Füsioloogia eksami
küsimused1. Füsioloogia mõiste. Homöostaas.Füsioloogia on bioloogias
ja meditsiinis
õpetus organismi
ja selle elundite
talitusest ja funktsioonidest.
Homoöstaas on organismi sisekeskkonna suhteline püsivus.
Konstantsena hoitakse:
- glükoosi kontsentratsioon
- erinevate ioonide kontsentratsioon (nt. naatrium , kaalium , kaltsium )
- süsihappegaasi kontsentratsioon
- vee- ja osmoregulatsioon (vee ja lahustunud aine vahekord )
- temperatuur
- pH (happe ja leelise vahekord)
Füsioloogia on õpetus
elusorganismide talitlusest ja
nende seosest ümbritseva keskkonnaga. Talitlust ei saa mõista ilma
organismide ehitust uuriva õpetuse – anatoomia – aluseid
teadmata. Füsioloogia on bioloogias
ja meditsiinis
õpetus organismi
ja selle elundite
talitusest ja funktsioonidest.
Homoöstaas on
bioloogiliste süsteemide
(elusorganismide) võime säilitada neis toimuvate protsesside
tasakaalu, vältida süsteemi põhiomaduste eluohtlikke
kõrvalekaldeid ning kohaneda ümbritsevate tingimustega, et tagada
eluks vajalik sisekeskkonna suhteline püsivus. Suuruste suhtelise
püsivuse hoidmine toimub organismis tänu nende ja paljude teiste
füsioloogiliste näitajate täpsele regulatsioonile, hoitakse
püsival tasemel rakkude ja kudede struktuur ja füüsikalis-keemilised
omadused. Homoöstaas
saavutatakse regulatsiooni kaudu:
närvisüsteemi-,
humoraalne - ja
autoregulatsioon .
Konstantsena hoitakse:
- toitainete ja jääkainete kontsentratsioon
- erinevate soolade/ioonide kontsentratsioon (nt. naatrium, kaalium, kaltsium)
- süsihappegaasi ja hapniku kontsentratsioon
- vee- ja osmoregulatsioon (vee ja lahustunud aine vahekord), maht, temp, rõhk
- temperatuur
- pH (happe ja leelise vahekord)
2. Organismi talitluste regulatsiooni üldised põhimõtted.
Rakkudevaheline kommunikatsioon füsioloogia kontekstis. –
Refleks on organismi sihipärane kohatumisreaktsioon, mis
toimub refleksikaare kaudu,
vastuseks sise- või väliskeskkonnast
pärinevatele stiimulitele (ärritajatele). Refleks avaldub mingi
elusdi, elundsüsteemi või kogu organismi talitluse muutuses,
refleksi anatoomiliseks substraadiks on
refleksikaar . Refleksikaare
moodustab
sensor e
retseptor ,
aferentne juhtetee
(
sensoorne neuron )
refleksikeskus (KNS)
eferetsne juhtetee
ja
efektorelund. Nii aferentses kui eferentses refleksikaare
osas võivad olla vahele lülitatud ka sisesekretoorsed näärmed,
sellisel juhul jõuavad esmase ärritaja mõjul tekkinud
impulsid refleksikeskusesse ja refleksikeskusest välja
saadetud impulsid
efektorelundini ühe või mitme hormooni vahendusel.
Reguleerimiskontuuri põhiplokkideks on
reguleeritav
süsteem ja
regulaator .
Andur mõõdab
reguleeritava suuruse
tegelikku väärtust antud hetkel ja
edastab selle regulaatorile. Reguraatorile on ette antud
reguleeritava suuruse
nõutav väärtus, kui reguleeritava
suutuse tegelik ja nõutav väärtus üksteisest erinevad, on
tegemist
reguleerimishälbega. Selle põhjal töötab
regulaator välja
reguleertoime, mis mõjub
reguleeritavat
süsteemi. Nii reguleerimiskontuuri kui refleksikaare kaudu
toimuva regulatsiooni juurde kuulub tagasiside, mille vahendusel
antakse regulaatorile informatsiooni saavutanud tulemuste kohta, et
eesmärgini jõudmiseks vajadusel tegevust
korrata .
Faktorit või
faktoreid, mis põhjustavad reguleeritava suuruse kõrvalekaldumise
selle nõutavast väärtusest, nim
häiringuks.
Reguleertoimega saavutatud reguleeritava suuruse muutust kontrollib
andur, saadud informatsiooni põhjal käivitub tagasiside, mis
võib olla kas positiivne või negatiivne. Kogu organismi käitumist
kontrollivad samuti meie meeled,
andes teada tegevuse efektiivsusest.
Negatiivne tagasiside avaldub selles, et reguleeriva suuruse
tõus või langus kutsub esile reguleeritava süsteemi vastuse, mis
muudab või püüab muuta
reguleeritava suuruse tegelikku
väärtust esialgsele nihkele vastupidises suunas, nii et see
võimalikult vähe erineks reguleeritava suuruse etteantud
väärtusest. Selline
reaktsioon tagab süsteemi stabiilsuse –
stabiliseerib süsteemi.
Positiivne tagasiside avaldub selles, et reguleeritava suuruse
tõus või langus kutsub esile reguleeritava süsteemi vastuse, mis
muudab või püüab muuta
reguleeritavat suurust esialgse nihkega samas suunas. Selline
reaktsioon võib viia süsteemi
tasakaalust välja, võib süsteemi destabiliseerida. Sageli on aga
positiivne tagasiside kiire vastuse saamiseks, eriti reguleerimise
alguses, vajalik. Reguleeritava suuruse ülemäärase muutuse
vältimiseks tuleb positiivse tagasiside ahel reguleerimise teatud
etapil kas reguleerimiskontuurist endast või väljastpoolt tuleva
mõjutustega katkestada.
Ennetavside põhjustab reguleeritavas süsteemis muutused, mis
püüavad ära hoida reguleeritava suuruse nihet enne, kui häiring
on mõju avaldanud. Niiviisi valmistatakse
organimsi eelseisvaks
tegevuseks ja ümbritsevate tingimuste muutuseks ette ning hoitakse
ära reguleeritava suuruse suuremad
nihked või viiakse need
kiiremini vastavusse organismi
vajadusega .
- Teatud muutujate muutmine soovitud eesmärgile orienteeritud viisil
- Teatud parameetreid hoitakse teatud kitsaas vahemikus
- Kohastumine ja komplekse termodünaamiliselt avatud struktuuri hoidmine
- Parameetri tasakaalus hoidmine toimib vaid siis kui parameetri suurenemisest ja vähenemisest tingitud mõjud on tasakaalus
- Homeöstaas
- Parameetrid võivad olla ruumiliselt eraldatud sellepärast regulatsioon üle terve organismi
Rakkudevaheline kommunikatsioon füsioloogia kontekstis.3. Autonoomse närvisüsteemi (ANS) määratlus ja
üldiseloomustus. ANS-i sümpaatiline ja parasümpaatiline osa: anatoomiline struktuur, neuromediaatorid ja retseptorid ,toimesihtorganitele. Soole närvisüsteem.Autonoomne ehk
vegetatiivne närvisüsteem reguleerib ja koordineerib
siseelundite talitlust. ANS kaudu juhitavad funktsioonid ei allu
tahtele. ANS effektoriteks on südamelihas,
silelihased ja näärmed.
Sisekeskkonna stabiilsus sõltub suuresti ANS-st. ANS-l on 2
neuroniline ühendus kesknärvisüsteemi ja sihtorgani vahel.
Sünapsid 1. ja 2. neuroni vahel paiknevad närvisõlmedes ehk
ganglionides.
Vegetatiivse närvisüsteemi keskuste närvirakkudest
lähtuv preganglionaarne närvikiud lülitatakse vegetatiivses
ganglionis ümber teisele närvirakule, mille
akson postganglionaarne
närvikiud jõuab innerveeritava elundini. Preganglionaarne neuron on
kergelt müoliniseerunud.
ANS põhiline inegratsiooni
tsenter on hüpotaalamus.
ANS-i sümpaatiline ja parasümpaatiline osa: anatoomiline
struktuur, neuromediaatorid ja retseptorid, toime sihtorganitele.
Vegetatiivne närvisüsteem jaguneb: sümpaatiliseks ja
parasümpaatiliseks osaks. Üldiselt teenivad nad sama siseorganit,
kuid põhjustavad vastupidist effekti.
- Sümpaatikus funktsioneerib intensiivselt äkilistes kriisiolukordades ( fight or flight ). Vereringe aktiveerub , südame löögisagedus kiireneb ja suureneb löögimaht, naha ja siseelundite veresooned ahenevad ning vererõhk tõuseb. Südame ja töötavate luustikulihaste veresooned laienevad . Peente bronhiarude silelihaskiud lõtvuvad ja hingamisteed avarduvad.
- Sümpaatikus aeglustab seedekanali motoorikat ja eritamist. Sümpaatikuse ärritus laiendab pupille, suurendab higi eritumist. Sümpaatikus kontrolllib termoregulatsiooni vastusena kuumale, renniini vabanemist neerust ja mtaboolset effekti.
- Parasümpaatikus domineerib kui kogutakse jõudu: magades ja toitu seedides. Parasümpaatikus aeglustab südame löögisagedust, kiirendab seedetegevust ja eritamist ning mõjutab osaliselt ka kusepõie tühjenemist.
Soole närvisüsteem. - Mao- ja sooletrakt koosneb mitmesugustest efektorsüsteemidest, nagu silelihased, sekretoorne epiteel , resorbeeriv epiteel, vaskulaarne ja endokriinne süsteem.
- Nende efektorsüsteemide talitluse regulatsioon on soole närvisüsteemi, soolevälise sümpaatilise ja parasümpaatilise närvisüsteemi ning vistseraalsete spinaalide ja vagaalsete aferentide funktsiooniks.
- Mao- ja sooletrakti soolevälise innervatsiooni väljalülitamine ei mõjuta enamikku mao ja soolestiku elementaarsetest funktsioonidest
- Neuronid jaotuvad aferentseteks neuroniteks, interneuroniteks ja motoorseteks neuroniteks.
- Soole närvisüsteem sisaldab sensomotoorseid programme soole efektorsüsteemide talitluse regulatsiooniks ja koordinatsiooniks.
- KNS avaldab sellele lokaalsele süsteemile ainult moduleerivat mõju.
- Efektorelundi läheduses paiknevad refleksiahelad, mis kohandavad jooksvalt efektorsüsteemi käitumise tingimustele soole valendikus.
- KNS saab infot vistseraalsete aferentide kaudu ja kohandab seedetrakti funktsionaalset seisundit vastavalt organismi seisundile.
KNS otsene neuraalne kontroll on rohkem välja kujunenud seedetrakti
algus- ja lõpposas (toidu vastuvõtt, tühjendusfunktsioon)
4. Lihasraku membraani bioelektrilised omadused. Müoneuraalne
sünaps. Lihasraku ehituslikud iseärasused. Lihaskoe põhitüübid.Membraani laeng on positiivne väljaspool (rohkem Na+ ) ja negatiivne
lihasraku sees. Sellega on tagatud potentsiaalide vahe ehk membraani
polariseeritus, mis on vajalik aktsioonipotentsiaali tekkeks.
Müoneuraalne sünaps on koht kus
motoneuron kohtub lihaskiuga
(eraldatud mulguga, mida nim neuromuskulaarseks piluks).Motoorset
lõpp-plaati ümbritseb sarkolemmist
tasku , mis on moodustunud
motoneuroni ümber. Motoneuronist vabaneb atsetüülkolliini, mis
põhjustab lõpp-plaadi potentsiaali (EPP) e lihasraku
depolarisatsiooni.
Motoneuronit koos lihaskiuga nim moroorseks ühikuks. Ühe
motoorse ühiku stimuleerimine põhjustab nõrga kontraktsiooni terves
lihases .
Kokkutõmme (tõmblus) koosneb kolmest faasist: latents- paar ms
pärast stimulatsiooni kuni erutuse/kokkutõmbe ilmumiseni.
Kontraktsioon - ristsillad on aktiivsed, lihas on lühenenud, kui
pinge on piisavalt suur ületamaks laengut. Lõdvestus
(puhkeperiood)- Ca2+ pumbatakse tagasi sarkoplasmaatilisse
retiikulumi ja lihas pinge
alaneb basaalsele
tasemele .
Lihasraku ehituslikud iseärasusedLihasrakk
koosneb - Lihasfiiber ehk
rakk , on sisse pakitud endomüüsiumi
poolt. Lihaskimpe ümbriteb perimüüsium. Epimüüsium katab kogu
lihast. Lihaskiu membraani nim sarkolemmiks , tsütoplasmat
sarkoplasmaks ja ER=SR (müofiiber). Kontraktiilseks üksuseks
müofiibris on sarkomeerid, Need koosnevad
aktiinist (
peened -
troponiin , tropomüosiin) ja müosiinist (paksud filamendid).
Sarkomeeris on M,H,Z- jooned ja A,I vöödid.
Aktiin libiseb müosiini
suhtes ja lihas lüheneb
Lihaskoe põhitüübid:•
Skeletilihased - Kinnituvad luudele (kõõluste abil)
- Palju perifeerselt paiknevaid tuumi
- Ristivöödilisus, tahtlikud ja mittetahtlikud (refleksid)
- Skeletilihaste stuktuur: * lihaskiud ehk rakud (arenevad müoblastidest, nende arv on konstantne ) *sidekude *närvid ja veresooned
- Inimkehas on rohkem kui 400 skeletilihast (40-50% kehakaalust)
Skeletilihase funktsioonid
- Lihasjõu produktsioon liikumiseks ja hingamiseks
- Lihasjõu produktsioon asendi säilitamiseks
- Soojaproduktsioon külma stressi puhul
• Silelihased
- Õõnesorganite seinad, veresoonde, silmas, näärmetes, nahas
- Üksik tsentraalselt paiknev tuum
- Mittevöödilised, tahtele allumatud, mulkühendused vistseraalsetes silelihastes
- Koosnevad käävi-kujulistest kiududest, mille diameeter on 2-10 m ja pikkus mõnisada mikromeetrit.
- Puuduvad skeletilihasele iseloomulikud tugevad sidekoelised ümbrised, kuid omavad umber õhukest endomüüsiumi
- Üldiselt on organiseeritud kahe kihina ( pikki - ja ringi-suunalisena) lähestikku asetsevate kiududena
- Asetsevad õõnesorganite seintes (v.a. süda)
- Omavad skeletilhastele sarnast kontraktiilset mehhanismi
Silelihase funktsionaalsed omadused• Mõnedele vistseraalsetele
lihastele on
omased autorütmilised
kontraktsioonid • Omavad võimed reageerida kontraktsiooniga vastuseks järsule
venitusele, aga aeglasel venitusel suurendavad nad oma pikkust
• Iseloomulik on suhteliselt konstantne pinge: silelihase
toonus • Kontraktsiooni
amplituud ei muutu
ehkki lihaspikkus võib
varieeruda
Silelihaste regulatsioon• Innerveeritakse
autonoomse närvisüsteemi poolt
• Virgatsaineteks on siin atsetüülkoliin ja
noradrenaliin • Olulisemateks hormoonideks on
adrenaliin ja oksütotsiin, aga ka
mitmed gastrointestinaalsed hormoonid
• Plasmamembraanil
paiknevad retseptorid, millega
virgatsaine või
hormoon seostuvad,
determineerivad, lihasvastuse
• Südamelihased
(vt täpsemalt punkti nr 10)
– Süda
– Üksik
tsentraalselt paiknev tuum
– Ristivöödilisus,
mittetahtlikud,
tihedad ühendused rakkude vahel
5. Lihaskontraktsiooni molekulaarne mehhanism , selle iseärasused
erinevat tüüpi lihasrakkudes. Lihaskontraktsiooni energeetika .Nii sile kui ka vöötlihases on mehhanism sarnane.
Lihase lühenemisel nihkuvad aktiinifilamendid müosiinifilamentide
vahele. Aktiini ja müosiini haakumine toimub müosiini moodustatud
ristsildade kohal.( Kannavad ka adenosiintrifosfaati- kutsub esile
ATP muutumise ADPks).
Skeletilihase kontraktsiooni vallandavaks faktoriks on motoorse närvi
kaudu
leviva aktsioonipotensiaali jõudmine närv-lihas sünapsini.
Presünapsi vesiikulitest vabaneb atsetüülkoliin
seondub postsünapsi membraani N-kolienergiliste retseptoritega, mille
tagajärjel aktiveeruvad Na kanalid.
6. Kehavedelikud: jaotus, keemiline koostis ja ainete tsirkulatsioon . - Päritolult on nad näärmete sekreedid, filtraadid või ka mitme samaaegselt toimuva protsessi resultandid
- Nende koosseisus harilikult mitmed komponendid ehk teisiti öeldes ei leidu organismis kusagil vett ilma lahustunud komponentideta.
- Täidavad mitmekesiseid ülesandeid sõltuvalt kehavedeliku komponentidest.
Vesi organismis - Täiskasvanul 60 % kehamassist, so 70 kg inimesel 42 l
- Sellest 2/3 (~28 l) moodustab intratsellulaarne vedelik ja 1/3 (~14 l ) ekstratsellulaarne vedelik, mis täidavad vastavalt intratsellulaarse ja ekstratsellulaarse vedelikuruumi.
- NB! Kehavedelikud kujutavad endast paljukomponendilisi vesilahuseid.
Intratsellulaarne vedelik - Intratsellulaarne vedelikuruum ei ole kompaktne, vaid moodustub kõikides organismi rakkudes olevate vedelikuruumide summana
- Tsütosooli keemiline koostis on teatud ainete suhtes küllalt stabiilne, mis võimaldab tekkida füsioloogiliselt olulistel gradientidel.
- Rakkude sees on membraanidega ümbritsetud ruumid (kompartmentid), mille keemiline koostis võib tsütosooli omast oluliselt erineda.
- Et bioloogilised membraanid on poolläbilaskvad, siis osmoos on oluline protsess, mis mõjutab vee liikumist intra - ja ekstratsellulaarse vedelikuruumi vahel.
Ekstratsellulaarne vedelik - Ekstratsellulaarsest vedelikust 4/5 (~11 l) on interstitsiaalne ehk koevedelik ja 1/5 (~3 l ) vereplasma
- Ekstratsellulaarse vedeliku hulka loetakse ka transtsellulaarne vedelik: tserebrospinaalvedelik, eksokriinsete näärmete sekreedid, silmakambrite vedelik jt)
Ainete liikumine vedelikuruumides ehk tsirkulatsioon: - Vedelikuruumide sees difusioon
- Vedelikuruumide vahel:
- Ekstratsellulaarne vedelik – rakud: osmoos
- Vereplasma – interstitsiaalne vedelik: difusioon ja filtratsioon
7. Vere üldiseloomustus. Vereplasma iseloomustus.Veri on vedel sidekude. On vahendajaks kõikide kudede vahel.
Vere ülesanded:A)transpordifunktsioon-kannab kopsudest hapnikku ja seedetraktist
imendunud aineid kudedesse. Toitainete oksüdatsioonil vabanenud CO2
viib veri kopsudesse ja teisi ainevahetuse jääke neerudesse. Veri
toimetab hormoone toimekohtadesse. Vere vahendusel jaotatakse
ühtlaselt
soojus .
B) kaitsefunktsioon- veres peituvad
verelibled koos aintikehadega
teevad
kahjutuks haigustekitajad. Vere hüübimine kaitseb
verekaotuse eest.
C) Homoöstaas- veri puhverdab aluselisi ja happelisi aineid, mis
tekivad ainevahetuse käigus. Veri reguleerib oma mahu kaudu
organismi soolade ja vee sisaldust.
Vere hulk, maht, koostis-Inimese kehamassist 6-8%. Keskmiselt 5l verd inimeses.
Koosneb vereplasmast ja verelibledest (hemotsüüdid). Verplasmat
54-59%, liblesid 41-46%
Hematokrit nätab kui suure osa moodustavad
vererakud vere
kogumahust.
Vereplasma, selle koostis, omadused-Vereplasmas on 90-91% vett, 6-8% valke, ja madalmolekulaarseid
aineid.
Värvus
kollakas .
Tihedus
viskoossus 1,025...1,029 ja 1,9...2,6
Osmootne rõhk 768...819 kPa
Leeliselise reaktsiooniga, pH 7,4
Osmomolaarsus 300mosm/l
Vereplasma valgud , fraktsioonid, hulk-Valkusid on 65-80g/l, jaotatakse albumiinideks ja globuliinideks.
Albumiine 35-45 g/l ja globuliine 24-37 (+ fibrinogeeni 1,5-4,5)
Vereplasma valgud kannavad neg.laengut ja käituvad hapetena.
Omavaheline suhe- 1,2-2,0/l
Vereplasma valkude ülesanded-Olulised
vere ja kudede vahelises vee-ja ainevahetuses.
Vereplasma valkude osmootne rõhk (kolloidosmootne/onkootne rõhk)
keskmiselt 3,3 kPa.
Sellest oleneb vere ja kudede vaheline
ainevahetus ja esmasuriini
teke neerudes.
Võtavad osa
ainete transpordist veres-albumiinidega on seotud
kaltsium,
rasvhapped ,mõned
ravimid . Globuliinidega
kortisool ,osa
lipiide ,rauda, vitamiiine.
Vereplasma valkude tähtsus
organismi kaitsereaktsioonis- suur
osa antikehi on globuliinide fraktsiooni kuuluvad
immuunglobuliinid .
Vereplasma valgud moodustavad
osa vere puhversüsteemist.
Valgu molekulid on võimelised
reageerima nii aluste kui hapetega ja
võtavad osa vere happe-leelise tasakaalu säilitamisest.
Vereplasma valkudest oleneb
vereplasma viskoossus, on ka
teatud
valgureserviks.
Vereplasma madalmolekulaarsed ained-Ligikaudu 20g/l , mille konsentratsioon on 290 mmol/l.
Anorgaanilisi (ioonidena) 9g/l orgaanilisi 10g/l.
Anorgaanilised määravad
vere osmootse rõhu, mille kaudu
reguleeritakse vee ja soolade tasakaalu organismis.
(Cl 3,60 g/l ; HSO4 0,02 g/l : valgud 65-80 g/l)
Orgaanilised ained-glükoos,rasvhapped,piimhape. Ainevahetuse
lõpp-produktidest
kusiaine ,kusihape,ammooniumi.
( Na 3,22 g/l ; Ca 0,10 g/l ; Mg 0,02 g/l)
8. Ülevaade vererakkude (erütrotsüüdid, leukotsüüdid ja
trombotsüüdid) talitlusest.
Verelibled/vererakud/hemotsüüdid-Jaotatakse punalibledeks e erütrotsüütideks ja valgelibledeks e
leukotsüütideks ja vereliistakuteks e trombotsüütideks.
Hulk veres suur ja konstantne. Vanad rakud asendatakse uutega.
Erütrotsüüdid-Hulk
4,5-5,5 *10ˇ12 liitri vere kohta.
Kaksiknõgusa ketta
kujulised , tuumata. Keskmine eluiga 120 päeva.
Punaliblede püsimine suspensioonina oleneb
vereplasma
viskoossusest ja erütrotsüütide massi ja pindala
suhtest ,
elektrostaatilistest
tõmbe-ja tõukejõududest.
Hüübimatuks muudetud vere punalibled settivad, hinnatakse
erütrotsüütide
settimise kiirust. Põletikulise haiguse korral
settimine kiireneb. Normaalselt oleks see 2..15mm/h.
Massist
30% hemoglobiini , mis transpordib hapnikku. Keskmine
hulk
meestel 140-170g/l,
naistel 120-160g/l.
Punaliblede purunemisel vabaneb
hemoglobiin , nim.
hemolüüsiks.
Hemolüüsi põhjustavad temperatuuri kõikumised, hüptoonilised
lahused , mehhaanilised
faktorid . Keemilised-lipoide
lahustavad ained
ja bioloogilised-toksiinid.
Leukotsüüdid-Liitris veres
6-10 * 10ˇ9Tuumaga rakud, suuruselt ja funktsioonilt erinevad.
Jagatakse
granulotüütideks(65%) ja
agranulotsüütideks(25-35%). Olenevalt graanulite
sisaldamisest.
Graanulite värvumise järgi eristatakse basofiilseid,eosinofiilseid
ja neutrofiilseid granulotsüüte.
Agranulotsüüte jaotatakse lümfotsüütideks ja monotsüütideks.
Üks osa leukotsüüte
fagotsüteerivad baktereid ja võõrkehi.
Peamiselt neutrofiilsed granulotsüüdid.
Eosinofiilsete gr.hulk
suureneb allergiliste reaktsioonide
korral.
Basofiilsete terakesed sisaldavad hepariini ja
histamiini , mis
laiendavad veresooni ja
parandavad verevoolu põletikukolletes.
T ja B Lümfotsüüdid-T lümfotsüüte u 70-80% lümfotsüütide koguarvust. Vastavalt
funktsioonile eristatakse:
T-hävitajarakud- vabastaavad lümfokiine ja hävitavad
patogeense faktori.
T-abistajarakud-vallandavad B rakkude võime muutuda
plasmarakkudeks ja produtseerida antikehasid.
T-pärssijarakud-muudavad B ja T lümfotsüütide aktiivsust
ja reguleerivad selle kaudu immuunreaktsioonie
T-mälurakud-jäävad lümfisõlmedesse aastateks pärast
infektsiooni ja produtseerivad väikesel hulgal antikehasid. Sama
patogeense faktori
sattumisel organismi valmib suurel hulgal
T.rakkusid,mis tagavad kiire immunoloogilise vastuse.
B-lümfotsüüte umbes 15% lümfotsüütide üldarvust. Jaotatakse
plasmarakkudeks ja
B-mälurakkudeks (antigeenide
retseptorid)
B-lümfotsüüdid
sekreteerivad immuunglobuliine. Enamasti
vajavad T-abistajarakkude abi.
Trombotsüüdid-Vereliistakuid liitris
2,5-3*10ˇ11Tuumata.
Sisaldavad vere hüübimiseks olulist trombotüütide faktorit
(fosfolipiide) ja veresooni ahendavat ainet serotoniini, samuti
adrenaliini, noradrenaliini, histamiini jne.
9. Vere hüübimise füsioloogia.Vigastuse tagajärjel vabanevad trombotsüütidest
vasokonstriktoorsed ained, mis ahendavad vigastuse kohal
veresooni. Trombotsüüdid kleepuvad kokku ja liibuvad
vigastatud kohale. Tekib
valge tromb, mis vähendab
verejooksu haavast.
Samal ajal käivitub verehüübe-
punase trombi teke, mille
põhireaktsiooniks on plasmavalgust fibrinogeenist trombiini toimel
lahustumatu fibriini tekkimine.
Vere hüübimisel on eristatavad
aktivatsiooni-,koagulatsiooni-,fibrinolüüsifaas.
Aktivatsioonifaasis-mitmete faktorite järkjärgulise
aktiviseerimise tulemusel muudetakse toimivaks X e
Stuarti-Proweri
faktor. Seda saab algatada seesmise ja välimise tee kaudu.
Seesmise tee korral: kõrgmolekulaarne
kininogeen ja
prekallikreiinist tekkinud
kallikreen aktiveerivad
XII
faktori, mis kõrgmolekulaarse kininogeeni osalusel aktiveerib
XI
faktori ja see koos
VII faktoriga põhjustab
IX faktori
aktiveerumise. Aktiivne
IX faktor koos Ca,fosfolipiidide ning
aktiivse
VIII faktoriga aktiveerivad
X faktori. Kõik
faktorid esinevad veres.
Välimise tee:vigastatud koest pärit fofolipiidid ja
Ca-
ioonid aktiveerivad VII faktori. VII faktorkoos koe
tromboplastiini (III faktori) Ca ja fosfolipiididega muudab
aktiivseks X faktori.
Seesmise kui välimise tee kaudu aktiivseks muudetud X faktor koos
fosfolipiidide,Ca, ja aktiveeritud V faktoriga
(aktseleraatorglobuliiniga) põhjustavad protrombiini muutumise
trombiiniks.
Koagulatsioonifaasis tekib fibrinogeenist trombiini toimel
fibriin , mis moodustab
võrgustiku, kuhu takerduvad
verekehakesed.
Tekib punane tromb, mis
suleb verevoolu.
Trombiin aktiveerib fibriini stabiliseeriva
XIII faktori, mis
muudab lahustuva fibriini monomeeri lahustumatuks fibriini
polümeeriks.
Fibrinofüüsifaasis-
plasmiin lõhustab veresoonde
jääva ja verevoolu takistava fibriini.
Plasmiin on
proteaas ,mis tekib vereplasma inaktiivsest
plasminogeenist.Plasminogeeni aktiveerib
plasma kallikreiin ning
aktiveeritud XII faktor.
Plasminogeeni võib aktiivseks muuta ka neerudest uriini üleminev
urokinaas, väljaspoolt organismi Beeta-hemolüütiline stretokokk.
Mõne hüübimisfaktori puudumisel tekib hemofiilia.
Vere hüübimist takistavad ained on
antikoagulandid .
Soolad (
naatriumtsistraat), hepariin,
hirudiin , dikumariin.
10. Südame ja vereringe füsioloogia. Südame ehitus,
südamelihase omadused. Südameerutustekke ja erutusjuhtesüsteem. Elektrokardiograafia.Vere liikumine südames.Südametsükkel-
Südame parem ha vasak pool on vereringes järjestikku ühendatud
pumbad . Vere ühesuunalise liikumise tagavad kodade ja vatsakeste
vahel asuvad kodedevatsakeste e atrioventikulaar e.hõlmased
klapid ,
ning poolkuuklapid, e.semilunaarklapid.
Vasaku koja ja vatsakese vahel on
mitraalklapp ning parema koja ja
vatsakese vahel kolmehõlmane
klapp .
Südamelihase süstolid ja lõõgastumised e.diastolid vahelduvad
korrapäraselt, moodutades ühe südametsükli. Selles võimalik
eristada kodade ja vatsakeste tsüklit.
Südametsükkel arvutatakse südamelöögisagedus / sekunditega
minutis .
Erinevad rõhud parema ja vasaku vatsakese vahel- maksimaalne
süstoolne rõhk paremas vatsakeses 25mmHg ja vasakus 120mmHg.
Kodade tsükkel-
Kodade süstol, mille kestus umbes 0,1seki lisab
diastoli ajal
täitunud vatsakesse täiendava koguse verd, mis moodustab 8-10%
vatsakeses
olevast verest.
Kodade süstol lõpetab vatsakeste täitumisfaasi, mille jooksul on
kummassegi vatsakesse voolanud 70-80 ml verd. Nim.täitumismahuks.
Algab kodade
diastol , mis kestab kuni 0.9sek.
Kumbki
vatsake sisaldab 150ml verd, so.vatsakeste lõppdiastoolne
maht.
Vatsakeste tsükkel-
Kodade süstolile järgneb vatsakeste süstol.
Algatajaks vatsakeste
lihaseni jõudnud erutusimpulss, Q-
saki algus. Vatsakeste süstol
algab asünkroonse kontraktsioonifaasiga 0,05seki,
atrioventikulaarklapid on veel avatud. Vatsakestesisene rõhu tõus
põhjustab vere liikumise kodade suunas,atrioventikulaarklapid
sulguvad takistades vere tagasivoolu kodadesse. Sulgumisega kaasneb
esimese südametooni teke.
Pärast atrioventikulaarklappide sulgumist on vatsakeste õõs
suletud.
Südamelihaste jätkuva kokkutõmbe tõttu tõuseb rõhk
vatsakestesiseselt järsult, maht ei muutu. Nim.isomeetrilise
kontraktsiooni faasiks. 0,05seki.
Kui vasaku vatsakese siserõhk ületab rõhu
aordis , 80mmHg ja
paremas vatsakeses on rõhk suurem rõhust kopsuarteris 8mmHg,
avanevad poolkuuklapid ja veri surutakse aorti ja kopsuarterisse.Vere
väljutusfaas 0,25seki.
Süstol kokku kestab 0,35 seki.
Puhkeolekus paisatakse ühe süstoliga kuni 80ml verd,mida nim.südame
löögimahuks. Pärast väljutusfaasi lõppu
vatsakestesse jäävat
mahtu nim lõppsüstoolseks mahuks.e.jääkmahuks.
Arvutatakse kuni löögisagedus 70x/min ja maht 70ml= 70*70=4900ml,
e. südame
minutimaht Q.
Väljutusfaasis rõhk aordis 120-130mmHg,kopsuarteris 20-25mmHg.
Süstoolse rõhu väärtused.
Väljutusfaasi lõppedes vatsakeste lihas lõõgastub.
Vatsakeste siserõhk langeb.
Järgneva prodiastoli 0,04sek sulguvad poolkuuklapid, mis takistavad
vere tagasivoolamist vatsakesse.Sellega kaasub teise südametooni
teke. Algab diastol. Järgneb
isomeetriline lõõgastumise faas.0,05
seki. Rõhk vatsakeses langeb peaaegu nulli. Kui rõhk osutub
madalamaks rõhust kodadest avanevad antrioventikulaarklapid ja algab
vatsakeste täitumisfaas.
Südame löögisageduse tõus viia diastoli lühenemisele.Kui südame
löögisagedus ületab 180 lööki minutis, kogu tsükkel kestab
.0,3seki, siis viib see minutimahu vähenemiseni, sest süda ei jõua
verega täituda.
Südamelihas-müokard- ühendab sile-kui ka vöötlihase omadusi.
Ehituselt on südamelihas sarnane vöötlihasele.
Müosiini ja aktiinifilamente sisaldavad lihaskiud on skeleti
omadest lühemad ja kõrvalharude kaudu ühendatuna moodustavad süntsüütiumi.
Sarnasus silelihasega-müokardi kokkutõmbeid algatavad südames
endas
tekkivad impulsid. Koerakkude sellist omadust
nim.automaatsuseks, seetõttu süda võib töötada ka väljaspool
organismi.
Südame erutustekke ja erutusjuhtesüsteem-,mis koosneb atüüpilisest lihaskoest,kuuluvad sinuatriaal-ja
atrioventikulaarsõlm,
Hisi -kimp, Hisi
kimbu sääred ning
erutusjuhtesüsteemi lõppharudene Purkyne kiud.
Sinuatriaalsõlm paikneb parema koja seinas õõnesveenide
suubumiskohal. Seal tekib südame kokkutõmbeid käivitav
erutus , mis
levib mööda kodade muskulatuuri kodade ja vatsakeste
piiril asuva
atriventikulaarsõlmeni. Sealt edasi Hisi kimpu, selle sääri ja
Purkyne kiudusid pidi vatsakese
lihasele .
Erutuse tekke ja
levikuga kaasneb südamelihase kokkutõmme e.süstol.
Kokkutõmbe ajal ei vasta südamelihas teatud aja jooksul
ärritajatele uue erutuse tekkega. Seda ajavahemikku nim. Absoluutse
refraktaarsuse
perioodiks .
Puhkeolekus tekib 60-70
impulssi sinuatriaalsõlmes minutis,
südametsükli kestus 0,85-1 sek, kestab absoluutne refraktaarsuse
periood 0,2 seki.
Sellele järgneb suhtelise refraktaarsuse periood e.erutuvuse
järk-järguline
taastumine . Kestab 0,02-0,05 seki. Selle jooksul
võib uue erutuse esile kutsuda
normaalsest tugevama ärritajaga.
Südamelihas uutele ärritajatele ei reageeri refraktaarsuse ajal,
see tagab südame rütmilisuse.
Erutuse levimiskiirus- südame eri osades erinev. Kodades 1m/s,
antrioventikulaarsõlme ülaosas 0,02-0,05 m/s, ülejäänud
erutusjuhtesüsteemis 2-4m/s, vatsakeste lihases u 1m/s.
Erutuse juhtimise aeglustumine atrioventikulaarsõlme ülaosas hoiab
ära kodade ja vatsakeste samaaegse kokkutõmbe. Vasakesed
kontraheeruvad pärast kodasid.
Südamelihase kokkutõmbeid algatanud erutus avaldub spontaanselt
tekkiva elektrilise potensiaali muutusena sinuatriaalsõlme rakkude
pinnamembraanil, mille väärtus -50..-60mV.
Suhtelist puhkeolekut iseloomustab aeglane diastoolne
depolarisatsioon , mis vallandab kiire depolarisatsiooni- ja
repolarisatsioonifaasi.
Antrioventikulaarsõlmel on iseloomulik potentsiaalimuutus samaugune
kui sinuatriaalsõlmel. Vatsakeste müokardi pontentsiaalil on püsiv
polarisatsioonifaas, kiire depolarisatsioon ja platookujuline
repolarisatsioonifaas, mille jooksul on vatsakeste lihas
refraktaarperioodis ega võta vastu uusi ärritajaid.
Elektrokardiogramm-
Kui südametegevusega kaasnevaid elektrinähtusi registreeritakse
keha pinnalt. EKG.
Tavaliselt kätele ja
jalgadele kinnitatud eletroodide abil.
Standardsete jäsemelülituste abil.
I lülitus-parem ja vasak käsi
II lülitus parem käsi, vasak jalg
III lülitus vasak käsi ja jalg.
Erinevaid väljalööke tähistatakse P,Q; R, S, T sakkidega.
P-
sakk -erutuse levikul kodades, Q;R;S sakk (QRS
kompleks ) vastab
vatsakeste depolarisatsioonile. T-sakk näitab vatsakeste
repolarisatsiooni.
P-saki algusest kuni Q-saki
alguseni mõõdetud aeg vastab erutuse
levikule kodadelt vatsakestele.
Q-saki algusest T-saki lõpuni (Q-T
intervall ) mõõdetud aeg on
vatsakeste elektriline süstol.
Võimalik iseloomustada müokardi seisundit,erutusjuhtesüsteemi.
Esimesena Einthoven.
Südamelihase klokkutõmbejõud sültub kontraktsioonieelsest
pikkusest, südamelihase energia-ning hapnikuvarudest. Kui need ei
muutu, süda vastab sama tugeva kokkutõmbega.
11. Veresoonkond. Veresoonte jaotus funktsiooni järgi. Vererõhk
ja selle regulatsioon. Vererõhu mõõtmine.Veresoonte jaotus funktsiooni järgi-Oleneb asukohast,läbimõõdust ja seina ehitusest.
Eristatakse jaotus-,kogumis-,takistus-,vahetus-,
mahtuvus -,
elastsus -,
šuntfunktsioniga veresooni.
Jaotus ja kogumisfunktsioon- vere kiire transport. Aort,
suured ja väiksesed
arterid , väikesed ja suured veenid.
Takistusfunktsioon-avaldavad verevoolule teatud
takistust.Arteriool,prekapillaarid,kapillaarid.
Arterioolidel ja prekapillaaridel tänu silelihaskihile suurem osa.
Aterioorid peened veresooned, reguleeritava toonusega
silelihaskihiga.
Vahetusfunktsioon-erinevate koealade varustamine toitainete ja
hapnikuga, ning CO2 ja teiste ainevahetuse jääkide äraandmine
kudedelt verele.Aeglane vool.
Prekapillaarid,kapillaarid,
veenulid -mikroringe.
Mahtuvusfunktsioon-vereringe madalrõhu süsteemi veresooned.
Madalrõhuga
alas on ringluses olevast veres ligi 60%. Veenide
õhukese seina tõttu muutub nende maht väikeste muutuste korral.
Elastsusfunktsioon-südamest väljuvad suured arterid.
Elastsus tagab vererõhu püsimise ja vere pideva liikumise ka
diastoli ajal.
Šuntfunktsioon-täidavad arterite ja veenide vahelised
ühendused-arteriovennoossed anastomoosid. Kui mõne koepiirkonna
verevajadus väheneb, sulguvad prekapillaarsed sfinkterid ja veri
suunatakse ilma kapillaare läbimata otse veenidesse.
Vererõhu regulatsioon-Oleneb vereringes oleva vere mahust ja vere viskoossusest, südame
minutimahust, veresoonte (eriti arterioolide ja kapillaaride)
takistusest. Kõik mis suurendavad südame minutimahtu, veresoonte
takistust, tõstavad vererõhku.
Piklikajus ,veresoonte toonust reguleerivatel närvikeskustel,
eristatakse veresooni ahendava e vasomotoorseid ja laiendava
e.vasodilatoorse mõjuga osa.
Regulatsioonis osalevad paljud refleksid.
Rõhuretseptorid paiknevad aordikaares, kemoretseptorid karotiid ja
aordigloomustes.
Neilt viiaks
veresoonkonna ja südame talitlus
vastavusse
verevarustuse vajadustega.
Vererõhu ülemäärane tõus vallandab depressoorse
refleksi-
perifeersed veresooned laienevad, takistus verevoolule
väheneb, südame löögisagedus ja maht vähenevad.
Vererõhu liigne langus vallandab pressoorse refleksi-
vastupidine depressoorsele.
Vererõhu mõõtmine-Otseselt või kaduselt.
Otsene vererõhu mõõtmine-nim.invasiivseks. Inimesel viiakse
veresoonde manomeetriga ühendatud kanüül.
Kaudsel mõõtmisel-mitteinvasiivne-avaldatakse veresoonele
vasturõhku
veresoont ümbritsevate kudede kaudu. Kasutatakse
vererõhu vasturõhu põhimõtet.
Korotkovi meetod-mõõdetakse arteriaalset vererõhku õlavarrearteris
sfügmanomeetri abil. Õlavarrearter ümber asetatakse manomeetriga
ühendatud
mansett . Leitakse arteri asukoht, asetatakse stetoskoobi
otsik. Tõstetakse manseti rõhku kuni see ületab arterisese rõhu,
mansetialune arter sulgub,
verevool lakkab. Avatakse
ventiil ,
langetatakse aeglaselt rõhku mansetis. Atreriaalsest rõhust
madalama mansetirõhu juures läbib veri arterit, verevool kireneb,
tekivad keerisvoolud, millega kaasnevad Korotkovi toonid.Toonide
ilmumisel fikseeritakse manomeetri näit, mis vastab süstoolsele
arteriaalsele rõhule.
Rõhu edasisel langusel toonid alguses tugevnevad,siis
kahanevad või
kaovad. Sel hetkel vastab rõhk mansetis diastoolsele ateriaalsele
rõhule.
12. Hingamise füsioloogia. Kopsude ventilatsioon ,
hingamismehaanika, kopsude mahud ja mahtuvused. Surnud ruumid ja
alveolaarventilatsioon. Kopsude verevoolutus.1.
gaasivahetus kopsudes e. väline
hingamine , mille käigus
uuendatakse kopsude ventilatsiooniga osa
alveoolides olevast
gaasisegust. Kopsukapillaaride gaasivahetustsoonis olev veri rikastub
hapnikuga ning annab ära süsinikdioksiidi
2.
gaaside difusioon alveoolide ja vere vahel3.
hapniku ja süsinikdioksiidi transport verega4.
gaaside difusioon kudede ja vere vahelDaltoni seadus ja gaasi osarõhu arvutamine gaasisegus:
P = PN2 + PO2 + PH2O
PX = FX * (PB – PH2O)
Henry seadus:
[
gaas ] = α/760 * Pgaas
Grahami seadus:
DO2 = 1/√32 = 1 : 5,65 = 0, 176
DCO2 = 1/√44 = 1 : 6,63 = 0,150
DO2/DCO2 = 0,176/0,150 = 1,2
Boyle ’i seadus:
P * V =
const (sümbolite tähendused lk 78, 79)
Gaasivahetus kopsudes, kopsude ventilatsioon:Hingamisteid jaotatakse ülemisteks (nina- ja neeluruum) ja
alumisteks (alates hingetorust), nende
piiriks on häälepilu.
Kopsu katva
pleura e. kopsukelme ja
rindkere seina seesmist pinda
vooderdava rinnakelme vahele jääb kapillaarne ruum – kopsukelme-
e. pleuraõõs, mis on täidetus üliõhukese vedelikukihiga.
Atmosfäärirõhust madalama rõhu tõttu pleuraõõnes on
kopsud kogu aeg teatud ulatuses väljavenitatud.
Intrapulmonaalse (kopsusisese) ja intrapleuraalse (pleuraõõnesisese)
rõhu diferentsi nimetatakse transpulmonaalseks rõhuks. (rõhu-mahu
diagramm lk 84)
Sissehingamisel rindkere maht suureneb,
hingamisteedes langeb rõhk
atmosfäärirõhust madalamale ja õhk voolab kopsudesse.
Väljahingamisel rindkere maht väheneb, selle tagajärjel
intrapulmonaalne rõhk tõuseb, ületab atmosfäärirõhu ja üks osa
hingamisteedes olevast gaasisegust surutakse välja. Sissehingamisel
on roided tõstva funktsiooniga välitse roietevahelihaste
kokkutõmme, rinnaõõne maht suureneb eest taha suunas,
diafragma kontraktsioonil tema
kuppel lameneb ja rinnaõõne maht suureneb ka
vertikaalsuunas. Tavalise sügavusega, puhkeolekule iseloomuliku
väljahingamise korral
taastub rindkere maht tema raskuse ja
elastsuse tõttu, sel puhul öeldakse: väljahingamine on passiivne.
Sügavamal väljahingamisel ja hingamisteede takistuse suurenemise
korral võtab roiete langetamisest osa sisemiste roietevahelihaste
kokkutõmme. Sügaval ja raskendatud hingamisel muudavad rinnaõõne
mahtu ka hingamise abilihased:
kaelalihased , saaglihas, rangluualune
lihas, kõhulihased jt.
Kopsude alveolaarventilatsioon. Surnud ruumid:Üks osa hingamismahust täidab hingamisteede selle osa, milles
gaasivahetust ei toimu, sinna kuuluvad nina- ja neeluruum, hingetoru
ja hingamisteed kuni
terminaalbronhioolideni . See on
anatoomiline
surnud ruum, kus sissehingatud õhk puhastub suurematest
tolmuosakestest, soojeneb kehatemperatuurini ja küllastub veeauruga.
Kui alveoole ümbritesevates
kapillaarides puudub verevool, siis
nende alveoolide ventileerimisel gaasivahetus alveolaarõhu ja vere
vahel pole võimalik ning tekib
alvolaarne surnud ruum.
Anatoomiline ja alvolaarne surnud ruum kokku moodustavad
funktsionaalse surnud ruumi. Tervel inimesel on alvolaarne
surnud ruum väga väike ning anatoomiline surnud ruum võrdub
funktsionaalse surund ruumiga.
Seda osa kopsude ventilatsioonist, mis osaleb gaasivahetuses, nim
alveolaarventilatsiooniks (VA). VA saab leida,
kui on teada
hingamismaht ,
anatoomilise surnud ruumi suurus ja
hingamissagedus : VA = (VT –VD) *
fr Alveolaarventilatsiooni saab mõõta ka gaasivahetuse
kaugu nn
Bohri valemi alusel. Määramise põhimõte seisneb selles,
et kopsude üld- ja alveolaarventilatsiooniga äraantud
süsinikdioksiidi hulgad on võrdsed. Surnud ruumi ventilatsiooniga
CO2 organismist välja ei tooda. CO2 hulk
väljahingatud õhus võrdub kopsude minutiventilatsiooni ja
väljahingatud õhu CO2 fraktsiooni korrutisega. See CO2
kogus tuuaksegi alveolaarventilatsiooniga organismist välja. Selle
tõttu on CO2 hulk väljahingatud õhus võrdne ka
aveolaarventilatsiooni ja alveolaargaasis oleva CO2
fraktsiooni korrutisega. Tavaliselt moodustab avleolaarventilatsioon
kopsude üldventilatsioonist > 0,7 ja VD/VT
= naeru ,
nutu, punkpillide mängimise jm tegevuse korral muututda kopsude
ventilatsioon ja väljahingatava õhu hingamisgaaside fraktsioonid,
mitte aga nende hulgad. Juhul, kui ainevahetuse tase ei muutu, on
korrutis VA * FACO2 = const. See tähendab, et
ainevahetusele mittevastava kopsude ventilatsiooni tõusuga kaasub
CO2 fraktsiooni langus ja vastupidi. (vt. veel valemeid lk
85, 86)
Kopsude mahud ja mahtuvused:Hingamismaht (VT) on tavalisel hingamisel ühe korraga sisse-
või väljahingatud õhu hulk. VT suurendamisekso n
reservid nii
välja- kui sissehingamisel. Väljahingamise suunas on reserviks
ekspiratoorne reservmaht (ERV), s.o õhu hulk, mis saadakse
pärast tavalist väljahingamist maksimaalse sügavuseni välja
hingates, ja
sissehingamise suunas on reserviks
inspiratoorne reservmaht (IRV), s.o õhu hulk, mis saadakse pärast tavalise
sügavusega sissehingamist maksimaalse sügavuseni sisse hingates.
Pärast maksimaalse sügavusega sissehingamist maksimaalse sügavuseni
välja hingaryd ühu hulk on
ekspiratoorne vitaalkapatsiteet
(EVC) ja pärast maksimaalse sügavusega väljahingamist maksimmaalse
sügavuseni sisse hingatud õhu hulk on
inspiratoorne
vitaalkapatsiteet (IVC). Pärast maksimaalset väljahingmaist
jääb kopsudesse ruumala, mida nimetatakse kopsude
jääk- e
residuaalmahuks (RV). Maksimaalse sissehingamise järel on
kopsudes ruumala, mida nimetatakse
kopsude kogumahtuvuseks e
totaalkapatsiteediks (TLC). Pärast tavalise sügavusega
väljahingamist kopsudesse jäävat ruumala nimettaakse
funktsionaalseks residuaalkapatsiteediks (FRC). Selles
ruumalas uuendatakse iga gingamisega osa alveolaargaasist, mis
moodustab puhverruumi väljisrõhu ja vere vahel. Ruumalasid, millel
puuduvad tinglikud alajaotused, nimetatakse
mahtudeks, mitmest
mahust
koosnevaid ruumalasid aga
mahtuvuseks e kapatsiteetideks.
Kopsude elulist mahtuvust e
vitaalkapatsiteeti ja selle
alajaotusi – ekspiratoortset reservmahtu, hingamismahtu ja
inspiratoorset reservmahtu saab registreerida spirograafi abil.
Kopsude residuaalmahu määramise tuntumad meetodid põhinevad mingi
testgaasi kopsudesse sisse- või kopsudest väljauhtmisel.
Alveolaarventilatsiooni ja kopsude verevoolutuse suhe:
Hingamisgaasid difundeeruvad läbi alveolaarmembraani ja
venoosne veri arterialiseerub siis, kui ventileeritud alveoole ümbritsevad
verega läbivoolutaud kapillaarid, see sõltub kopsude
alvolaarventilatsiooni (VA) ja kopsude verevoolutuse e
perfusiooni (Q) suhtest. Kui puhkeolekus on kopsude
alveolaarventilatsioon umbes 4...5 l/min ja kopsude verevoolutus
5...6 l/min, siis VA/Q = 0,8...1,0.
Alveolaarventilatsiooni ja kopsude perfusiooni vastavusest oleneb
vere arterialiseerumine. Mitteventileertavaid alveoole (VA
= 0) ümbritsevatest kapillaaridest läbi voolav veri ei
arterialiseeru ning lisandub venoosse nn šuntverena arteriaalsele
verele, nendes alveoolides on VA/Q = 0. Selliste
alveoolide ventilatsioon, mille kapillaarides puudub verevool (Q =
0), moodustab alvolaarse surnud ruumi ja nendes on VA/Q =
∞.
Selgub , et VA/Q
suhtarv võib teoreetiliselt jääda
vahemikku nullist lõpmatuseni. Lokaalsete faktorite mõjul, kus
peamist osa mängivad O2 ja CO2 osarõhud,
reguleeritakse verevoolu ja ventilatsiooni nii, et verega
voolutatakse läbi just neid alveoole, mida ventileeritakse ja
ventileeritakse neid alvoole, mille kapillaarides voolab veri.
Normaalse alveolaarventilatsiooni ja kopsude verevoolutuse korral on
kõige enam alveoole, kus VA/Q = 0,9.
13. Hingamisgaaside difusioon kopsudes ja nende transport verega.
Hingamise regulatsioon.Hingamise üldine iseloomustus. Gaasivahetus
organismi ja teda ümbritseva keskkonna vahel.Hingamise “etapid”.Gaasivahetus välisõhu ja alveolaargaasi vahel hoiab viimasesCO2
osarõhu madalama ja O2 osarõhu kõrgema kui venoosses veres.
Venoosses veres on CO2 osarõhk kõrgem kui alveolaargaasis ja
alveolaargaasi O2 osarõhk kõrgem, CO2 difundeerub verest
alveoolidesse ja O2 alveoolidest verre
veri arterialiseerub. Kui ei esine difusioonihäireid, siis osarõhud
võrdsustavad.
Veri kannab O2 füüsikaliselt lahustununa ja
hemoglobiiniga seotult.
Füüsikaliselt lahustunud hapnikku on vähe, suurem osa kantakse
hemoglobiiniga seotult. Hemoglobiin koosneb neljast
polüpeptiidahelast, igaüks neist sisaldab heemi, igas heemis
kahevalentne rauaaatom., O2 seotakse kergesti ilma rauavalentsi
muutmata ja hemoglobiin muutub oksühemoglobiiniks. Üks
mool hemoglobiini seob täieliku küllastuse korral 4 mooli hapnikku.
CO2 transport veres: lahustunult vereplasmas ja erütrotsüütides(kogu
hulgast u 10%), seotult valkudega nt hemoglobiiniga(11% kogu hulgast)
plasmavalkudega u 1% ja ka vesinikarbonaadina vereplasmas( kõige
rohkemja erütrotsüütides, väga vähe ka dissotseerumata
süsihappena.
Vesinikarbonaadina vereplasmas: kudedes tekkinud CO2 difundeerub
verre ja sealt erütrotsüüti, kus CO2st tekib süsihape.
Reaktsiooni kiirendab karboanhüdraas.
HCO3 ioon difundeerub
erütrotsüüditest välja kontsentratsioonide diferentsi tõttu.
Erütrotsüüti liiguvad negatiivse laengu asendajana kloori ioonid
ja järgneb vesi. HCO3- asendumist klooriga nimetatakse Hamburgeri
nihkeks Kopsude CO2 vahetus on
eelpool kirjeldatud.
Kopsude ventilatsiooni reguleerib piklikajus paikenv
hingamiskeskus ,
kus eristatakse sissehingamislihaste tööd juhtivaid inspiratoorseid
ja välja hing lihaste tööd juhtivaid ekspiratoorseid
neuroneid .
Neile
alluvad motoneuronid
seljaaju IV-VII ja kaela ja I-VII
rinnasegmentide eessarvedes.Neuronite aktiivsuse rütmi-
tsentraalse rõtmogeneesi( selle kujundamine on autonoomne, mõjutab
perifeersetelt retseptoritelt ja KNS osadelt saadav info) abil
viiakse hingamine organismi ainevahetusega vastavusse.
Gaasivahetus kopsudesgaaside
difusioon alveoolide ja vere vahelO2
ja CO2 transport verega gaaside
difusioon kudede ja vere vahel.
Atomosfäärirõhust madalama rõhu ( mäed etc) puhul tekib
hapnikuvaegus õhurõhu languse tõttu ja hemoglobiini küllastatus
O2 väheneb.
Kõrge rõhu puhul hakkab lämmastik kudedes rohkem ning see kutsub
esile muutuseid organismis nt
kohatu rõõmutunne 45 m sügavusel,
sügavamal juba uimasus etc. Kui rõhuvähenemine on liiga kiire
hakkab lämmasti kudedest väljuma mullikestena ja võib ummistada
elutähtsate organismide veresooned
14. Seedimise üldine iseloomustus, olulisemad seedeprotsessid.
Süsivesikute, valkude ja lipiidide seedimise üldine iseloomustus.
Seedimine suus ja maos.Toiduga saadavaid aineid organism enamikul juhtudest väliskeskkonnas
esinebal kujul ei
omasta . Toitu töödeldakse seedekulglas
mehaaniliselt ja segatakse seedenõredega, mille ensüümid
lõhustavaf toitaineid nii, et need muutuvad imenduvaks ja
organismile vastuvõetavaks, valgud ja
lipiidid kaotavad liigispetsiifilisuse.
(Pole kindel olulisemates seedeprotsessides aga ehk on need)
Olulisemad seedeprotsessid: Seedimine suus,
neelamine ,
seedimine maos, seedimine
peensooles , maksa osa seedimises, seedimine
tühi- ja niudesooles, seedimine jämesooles.
Süsivesikute lõhustamine ja imendumine : süsivesikutega
kaetakse 50...60% kogu ööpäevasest energiakulust. Polüsahhariidid
(tärklis, glükogeen,
tselluloos ),
disahhariidid (sahharoos,
laktoos , maltoos),
monosahhariidid ( glükoos,
fruktoos ). Süsivesikud
imenduvad monosahhariididena.
Kiudained lõhustatakse osaliselt alles
jämesoole bakterite poolt.
Kiudained seovad rohkesti vett, stimuleerivad soole motoorikat. Sülje
ja kõhunäärme α-amülaas lõhustab polüsahhariidid kuni
disahhariidi – maltoosini. Amülaas inaktiveeritakse maos soolhappe
toimel, disahhariidid jõuavad lõhustamatult
peensoolde .
Süsivesikute lõhustamise lõpetavad peensoole limaskesta
epiteelirakkudes tekkivad ensüümid – disahharidaasid: maltaas,
sahhraas, laktaas.
Aineid transporditakse läbi raku, s.o transtsellulaarselt.
Enterotsüüdi plasmamembraani apikaalsetes ja basolateraalsetes
piirkondades on erinevad kandurid e transporterid. Monosahhariidide
kergendatud difusioonis osalevad kandurid kuuluvad GLUT perekonda.
Valkude lõhustamine ja imendumine: valku lõhustavaid ensüüme
võib jaotada proteaasideks ja peptidaasideks. Proteaasid lõhustavad
valkusid väiksemateks peptiidideks, mida peptidaasid omakorda
väiksemateks koostisosadeks lõhustavad. Peptidaase jaotatakse
endo -
(lõhustavad peptiidsidemeid aminohapete sees) ja eksopeptidaasideks
(vabastavad üksikuid
aminohappeid ). Valgud imenduvad aminohapetena.
Lipiidide lõhustamine ja imendumine: lipiidide hulka kuuluvad
triglütseriidid,
fosfolipiidid , tsüklilised lipiidid ja
lipiididesarnased
rasvlahustuvad vitamiinid . Lipiide lõhustavad
ensüümid on
lipaasid . Lipiidide lõhustumisele aitab kaasa ensüüme
mittesisaldavad
sapp ja kolipaas. Lipiidide imendumine toimub lihtsa
difusiooni teel läbi enterotsüüdi apikaalse membraani.
Seedimine suus: seedimine algab suus, kus toit peenestatakse
ja segatakse süljega ning muudetakse neelatavaks. Süljenäärmete
sekreeti – sülge – produtseerivad 3 paari suuri
(kõrvasüljenäärmed, keelealused ja lõuaalused näärmed) ning
hulgaliselt suuõõne
limaskestas asuvaid väikesi süljenäärmeid.
Sülg on kergelt
leeliseline vedelik, sisaldab 98...99% vett ja
1...1,5% soolasid ning orgaanilist ainet. Ensüümidest on süljes
süsivesikuid lõhustavat α-amülaasi ning keelepära piirkonnas
asuvatest Ebneri näärmetest pärinevat, lipiide lõhustavat
lingvallipaasi. Süljes lahustunud ained
toimivad maitsmissensoreile.
Sülg vähendab organismi sattuva nakkuse vüimalust. Limaaine
mutsiin muudav toidupala libedamaks ja neelatavaks. Sülg hoiab suu
limaskesta niiskena, mis on vajalik kõnemisel. Süljehulk oleneb
toidu koostisest ja veesisaldusest, seda eraldub ka mittesöödavate
ainete
suhu sattumisel, viimasel juhul on süljel loputusfunktsioon.
Sülje
sekretsioon algab juba toidu nägemisel ja haistmisel ning
ärritajate mõjul. Keskmine sülje hulk ööpäevas 1...1,5 liitrit.
Seedimine maos: Magu on toidu reservuaariks. Maos jätkub
süsivesikute lõhustumine süljeensüümide toimel seni, kuni seda
võialdab
maomahla pH (kui pH45° ja delta kiududega. Mehanosensitiivsed notsitseptorid ühenduvad
A-delta kiududega. Polümodaalsed notsitseptorid on tundlikud
mehaanilistele, termilistele, keemilistele stiimulitele, ühenduvad
C-kiududega. Siseorganites on
vaikivad notsitseptorid, mille lävi
langeb tunduvalt põletiku mõjul.
Temperatuurimeele sensoriteks on nahas paiknevad
külma-, sooja-
ja valusensorid.
Külmasensoriteks on
spetsiifilised Aδ müeliniseeritud,
hargnevad närvilõpmed, mille harud
tungivad epitermise basaalsesse
ossa. Külmasensoreid on 3-10 korda rohkem kui soojasensoreid.
Erinevatel kehapiirkondades on sensorite tihedus erinev.
Soojasensoreid on vähem kui külmaseonsoreid. Sooja vastuvõtuga
seostatakse vabu närvilõpmeid, mis juhivad C tüüpi närvikiudude
kaudu erutust.
Sensorite erutumise mehhaniski seotatakse nende ainevahetuse
intensiivsuse muutusega temp mõjul. Temp muutus 10 kraadi muudab
enam kui kahekordselt intratrellulaasete keemiliste reakstioonide
kiirust. Termosensorite ärritamisel antakse erutus edasi mööda
erinevaid aferentsed närvikiudusid:
Temp alla 15 kraadi – erutuvad külma suhtes tundlikud valusensorid
Külmasensorid alustavad impulsside väljasaatmist 10 kraadist alates, maksimaalne vastus saadakse 25 kraadi juures, sensorid lõpetavad impulsside väljasaatmise 43 kraadi juures
Alates 30 kraadist alustavad impulsside väljasaatmist soojasensorid
45 kraadi juures stimuleeritakse juba kuuma suhtes tundlikke valusensoreid.
ÜLEJÄÄNUD KÜSIMUSTE VASTUSEID EI LEIDNUD! KUI KEEGI NII TARK ON
JA LEIAB/TEAB NEID SIIS ANDKE MULLE KA TEADA! KADRI
Organismis on olemas nn. Antinotsitseptiivsed mehhanismid , nende neurokeemia on tihedasti seotud endogeensete opioidide (endorfiinid)
ja endokannabinoididega (anadamiid) ning alaneva pidurdussüstemiga.
Valude pärssimine valuvaigistitega: mittenarkootilised, nt
atsetüülsalitsüülhape, ja narkootilised, nt morfiin ,
valuvaigistid. On olemas veel lokaalsed ja üldanesteetikumid.
Lokaalsed anesteetikumid toimivad konkreetse piirkonna
valuretseptoritele ja kiududele, üldanesteetikumid kõrvaldavad
teadvuse.
Endogeensed ja eksogeensed opiaadid (nt morfiin, st
oopiumipreparaadid), mõjuvad samadele piirkondadele, millele ka
elektr. stimulatsioon . Organism ise on võimeline samuti tootma endogeenseid opiaate (nt enkefaliin, avast .1975). Hiljem on leitud
ligi 20 proteiini, mis aktiveerivad opiaat-retseptoreid.
Kokkuvõtteks: ajus on nn alanev valu reguleeriv süsteem, mida võib
otseselt mõjutada kas el.stimulatsiooniga v morfiini-taoliste
ainetega. (aju ise aktiveerib seda süsteemi neurotransmitterite
klassiga, mida nim endogeenseteks opiaatideks – platseebo -katsed!).
22. Nägemismeel. Silma ehitus. Nägemisteravus. Silma võrkkest
ja tema retseptorid.
Biokeemilised protsessid kolvikestes ja kepikestes.
Nägemisinformatsiooni vahendavad
juhteteed . Nägemiskeskused ajukoores . – Nägemismeeleelundiks
on silm, mille valgustundlikud sensorid – kepikesed ja kolvikesed – asuvad võrkkestas. Silma optiline süsteem tagab
valguskiirte fokuseerimise võrkkestale, kus tekib vähendatud
ümberpööratud kujutis. Sensorirakkudes valguse toimel tekkinud
sensoripotentsiaalid kutsuvad nägemisnärvis esile
aktsioonipotentsiaalid, mis juhitakse nägemismeele tsentraalseid
teid pidi ajukoore kuklasagarasse, kus teadvuse tasemel tekib
nägemisaisting ja –taju. Inimene tajub valgusena 400-750 nm
pikkusi elektromagnetlaineid.
Silma optilise süsteemi moodustavad: sarvkest , eeskamber,
lääts ja klaaskeha . Lisaks nimetatud valgustmurdvatele
struktuuridele kuulub optilisse süsteemi veel silmaava e pupill ,
mille kaudu reguleeritakse silma langeva valguse hulka. Silma
optilise süsteemi valgustmurdvat võimet mõõdetakse dioptriates.
Silmas on valgustmurdvaid keskkondi mitu.
Silmamuna ehitus. Silmamuna koosneb kestadest ja sisust.
Silmakestad on: fibroos -, soon- ja võrkkest.
Fibrooskesta eesmine 1/6 on sarvkest ja ülejäänud 5/6 kõvakest e skleera.
Soonkestal on 3 osa:
Vikerkest annab silmadele värvi ja võib olenevalt pigmendi hulgast olla tumedam või heledam. Vikerkesta keskkele jääb silmaava, mida ümbritsevad sulgur- ja laiendajalihas
Ripskeha, milles paikneb ripslihas
Pärissoonkest sisaldab veresooni, mille kaudu toidetakse võrkkesta epiteeli
Võrkkestas e reetinas on mitmeid erinevaid rekukihte
Melaniini sisaldav pigmentepiteel, milles valgus neeldub ja seetõttu ei teki silmasisest peegeldust
Sensorirakud: kepikesed ja kolvikesed. Võrkkesta perifeerias piknevad kepikesed, kolvikesed on koondunud kesklohku kollatähni piirkonda
Horisontaalrakud
Bipolaarsed rakud
Ganglionirakud, mille jätked moodustavad nägemisnärvi
Nägemisnärvi reetinast väljumise koht on pimetähn, kuna seal
puuduvad sensorid ja valgustundlikkus.
Nägemismeele iseloomustamiseks mõõdetakse nägemisteravust ning
nägemis- ja vaatevälja suurust. Nägemisteravuse määrab
väikseim kahe punkti vaheline kaugus, mida silm on võimeline eristama . Normaalne silm eristab kaht punkti millelt lähtuvate kiirte nurk on 60 kaarsekundit s.o 1 kaareminut.
Retseptiivseks väljaks on see võrkkesta ala, mille
ärritamine valgusega põhjustab ganglionirakkudelt lähtuva
impulsatsiooni tõusu või languse. Ganglioniraku retseptiivsed
väljad on sõõrjad, nendel on tsentrum ja perifeeria . Valgusele reageerimise järgi on neid kahte tüüpi:
Retseptiivväljad, mille tsentrumi ärritamine valgusega põhjustab sealt lähtuvate impulsside sageduse tõusu, samal ajal perifeeriast välja saadetud impulsside arv väheneb. Retseptiivvälja perifeeria ärritamisel on olukord vastupidine. Need retseptiivväljad reageerivad valguse sisselülitamisele.
Retseptiivväljad, mille tsentrumi ärritamisel valgusega sealt lähtuvate impulsside sagedus langeb, samaaegselt perifeeriast lähtuvate impulsside sageuds tõuseb. Retseptiivvälja perifeeria ärritamisel tõuseb tsentrumist väljasaadetud impulsside sagedus. Need retseptiivväljad reageerivad valguse väljalülitamisele.
Seega algab erineva tugevusega valgustatud pindade eristamine juba
võrkkestas. Valguse intensiivsuse järsust suurenemisest
signaliseerivad on-tsentrumi gangloinirakud, mis nõrgas valguses
ainult üksikuid impulsse välja saadavad . Valguse intensiivsuse
järsust nõrgenemisest signaliseerivad off-tsentrumi rakud,
millelt lähtuv aktiivsus on madal tugeva valguse juures.
Võrkkestas on valguse suhtes erinevate absoluutlävedega sensorid:
kepikesed ja kolvikesed.
Kepikeste abil nähakse hämaras ja värvusi ei eristata, seda
nim skotoopiliseks nägemiseks. Kepikesed sisaldavad
nägemispurpurit e rodopsiini, mille punane värvus tuleb
sellest, et ta neelab tugevasti rohelist ja sinist valgust.
Kolvikestega nähakse valges ja eritatakse värvusi, see on
fotoopiline nägemine. Värvuste nägemist seletatakse sellega, et kolvikesed sisaldavad kolme erinevat fotopsiini,
mis neelavad sinist, rohelist ja punast valgust.
Pimeduses on kepikeste Na+-kanalid avatud, erinevalt
teistest närvikoemembraanidest, kus Na+-kanalid on
puhkeolekus suletud. Avatuna hoiab neid intratsellulaarne cCMP. Valguskvant aktiveerib rodopsiini, mis koosneb opsiinist ja
11-cis-retinaalist, 11-cis-retinaal muutub
11-trans-retinaaliks. Reaktsiooni võib nim stereoisomerisatsiooniks,
sest muutub aine konfiguratsioon, ilma et aine keemiline koostis
muutuks. 11-trans-retinaal seondub membraani G-valgu transdutsiiniga,
mis aktiveerib omakorda cGMP fosfodiesteraasi ja cGMP muutub
5-GMP-ks. cGMP kontsentratsiooni languse tõttu sulguvad
naatriumkanalid. Na+ sissevool väheneb. Kaaliumkanalid,
millel puuduvad väratid, jäävad avatuks. Tulemuseks on
sensorimembraani hüperpolarisatsioon, mis on seda
ulatuslikum, mida tugevam on valgus. Pimeduses on sensor osalise depolarisatsiooni seisundis ja saadab pidevalt välja pidurdavaid
impulsse võrkkesta bipolaarsetele rakkudele. Kui valgus sensori
hüperpolariserib, siis vähenevad pidurdavad mõjud bipolaarsetele
rakkudele ja need aktiveeruvad.
Valguse „transleerimist“ närvirakkude elektrilisteks
potentsiaalideks nim fototransduktsiooniks.
Bipolaarsed rakud on sünaptilises ühenduses ganglionirakkudega.
Nägemisnärv, mille moodustavad ganglionirakkude jätked,
sisaldab enam kui 106 kiudu . Võrkkestast nina poolt
pärinevad nägemisnärvi kiud ristuvad nägemisristmikus,
oimu poolt tulenevad kiud ei ristu. Nina ja oimu polt tulnud
närvikiud moodustavad ühinemisel nägemistrakti, mis suundub
keskajus asuvasse külgmisse põlvikkehasse, kus toimub
ümberlülitus nägemiskorteksisse viivatele neuronitele. Keskajus
asuvasse närvikeskuste kaudu juhitakse ka silmaliigutusi ja
pupillireaktsioone.
Nägemis- e visuaalkorteks. Primaarne nägemiskorteks asub
kuklasagara kannusvao piirkonnas. 1. Visuaalne ala – V1 saab
informatsiooni kontralateraalse nägemisvälja alalt.
Visuaalkorteksis on 2 mm paksune ja organiseeritud vertikaalsete
sammastena, mida nim orientatsiooni silmadominantseteks
sammasteks.
2. visuaalne ala V2 võtab vastu teatava oreintatsiooniga
kontuure ja joonekatkestusi.
Eristatakse veel ka liikumistundlikkuse ja värvuste nägemisega
seotud nägemiskorteksi alasid.
23. Kuulmismeel. Õhuvõnkumisi vahendav aparaat keskkõrvas. Teo
anatoomiline ehitus.Karvarakud teos. Õhuvõngete muundamine kuulmisnärvi elektrilisteks signaalideks.Kuulmisinformatsiooni
vahendavad juhteteed ja selle informatsiooni töötlemisega tegelevad
ajupiirkonnad. – Kuulmismeeleelundiks on kõrv, millel
eristatakse välis-, kesk- ja sisekõrva. Sisekõrvas asuvad
sensorirakud. Nendelt lähtuvad impulsid suunduvad kuulmisnärvilt
kuulmismeele tsentraalseid teid pidi kuulmiskorteksisse ülemises
oimukäärus.
Keskkõrv. Trummikile võnkumised antakse kuulmeluukeste –
vasara, alasi ja jaluse – kaudu edasi ovaalaknale.
Tummikilele kinnitunud vasar liigestub alasiga, mis on omakorda
ühenduses ovaalaknale fikseeritud jalusega. Kuulmeluukeste liikuvust
reguleerivad trummikilepingutaja- ning jaluselihas .
Tummikile pindala on ligikaudu 3,2 mm2. Trummikile ja
ovaalakna pindalade suhe on seega 1:22. Kuulmeluukeste kangide
süsteem ja võnkuvate membraanide suhe võimendab trummikile poolt
vastuvõetud võnkumist umbes 22 korda (võnkeamplituud väheneb,
võnkumiste energia suureneb). Ovaalakna membraani võnkumised
antakse edasi sisekõrva täitvale vedelikule ja seal olevatele
sruktuuridele.
Sisekõrva moodustab luuline kanal , millel on kaks ja pool
keerdu, seda nim teoks. Basilaarmembraan jaotab selle
kanali esikuastrikuks ja trummiastrikuks, mis sisaldavad
perilümfi, vestibulaar e Reissneri membraan eraldab esikuastrikust
teojuha , millesse jääval basilaarmembraani osal asub Corti elund , kus paiknevad sensorirakud.
Ovaalakna membraani võnkumised antakse eskikuastrikku täitva
perlümfi kaudu edasi teojuha endolümfile. Esikuastrik on
teotipu kohal helikotreemi kaudu ühendatud tummiastrikuga,
selle kaudu kanduvad võnkumised sise- ja keskkõrva piiril oleva
ümaraknani. Teojuha on täidetud endolümfiga, mille koostis
on sarnane rakusisese vedelikuga. Teojuha ja tasakaalumeele juurde
kuuluva ümarkotikese vahel on ühendus- e reunientjuha.
Corti elund. Trummikile võnkumised antakse kuulmeluukeste,
ovaalakna membraani ja esikuastrikku täitva perilümfi kaudu edasi
endolümfile kogu teokanali ulatuses. Mida madalamad on vastuvõetud
helid, seda pikem peri - ja endolümfi sammas kaasa võngub. Kõrgemate
helide korral võngub kaasa ainult ovaalakna lähedal oleav perilümfi
sammas. Basilaarmembraanil asuvad karvarakud, mille 4-5
mikromeetri pikkused jätked kontakteeruvad võnkumisel
kattemembraaniga. Tekib sensoripotentsiaal, mis kutsub teonärvis
esile erutuse (aktsioonipotentsiaalide jada). Sisekõrva
universaalseks sensoorse raku tüübiks on karvarakk, mis ühendub
sünaptiliselt aferentse närvikiuga. Igal rakul on ~60-80
stereotsiili ja 1 teistest pikem kinotsiil. Karvarakkudel on
märkimisväärne spontaanaktiivsus. Karvakeste liikumine kinotsiili
suunas aktiveerib raku ning liikumine vastupidises suunas pidurdav raku aktiivsust. Teatud kõrgusega helile reageerivad kindlad rakud.
Corti elundis eristatakse ligikaudu 3500 sisemist
karvarakku ja 3-4 reas asuvat ümbes 20000 välimist
karvarakku. Need on ühenduses närvilõpmetega, mis viivad
erutuse spiraalganglionini. Seal asuvate neuronite jätked
moodustavad teonärvi, mis esikuteonärvi koosseisus viib
signaali teonärvi selgmise ja kõhtmise tuumani.
Kuulmismeele tsentraalsed teed. Teonärvituumadest algavad
närvikiud moodustavad trapetskeha, seal lähevad kuulmisteed üle
teise ajupoolde ja ülenevad lateraallinguna. Trapetskeha ja
lateraallingu tuumades toimub ümberlülitus kolmandatele
närvirakkudele, mille jätked lõpevad ülemises oimukäärus asuvas kuulmiskorteksis.
24. Tasakaalumeel . Labürindi ehitus ja füsioloogia.
Vestibulaarsed karvarakud. Vestibulaarsest labürindist lähtuvad
juhteteed ja ajukeskused, mis on seotud vastava informatsiooni
töötlemisega. – Tasakaalumeele- e vestibulaarelundi,
mis paikneb oimuluu püramiidi osas, moodustavad esik ja kolm
poolringkanalit. Esikus asuvad mõik ja ümarkotike. Neid
nim ka tähnielunditeks, sest sensorirakud asuvad mõigu ja
ümarkotikese tähnidel. Luu poolringkanaleid vooderdavad
kilepoolringkanalid, nendevaheline ruum on täidetud perilümfiga.
Kolm poolringkanalit – ülemine, tagumine ja külgmine –
on üksteise suhtes paigutatud perpendikulaarselt. Ülemine,
mille kumerus on suunatud püramiidi pinna poole, asub
frontaaltasapinnas, on selle suhtes aga 45 kraadise nurga all
tahapoole painutatud. Tagumine paikneb sagitaaltasapinnas ja
on selle suhtes 45 kraadi külgsuunas kallutatud. Ülemisel ja
tagumisel poolringmanalil on üks ühine säär. Külgmine
poolringkanal on horisontaaltasapinna suhtes 30 kraadi taha ja
allapoole painutatud. Poolringkanalid algavad seega esikust 3
suudemga, suubuvad sinna ülemise ja tagumise poolringkanali ühe
ühise sääre tõttu 2 suudmega. Poolringkabaleid täidab endolümf.
Kuna kanalid on kolmes eri tasandis , tekib nendes eri suundades
toimuval pöördliikumisel erineva tugevusega endolümfi liikumine,
millele reageerivad sensorirakud.
Poolringkanalitel eristatakse ampullaar- ja lihtsäärt.
Ampullaarsäär on nime saanud oma laienenud osa – ampulli –
järgi, kus asuvad sensorirakkude kogumikud – ampulliharjad.
Sensoriraku pealispinnal paiknevad peened karvakesed , sellepärast
nim neid karvarakkudeks. Karvakeste hulgas eritstatakse umbes
60-80 astmeliselt pikenevat stereotsiili ja üht, teistest
pikemat kinotsiili. Ampullihari ulatub kanali õõnde umbes
kolmandiku ulatuses, tunderakkude karvakesed on pintslitaoliselt sisestatud sültjasse, kristalle mittesisaldavasse massi –
moodustub kuppel, mis täidab kanali peaaegu kogu ulatuses.
Horisontaalsetes poolringkanalites on kõik sensorid orienteeritud selliselt , et kinosiil jääb mõigu poole. Sensoripotentsiaal
tekib ja aktiivsus aferentses närvis suureneb siis, kui kuppel
kooldub mõigu suunas, s.o utrikulopetaalselt. Poolringkanalite
meelerakud reageerivad pöördliikumisel tekkivale endolümfi
liikumisele, kuppel koos sensorirakkude karvakestega paindub ja
põhjustab sensoripotentsiaali tekke. Ärritajaks on endolümfi
liikumine.
Tähnelund. Tasakaaluelundi teise osa sensorirakud paiknevad esiku e vestiibuli mõigus ja ümarkotikeses, selletõttu nim
seda ka vestibulaarelundiks. Mõik paikneb horisontaal-,
ümarkotike vertikaaltasandis. Raskusjõu muutumisel,
sirgjoonelisel kiirendusel paikneb otoliitmembraan sensorirakkude
karvakeste suhtes ümber ning annab andmeid keha- ja peaasendi
muutuse kohta. Seega on tasakaalumeele selle osa adekvaatseks
ärritajaks raskujõud.
Tasakaalumeele tsentraalsed teed. Esimene neuron on
vestibulaarganglionis, sellest suundub neuroni perifeerne jätke tähnielundi või poolringkanali ampulli sensorirakku. Tsentraalne jätke moodustab VIII peaajunärvi teise osa –
esikunärvi-, mis suundub piklikajus asuvatesse tuumadesse.
Osa esikunärvi kiude läheb otse väikeajju. Esikunärvi tüvi
jaguneb esikünärvi lateraalses tuumas lõppevaks alanevaks ning
esikunärvi ülenevaks osaks. Tähnileundist pärinevad kiud jõuavad
alumisse, osalt ka mediaalsesse tuuma, poolringkanalitsest pärit
kiud suunduvad ülemisse ja osalt mediaalsesse tuuma. Esikunävi
tuumadest kulgevad närvikiud edasi resikulaarformatsiooni,
väikeajju, silmalihaseid innerveerivate närvituumade juurde ja
seljaajju. Retikulaarformatsiooni suunduvad teed mõjutavad
retikulaarformatsiooni-seljaaju kulgla kaudu seljaaju motoneuroneid.
Väikeajju tuleb osa närvikiudusid otse vestibulaarganglinost, osa
tuleb sinna ka esikunärvi mediaalsest ja alumisest tuumast. Need
lõpevad väikaju tuumades, mida nim sõlmekeseks ja
tätrakeseks. Üks osa ühendusteid läbib talamuse ja jõuab
ajukoorde. Ülalnimetatud teede ning esikunärvituumade ja
silmalihaseid innerveerivate närvituumade vahelise ühenduste kaudu
kulgevad vestibulaarrefleksid silmamuna välislihastele.
Pöördliikumisel tekivad pea ja silmade nõksuvad liigutused –
nüstagm -, millega tagatakse nägemisorientatsioon ruumis.
25. Haistmismeelega seotud retseptorid. Haistmismeelega seotud
juhteteed ja ajupiirkonnad. – Haistmismeele sensorid –
haistmisrakud – paiknevad ninaõõnes ülemise karbiku serval asuvas haitsmisretseptoris. See piirkond, mida katab 4-5 cm2
suurune haistmisepiteel, on peamistest hingamisteedest kõrval.
Inimesel on kokku ~107 haistmisrakku. Haistmisrakud
uuenevad pidevalt, eluiga on keskmiselt 30-60 päeva. Õhk satub
sinna difusiooni teel või kiirel ja sagedal hingamisel. Inimesel on
erinevatel andmetel 40-100 miljonit haistmisrakku, raku pinnal
paikneb 6-12 aktiivselt liikuvat karvakest e tsiili. See
soodustab nende kontakteerumist lõhnaainetega. Haitsmirakud on
primaarsed bipolaarsed meelerakud, mille tsentraalne osa läheb üle
peeneks jätkeks. See põimub teiste samalaadsetega 15-20
haistmisnärviks. Haistmisnärvid sisenevad sõelluu mulkude
kaudu koljuõõnde ja lõpevad haistmissibulas.
Haistmisrakkude neuronite lõppharud moodustavad haistmissibulas
asuvate mitraalrakkude tugevasti hargnevate dendriitidega
ühinedes haistepäsmakesi.
Mitraalrakud on haistmistee teisteks neuroniteks, mis on dendriit -dentriit sünapsite kaudu ühendatud
periglomerulaarrakkudega. Mitraalrakud on sünaptilises
ühenduses ka sõmerrakkudega. Periglomerulaar- ja sõmerrakud on
pidurdusvaheneuronid, kuna mitmed eferentsed impulsid KNS-st mõjuvad
oste neile. Mitraalrakkude neuriidid moodustavad haistmistee
põhimassi ning jagunevad tagapool mediaalseks, vahelmiseks ja
külgmiseks jutiks, mis üksteisest eemaldudes paiknevad
samanimelistel rudimentaarsetel hasitmiskäärudel. Tekkinud piirkond
kannab nime haistekolmnurk. Haistmistee suundub
frontaalsagaras paiknevasse haistmiskoorde.
Kolmanda neuroni kehad paiknevad haistmissibulast tagapool olevates haistesagarate osades, sealt lähtuvad neuriidid mooddustavad
kortikaalseid ja subkortikaalseid ühendusteid. Haistmisteed ulatuvad
ka limbilisse süsteemi, hüpotalamusse ja suurajukoorde, mille kaudu
tekib hasitmistaju teadvuse tasemel.
Lõhnaainete vastuvõtmine võib olla seotud sensori ehituse
erinevustega. Sellisel juhul seotakse lõhnaaine spetsiifilise kuju
ja suurusega just selle molekuli äratundmiskohta, see põhjustab Na+
- või K+- kanali avanemise ja sensoripotentsiaali tekke.
Signaaliülekanne on seotud G-valkude, cAMP ja viimastest sõltuvate
katioonkanalitega, kuid leitud on ka teiste signaalainete olemasolu
(IP3, cGMP, CO). Retseptori aktivatsiooni tulemuseks on
membraani depolarsiatsiooni ja AP genereerimine aksonil.
Haistmismeele sensorid ehk haistmisrakud paiknevad ninaõõnes
ülemise ninakarbiku serval asuvas haistmisregioonis. Õhk sattub
siia põhiliselt difusiooni teel või kiire ja sageda hingamise abil.
Haistmisrakke on 40-100 miljonit. . Lõhnaainete vastuvõtt on seotud
sensori ehituse erinevustega. Lõhnaaine seotakse spetsiifilise
suuruse ja kujuga vastavasse molekuli äratundmiskohta, kus tekib
sensoripotentsiaal ( Na ja K kanalite teema) Haistmisrakud on
primaarsed bipolaarsed meelerakud, mille tsentraalne osa läheb üle
peeneks jätkeks, mis põimub teiste samalaadsetega haistmisnärviks.
(Dendriitidel on retseptorid lõhnaainetele, aksonid suunduvad
kimpudena haistmis sibulasse)Haistmisnärvid liiguvad mulkude kaudu
koljuõõnde, kus nad lõppevad haistmissibulas.Haistmisrakkude
neuronis on haistmissibulas tugevasti seotud mitraalrakkude
dendriitidegaja moodustavad haistmispäsmakesi.Mitraalrakud
moodustavad haistmistee põhimassi ja nad on veel ühenduses ka
pidurdusvaheneuronite: periglomerulaar ja sõmerrakkudega. See
piirkond kannab nime haistekolmnurk. Haistmistee suundub
frontaalsagarasse, haistmiskoorde. Haistmisteed ulatuvad veel ka
limbilisse süsteemi, hüpotaalamusse ja suurajukoorde, mille kaudu
tekib haistmistaju teadvuse tasemel Kes tahab veel rohkem lisa teada,
siis sinine raamat lk 210-212
26. Maitsmismeelega seotud retseptorid. Maitsmismeelega seotud
juhteteed ja ajupiirkonnad. – Maitse- e maitsmissensorid
asuvad keele pealispinna keelenäsadel. Eristatakse seen -, leht-
ja vallnäsasid, mis moodustavad maitse- e maitsmisrakud.
Maitsmiskarikate ümbruses paiknevad näärmerakud, lahustunud ained
jõuavad sensoriteni difusiooni teel. Sensorrakkude äär moodustab
mikrohattusid, mis suurendavad oluliselt kontaktipinda
vesikeskkonnaga. Maitsmisrakud uuenevad pidevalt.
Seennäsasid on arvult mõnisada, nad asuvad keele eesosal ja
sisaldavad 1-5 maitsekarikat, mis reageerivad magusale,
soolasele ja ka hapule. Lehtnäsad paiknead keelepäral ja
keele külgmistel osadel ning reageerivad hapule. Vallnäsad
(igaühes umbes 200 maitsekarikat) asuvad keele tagumisel osal
(keelepäral). Maitsekarikaid esineb ka epiglottisel, suulael ja
söögitoru ülemise kolmandiku limaskestal. Kokku on
maitsmiskarikaid umbes mõni tuhat . Tipus olevad mikrohatud moodustavad vastuvõtva välja: Moodustavad sünapsi sensoorsete närvidega oma põhimikul: Tugirakud; Basaalsed rakud.
Maitseraku distaalne osa on kaetud mikrohattudega. Molekul ,
mis ärritab maitsesensorit, seondub spetsiifilises äratundmiskohas,
selle tagajärjel tekib ioonkanalite läbilaskvuse ja
membraanipotentsiaali muutus. kui sensoripotentsiaal saavutab
lävitugevuse. Siis vallanduvad aktsioonipotentsiaalid, mis levivad
mööda maitsmismeele sensoritega ühenduses olevaid närvikiude.
Erinevad maitsekvaliteedid realiseeruvad erinevate neurokeemiliste
radade kaudu. Adekvaatne stiimul depolariseerib sensoorse raku
membraani, tekib AP ning selle tulemusena avanevad pingeletundlikud
Ca2+ kanalid, mille tulemusena vallandub neuromediaator
(põhiliselt glutamaat) ning sensoorsel kiul tekib AP. Keele
eesmiselt osalt annavad maitsetundlikkust edasi näonärvi
perifeersed jätked, rakukehad asuvad geniikuliganglionis,
tsentraalsed jätked lõpevad maitsmistuumas, mis asub solitaartrakti
e üksikkulgla tuuma ülemises osas. Keele tagumiselt osalt
juhib maitsetundlikkust keele-neelunärv, mille tsentraalsed
jätked lõpevad samuti maitsmistuumas. Üksikkulgla tuumast
suunduvad närvikiud mediaallingu kaudu talamuse kõhtmisse ja
posteriomediaalsesse tuuma ning läbi sisekihnu suurajukoore
posttsentraalkääru piirkonda.
Maitsmissensorid paiknevad keele pealispinna keelenäsadel või selle
lähedases ümbruses spetsiaalsed epiteelrakud .Keelenäsadel
eristatakse seen -,leht-ja vallnäsasid, mis moodustavad
maitsmiskarikaid( ühes karikas u 100 rakku)
Maitsmiskarikate ümbruses paiknevad näärmerakud, lahustunud
ained jõuavad sensoriteni difusiooni teel.Seennäsad: arvult
mõnisada, keele eesosal, 1-5 maitsekarikat, reageerib magusale,
hapule ja soolasele. Lehtnäsad: keelepäral ja külgmistel osadel,
reageerib hapule. Vallnäsas on u 200 maitsekarikat, keele tagumisel
osal.Maitsesensorid on molekulide suhtes spetsiiflised. Juhul kui
saavutatakse sensoripotentsiaal, vallanduvad AP-d, mis levivad mööda
maitsemeele sensoritega ühenduses olevaid närvikiude. Keele
eesmiselt osalt annavad maitsetundlikkust edasi näonärvi
perifeersed jätked, mille rakukehad on geniikuliganglionis,
tsentraalsed jätked lõppevad maitsmistuumas, mis asub
solitaartrakti e üksikkulgla tuuma ülemises osas. Keele-neelunärv
juhib keele tagumise osa maitsetundlikkust, ka siin lõppevad
tsentraalsed jätked maitsmistuumas. Üksikkulgla tuumast liiguvad
närvikiud mediaallingu kaudu talamuse kõhtmisseja
posteriomediaalsesse tuuma ning sisekihnu läbi suurajukoore
posttsentraalkääru piirkonda. Muud juttu tajumiste kohta võib
lugeda sinine raamat lk 214
27. Refleksid: nende määratlus, võimalikud klassifitseerimise
viisid. Reflekside osatähtsus organismi motoorses talitluses.
- Tingimatu refleks – organismi käitumisreaktsioonid, mis on määratud organismi geneetilise programmiga, ilma et ümbritsev keskkond nendele olulist mõju avaldaks. Neid reflekse kutsuvad esile tingimatud ärritajad. Inimesel näiteks imemise, haarde-, kõhimise, aevastamise, silmasulgemise jmt refleksid
- Tingitud refleks – selle väljakujundamisel on oluline meeles pidada, et:
- Tingitud refleks kujuneb välja tingimatu või varem omandatud tingitud refleksi alusel
- Ärritaja, millele tahetakse välja kujundada tingitud refleksi, ei kutsu algul esile mingit kindlat vastust, põhjustab üldise orienteerumisreaktsiooni (silmade ja pea pööramine, liikumine ärritaja suunas). Sellised ärritajad on neuraalsed ja kaotavad tähenduse, kui neid ei kinnitata tingimatu ärritajaga.
- Indiferentne (neuraalne) ärritaja peab eelnema tingimatule ja toimima mõnda aega koos viimasega
- Tingitud refleksi väljakujunemiseks on vaja neid katseid teatud vaheaja möödumisel korrata
- Tingitud refleksi väljakujundamise ajal ei tohi samal ajal eineda teisi tugevaid ärritajaid.
Kui tingitud regleksi pikka aega ei kinnitata, siis see kustub ,
väljakujunenud seos kaotab oma bioloogilise tähenduse. Kui tegevuse
resultaat ei vasta püstitatud eesmärgile, vallandub
orienteerumisreaktsioon, mis viib organismi käitumisprogrammi
muutmisele.
- Operantne tingitud refleks – tingitud refleksi erivorm, kasutatakse ühe võimalusena loomadega suhtlemisel. See kujutab endast sisuliselt katseloomal või linnul tingitud refleksi väljakujundamist, millest saab omapärane suhtluskeel . Katseloomal kujundatakse mingile kindlale ärritajale välja motoorne tingitud refleks.
28. Aju funktsionaalsete süsteemide ülesehituse põhimõtted.
Kesknärvisüsteem koosneb neuronitest ja neid toetavatest gliiarakkudest . Perigeerias asuvate sensorite ja efektoritega on KNS seostatud aferentsete (sensoorsete) ja eferentsete ( motoorsete ja
sekretoorsete) närvikiudude kaudu. Neuronite kehad moodustavad
kogumeid – tuumad , neid ühendavad aksonite poolt moodustatud
närvikiudude kimbud – juhteteed e traktid e kulglad. Juhteteed
jaotatakse alanevateks (kannavad sensoorset infot) ja ülenevateks
(kannavad eferentseid signaale). Üks osa närvirakke võib
sekreteerida neurohormoone.
Inimese aju funktsioonid on piirkonniti erinevad. Mingi talitluse
funktsioneerimisest ja juhtimisest osa võtvad neuronid moodustavad
närvikeskuse, sinna jõuab info, seda analüüsitakse, võetakse
vastu otsus, saadetakse see efektorneuronitele ja sealt
täidesaatvatele elundiele
Närvikeskustes levib erutus ühes suunas – sensoriga ühenduses
olevalt neuronilt efektorneuronile. Erutuse juhtimine närvikeskustes
aeglustub, sest mediaator peab difundeeruma läbi sünapsipilu ja
esile kutsuma postsünapsimembraanil kas erutava või pidurdava
postsünaptilise potentsiaali.
Erutus võib närvikeskustes summeeruda. See võib olla:
- Ajaline summatsioon – ühekordne alalävine ärritaja erutust esile ei kutsu, erutus tekib siis, kui sama tugevusega ärritajat korrratakse suurema sagedusega. Rakumembranidel tekivad hüpopolariseerivad potentsiaalid , mis summeeruvad ja saavutavad lõpuks kriitilise depolarisatsiooni läve.
- Ruumiline summatsioon – samaaegne kahe või enama piirkonna sensorite ärritamine alaläviste ärritajatega vallangab erutuse. Ühe piirkonna sensorite ärritamine alalävise ärritajaga erutuse teket ei põhjusta.
Erutuse levikus avaldub:
- Divergents – ühe närviraku aksoni kaugu sisenev erutus algatab erutuse paljudes närvirakkudes
- Konvergents – erutus koondub suuremalt arvult närvirakkudelt väiksemale arvule.
Rütmi transformatsioon – keskusesse saabuvate ja väljuvate
impulsside arv ei ole üks ja sama. Sagedus võib nii suureneda kui
ka väheneda.
Kord tekkinud erutus võib püsida keskuses ka pärast ärritaja
lakkamist, seda kutsutakse järeltoimeks. Erutust on võimalik
ringluses hoida neuronite ahelate vahendusel nii, et jõuab lõpuks
tagasi sinna kus tekkis. Sellist ringlust nimetatakse
reverberatsiooniks. See on oluline mälu mehhanismideks info
läbitöötamisel ja säilitamisel.
Pidurdusnähud – erutuse leviku blokeerimine võimalik tänu
pidurdusneuronitele.
- Presünaptiline pidurdus – väheneb mediaatoraine vabanemine presünapsis ja takerdub erutuse läbiminek. Blokeeritakse erutus enne närvirakuni jõudmist.
- Postsünaptiline pidurdus – vallanduv mediaatoraine kutsub esile postsünapsimembraanil K+ või Cl- ioonide läbilaskvuse suurenemise – tekib pidurdav postsünapsiline potentsiaal.
- Lateraalne pidurdus – pidurdus lähtub sama sensoorse tee kõrvalasuvatest neuronitest.
29. Perifeerse närvisüsteemi anatoomilis-funktsionaalne
iseloomustus. Spinaalnärvid.
Kraniaalnärvid.
Kesknärvisüsteem koosneb neuronitest ja neid toetavatest
gliiarakkudest. Perifeerne närvisüsteem koosneb 12 paarist
kraniaalnärvidest
ja 31 paarist seljaajunärvidest.
See närvisüsteemi osa on vahendavaks lüliks (välis)keskkonna ja
KNS vahel. Perifeerne närvisüsteem jaguneb somaatiliseks
ja autonoomseks
e. vegetatiivseks närvisüsteemiks(ANS).
Somaatiline närvisüsteem on seotud info juhtimisega vastavatelt
retseptoritelt
KNS poole (aferentne e. sensoorne osa) ja kesknärvisüsteemist
lihaste, liigeste ,
mõningate näärmete
või muude efektororganite suunas ( eferentne e motoorne ja
sekretoorne osa). Neuronite kehad moodustavad tuumi ning neid
ühendavad närvikiudude kimbud juhteteed e traktid e kulglad:
Ülenevad juhteteed: sensoorsete infot kandvad
Alanevad juhteteed: eferentseid signaale edastavad
Seljaaju hallaine teatavaid piirkondi koos nendele vastavate 31 paari
seljaajunärvidega nimetatakse segmentideks. Sellega, et
refleksikeskused asuvad teatud segmendis, on võimalik hinnata
seljaaju funktsiooni (põlverefleks).
Spinaalnärvid: Spinaalnärvid on paarilised närvid (31 paari). Iga
spinaalnärv jaguneb kohe peale lülisambakanalist väljumist ventraal - ja dorsaalharuks. Spinaalnärvi dorsaaljuured (tunde ehk
toomajuured) koosnevad aferentsete närvikiudude kimpudest ,
ventraaljuured ( motoorsed ehk viimajuured) – eferentsete
närvikiudude kimpudest
Seljaaju kaudaalsest osast väljuvad nimme -, ristluu- ja õndranärvide
juured kulgevad allapoole püstiselt, moodustades koos lõppniidiga
närvijuurte kimbu – nn. Hobusesaba. Pärast seljaju väljumist
liituvad dorsaal - ja ventraalnärvijuur vastavate lülidevaheliste
mulkude piirkonnas spinaalnärviks. Enne ventraaljuurega liitumist
moodustab iga dorsaaljuur närvisõlme – spinaalganglioni
(aferentsete neuronite kehade kogumik)
Ventraalharud on kõige jämedamad ja moodustavad omavahel ühinedes
närvipõimikuid , mis on tekkinud seoses jäsemete arenguga (kaela-,
õla-, nimme- ja ristluupõimik). Põimikust lähtuvad naha-, lihase-
ja seganärvid. Rinnanärvide ventraalharud põimikuid ei moodusta ja
kulgevad roietevahemikes
Ventraalharud innerveerivad kõhtmist kerelihastikku (kaela-, rinna-
ja kõhulihaseid), jäsemete lihaseid ning nende piirkondade
liigeseid ja nahka. Dorsaalharud innerveerivad selgmist
kerelihastikku (süvasid seljalihaseid), selja nahka ja
lülidevaheliiigeseid
Nervus craniales - kraniaalnärvid Saavad alguse
peaajust ja mis on nummerdatud vastavalt lähtekohale eest poolt taha
poole.
Haistenärvid (S)
Nägemisnärv (S)
Silmaliigutaja närv (M)
Sisaldab
ka parasümpaatilisi kiude. Liigutab kõiki (va kahe) silmamuna
liigutavat lihast silmakoopas.
Plokinärv (M)
Innerveerib
ülemist põikilihast silmamunas
Kolmiknärv (MS)
Tundeerutused
lauba, pealae, silmakoopa ja ninaõõne piirkonnast .
Erutused
põse, ninaõõne, ülalõualuu, suulae piirkonnast
Tundeerutused
alalõua, oimu, suuõõne alaosalt ja keelelt, alalõua hammastelt.
Innerveerib mälumislihaseid.
Eemaldajanärv (M)
Innerveerib
silmamuna külgmist sirglihast.
Näonärv (MS)
Valdavalt
motoorne, innerveerib miimilisi lihaseid. Sensoorsed edastavad
maitseerutusi.
Tasakaalu-kuulmisnärv (S)
Kaksiknärv,
tasakaaluerutusi ja kuulmiserutusi.
Keele-neelunärv (MS)
Maitseerutusi
keele tagaosast, motoorne osa neelamislihaseid
Uitnärv (MS)
Peamiselt
parasümpaatilistest kiududest
innerveerivad pea kogu vegetatiivset osa. Motoorsed innerveerivad
kõri, neelu alumisi lihaseid. Sensoorsed toovad erutusi välisest
kuulmekäigust, neelust, kõrist.
Lisanärv (M)
Innerveerib
peapöörajat ja trapetslihast, teine haru liitub uitnärviga.
Keelealune närv (M)
Innerveerib
keele lihaseid.
M – motoorne närv, S – sensoorne närv, MS – mõlemad
30. Seljaaju ehitus ja talitlus
Seljaaju asub lülisambakanalis ja koosneb närvirakkude kogumikest
(hallaine) ja seda ümbritsevate närvikiudude ( valgeaine ) poolt
moodustatud juhteteedest. Hallainel eristatakse ees- ja tagasarvi,
rinna- ja nimmepiirkonnas veel külgsarvi.
Seljaaju ülesanded:
Reflektoorsete reaktsioonide keskuste funktsioon – toimuvad refleksid, mis ei vaja kõrgemate ajuosade osavõttu
Juhtefunktsioon – vahejaam erutuse ülekandel teistesse närvikeskustesse.
Seljaaju teatavaid piirkondi koos neile vastavate 31 paari
seljaajunärvidega nimetatakse segmentideks. Vastavalt seljaaju poolt
innerveeritavatele piirkondadele saab keha jagada segmentideks –
metameeridek – teatud seljaaju osa kontrollib teatud keha osa, saab
kontrollida seljaaju tööd (põlverefleks)
Seljaaju saab impulsse:
- Puutesensoritelt
- Valusensoritelt
- Temperatuurisensoritelt
- Propriosensoritelt
- Vistserosensoritelt
Need impulsid liiguvad seljaaju selgmisse juurde – spinaalganglioni
(moodustunud närvirakkudest) – sensoorsed närvikiud. Impulsid
liiguvad seejärel tagasarvest vaheneuronile ja sealt eessarvedele
(seljaaju kõhtmised juured), millest väljuvad motoorsed närvikiud.
Pärast seljaajust väljumist ühinevad motoorsed ja sensroosed kiud
seljaajunärvide paariks.
Juhteteedest eristatakse:
- Ülenevad – sensoorseid impulsse kandvad, perifeeriast seljaajju sisenevad. Tähtsamad asuvad selgmiste ja külgväätide eesmistes osades, nad on:
- Seljaaju- piklikaju kulgla – moodustub spinaalganglionide närvirakkude jätketest, jaguneb:
- Õrnväät – juhib alakeha lihaste ja liigeste impulsse
- Talbväät – juhib ülakeha lihaste ja liigeste impulsse
- Seljaaju-talamuse kulgla – külgmine ja kõhtmine – talamuse kaudu suurajukoorde.
- Seljaaju-väikeaju kulgla – selgmine ja kõhtmine – impulsid väikeajju
- Seljaaju- katendi kulgla – viiakse valu keskaju piirkonda
- Alanevad – kõrgematelt ajuosadelt pärinevad enamasti motoorseid impulsse kandvad. Paiknevad seljaaju valgeaine ventraal- ja külgväätide mediaalses osas, kannavad eferentseid impulsse ja juhivad erutust peaajust seljaaju eessarvede motoorsete rakkudeni, kust liigub edasi skeletilihasteni. Tähtsamad alanevad juhteteed on:
- Püramiidi süsteem – selle kaudu juhitakse meie tahtlikke liigutusi. Peaaju motoorse ala püramiidrakkude jätketest liiguvad seljaaju eessarvede motoneuronitesse.
- Mittepüramiidisüsteem – seovad ajukoorealuseid tuumasid peaajunärvide tuumade ja seljaaju eessarvede motooresete rakkudega. Selle kaudu reguleeritakse lihastoonust ja koordineeritakse keha asendi säilitamist. Juhitakse automaatseid liigutusi. See jaguneb:
- Punatuuma-seljaaju kulgla – selle kaudu allutatakse seljaaju motoneuronid juttkeha ja väikeaju kontrollile .
- Külgmine ja kõhtmine retikulaarformatsiooni-seljaaju kulgla – seob ajutüve retikulaarformatsiooni ja seljaaju essarvede motoorseid rakke
- Esiku-seljaaju kulgla – reguleeritakse tasakaalumeele impulssidega lihaste toonust tasakaalu säilitamiseks
- Katendi-seljaaju kulgla
31. Piklikaju, ehitus ja talitlus. Retikulaarformatsioon e.
võrkmoodustis.
Piklikaju läbivad alanevad ja ülenevad juhteteed, millest üks osa
ümber lülitatakse.
Piklikaju närvikeskuste kaudu reguleeritakse:
- Hingamiselundite
- Südame ja veresoonkonna talitlust
- Paiknevad peaajunärvi (IX,X,XI,XII) ehk keele, neelu,uit,lisa- ja keelealuse närvi tuumad.
- Piklikaju närvikeskuste kaudu toimuvad reflektoorsed tegevused- imemine,neelamine,oksendamine, aevastamine,köha.
Retikulaarformatsioon:
Nimetus tuleneb sellest, et ajutüve keskosas asuvad hajutatult
närvirakkude kogumikud, mis on läbi põimunud võrgustikku
meenutavate närvikiududega.
Retikulaarformatsiooni ülenevad mõjud:
Aferentsed teed toovad infot
seljaajust,väikeajust,talamusest,hüpotalamusest,basaalganglionidest,peaajukoorest,
meelesüsteemidest.
Aferentsed kiud moodustavad RF neuronitel sünapse.
RF mõju on oluline
ärkvelolekuks,tähelepanureaktsioonide,orienteerumisreflekside
teostamiseks.
RF abil saab muuta meelesüsteemidelt lähtuvat sensoorset infot kuni
kortikaalsete mõjudeni välja. RF erutavaid mõjusid
tasakaalustatakse pidurdavatega, nii hoitakse ära suurajukoore
hüperaktiivsuse teke.
RF võtab osa kogu organismi vegetatiivsete ja homöostaatiliste
reaktsioonide korraldamisest. RF kaudu kulgevad paljud
destsendeeruvad mõjustused, mis muudavad seljaaju reflekse.
RF alanevad mõjud:
Võivad olla pidurdavad või erutavad. Pdurdavad mõjud piklikaju
piirkonnast. Erutav tsoon algab vaheaju keskosast, haarab keskaju ja
jõuab seljaajuni.
Arvatakse et RF kaudu muudetakse seljaaju A+(alfa) motoneuronite
aktiivsust.
A+ gamma -motoneutronite kaudu realiseeruval RF mõjul oluline koht
keha asendit,lihastoonust,liigutusi reguleerivates
kohastumisreakstioonides.
32. Ajusilla ja väikeaaju ehitus ning talitlus.
Ajusild kuulub tagaaju koosseisu, nagu ka väikeaju. Sillast saavad
alguse V,VI,VII,VIII peaaju närv
e. kolmik -,eemaldaja-,näo-,esikuteonärv, mille tuumad asuvad
tagaajus. Sillas paikneb trapetskeha ja üks osa hingamisneuronite
tegevust reguleerivaid närvirakke.
Väikeaju:
Koosneb kahest poolkerast ja neid ühendavast
ussist.Poolkerasid katab väikeajukoor. Väikeaju valgeaine on
ühendatud teiste ajuosadega alumiste, kesksete ja ülemiste
väikeajujalakeste kaudu. Olulisemateks ühendusteedeks on
seljaaju-väikeaju kulgla (toob impulsse) ja esiku-väikeaju kulgla
(seob tasakaalumeele esikuteonärvi tuumasid väikeajuga), mille
kaudu tuuakse infot pea,keha, jäsemete asendi kohta. Tähtsamad
juhteteed seovad väikeaju kesk-,piklik-,seljaajuga, tähtsamad
ühendusteed suunduvad hüpotaalamusse,talamusse,punatuuma,piklikaju
ja RFi tuuma.
Väikeaju kaudu reguleeritakse automaatset motoorikat ning tahtlike
liigutuste ulatust ja jõudu, korrigeeritakse aeglasi liigutusi,
programmeeritakse kiireid liigutusi.
Kahjustuse korral häiritud liigutuste koordinatsioon , esineb
ataksia, tasakaaluhäired,astaasia(seismisraskused),
atoonia(lihastoonuse langus), asteenia (kiire
lihasväsimus),adiadohhokinees.
33. Keskaju ehituslikud ja talituslikud iseärasused.
Keskaju koosseisu kuuluvad suurajujalakesed,katteplaat,punatuum ja
mustaine. Siin asuvad III ja IV peaajunärvi
(silmaliigutaja,plokinärvi) tuumad.
Katteplaadi ülaküngastes lülitatakse ümber nägemismeele ja
alaküngastes kuulmismeele tsentraalseid teid.
Keskajus asub pupillirefleksi keskus.
Mustaju kaudu reguleeritakse neelamis ja mälumisliigutusi.
Punatuum seostub punatuuma-seljaaju kulgla ja ajukoore, koorealuste
tuumade ning väikeaju kaudu seljaaju eessarvede motoorsete
rakkudega, mille abil korrigeeritakse lihastoonust.
34. Vaheaju ehituslikud ja talituslikud iseärasused. Hüpotalamus. Talamus .
Vaheajju kuuluvad talamused ja hüpotalamused, ka kolmas ajuvatsake.
Talamustesse koonduvad aferentsed impulsid, nende kaudu kulgeb
meeleelunditelt saadud info kõrgematesse ajuosadesse. Talamuste taga
paiknevad põlvikkehad, mida läbivad nägemis-ja kuulmismeele tee.
Lisaks asuvad talamuses piirkonnad, mis on seotud ajukoore
assotsiatsiooniväljadega.
Talamuse kaudu mõjutatakse tahtmatuid, emotsioone väljendavaid
liigutusi.
Hüpotalamus asub talamusest allpool, on vegetatiivsete
funktsioonide kõrgemaks keskuseks, mille kaudu reguleeritakse
ainevahetust, kehatemperatuuri,toitekäitumist.
Siin asuvad osmootse rõhu sensorid, mis hoiavad tasakaalus vee ja
soolade sisaldus.
Oluline roll homöostaasi säilitamisel,kuna on neurohormoonide
vahendusel seotud hüpofüüsiga.
Hüpotalamuses produtseeritakse riliising- ja inhibiitorhormoone, mis
soodustavad/pidurdavad hüpofüüsi hormoonide teket.
35. Limbiline süsteem ( mandelkeha ja hipokampus).
Mandelkeha ja hipokampus on vaheaju piirkonnad.
Mandelkeha ärritus põhjustab
raevu-,põgenemisreaktsioone,muutusi vegetatiivses sfääris- pupilli laienemine, vererõhu tõus, südame löögisageduse
tõus,hingamissageduse tõus, süljeeritus,närimisliigutused.
Hipokambi keemiline ärritamine kutsub esile labiilsuse
suurenemise, kergesti vallandub raev ja agressiivsus , on seotud
afektsete reaktsioonidega.
Limbilise süsteemi ja hüpotalamuse vahendusel reguleeritakse
siseelundite aktiivsust ja sisenõrenäärmete tööd, selle kaudu
homöostaasi. Seotud on ka emotsioonid -rõõm, mure, lõbu- ja
norutunne,viha, hirm. On seotud vegetatiivsete reaktsioonidega seoses
emotsioonidega ( higistamine , kahvatumine jne).
36. Ajukoore ehituslikud ja talitluslikud iseärasused. Basaalganglionid . Elektroentsefalograafia .
Suurajukoor- sellega on seotud teadvus, mõtlemine, mälu,
meeleelundite talitlus, aistingute ja tajude teke, õppimine.
Suuraju poolkerade pind on liigendatud sagarateks ja käärudeks.
Selle tõttu on aju pindala 0,2m2.
Suurajukoorel eristatakse projektsiooni - ja assotsiatsiooniväljasid.
Sensoorsed on projektsiooniväljad, kuhu saabuvad aferentsed impulsid
meeleelunditelt, motoorsed need alad, kus suurajukoores paiknevatelt
neuronitelt algavad tahtlike liigutustega seotud püramiidsüsteemi
teed. Tundlikkuse alusel jaotatakse aju 52 väljaks.
Sensoorne informatsioon jõuab suurajukoorde pärast ümberlülitusi
ja töötlusi. Infotöötlusse seoses aistingute ja tajude tekkega on
haaratud assotsiatsiooniväljad, millel ei ole domineerivat
sensoorset ega motoorset funktsiooni.
Motoorsed projektsiooniväljad koondunud otsmikusagara pretsentaalkääru piirkonnas. Siin asuvad närvirakud saadavad
aksonid madalamatele aju-ja seljatüves asuvatele keskustele,
moodustub püramiidsüsteem, millega ühe poolkera tegevus juhib
vastaskehapoole motoorikat.
Assotsiatsiooniväljad saavad infot talamuselt või
projektsiooniväljadelt.Moodustavad kuni pole ajukoore pinnast.Nende
vahendusel kujunevad ärkvelolek,tähelepanuvõime,õppimine, mälu,
tegelikkuse tunnetamine , sihipärane käitumine.
Basaalganglionide hulka kuuluvad kahkjaskera ja juttkeha.
Mõnikord loetakse juurde kuuluvateks ka punatuuma ja mustainet.
Kahkjaskera on on ühendatud kooriku ja sabatuumaga. Tema alanevad
teed seovad teda hüpotalamuse piirkonna RFiga, silmaliigutajanärvi
tuumaga, punatuumaga, mustainega, ajusilla ja piklikaju RFga ning
näonärvi ja keelealusenärvi tuumaga.
Basaalganglionid võtavad koos väikeajuga mittepüramiidsüsteemi
koosseisus osa keha asendi säilitamisest, liigutuste automaatsuse ja
koordinatsiooni regulatsioonist. Tagatakse liigutuste sujuvus.
Basaalganglionide kahjustus avaldub parkinsonismina, esinevad
tahtmatud liigutused. Mis on seotud mustaine, punatuuma ja juttkeha
funktsioonihäiretega. Esinevad ka treemor ja rigiidsus .
Elektroentsefalograafia-ajutegevusega seotud elektriliste
potentsiaalide registreerimine ajafunktsioonina. EGG
registreerimiseks asetatakse elektroodid peanahale, ühendatakse
võimendaja ja registreerijaga. EEG, mis iseloomustab neuronite
seisundit, iseloomustatakse alfa, beeta, delta ja teeta
rütme.Puhkeolekus ilmneb alfarütm, silmade avamisel,valguse toimel,
mõtlemisel beetarütm. Delta ja teetarütm esinevad une, narkoosi ja
teatud haiguslike seisundite korral.

















































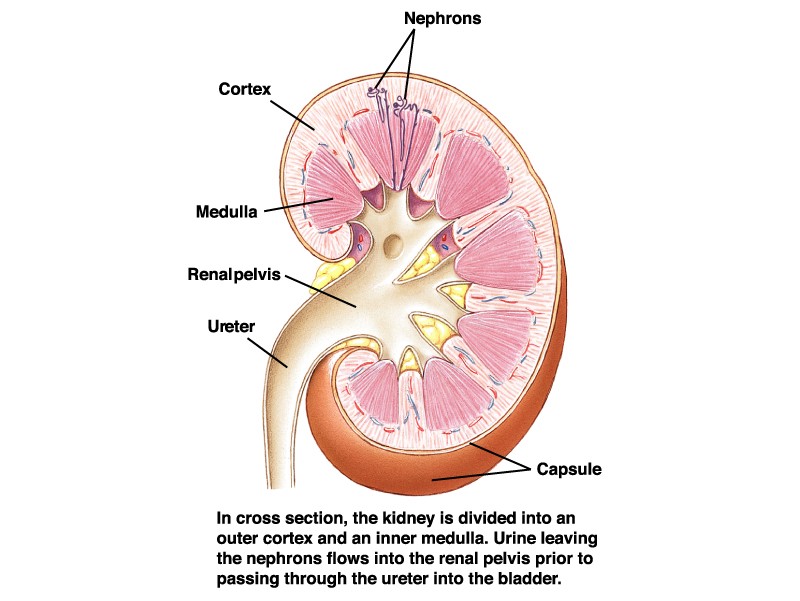




Kõik kommentaarid