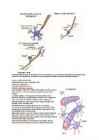
NÄRVISÜSTEEMI BIOLOOGILISED ALUSED EKSAM 1. Närvisüsteemi areng sünnieelsel perioodil 18-28'ndal päeval hakkab moodustuma närvisüsteem ja see hakkab juhtima teiste organite tegevust. Kahe ja poole nädala vanusel lootel tekib keha dorsaalküljel ektodermi paksend neuraal- e medullaarplaat, mis kiirelt muutub neuraalvaoks ja seejärel sulgub neuraaltoruks. neuraaltoru seintest kujunevad närvi- ja gliiarakud KNS-s, ruumidest neuraaltoru sees areneb välja ajuvatsakeste süsteem.Neuraaltoru kaudaalne osa on algmeks seljaajule ning rostraalne osa peaajule. Neuraaltoru
Meelesüsteemi talitlus Sensoris muudetakse ärritaja energia sensorimembraani permeaabluse muutuste kaudu sensoripotentsiaaliks (SP) transduktsioon 1) SP tekib sensorimembraanil ja on lokaalne potentsiaal 2) muutub astmeliselt ja sõltub ärritaja tugevusest 3) levib mööda membraani elektrotooniliselt 4) summeerub nii ajaliselt kui ruumiliselt 5) vallandab sensoriga ühenduses olevas aferentses närvis AP tekke ja kannab seetõttu nimetust generaatorpotentsiaal SP põhjustab sensoriga ühenduses olevates närvikiududes aktsioonipotentsiaalide tekke(transformatsioon), mis juhitakse aferentsete juhteteede kaudu meelesüsteemi kuuluvatesse erinevatesse KNS osadesse. Sensorite jaotus vastavalt asukohale · eksterosensorid kohandunud välismaailma ärritajate vastuvõtuks; neid võib jaotada veel ärritaja ja sensori vahekauguse alusel
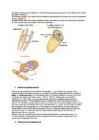
NÄRVISÜSTEEM SYSTEMA NERVOSUM Mõisted NEURON - närvirakk + jätked SÜNAPS - neuronite kontakt, kus erutus kandub ühelt neuronilt teisele v lõppelundile MEDIAATOR - e neurotransmitter - närviraku impulsi toimel sünapsis moodustunud keemiliselt aktiivne aine, mille varal toimub erutuse ülekanne (atsetüülkoliin, noradrenaliin) - nr jätke, mida mööda juhitakse erutus neuroni suunas: lühike puuvõratoline või DENDRIIT niitjas - neuroni jätke, mida mööda juhitakse erutust neuronist välja / neuroni jätke, mis juhib AKSON närviimpulsse nr-st kas teise nr, moodustades sünapsi või efektoorse lõppelundi kaudu lõppelundisse, nt lihasesse - närvisüsteemi tugirakud (kaitse-, tugi-, toitev ja AV-funktsioon) NEUROGLIIA - e närvisõlm e närvitänk e tänk -
a. on see toidulisandina müügil USA-s. Postsünaptilise potentsiaali pikkus oleneb mediaatori eemaldamisest postsünaptilisest pilust. Mediaatori eemaldamine on erinevate mediaatorite jaoks erinev, osad (nt ACh) hüdrolüüsitakse sünaptilises pilus, teised (nt serotoniin) seotakse taas presünaptilises rakus ja kasutatakse hiljem uuesti. 3. Refleksi ja refleksikaare mõiste ja osad. Refleks on organismi vastus ärritusele. Selle kaudu närvisüsteem reguleerib org talitust. Refleks realiseerub mööda refleksikaart. See on tee mille erutus läbib vastuvõtvalt retseptorilt (sensorilt) kuni täidesaatva elundini. Refleks toimub teatud aja jooksul, seda 1 nim refleksiajaks. Erutuse ülekanne toimub kas närvilt närvile või närvilt erudeeritavale elundile. Olgu selleks lihas vms. erutuse ülekande kohta kutsutakse sünapsiks.
FÜSIOLOOGIA (KKSB.02.046) EKSAMIPROGRAMM - kevad 2013 Närvisüsteemi talitlus (I kontrolltöö osa) Närvisüsteemi üldine ülesehitus ja eri osade peamised ülesanded. Kesknärvisüsteem: pea- ja seljaaju. Perifeerne närvisüsteem: aferentne e. sensoorne ja eferentne e. motoorne osa; eferentse osa jagunemine somaatiliseks motoorseks ja autonoomseks närvisüsteemiks; autonoomse närvisüsteemi sümpaatiline ja parasümpaatiline osa. Autonoomse närvisüsteemi troofiline ja funktsionaalne mõju siseelundite talitlusele. Neuronid ja neurogliia rakud. Neuroni üldine ehitus. Neuronite tüübid: funktsiooni alusel, struktuuri alusel. Aksoni üldine ehitus.
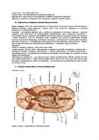
Närvisüsteem: Kesknärvisüsteem , Seljaaju, Peaaju, Perifeerne närvisüsteem Somaatiline: närvid Autonoomne ehk vegetatiivne närvisüsteem Sümpaatiline Parasümpaatiline Seljaaju ( medulla spinalis ) KNS vanim ja madalam osa Funktsioonid: Reflektoorne – seljaajus paiknevate motoorsete keskuste vahedusel toimuvad spinaalrefleksid, mis ei vaja kõrgemate ajuosade osavõttu Juhtefunktsioon – seljaaju on vaheajaam erutuse juhtimisel kõrgematesse keskustesse ja kõrgematest keskustest teostusorganitesse Seljaaju ehitus Võrreldes loomadega on inimese seljaaju väiksema iseseisvuga. Seljaaju kaal
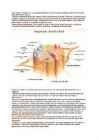
närvikiud - pikk närviraku jätke; koosneb kesksest telgsilindrist ja seda ümbritsevast neurilemmist innerveerima s.o. närvidega varustama aferentne e. sensoorne (tooma)närv - tundenärv eferentne e. motoorne (viima)närv Närvisüsteemi ülesanded, ehitus ja jaotus http://www.sruweb.com/~walsh/nervous_syste m.jpg Närvisüsteem (NS) on organismi juhtiv regulatsioonisüsteem. Ta täidab kaht olulist ülesannet : 1. loob organismi sideme väliskeskkonnaga 2. ühendab ja kooskõlastab keha organsüsteemide tegevust organismis kui
1. KÄITUMISE BIOLOOGILISED ALUSED Inimese käitumine alates lihtsamatest asjadest (silmapilgutamine) ja lõpetades kõige keerulisematega (probleemide lahendamine), sõltub meie sees toimuvate protsesside integreerumisest. Integreerimist e ühendamist teostab närvisüsteem (NS). Et paremini mõista inimkäitumist, tuleb tunda selle aluseks olevaid bioloogilisi protsesse. Teadusharu, mis uurib organismi bioloogiliste ning psüühiliste ja käitumuslike aspektide vahelisi seoseid, kuuluvad psühhobioloogia valdkonda. Need on neurofüsioloogia, sotsioloogia, psühhofarmakoloogia jpt. Närvisüsteem NS ülesanne on reguleerida ja koordineerida organismi elundite talitlust ning kohandada seda sise- ja väliskeskkonna muutustele.






































































Kõik kommentaarid