a variety not recognised
in Britain at the time and considered to be a badly bred chinchilla
or poor quality smoke. Chinchilla and shaded silver are genetically
tabby cats, smokes are genetical solid colour cats; this difference
was not understtod in the early 1900s. According to Milo Denlinger in
1947, "Masked silvers are a new variety and very few are bred."
Denlinger went on to describe the variety: "The ideal masked
silver is a very beautiful animal; in colouring or, I should say,
marking, they should resemble the Siamese Cat; that is to say, they
should have a black mask, or face, black feet, and legs. The body
should be as pale a silver as posible." The eyes were to be deep
golden or
copper . Several authors have observed that the description
of the masked silver resembles that of the Siamese.
OTHER
COLOURPOINTSIn
addition to the recognised breeds, the colourpoint pattern is
recognised as a permissible colour/pattern in many other breeds. In
most cases the cat is simply described as a "colourpoint
'breed-name' ". The colourpoint Japanese Bobtail has only
recently found its way into the West although it has existed for
decades in
Japan and a similar breed known as the
Mekong Bobtail
(formerly the Thai Bobtail) is known in Russia. Mekong Bobtails also
occur naturally among feral cats in Malaysia and Thailand.
In
others the
prefix "Si" is used e.g. the Si-Rex is a
colourpointed Rex and is further identified as Cornish Si-Rex or
Devon Si-Rex. It exists unofficially or experimentally in a few
breeds e.g. Si-Manx (Manxamese). In the American "Cats Magazine"
of June 1965 there was a photo of two Blue-Point rex-coated kittens
born to a blue self (Cornish) Rex in August 1964. Their pedigree
showed that four generations back, there had been a Blue Point
Siamese outcross in the breeding program. In addition, one ancestor
was an
Oregon Rex mutation owned by Mrs Mildred Stringham of
Warrenton, Oregon. A Si-Rex was being developed in England at the
same time, by Mrs Ashford (Annelida prefix).
At
a Feline Fantasy staged by the Silvergate Cat Club of San Diego, USA.
in 1952, a "Sianx" cat was exhibited. This was a
Siamese-Manx cross with Seal Point colour and type, sans tail.
However, the Western
province Cat Club of Cape Town claimed
precedence in Sia-Manx breeding. The late Father Fowler's Sia-Manx
was exhibited at their show during 1952, some months before the cat
at the San Diego show, and resembled a Siamese Seal Point with
definitely high hind quarters and complete absence of tail. He was
described as a very charming person by Miss P. Ashby-Spilhaus, the
Registrar of the South African Cat Union.
The
colourpoint trait can be introduced into any breed by crossing it
with an existing colourpoint breed, although the first generation
kittens will not show the trait. The gene for colourpoint has also
entered the gene pool of free-breeding cats and there are many
pretty "look-alikes" to be found in shelters and humane societies.
Although not registrable as Siamese, Himalayan etc, they are still
good pets and can be shown in household pet classes.
An unexpected colourpoint (tabby point) kitten born among a litter of black-and-white bicolours. The mother was a non-pedigree and the father was suspected of being a Birman cat that lived in the area.
Below
are some non-pedigree colourpoints. Capi ("Paws") is a male
Siamese mix who resembles a Snowshoe. He was photographed by Koraljka
Polack on Cherso island (north Adriatic sea) in 2007. There are
several "Siamese mongrels" in the area due to a Siamese cat
that lived in the area some years ago. This bred with stray and feral
cats. The third photo is a random-bred male colourpoint.
COLOUR
DILUTION IN CATS: BLUE DILUTION, PINK-EYED DILUTION, CHEDIAK-HIGASHI
SYNDROME DILUTIONInformation
collated 2015, Sarah HartwellBroadly
speaking, dilution refers to making a colour lighter – like mixing
a coloured paint with white paint.
COMPARISON STUDIESThe
colour of cat fur is due to the size, shape and distribution of the
melanin (black pigment) or phaeomelanin (red pigment) granules in the
hair. The hair has a central shaft (cortex) and
outer sheath
(
medulla ). Melanin granules may be distributed differently in cortex
and medulla. Granules in the medulla can be a different size and
orientation to those in the cortex. It is the combination of
location, size, shape and density of melanin granules that gives
different colours.
The
Structure Of A Hair Shaft
Simplified
Diagram Of Pigment Granules In Cat Hair - based on the microscopy
examination photos
Prieur
& Collier (1981) compared the melanin granules in hair of black,
smoke, blue, Chediak-Higashi-smoke, and pink-eyed dilution cats. A
portion of the preserved skin of the pink-eyed dilute cat was used in
this study.
- Black cat hair (non-agouti) contains numerous small dark brown to black melanin granules uniformly distributed throughout all portions from base to tip of the hair.
- Smoke cat hair (non-agouti + inhibitor (silver) gene) has a scarcity of melanin granules at the base of the hair.
- Blue cat hair (non-agouti + dilution) has larger basic melanin granules, but still relatively regularly shaped, and a non-uniform distribution of granules.
- Chediak-Higashi Smoke cat hair trait has even larger melanin granules than the blue dilution, and they are relatively irregularly shaped. All of the cats were Persian and resembled blue smoke Persians.
- The pink-eyed dilute cat hair had very small yellowish-brown melanin granules and very few larger granules. Under the microscope, some lighter banding could be seen on the hairs, but the tip was coloured. The base of the hair was paler than the tip (agouti banding).
-
Prieur DJ, Collier LL. Morphologic
basis of inherited coat-
color dilutions of cats. J Hered. 1981 May-Jun;72(3):178-82.
BLUE
DILUTIONThe
normal dilution seen in cats is "blue dilution" that
changes black to grey (blue), chocolate to lilac, and cinnamon to
fawn. Blue dilution affects the skin and coat colour by altering the
pigmentation granules, but leaves the eyes pigmented.
How
Blue Dilution Affects Different Colours
Prieur
DJ, Collier LL. Maltese dilution of domestic cats. A generalized
cutaneous albinism lacking ocular involvement. J Hered. 1984
Jan-Feb;75(1):41-4. "The Maltese dilution is an autosomal
recessive trait of cats that dilutes black cats to blue, and orange
cats to cream. The pigmented cutaneous and ocular tissues of Maltese
dilution and control cats were examined and compared by light
microscopy. Most of the melanin granules in all of the pigmented
cutaneous tissues of the Maltese dilution cats were aggregated
together into large clumps. However, none of the intraocular tissues
containing melanin producing cells of either neural crest or optic
cup embryologic origin contained clumped melanin granules. It is
concluded that the Maltese dilution trait is a
unique generalized
albinism without ocular involvement."
PINK
EYED DILUTIONPink-eyed
dilution is found in a number of mammals but there is only one good
account of it in cats. It changed black to a fawn/bluish-tan colour
and the eyes had a red pupil and gold flecked red/pink irides (the
coloured
section around the pupil). There may have been occasions
where pink-eyed dilution has been confused with pink-eyed albino
which is reported sporadically in cats.
Analysis
of hairs from the preserved skin of the female pink-eyed dilute
showed very small yellowish-brown melanin granules and very few
larger granules. The base of the hair was paler than the tip and some
hairs had lighter banding (agouti banding) that was only visible
under magnification. The tip of the hair was coloured.
For
those with a scientific background, I’ve included details from the
two known studies of the single confirmed cat with this mutation.
Pink-Eyed
Cats /Albino SeriesTo
untangle the different pink-eyed mutations, here is a short
description of the albino series in cats showing where pink-eyed
albino fits in. It is the most recessive of the albino alleles (gene
variants). A pink-eyed albino has an unpigmented white coat. A
pink-eyed dilute has a coloured coat (right now we don’t know how
pink-eyed dilution interacts with dominant white (white masking)).
- C/C, C/cb, C/cs, C/ca, C/c - full colour
- cb/cb, cb/ca - Burmese colour restriction (temperature- sensitive )
- cb/cs - Tonkinese Colour restriction (temperature-sensitive)
- cs/cs, cs/ca, cs/c - Siamese Colour restriction (temperature-sensitive)
- ca/ca - Blue-eyed Albino (not temperature-sensitive)
- c/c - Pink-eyed Albino (not temperature-sensitive)
Temperature-sensitive
albinism in cats is caused by tyrosinase (TYR) mutations. For the
molecular genetically minded, it is a cytosine deletion in the TYR
gene. Cat fanciers refer to it as the "C" locus (a sort of
genetic "address"). Albino cats have been reported in
scientific literature (Todd 1951; Turner et al. 1981) but their
eye-colour has not always been well described. Blue-eyed albinos have
not been properly distinguished from pink-eyed albino cats (Bamber &
Herdman 1931; Todd 1951; Leventhal 1982; Leventhal et al. 1985). This
made it unclear as to whether there were more than one "one non
temperature-sensitive albinism alleles" in cats i.e. both
blue-eyed and pink-eyed. Lyons believed the single report of a
red-eyed albino cat to be erroneous (Imes et al. 2005), but since
2005 there have been more reports and photographs of pink-eyed albino
cats (this mutation has entered the Bengal breed via the Asian
Leopard Cat). Because of the cat's eye structure, pink-eyed albinos
often appear lilac-eyed with a red pupil, however the coat is always
white. Blue-eyed albinos have pale blue irides and must not be
confused with "blue-eyed white" cats that have the white
masking gene and vivid blue eyes.
How
is this relevant? A test-mating of a female pink-eyed dilute to a
colourpointed cat demonstrated that the pink-eyed dilution is
recessive, but it's not on the "C" locus and therefore
isn't related to the known albino series. This makes it interesting
to cat fanciers as a potential new variety, providing it doesn't have
any damaging side-effects. This was documented by Neil B Todd in
1961:
A
Pink-Eyed Dilution in the Cat - Neil B Todd
September
1961, 52 (5) - Journal of HeredityIt
was the author 's original intention to report in detail on the
genetic basis, if any, for the cat to be described below. However,
her untimely death due to an intestinal obstruction ended the work
almost before it was started.During
the summer of 1958 three kittens from a litter were turned over to
the SPCA in Springfield, Massachusetts. Their estimated date of birth
was April 1958. Two of these animals were apparently quite ordinary,
although not detailed description of them was available. The mother
was presumably normal; the father was unknown. The third kitten, a
female, had pink eyes (although a small amount of yellow pigment was
present in the iris ) and a light tan coat. She was a blotched tabby
(dominant) with moderate white spotting (dominant) and had a slight
kink at the tip of her tail. Her hair was short and dense with a fine
texture. In all other respects she conformed to the wild type. It is
suspected that she had some visual impairment from the rather clumsy
way she would run into things and from her hesitancy to jump from a
height which would not disturb most cats.The
animal apparently never came into heat spontaneously during the first
three years of her life, but this may have been due to her caged
existence rather than to any inherited or congenital defect. She was
treated with hormones in January, 1961, and subsequently mated to a
chocolate point Siamese. This cross was made with the purpose of
discovering if any relationship existed between her phenotype and the
albino series of alleles. A chocolate point Siamese is homozygous
recessive for a pigment-determining allele at the albino locus
(cbcb),
for brown (bb) and for non-tabby (aa). [Note "+" meant
"wild type" in the genetic nottion of tht time.]The
cross produced a litter of three kittens, born 10 days prematurely
with the result that none survived. However, all were fully pigmented
tabbies and therefore of the genotype +acb+bb+aa.
From the data it can be concluded that the phenotype of the mother
was due (if indeed genetically determined ) to the homozygous
recessive condition at some locus other than that of the albino
series of the black-brown alternatives.[Modern
notation for the offspring: Aa Bb Ccs Dd oo T-
(I’ve used “T” on its own because the report doesn’t say if
the tabby is classic (Tc) or mackerel ( TM )]Neil
B. Todd
The Biological Laboratories, Harvard University And
that seemed to be the end of it until 2015 when Kathryn
Eden posted
photos of Milkdud, a Donskoy (Don Sphynx kitten) whose colour defied
analysis. Born to black parents, Milkdud’s genetic test results
were:
- Agouti Result: a/a - Non-agouti (solid colour)
- Albino Result: N/N - No copies of albino allele are present.
- Amber Result: E/E - No copies of the mutation for Amber.
- Brown Result: B/B - Full color black, does not carry brown or cinnamon.
- Colourpoint Result: C/C - Full colour, cat does not carry Burmese or Siamese alleles.
- Dilute Result: D/D - Full (Dense) color. Cat does not have the dilute allele.
Milkdud's
skin was closer to a fawn colour and his eyes had yellow irides with
red pupils. There is no genetic test for pink-eyed dilution. Photos
exist of the original pink-eyed dilutes hairs under high
magnification, so that is one route to investigate (the Don Sphynx
has slight furring on the tail). The other is to breed furred cats
with the same colour and eye colout and to analyse their fur. While
the genetic basis isn't yet known, visually Milkdud appears to be a
pink-eyed dilute and hence a new
opportunity for the cat fancy.
Anna
Kholmska, in Ukraine, has bred 4 generations of these cats (over 7
years) to establish a breeding line. All have been healthy. The
unidentified colour is known in discussion (during 2015) as "Ukraine
chocolate." From the visual appearance, it closely resembles the
description of pink-eyed dilution. What is exciting, is the
possibility of a lost phenotype reappearing in cats. The colour also
needs to be bred into furred cats for study. Anna notes that the
chocolate colour (b at the TYRP1 (B) locus) is not accepted in the
Donskoy, although it can be very attractive. For more than seven
years, Donskoy Discovery has bred show-quality cats, including some
that resembled chocolate colour. Unlike Chocolate Orientals, the
Donskoys had an unusual eye colour that appeared pink in some
lighting conditions. This was initially attributed to a combination
of the chocolate gene with the dominant hairless gene, but the work
of Kathryn Eden and Anna Kholmska and genetic testing has shown that
the chocolate allele is absent. The cats need to be tested to confirm
the presence of the dominant B allele (black). The coat and eye
colour could be caused by a novel dilution of black (if B is present)
or by some other mutation (if B is absent). All of the cats share a
common ancestor. Their genotype (genetic makeup) shows they should be
solid black but their phenotype (visual appearance) is chocolate.
This indicates a diluted-colour but the cats do not have the dilution
gene (d/d) at the D locus. In any case, “d” causes blue dilution
i.e. black to grey. “d” is one of the three genes involved in
melanosome transfer and translocation. The unexplained “Ukraine
chocolate” seems due to a gene for which there is no test (at least
in the cat). Possible candidates are the other two melanosome
transfer and translocation genes: Myosin Va (MYO5A) and RAB27A. Genes
involved in the synthesis of eumelanin other than TYRP1, for example,
TYRP2, are also possibilities. The other known dilutions in the cat
are the extinct Barrington Brown cats (none of which were bred
outside of a laboratory colony) and the single example of a pink-eyed
dilution cat.
UNDETERMINED
DILUTIONThere
are 4 cats of Thai descent whose dilution factor hasn’t been
genetically identified.
The
first of these is Mai Thong. In January 2015, I received information
and photos of a native Thai cat that TIMBA hoped to use in the
Suphalak breeding programme. Nolan Betterley lives in Thailand and is
working on the breeding project to resurrect some of the rare cats
that are described in the ancient cat
poem manuscripts. Breeders
working on the Suphalak, which is a solid (i.e. no Burmese pointing)
chocolate self, have searched all over Thailand to find cats suitable
to be used in the breeding program. They found cat, Mai Thong, aged
approx 7 months, on the
street . Nolan hoped that she was either
cinnamon or lilac (these are recessives). DNA was tested at UC
Davis and came back as aa BB CC DD (Non-Agouti, Black, No Colourpoints, No
Dilution), which should produce a black cat. However Mai Thong is
very obviously not black, but has an orangey colour. The gene
candidates are a non-extension gene (since Russet Burmese exist) or a
different form of dilution.
The
remaining 3 cats that failed the test for the Suphalak breeding
programme were also street cats; two were brown and one was tortie.
They are now in the USA with two different breeders. The plan was to
mate them and test offspring, but this has not happened. Doug Schar
has the tortie sister. The owner of the other 2 cats has not allowed
access to a male sibling for test-mating. UC Davis found no mutation
of the TYR gene for these cats. Unfortunately, no other labs appear
interested in following up to see what mutation is present..
CHEDIAK-HIGASHI
SYNDROME (CHS) DILUTION (CHEDIAK-HIGASHI SMOKE)This
has only been found in Persians and manifested as blue smoke fur with
yellow to light-green eyes (normal blue smoke Persians have darker
fur and orange/copper eyes). Their eyes reflected red and tended to
be sensitive to light. The cats developed cataracts as early as 3
months old. Melanin pigment granules in all tissues, not just the
fur, were abnormally in size and shape due to combining with
abnormally large lysosomes (part of a cell's digestion mechanism).
CHS cats tend to develop kidney lesions, and have impaired
vision due
to the abnormal pigmentation in the retina. The University of Sydney,
Australia's Faculty of Veterinary Science defines CHS as a "rare,
lethal
disorder " but it better meets the criteria for an
"impairing mutation" as its effects can be mitigated with
proper care. Affected cats are highly susceptible to infection, due
to the abnormally large lysosome in their white blood cells.
Bacterial infections occur repeatedly because the white blood cells
cannot
respond normally to infection; antibiotic treatment is
required to combat infection. Prolonged bleeding tends to occur
because of abnormal blood platelets delaying clotting. In the
laboratory, CHS cats had a longer period of eye-flicker, compared to
non-CHS Persians, after being spun around.
A
Blue Smoke Persian and a Chediak-Higashi Smoke Persian
Despite
the many side effects, cats given appropriate care can have a good
quality of life. Because it affects health, the trait should not be
deliberately propagated as a colour curiosity. For those with a
scientific background, here are several references and abstracts
about the condition.
Collier
LL, King EJ, Prieur DJ. Aberrant melanosome development in the
retinal pigmented epithelium of cats with Chediak-Higashi syndrome.
Exp Eye Res. 1985 Sep;41(3):305-11. "Chediak-Higashi syndrome is
a genetic disorder characterized by greatly enlarged cytoplasmic
granules, including lysosomes and melanosomes. Eyes of humans and
animals with Chediak-Higashi syndrome are hypopigmented to various
degrees. Intraocular melanin granules vary in size, with some being
massively enlarged. Electron microscopic examination of retinal
pigmented epithelium of kittens with Chediak-Higashi syndrome
disclosed a number of abnormalities of premelanosomes and
melanosomes. Few premelanosomes were present. Most of the melanin
granules were giant
sized , but their structures varied. Some of the
giant granules were composed of several premelanosomes and
melanosomes in different stages of maturation. Others contained
randomly oriented melanofilaments between melanosomes. There were
also complex giant granules consisting of both melanosomal and
lysosomal components. Inappropriate
fusion of cytoplasmic granules
appears to be the most likely mechanism for formation of the giant
granules. Fusion of premelanosomes with lysosomes and resultant
destruction of the premelanosomes probably is a major cause of the
ocular hypopigmentation of Chediak-Higashi syndrome."
Hargis
AM, Prieur DJ. Animal model:
renal lesions in cats with
Chediak-Higashi-Steinbrinck syndrome. Am J Med
Genet . 1987
Jan;26(1):169-79. "In this study designed to characterize the
renal lesions in cats with the autosomal recessive
Chediak-Higashi-Steinbrinck syndrome (CHS), the renal tubular
epithelial cells of CHS cats were examined by light microscopy. The
lesions observed were similar to those in the renal epithelial cells
of other species with CHS and were consistent with those reported
previously in other tissues of CHS cats."
Kramer
JW, Davis WC, Prieur DJ. The Chediak-Higashi syndrome of cats. Lab
Invest . 1977 May;36(5):554-62. "Three cats with Chediak-Higashi
syndrome were found in a single line of 27 Persian cats, and three
additional affected cats were produced from two prospective breedings
of the original line. The disorder was characterized genetically as
an autosomal recessive condition. All cats in the line with the
combination of yellow eye color and "blue smoke" hair color
exhibited the disorder. Four of the five cats examined had bilateral
nuclear cataracts as early in life as 3 months of age. No increased
susceptibility to infectious disease was observed. A bleeding
tendency was noted. Light microscopic studies of hair and skin
revealed enlarged melanin granules. These manifestations were similar
to those in man, mink, cattle, mice, and the killer whale with
Chediak-Higashi syndrome. Cats are the sixth species in which this
genetic disease has been reported." [Note: later studies of CHS
cats found the immune system to be impaired]
Prieur
DJ, Collier LL. Morphologic basis of inherited coat-color dilutions
of cats. J Hered. 1981 May-Jun;72(3):178-82. "The melanin
granules in hair of black, smoke, blue, Chediak-Higashi-smoke, and
pink-eyed dilution cats were studied. The hair of black cats contains
numerous small dark brown to black melanin granules uniformly
distributed throughout all portions. The basis for the dilution in
smoke cats is a paucity of melanin granules in the basal portions of
the hair. Blue cat hair has a larger basic melanin granule, some very
large but relatively regularly shaped granules, and a non-uniform
distribution of granules. The granules in the blue cat hair resemble
those in the hair of dilute mice. The Chediak-Higashi trait causes
even larger basic melanin granules than the blue dilution and
enlarged and relatively irregularly shaped granules. The melanin
granules in the hair of the pink-eyed dilution cat are very small,
and are yellowish brown compared to the dark brown to black of those
of black, moke, and blue cats."
Prieur
DJ, Collier LL. Inheritance of the Chediak-Higashi syndrome in cats.
J Hered. 1981 May-Jun;72(3):175-7. "The phenotypes with respect
to the Chediak-Higashi syndrome (CHS) of 245 cats from 67 matings
were analyzed. It was determined that the gene for feline CHS, like
that in other species with CHS, is inherited in an autosomal
recessive manner with complete penetrance. We propose the symbol ch
for the gene for CHS in cats. "
EYE
COLOURS
Copyright
2009, Sarah HartwellWild
cats in temperate regions generally have hazel eyes, but domestic
cats' eye colours vary from blue, through green to yellow, orange and
brown. Within each of those colours there is wide variation in hue
and intensity. The colours are not discrete, but form a continuum,
for example between blue and green there are "sea-green"
and aqua while between green and yellow there are
lemon and
lime shades. The coloured part of the eye is called the iris, a word that
means "rainbow".
Some
eye colours are linked to coat colour, for example colourpoint
(Siamese pattern) cats have blue eyes. In most breeds, various eye
colours were possible so breeders set chose the eye colour that
harmonised with the coat colour and wrote that into the breed
standard. While breeders might prefer certain pedigree black cats to
have vivid orange eyes, in the general cat population black cats can
also have green or yellow eyes.
HOW
EYES GET THEIR COLOURThere
are two major factors that influence feline eye colour: iris
pigmentation and blue refraction.
The
iris has 2 layers containing pigment-producing cells (melanocytes).
The outer layer is the stroma, consisiting of consists of loosely
arranged cells.
Underneath the stroma is the epithelium which
contains tightly packed cells. Both the stroma and epithelium produce
pigment, but in different amounts. The pigmentation of the iris is
caused by melanin and the colour ranges from lemon yellow to hazel to
deep orange or brown. In addition to this, the "transparent"
structures of cats' eyes absorb and refract light much like a sheet
of glass. Viewed head on, glass appears colourless but viewed edge-on
it is greenish or bluish. The size, spacing and density of the fibres
in the stroma determines how it reflects and refracts the light,
determining the shade of blue. Just like glass, the transparent
structures in cats' eyes are transparent blue, ranging from
practically colourless to deeper blues and violet-hued blues. The
combination of the blue refraction and iris pigment produces the
overall eye colour. Looking into a cat’s eye is like looking
through a blue-tinted window; the blue tint affects how we see the
other pigments present in the iris.
The
iris pigmentation and the transparent structure pigmentation are both
controlled by multiple genes in different
places on the chromosomes
(polygenes). Littermates can inherit different mixes of these genes
from the same parents and have different eye colours. Different mixes
of polygenes causes the continuum of shades.
The
type of colour and the intensity depends on the number of melanocytes
in the eye and how active they are. If there are no melanocytes, the
eye appears blue (or in rare cases, pink which is due to the colour
of the blood vessels). A low number of melanocytes gives green. A
high number of melanocytes gives orange. A second factor is how much
pigment those melonocytes produce (
activity ). Those that produce less
pigment give a lighter shade, while those that are more active and
produce a lot of pigment give the deeper shades. There is no single
inactive/active
switch , so there are varying shades inbetween. Eyes
with a low numbers of melanocytes (green) can range from pale green
(less active melanocytes) to
strong green (industrious melanocytes).
Similarly , the yellows range from light amber to deep copper. The
activity levels of melanocytes is genetically determined so it is
possible to selectively breed for either deeper or paler eye colours.
The way that light passes through the various blue-tinted structures
gives us the
final visible colour.
Kittens
are born with blue eyes. The adult eye colour develops around 6-7
weeks old, but may not reach its adult hue until 3-4 months old.
During a cats' lifetime, illness and injury can cause variations in
eye colour, but there is no truth in the old wives' tale that feeding
a cat on
fish will cause its eyes to change colour! That tale is due
to old wives weaning kittens (probably onto boiled fish in the days
before commercial cat foods) around the time the eye colour naturally
changed.
There
is also some pigmentation on the inside of the eye and a highly
reflective area behind the retina called the Tapetum Lucidum. This is
an iridescent layer of tissue that reflects light back through the
eye. and helps cats see in low light - it also causes eye
shine in
flash photos. Most blue-eyed white cats lack the Tapetum Lucidum;
their eyes reflect a dull red colour (caused by the blood vessels)
rather than an iridescent eye-shine.
THE
BLUESKittens
are born with blue eyes and this is not a guide to their adult eye
colour! In a few breeds, the blue colour is
permanent , but in most it
will be replaced by another colour starting around 6 weeks old. The
blue colour depends on the intensity of blue refraction.
Blue
eyes are a feature of colourpointed cats where the colour is linked
to the temperature-dependent albinism that produces the colourpoint
pattern. The blue colour varies in hue and intensity and breeders
must selectively breed cats with the best eye colour to try to
pass on that colour to the kittens.
Blue
eyes also occur in white cats and are associated with the epistatic
white (often called dominant white) gene which is quite different
from albinism because it masks an underlying coat colour. The white
spotting gene also affects eye pigment if there are white patches
around the eyes. White cats can have one or both eyes blue. The
non-blue eye will be whatever colour the breed standard requires for
example in white Persians, the other eye is vivid orange. In the
randombred odd-eye white cats, the non-white eye might be green or
yellow or even a different shade of blue. For more on blue-eyed white
cats
White
Cats, Eye Colours and Deafness.
The
Ojos Azules is characterised by cornflower blue eyes, but the
mutation is linked to lethal deformities if a kitten inherits 2
copies of the mutated gene. In this breed, the eye colour is
independent of the coat colour. Other blue eye mutations that are
independent of coat colour crop up from time to time. To date, most
have been a pale blue rather than an intense blue and none of these
other mutations have been deliberately bred.
A
grey bicolour (tuxedo pattern) cat with intense sapphire blue eyes
was found on a country road in Windellama, New South Wales, Australia
by A M Schnieder. Since the Ojos Azules has not been imported into
Australia, this represents a spontaneous mutation among the feral
population. Whether it is the same as the American Ojos Azules
mutation is not known.
GREEN
AND AQUAGreen
ranges from a yellowish-green through gooseberry-green to deeper
greens and blue-toned greens. Some of the older standards spoke of
grass-green eyes, indicating the green be as rich as possible. Green
tints and flecks also appear in some hazel brown eyes.
The
Chinchilla Longhair (Chinchilla Persian) and its shorthaired
equivalents are notable for their black-rimmed sea-green eyes. Aqua
(blue-green) is found in the Tonkinese which is intermediate in type
and colour/pattern between Siamese and Burmese cats. The Siamese has
blue eyes while the Burmese has orange eyes.
YELLOW,
ORANGE/AMBER AND BROWNYellow
varies from a pale lemon to more vivid hues. There is overlap between
the yellows and browns (pale hazel or tan) and also between yellows
and greens.
While
the boundary between yellow and green may be fuzzy, there is no
mistaking an orange-eyed cat. The colour is often described as
coppery. Many of the breeds developed by early cat fanciers in the UK
called for orange eyes as the pale greens and yellows were considered
undistinguished, while bright orange complemented the coat colour.
Brown
covers a wide range from hazel (the normal colour of temperate
climate white cats) through to darker browns. Some browns appear
tinted or flecked with green, orange or yellow.
ODD
EYES (HETEROCHROMIA)The
scientific term for odd eyes is heterochromia iridium (different
coloured irides). It can be genetic (inherited), congenital
(development defect) or acquired (illness, injury, medication). In
the cat fancy, odd-eyes means having one blue and one other-colour
eye. Odd eyes are most common in epistatic white cats where one eye
is blue and the other is orange, yellow, brown or green. Pedigree
odd-eyed white cats have one blue and one orange/amber eye, but in
randombreds the non-blue eye may be yellow, green or brown. Another
form of odd eyes is found in white cats where one eye has a tapetum
lucidum (green eye shine in the dark) and one eye lacks the tapetum
(red eye shine). These different eye shines are seen in reflected
headlights or flash photography. Bicolour cats with a high degree of
white on the face may also have odd eyes.
Heterochromía
from birth is common in cats (blue/amber), horses (blue/brown) and
some breeds of dog (blue/brown). It is uncommon in humans. In cats,
there is no specific gene for "odd eyes," but it is
associated with the white spotting gene and the white masking gene
(epistatic white/dominant white). These genes prevent pigment
production in the embryonic stage, which includes the pigment of the
iris. If some melanocytes (pigment cells) remain active in the eye
area, this is sufficient to make the eye green, amber, brown or
yellow etc. Depending on the activity level of the melanocytes, the
eye colour ranges from blue with other tints through to fully
coloured, and from pale through to intense colour. During the cat's
lifetime, the effect may be incremental so that the intensity of eye
colour deepens, gradually replacing the blue colour.
Heterochromia
is mostly found in cats with white coats or white patches.
Occasionally it is seen in coloured cats. When present from birth, it
can be a result of congenital abnormality (birth defect), mosaicism
(merged embryos), or a somatic mutation in some cells that form the
skin and eye (birthmark). These are not hereditary and this form of
heterochromia is not passed from parent to offspring.
Odd-eyed tortoiseshell cat.
Odd-eyed white cat photographed by Rodrigo Arancibia in Iquique, Chile. She shows no
sign of deafness. her kittens are solid white, indicating the dominant white gene.
Less
commonly , other colour cats may have odd eyes for example a tortie
with one yellow and one blue eye. This can be due to the eyes
developing differently in the embryo. Perhaps the cat has a white
spotting gene that hasn't showed up on the fur, but has affected the
eye. Below is another odd-eye tortie: a 10 year old tortie/tabby
called Debby photographed by Marjan Boonen in The Netherlands. Since
tortie cats' patched colouration is due to mosaicism, it's possible
this has extended to the eyes resulting in two different colours
Occasionally
the different colour eyes are due to an eye injury or illness that
has damaged the iris or one eye. In this case the trait cannot be
passed on.
DICHROIC
(DICHROMATIC) EYESPartial
heterochromia (dichroic eyes) occurs when different amounts of
melanocytes, or melanocytes with different activity levels, are
present in distinct areas of one iris. This gives the “pie slice”
or “coloured halo” effects. Dichroic/dichromatic eyes have two
colours in one iris. For example there may be a yellow ring around a
green iris. This is considered a fault in show cats, but can be very
attractive.
Less
commonly, there may be a distinct differently coloured section (like
a slice of a pie) in the iris, for example a brown area in a blue or
green eye. This has been reported in some white cats. Instead of
affecting the pigmentation of the whole iris, the white gene affects
only part of the iris. One or both eyes can be affected; if both are
affected there may be a mirror image effect. These are sometimes
called "weird-eyed" cats because they are not odd-eyed in
the normal
sense .
The
photo is "Gilbert" who has one blue eye and one dichoric
eye.
LILAC
/ BLUE-EYED ALBINOAlbinism
is found in the Siamese to closely related Oriental breeds due to
inbreeding.
Emma Stothers (2013) provided photos of her albino
Oriental Longhair, Pangur. Pangur's eyes look like faded lilac-blue.
On close inspection, the pupil is a dull red, while what looks like
lilac is just the pale blue refraction of the lens overlaid on a
pinkish pupil. This makes Pangur very photo-sensitive. Pangur's
parents are half-siblings with the same father.
PINKPink
is not a normal eye colour in cats. It is found in pink-eyed albinos
and is due to complete lack of pigment in the eye combined with very
little blue refraction. True albinos entirely lack melanin and the
iris is pale pinkish-white. Because of blue refraction, the eyes
usually have a bluish hue, result in a blue-eyed albino (as with
Pangur above). Very rarely the transparent structures are colourless
and a pink-eyed albino results.
It
was once believed impossible for a cat to have pink eyes due to the
structure of the eye and true albinos had pale blue eyes, but there
have now been several pink-eyed albinos reported in recent years.
Albinism has been recorded in the Bengal breed and in its wild
parent, the Asian Leopard Cat (shown here).
Pink-eyed
dilution has been reported once in the cat (Todd NB. 1961. "A
pink-eyed dilution in the cat" Journal of Heredity 52, pg 202).
The type of dilution found in cats is blue dilution which turns black
into grey. A second type of dilution seen in some mammals is
"pink-eyed dilution" which gives a bluish-tan/fawn coat and
also depigments the eye, giving a pink or ruby appearance. This has
been reliably reported only once in cats when a pink-eyed female with
a light tan coat was produced (none of her kittens survived so
pink-eyed dilution in cats seems to have been lost).
Pale
irises and underpigmented retinas also occur as part of
Chediak-Higashi syndrome along with a pale coloured coat (seen in
Blue Smoke Persians) .
EYE
SHAPESIn
addition to the different colours, there are several distinct eye
shapes. These are shown in slightly exaggerated form in the chart
below.
TORTOISESHELL
AND TRI-COLOUR CATS
Copyright
2002 - 2013, Sarah HartwellBecause
of the way tortoiseshell is inherited, almost all tortoiseshell and
calico cats are female. The very few male tortoiseshell cats are
caused by genetic aberration or development abnormalities in the
foetus. These are discussed in detail in "
Mosaicism,
Tortie Tomcats, Genetically Impossible Kittens and Gender Anomalies".
This article only considers normal tortoiseshell cats.
Jean
Bungartz referred to tortoiseshell-and-white cats as "Spanish
cats" in his 1896 book "Die Hauskatze, ihre Rassen und
Varietäten" (Housecats, Their Races and Varieties) in "
Illustriertes Katzenbuch" (An Illustrated Book of Cats) though
he said it was hard to explain why they got the name as they were not
restricted to the Pyrenean peninsula.
HOW
TORTOISESHELL PATTERNS OCCURThe
ginger colour of cats (known as "yellow", "orange"
or "red" to cat breeders) is caused by the "O"
gene. The O gene changes black pigment into a reddish pigment. The O
gene is carried on the X chromosome. A normal male cat has XY genetic
makeup; he only needs to inherit one O gene for him to be a ginger
cat. A normal female is XX genetic makeup. She must inherit two O
genes to be a ginger cat. If she inherits only one O gene, she will
be tortoiseshell. The O gene is called a sex-linked gene because it
is carried on a sex chromosome. Tortoiseshell cats are therefore
heterozygous (not true-breeding) for red colour.
The
formation of red and black patches in a female with only one O gene
is through a process known as X-chromosome inactivation. Some cells
randomly activate the O gene while others activate the gene in the
equivalent
place on the other X chromosome. This only shows up
visibly in skin cells as these produce pigment. This occurs early on
in the embryo and as skin cells multiply, they form patches. The skin
is a mosaic of cells where some have the O gene active (making ginger
pigment) and some do not (making black pigment). This can only happen
in cats with two X chromosomes. Male cats only inherit one X
chromosome so this is active in all skin cells as there is nothing
equivalent on the Y chromosome which could "switch off" the
O gene.
There
are two main theories regarding brindled torties and patched torties.
One (the "early/late deactivation theory") suggests that
the time at which X chromosome deactivation occurs during foetal
growth determines whether the cat has well defined patches or is
brindled with intermixed black and orange hairs. Skin cells multiply
during growth and spread out across the skin; as the embryo grows the
skin cells multiply. If deactivation occurs early on each pigment
cell has
room to multiply into, a "red" cell will multiply
into more red cells while a "black" cell will give rise to
patches of black. If it occurs later, the patches are smaller as the
cells have less room to multiply into; some "patches" will
be no larger than a single hair! All "red" cats are red
tabbies - where there are large red patches, the tabby pattern will
usually be discernible.
All
Red Cats Are Red Tabbies? Strange
as it may seem, all red cats are actually red tabby because the
non-agouti gene (the gene that turns a tabby into a self/solid
colour) does not affect the way red pigment is deposited. A variety
of other genes, called polygenes or modifiers, control the intensity
of colour and contrast between markings and background colour.
Selective breeding has reduced the tabby markings to produce a cat
that looks solid red by breeding from those cats with the least red
markings (cats with "low contrast" between markings and
background colour). Because the non-agouti gene does not work on the
red pigment, red tabby ghost markings can never be completely
eliminated and may be seen on the tail, legs and forehead and as a
darker region along the spine. Even though red cats are registered as
"red self" they are still red tabbies, albeit red tabbies
with very reduced markings. Because they lack the polygenes for high
contrast between markings and background colour, their offspring also
appear to be red self. More information can be found in Robinson's
"Genetics for Cat Breeders".The
other theory regarding brindling and patching (the "migration
theory") is that brindled torties occur when there are more
pigment producing cells produced from the neural crest (which becomes
the back and spine area). The cells are assumed to have undergone X
chromosome deactivation before migration. The migrating cells carry
either O (red) or o (black) and they migrate at the same rate into
their final positions. Where there are many pigment producing cells,
there is more competition (
imagine cells jockeying for
position on
the skin) and they become intermingled. Where there are fewer pigment
producing cells, each cell has room to grow into patches of colour
(imagine
plants forming clumps in a
flower border).
As
an interesting side note, human women can also have X-inactivation.
Although there are no tortoiseshell women, there is an uncommon
condition called anhidrotic ectodermal dysplasia caused by a faulty
gene on the X chromosome. The faulty gene reduces the number of sweat
glands per
square centimetre on the skin. In women who inherit one
normal gene and one faulty gene; their skin may be divided into
hundreds of small patches which only become visible when they sweat.
The normal patches of skin sweat normally, but the patches derived
from cells with a faulty gene don't.
To
be completely accurate, O = orange and o = "non-orange"
meaning that o allows whatever colour is carried on other chromosomes
to be displayed (black, blue, chocolate etc). The O gene converts
normal black/chocolate/cinnamon etc pigment into red pigment. For the
sake of simplicity, this article refers to o as "black"
since that is the non-orange colour most commonly seen on tortie and
calico cats.
TORTOISESHELL-AND-WHITE
(CALICO) CATSThe
white patches in tortie-and-white (tricolour, calico) cats is caused
by the piebald spotting gene discussed in
Beautiful
Bicolours.
This is a semi-dominant gene with very variable expression ranging
from
nearly all white to nearly all coloured with only a few white
hairs. The gene affects the embryo cells which will become
pigment-producing skin cells. These cells are initially formed along
the "neural crest" - the embryo's backbone area - and
migrate to all over the body during formation of the skin. Where
these pigment producing cells fail to get in position before the skin
is fully formed, there will be areas of skin which lack pigment
producing cells i.e. white areas. White areas are usually the areas
furthest from the cat's backbone - paws, belly, chest and chin -
these areas take longest to reach.
One
effect of white spotting in tortoiseshell cats is to change the
pattern from brindled to patched. Tortie cats with little or no white
tend to have brindled coats. However, the more white there is, the
more the black and white will also be separated out into patches
instead of being intermingled. The phenomenon of tortie-and-white
cats having better defined patches of colour is familiar to most cat
lovers.
In
the developing embryo, the pigment forming cells migrate from the
neural crest. If the "migration theory" is right, cells
which activate O (red) and those which activate o (black) appear to
migrate at the same speed, leading to a brindled pattern. If the
embryos also inherit the gene for white spotting, the fur develops as
patches of colour. The bigger the white areas, the bigger the and
better defined the separate patches of black and red. The presence of
the white spotting means fewer pigment producing cells and less
competition between them as they migrate into position. One or two
cells reach an area and these multiply in situ to form a patch of
colour (a clonal patch).
Red
Tabby Markings in Tortie and Tortie-Tabby Cats.
Where a tortie (or calico) cat has the non-agouti gene it will appear
to have solid patches of colour. Due to the quirk of the red pigment
being unaffected by the non-agouti gene, there may be tabby markings
visible in the red areas. How much red tabby pattern is visible is
due to other genes called polygenes that also affect the depth and
contrast of the red colour (the
illusion of solid red is due to low
contrast between background and markings). Where a tortie (or calico)
has the agouti gene instead, the tabby markings will be visible in
all of the coloured patches. In a tortie cat, the non-red colour will
be a solid colour. In a torbie cat, the non-red colour will also have
tabby markings. More information can be found in Robinson's "Genetics
for Cat Breeders".
TORTIE
ABYSSINIANS AND TORTIE TICKED TABBIESThere
is another gene which gives rise to the red colour. This is the
non-sex-linked red ("sorrel") found in Abyssinian cats and
other ticked breeds and it does not produce tortoiseshells.
Genetically, the Abyssinian pattern is not a solid colour, but is a
form of tabby, called ticked tabby, and is described in
Striped
and Spotted Cats.
The ticked pattern changes the visual appearance of the cat's colour.
Although some registries call this colour "red"
("non-sex-linked red" in Australia), many registries prefer
to call it "sorrel" to avoid confusion with the sex-linked
red gene described earlier. Genetically, Abyssinian "non-sex-linked
red" is the equivalent of cinnamon in non-ticked cats. It
appears as a reddish-brown due to the ticked pattern of the hair.
Chocolate Abyssinians appear a deeper red which can be differentiated
from sorrel by the chocolate's darker tail tip.
Strictly
speaking, because Abyssinian is a tabby pattern, the tortie versions
should be termed ticked torbies (tabby-torties) rather than ticked
torties. This photo shows Harley (Harlequin) and Spot (provided by
owner Lizzi) who were resuced as feral kittens. Harlequin is a ticked
torbies, while littermates Spot is ticked tabby with white, and Blaze
and Mr Grey are mackerel tabbies with white. The two mackerel tabbies
are quite lightly marked on a ticked background and show some
rufousing (red tints).
The
true sex-linked red Abyssinian is bright orange with a red tail tip.
It is important for breeders to know whether they have sex-linked red
or non-sex-linked red as this will affect the breeding program. Where
there is sex-linked red, there can also be tortoishells. Tortie
Abyssinians occur, but since the breed does not permit white, these
are always brindled. The combination of brindling and ticking can
make it almost impossible to determine whether a female is tortie or
not just from a visual inspection. Sometimes a female Abyssinian is
only known to be a genetic tortie when she produces a mix of red and
black kittens!
Ticked
tabby also occurs in non-pedigree cats. In randombred cats with the
ticked tabby pattern, white markings may be present as well,
resulting in "ticked tortie tabby and white" (ticked
calico). The photo shows part pedigree tortoiseshell ticked tabby
kittens with white markings.
DIFFERENT
TORTOISESHELL COLOURSThe
basic tortoiseshell combination is black and red. Because red does
not occur as a true solid colour, the "red" patches are
actually red tabby and this will be more apparent on some cats than
on others. The addition of a dilution gene gives rise to the other
tortoiseshells combinations. In Burmese cats, where there is a
different dilution gene at work, diluting black into brown.
The
action of caramel is noted as theoretical, in reality the colours may
be indistinguishable from other colours unless the presence of
caramel is known from the cat's pedigree; the entries are based on
data from solid colour cats. The colour intensifier is also
theoretical, based on the possible "indigo" gene which
turns blue into a deeper richer colour. This could equally be due to
polygenes (multiple genes having a cumulative effect). I have
included it after seeing one cat which was dark-blue/hot-cream
tortoiseshell i.e. where both colours were intensified.
NON-DILUTE COLOURDILUTE VERSIONCARAMELISED VERSION OF THE DILUTE VERSION (DOUBLE DILUTION)INTENSIFIED VERSION OF THE DILUTE VERSION
("INDIGO" GENE)Tortoiseshell
Black/Orange Tortoiseshell
(Brown Tortoiseshell in
Burmese cats)Blue-Cream Tortoiseshell
Caramel/Apricot
Indigo/Hot Cream (dark-blue and hot cream) (theoretical)
Cinnamon Tortoiseshell
Milk chocolate and cream
Fawn Tortoiseshell:
Fawn and cream
Caramel/Apricot
Chocolate (Chestnut) Tortoiseshell
Warm milk chocolate, red, and cream
Lilac (Lavender) Tortoiseshell:
Frosty lilac-grey and cream
Taupe/Apricot (theoretical)
Above
are black (smoke), solid chocolate and chocolate tortie kittens. The
black is shown for colour comparison. Chocolate and
choc -tortie are
more common in pedigree cats than in randombreds, but these kittens
were born to non-pedigree parents. Photo courtesy of Leva Cygnet.
In
Siamese cats, only the points (legs, tail and face) show as
tortoiseshell and because the gene causing the Siamese pattern turns
black into dark brown, the tortoiseshell pattern will contain dark
brown rather than jet black.
TORTOISESHELL
TABBY AND TORTOISESHELL TIPPEDTortoiseshell
combines with the tabby pattern to create tortoiseshell-tabbies, also
known as patched tabbies or (in the US) torbies. The tabby pattern
may be classic (blotched), mackerel or spotted.
In
a usual tortoiseshell, the black patches become brown tabby (i.e..
black markings on brown background), the red patches become more
obviously red tabby (red patches on paler red background). Patched
tabbies occur in other tortie colours e.g. blue-cream patched tabby.
In
combination with silver, this would produce tortoiseshell markings on
a silvery background e.g. black-silver (black markings on pale grey)
and red-silver patches; though such combinations are not common.
The
tortoiseshell markings can also be combined with the genes for
tipped, smoked or shaded in which case the tortoiseshell colours are
restricted to the hair-tips rather than going all the way to the
root. This gives the effect of a shimmering tortie pattern on a pale
undercoat.
BREEDING
TORTOISESHELLSIn
order to produce tortoiseshell kittens, at least one of the parents
must carry the O gene. Red male and red female only produce red
kittens. The combinations which produce tortoiseshell kittens are red
male and non-red female, red female and non-red male. The following
can also produce tortoiseshell kittens: tortie female and non-red
male, tortie female and red male. It also depends on some kittens
being female.
Parents
Kittens
Female
Male
Males
Females
Red
Red
Red
Red
Black
Red
Black
Tortie
Tortie
Red
Red
Red
Black
Tortie
Tortie
Black
Red
Tortie
Black
Black
Red
Black
Red
Tortie
Black
Black
Black
Black
This
is what is happening when a tortoiseshell female is mated to either a
red or a black male. Because a tortie female is heterozygous, she can
pass on either the gene for red or the gene for non-red (black). The
colour of the kittens depends on whether they are male or female and
what gene, if any, they inherit from the male.
GENES FROM RED MALE
GENES FROM TORTOISESHELL FEMALE
GENES FROM BLACK MALE
GENES FROM TORTOISESHELL FEMALE
O (red)
o (non-red [black])
O (red)
o (non-red [black])
O (red)
X chromosome
OO
Red female
Oo
Tortie female
o (non-red [black])
X chromosome
Oo
Tortie female
oo
Black female
No gene (Y chromosome)
O-
Red male
o-
Black male
No gene (Y chromosome)
O-
Red male
o-
Black male
Because
other conditions can give the impression of being tortoiseshell, some
matings which theoretically have a high probability of producing
tortie kittens will only ever produce red or black kittens. At the
other extreme, a genetically tortoiseshell cat may appear to be solid
ginger or solid black because of the way the X chromosomes activated
- these cats will produce unexpected tortie kittens in appropriate
matings.
CLONING
TORTOISESHELLSThis
may seem an odd topic, but if you clone a tortoiseshell cat you will
end up with a cat of one or other of the constituent colours and not
a tortoiseshell clone. If you clone a red/black tortoiseshell, the
clone will be either red (ginger) or black.
DNA
tests on the tabby-and-white cloned kitten ("Cc") proved
that she was a clone of her tortie-and-white genetic mother (the cat
whose cell was used to create Cc). The pattern of pigmentation in
multicoloured animals is the result of genetic factors combined with
developmental factors within the womb. This means bad news for owners
who want an exact replica of a tortoiseshell or calico cat. Why isn't
Cc also tortie-and-white? The
answer is due to ‘X-linked
Inactivation'.
Tortoiseshell
cats have two X chromosomes, one carrying the gene for orange coat
colour and the other carrying the gene for black coat colour. As the
embryo develops, a process called ‘X-linked inactivation’ occurs
in its tissues. One or the other X-chromosome in every cell in a
tortie cat embryo is randomly inactivated. This only shows up in
pigment producing cells, producing the familiar tortie effect.
Regardless
of which cell was used to produce Cc, because that cell is already an
adult cells, one or other of the cell's X chromosomes would have been
inactivated while the
donor cat was
an embryo. Cc had an equal chance of being orange-tabby-and-white or
black-tabby-and-white, but would never be tortie-tabby-and-white.
Unless a way can be found to undo X-linked inactivation at the embryo
stage, owners wishing to clone a tortoiseshell cat will have to
settle for a cat of a different colour entirely. If the X-linked
inactivation can be reset, the inactivation is a random process so
the clone will have the right colours, but not in the same places as
the donor cat - it may have well-defined patches of colour while the
donor cat was thoroughly brindled.
TRI-COLOUR
BENGALSConfusingly
for cat lovers, tri-colour Bengals are not tortoiseshell cats.
In the Bengal breed, tri-colour refers to a cat with black rosettes
on a golden brown background. The centre of the
rosette is generally
a darker version of the background colour, hence the cat has three
colours: pale background, black markings, dark rosette centre.
Tortoiseshell
Bengal variants have occurred during breed development, but it is not
an accepted colour in a breed which aims to re-create a wild look.
ANOMALOUS
TRI-COLOURSA
number of anomaous tricolours are discussed in "
Mosaicism,
Tortie Tomcats, Genetically Impossible Kittens and Gender Anomalies".
These include a white, black and grey tricolour and a white, red and
grey tricolour. A further unusual tricolour is best described as a
"white brindle".
The
"white brindle" was one of the most striking
"tortoiseshell" cats I have seen. Imagine a brindled
tortoiseshell in which all the black areas have been converted to
white. The result was a cat with intermixed fur of white, cream and
ginger; the fur being brindled like a tortie rather than being
patched like a red-and-white bicolour. This was seen while on holiday
(either Turkey or Tunisia) and unfortunately I didn't have a camera
with me. I have never seen anything like it since. A likely
explanation is a genetic mutation in the fertilised egg had caused a
pigment production fault in the areas which should have been black.
It would have been interesting to see if the trait could have been
perpetuated.
One
reader suggestion for the "white brindle is a
red-tabby-and-white smoke shorthair. This would give a similar
effect, but the colour in a smoke cat does not go down to the roots
and the brindled effect would not be so thorough. I am currently
inclined to think that the cat was a conventional brindled tortie
which was not producing black pigment.
WHITE
CATS, EYE COLOURS AND DEAFNESS
2001
- 2016, Sarah HartwellThis
page has two linked topics - the different causes of blue eyes and
white fur in cats is one, the other is the relationship between eye
colour, coat colour and deafness. This means some information in one
section might also be repeated or summarised in onther section (that
way you can read a section and not have to read the whole page).
ALBINO
OR JUST WHITE?A true albino catThe eyes are unpigmented and appear pink. The skin is pale pink.
A true albino kitten, age 7 weeks, owned by Michelle R. McGaha. The eyes are unpigmented and appear pink. The skin is pale pink. The mother is a tortie Manx, the father is unknown. He is very light sensitive and will need sun-
protection on his ears to guard against skin cancer (all white-eared cats have a risk of skin cancer if they sunbathe too much).
Contrary
to popular belief, white cats with blue, orange or green eyes are not
albino. Albino cats, such as the one in this photo, have pink
(unpigmented) or bluish-pink eyes and, like most albino animals,
their eyes are sensitive to light. The white colour in cats is due to
a gene that masks any other colour genes (this is why white cats can
have non-white or bi-colour kittens). Albinism is a different
mutation that causes the absence of colour, not the covering up of
colour. When owners talk about having green-eyed or orange-eyed
albino cats, they mean green-eyed or orange-eyed "dominant
white" cats. In this context, dominant means a gene that masks
other genes, it does not mean the cat has a dominant personality.
Recessive
white is an alternate name for the blue-eyed/pink-eyed albino gene
which is part of the colourpoint series of genes. The white spotting
gene can sometimes produced solid white cats and is also sometimes
called recessive white.
It
is also a fallacy that "all blue-eyed white cats are born deaf".
All kittens are born blue-eyed and with their ears folded down.
Whether they will stay blue-eyed and whether they will be deaf can
only be ascertained after a few weeks. Not all blue-eyed white cats
will be deaf (as this article explains).
DOMINANT
WHITEDominant
white (more properly "epistatic white" since it occurs on a
different gene to the black-based or red-based colours) denoted by
the gene symbol "W", is the colour associated with deafness
in cats. Dominant white masks all other colours and cats may have
blue, orange or odd eyes. Those with blue eyes have a high chance of
deafness. Those with one blue eye have a high chance of deafness on
the blue-eyed side. Those with orange eyes are far less likely to be
deaf. Some dominant white kittens are born with smudges of coloured
fur on top of the head where the colour is incompletely masked, this
smudge of colour usually disappears by adulthood, but kittens with
colour smudges are more likely to have normal hearing. These cats are
not albinos; genetically they can be any colour, but the white is
dominant to those other colours (albinism is an absence of other
colours).
Lacy has the dominant white gene associated with blue eyes and deafness (Lacy is deaf). Lacy owns Greg Schultz who refers to her as "The Queen".
Mikey has the dominant white gene, but has green eyes rather than blue. He is also deaf. Mikey is owned by Jennifer Moore (ibjennyjenny).
In
some animals, the blue-eyed white trait is sex-linked (carried on the
X chromosome) and is found in males more often than in females. In
cats, the gene for white is carried on an autosome (a chromosome
other than the X or Y sex chromosomes) and the trait occurs equally
in male and female cats. Blue-eyed white is not sex-linked in cats.
WHITE
SPOTTINGThe
gene for white spotting, denoted by the gene symbol "S",
can also create the impression of a self white cat. This gene is
semi-dominant and is variable in the way it is expressed - a cat may
have no visible white spots or may be wholly white and all stages in
between those two extremes. Unlike dominant white, white spotting is
not linked to deafness.
Some
white cats are due to the very variable expression of the gene for
white spotting. The diagram above shows a typical progression from
solid colour through to solid white caused by this gene. The number
by each diagram is the "Grade" of spotting from Grade 0 (no
white spotting) through to Grade 10 where white spotting has obscured
all of the base colour. Grade 10 white spotted cats resemble solid
white cats, but are rarely deaf. Again, small spots of colour may be
discernible on cats that appear solid white due to the white spotting
gene.
ALBINO
CATS (RECESSIVE WHITE)Albino
is generally thought of as pure white, but the situation in cats is
more complex. There are five known alleles for albinism: blue-eyed
albino ("ca"),
pink-eyed albino ("c"), Burmese pattern ("cb"),
Siamese pattern ("cs")and
full colour (non-albino, "C"). Full colour is dominant to
all of the other four alleles. Burmese pattern is incompletely
dominant to Siamese pattern; cats that inherit one of each of those
genes will be intermediate in pattern and is known as Tonkinese. A
quirk of the Siamese form of albinism is that it is temperature
dependent with warm areas of the body being paler than cooler areas.
For this reason, it is often described as "colour restriction"
rather than albinism. Pink-eyed albino appears to be recessive to all
of the other albino mutations. Albinism is not linked to deafness in
cats ("dominant white" (W) is the gene linked to deafness).
Full
albinism - also known as recessive white - is rare. When present it
affects the structure of the tapetum of the eye so tht it reflects
red/pink instead of reflecting green.
In
1927, judge Mrs Basnett reported on the Paris Cat Show held on
14th and
15th of
January by the Cat Club de France and wrote "One lady brought
for my inspection a Siamese Albino of about 4 months; I could not see
a trace of any shading anywhere on the white coat, tail, legs, ears
or mask; the eyes were a very beautiful blue, and their fiery centre
seemed to be accentuated by the absence of all shading. In appearance
it was a very typical Siamese with the long
sleek body and whip tail,
and a very beautiful wedge-shaped head and face." Albino Siamese
have also been purposely bred; they are completely white Siamese-type
cats with bluish-pink eyes (true "pink eyes" are uncommon
due to the physical structure of a cats' eyes). The "European
Albino" bred in
Belgium is a European shorthair type white cat
with ruby-red eyes which have pale translucent blue irises. The
albino cats reported in Europe and the USA seem to be intermediate
between pink-eyed albino and blue-eyed albino. A true pink-eyed
albino was reported in 1931 and again in 1980s in the USA.
Albino
Siamese had pinkish-blue eyes rather than the clear blue related to
dominant white in the Foreign White breed. The mutation for "true
albinism" was reported when a Chocolate Point female bred to her
Chocolate Point son (this line became quite inbred in order to
produce the albino cats). The recessive white gene has continued to
lurk in the Chocolate and Cinnamon Oriental gene pool, where it somes
manifests by affecting the eye structure and producing blue-eyed
Orientals. When pink/red eye shine shows up in breeds of cat, or in
randombreds, it is most likely due to chocolate or cinnamon in their
ancestry. It has been observed in some oriental cats and in
chocolate/cinnamon Devon Rex, Affected cats are not necessarily
wholly or partly white and the only hint of this gene seems to be the
red eye shine instead of green eye shine! Why does it only visibly
affect eye structure? All
multi -celled creatures are mosaics where
genes can be switched on in some cells but not in others. For some
reason, during the development of the embryo, the lurking albino gene
got switched on in those cells that formed the eyes.
OTHER
ALBINISMAn
albino kitten was born at Chelmsford Cats Protection League shelter
in the 1990s and required handrearing. It was described as white
furred and having very pale pink ears, nose leather and paw pads
though I have no information on its long term survival. Albino
kittens have turned up more recently in the Bengal breed,
unsurprising since albinism is found in the Asian Leopard Cat (the
wild parent of the Bengal).
In
the Ojos Azules cat, normally typified by bright blue eyes in
combination with colours other than white or colourpoint, the
homozygous form of the gene has caused dead albino kittens.
WHITE
CATS AND DEAFNESSA
few years back I was asked three related questions on a newsgroup.
This article is adapted from my answer.
-
Are white cats, particularly blue eyed white cats, always deaf or is
this an old wives' tale?
- Is deafness linked only to
odd-eyed white cats?
- Some blue-eyed whites aren't deaf -
why?There
is an established link between the white coat color, blue eyes and
deafness. The tapetum lucidum is generated from the same stem cells
as melanocytes (pigment cells). The blue eyes in a piebald or
epistatic white cat indicates a lack of tapetum. Deafness is caused
by an absence of a cell layer in the inner ear that originates from
the same stem cells as well. In odd-eyed white cats, the ear on the
blue-eyed side may be deaf, but the one on the orange-eyed side
usually has normal hearing. Not all blue-eyed whites will be deaf
since there are several different genes causing the same physical
attributes (whiteness, blue-eyedness) so it all depends on the cat's
genotype (its genetic make-up) not its phenotype (its physical
appearance). Some people claim that 99% of blue-eyed white cats are
deaf. This is inaccurate because blue-eyedness and whiteness can both
be caused by different genes. It all depends on what genes the cat
has inherited. These are the actual figures from scientific studies
around the world. The percentages are given in ranges because results
are different in different areas, partly because of the different
genes found in the cat population. Where a cat is classed as deaf,
the deafness may affect one or both ears.
- 95% of the general cat population is non-white cats (i.e. not pure white) and congenital deafness is extremely rare in non-white cats.
- 5% of the general cat population is white cats (i.e. pure white). 15-40% of these pure white cats have one or two blue-eyes.
- Of those white cats with one or two blue eyes, 60-80% are deaf; 20-40% have normal hearing; 30-40% had one blue eye and were deaf while 60-70% had one blue eye and normal hearing.
- Of the 5% of white cats in the overall population, 60-80% had eyes of other colors (e.g. orange, green). Of those 10- 20% were deaf and 80-90% had normal hearing.
- Deaf white cats with one or two blue eyes account for 0.25 - 1.5 of total cat population
- Total number of cats with white coat and blue eyes account for 0.75 - 2.0% of total cat population
It
is evident from those studies that blue eyed whites exhibit a higher
incidence of deafness than do orange/green eyed whites or non-white
cats! But not all blue eyed whites are deaf and here's why:-
There
is a known link between white coat color, blue eyes and deafness -
but since the coat and eye color can be caused by different genes it
means that only
some blue
eyed whites are deaf. There is a gene/gene complex which causes white
coat, blue eyes and deafness, but not all cats get their white coat
and blue eyes from that particular gene, so not all white cats will
be deaf.
If
the cat is a Foreign/Oriental White, it carries the gene for 'Siamese
Blue Eyes' which is not linked to deafness (the gene for Siamese Blue
Eyes is linked to cross-eyes instead). Siamese blue eyes have a
reflective tapetum, but this is depigmented because the Siamese
colour is caused by albinism. This depigmentation gives the red-eye
with flash cameras. Random matings can mean that this gene sometimes
appears in non Oriental-looking cats which have colorpoint cats in
their ancestry.
Crystal (owned by
Alana Harley, Vancouver Island, Canada) has two different types of blue eye.
As this photo shows, one eye has normal green eye-shine and the other has pink eye shine. Depigmentation gives the red-eye with flash cameras
Albino
cats are also white. True albinism causes pinkish eyes, but some
albinos or partial albinos have pale blue eyes. There are too few
albino cats studied to draw
firm conclusions, but this mutation is
not necessarily linked with deafness. It is also hard for a cat owner
to determine whether their cat is a blue-eyed albino cat rather than
an ordinary blue-eyed white.
Jamie Linton, who volunteers at a shelter in California, came across this blue-eyed white young cat. The blue is restricted to an inner ring around the pupil, fading to white around the outside of the iris. It wasn't known if the cat was deaf. The most likely explanation seems to be that pigment remained concentrated at either end of the iris muscles as the eye grew, rather than being distributed evenly - a bit like pulling toffee where the middle section goes pale as it it stretched and the more intense colour remains at either end.
There
is also a gene for blue eyes which is inherited separately from coat
color. This is the gene responsible for the Ojos Azules breed. If the
cat is white colored, there is no easy way of telling whether it has
the blue-eyed-deaf-ear type gene or the Ojos-Azules-blue-eyes type
gene. It's only possible to
tell that a cat has this particular gene
if the cat is non-white and has blue eyes. Genes for blue eyes
independent of coat color may be more common than previously
realised. I have encountered three blue-eyed random-bred cats (one
ginger, one silver tabby, one brown tabby) in one UK town between
1989 and 1995. This may be the same gene as Ojos Azules, or it may be
due to different gene mutations. Other types of blue eye are being
discovered in all-white cats in Asia, the blue is different than
Siamese Blue Eyes, but does not seem to cause deafness.
The Lady Chablis, belonging to Denise Parmentier, is an odd-eyed white. One eye is yellow-green and the other is blue. She has perfect hearing.
Odd-eyed white cat photographed by Rodrigo Arancibia in Iquique, Chile. She shows no sign of deafness. her kittens are solid white, indicating the dominant white gene.
The
white coat can be caused either by a gene for white coloration or by
a gene for 'white patching' - sometimes the white patching is so
extensive that the cat appears solid white. If the white cat
exhibited a few colored hairs or a smudge of color on its body
(usually on the head) as a kitten, then it should have normal hearing
even if it has blue eyes because it has inherited a non-deafness
causing gene for white coat! However, blue-eyed bicolour cats occur
and the more white they have in the region of their eyes and ears,
the more likely they are to be deaf. This is because one of those
genes for white affects the development of both the eyes and ears -
it causes lack of eye pigmentation (i.e. blue eyes) and deafness -
and since the eyes and ears are close together, if that gene affects
that area of the body, it is likely to affect both sense organs
(hence odd eyed whites may be deaf on the blue-eyed side).
Aries, a blue-eyed silver-tabby-and-white Norwegian Forest Cat bred by Raquel Ortega Cormenzana of MONTEGANCEDO*ES (Madrid). Sometimes bicolour cats with white faces have blue eyes. Aries has has perfect hearing because the white does not extend to the ears.
Congenital
deafness can also be caused by a variety of hereditary factors, just
as in humans. Due to random mutation or the wide gene pool, cats of
any color can be born deaf (probably less than 1%) - including
orange-eyed whites. Hence deaf white cats with non-blue eyes can
occasionally appear and in these cases the deafness is not linked to
coat color. Deafness can also be caused by illness or injury, so a
person adopting a deaf cat may not know whether the cat was born that
way or became deaf later on.
There
are also green-eyed white cats, the Russian Angora is green-eyed and
white is a favorite color. Green-eyed white cats have a
lower incidence of deafness than blue-eyed white cats because the gene for
white they carry does not normally affect their eye color. It is
similar for orange eyed whites; they rarely have congenital deafness.
Genetic expression is very variable and
orange-eyed/odd-eyed/blue-eyed whites are interbred in many breeds -
hence up to 20% of non-blue-eyed, white cats may have some degree of
hearing impairment depending on what gene is causing them to have a
white coat.
So
overall, blue-eyed white cats
stand a higher than usual chance of
being deaf; but they are not guaranteed to be deaf. Odd-eyed white
cats
may be
deaf on the blue-eyed side. If you have a deaf white cat, it is not
advisable to breed from it as this would pass the trait along. Deaf
white cats are banned from exhibition or breeding by some fancies in
Europe and there is a move to
reduce or eliminate this trait from
British breeding lines of various breeds. Deafness can cause problems
because a cat cannot hear
danger approaching. It can cause problems
to breeders because deaf female cats cannot hear their kittens crying
out and may neglect them. Deaf kittens cannot hear their mother
calling to them and may get lost. Deaf cats also seem to have no
volume control when meowing. For more information see
Living
With a Deaf Cat.
NOTES
1. THE "ALBINO SIAMESE"Feline
geneticist Don Shaw, writing in Cats Magazine, September 1972 to
February 1973, looked into the so-called Albino Siamese and wrote
that not all so called Albino Siamese were true genetic albinos and
that many are blue-eyed whites of Siamese type (what we now call
Foreign Whites). However, some Albino Siamese had pinkish-blue eyes
rather than the clear blue of other blue-eyed white breeds. His
research found that early registrations of "Albino Siamese"
mentioned "Chinese Whites" - reportedly cats of Asian
origin with Malay type and apparently blue-eyed whites having the
piebald (white spotting) gene. Unlike other albino animals, the cat's
eye structure always gave bluish tone, even to the pink or ruby eyes
normally associated with albino animals; this made true albinism in
cats difficult to detect.
A
mutation for "true albinism" was reported when a Chocolate
Point female bred to her Chocolate Point son. Most of the early
breedings to produce these reported "Albino Siamese" were
within the same Chocolate Point line, meaining the cats were
relatively inbred. It had always been suggested that the mother/son
mating resulted in doubling up of a recessive allele for albinism,
however Shaw challenged this theory.
A
Chinese White, and thus white spotting, had been introduced into the
Chocolate Point Siamese line and this Chinese White may have carried
both white spotting and albinism. If so, the apparently true albinism
in Albino Siamese would have been introduced from outside and not
occurred as a mutation within the breed. Alternatively, he had noted
that Chocolate Point and Lilac Point Siamese seemed to have a pink
glow to their otherwise blue eye which wasn't seen in Seal Points or
Blue Points.
Shaw
believed the
supposed Albino Siamese were blue-eyed whites resulting
from the white spotting inherited from the Chinese Whites PLUS the
pinkish tone due to being homozygous Chocolate Point Siamese (this
would be masked by the white spotting) - this combination mimicked
true albino. Since the alleged Albino Siamese resulted from
inbreeding Chocolate Point Siamese, so Shaw reasonably expected the
offspring to have the same pinkish-blue eyes he had observed in
Chocolate Points along with a double dose of white spotting that
produced a wholly white coat.
The
possible alternatives presented by Shaw were that Albino Siamese
might be true genetic albinos or they might simply be blue-eyed
whites with pale blue, slightly pink-tinted, eye colour. He didn't
have enough breeding data to determine which, but there was enough
data to demonstrate the presence of white spotting genes in that line
(white toes, lockets etc). Since Shaw’s time, true albinism has
been seen in cats (the eyes being pink with only a slight blue tint).
It has been recorded in the wild in the Asian Leopard Cat and also in
the domestic descendants of Leopard Cat hybrids, the Bengal.
Albinism
has spread from the Siamese to closely related Oriental breeds. Emma
Stothers sent me this report of her albino cat in 2013. Pangur is an
oriental longhair with very pale skin and nose. Her paw-pads and
ears, are pink-tinted white threaded through with red veins. Her eyes
look like faded lilac-blue (Emma describes it as an Elizabeth Taylor
colour) and when you look closer, you can see that the pupil is
a dull red, while what looks like lilac is just the pale blue
refraction of the lens overlaid on more red. You can very clearly see
the blood vessels beneath the iris which can be a bit creepy, because
when the light hits Pangur in the right way, her
entire eye turns a
muted pinkish red. In the dark, they reflect bright red.
Pangur
is an indoor cat, and has only been into the backyard under
supervision. In direct sunlight, she's unable to open her eyes, and
has to squint through little slits. She also squints when looking out
of
windows . She's very sensitive to light, and hates being
photographed using flash. In contrast to her poor daytime vision, she
can see very well in the right lighting. Her hearing has been tested
by the vet and is perfect. Emma has checked Pangur’s pedigree and
found inbreeding. Pangur has two grandmothers, but only one
grandfather , which means her parents are half-siblings. It seems
likely that the grandfather passed on his recessive albinism gene to
Pangur’s parents, and Pangur inherited two copies of the gene. One
of her sisters appears to be albino, but her other siblings have
neon-bright cyan eyes (i.e. the blue colour normal for foreign white
cats).
NOTES
2. OTHER BLUE-EYED CATSNot
all blue-eyed cats are white, nor are all blue-eyed non-white cats
Ojos Azules. The blue-eyed trait turns up surprisingly often in
random-breeding cats. Below: silver tabby female feral cat,
black-and-white male, red tabby (with white bib) male, red tabby
longhair male, odd-eyed tortie, black-and-white male.
CURLED,
CURVED AND FOLDED EAR CATS
Copyright
2002-2011, S HartwellCats
generally have prick (upright ears) which are triangular in shape.
The size, set and exact shape varies from the small ears of Persian
cats to the "bat-like" flared ears of some Siamese cats.
Some breeds require the base to be more widely flared than others.
Some breeds have ears placed close together high on the head, others
have wide-set ears with a broad expanse of forehead between. The tips
may be rounded, pointed, tufted or fringed depending on the breed.
There
is also a rare genetic condition causing the cat to have a second,
smaller, pair of ears behind the normal ears. These are apparently
non-functional and may be no more than ear flaps with no middle or
inner ear section. Though affected cats are generally physically
healthy, some have reportedly been lethargic or sluggish which
suggests that the condition might affect the brain. This rare
condition must not be confused with
accounts of "four-eared
cats" from China - these were Persian-type cats where the fur
inside the ear gave the impression of extra ear-flaps.
In
addition to variations on a general theme, there are two distinct ear
shapes - folded forwards and curved backwards. Both mutations (or
very similar mutations) have occurred more than once.
FOLDED
EARSThe
Sumxu (Chinese Lop) is now regarded as extinct, but was once found in
the area around
Peking , China. Descriptions of the Sumxu as
longhaired cats with glossy black or yellow coats and pendulous ears
derive from a series of mistranslations and the confusion of two
entirely different animals! A Polish Jesuit missionary to south
China, Michael Boym (ca. 1612-1659), first described the Sumxu in his
illustrated book Flora Sinensis (1656) in which he also described
Chinese fauna. "Sum Xu" was the Portuguese rendering of
songshu, meaning "
pine rat". Though Boym's illustration
resembles a squirrel, the description indicates the Yellow Throated
Marten (Martes flavigula) found in the region. Boym wrote that the
Sumxu was a pretty yellow-and-black animal that was commonly tamed
and wore a silver
collar . They were valued as hunters of mice and
sold for up to 9 silver coins (indicating their usefulness or
rarity).
The
cat-like Sumxu was described in early 1700s as a curiosity, and in
1796 when a droop-eared cat brought back from China. But how did the
Sumxu name end up attached to a variety of cat?
This
engraving is from German Jesuit Athanasius Kircher's book "China
Monumentis, Qua Sacris qua Profanis" (1666). The book is written
in
Latin and describes the Sumxu as being like a cat. The engravings
would have been done from descriptions or rough sketches rather than
from life. Kircher was a scholar rather than a missionary and he
relied on the expert knowledge of those who had travelled to China,
such as Boym and one of Kircher's
former students,
Martino Martini .
Kircher's description of the sumxu referred to it as a domestic
animal similar to a cat (in Europe, martens were also known as "tree
cats"). He described it as black and saffron coloured with
splendid hair. Following Boym's description, Kircher also mentioned
that the Chinese tamed this creature and put a silver collar around
its neck. It was an avid hunter of mice and was rare and valuable.
The creature depicted, while similar to Boym's illustration, was the
size of a small bear and is shown chasing deer, rather than mice! In
this it resembles a wolverine, rather than a marten.
Italian Jesuit Martino Martini had visited China in the 1650s and published
Novus Atlas Sinensis in 1655. In the section on Peking Province,
Martini described a
race of white, long-haired and long-eared cats
found in the region. He wrote that the cats in that province were
white all over with long hair and long ears like lap-dogs and that
these well-fed cats did not
catch mice, but were
ladies ' lap-cats.
Martini became the definitive source of information about China and
his description was used in 1673 by John Ogilby and by others who
described the long-eared, milk-white cats as being companions and not
hunters (there being other cats that were good mousers). The cats
were likened to the Maltese lap-dog and to spaniels. It retrospect,
it is likely that these white cats were also blue-eyed, a combination
of traits associated with deafness in cats - quite likely they
couldn't hear the mice in order to investigate mouse bolt-holes! If
this was the same mutation as the
Scottish Fold , it is associated
with sometimes crippling changes to the tail, hind legs and vertebrae
which would make an affected cat reluctant to move around.
In
1736-37, Martini's work was superseded by that of French Jesuit du
Halde's
works on the Chinese Empire. This compiled
material from
French Jesuits and re-used some of Martini's book, including the
description of the long-eared white cats:
"Among
the animal species one finds singular (i.e. unique) cats which the
Chinese ladies seek out to provide them with amusement and feed with
plenty of delicacies: they have long fur and drooping ears." This
remained the standard reference book for many years and was reused by
other authors, including the passage about the lop-eared cats. For
example John Green's book described the lop-eared cats of Peking
Province as "a particular Sort of Cats, with long Hair, and
hanging Ears, which the Chinese Ladies are very fond of." This
was later
translated back into French by abbé Prevôt as
"among
the animals one finds a singular species of long-haired cats with
hanging ears, which the Chinese ladies greatly liked."In
volume 4 of his "Histoire Naturelle" (?1767), Georges Louis
Leclerc,
Comte de Buffon, wrote "The Natural History of The
Cat". Buffon noticed that, unlike the many breeds of dog, there
were few distinct races of domestic cat: Spanish, Syrian and Khorasan
(Persian). He believed the long-haired lop-eared Chinese cat could be
a fourth distinct type.
"They
(cats) are in nature much more constant , and their domesticity is not
as complete nor as universal as that of the dog, so it is not
surprising they are less varied in type. Our domestic cats, though
different from each other in colour, do not form distinct and
separate races. Only the climates of Spain , Syria or Chorazan
( Persia ) have produced constant varieties; to find another, one must
join the climate of the province of Pe-chi-ly ( Northern Zhili) where
there are cats with long hair and hanging ears, much liked by the
Chinese ladies." Buffon
was translated into English in 1781 by William Smellie. According to
Buffon,
"Our
domestic cats, though they differ in colour, form no distinct races.
The climates of Spain and Syria have alone produced permanent
varieties: To these may be added the climate of Pe-chi-ly in China,
where the cats have long hair and pendulous ears, and are the
favourites of the ladies. These domestic cats with pendulous ears, of
which we have full descriptions, are still farther removed from the
wild and primitive race, than those whose ears are erect ."In
a supplement, Buffon added that there was some doubt as to whether
the Sumxu was a cat or some other animal:
"I
formerly remarked, that, in China, there were cats with pendulous
ears. This variety is not found any where else, and perhaps it is an
animal of a different species; for travellers, when mentioning an
animal called Sumxu, which is entirely domestic, say, that they can compare it to nothing but the cat, with which it has a great
resemblance. Its colour is black or yellow, and its hair very bright
and glittering. The Chinese put silver collars about the necks of
these animals, and render them extremely familiar. As they are not
common, they give a high price, both on account of their beauty, and
because they destroy rats."Buffon's
source was Prevot, whose source was Green, whose source was de Halde,
whose source was Martini's 1655 work. Buffon noticed a process we now
call neoteny - domesticated animals
retain juvenile features into
adulthood. Thus wolves ahd pricked ears and domestic dogs often had
softer, hanging ears. Buffon believed this could apply to cats as
well and that the mild climate and ancient civilisation of China was
conducive to domestic cats developing a similar trait to domestic
dogs. In his work, Buffon also mentioned the sumxu as a pretty
domestic animal of China, not unlike a cat. In his Vol VIII
(published 1777), Buffon goes so far as to suggest the lop-eared cat
of China is a different species from the ordinary domestic cat and
that it might therefore be the cat-like sumxu:
"We
said (volume VI, page 14) that China has a race of cats with hanging
ears; perhaps this variety is found nowhere else and is a species to
the (domestic) cat, because Travellers speak of an animal called the
Sumxu, which is completely domestic in China and which is in many
respects like a cat. In colour it is black and yellow with extremely
glossy fur. The Chinese give them silver collars around their necks
and make them very tame . As they are uncommon, they are expensive;
not only because of their beauty, but because they most cruelly wage
war upon rats."This
seems a big leap (and an erroneous one) from Boym's black-and-yellow
squirrel-like creature to Martini's white lop-eared cat, however du
Halde had omitted to
mention in his
compilation that the lop-eared
cats were milk-white! While Boym's sumxu was valued as an avid
mouser, Martini's lop-eared white cats were not interested in mice
and were kept as lap-cats, another piece of information du Halde
omitted in his compilation. Boym's illustration of the sumxu ignores
its ears, while Martini described them as the defining feature of the
white cats. Boym didn't liken the sumxu to cats and it is Kircher who
made that comparison. The sumxu (probably the Yellow Throated Marten)
was a creature of south China while the lop-eared cats were from
Peking Province in the north. Buffon's leap was further
misinterpreted by later authors such as French
writer Anselme Gaëtan
Desmarest who combined the name "sumxu" with the
description of the lop-eared cat (its white colour being unknown to
him) as
"hanging-ear
cat, fur long and fine, black or yellow, domesticated in China in
Peking province (Pe-chi-ly) under the name of sumxu". This
was perpetuated through the
19th and early
20th centuries, especially
by cat fanciers looking for new and exotic cats to
import .
When
China reopened to foreigners after the Opium Wars, a number of
missionaries, entrepreneurs, cat fanciers and researchers went
looking in vain for the lop-eared cat. In his book "Variation of
Animals and Plants Under Domestication" Charles
Darwin referred
briefly to a drooping eared race of cats in China. In "The Cat"
by Lady Cust (1870) it states "Bosman relates that in the
province of Pe-chily, in China, there are cats with long hair and
drooping ears, which are in great favour with the Chinese ladies;
others say this is not a cat but an animal called 'Samxces'"
In
his 1885 book "The Cat: Natural History, Husbandry ["Hygiene"]
and Illnesses" Gaston Percheron suggested the lop-eared cat
might be a hybrid between the cat and a marten:
"Certain
respected naturalists even claim even that the [domestic cat] mates
with the marten and produces among the hybrids those like the latter in colour and fur. In this way they explain the hanging-ear cats of
China [...]" but instead of being fed delicacies by its
mistress, the cat has become a delicacy to be eaten, "The
Chinese Cat. It has long, silky hair and hanging ears, like those of
a badger. Its flesh is highly esteemed by the inhabitants of the
Celestial Empire. As with the dog, it is treated with great
solicitude by feeders and fatteners of this country and when it is
fattened, it appears alongside swallows' nests on well-served tables.
" As
a side-note, in 1926, Siamese cat fancier Lilian J Veley remained
adamant that Siamese cats were not merely a form of domestic cat, but
resembled an unknown type of viverrine that lived in the region (the
most likely contender again being the Yellow Throated Marten) in
their sable-and-dun colour and "marten-like" face! Maybe
"Sumxu" and "Siam" sounded similar to her ears.
The
Chinese, or Hanging Ear, Cat according to "Die Hauskatze, ihre
Rassen und Varietäten" (Housecats, Their Races and Varieties)
from "Illustriertes Katzenbuch" (An Illustrated Book of
Cats) written and illustrated by Jean Bungartz, published in
Berlin in 1896 (There is no published English translation of this book, this
gives the
gist of the text)
The
Chinese or Hanging-Ear cat is most interesting, because it provides
proof that by continual disuse of an organ, the organ atrophies. So
with the Chinese cat the hearing and/or the ears have deteriorated.
Michel says the Chinese, not only admire the cat in porcelain, but
also value it for culinary reasons. The cats are regarded as special
titbits and enjoyed particularly with chains (noodles?), with rice".
This cat is bred particularly for the purpose of meat production, and
is a preferred Chinese titbit; this is not unusual if one considers
that the Chinese consume much the sight of which turns the stomachs
of Europeans. The poor creature is locked up in small bamboo cages
and much like a kind of geese fattened with plentiful portions.
Extensive trade is carried on with other parts of Asia and the
Chinese allow no tomcats to be exported so there is no interference
in this lucrative source of income.Due
to the restrictive conditions that have deprived the cat of its
actual use, its hearing decreased because it was no longer needed as
for hunting its own food. With no need for watchfulness, it was
useless to have sharp hearing to listen for hidden things so the
hearing became blunt and in natural consequence the ear lost its
upright nature, gradually become lower and becoming the hanging ear
that is the characteristic feature of the Chinese cat. At first
impression this is a surprising and amusing look, but is impression
is lost with closer examination. If one ignores the characteristic of
the ears, one sees a beauty similar to the Angora cat: a long, close
coat of hair, albeit less rich, covers the body. The hair is
silky- soft and shining and the colour is usually a light yellow
(isabelline) or a dirty white yellow, although some have the usual
colouring of the common house-cat. In size it considerably exceeds
house-cats and is stronger. The ears hang completely, as with our
hunting dogs and are large in relation to cats.Although
the Chinese cat is found in considerable numbers in its homeland, it
rarely arrives at European animal markets. Only one such cat has
reached us in the flesh; we acquired this years ago when a sailor
returning from China brought it into Hamburg . The accompanying
illustration is based on this. In character it is like the Angora cat
and somewhat inactive. It also prefer to live by a warm fire, is a
little sensitive to attention , hears badly and is at its most
animated when it sees the milk or food. Apart from its unusual ears,
it does not have any really attractive characteristics and is a
strange representative of the house cat.Though
reports refer to the Chinese Lop having pendent or pendulous ears
(suggesting abnormally long or
floppy ears e.g. like a labrador dog)
this is probably an exaggeration. In all likelihood, and in the
absence of any current examples or pictorial evidence, the ears were
folded in a manner similar to the Scottish Fold. In Frances Simpson's
"The Book of the Cat" (1903), contributor H C Brooke wrote
"There is said to be a variety of Chinese cat which is
remarkable for its pendent ears. We have never been able to ascertain
anything definite with regard to this variety. Some years back a
class was provided for them at a certain Continental cat show, and we
went across in the hope of seeing, and if possible acquiring, some
specimens; but
alas the class was empty! We have seen a stuffed
specimen in a Continental museum, which was a half long-haired cat,
the ears being pendent down the sides of the head instead of erect;
but do not
attach much value to this."
In
1926, Brooke wrote that "for donkey's years" Continental
cat shows had offered prizes for the
Drop -eared Chinese Cat. On each
occasion, the cat failed to materialise and Brooke considered it to
be mythical. Other writers suggested it was the result of haematomas
causing the ears to fold or crumple. Brooke noted that although
no-one ever saw the cat itself, one always met "someone who
knows someone whose friends has often seen them". He had been
assured by a Chinese gentleman he had met only once that "he
knew them well". HC Brooke, and other fanciers, made enquiries
of the Chinese Embassy, of Hagenbeck's (a major Hamburg animal dealer
at the time) and of a "certain well known author, who has lived
for years in China and knows that country well", but to no
avail. The American
Express Company had instructed their
representatives at Shanghai and Peking to make enquiries, again
without success. None of the wild animal dealers knew of the Chinese
Lop.
The
German naturalist, Brehm, had given a very detailed description of
the cat in the 1700s. Brehm was usually very accurate. In 1882,
Brooke had seen a stuffed specimen in a Continental museum. The
specimen was "half-coated with yellowish fur". He admitted
that it might have been a
fake or a cat with its ears deformed by
canker (i.e. cauliflower ears) that had been presented in all good
faith. All avenues of enquiry exhausted, Brooke declared the Chinese
Drop-eared cat extinct. The last reported sighting of the Chinese Lop
seems to have been in 1938 when a droop-eared cat was imported from
China. On that last occasion the mutation was thought to be
restricted to white longhaired cats. It is hard say for certain
whether these were isolated cases or whether the Chinese Lop was a
genuine variety. It cannot even be said with certainty that the trait
was an inherited one.
Cats
with folded ears have been reported in the Hebrides, Germany and
Belgium, but were apparently regarded as no more than isolated
curiosities and not bred.
The
only widely recognised fold-eared cat is the Scottish Fold and its
longhaired version, the
Highland Fold (Coupari). These have ears
which form a close-lying cap. Also known as Scottish Lop, the
Scottish Fold traces back to a female white shorthair
farm kitten
discovered near Coupar
Angus , Perthshire, Scotland in 1961. This cat,
Susie, produced some fold-eared kittens. One of these, Snooks, became
the founding mother of the breed after Susie's unfortunate death in
1963. By 1967, numerous "Lop-eared Cats" had been born
however the UK cat registries declared the trait to be a deformity
and refused to accept the "Scottish Fold" for breed status
because of potential ear problems (an inconsistency considering they
recognised the tailless Manx). Interest in Scottish Folds in the USA
led to breeding being continued there and it was recognised as a
breed in 1973. The cat does not suffer undue ear problems and one
British registry accepted it in 1983.
The
Coupari is the name given to Longhair Folds in the UK, although these
are known elsewhere as Highland Folds. The argument is that Coupar
Angus is not in the Highlands, however Highland Fold is no more
inaccurate than other breed names e.g. Balinese, Tonkinese.
Longhaired cats were present in the original Scottish Fold, but the
shorthaired variety was preferred because the ear shape was more
visible. It was developed in the 1980s and recognised in 1986. It is
known by various names: Coupari (in UK), Highland Fold, Longhair Fold
and Scottish Fold Longhair.
Kittens
are born prick-eared and start to develop the folding at around 4
weeks old. The fold is fully developed at around 3 months of age. The
gene causing the folded ear trait is a dominant gene which causes
skeletal problems if the cat inherits two copies of the gene.
Scottish Folds are therefore always bred to prick-eared to keep the
incidence of problems to a minimum. The problems are a thickened tail
caused by tail vertebrae fusing and thickened legs with swollen feet
due to overgrowth of cartilage around the paws. These side-effects
cause problems with walking.
Prick-eared
Scottish Folds gave rise to the Scottish Shorthair in Australia. This
breed (with longer tail and legs, and different coat texture to the
British Shorthair)was recognized and named by the
Queensland Independent Cat Council (
QICC ) as a result of a submission by Rona
Sandilands at a
meeting at then QICC-
president Pat Mercer’s home
attended by prominent Scottish Fold and British breeders, including
Lillian Carter founding breeder in Queensland. The first Scottish
Shorthair exhibited was Ptah Dunstan Gold, bred by Lillian Carter and
owned by Rona Sandilands. During the 1980s he was exhibited at the
Queensland Longhair Cat Club show, achieving a Reserve in Group 3
under Victorian Judge Marie Orchard. There is also a Scottish
Longhair.
The
Poodle Cat (Pudelkatze) is essentially a breed developed from the
Scottish Fold and Devon Rex to create a curly coated fold-eared cat -
a curly-coated Scottish Fold or fold-eared Devon Rex. It was
developed initially in Germany where its future is threatened by
rulings prohibiting the breeding of cats with harmful defects. This
ruling affects Scottish Fold cats because of the skeletal
abnormalities which can occur. The breed is attractive and if
breeders are careful to breed only from healthy cats, there is no
reason it should not be accepted. Astonishingly, there appears to be
the intention of adding Manx into the mix so that the cats are
tailless as well. This would create a more dangerous mix of
semi-lethal genes since the Manx condition can cause other skeletal
abnormalities.
Back
in 1981, Phyllis Lauder wrote in "The British, European and
American Shorthair Cat" wrote of news from Australia of
"interference not beneficial to the domestic shorthairs". A
correspondent, Mrs Batten, had been asked for her views on the idea
of crossing Manx with Scottish Folds. Scottish Folds had not yet been
recognised. Lauder wrote that the idea was born of "love of
change for its own sake and by the
desire to meddle" since a
Manx/Fold cross would not only produce a freakish-looking cat, it
would be a tragedy for the cats - the two breeds had enough to
contend with in refuting charges of deformity.
The
unrecognised Oriental Fold is a Siamese/Oriental type cat with folded
ears (I believe it came from attempts to breed colourpoint Scottish
Folds). The
Hemingway Fold is an unrecognised variety of fold-eared
cat with extra toes. The term "Hemingway" is often used in
America to describe polydactyl cats (mitten cats). It results either
from a spontaneous folded ear mutation in a polydactyl cat or from
accidental or deliberate crossed of Scottish Folds and polydactyls,
possibly for curiosity or as attractive pets.
A
fold-eared cat aiming for recognition is the Foldex or Exotic Fold.
The Foldex is a cat of Exotic (shorthaired Persian) body type
developed through crossing Scottish Folds with Exotics. It has a
shorter nose than the Scottish Fold but not as short as the Exotic
Shorthair. In other respects it is a fold-eared version of the Exotic
Shorthair. Facially, the Foldex looks like "a little furry owl".
Like the Scottish Fold, it is prone to the same bone deformities and
produces some non-folded offspring.
In
2005, the Ukrainian Levkoy Cat, a fold-eared naked cat, was created
using the Don
Sphinx and Scottish Fold. The Ukrainian Levkoy is less
extreme in body and face type than the Don Sphinx (the face is wider
and ronder) and the ears do not fold tightly to the skull as in the
Scottish Fold, but stand out from the head and fold closer to the
tips. It also occurs in velour and prick-eared forms.
In
1986 there was a report of a "Hebridean Cat". The owner
said that the
tiny ears were a feature of Hebridean and in addition
the ears were folded like those of a Scottish Fold. No more was heard
of the Hebridean breed.
In
the mid-1990s, a fold-eared black cat turned up in a feral cat colony
in Essex, England. It was not possible to determine whether the trait
was inherited or was a birth defect. It is extremely unlikely to be
due to a free-ranging Scottish Fold as these are uncommon in Britain
- there would certainly be no free-ranging unneutered Scottish Fold
males around.
In
2007, Vasilis Lekkas from Athens (who is deeply involved in the
recently formed
Greek network for the preservation of domestic
livestock including cats) reported possible folded ears on the Greek
island of Myconos, but these may have been due to injury.
Lizzie
Ellis (The Feline Rescue Association Inc, Maryland, USA) provided
this photo of a folded ear apparently caused by frostbite. The ears
had to be surgically removed. This shows how inury, including
haematoma ("cauliflower" or collapsed ears from fighting)
and frostbite can mimic folded ears at a casual glance.
CURL -EARED
CATSThe
first recognised curl-eared breed was the American Curl. The ears are
curled or swept back on themselves and feel rigid to the
touch . The
originate from a stray kitten, Shulamith, in 1981. Shulamith was a
black longhair female with strangely curled ears. She went on to
produce kittens, some of which also had curled ears. These attracted
attention when shown at a cat show in 1983 and in 1985/6 the American
Curl breed was recognised.
The
mutation is a dominant gene so litters will often contain a mix of
curl-eared and prick-eared cats. It is impossible to tell which
kittens will develop a good curl - they are all born prick eared. The
ears curl up tightly over the next few days. During the next four
months, the tight curl starts to relax until the final semi-curled
state is reached. As far as is known, the gene does not cause
detrimental side-effects.
The
Hemingway Curl is a localised variety rather than a breed.
The
first Hemingway Curl appeared as a spontaneous curl-eared mutation in
a colony of polydactyl cats known as Hemingway cats (after the
polydactyl cats of Ernest Hemingway) on Key West. Hemingway Curls
have since been crossed with African Jungle cat hybrids to produce
the Jungle Curl breed. It occurs in shorthair and longhair versions
and may
attract sufficient interest for a breed to be established.
Having
established the curl-eared trait, it became possible to cross
American Curls with other breeds to create new varieties e.g. the
Kinkalow which is a mix of American Curl and Munchkin to produce a
short-legged curl-eared cat.
The
Ruffle was, according to one source, an accidental side-effect
resulting from attempts to improve the ear size of American Curls by
introducing the Cornish Rex into a breeding line. Whether the Cornish
Rex is an allowable outcross for the American Curl is another matter
entirely. According to another source, the original mating was a
deliberate attempt to produce a new breed. Progeny from this breeding
line was apparently acquired by a breeder unaware of the Curl-to-Rex
matings which had the side-effect of introducing the recessive gene
for curled fur. The gene remained hidden until two carriers were
mated and produced curly coated American Curls some generations
later. The rippled effect of the fur reminded the breeder of a brand
of potato snack hence the name Ruffle. Unfortunately the breeder's
circumstances changed and further development of the Ruffle breed
could not be funded. Interestingly, the Rex coatd in American Curls
was mentioned as early as 1991 by feline geneticist.
An
experimental breed called the Jaguarundi Curl (shorthaired and
longhaired) was reported in 2001, but its existence is unconfirmed.
The Jaguarundi is a South American felid which is very un-cat-like in
shape. It is suspected that the reporter had confused it with the
Jungle Curl. The Jungle Curl does exist and is a hybrid of Jungle Cat
(F chaus) and American Curl or Hemingway Curl with Serengetis, Jungle
cat hybrids, pure Jungle Cats, Bengals, Egyptian Maus, Abyssinians
and other domestic shorthairs contributing their genes to the mix. It
is still in the developmental stage to produce wild looking cats,
preferably with a spotted or rosetted pattern, with curled ears.
Several breeders in Illinois are working with a curl-eared polydactyl
cat called Impians (formerly called Tulips). Impians were originally
developed in the 1990s by crossing American Curls with polydactyls,
creating a harlequin patterned semi-longhaired breed. The markings,
which can be any colour, are restricted to the head, down the spine,
shoulders,
hips and tail.
The
Australian Curl was a breed that never was, rather it was a single
female cat,
Matilda , which was discovered in 1996 and who failed to
pass on the trait. The curl of the ears apparently differed from that
of the American Curl though no precise details were provided. She was
mated to a normal eared cat of similar type, but none of the kittens
were curl-eared. She suffered severe illness after the birth (1997)
and could not be bred from again. No back-crossing could be attempted
to determine whether the gene was carried as a recessive. Although
none of the offspring had curled ears, they were not bred to each
other (or it was not possible to do so) to determine whether they
carried the trait was carried in recessive form. It is possible that
a breed opportunity has been missed due to Matilda's unfortunate
illness.
In
2007, Vasilis Lekkas from Athens (who is deeply involved in the
recently formed Greek network for the preservation of domestic
livestock including cats) reported "In the islands of Paxoi in
the Ioania Sea in western Greece, an otherwise conventional cat with
curled ears is common. Locals consider it their "normal"
cat and view as unusual cats of the mainland or other islands. These
cats tend to be white bodied with restricted coloured patches. It
appears the mutation may be similar to American Curl. There are two
Paxoi islands, but is unclear, which one the report relates to."
A young cat with a moderate curl has also been reported.
The
Elf breed was initiated in 2006 with the first full hairless,
curl-eared Elf being born in 2008. It combines the Sphynx
hairlessness with the American Curl's ear conformation. Permissible
out-crosses include non-pedigree domestic shorthairs. They have a
sturdy, athletic build, are sociable, intelligent, inquisitive and
people centred. Half-Elf cats e.g. furred Elf variants result from
outcrosses to diversify the gene pool.
The
Dwelf also combines curl ears with hairlessness, but adds Munchkin
and Highlander (a bobtailed, curl-eared polydactyl breed) to the mix.
These combine hairlessness, curled ears and short legs. Non-standard
(long legged) Dwelfs may resemble the Elf breed.
IRISH
SHORT-EARED CATDescribed
in a light-hearted book depicting cats which had been "painted"
with vegetable dye (in fact all painted cats were
photo-manipulations), the Irish Shortear is a mutant-eared cat that
never was. This footnote has been included because people have asked
where to find an Irish Shortear!
The
book in question was the spoof "Why Paint Cats" and the
fake breed among genuine breeds was the Irish Shortear. It was
described as Burmilla x Scottish Fold with large protuberant eyes,
short ears and very relaxed nature due its excellent vision. The
photo was that of a Brown Burmilla which was been edited to give the
cat larger eyes, narrower chin and short ears (the original ear tip
had been photographically enlarged and grafted back onto the face).
The face was then grafted back onto the image of the cat's body. One
giveaway was that the facial area had a "floating effect",
which is common in photo-composites. A Scottish Fold would have given
folded not shortened ears. In addition, the partial-dominant
"macro-retinal" gene mentioned in the text is fictional
(and it should have been a macropthalmia gene since the retina is the
back of the inside of the eye).
BOBTAILED
AND CURLY TAILED CATS
Copyright
2002 - 2013, S HartwellTHE
TAILThe
normal tail has 21 - 23 vertebrae on average with the normal range
being 18 - 28 vertebrae. It is on average 25 cm (10 inches) long, but
can range from 20 cm (8 inches) to 30 cm (12 inches) with a few
exceptional specimens having 35 cm (14 inch) tails
There
are various mutations affecting the tail and in 1940 American
zoologist Ida Mellen wrote of oddities in cats' tails including
kinked, bobbed, curled and even double tails. Curly tailed cats were
known in China in the 12th Century (probably bobtails) and ringtail
cats were known in the USA sometime prior to 1940. In 1868, Darwin
wrote in
The
Variation of Animals and Plants Under Domestication "Thropughout
an immense area, namely the Malayan archipelago, Siam, Pegu, and
Burmah, all the cats have truncated tails about half the proper
length, often with a sort of knot at the end."
Tailless
and cats have occurred periodically through random mutation. They
have been reported in
Bosnia , Burma, China, Crimea,
Java , Malaya,
Denmark, Nova Scotia and Thailand though some of these will have been
bobtails. Bobtail cats occur throughout Asia and into Russia, with
more recent mutations occurring in the USA. In a 1949 study, Searle
found no kinked tails in London cats. He studied Singapore's cats in
1959 and eventually concluded that stubby-tailed cats were common in
Singapore, but in general was rare in Europe except for the Manx
breed where the tail was often completely absent. The Manx and the
Asian bobtails are due to different mutations. Research indicates
that the “tailless gene” has 4 alleles i.e. there are four
different versions of the same gene. The Manx breed is based on one
of those mutations, and that mutation can have side effects such as
spina bifida. The PixieBob is based on a different mutation of the
same gene; that mutation has a different appearance from the Manx
mutation. The American Bobtail, which occurred spontaneously in a
different geographic area, may be based on the same, or a different
mutation of the same gene. The Kurilian Bobtail is due to an
incomplete dominant gene. However, the similarly named Karelian’s
bobbed tail is due to a recessive gene, as is the Japanese Bobtail.
BOBTAILED
CATSThe
bobtailed mutation is not related to the Manx mutation. It has
occurred independently in various geographic areas. It is widespread
in Asia and parts of Russia. An Abyssinian Bobtail has been reported
in non-pedigree Abyssinian-type cats (location not specified). A
Spanish bobtail has also been reported. Cats with knotted, shortened,
kinked and pom-pom tails are relatively common in Tenerife and the
Canary Islands, the mutation apparently having become established due
to the isolation of the gene pool (tourists frequently mistake the
trait for breakage). Similar mutation have occurred spontaneously in
the USA. There are also purported
Bobcat hybrids that have inherited
the short tail from the wild parent. According to leading feline
geneticists, the genes governing tail conformation are
located on a
mutation hotspot.
According
to an earlir anatomist, Sir Richard Owen, and to
Professor H N
Moseley, the kink in the Siamese's tail (a mild form of the bobtailed
trait) was the relic of a prehensile tail, possibly inherited from
civet ancestors (though neither civets nor genets had prehensile
tails)! Others asserted that the kink was due to intercrossing the
Siamese with the "common strain" however Lilian J Veley
wrote in 1926 that this could be discounted since there was no other
cat known in Siam, "common" or otherwise that had ever
possessed an original kink, making it a folly to try to eradicate the
trait. Presumably Ms Veley had not encountered the numerous bobtailed
and kink-tailed street cats in Thailand. In "Our Cats" of
May 1901 there was information on the Siamese's tail from interviews
with the King of Siam and his Private Secretary. While the cats in
the Royal Palace apparently had no kinks in their tails, occasionally
a "tramp cat of Malay origin" strayed in and the resultant
crossbred cats had kinked tails.
In
1783, Willian Marsden, Fellow of the Royal Society and late Secretary
to the
President and Council of Fort Marlborough wrote in "The
History of Sumatra" of the Malay Cat: "All their tails
imperfect and knobbed at the end." In 'The Variation of Animals
and Plants under Domestication", Darwin wrote "throughout
an immense area, namely, the Malayan Archipelago, Siam, Pequan, and
Burmah, all the cats have truncated tails about half the proper
length, often with a sort of knob at the end. […] The Madagascar
cat is said to have a twisted tail." Another writer and
traveller, Mivart, had corroborated the statement regarding the Malay
cat, of which he said the tail "is only half the ordinary
length, and often contorted into a sort of knot, so that it cannot be
straightened […] Its contortion is due to deformity of the bones of
the tail". Joseph Train had also mentioned th Malayan cats,
comparing them with the Manx: "The Manks rumpy resembles some
what in appearance the cats said by Sir Stamford Raffles to be
peculiar to the Malayan Archipelago." Sir Stamford Raffles' name
is closely associated with Singapore.
HC
Brooke reported seeing three spotted tabby Malay cats in
Holland in
the 1880s and some very similar specimens at the Jardin
d'Acclimatation in Paris around the same time. "The tails of
these [Paris] cats, about three or four inches long, were tightly
screwed, or at least the tail formed three complete revolutions. The
'
screw ' tail, as also the spotted type of colouration, appear to be
becoming very rare." In 1889, Mr O Gould had apparently taken
four Siamese to Ceylon and soon Ceylon was "overspread"
with their progeny, known as "Gould's Cats" and many had
kinked tails in spite of there being no Malay cats for them to
interbreed with. Lilian J Veley wondered if the Malay cat's kink had
come from the Siamese rather than the other way round (she considered
the Siamese to be the older type). She also implicated the Korat "a
blue variant of the Siamese, to which our 'blue-pointed' freaks are
due."
Jean
Bungartz described several bobtailed cats in his 1896 book "Die
Hauskatze, ihre Rassen und Varietäten" (Housecats, Their Races
and Varieties) in " Illustriertes Katzenbuch" (An
Illustrated Book of Cats). He noted that according to Brehm Martens,
on the Sunda Isles and in Japan there were cats with different
lengths of tail. Kessel had told Weinland that there were
short-tailed cats on Sumatra. Bungartz added that the cats of
Cochinchina had only a short, curled tail and the Madagascar cat had
a turned, knotted tail.
R
Shelford, former Curator of the Sarawak Museum wrote in his book "A
Naturalist in Borneo" "It may be mentioned here that the
domestic cat of the Malays is quite a distinct variety [...] it is a
very small tabby with large ears and a body and hind-legs so long
that it lacks all grace. The tail is either an absurd twisted knot or
else very short and terminating in a knob; this knotting of the tail
is caused by a natural dislocation of the vertebrae so that they join
onto each other at all sorts of angles." The length of hind-leg
was a trait shared by the Manx, leading some cat-fanciers to believe
that the two were related.
Miss
Lowndes, daughter of the novelist Mrs Belloc Lowndes described a
Malay kitten that she had acquired. It had recently arrived, along
with its mother, from the Straits Settlements. "It has a
triple-kinked tail. It is, unfortunately, not of the spotted kind,
but these seem to be very rare nowadays." More information was
provided by Mr Boden Kloss, Director of the Raffles Museum and
Library at Singapore "The tail which distinguishes these cats
may be clubbed or kinked, very short or of medium length, and the
animals themselves of many colours - plain, piebald, or patterned."
He also wrote "A fair proportion of the cats of Singapore seen
in native villages are short-tailed animals with a kinked tail. There
would [be], I should say, three or four kinks. In colour they may be
tabby, or boldly black and white. As a point of interest it may be
noted that
Felis planiceps [
Flat -Headed Cat], one of the wild species
of the peninsula, tends to resemble the domestic Malay cat in the
matter of tail." The cat writer HC Brooke, who had an interest
in the Malay cat, wrote that F planiceps and the domestic cat were
unlikely to be
inter -fertile.
Mr
H O
Forbes had exhibited a bobtailed Malay cat to the Liverpool
Biological Society and shown the cause of the knotting to be the
development of wedge-shaped cartilages between the tail vertebrae.
Forbes attempted to link the Malay bobtails to the bobtailed cats
found in part of
Portugal . In the 1920s, Forbes wrote "My
remarks referred to the interest I had in exhibiting the creature's
skin from the occurrence in the East of what I had noted as extremely
common in the cats of Portugal when I lived there about 1876. The
kink, I was told was then believed to have become hereditary, from a
custom long practised by the Portuguese of pinching or breaking the
tails of the new-born kittens, and it would be of special interest if
it could be established that the kink in the Malayan cats' tails had
been communicated to them through those imported by the early
Portuguese into the East. If I can trust my memory the tail of this
cat, though short and kinked had the full number of vertebrae, some
of them reduced and wedge shaped." Others disputed his theory as
the trait had been reported in the Malay cat since at least 1783 and
no amount of tail-pinching would cause the trait to become
hereditary!
Kuantan, Malaysia. Feral male with perfect pom-pom tail.From Lake Chini, Kuantan area, local domestic male with bobtail and extra toes (not visible). His offspring ranged from bobtail, kinked tail to two-thirds normal length tail. The
bobtail trait ranges from a normal-length tail with a distinct kink,
through to a short twisted pom-pom and just about anything between
those two extremes. The degree of kink is variable and the vertebrae
are affected so that the tail cannot be straightened. It is often
possible to feel a bony knot inside the kink where vertebrae have
fused. The bobtail mutation is widespread throughout Asia, extending
as far as Russia. It is well know that early Siamese cats had kinked
full-length tails and this is still seen in Siamese-type cats in
Thailand, but has been bred out of pedigree Siamese cats.
Colourpointed cats were kept by Thai royalty and legend has it that a
princess entrusted her rings to a palace cat while she bathed. She
threaded them on the cat's tail and the cat knotted its tail so the
rings did not fall off. The kink therefore marks where it knotted its
tail. Unlike the Manx mutation, there appear to be no detrimental
effects.
The
Madura or Buso cats are a closed colony of bobtailed blue cats with
green eyes which live on the tiny and isolated island of Ra'as off
the coast of the Indonesian island of Madura (it is a 6 hour ferry
ride from Madura to Ra'as). There are reckoned to be less than 100 of
these cats and they are in danger of extinction, in part due to local
beliefs that prevent the exportation of fertile cats. The Madura
breed and its decline have been documented by Dr Ronny Rachman Noor
(Faculty of Animal Science, Bogor Agricultural Universtity). The cats
are known to the islanders as "Kucing Buso" (grey cat) and
the true Madura cat is blue self, suggesting they may have come from
Korat stock taken to the island on trading ships. There is some
mythology surrounding the breed, just as there was about the "Royal
Siamese". The true Madura cat, originating from Ra'as, may only
be kept by high-ranking people such as religious leaders, high rank
government officials and informal leaders. Anyone attempting to
smuggle a cat from Ra'as will find their boat sinking. The cats'
posture and the triangular facial shape are similar to the wild cat
and leopard. The cats are large with a medium length tail with a
visible bend or kink at the end. The fur is relatively thick. The
true Madura cat is solid grey known as "buso" often with
lavender nose leather. A few so-called Madura cats on Madura can be
found with the brown sepia, mink and colourpoint patterns as well as
blue bicolours due to matings with local cats. A brown mink "Madura
cat" exhibited at a cat show in Surabaya was of Japanese Bobtail
type. The declining numbers appear due to people trying to export the
cat out of the island and to a high mortality rate among breeding
cats, both on Ra'as and on Madura, indicating either a lethal gene or
a high degree of inbreeding. Cats brought out from Ra'as are
castrated due a local belief that the fertile Buso cat may only be
kept at the Ra'as island (castration in itself may
pose dangers of
infection). To preserve this cat as a distinct type and avoid
extinction, breeders would need to establish breeding lines which
would mean obtaining unneutered cats. The restriction on who may own
a true Madura cat also limits ownership and controlled breeding.
There
are various
tales associated with the Asian bobtailed cats. The
Siamese tail kink, for example, was supposedly due to a princess
threading her rings on the cats' tails for safekeeping and the loyal
cats kinked their tails to keep the rings in place. Siamese were also
believed to be royal palace cats. Likewise Madura cats may only be
owned by people of high status. A Malaysia belief is that if a
kittens tail is cut off and buried under the doorstep, the cat will
not stray from home. Another is that monks cut the tails off of cats
so that the cats do not go to heaven. A cat with a stumpy tail is not
perfect and imperfect creatures cannot go to heaven. I examined a
tabby cat brought back from Japan whose owner claimed that its tail
had been cut off by monks during kittenhood. I found evidence of
knots and kinks in the remnant of tail which told me it was a natural
bobtail. Also, the tail ended quite normally in a black tip. This was
a perfectly normal genetic bobtail, though the owner preferred to
think she had rescued an abused cat.
In
1988, the Cat Association of Britain finalised the standard for the
"Oriental Bobtail"; a cat of oriental (or foreign)
conformation and coat, but with a bobbed tail. Since then, little or
nothing has been heard of this breed.
The
most famous bobtailed breed is the Japanese Bobtail found in both
shorthair and semi-longhair varieties. This is due to a recessive
gene. It has appeared in ancient Japanese art and has evidently
existed back as far as the 6th century. It is claimed that it was
introduced into Japan from China at that time, which corresponds to
Ida Mellen's account of curly tailed cats in China. Once the pet of
Japanese nobles, it eventually spread to the general population but
was not considered anything more than a common moggy. It attracted
the interest of American breeders in 1968 and was recognised in the
US in 1978. The longhaired version had always existed (longhair is
due to a recessive gene) but was not given breed status until 1991.
In
a study comparing the genetics of several breeds (Lipinski MJ, et
al., The ascent of cat breeds: genetic evaluations of breeds and
worldwide random-bred populations, Genomics (2007), the Japanese
Bobtail was found to be more closely related to Western cats than to
Asian cats, though the breed showed some Asian genetic influence.
Cats are not indigenous to Japan, but reached the islands along Asian
trade routes hundreds of years ago. However, the Japanese Bobtail was
imported into the USA in 1968 and developed as a breed in the USA. As
a result, their gene pool was largely influenced by European/American
cats rather than by Asian cats. It would have been interesting to
compare the Japanese Bobtail to the indigenous Russian Kuril and
Karel bobtails which (visually) are of Western European, rather than
Asian, type.
Distinguished
by its bobtail and high-cheekboned triangular face. Close-lying,
silky short hair. The tail is 10-13 cm in length if fully extended,
but due to the kinked structure it appears only 4-7 cm long. The tail
hair often grows straight out in all directions, giving it a
rabbit -like fluff-ball or pom-pom appearance. In Japan it is found in
all colours including agouti (Abyssinian pattern) and colourpoint
though these are not accepted in the Western fancy.
In
2001 a Singapura Bobtail was reported in a breeding programme. The
bobtail kitten appeared in a Singapura litter, tracing back to a
part-tailed foundation cat. Inbreeding can cause recessive genes to
reappear. The kitten initially appeared to be tailless, but proved to
have a stumpy tail similar to that of the Japanese Bobtail. This is
not surprising since there are numerous bobtails in Singapore.
Street cats from Singapore and Johor Bahru (the town at Malaysian end of causeway to Singapore) showing the typical bobbed or kinked tail of local felines. The
bobtail trait is also widespread in parts of Russia and following the
break with communism, a cat fancy developed in Russia and several
bobtailed breeds are under development. The Kurilian’s bobbed tail
is due to an incomplete dominant gene, while the similarly named
Karelian’s bobbed tail is a recessive gene.
The
Karel Bobtail (Karellian) is a shorthaired/semi-longhaired breed
which occurs naturally along the coasts and islands of Lake Ladoga.
The mutation is claimed to be identical to the Japanese Bobtail.
These are elegant, svelte cats with lifted rumps and short pompom
tails (4-13 cm). The Kuril Bobtail (Curilsk) is smallish, compact and
cobby with a short (5-13 cm) "bob" or "pompon"
tail . It has shorthair and semi-longhair forms. The Mekong Bobtail
(formerly the Thai-Bob or Thai Bobtail) is a medium-sized Russian
breed resembling the Traditional style (Apple- or Round-head) Siamese
in all non-mitted colorpoint varieties. The tail is short (3-11 cm),
bobbed and its outline is smoothed by the coat. The Toy-Bob or Toy
Bobtail (1986) is a Russian miniature breed, no larger than a normal
3-4
month kitten. Toybobs have short, solid bodies and excellent
muscles, with short straight or curved tail-remnant (3-7 cm),
straight or corkscrewed, covered with fur in a "pompon" or
"brush" effect. All of these were recognised in the 1990s,
but apart from the Toy-Bob have existed for far longer.
Confusingly,
there is another variety known as a Thai Bobtail. It is a naturally
occurring bobtailed variety of Oriental/Burmese type. These are
mostly, but not exclusively, colourpointed and are found in Thailand
and Malaysia. The Malay Cat reported in the Malaysian peninsula in
1881 and found throughout Malaysia is similar to Japanese Bobtail,
but has not been adopted as a formal breed.
The
second officially recognised bobtail breed is the American Bobtail
bred from a foundation cat discovered in the 1960s in Arizona. This
is a dominant gene mutation. It occurs in shorthair and semi-longhair
varieties and has a powder puff tail up to one third normal length.
It was reputed to have bobcat blood (based on purely circumstantial
evidence), but it may be a Manx-type mutation since some lines
produce a range of tailless, rumpy, stumpy, longy and kink-tail cats.
To avoid mixing up different mutations, the American Bobtail is never
bred with either the Manx or the Japanese Bobtail. This is the
official American Bobtail breed, however the trait has evidently
occurred independently several times. The Sno-Bob is a colour variety
of American Bobtail. It apparently resembles the Alaskan Bobcat being
pale in colour with darker ear tips and darker bob-tail.
In
1940, American zoologist Ida Mellen wrote "The American Domestic
Bobtail Cat of the New England and Middle Atlantic States (called the
Rabbit Cat) traces its ancestry to the Manx cat, but the distribution
of tailless cats is wide, covering the Crimea and other parts of
Russia, the Bismarck Archipelago, the Malayan Archipelago, Burma and
Siam" However nothing more has been said about the purported
American Domestic Bobtail and it may have been no more than a few
isolated Manx cats. The cats from Malaya, Burma and Siam are related
to the Japanese Bobtail rather than the Manx.
There
have been claims of an American Bobtail cat breed which resulted from
bobcat/domestic cat crosses in the late 1970s. The breeder indicated
that bobcats will mate with female Siamese cats because the
scent of
a Siamese female in season resembles that of a sexually receptive
female bobcat. Bobcats can and do mate with domestic cats to produce
at least some fertile offspring.
The
Si-Bob (Si-Bobtail) is not a formal breed. It is the colloquial name
for colourpointed cats of Japanese Bobtail or American Bobtail type.
These are not recognised as part of the main breeds and their
breeding is not actively encouraged, but they are attractive
crossbreds not unlike the Thai Bobtails. Si-Bobtails occur naturally
in Japan.
The
PixieBob (1995) was, and sometimes still is, reputed to be derived
from natural bobcat/domestic hybrids and to have inherited its tail
from the bobcat. It is a polydactyl (extra-toed) breed derived from
"Legend Cats" and in some respects it resembles a bobcat.
The presence of bobcat genes has not been confirmed though the cat
certainly resembles the wild species in several respects. On the one
hand there is a persistent tale of "Legend Cats" - cats
that are the result of natural cross-breeding between the two
species. On the other hand there are cat fancies that
refuse to
recognise hybrid breeds. This leaves a few PixieBob enthusiasts both
promoting the Bobcat influence (some claiming 25% bobcat blood) and
simultaneously playing it down! Apparently genetic markers for the
Bobcat do not show up in modern PixieBobs, although this may be due
to the genes being diluted by several generations of backcrossing to
domestic cats.
There
is an unconfirmed version known as the
Munch -Bob which is a
Munchkin/PixieBob cross to produce a short-legged bobtail. In theory,
any of the bobtailed or tailless breeds could be used to produce
bobtailed/tailless Munchkins.
Not
to be confused with the PixieBob is the
Poly -Bob (1998) which is
linked to the infamous Twisty Cats (cats bred for detrimental
deformities). Poly-Bobs are not a recognised breed. They are
bobtailed polydactyls which carry the harmful form of polydactyly
which also causes gross deformities of the foreleg and front paws,
including vestigial, absent or deformed leg bones and flipper-like
forelimbs. No attempt has yet been made to eliminate these harmful
effects and they represent the darker side of cat breeding
(See Polydactyl
Cats). Occasional tailless cats suggest a Manx-type mutation in
which case the breeding line contains semi-lethal genes as well as
genes for gross deformity.
The
current trend to developing hybrid domestic breeds by crossing
domestic cats to wild species may have contributed the bobtailed
trait to one or two American cat breeds. In the American Bobtail the
evidence is anecdotal and based purely on the cat's appearance, in
the PixieBob it is unconfirmed. In the following, the hybridization
is deliberate.
The
next group of bobtails are the "Lynx" group of bobcat
hybrids developed during the 1990s. Although it is speculated that
the
Desert Lynx breed group (American Lynx, Desert Lynx, Highland
Lynx,
Alpine Lynx and Mohave Bob) is derived from bobcat crosses DNA
testing has not confirmed bobcat ancestry. The American Lynx is a
bobtailed spotted shorthair originally claimed to be derived from
bobcat/domestic crosses. The Desert Lynx is speculatively a
Manx/bobcat hybrid (some early advertisements stated
Abyssinian/bobcat) which once claimed 12.5% bobcat (i.e. bobcat
within last 3 generations) and the tail types range from absent,
through rumpy-riser to hock length. The Desert Lynx comes in
shorthaired and semi-longhaired varieties. The Highland Lynx was said
to be a hybrid of Jungle cat hybrids and bobcats. The Alpine Lynx is
a white version of the Highland Lynx. The Mohave Bob breed is a rexed
Desert Lynx developed by crossing Desert Lynx with Selkirk Rex to
introduced the rexed coat type. Mohave Bobs, Highland Lynx, Desert
Lynx, and Alpine Lynx may be bred together, with offspring registered
according to their appearance. Solid white offspring with either
straight or curled coats and either straight or curled ears are
considered Alpine Lynx. Non-white kittens with rexed coats are
considered Mohave Bobs. Non-white kittens with straight coats and
straight ears are considered Desert Lynx. Non-white kittens with
straight coats and curled ears are considered Highland Lynx.
In
spite of their bobcatty appearance, DNA testing of cats within the
Desert Lynx grouping has not found bobcat markers and these cats are
considered to be wholly domestic cats by registries.
The
Jungle Bob is a mix of F chaus (Jungle Cat) and PixieBob producing
Jungle Cat type cat with a bobbed tail. There are also alleged
Bengal/Bobcat hybrids in existence.
As
well as the main groupings of bobtailed cats, the trait crops up
elsewhere through random mutation. In the 1980s a "Spanish
Bobtail" was reported, though the exact location in Spain was
not given. In 2004 I received information on a localised strain of
bobtailed cats in one part of Spain. According to Russell Meyers, a
bobtail mutation has become
fixed in cats around the Spanish villages
of La Drova and Barx in the mountains in the Gandia area near
Valencia. There he encountered two kittens with deformed tails. one,
a male called Stitch, had a half-length tail that bent back upon
itself. The other, Milo, had a knobbly bit at the end of her tail.
The trait is known in the area as "Barx tail", Barx being a
neighbouring
village . Meyers came across a large number of cats in
the area with similar tails. The most likely scenario is that a
tomcat carrying the mutation fathered most of the kittens during one
or more breeding seasons. He was not necessarily the cat in whom the
mutation originally occurred, but by fathering the majority of
kittens his genes - including the mutation - would have become
widespread and the trait would have shown up in later generations
through inbreeding.
In
2006, TICA proposed to clamp down on certain breeding
trends including new bobtailed breeds created by crossing existing breeds
together. Their Genetics Committee report stated:
"The
Committee proposes that TICA does not accept any proposed breeds for
Registration Only status that do not exhibit novel mutations. The
current mutations would be reserved for currently recognized breeds
exclusively. This would end the seemingly endless applications for
"munchkinized" new breeds, and then deter the inevitable
introduction of "rexed", "Bob-tailed" and
Poly-ed" everything else."A
bobtail breed in development is the
Tennessee Bobtail. A description
of its origins and type is provided by Patty Shane. Scattered
throughout the Southern USA are a variety of bobtailed cats. The more
common type is the Manx-type with tails ranging from rumpy to full
and bodies ranging from the cobbier high-rumped Manx-type to a more
svelte build as a result of random breeding. Some have ear tufts. The
less common type is the Oriental (Japanese) Bobtail type. These lose
the distinctive bobbed tails when out-crossed to other cats, although
the kittens may have kinks or curves in the tail. The bobtail shows
up when they are bred to each other. They are long,
tall cats with
more Oriental features and conformation and level backs in spite of
their long back legs. When bred together, these create a long, tall
cat, with some individuals having a broader, heavier frame and others
leaning towards one or other of the parental types. The goal is a
long, tall 6lb - 15 lb cat with the high, angled hind-legs of the
Oriental type. The head should be apple or pear shaped, the ears
straight and moderate sized (with tufts permitted) and the eyes
almond (preferred) or lemon shaped in any colour. Tails range from
tailless to full tailed and included bobs and single or multiple
kinks or curves. A natural bobtail with kink(s) and/or curve(s) is
preferred over tailless or long-tailed cats (the standard prohibits
the
docking of long tailed kittens). They come in any fur-length,
colour and pattern with tabby/spotted preferred. Hairless, curly or
wirehair is not allowed.
Two Tennessee Bobtails - Siam and Miss Hissy. Although tabby/spotted is preferred, the colourpoint pattern also occurs. Photos copyright Patty Shane.At
present, foundation Tennessee Bobtails are being registered with
REFR. These are random-bred natural bobtail cats that meet the
desired type. Cats meeting the Tennessee Bobtail type and belonging
to a known bobtailed breed are being used to help establish
bloodlines. Where a Manx-type cat is used, it is outcrossed to
another type of bobtail to prevent Manx syndrome (skeletal and neural
tube defects) as a result of Manx-Manx matings. I don't yet know how
this breed will be visibly distinct from the American Bobtail.
In
2007, Vasilis Lekkas from Athens (who is deeply involved in the
recently formed Greek network for the preservation of domestic
livestock including cats) reported One complete bobtail and four
cases of shortened tails, about half the normal size, where the tail
ends were blunt and thick rather than tapering as in regular cats.
In
2011 a colony in Canada was report to have a short-tailed gene not
passed on as a dominant. Some lines of Bengal have had
issues with
both spina bifida and with spontaneously producing
bobtailed/corkscrew-tailed kittens accompanied by other defects
including still-born kittens with intestines outside the body. Less
affected cats tended to have a
lump or bump at the base of the tail.
Whether this defect is akin to recessive gene found in the colony in
Canada, or to an incomplete dominant,had not been established.
CURLY
TAILED CATSCurly
tailed cats have occurred throughout history and around the world
though only recently has any interest been shown in perpetuating the
trait in the form of the Ringtailed
Sing -a-
Ling (which became the
American Ringtail in 2004). Many of the 1990s reports of curious
tails confuse curly tails with bobtails. Some suggested that the cats
are "longtailed bobtails" i.e. have a kinked section of
tail close to the body, combined with a full-length tail with normal
vertebrae beyond the kinked section; the kink would lift the tail
upwards and the normal section would hang over the back or flank. In
"Variation of Animals and Plants Under Domestication"
Charles Darwin wrote: " I have seen a cat which always carried
its tail flat on its back when pleased."
Although
judges examining the early American Ringtails had reported the trait
to be very rare, this is not strictly correct. Judges see pedigree
cats and pedigree cats are bred for uniformity and adherence to a
standard. This means rogue genes have been eliminated generations
ago. The genes for curly tails would have been weeded out of pedigree
lines. In addition, a curly tail would be a deviation from the
standard (and might be considered a sign of genetic impurity in the
breed) hence the cats would not be entered in shows, except possibly
in household pet classes. The trait is found with greater
frequency in random breeding cats but until recently was seen as no more than a
charming oddity. I was
surprised to be told that the trait is so rare
as I had already encountered several curly tailed cats and kittens
(nicknamed pigtailed kittens) in Chelmsford, Essex, England. Their
curly tails were quite distinct from the kink found in bobtails.
In
the Jan/Feb 1998 issue of the Cats Protection League magazine "The
Cat" there is an account of further curly tailed kittens from
Basildon, a town only about 13 miles from Chelmsford:-
"At
the time of writing, our latest little problems are four nine-week
old kittens who have curly tails. Our vet has advised us that their
tails are deformed and not growing properly and will have to be
removed when the kittens are a little bit older. Our vets have said
that once their tails are removed, they will be able to live
perfectly normal lives, although they will look like little Manx
cats."The
report would have been written in November/December 1997 to make the
Jan/Feb 1998 edition. The accompanying photo showed the kittens
sitting on a cushion; their curled tails were not visible. A follow
up report in a later issue merely noted that the kittens' tails had
been removed and the kittens had found homes. The only other recorded
case of a whole litter of curly tailed kittens was a case reported to
Sue Manley (who is establishing a curly tailed breed) and was a
litter born to a barn cat in Oregon.
Ida
Mellen's 1940 discourse on oddities in cats included a picture of a
curly tailed cat from Jenkintown, Pennsylvania. This ginger and white
cat's long tail clearly forms a tight spiral. Phyllis Lauder, writing
in 1981 about Shorthaired cats, noted that an ACA directive stressed
that the tail shall not be carried over the back:
"This
last is not a common fault, but it certainly looks strange when it
occurs; there is a Siamese neutered pet who carries his tail as would
a Pekinese dog, and the effect is to make people laugh!" Several
curly tailed cats have been reported in random-bred cats in Britain
in the 1990s and early 2000s. A curly-tailed tabby and white male cat
was reported in Perthshire, Scotland in 1986. The degree of curl
ranges from loose through to a tight corkscrew. In one case, the tail
curled at the base and for the rest of its length lay flat along the
spine.
In
the 1980's, Katrina Lee of Washington, DC, vacationed on the island
of Guadeloupe . In a small restaurant on the lower island she saw
about half a dozen various colored cats with corkscrew tails. The
tails were not looped over, but stuck out from the body like a normal
cat's tail while looking like they'd been wrapped around something to
make them spiral!
In
Britain, one curly-tailed cat has achieved fame with South Ribble Pet
Cat Club. Raffles is a white semi-longhair born in June 1998 and
homed via a cat shelter in Oldham. When sitting, his tail forms a
curl behind him. Another British curly tail is Sprocket found at
Spaghetti Junction (Gravelly Hill motorway interchange in the
Midlands). She was taken to Erdington Cat Rescue and it seemed that
some prospective adopters didn't want her because she had a curly
tail and it was perceived as a deformity or health problem.
Since
the breeding programme for the American Ringtail began, reports of
curly tailed cats have come in from around the world, indicating that
it is not as rare as previously claimed and that there might be
several different genes involved. It is now believed to be a
polygenic trait i.e. several genes interact to dictate the type of
curl and the degree of
curling . Though the trait seems to be
polygenic, some curly tailed males pass the trait onto more than 50%
of the offspring while females seem to pass it on to a lesser degree.
This suggests sex-linkage. There might be several different gene
mutations producing a visually similar effect.
Unlike
the bobtail where the vertebrae are fused into permanent kink and
motion is limited, curly tailed cats have mobile tails and no fusion
of vertebrae. The trait is hereditary, possibly as a result of
several interacting genes which govern the type and degree of curl,
and not a birth defect as once thought. Generally, the tail curls up
and over in an arc or full circle, coming to lie against one or other
flank as the cat walks. Owners of curly-tailed cats are often
intrigued as to whether similar cats are "left-handed" (the
tail tip on the left flank) or "right-handed". Sue Manley,
breeder of American Ringtails (formerly Ringtailed Sing-a-Lings)
produced another unusual tail alongside the normal ringtails. She
described it as "baboon tail". The cat holds it curled up
in a compete ring underneath him while he stalks around or hunts his
toys. He then brings it up over his back when he is petted.
One
peculiar curly tail was seen in a cat brought to a cat shelter in
Chelmsford, Essex, England in 1999 or 2000. This was a black female
cat whose tail curled up over her back and then lay flat along her
spine. It gave her the appearance of a Rhodesian Ridgeback dog. The
tail was mobile, but returned to that position when she was at rest.
Over the years several "pig-tailed" kittens have turned up
in the area, either as spontaneous mutations or due to genes in local
feral colonies. The occurrence of several curly-tailed cats and
pig-tailed kittens around the Chelmsford and Basildon areas in a six
year time frame suggests a curly-tail hotspot in that part of Essex.
Between
2000 and 2002 there were intermittent reports of curly-tailed cats
and also of people seeking curly-tailed cats as companions for, to
replace , an earlier curly-tailed cat. Most reports came from the USA
(sadly from unidentified locations). These included a grey tabby
female with a curly tail, and a grey shorthair female with a tail
that curled over to rest on her back. The latter cat's tail was
described as spring-loaded, quickly returning to its curled position
if the tail is straightened by hand during petting. This led to a
suggestion that the trait was associated with grey cats.
A
red tabby male from Missouri was reported to have a cute pigtail
generally held curled up and over his back in a large perfect loop.
His littermates had normal tails. His tail sometimes unwound to lie
flat on his back or was held in a more conventional unwound posture
during play. As the cat aged, the tail was held curled more often,
suggesting a gradual loss of flexibility. The owner stressed that
this caused the cat no discomfort. A cream tabby female developed her
curly tail relatively late in life. She apparently had a near normal
tail until she reached 8 or 9 months old, after which it became
progressively more curled, forming an up-and-over curl with the tip
resting on her left flank. This again suggested a loss of flexibility
as the cat aged.
The
majority of curly-tailed cats are random-bred cats since purebred
cats have necessarily lost a degree of genetic diversity in order to
produce consistent conformation. In 1997/98 a curly-tailed Turkish
Van was reported. A Singapura with an unusual tail was reported in
2002. The tail was described as very flexible and he could curl it
round. It is not like American Ringtails, as it does not curl as
much, nor does it curl up and over. It is held low and curled in a
loop at his side. An American Keuda with a similar tail
("Curlietail") was reported at around the same time. She
double curled the tail when she sat down, but at 5 months old she
does not curl it as much. Again, it would appear that flexibility is
lost as the cat matures. Since the establishment of a breeding
programme, it seems that curly tails of varying degrees are more
frequent that initially suspected. What was once seen as a fault now
has a serious following. Curled tails have also turned up in Russian
Blues, Siamese/Orientals, Ocicats, Persians, Ragdolls, Scottish Fold,
Devon Rex and Bengals (interestingly, several of those breeds have
Siamese blood in their ancestry).
The
photos above, very clearly showing the curly tail trait, are of
Barbara Clark's 4 year old curly-tailed cat "Bunzy". His
brother has a normal straight tail. Bunzy and his brother were
adopted from a cat shelter in Utah, USA at 8 weeks old so nothing is
known of their family history. Bunzy's tail clearly loops over to
rest on his left flank with the tail tip pointing outwards.
A
gallery of curly-tailed cats sent in by visitors to Messybeast can be
seen at Curly
Tailed Cats (a separate page has been created to improve
loading times).
WHITE
RINGTAILIt
should be noted that historical reports of the "White Ringtail"
cat refer to the Turkish Van with its ringed or banded tail, not to a
race of curly tailed cats. Some early mentions of curly tailed cats
will have referred to the bobtailed cats common in parts of Asia.
Also, very obese cats may hold their tails in a curled tail position
for the simple fact that rolls of fat at the base of the tail prevent
it from reaching any point below horizontal.
LONGHAIRED
CATS
Copyright
2002-2015, Sarah HartwellThe
gene for longhair in cats is recessive i.e. a needs 2 copies of the
gene in order to have longhair. A cat with one copy of the gene is
shorthaired, but can pass the longhaired trait on to its offspring -
the carried gene will only come to light if the cat is bred to
another
carrier of the gene or to a longhaired cat.
Remains
of early cats can't tell us about fur length, but cats depicted in
early Egyptian art are shorthairs and the parent species of the
modern domestic cat, the African Wild Cat, is shorthaired. The
shorthair coat ranges from plush and dense as in British Shorthairs
and sleek and close-lying as in the Oriental breeds, with a variety
of intermediate coat types depending on actual fur length and whether
or not there is a woolly undercoat. There are a number of theories as
to how the longhaired domestic cat arose.
Hybrid
Origins?In
the 1800s, Pallas suggested that Angora and Persian cats were
descended not from Felis silvestris but from Pallas’s cat (Felis
manul) which he discovered and named. This suggestion was quoted by
Darwin in 1868 who wrote, "
The
large Angora or Persian cat is the most distinct in structure and
habits of all the domestic breeds; and is believed by Pallas, but on
no distinct evidence, to be descended from the Felis manul of middle
Asia.".
There is anecdotal evidence that Pallas cats can interbreed with
domestic cats to produce offspring, but to introduce the gene, the
hybrid offspring would have to be fertile and be bred back to
domestic cats. In 1907, Pocock described the various English domestic
cats for Royal Zoological Society and strongly refuted the Pallas cat
theory since the skull of Pallas’s cat differed from that of the
Angora or Persian of Pocock's time. Modern genetic studies have also
shown that the Pallas cat did not contriubte to the gener pool of the
modern domestic cat.
There
are claims that the Persian is descended from the
Sand cat (F
margarita) based on that fact that both have long hair covering the
paws which forms a pad over the soles. Measurements of the length of
the hair on the body and feet of both Persians and Sand cats does not
uphold this claim. In addition, the long hair on the feet of modern
Persians is due to their long coat; in the Sand cat it is a feature
since the cats are otherwise shorthaired. Again, there is no genetic
evidence of Sand Cats contributing to the domestic cat gene pool
(although hybrids have been bred more recently.
Single
Origin or Multiple Mutations?A
more plausible explanation is a gene mutation in a group of cats with
the foreign conformation. Inbreeding would have allowed the trait to
become fixed. This mutation may have originated in Asia Minor and in
a book published in 1876, long-haired cats were referred to as
Asiatic cats.
From
historical writings, the longhair mutation either arose in three
separate areas (Russia, Persia (
Iran ) and Turkey), or that the
longhair mutation originally occurred in Russia and that Russian
Longhairs (Siberian, Russian Angora) spread from Russia into Turkey
(becoming Angoras), Persia (becoming Persians) and surrounding
countries and south east Asia (introducing the gene into native cats
to create Longhair Japanese Bobtails) by
land and sea trade routes,
retaining many of their
cold -climate adaptations: a heavier body,
dense undercoat and coarser hair. If so, all longhairs are derived
from the Siberian cat. Yet another view is that long hair arose and
was developed in Turkey and transported on land and sea trade routes
to Europe, the Middle East and Far East. Much of this is based on the
supposed origins of the modern Persian breed which was derived from
Turkish Angoras, Russian Longhairs and purported Persian cats. Long
fur in domestic cats appears to be an adaptation to cold as the paws
do not have long fur underneath; in desert species of wildcat long
hair is found on the paws and protects against hot surfaces.
It
is possible that the longhair mutation has occurred not once, but
many times and that it is still occurs. Some genes are mutational
hot-spots e.g. identical mutations for curly hair and for bobbed
tails have occurred (and still do) independently in differently
locations. Parallel evolution means that similar cold environments
could lead unrelated cat populations to evolve similar traits through
natural selection in a relatively short period of time. Long hair
mutations are seen in other cat species e.g. the cheetah has produced
semi-longhair "woolly cheetah" variants in the past.
Longhairs
range from "woolly" usually due to the undercoat) to fine
and silky and the length itself varies from the
flowing coat of the
modern Persian to the semi-long coat of Turkish breeds.
Semi-longhaired variants in normally short-haired breeds are
generally blamed on recessive genes, but could be due to more recent
mutations.
The
first longhairs recognised by the western cat fancies were Angoras
and Persians whose histories are intertwined. The Persian group is
defined as all varieties developed from cats imported from Turkey,
Persia (now Iran), Afghanistan and Russia at the turn of the
18th and
19th centuries. Brian Vesey-Fitzgerald "Cats" (1958)
mentioned the belief, held by some, that a female Black Persian was
imported direct from the Shah of Persia himself.
EARLY
LONGHAIRED CATS: ANGORAS AND PERSIANSLonghaired
cats were first seen in Europe in the 1500s, first in Italy (1521)
and then in France. They were named after the Turkish city of Angora
(Ankhara). The first documented ancestors of the Persian were
imported from Persia into Italy in 1620 by Pietro
della Valle, and
from Turkey into France by Nicholas-Claude Fabri de Peiresc at around
the same time. From France they soon reached Britain. The longhaired
cats from Persia were interbred with Turkish Angoras. A tabby Angora
cat is depicted in Buffon’s "Natural History" (1756). Its
head length and conformation are generally similar to the Turkish
Angora except that its ears are set lower on the head.
The
first longhairs in Britain were variously described as Angoras or as
French cats (the latter generally being white). They were also known
as Chinese (some longhairs were imported from China where they were
known as four-ear cats due to the furnishings of the ear), Russian
(the Russian Longhair or Russian Angora was known) and Indian. It was
the existence of longhairs in China which led some to speculate that
the longhair trait entered the domestic cat population through
hybridisation with the Pallas cat. It is suggested that longhairs
reached China from Iran (Persia), as a gift of the king of Persia.
The
eighteenth-century French naturalist the Comte de Buffon quoted the
16th/17th Century Italian traveller Pietro della Valle:
‘In
Europe there is a species of cats which properly belong to the
province of Chorazan the USSR and Afghanistan. Their beauty consists
in the colour of their hair, which is grey, and uniformly the same
over the whole body, except that it is darker on the back and head,
and clearer on the breast and belly, where it approaches to
whiteness. Besides, the hair is fine, shining, soft as silk , and so
long, that, though not frizzled, it forms ringlets in some parts, and
particularly under the throat. The most beautiful part of the body is
the tail, which is very long, and covered with hair five or six
inches in length. They extend and turn it upon their back, like the
squirrel, the point resembling a plume of feathers. They are very
tame; and the Portuguese have brought them from Persia into India.’
From this description it appears, that the Persian cats resemble, in
colour, those we call Chartreux cats, and that, except in colour,
have a perfect resemblance to the cat of Angora.Buffon
had not seen these "Persian" cats himself, but he was
familiar with the Angora in France and he apparently believed there
was no difference between an Angora and a Persian except in colour
(the cat was possibly a black smoke or blue smoke Angora). He named
them "Catus Angorensis".
An
apparently now extinct longhaired cat from China was known as the
Sumxu, a fold-eared cat which was reported from the area around
Peking, China. It was a longhaired cat with white fur and pendulous
ears. It was described (mostly with regard to its folded ears)
several times in the 1700s, with the final report being in 1938;
though evidently extinct it indicates the presence of longhairs in
China.
In
the late 18th century, longhair cats with coarser, denser coats, and
a stockier build were imported into Britain from Persia, Afghanistan
and Russia. Cats imported from Turkey were mostly whites with a
short, soft, silky top coat and little undercoat (and no woolliness
in the undercoat). Those from Russia, Afghanistan and Persia were
mostly black or blue and less foreign in type. The Angora has a ruff,
breeches and a plumy tail, but unlike the later Persian, its coat
followed the lines of its body due to the absence of woolly "padding"
beneath. The Russian Angoras had green rather than blue, eyes.
A
letter from M Lottin de la Val, President of the
Imperial Acclimatation Society, to the President of the French Zoological
Society in 1856 stated,
"When
you recently did me the honour of calling on me, you imparted the
recently held view that the so called 'Angora' cat does not exist or
could not exist except in the vicinity of ancient Ancyra. I hasten to
dispel this illusion. I myself came upon specimens of that lovely
feline species in the great Armenian plateau, at Erzerum, where the
climate is greatly different from that of Angora. The species is very
numerous at Mourch in Kurdistan, where it is the dominant variety. I
also found it at Billis and in the pashalik of Bayazit.. The finest
specimens, however, which I saw belonged to the Archbishop of Van, a
town in the east of Kurdistan, on the frontier of Azerbaidjan. He had
three of them, one pearl grey, one orange-hued with black and white
flecks, and a third, which was completely white. Their fur was
magnificent, though there was thought to be nothing to be surprised
at in them, as such cats are common in Kurdistan. I also saw some at
the residence of Khan Mahmoud, Prince of Hekiars, at Alpeit. I can
not recall having seen any in Persia, though, had I thought that
scientists might have been interested, I would have taken care to
seek them out, busy as I was. But what will surprise you most of all
is that despite the high temperatures prevailing, one should find
Angora cats at Baghdad, though certainly these are not so fine as
those to be found on the northern slopes of the medique and Taurus
mountains, though whether the difference is due to the hot atmosphere
or the hostility of the people of Baghdad, I cannot say. You will no
doubt settle that point better than I could, all I can say is that
the people of Baghdad are in constant warfare with their cats,
maintaining, not without good reason, in my opinion, that they bring
the plague, because of their fur coats and their habits."While
these longhairs might not have been appreciated in their native land,
their charms were not lost on British cat fanciers. In "Our
Cats" in 1889, Harrison Weir wrote of the various longhaired
cats:
"There
are several varieties - the Russian, the Angora, the Persian, and
Indian. Forty or fifty years ago they used to be called French cats,
as they were mostly imported from Paris - more particularly the
white, which were then very much in fashion." AngorasIt
was not until the mid-l9th century that cat enthusiasts distinguished
between Turkish Angoras and other long-coated cats coming from Persia
and Russia. According to the “Illustrated Natural History” by the
Rev JG
Wood (1853,
1874 ):
There
are many varieties of the Domestic Cat, of which the most conspicuous
are the MANX CAT and the ANGOLA . [...] the Angola Cat being gorgeous
in its superb clothing of long silky hair and bushy tail [...] A fine
Angola Cat is as handsome an animal as can be imagined, and seems
quite conscious of its own magnificence. It is a very dignified
animal, and moves about with a grave solemnity that bears a great
resemblance to the stately march of a full-plumed peacock conscious
of admiring spectators. It is one of the largest of domestic Cats,
and in its own superb manner will consume a considerable amount of
food. Wood,
a naturalist rather than a cat-fancier, evidently meant the Angora.
In
1868, an English writer described the Angora as "
a
beautiful variety with silvery hair of fine texture generally longest
on the neck but also on the tail".
In the early days of the cat fancy, longhaired breeds competed
against each other; the Persian was preferred over the Turkish Angora
and the Russian Longhair (Russian Angora), so much so that any
resemblance to the Angora was later frowned upon by cat fanciers.
In
1870, the Honourable Lady Cust produced a book called "The Cat"
(in which she perceptively wrote "In the present day, a love for
cats appears chiefly permitted to 'elderly spinsters,' and is often
even ridiculed") in which she described the longhair thus:
"Of
the wild cat, there is supposed to be only one species, which extends
with trifling variety of colour over all parts of the world, the
difference from the tame variety being more in the internal than in
the external structures, its intestines being the shortest and
smalles of all the quadrupeds. […] Hiertro dello Valli evidently
means the Angora kind when he says 'There is in Persia a cat
(particularly in the province of Choragan) of the figure and form of
our ordinary ones, but infinitely more beautiful in the lustre and
colour of its skin. It is of a grey blue, without mixture, and as
soft and shining as silk. The tail is of great length and covered
with hair six inches long, which the animal throws on its back like a
squirrel.'"Jean
Bungartz described the Angora (Felis maniculata domesticus
angorensis) in his 1896 book "Die Hauskatze, ihre Rassen und
Varietäten" (Housecats, Their Races and Varieties) in "
Illustriertes Katzenbuch" (An Illustrated Book of Cats) as the
most beautiful and best known of the foreign cats and originating
from high Asia. He summed up the debate over whether the longhair
meant it was related to Pallas's Manul or was a housecat adapted to a
cold, mountainous climate. He added that there were bluish-grey
Angoras in the south Siberians. Bungartz wrote that it could not be
proven whether it really came from Angora or just got that reputation
because of Angora goats and rabbits from that region. He noted that
white and silver (chinchilla?) were most popular, followed by blue,
black, "grey-touched" (shaded silver?) and isabelline
(cream) while other colour occurred through addition of other blood.
He noted that crosses between Angoras and usual housecats lacked the
full, rich, silky hair. It was a
favourite , and very indulged,
salon-cat in Bungartz's time and he described it as calm,
aristocratic, attached to humans and a coddled lap-
child ! He also
noted the need to groom its hair to prevent matting ("a
horrible, inextricable felt ball") and the need to wipe the eyes
clean.
In
a footnote to his lengthy notes on the Angora, Bungartz added a short
statement about the Persian: "The Khorassan or Persian cat seems
to be a modification of the Angora cat, their hair is somewhat more
woollier and curlier, but nevertheless still especially long. The
colour is dark bluish gray. In terms of beauty, she is quite close to
the Angora cat, but is far rarer. "
By
1903 the Persian had replaced the Angora and by the 20th century the
Turkish Angora was virtually unknown outside of Turkey. In Turkey,
they were seen in many colours e.g.
sarman (red
tabby),
teku (silver
tabby),
Ankara kedi/Van kedi (odd-eyed
white), brown tabby, selfs (solids), torties and tortie-and-white
although some cat fanciers still claim that the white Angora is the
only pure native Turkish Angora. There is a true-breeding colour
variant found near Lake Van originally known in Britain as the
Turkish Cat (or Turkish Swimming Cat) and later as the Turkish Van.
In Turkey the name "Van kedi" (Van cat) refers to the
highly prized odd-eyed white. Turkish
Vans are auburn (red) and white
with the colour restricted to around the ears and on the tail. They
are also known to
swim in shallow pools and streams.
In
their study comparing the genetics of several breeds (Lipinski MJ, et
al., The ascent of cat breeds: genetic evaluations of breeds and
worldwide random-bred populations, Genomics (2007), the American
Turkish Angora breed and the American Turkish Van breed were found to
be distinct from each other. The American Turkish Van was related to
Egyptian random-bred cats while the American Turkish Angora was
closer to random-bred cats from Tunisia and Turkey. Lipinski
suggested that cats may have spread into different regions of Turkey
from different directions: into the Lake Van region from
Egypt via
land trade routes and into the Ankara (Angora) region from Tunisia
via sea routes. Cat fancy history also refers to "Russian
Angoras" suggesting a historical link between Russian and
Turkish longhaired cats. Studies of Turkish Angoras maintained as a
pure breed at the Ankara Zoo found that the American "Turkish
Angora" is not closely related to the authentic Turkish cats.
The American version was bred from Persians and other cats to
"resemble" the Angora and is physically and genetically
different from true Turkish cats.
Angoras
were extinct in Britain until after World War II. In the 1950s and
1960s, Turkish cats were taken to North America, Sweden and Britain,
but only the Van cats were bred and recognised in Britain. In the US,
white Turkish Angoras were recognised in the early 1970s and other
colours in 1978. They are now recognised in a full range of colours
including black, blue, tabbies, tortie-and-white and smoke colours.
Turkish Angoras have silky coats and gentle dispositions.
Meanwhile ,
in Britain the name "Angora" was given to an impostor! In
1977, a breed confusingly known as the Angora was recognised in
Britain. Although similar to the Turkish Angora, it is bred in a
wider range of colours (including chocolate and lilac, which come
from Siamese ancestry), its voice is similar to that of the Siamese
and it is more fecund than the Turkish Angora. The British Angora is
a Foreign Longhair bred to recreate or resemble the Turkish breed
rather than reintroduce genuine Turkish cats. It is related to the
Oriental and Balinese. Elsewhere in Europe it is known as the
Javanese or
Mandarin ; in the USA it is a Foreign Longhair. The
British Angora is more foreign in type with a longer, narrower head
and larger ears than the Turkish Angora.
This
situation was rectified in 2002 when Britain's GCCF came into line
with other registries and called the British variety Oriental
Longhairs, thus removing confusion with the original Turkish Angora
cats.
The
possibility of importing Turkish Angoras was apparently considered,
but bureaucracy prevailed since the British GCCF would apparently not
accept the documentary evidence (verification of breed) supplied by
the Ankhara Zoo. If true, then the British cat fancy places more
importance of paperwork than on actual physical felines; a serious
drawback where naturally occurring varieties are concerned!
Importation of either Turkish or American lines of Turkish Angora
would mean 6 months quarantine.
Breeders
of the British Angora have attempted to recreate the characteristics
of the original Turkish cats. Those working with Turkish Angoras/Vans
have worked to preserve characteristics of those naturally occurring
breeds and the Turkish Angora has prior claim to the Angora name.
Luckily the genuine Turkish Angora has since returned to Britain.
The
most obvious differences between the Turkish Angora, British Angora
and Persian are the coat, head shape and general conformation. The
British Angora is an oriental cat - long-bodied with a longer,
wedgier head and larger ears. The Turkish Angora is less foreign in
build, with a wider, shorter head and smaller ears. The Persian has a
wide head, flat face, small-ears and extremely cobby build. In
addition, the Persian has longer fur with a woolly undercoat and a
tendency to tangle or knot. The Angora coat is silkier without the
woolly undercoat; the full Angora coat is seen in the
winter - in
summer, they moult and may look like shorthairs with fluffy tails.
In
an article in the Colourpoint, Rex-coated and AOV Club's Journal
(reported in 1981 by cat breeder Phyllis Lauder), Robin Sims made the
point that though the terms "Persian" and "Angora"
were often used interchangeably, the cats concerned were not the same
and that the flowing coats of the cats imported into Europe from Iran
were not governed by the same gene as that concerned with the cats
imported from Ankara. Sims made it clear that the Turkish Van were
Angoras, and held the opinion that it was unfortunate that in the
early 1900s the Angora was used to improve the coats of the Persians.
PersiansIn
1868 Charles
Ross described the Persian as "
a
variety with hair very long and very silky, perhaps more than the Cat
of Angora; it is however differently coloured."
The cats most likely to win prizes at early cat shows were those
which combined the Angora's longer fur with the Persian's shorter
faces. The Turkish blue-eyed white cats were crossed with
shorter-nosed blacks, blues and fawns (creams). The whites became
shorter-faced, but the distinctive blue eyes were often replaced by
orange eyes or odd eyes. Another reason for cross-breeding might have
been to tackle the problem of deafness in blue-eyed white Angoras.
In
the 19th century, Harrison Weir described Persians in white, black,
blue, grey, red and "any other" self colour; tabbies were
recognised in brown, blue (black markings on blue background,
possibly black-silver tabbies), silver and light grey-and-white
(probably blue-silver tabbies). In 1872 Weir described smoke
Persians, "
a
beauty was shown at Brighton, which was white with black tips to the
hair, the white being scarcely visible unless the hair was parted".
The following year these were numerous enough to have their own
classes. The British National Cat Show of
1879 included a Persian
with a coat of "
strangely
graduated grey"
(possibly chinchilla or shaded silver). Chocolates, lilacs and
tortoiseshells were not described. Creams, then called fawns, or
derisively as "spoiled oranges", were first recorded in
1890 and were darker in colour than they are today. The first Smoke
Persian Champion (1893) was Mrs H. V. James's Backwell Jogram and it
is worth noting that he is completely different to the modern Smoke
Persians and (judging from photos) bears a striking resemblance of
type to the modern Siberian Longhair.
In
her 1903 "The Book of the Cat", Frances Simpson did not
distinguish between the Persian and Angora and did not mention the
Russian Longhair. She wrote
in
"In classing all long-haired cats as Persians I may be wrong,
but the distinctions, apparently with hardly any difference, between
Angoras and Persians are of so fine a nature that I must be pardoned
if I ignore the class of cat commonly called Angora, which seems
gradually to have disappeared from our midst. Certainly there is no
special classification given for Angoras, and in response to many
inquiries from animal fanciers I have never been able to obtain any
definite information as to the difference between a Persian and an
Angora cat. Mr Harrison Weir, in his book on cats, states that the
Angora differs somewhat from the Persian in that the head is rather
smaller and ears larger, fur more silky with a tendency to
woolliness."She
considered the Persian to be less reliable in temperament than the
English short-hairs, but considered them more intelligent and also
keen hunters. However, they were less healthy than short-hairs and
the longest haired kittens were the most difficult to rear. She
attributed these health problems to in-breeding. Persians were
disadvantaged by the fact that cat shows were held in the summer
months (many were held in marquees), at a time when long-haired cats
were moulting and tended to look
moth -eaten and unkempt. This, she
felt, held them back - not that the numbers shown upheld this
opinion, long-hairs outnumbered English short-hairs by about four to
one!
According
to John Jennings "Domestic or Fancy Cats", published a
little earlier than Simpson's work,
"Of
the many varieties or breeds of the cat with which we are now
familiar, it must be remembered that, however crossed, selected,
re-crossed, domesticated, or what not, we have but two breeds on
which the super -structure of what is known today as the
'classification of varieties' has been reared - viz, the long-hair or
Eastern cat, and the short-hair or European." Jennings
found little difference in the skull length between those long-hairs
and short-hairs; indicating that the Victorian long-hairs had not yet
become the snub-nosed cats familiar to 21st Century
cat lovers.
When
Weir drew up his "points of excellence" for judging cats in
1889 he defined differences between the Angora and the Persian.
Pocock later stated (1907) that Persians had a marked shortening and
widening of the face i.e. it had diverged from the Angora. In
addition, the Angora had sleeker fur, lacking a woolly undercoat. By
1903, Simpson had effectively dismissed the Angora cat in favour of
the Persian type. Only in very recent times has the true Turkish
Angora come back from being thus relegated in Britain.
By
1901 the colours were black, white, blue, orange (red selfs, red
tabbies), cream or fawn, sable, smoke, tabby, spotted, chinchilla,
tortoiseshell, bicolour and tricolour (tortie-and-white). True
spotted Persians were described as being rare or non-existent; almost
all had some tabby lines and rings. In fact the long flowing coat of
the Persian does not lend itself well to spotted or mackerel tabby
patterns. The sables were described in a 1903 book as "
a
kind of brown tabby ... These cats have not the regular tabby
markings, but the two colours are blended one with another, the
lighter sable tone predominating"
which suggests shaded goldens. The orange (red self) was always
popular and by 1915 there were classes for red self or shaded and for
red tabbies. Blue-creams were recognised in 1929.
Persians
arrived in North America from Europe towards the end of the 19th
century and by the turn of the century they were eclipsing the native
Maine Coon. Frances Simpson was an early champion of Persians and
devoted considerable space in her 1903 book "The Book Of The
Cat" to Persians "distinguished by unusually long coats,
round heads, tiny ears, and wonderful toe tufts" (she also
included the Maine Cat in her book as we shall see later on).
Simpson
wrote
"A
gentleman who has lived for ten years in Assam says that he never saw
in that part of India any long-haired cats except blue-eyed whites"
although these seemed mainly to belong to English colonials, rather
than being native cats. At around the same time, Mrs. Clinton
Locke ,
president of the Beresford Cat Club, wrote in a letter to "Our
Cats"
"The
first white Persian I ever owned was brought to me many years ago
from Persia by a distinguished traveller, and its eyes were amber".The
White Persian was especially popular:
"There
is always a keen demand for white kittens, either as pretty pets or,
if with correct-coloured eyes, for breeding purposes, and, doubtless,
when more encouragement is given to this beautiful variety, there
will be an increase of fanciers of the white cat, whose praises have
been sung in fairy tales, nursery rhymes, and by novelists who have a
weakness for describing interiors with a beautiful white Persian cat
reclining on the hearthrug.""Whereas
formerly blue eyes were considered quite a rarity, now it is seldom
we see any yellow-eyed white cats exhibited at our principal shows
[...] It is easy to tell whether the baby blue eyes are likely to
retain their colour or turn yellow. If at about three weeks or a
month old the blue becomes tinted with green, then surely but sadly
may we make up our minds that these kittens have not a distinguished
career before them, for they will see and be seen with yellow eyes."
A well-known authority on cats, wrote in a letter to one of the cat
papers, "A few years ago white cats with green or yellow eyes
frequently were prize-winners, and a blue-eyed white was looked upon
as a rarity. Now blue eyes have it all their own way." Occasionally
green-eyed White Persians turned up, hinting at Russian Angora
ancestry.
Imported
longhairs (probably then of Angora type) were considered better
quality than British-bred cats: "the most perfect type of a
white Persian is assuredly to be found amongst the imported cats;
there is a certain beauty of form and silkiness of fur which is not
possessed by the specimens bred in this country [...] These imported
cats are often of a rather savage disposition, and, although they can
be
sweet -tempered enough with human beings, they are extremely fiery
with their fellows."
Angora cat by J B Huet, 1808 "
Collection des Mammiferes de Museum d'Histoire Naturelle" (Paris)
Weir's idealised longhair, 1889
In
1906 a Canadian writer described the conformation of one Persian
thus: "
His
head is magnificent, and he is short on the leg, has plenty of bone"
and the best Persians were reckoned to come from Britain. In the
1960s, Fernand Mery wrote that the development of longhairs into
extremely cobby cats was due to either the British climate or was a
British preoccupation since the British had done the same thing with
the Chow dog breed.
Persians
were also favourite photographic subjects as Sydney W France's noted
(with some dismay) in his book "Siamese Cats" (1949) though
he was good enough to list the various colours of Persian:
"Take
a look at any cat magazine, or at any cat photographs you may see in
the papers or periodicals. Mostly of the long-haired cats, are they
not? Chinchillas, Blue Persians, Biscuit, Smoke, White Persians." The
"Biscuit" colour sounds to be cream or fawn. Grace Cox-Ife,
author of "Questions Answered About Cats" (1947)
lists the
official Persian colours as Black, White (blue-eyed, orange-eyed),
Blue, Red Self, Cream, Smoke, Silver Tabby, Brown Tabby, Red Tabby,
Chinchilla, Tortoiseshell, Tortoiseshell-and-White and Blue-Cream.
In
a study comparing the genetics of several breeds (Lipinski MJ, et
al., The ascent of cat breeds: genetic evaluations of breeds and
worldwide random-bred populations, Genomics (2007), the Persian breed
was more closely related to random bred cats of Western
Europe/America than to random-bred cats from the Near East. Their
study also found the Persian to be derived from multiple lineages.
This accords with breed histories describing it as derived from
Turkish, Russian and British longhaired cats. Although Lipinski's
team were surprised by the result, it did not come as a surprise to
those versed in Persian breed history!
The
modern Persian has evolved to have longer hair, a denser undercoat, a
wider, flatter face, shorter muzzle, smaller wide-set ears, shorter
body and tail and large round eyes. The lighter-bodied type of the
early Angoras was lost and the modern Persian bears little or no
resemblance to the original imports. However, the early ancestors of
the Persian are once more being recognised in the form of the Turkish
Angora and the Siberian.
In
Britain, the cats became known as Longhairs, though most still call
them Persians or Persian Longhairs to distinguish them from other
long-haired breeds. In the USA, Persian is the official breed name
although some American associations do not recognise the chocolate or
lilac solid (self) colours introduced via Himalayan (Colourpoint
Longhair) breeding programmes; these are known as Kashmirs. In some
American registries, non-colourpointed cats of Himalayan origin are
called the Himalayan Reflections or Self Himalayan. Thankfully some
have seen fit to sort out the muddle so that Persians and Himalayans
can be interbred and some offspring are known as Colourpoint Carriers
(CPCs). The history of the Himalayan is in Colourpointed
and Masked Cats.
A 1950s colourpoint longhair and the 1990s equivalent
The
British system had its own perversities - each variety of Longhair
being regarded as a separate breed, not as a colour variation of a
single breed. The Chinchilla is therefore seen as a different breed
to the Blue Longhair! Conformation does differ across the Longhair
"breeds" and some of the Chinchillas remained less
ultra-typed than the selfs. Chinchillas were reckoned to be several
years behind other Persians in terms of extreme-typing.
During
the mid 1970s and early 1980s, American Persians changed
dramatically. The pre-1980s look had heavy brows, flat-topped heads
rather than the domed heads. The "sweet, open-expression"
was lost as fanciers pursued the extreme head type (ultra-type).
These were dubbed "pigs" or as having a "piggy
expression." The nose became
narrow and ultra-high; the breaks
above the eyes were
moving upwards into the foreheads; the eyes were
tiny and the jaws often maloccluded to produce a frowning mouth. It
is suggested that Peke-Face reds Persians were bred to other colour
Persians to produce the piggy look and the teardrop-shaped eye of
modern ultras is attriubted to the Peke-face cats. The new look was
deemed an exciting development and open-ended standards favoured
these cats on the showbench. During the 1990s, both open-face and
Persian pigs were advertised, but the trend is now back to the
healthier open-face cats.
True
Peke-face Persians (solid red and red tabby) generally conformed to
the red Persian standard, but only 98 had been registered with the
CFA beteen 1958 and 1993 (the 3 registered between 2000 and 2002 may
not have been genuine Peke-face Persians). They had higher ears and a
different skull structure that produced a very round head with a
strong chin and very wide-set eyes. The nose was depressed and
indented between the eyes. The muzzle was wrinkled and there was a
horizontal break located between the usual nose break and the top
dome of the head. This second break created half-moon boning above
the eyes and an additional horizontal indentation in the center of
the forehead. The Standard for the Peke-Face called for a brow ridge,
dimple, and a double dome. The term peke-face is often incorrectly
used to describe extreme-type Persians.
While
American breeders were the first to breed the Persian to extremes,
practically eliminating the cat's muzzle, British breeders are now
following suit with ultra-typed Persians which find favour on the
showbench but less favour with the general public who prefer the
older, less extreme facial conformation. Personally I find the
ultra-typed Persian ugly and they are more prone to breathing
problems and tear duct problems than those of more moderate type. In
the USA, the older type is now preserved as a traditional style
Persian (bizarrely the breeder does not like the actual breed name
used in reviews in case it erodes, rather than promotes, the breed).
For a short while, American breeders developed the Peke-Face Persian.
This arose as a mutation in red/red-tabby Persians and suffered from
breathing problems, tear duct problems, high palate (making suckling
difficult) and sometimes the head was too massive for kittens to be
born naturally (Caesarian section needed). These have now vanished,
but the ultra-types are becoming almost as extreme.
In
1872, Dr Gordon Stables, in his review of cat shows, had noted that
"urbanity of countenance" should not be overlooked when
judging Red Tabbies; the modern equivalent would be "a pleasing
expression". In 1981, Phyllis Lauder noted that the Peke-faced
Persian (bred only in the USA) was a solid red or red tabby longhair
(there being no Peke-faced Exotic Shorthair equivalent) with a very
different expression to that of a regular Persian. Like Pekinese
dogs, they had prominent eyes, sometimes described as "owl-eyed".
It was debatable whether these cats (or indeed the modern
ultra-types) could be said to have a pleasing expression.
The
Persian is still
evolving . The Mink Longhair (Mink Persian) is a
Persian Longhair in the Tonkinese (mink) colour series. Similar cats
have been experimentally in different countries under a variety of
names and since about the 1960s. They are attractive cats, but have
yet to attract a serious following or achieve formal recognition.
Other names used for this variety includeBurmalayan, Himbur, Iranese,
Layanese, Silkanese, Tibetane/Tibetaan and Tonkalayan (some resemble
Persians, other resemble Tonkinese). Mink Persians would seem a
logical and attractive development. The Napolean Cat Longhair is a
short-legged Persian bred through outcrossing Persians to Munchkins
in the 2000s.
To
add confusion to the Persian story, there is also the Exotic
Longhair. Longhaired individuals are sometimes born to Exotic
Shorthair parents due to the recessive nature of the gene for
longhair. In many registries, they are not accepted in the Persian
class due to their non-Persian parentage. In addition they may have a
relatively poor coat compared to Persians born of Persian parents.
However, they are accepted as Exotic Longhairs by some registries in
the USA/ The original proposed name was
Tiffany , due to the original
conception that the Exotic Longhair was to be allowed in silver
colurs only.
Russian
LonghairsHarrison
Weir, founding father of the cat fancy and organiser of the first
formal cat show (also writer of the first standards for pedigree
cats), wrote about the Russian Longhairs in "Our Cats" in
1889. The cat he described differs from the modern Siberian cat, for
example it was woollier, although cats of the old Russian Longhair
type apparently still exist among domestic longhairs in parts of
Russia.
The
Russian Longhair male described by Weir differed from the Angora and
the Persian in many respects, being
"larger
in body with shorter legs. The mane or frill was very large, long,
and dense, and more of a woolly texture, with coarse hairs among it;
the colour was of dark tabby, though the markings were not a decided
black, nor clear and distinct; the ground colour was wanting in that
depth and richness possessed by the Persian, having a somewhat dull
appearance. The eyes were large and prominent, of a bright orange,
slightly tinted with green, the ears large by comparison, with small
tufts, full of long, woolly hair, the limbs stout and short, the tail
being very dissimilar, as it was short, very woolly, and thickly
covered with hair the same length from the base to the tip, and much
resembled in form that of the English wild cat."According
to Weir, its motion was less agile than other cats and it did not
care for warmth, preferring to be outdoors in the coldest weather.
Another peculiarity was that it seemed uninterested in hunting
birds .
Its habits were not like those of its companion shorthaired cats. It
attached itself to no person, but was inseparable from a shorthaired,
silver-gray tabby female. This pair produced a single black-and-white
kitten. The kitten inherited the woolly coat, had somewhat of a mane
and a short bushy tail. It also seemed uninterested in birds, but
would attack rats!
"I
have seen several Russian cats, yet never but on this occasion had
the opportunity of comparing their habits and mode of life with those
of the other varieties; neither have I seen any but those of a tabby
colour, and they mostly of a dark brown. I am fully aware that many
cross-bred cats are sold as Russian, Angora, and Persian, either
between these or the shorthaired, and some of these, of course,
retain in large degree the distinctive peculiarities of each breed."
However to the practised eye, it was possible to distinguish whether
the cat was Angora, Persian of Russian. Unfortunately, these were all
judged together in a longhaired class and not as distinct breeds;
something which ultimately led to the loss of the Angora and Russian
types.I
have seen some 'first-cross cats' that have possessed all, or nearly
all, the points requisite for that of the Angora, Persian, or
Russian, while others so bred have been very deficient, perhaps
showing the Angora cross only by the tail and a slight and small
frill. At the same time it must be noted, that, although from time to
time some excellent specimens may be so bred, it is by no means
desirable to buy and use such for stock purposes, for they will in
all probability 'throw back' - that is, after several generations,
although allied with thoroughbred, they will possibly have a little
family of quite 'short-hairs'. I have known this with rabbits, who,
after breeding short-haired varieties for some time, suddenly
reverted to a litter of 'longhairs'; but have not carried out the
experiment with cats."Weir
noted that he had never seen tabbies among the shorthaired Russian
cats, but the Russian Longhairs were all brown tabbies except for two
black Russian Longhairs, which he supposed were the offspring of
tabby or grey parents. He did not recall having seen any white
Russian Longhairs and wrote
"I
should feel particularly obliged to any of my readers who could
supply me with further information on this subject".In
1926, Dr Jumaud's book "Les Races des Chats" (The Breeds of
Cats), which was based largely on the works of Professor Cornevin of
Lyons, described the Carthusian cat (felis catus carthusianorum) and
Tobolsk cat. The Carthusian was apparently the "Maltese cat"
known the the Americans, though Jumaud's description referred to a
large head with large, full eyes, short nose and small, erect ears.
Its coat, he said, was half long and woolly and the colour was grey
with bluish reflections. However, there was another variety of
Russian cat known as the Tobolsk variety: "This variety,
described by Gmelin, exists in
Siberia , and is sometimes called the
Tobolsk cat. It is larger than our common cat, and somewhat resembles
the Carthusian in shape. The head is large, with big eyes, short
nose, and small erect ears. Coat: as is fitting for an animal of a
cold country, the Tobolsk cat has long fur, longer than that of the
Chartreuse cat. Its texture is woolly, and in colour, uniformly
reddish."
In
1927, Mrs Amy
Lawrence wrote "In the Natural History Museum
[South Kensington, London] there is an enormous cat which is said to
be a 'Russo-Persian' cat. It has an immense coat, and is similar in
every way to a Persian long-hair, except that it is larger than any
specimen I have ever seen. An old
uncle of mine possessed what HE
called a Russian cat, also a long-hair with immense coat and very
large." However the only "Russian" cats Mrs Lawrence
had seen at cat shows was the small short-haired Russian Blue that
looked like a blue Siamese cat! Her uncle's huge Russian cat had been
a tabby.She wondered "Do Blue Russians really come from Russia,
and if so, then where do those immense long-hairs come from, and why
were they called Russians even by Museum authorities?"
NATURAL
LONGHAIRS: MAINE COON, NORWEGIAN FOREST CAT, SIBERIAN, GERMAN
LONGHAIRIn
hotter climes, a short sleek coat is an
advantage e.g. Siamese,
Burmese. In harsh conditions, longhair cats fare better - they are
insulated against the cold and their outer coat may have water
repellent qualities. "Refrigerator Cats" were a strain
allegedly developed in 19th Century Pittsburgh to control vermin in
refrigeration plants, but actually the product of a newspaper story.
Natural selection supposedly produced a race of "Eskimo cats"
which were at home in the cold, having heavily furred coats, thick
tails like Persians and tufted, lynx-like ears. Although the idea of
natural selection favouring thick fur for a harsh climate is sound, a
little investigation shows that no such race of cats existed.
The
American Maine Coon arose through natural selection in conditions
which favoured robust, longhaired cats. Accounts suggest that some of
the cats taken to America were longhairs e.g. Turkish Angoras i.e.
the longhair gene was imported into America. These would have
interbred with the various other cats taken there and over many
generations the harsh winters would have selected in favour of
longer, thicker, more protective coats.
In
Europe, the same natural selection process gave rise to the Norwegian
Forest Cat, the Siberian and the Rugkatt. To the casual observer
these resemble the Maine Coon, but the conformation and fur type
differ. A "Russian Angora" cat has been described as
similar to the Turkish Angora, but with green eyes i.e. naturally
occurring semi-longhaired version of Russian Shorthairs (the group
comprising Russian Blues, Whites, Blacks etc).
Lipinski
et al found the Norwegian Forest Cat, Persian, and Siberian each
showed subdivisions within each breed, indicating multiple lineages.
This is not surprising in the Norwegian and Siberian as these come
from a broad base of random-bred cats that have relatively recently
been recognised as a breed.
Maine
CoonThe
Maine Coon, or Maine Cat, is one of the oldest natural breeds of
North America, and is regarded as originating from the state of
Maine. Is has also been known as American Longhair, American Shag,
American Forest Cat, American Snughead and Maine Trick Cat.
There
are various explanations of its origin. One is that it is a
racoon/cat hybrid. Another is that Marie Antoinette sent her Angora
cats to safety in the USA and that these cats interbred with the
shorthaired domestics. Another is that New England sailors took home
Angoras from Turkey in the late 17th century. Due to its tufted ears
and large size (though not as large as some media reports would have
us believe), others believe that the cats descend from North American
bobcats or bobcat/domestic cat hybrids or, even more implausibly, as
a hybrid between domestic cats and lynx. The misconception that it is
a lynx hybrid is unfortunately still perpetuated by some credulous
cryptozoologists.
Most
likely, it derives from a mix of longhaired and shorthaired cats
taken to New England by colonists and as ships' ratters. The rugged
longhaired cats of Scotland,
Norway and Russia are good candidates
for some of its ancestry with the addition of Persians and Angoras.
In the late 18th century Maine was a major
ship -
building , sailing and
trading state. Trading ships would have carried a variety of animals
including European cats, both as pets and as ships’ ratters and
mousers. The Maine Coon would have evolved from these.
Maine
Coons were well-established by the early 1800s and had evolved into a
hardy, handsome breed of domestic cats and excellent hunters. They
had a rugged coat and build and were tough enough to withstand the
harsh winters. They were also large - both tall and long-bodied. The
long, flowing fur is relatively heavy and shaggy, shorter on the
shoulders and longer on the belly and tail. Maine Coons also have a
well-developed ruff, broad muscular chest, strongly boned legs and
relatively long, square-muzzled head with slightly concave profile.
They
were first recorded in cat literature in 1861 and became popular
competitors at early cat shows in Boston and New York. A Maine Coon
won the 1895 Madison Square Gardens show. It was described in "The
Book of the Cat" in 1903. And by 1906 there were 28 registered
Maine Coons, but interest was waning. The Maine Coon's popularity as
show cats declined when Persians arrived and though they remained
popular as pets, they were largely ignored by cat fanciers and
breeders until the early 1950s. In 1967 they were recognised as a
breed (1976 by the CFA).
The
first Maine Coon in Europe was a pregnant female taken to Austria
from Canada in 1953 or 1954 and her progeny were known in Germany as
American Forest Cats. They reached Britain in 1984 and are now
popular in Britain, Europe and Australia.
The
are found in almost all patterns and colours; the only ones not
permitted are those indicating hybridisation with colourpointed cats
i.e. chocolate, lavender or Siamese-pattern. Brown tabby is the most
popular colour. Polydactyly was found in early Maine Coons but was
discouraged. In the late 1990s, some breeders became interested in
reinstating the trait.
The
early Maine Coons were documented by Frances Simpson who was an early
champion of longhaired cats. In "The Book Of The Cat"
(1903) she included a chapter about Maine Cats from American breeder
F. R.
Pierce . In childhood (1861) Pierce had owned a longhaired
black-and-white, "
Captain Jenks of the Horse Marines" and,
like Simpson, was evidently a longhair enthusiast. Pierce did not
know Jenks' ancestry, but assumed that longhairs entered Maine (a
major ship-building and seafaring region) much in the same way, and
at about the same time, that they reached England. The major
difference was that the Maine cats were largely left to their own
devices while the British cats were being selectively bred.
In
1869, Pierce saw a pair of blue-eyed white Persian kittens that
landed from a foreign
vessel which had put into a seaport town for
repairs. These had been acquired by a sailmaker making repairs to the
ship from the cook who owned a Persian female which had produced
kittens. The two cats were both kept for 2 or 3 in the hope of
getting a good male for neutering (as a pet); all the female kittens
being destroyed! When the desired male arrived, the original pair
were sent to a relative in the country. However, during those 3 years
they had evidently met up with local cats since longhaired blue-eyed
white kittens began to appear in unexpected places. Lack of selective
breeding meant the strain generally vanished, only to reappear later
on. Pierce owned ones such cat, Dot, said to be as good a specimen of
Persian as the one that came from the original kittens eleven years
previously.
One
Mrs
Thomas , also of Maine, wrote that her cat was descended from a
blue-eyed white brought to Rockport, Maine on a ship from France.
That line of whites, while in the same locality, was quite distinct
and unrelated to Pierce's white longhairs. Pierce wrote on a little
island well off the coast and inhabited by only three families, there
were pure white blue-eyed Persian cats, but was apparently unable to
obtain one of these cats.
Another
early champion longhair was Richelieu, owned by Mr. Robinson, of
Bangor, Maine in 1884. Richlieu was described as a silver or bluish
tabby, very lightly marked, but rather a coarse-grained variety - "a
drug store cat" (moggy). At that time Maine, near the coast, had
many fine specimens of the longhaired cats, particularly brown
tabbies. The Maine cats were not considered valuable at the time.
From the coastal towns and cities, the longhaired cats spread inland.
Around 1895/6 the "cat fad" struck the Middle West and cats
from Maine were being acquired by enthusiasts inland of Maine, with
considerable sums being paid even for poorer quality or mongrel
kittens.
Many
of the prize-winning Maine cats of the mid-to-late 1890s were
described as being of Persian type. At the turn of the 20th century,
smokes, silvers and chinchillas were uncommon. The most common
colours were whites, blacks, blues, oranges and creams, plus tabbies.
A line of creams was founded by a fine cream apparently brought from
an unspecified Mediterranean port by one Captain Condon in the 1880s.
The Maine cat was distributed along the coast, and for about 60 miles
inland, but were not then common in the less populous northern
portion of that State.
Norwegian
Forest CatAlthough
in some
ways it resembles the Maine Coon, the Norwegian Forest Cat
(Norsk Skaukatt or Skogkatt) is a Scandinavian breed which evolved in
the cold northern climate of Norway. Generations of living in the
cold and wet gave rise to a cat with a heavy, weather-resistant coat
and full ruff. The woolly undercoat provides warmth while a
medium-long, glossy outer coat resists rain and snow. It differs from
the Maine Coon in several respects - including back legs slightly
longer than the front. It is an excellent climber.
Longhaired
cats are mentioned in Norse mythology and in
books of Norwegian fairy
tales written 1837 and 1852 which describe it as having a long, bushy
tail. Due to its resemblance to the Maine Coon and Scottish Wildcat
some have suggested the Vikings took cats to Scotland (where they
interbred with wild cats) and to North America on
Viking longships.
This theory is unlikely. Domestic cats did not
arrive in America
until European colonists arrived; there is no archaeological evidence
of domestic cats in supposed early Viking
sites in America and
reports of native cats almost certainly referred to the racoon. The
Scottish Wildcat has an intractable temperament and cannot be
reliably domesticated, hybrid offspring inherit this wildness.
Other
theories suggest it derived from Angora cats which arrived at
Norwegian
ports as ships’ cats and interbred with native Norwegian
shorthairs, or that Crusaders took British shorthair domestic cats
and longhaired cats to Norway. There is a misconception, fostered by
credulous cryptozoologists, that it is a cross between domesticated
longhairs and Scottish Wildcats. Norwegian longhairs would have to
swim a long way to meet up with Scottish Wildcats! TIt is not a
hybrid with European Wildcats nor with European Lynxes. Crosses
between domestic cats and European Wildcats are closer in type to the
wildcats - wild in temperament and shorthaired. Most likely, it
evolved naturally as an adaptation to harsh wintry conditions.
Breeding
of the pedigree Norwegian Forest Cat from semi-wild outdoor cats and
farm cats began as early as the 1930s, with the cats being exhibited
in Oslo before the Second World War. In 1963 it was shown under the
name Skogkatt. The breed was revived in the early 1970s; in 1972 it
was formally recognised in its home country, serious breeding began
in 1973.
Many
of the foundation cats came from near the Swedish border. Swedish
breeders declared that the breed belonged to Sweden as much as to
Norway. But the Norwegians disputed this and it was agreed that
foundation cats must come "straight from the Norwegian forests".
While single Skogkatts might cross the Swedish/Danish border while
hunting in the woods no entire (unneutered) registered cats were
allowed out of Norway until the breed was recognised. It was
eventually allowed to be exported and was recognised in Europe in
1977 and became popular in the USA during 1985 and Britain in 1987.
Siberian
and other Russian LonghairsIn
the 19th Century, Russian Longhairs were described as distinct from
the distinct from the Persian or Turkish Angora; and these are
described earlier in this article as they contributed to the Persian
type. The Russian cats had a larger body with shorter legs, a woolly
coat with coarse hairs among it, a large mane and short thickly
furred tail. In Victorian times, it was extensively crossed with the
Persian and the Angora and was lost as a distinct type. It continued
to breed naturally in Russia as the Siberian cat. The Russian Angora
is described as similar to the Turkish Angora, but with green eyes
instead of blue, orange or odd eyes.
The
first Siberian cats (Russian Longhairs) were imported into the USA in
1990 by Elizabeth Terrell of Louisiana, in exchange for Himalayan
cats sent to establish that breed in Russia. Before 1990, it was
almost unknown outside of Russia. The first colourpoint Siberians
(Neva Masquerades) were imported into the USA in 1997 and registries
are have recently accepted it following early controversy. The
history of the Neva Masquerade is in Colourpointed
and Masked Cats.
Cats
would have entered Russia from Europe, by land and sea, from Persia
(Iran) by overland trade routes and from the Far East on
board trading ships. The existence of bobtailed Russian breeds demonstrates
that cats from south east Asia reached Russia one way or another. The
cats of Russia and Siberia are little changed from mediaeval times,
perhaps earlier.
It
is suggested that the longhair mutation occurred in Russia and spread
from there to Turkey (Angoras) and Persia (Persians) or even that all
long-haired breeds ultimately have their origins in Russian cats. The
similarities between the Siberian and the Norwegian Forest Cat and
Maine Coon suggests a common origin and some researchers suggest that
the Siberian is the ancestor of both the Norwegian Forest Cat and the
Maine Coon. Equally, it could be parallel evolution where similar
environments have led to unrelated cat populations evolving similar
traits through natural selection.
The
Karel Bobtail (Karelian, Karellian) and the Kuril (Kurilian, Curilsk)
are both shorthaired/semi-longhaired Russian breeds with bobtails.
The American-bred Nebelung is longhaired version of the Russian Blue,
developed through outcrossing Russian Blues to other cats. Longhaired
Russian cats other than the Siberian exist naturally in their own
country. In 1993/4, the first Nebelung in the Netherlands arrived
from Russia. A purebred Russian Blue stud cat called "Timofeus"
turned out to be semi-longhaired. Although not found in modern
Nebelung pedigrees, Timofeus confirmed that the recessive longhair
gene was present among Russian Blues. In the following years, it
turned out that semi-longhaired Russian Blues were not uncommon in
Russia and some have become part of Nebelung breeding programmes in
other countries.
German
LonghairThe
German Longhair has waited a long time for international recognition
although a breed standard and scale of points has existed since 1929.
All that was missing until a few years ago was a registered breed
that corresponded to this standard. Following the 2nd World War,
which interrupted cat breeding in Germany, the only native German
longhaired cat breed was considered extinct.
Longhaired
cats in Germany were generically called Angoras and bred for colour,
not conformation. Biologist and zoologist Professor Dr
Friedrich Schwangart (1874-1958) criticized them as generally not meeting the
"Hochzuchtperser" ("high-bred Persian") standard
seen in British Persians, hence he created separate standards for the
Persian and the German Longhairs in 1929, describing the differences
between the two types. From that point, breeders of "Angoras"
had to decide whether to breed British-style Persians or more
natural-looking German Longhairs. Schwangart hoped the German
Longhair, with its silkier "wash and wear" hair would take
its place as a more natural counterpart to the Persian that had been
bred in Great Britain for decades. The German Longhair was first
exhibited and acknowledged nationally at the Exhibition of the
Federation for Cat Breeding and Protection in 1930 in Berlin. In the
following years it was frequently seen at cat shows and in 1932,
German Longhair "Fox of the Rhine Castle", owned by Dr
Heine in Leipzig, became a Federal winner.
With
the standards laid down in 1929, the longhaired cat types fell into 2
categories in the "Classification, Pedigrees and Systems of
House Cats" and these were not to be interbred in order to
maintain their distinct types (in Britain, early Persians, Angoras
and the native British Longhairs had been interbred to create a
single Persian Longhair). There was more information in Schwangart's
1932 publication "Formation and Breeding of House Cats (Results
and Problems)" which noted further longhair breeds being the
German Longhair founded by Schwangart himself and, in the previous 2
years, the Burma breed that appeared in Paris (i.e. the
Khmer/Birman). By the day's standards, both Persians and German
Longhairs had a compact conformation, short sturdy legs, a broad head
with relatively short, broad muzzle and moderately small ears (akin
to the European Wildcat). Both had rather short, beautifully carried
tails, level back and long fur (with age differences, seasonal coat
and pregnancy to be taken into consideration). And in both cases a
"half-Angora" type with svelte body or narrow, pointed face
were undesirable.
However,
the Persian was described as thicker-set with a rounded head and a
prominent forehead that fell abruptly to a broad, short muzzle giving
an "angry" expression (i.e. a shorter face). The Persian's
fur was denser and woollier with a well-developed ruff and the cats
were bred with cobbiness and size in mind. In contrast to the
Persian, the German Longhair had a more moderate head: a less
prominent, tapered forehead that curved gently up from a longer nose
with a more gentle slope. The conformation was less compact, the
movement more fluid and the tail longer than the Persian. In essence,
German Longhair did not permit the short face and prominent forehead
of the Persian and in profile the face resembled the Tabby Shorthair.
The German Longhair was found in the same colours and patterns as the
Persian: single/self colours (black, blue, cream, red and white),
bicolours, tortoisehsells (with or without white), "masks"
(colourpoints), smokes, Chinchilla (tipped), peach (goldens?),
silvers and both "tiger" (mackerel) and "marble"
(classic) tabbies.
In
"The Formation and Breeding of House Cats (Results and
Problems)" (1932) Schwangart suggested the head and face of the
German Longhair showed the influence of the large
Nordic form of F
silvestris (European Wildcat) resulting in a native Longhair that was
distinct from the Persian or Angora. The tiger pattern completed the
image of a German Longhair that might trace its ancestry, in part, to
a wild cat. It was already known that domestic cats and wildcats
could interbreed and some still believed that local races of domestic
cat had arisen independently from local wildcat species. In a last
work "
Overview and Description of Domestic Cat Breeds"
(1954) Schwangart described the German Longhair in detail, noting the
existence of intermediate forms between Persian and German Longhair
which were found in some of the colours, and the need to eliminate
the intermediates in order to restore the 2 breeds as distinct form
each other. It's clear that the Persian had been bred together,
perhaps due to the difficulties of maintaining breeds during wartime,
perhaps to improve the traits of one or other breed or perhaps
through ignorance that they had originally been separate breeds.
He
elaborated on the breed standard, though by then he may have felt it
a losing battle due to the increasing popularity of the Persian. In
the solid-colour German Longhairs, amber/yellow was the preferred
eye-colour, except in solid white cats where amber, blue or odd-eyes
were permitted. Deafness was a disqualifying fault in white cats
which were to be tested using a whistle out of the cat's sight. He
also mentioned the potential for degenerative problems, such as
deafness, related to "albinism" (blue-eyed white was
mistaken for albinism) so some indication that the cat wasn't albino,
such as a dark membrane, was desirable. The bicolour and tricolour
cats were to be more colour than white. The "masked" cats
were allowed to be less symmetrically marked than bi- or tri-colours.
This group included the "black and yellow" tortoiseshell
and the "Spanish" (tortoiseshell and white). The
tortoiseshells ideally were to have large patches of colour, but
Schwangart admitted that this was rare. In
parti -colour cats, the eye
colour was to reflect the predominating fur colour. The eye colours
of the Chinchilla (black-tipped), peach colours (goldens?), smokes
and silvery ones related to their coat colour (i.e. paralleling
shorthairs and Persians).
In
May 1935 the German Longhair was officially allowed to be bred under
the auspices of the "Katzenverein des Deutschen Reiches"
(Cat Club of the German
Reich ), which was the only breeding club at
this time. It was grouped in the longhair class together with the
Persian and Birman. In October 1939 it was recognised by the
Confédération Internationale Féline (CIF) as "Borealis"
or "Boreali" ("Northern"). The CIF. was the
predecessor of the Fédération Internationale Feline (FIFe) and had
been founded by the Societa Felina Italiana, the Cat Club of Paris
and the Fédération Suisse. The Second World War interrupted the
breeding programme and the German Longhair stagnated for several
years before apparently dying out. Dagmar Thies reported in 1979 that
Mrs R Aschemeier had managed to locate German Longhairs from original
bloodlines and had bred them at Blasheimer mill since 1968. These
cats were considered very typy representatives of the breed and their
descendents were useful in re-establishing the breed. By 2005 there
were a growing number of breeders interested in preserving or
recreating the German Longhair. They found foundation cats among
free-ranging farm cats that were close to Schwangart's German
Longhair standard. The foundation cats 5 of the remaining German
Longhairs descended from Mrs Aschemeier's cats from 1968 (and thus
preserving some of the genetic make-up of the original breed).
A
provisional German Longhair standard was registered with the World
Cat Federation in 2008 and based on the 1929 and 1954 standards. It
is the only longhaired cat developed on German ground and is the
longhaired "sister" of the European Shorthair breed which
it resembles in general conformation. it does not have the wide
muzzle of the Maine Coon or the straight nose line of the Norwegian
Forest Cat. The modern standard calls for a medium-size cat with a
long, rectangular, robust and supple figure. It differs from the
European Shorthair/Celtic Shorthair in having a deeper chest and
medium-length bushy tail that tapers to a round tip. The sturdy legs
are short to medium-long with large firm paws. The head is rounded,
but is longer than it is broad with medium-long and sloping nose with
slight stop (a pronounced stop is a fault). Strong chin and cheeks,
the latter suggesting the Nordic race of European Wildcat (F
silvestris silvestris). Ears are small to medium size, upright and
broad at the base with a rounded point. Eyes are round to oval, large
and slightly diagonally set; the colour relates to the fur
colour/pattern (or to predominant colour in parti-colour cats).
The
coat is medium long at the shoulders and shorter on the head. It is
longer on the flanks, back and belly and is particularly long at the
ruff, hind legs (britches) and tail. However the fur is
easy-maintenance, shining and not as woolly as the Persian. All
colours are accepted except chocolate, cinnamon and their dilutes
lilac and fawn (in both solids and in patterned cats). The
colours/patterns otherwise include self/solid, bicolour,
tortoiseshell, tortie-and-white, "masked", tipped, cream,
red, smokes, shaded, silvers and both mackerel and classic tabbies.
The personality is human-oriented.
At
the end of April 2012, it was formally recognized as the "
Deutsch Langhaar" (German Longhair) by the World Cat Federation (WCF) at
the general assembly with a revised standard that still is
nevertheless still based on Schwangart's description. The recognition
comes into effect at the start of 2013.
A
second longhaired German cat is the German Angora bred from
semi-longhair housecats and originally bred by Mrs Aschemeier who
retired from breeding in 2010 (Aschemeier always referred to her cats
as German Longhair, not as Angoras,
stating "Being experienced
in
keeping studbooks for hunting dogs from now on I kept records on
the progeny of my German Longhairs, so I could provide them with
pedigrees at any time"). The German Angora, bred since 2000
(breed club founded 2005), is not recognised by any cat association
and is trademarked instead. Breeders of the German Angora claim there
are no genuine German Longhair cats because they became extinct. They
describe the German Angora as a naturally occurring longhair in the
same way that the "Forest Cat" breeds are natural
longhairs. Until 2007, when German Longhairs were recognised, German
Angora and German Longhair were synonyms for one breed; the longhairs
that began in 1929. In 2007 the Board of Directors of the first
German Angora Cat Club dismissed this idea, resulting in a dispute
that divided the breeders' group. The German Angora became
trademarked and may only be bred in the 1st German Angora Cat Club.
It has not sought international recognition. Meanwhile, the German
Longhair is bred to conform to the old image. Both breeds are
described as very similar and both have been crossed to old-style
Persians to improve the conformation and coat.
THE
IRCA LONGHAIRS - RAGDOLLS, HONEYBEARS AND CHERUBIM CATS (RAGAMUFFINS)Baker's
Cherubim Cats included Ragdolls, Miracle Ragdolls, Honey Bears, Doll
Babies, Baby Dolls, Shu Schoos, Catenoids and Little Americans among
the fanciful names. The origins included fanciful claims of
secret government laboratories, infusions of skunk genes, human genes and/or
alien DNA while a Catenoid would supposedly produce Ragdolls
regardless of what you mated it to. Baker seemed increasingly
unstable; she distributed photos of herself amidst a hundred dead
kittens that she claimed were killed by rival breeders who broke into
her home. Disturbed cat fanciers were certain she had killed them as
part of a publicity bid and tried to figure how to get other cats
safely away from her (I recall the
pleas for information circulating
on mailing lists at the time). Perhaps mercifully for the cats, Baker
died in 1997. Her IRCA
organisation limped along for a few years, but
many breeders
quit or defected to conventional registries (which had
cat shows) and the IRCA trademark on Ragdolls lapsed in 2005.
Former
IRCA Ragdolls, Honey Bears (IRCA's version of the Persian) and
Miracle Ragdolls, not being accepted by other registries, were merged
and renamed RagaMuffins and registered under that name. Although bred
with the Ragdoll to begin with, the modern RagaMuffin has a shorter
nose with an obvious nose dip, and the eyes are walnut-shaped rather
than oval. RagaMuffin breeders have worked to produce a rounded, more
heavily boned cat by outcrossing to Persians/Himalayans and have used
domestic longhairs to give the breed a sound genetic footing. The
plushy coat is shorter and thicker than the Ragdoll's medium-long,
silky coat. Some registries only recognise non-pointed RagaMuffins
(feeling pointed varieties are to similar to Ragdolls), others also
recognise the pointed varieties.
Honey
Bears had been bred from a pair of Persian cats and could be
regidtered as Persians with the CFA. Ann Baker claimed skunk genes
were infused into the female and her kittens then resembled young
skunks , being born silver before turning black with stripes along the
head, back or underside (which to the rest of the cat world sounds
like shaded silvers developing their colour). The cats supposedly has
tails flatted on the sides (actually an illusion caused by long fur!)
that they held over their backs like skunks. Baker claimed the Honey
Bear looked like a Persian, but did not have a cat skeleton.
Essentially the cats were Persians and were even marketed as
non-matting Persian lookalikes, but Baker's breeding philosophies and
increasingly wild claims, led to them becoming unregistrable as
Persians with conventional registries. For all practical purposes,
Honey Bears were Persians, albeit less extreme in type than those
seen on the showbench. A few breeders persevered with them, but
otherwise they were absorbed into the RagaMuffin.
Miracle
Ragdolls ("highly upgraded Ragdolls") were mentioned in
some of Baker's IRCA advertisements and were also merged into
RagaMuffins after her death. Her press releases became increasingly
vitriolic towards the breakaway Ragdoll breeders and their
"half-bred" or "overbred" copies of her cats.
Catenoids and Little Americans also appeared in IRCA adverts close to
the end of Baker's life. Most of the other "breeds" seemed
to be products of a fertile imagination rather than distinct types of
cat.
SEMI-LONGHAIRS
(INTERMEDIATES)Semi-longhair
means intermediate between the thick, flowing coat of the
Persian/Longhair group and the short fur of the various Shorthair
breeds. By modern standards the parent of modern Persian longhairs,
the Turkish Angora is a semi-longhair. Semi-Longhair cats have coats
slightly shorter than the Persian, often lacking the undercoat of the
Persian. They range in type from the Oriental (e.g. Balinese) to more
robust types (Norwegian Forest Cat, Siberian) though all are more
moderate in type than the modern Persian.
According
to Phyllis Lauder, writing in "The British, European and
American Shorthair Cat" (1981), longhaired cats had long existed
among the non-pedigree population in the West - before Angoras and
Persians had been imported from the East - though they lacked the
tremendous length of the imported and carefully bred pedigree cats.
The prized "fluffy cat" found in the native population was
referred to by fanciers as an "intermediate" and it was the
considered opinion of breeders that the exhibition longhairs were
derived through crossing "intermediates" with imported
Persians and Angoras. Australian geneticist Mary Batten believed that
the fluffy moggy got his pretty coat from either the indigenous
Scottish wildcat or from cats imported from the Middle East by the
Romans (it is more likely to have come from later Viking imports).
She wrote "Almost certainly the factor which has produced the
'fluffy' coat is common to both sources". In fact the Scottish
Wildcat is not an ancestor of the modern domestic cat. Fluffy cats
had been present in the west for far too long to owe their existence
to relatively recent imports. Mrs Batten added that, in 1967, a
Tabby-point male bred by her was mated to a Chinchilla Longhair, and
produced kittens whose fur was of intermediate length, rather than
the expected shorthairs.
Lauder
noted that while pet classes were dominated by shorthaired cats, the
majority of the "longhairs" in pet classes were
"intermediates". By this she meant that they had
"Persian-type" fur as opposed to short, but the fur
bore no
resemblance to the tremendous pelage of the exhibition quality
longhair. She again noted that it was an unsolved question whether
the show longhairs were originally bred from cats imported from
Ankara or Iran (Persia), or whether they were the result of selective
breeding from the native "intermediates" that exhibited a
mutation for fur longer than that of the predominant shorthairs.
Lauder wrote "probably simply a variation - with a coat of
different length occurring in a predominantly short-haired
population. These 'intermediates' were much prized by their owners in
the early days of the century; people would say with pride 'He's
got
a
fluffy coat!"
Lauder added that though there had been surveys into cat colour
distribution, the provenance of the "fluffy" cats did not
seem to have been researched. According to Mary Batten: "It is
known that there are polygenes which influence shorthair coat-length.
These may also be present in longhair lines [...] It is arguable as
to whether the 'fluffy' cat received its coat-length from the Persian
or Iranian aristocrats or from the blending of cats brought from
these areas by Romans, and the indigenous Scottish wild cat. However,
almost certainly the factor which has produced the 'fluffy' coat is
common to both sources. It is
not an
allele of either longhair or shorthair, but is a separate gene or
polygenic series".
As
well as those domestic longhairs, "intermediates" or
"Semi-longhairs" arose in a number of breeds either by
spontaneous mutation or due to recessive genes introduced generations
previously. At first breeders sold the longhaired "sports"
as pets, or worse they destroyed the kittens so that other breeders
did not suspect any impurity in breeding lines. However, longhairs
continued to occur in these breeds and many have achieved recognition
in their own right. In other breeds, the longhair trait has been
deliberately introduced. A British Longhair known as the Lowlander is
now being bred (albeit not in Britain) which is less cobby and less
extreme than Persian Longhairs.
Some
have been bred to retain their natural characteristics (e.g. Birman,
Turkish Angora) while others are longhaired variants of shorthair
breeds e.g. Cymric (longhaired Manx), Scottish Fold Longhair,
Longhair Japanese Bobtail etc etc. Some are bred by crossing
different breeds: Tibetane/Tibetaan (longhair Tonkinese), Nebelung
(from Russian Blues and domestic longhairs).
In
some recently established shorthair breeds, recessive longhair genes
are now coming to light. The Bengal Longhair arose due to recessive
genes from foundation Abyssinian cats. It has a fluffier coat than
the Bengal and a plumy tail and distinct ruff. Although frowned upon
by Bengal breeders, some breeders are working with this as a distinct
variety. For the same reason, longhaired Ocicats appear in Ocicat
litters. The Australian Mist Longhair has also arisen due to
recessive longhair genes, again probably from Abyssinian foundation
cats. A Singapura Longhair has occurred, but the only
known
example was neutered. It was identical to the Singapura in all
respects apart from the semi-longhair coat which may have been due to
recessive genes or to a spontaneous mutation.
There
are many others, with new breeds appearing. Only a few are described
in any detail.
British
Longhair, Longhair Scottish Folds and Scottish LonghairLonghair
Scottish Folds are known as Coupari (after their place of origin,
Coupar Angus) in the UK and Highland Fold in the USA (although Coupar
Angus is not in the Highlands). It arose through matings with British
Shorthairs that carried a recessive longhair gene ("fluffies"
are sometimes born to British Shorthairs). It produces both
prick-eared and fold-eared variants. Prick-eared Scottish Folds gave
rise to the Scottish Shorthair in Queensland, Australia. It has
longer tail and legs, and different coat texture to the British
Shorthair. It also gave rise to the Scottish Longhair.
The
British Longhair (Britanica, Lowlander) is a cat with the British
Shorthair's conformation but a semi-long coat and arose from the
recessive longhair gene carried by some British Shorthairs and which
sometimes gives rise to "fluffies".
Colourpoint
Semi-LonghairsDescriptions
and histories of the Balinese, Birman and Ragdoll are
in Colourpointed
and Masked Cats.
Identical
in conformation to the Birman, but in different colours, is the
Tibetan, derived from Birman/Persian crosses when introducing new
colours into the Birman. It was recognised in Britain in 1986 and
should not be confused with the Dutch Tibetane (Tibetaan), a
longhaired Tonkinese cat bred in the late 1990s.
Various
mink-patterned longhairs and semi-longhairs have been bred during the
past few decades from Balinese/Burmese, Himalayan/Burmese,
Tonkinese/Persian or Tonkinese/Himalayan crosses . Some resemble
Persians, others resemble Tonkinese. The names used included
Burmalayan, Himbur, Iranese, Layanese, Silkanese, Tibetane/Tibetaan
and Tonkalayan. Although attractive, there has been insufficient
interest in perpetuating these.
Turkish
and Greek Semi-LonghairsApart
from the Turkish Angora, there are other longhairs originating from
Turkey and neighbouring Greece.
The
Turkish Van has existed near Lake Van in Turkey for centuries. In
1955 it was discovered by two British photographers who were given
two kittens and acquired three more later on. Recognition was
problematical at first since Turkey did not have a cat fancy and
there were no pedigree records for this naturally occurring breed.
While American cat fancies have mechanisms for accepting foundation
cats and developing naturally occurring varieties the British GCCF
appeared unable to cope with the physical reality of a cat unless it
was accompanied by a four generation pedigree! The Turkish embassy
provided documents stating that the cats represented a natural
Turkish breed, but this was not acceptable (even today it is a wonder
that new cat breeds ever get recognised in Britain). After being bred
by enthusiasts for the required number of generations, the "Turkish"
was recognised in Britain in 1969 and is now known as the Turkish
Van. A politically correct name for this cat is the Kurdish Van.
The
Van Kedi is an all-white Turkish Van originating from eastern Turkey
and should not be confused with the Turkish Angora. "Van Kedi"
is Turkish for "Van cat". In Turkey the self white Van Kedi
is prized and the auburn/white variety is held in less regard. The
most sought after is odd eyed white although blue eyed cats are also
considered special while amber eyed whites are the least sought
after. In Britain most matings are between an all-white cat and an
auburn/white to produce a mix of all-white and auburn/white offspring
(plus occasional cream/white offspring) with a mix of all three eye
colours. Outside of Turkey, the Van Kedi may be recognised as a
colour variant of Turkish Van rather than distinct breed.
It
is worth mentioning the Anatolian (Turkish Shorthair, Anadolu Kedisi)
which is a naturally occurring cat similar in type to the Turkish Van
with which it is allowed to breed. The Anatolian is found in all
natural colours, with and without Van markings. In the past, many
Anatolian cats were exported and registered as Vans or Angoras
although Dutch and German breeders are striving for purebred
Anatolians. The mistaken identity of Turkish Van suggests that they
produce semi-longhaired variants.
The
Aegean Cat is derived from naturally occurring cats of the Greek
Cycladic Islands. It is being developed by members of the fledgling
Greek Cat Fancy and is currently the only native Greek breed.
Selective breeding started in the early 1990s using native
semi-longhaired. These are of a light European/Continental type i.e.
neither cobby, nor oriental. The semi-longhaired is than that of
Turkish Angoras. All colours are found, with bi-colours
(colour-and-white) predominating.
Tiffany
(Chantilly), Tiffanie and Asian LonghairsThe
Asian Longhair (Tiffany, Longhair Burmese) is a longhair variety of
Burmese type and colour and was recognised in 1986. The British
breeding programme for Asian Shorthairs such as the Burmilla had the
side effect of bringing together the genes for long hair (from
Chinchilla Persians) and Burmese coat colour. This ultimately gave
rise to longhaired Burmese cats and to longhaired Burmese-type cats
in non-Burmese colours. At first the variety was known as the
Tiffany, but the Asian Longhair group encompasses a wider variety of
colours, paralleling that of Asian Shorthairs.
The
similarly named American Tiffany (Tiffany/Chantilly) is not related
to the British Tiffanie. It was developed in North America (late
1970s, early 1980s) from non-pedigree cats. It has a silky,
semi-longhaired coat in chocolate colour and superficially resembling
the Burmese, but is unrelated. At first they were thought to be
longhair Burmese so the name Tiffany was chosen in line with the
British cats. However Burmese kittens are born with lighter coats and
dark paw pads while Chantilly kittens are born dark with pink paw
pads. At first they were known as Foreign Longhairs, later as
Mahoganies, then Tiffany (in line with British Tiffanie) and later
Chantilly or Tiffany/Chantilly to reflect its non-Burmese origin. Its
exact history is not known, but it may have been a by-product of the
breeding program which gave rise to the British Angora Foreign
Longhair). They are recognised only in chocolate and lavender
colours.
The
Australian Tiffanie derives from the Burmilla breeding program in
Australia. Burmillas are Chinchilla /Burmese crosses. Subsequent
Burmilla-to-Burmilla matings may produce longhair kittens due to a
recessive gene. These are known as the Australian Tiffanie.
Oriental-Type
and Abyssinian-Type Semi-LonghairsThe
British Angora is known elsewhere as Javanese (reflecting its links
to the Balinese), Oriental Longhair and Mandarin. It may also have
been the ancestor of the American Tiffany/Chantilly. It was developed
from Abyssinian/Siamese crossings in 1973 which produced a chocolate
brown longhair with white roots to his fur although he was not
genetically a smoke. In 1974 it was remarked that he resembled the
old Angora cats. The breeding programme progressed slowly until it
was recognised in 1983. In 1984 it was decided that the Turkish
Angora and the British "Angora" were 2 distinct breeds. The
British "Angora" was improved to have true Oriental type
and in 1989 the Cat Association decided that "Angora" was
confusing since it was basically a longhaired Oriental. The name
Javanese was chosen since this name was already used in Europe for
Oriental Longhairs. It does, however. Clash with the American
Javanese which refers to red, tortie and tabby Balinese cats.
A
British breeder who had read reports on cat gene surveys in the late
1970s was inspired to recreate the coat patterns found in some cats
of the Seychelles. The breeding programme began in 1984 using two
tortie-and-white Persians and Siamese and Oriental cats. A breed
society was formed in 1989, but these cats remain rare. They are
essentially Oriental Longhairs with a pattern known to geneticists as
the Seychelles pattern and to cat-lovers as the Van pattern: white
body, coloured tail and splashes of colour on the head. Some have
small splashes of colour elsewhere on the body.
The
York Chocolate is an American breed
developed
in the 1980s/90s from domestic, non-pedigree cats. It is
distinguished by its semi-longhair soft, silky hair and chocolate
colouring. It has a long lean body reflecting some Siamese ancestry,
but is a large cat. In addition to solid chocolate, it comes in
chocolate and white bicolour, lavender (dilute of chocolate) and
lavender and white bicolour. It is rare in its home country, in part
because it is not considered distinctive enough.
The
Somali is a longhaired Abyssinian; the name reflects its link to that
breed. Longhaired "sports" sometimes occurred in Abyssinian
litters and one was exported to the USA as early as 1952. A
longhaired Abyssinian was exhibited in Australia as far back as 1965.
It was not until 1967 that they were bred in the USA in a deliberate
manner. In Britain, the longhaired variants were generally hushed up
until a breeder went public at a cat show in 1971. Always blinkered
in its outlook, the British cat fancy made a great effort to
eradicate the rogue longhair gene by clamping down on pedigrees in
which it had shown up. Somalis from the USA were imported into
continental Europe in 1977 although longhaired Abyssinians had
cropped up in litters prior to that. They were already being bred and
shown in Australia and New Zealand. Finally in 1982 (when the Somali
has achieved respectability overseas), the British cat fancy woke up
to the attractiveness of the Somali and cats from the USA were
imported.
The
Suqutranese was first shown in Britain in March 1990 and is a
sparkling white cat with Somali conformation, but nothing has been
heard of this variety since.
In
America, the Snow Cat (Alaskan Snow Cat) is similar to the silver
series of Somali recognised in the UK. It derives from crossed
between Silver Persians and Somalis and is intended to have heavier
boning, thicker fur and a rounder head than the Somali. The silver
Somali series of Abyssinian and Somali are both rare in the USA which
is very liberal when it comes to new conformations, but very
conservative when it comes to extending the colour range of existing
breeds. It seems that American cat fancies would rather recognise an
entirely new breed than allow new colours into an existing one!
Semi-Longhair
RexesTo
avoid unnecessary duplication, see Hairless
Cats and Curly Coated Cats for information on longhaired
cats which have curly hair. Some are due to recessive genes for
longhair in existing Rex breeds, others are due to the Rex mutation
spontaneously occurring in a longhaired breed. These include the
Angora German Rex, Bohemia Rex (Rexed Persian), LaPerm Longhair,
Longhair Devon Rex, Rexed Himalayan, Rexed Maine Coon. Combined with
Persian-type longhair and undercoat, the Rex mutation can be unruly
and unattractive - a Persian with a "bad hair day". With
semi-longhair or longhairs without a woolly undercoat, the fur is
soft, ringletted or wavy. In many cases, the curling of the hair
makes it appear shorter and denser than it really is.






































































































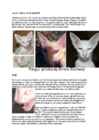




























































































Kõik kommentaarid