1
MOLEKULAARBIOLOOGIA .
1. Kui
aatom loovutab elektroni täielikult teisele aatomile, missugused keemilise sidemega on tegemist?
Ioonside, sellised ained lahustuvad hästi, kuna ioonide hüdratatsioonienergia on suurem kui kristalli võreenergia
2. Miks vesi on hea lahusti (solvent)?
Vesi on hea lahusti, sest ta
lahustab nii
tahkeid , vedelaid kui ka gaasilisi aineid. Vee
molekul moodustab
dipooli ning
aatomid omandavad
osalise laengu.
Polaarsete ühenditega moodustab vesiniksidemeid, mis tagavad
stabiilsust.
3. Termodünaamika II seadus.
Kõik protsessid kulgevad tasakaalu e. minimaalse potentsiaalse energia poole e.
entroopia kasvu suunas.
Entroopia (S) on korrastamatuse mõõt [J/mol*K], korrastatud madal entroopia. Isoleeritud süsteemid püüavad
korrastatud olekust korrastamata poole. Tasakaal on siis, kui entroopia on maksimaalne.Entroopia muutus on
null pöörduvate protsesside ja positiivne pöördumatute protsesside korral. Metaboolsed protsessid on üldjuhul
korrastavad, need saavad toimuda ainult siis, kui neid tasakaalustab keskkonna korrastamatus.
4. Mis on
kiraalsus ja kuidas seda kasutab loodus? ravimitööstus?
Kiraalsus on olukord, kus tetraeedriline sidemete
asetus (sageli asümmeetrilise süsiniku korral) võib
kolmemõõtmelises ruumis olla kaht erinevat moodi tekivad peegelpildid. Selliseid molekule kutsutakse
optilisteks või stereoisomeerideks. Enamik molekule rakus koosnevad vähemalt ühest asümmeetrilisest süsiniku
aatomist, mida kutsutakse ka kiraalseks süsiniku
aatomiks . Looduses on kõik
suhkrud D-isomeerid ja kõik
aminohapped L-isomeerid. D-L süsteemi aluseks on glütseeraldehüüdi paigutus (2C OH paremal/vasakul).
Ravimitööstus peab õigete ravimite tootmiseks arvestama ka kiraalsusi ja kasutama looduslikke, sest muidu
ravim ei toimi.
5. Eukarüootne
replikatsioon .
Mehanism ja läbiviivad ensüümid.
Replikatsiooni
mehhanism pärineb peamiselt viiruse
SV40 replikatsiooni uurimisest. Replikatsioon algab järjestusest, mida kutsutakse origin'iks, igas DNA-s on neid palju. DNA polümeraasid ei saa DNA-
ahelat lahti keerata. Replikatsioonis üks ahel pikeneb järjest (juhtiv ahel) ja teine pannakse kokku juppidest (mahajääv ahel), mis ühendatakse DNA-
ligaasi abil.
Helikaas keerab biheeliksi lahti kasutades ATP energiat. Primaas teeb lühikese RNA praimeri, mis on komplementaarnte templeit DNA-ga ja seejärejl hakkab polümeraas seda
pikendama moodustades lühikese 5'RNA-3'DNA tütarahela. Seejärel võtab polümeraas üle ja jätkab ahela
pikendamist. Pol on seotud templeitahelaga seotuse tõttu Rfc-valguga (replication
factor C), mis pöördudes
muutub PCNA-ks (proliferating cell
nuclear antigen trimeerne valk, mis ümbritseb tütarDNA-d). Tavaliselt on
mehhanism kahesuunaline replikatsioonikahvlid liiguvad kahes suunas.
Regulatsioon toimub kuuest MCM
valgust
koosneva helikaasi aktiivsuse kontrollimisega.
Ensüümid: · Helikaas keerab DNA biheeliksi lahti · DNA güraas e. topoisomeraas kompenseerib lahtikeerdumist (aitab struktuuri hoida) · DNA-polümeraasid vastutavad polünukleotiidahelate sünteesi eest 5'->3' suunas o - 4 subühikut, polümeraasi tootlikus, pole 3' eksonukleaasi aktiivsust o - vigade parandus o - replitseeriv ensüüm mitokondrites o - peamine polümeraas! Omab 3'-eksonukleaasi aktiivsust, teeb vähem vigu · DNA-primaas sünteesib oligonukleotiidse praimeri · DNA-
ligaas ühendab mahajääva ahela fragmente 2
6. Mis on Okazaki
fragmendid ja nende rolli DNA replikatsioonil.
Okazaki fragmendid on lühikesed DNA jupid, millest pannakse kokku ahel, mis on komplementaarne DNA
replikatsioonil mahajäävale ahelale. RNA-praimerid, millest fragmendid alguse saavad, lõigatakse ära ja
asendatakse pikeneva DNA ahelaga. Fragmendid ühendatakse DNA ligaasi poolt.
7. Kuidas tekkisid
pseudogeenid ?
Pseudogeen-
funktsionaalse geeniga homoloogiline järjestus, mida ei transkribeerita. Evolutsiooni käigus on
lisandunud neisse geenidesse geneetilise triivi (genetic
drift ) tulemusena rida muutusi, mis translatsiooni
enneaegselt termineerivad või inhibeerivad mRNA protsessingut, nii et need alad on muutunud
mittefunktsionaalseks
ehkki neilt transkribeeritakse
RNAd . Teine võimalus pseudogeenide tekkeks on RNA
pöördtranskriptsioon cDNAks ja viimase integratsioon genoomi
intron -vaba DNAna.
8. Missugused molekulaarsed mehanismid tagavad selle, et inimese genoomi 30,000-40,000 geeni
kodeerivad 100,000 ndeid erinevaid valke?
Alternatiivne splaissing kui tava-arusaama järgi kleebitakse küpse RNA saamiseks
eksonid lihtsalt kokku, siis
alternatiivse splaissingu käigus kasutab
rakk erinevaid eksonite kombinatsioone, et teha erinevaid küpseid
mRNA-sid ja valke.
Alternatiivne
splaising võib toimuda samas raku tüübis
vastuseks erinevatele keskkonna-ja raku arengut
määravatele signaalidele.
Post-translatoorsete modifikatsioonidega saab anda osadele
valkudele erinevaid funktsioone (nii, et need näivad
erinevate valkudena).
9. Missugusi geneetilise info vahetuse protsesse tähistavad a)
transformatsioon , 2)
transkriptsioon 3)
translatsioon . · Transformatsioon geneetilise informatsiooni ülekandumine ühest rakust teise rakust isoleeritud DNA abil. Pärilik muutus raku omadustes. Võõras DNA satub raku koosseisu ja
sellelt hakatakse geene eksprsesseerima. Eristatakse viirusliku päritoluga DNA lisandumist (transduktsioon), bakterite vahelist kontakti (konjugatsioon). Koekultuuris eukarüootsete rakkude
transformatsiooni kutsutakse transfektsiooniks. · Transkriptsioon mehhanism, mille käigus toodetakse DNA-lt ensüümide abiga RNA-d (tuumas) · Translatsioon mehhanism, kus ribosoomides (rRNA+valk) toodetakse mRNA pealt valku, kasutades selleks tRNA transporditud
aminohappeid .
10. Mis on TBP funktsioon initisiatsiooni kompleksi
tekkes ?
38kD TATAbox-siduvat valk (TBP); TBP on esimene valk, mis "istub" TATAbox promootorile. Kõik tänaseks
teadaolevad eukarüootsed TBPd on väga
homoloogsed C-terminuse 180AA ulatuses. Selle valguosa järjestus on
80% ulatuses identne pärmi ja inimese valkudel. TBP Nterminaalne domään on erinevatelt organismidelt pärit
TBP valkudel nii
pikkuselt kui järjestuselt väga varieeruv. Sellel on oluline roll RNAPolII-katalüüsitud
snRNAde geenide transkriptsioonis. TBP
monomeer pakkub moodustades sadul-struktuuri, kus molekuli 2 poolt
omavad diaadset (polaarset, kaheli) sümmeetriat, ent pole identsed. Nagu HMGI ja teised DNAd-
painutavad valgud , interakteerub TBP DNA väikese vaoga, painutades tugevasti kaksikheeliksit. DNAd-siduv valgu pind
on TBP puhul konserveerunud kõigis eukarüootides, mis selgitab ka TATAbox promootorelemendi kõrget
konserveerumist läbi evolutsiooni.
11. Kas transkriptsioonifaktorid (
aktivaatorid ) interakteeruvad otse(vahetult) TBPga?
Jah, Kui TBP istub TATAboxil, siis saab
TFIIB seonduda. TFIIB on monomeerne valk, mis on natuke väiksem
TBPst. TFIIB C-
terminaalne ots interakteerub kahel pool TATAboxi TBP ja DNAga, samas kui tema N-
terminaalne ots stabiliseerib tekkinud
interaktsiooni laiudes START-saidi poole. TFIIB sidumisele järgnevalt
toimub preformeeritud tetrameerse TFIIF ja RNAPolII sidumine. Viimase tulemusena paiguldub RNAPolII
START-saidile. Enamuse promootorite puhul peab kompleksile siduma veel kaks GTFi, et DNA
ahelad saaks
eralduda ja
toimuks matriitsahel
vabanemine . Esimesena
seondub tetrameerne TFIIE, mis tekitab
sidumisvõimaluse TFIIHle (samuti multimeerne faktor, mis koosneb 9 alaühikust). TFIIH kompleksi
seondumine lõpetab transkriptsiooni preinitsiatsiooni kompleksi
formeerumise in vitro.
12. Mis on transkriptsiooni aktivaatori 2 funktsionaalset domääni ja kuidas
mutatsioonid ühes või teises neist mõjutavad transkriptsiooni faktori omadusi?
N-terminaalne DNAd - siduv domään, mis seondub teatud DNA järjestustele, ja C-terminaalne aktivatsiooni 3
domään, mis interaktsioonis teiste valkudega aktiveerib transkriptsiooni.
N-terminuses asuva domääni
mutatsioon võtab ära võime DNA-le seonduda. C- terminaalse domääni
muteerimine vähendab transkriptsiooni aktiivsust. On leitud, et üks või enam transkriptsiooni aktivatsiooni
domääni on seotud järjestus-spetsiifilise DNAd-siduva domääniga linkeralade abil. Mõnedel juhtudel laiub
aktivatsioonidomään DNAd-siduva domääni sisse.
13. Missugused TAFid on TBPga vahetus interaktsioonis?
Olemas on 13 TAF-i, TAF TBP associated factor. TAF-1, TAF-2, TAF-6 (nõrgalt), TAF-11, TAF-12, TAF-13
14. Missugune TFII valk hüdrolüüsib ATPd ja mis protsessiga on tegu?
TFIIH helikaasne aktiivsus saab oma energia ATP hüdrolüüsilt. Selle energia abil keerataksegi START-saidi
piirkonnas DNA dupleksi lahti. See võimaldab RNAPolII-l avatud kompleksi vormis istuda matriitsahela
START-saidile. Kui ka rNTPd on keskkonnas, siis RNAPolII alustab matriitsahela transkribeerimist.
Transkriptsiooni käigus teine TFIIH alaühik fosforüülib paljudest erinevatest positsioonidest RNAPolII CTDi.
In vitro katsetest on selgunud, et TBP jääb TATAboxiga seotuks matriitsahela transkriptsiooni lõpuni, samas kui
ülejaanud GTFid dissotseeruvad kompleksist.
15. Võrdle transkriptsiooni initsitatsiooni protsesse prokarüootidel ja eukarüootidel.
Bakteriaalse polütsistroonse mRNA ribosoomaalsed sidumissaidid asuvad mRNA valkekodeeriva ala ehk
tsistroni ees. Translatsiooni
initsiatsioon võib
alata ükskõik milliselt neist paljudest ribosoomi sidumissaitidest,
mille tulemusena üks mRNA võib kodeerida erinevaid valke.
Enamike eukarüootsete mRNAde puhul määrab 5'-cap struktuur ära ribosoomi sidumiskoha ja translatsioon
algab lähimalt AUG koodonilt. Selle tulemusena translatsioon saab alata vaid ühelt saidilt. Paljudel juhtudel
eukarüootsed valku-kodeerivad primaarsed
transkriptid protsessitakse üht tüüpi mRNAks,
millelt sünteesitakse
ühte tüüpi valk.
Promootorid määravad ära transkriptsiooni initsiatsiooni koha ja suunavad RNA polümeraas II sidumist.
Eukarüoodi DNAs on leitud kolme tüüpi promootorjärjestusi. TATAbox, kõige
sagedasem , mis esineb tavaliselt
kõrge transkriptsiooni tasemega geenide promootorites. Mõnedel geenidel on Initsiaatoriga promootorid ja
teistele on iseloomulikud CpG (saarekestega) promootorid.
16. Mis on
sigma faktori funktsioon? Mis juhtub sigma faktoriga initsiatsiooni lõppedes?
See on prokarüootne transkriptsiooni initsiatsiooni faktor, mis võimaldab RNA polümeraasi spetsiifilist
kinnitumist geeni promootoritele. RNA polümeraas seob sigma faktori et moodustada RNA polümeraasi
holoensüümi kompleksi.
17.
Defineeri prokarüootne operon. Kirjelda geeni aktivatsiooni protsessi prokarüoodis trp või lac operoni näitel.
Geenide klaster, mis moodustab bakteriaalse operoni, sisaldab ühe
transkriptsiooniühiku, kuna ühelt promootorilt transkribeeritakse üks
primaarne
transkript . 1 geen = 1 valk, 1 operon = mitu valku.
18. Mis on peamine erinevus prokarüootse mRNA ja eukarüootse mRNA vahel?
Enamus eukarüootsetest
geenidest kodeerib monotsistroonseid mRNAsid
sisaldades pikki introneid. Paljud
bakteriaalsed mRNAd on
polütsistroonsed, st et üks mRNA
molekul (näit. Trp operoni kodeeriv
mRNA) sisaldab kodeerivaid järjestus, mis kodeerivad mitut, samas
bioloogilises protsessies osalevat valku. Eukarüootsed mRNAd on aga
monotsistroonsed, st iga mRNA molekul kodeerib ühte valku.
Polütsistroonsete ja monotsistroonsete mRNAde erinevus on seotud nende
mRNAde translatsiooniprotsesside fundamentaalse erinevusega. 4
19. Võrdle prokarüootsete ja eukarüootsete RNA polümeraaside struktuuri ja funktsiooni.
Eukarüootidel on kolme tüüpi tuumset RNA polümeraasi. Kõik kolm
polümeraasikompleksi sisaldavad kahte suurt ja kahte väiksemat kompleksi
valku, milledel on suur
homoloogia E. coli RNA polümeraasi , ' ja alaühikutega. Lisaks sisaldavad kõik 3 kompleksi ka hulgaliselt väiksemaid
alaühikuid. Mõnesid väiksemaid alaühikuid need
kompleksid jagavad, mõned
on
unikaalsed . RNA polümeraas I sünteesib ainult pre-
rRNAd . RNA
polümeraas II sünteesib mRNAsid and mõnesid väikseid tuuma RNAsid, mis
osalevad mRNA splaisingus. RNA polümeraas III sünteesib tRNAsid, 5S
rRNAsid, and mitmeid suhteliselt lühikesi stabiilseid RNAsid (U6, splaisingus;
7S,
transpordis ). Trankriptsiooni initsiatsiooni käigus fosforüülitakse RNA
polümeraas II suurima alaühiku karboksüterminaalne domään (CTD), korduv heptapeptiidne järjestus, mis jääb
fosforüülituks kuni matriitsilt transkriptsiooni lõpuni. Sarnaselt bakteri RNA polümeraasiga, initseerib RNA
polümeraas II geeni transkriptsiooni DNA matriitsilt teatud nukleotiidipaari (lämmastikaluste) juurest või
alternatiivina naabernukleotiidipaari juurest. 5'
nukleotiid , mis vastab matriitsahela sellele nukleotiidile, millelt
transkriptsioon algas, mRNAs cap'takse (lisatakse 7'-metüülguanolaat).
20. Joonista tüüpilise
PolII promootori struktuur ja kirjelda faktoreid, mis sinna transkriptsiooni initsiatsiooni käigus seovad. Mediaator
kompleks ja selle tähtsus. Promootorid on TATAbox,
Initsiaator , CpG. 1 TATA-binding
protein seondub, väänab DNA-d 2 TF II B seondub kõikides rakkudes TATA-sse ja initsiatsioonisaidi alasse (tetrameer) 3 polümeraas II saab seonduda, sest TFIIB stabiliseerib tekkinud nurka Tekib preinitsiatsiooni kompleks (PIC), kuhu seondub TFIIE 4 helikaasi aktiivsusega keerab THIIH (9meer) DNA ahelad lahku ja polümeraas saab mööda
matriitsi RNA ahelaid sünteesida ja peale sünteesi algust fosforüülib saba 5 kui saba on fosforüülitud, siis saab transkriptsioon toimuda, kui mitte, seisab paigal. 2 1 TF II D koosneb TBP-st ja 13 TAF-ist (aktiveerivad
faktorid , millest paljud seovad DNA-d, tähistavad või aitavad initsiaatoril elemendile istuda). Mediaator on suur valkkompleks, mis koosneb ~30 alaühikust. Mediaator moodustab silla DNAle seondunud aktivaatori ja promootoril 3 istuva RNAPolII vahel. Lisaks sellele on leitud, et ühel mediaatori alaühikutest on histoonatsetülaasne aktiivsus ning see võib funktsioneerida kui promootori hüperatsetüleerituse hoidja. 21.
PolI ja PolIII-sõltuv transkriptsioon. Mille poolest need erinevad PolII-sõltuvast transkriptsioonist. 4 Transkriptsiooni-regulaatorelemendid geenides, mida transkribeerivad RNA polümeraasid I ja III. Nende geenide initsiatsioonikomplekside kokkupanek algab teatud generaalsete transkriptsiooni faktorite sidumisega regulaatorelementidele. Transkriptsiooni-initsiatsiooni komplekside moodustumine RNAPolI ja RNAPolIII puhul on mõnes mõttes väga sarnane 5 RNAPolII-kompleksi moodustumisele. Samas aga iga kolmest RNAPol-st vajab oma polümeraas-spetsiifilisi generaalseid TF-id ning tunneb ära erinevaid DNA järjestusi. Veelgi enam, ei RNAPolI ega RNAPolIII vaja transkriptsiooni initsiatsiooniks ATP hüdrolüüsi, samas kui RNAPolII vajab. RNAPolI, mis sünteesib pre-RNAsid, ning RNAPolIII, mis sünteesib tRNAsid ning
lühikesi, stabiilseid RNAsid, seoselist transkriptsiooni initsiatsiooni on hästi uuritud pärmi
mudelil . On 5
selgunud, et ribosoomi osade:
tRNAde ja rRNAde süntees on tihedalt seotud raku kasvu ja jagunemise
mehanismidega. Kuigi palju on selles veel ebaselget.
22. Kus toimub transkriptsioon? Kus transkriptsioonifaktorid rakus paiknevad? Too näiteid, kuidas transkriptsioonifaktorite post-translatsioonilised modifikatsioonid mõjutavad nende a) DNAle sidumisevõimet ja b) nende võimet initseerida transkriptsiooni.
Transkriptsioon toimub eukarüootidel tuumas, prokarüootidel tsütoplasmas. Transkriptsioonifaktorid asetsevad
ka eukarüootidel tsütoplasmas, aktiivsed asuvad tuumas ja prokarüootidel tsütoplasmas. Mõnel juhul
transkriptsioonifaktorite tuuma transport
saavutatakse , kui neid ekspresseeritakse liitvalkudena, mis lokaliseerib
nad tsütoplasmaatilises membraanis. Kindlate signaalide korral hakkavad
spetsiifilised proteaasid neid lõikama
ja vabastavad transkriptsioonifaktori domääni, mis imporditakse tuuma. Teistel juhtudel reguleeritakse tuuma
transport post-translatsiooniliste modifikatsioonidega näiteks fosforüleerimine suurendab nende afiinsust
spetsiifilisele eksportiinile. Tuuma impordi ja ekspordi reguleerimine on võimas mehhanism, millega saab
kontrollida transkriptsioonifaktori aktiivsust ta saab siduda
geenidele ainult siis, kui on tuumas.
DNAd-siduvatest valkudest näit.
bHLHd ja bZip
valgud võivad anda alternatiivseid heterodimeere, sõltuvalt siis
nendega interakteeruvatest monomeeridest. Ka teistesse klassidesse kuuluvad transkriptsiooni faktorid võivad
moodustada heterodimeerseid komplekse. Mõnedel juhtudel heterodimeeride alternatiivsed vormid ei mõjuta
DNA-
sidumise spetsiifikat, pigem soodustab iga monomeeriga seotud aktivatsioonidomäänide alternatiivsete
kombinatsioonide teket ning seeläbi mõjutab tekkinud dimeerse transkriptsioonifaktori aktivatsiooniomadusi.
Individuaalsete Tfide aktiivsust saab reguleerida mitmeti. On teada ka inhibitoorsed valgud, mis seonduvad
bZip ja
bHLH valkudega ega luba viimastel interakteeruda DNAga. Rakkudes, kus need inhibiitorid
ekspresseeruvad, nad represseerivad nende faktorite (bZip ja bHLH) transkriptsioonilist aktiivsust.
Happelised aktivatsiooni domäänid on suhteliselt struktureerimata, juhusliku keerd-(coil) konformatsiooniga.
Need domäänid indutseerivad transkriptsiooni, kui nad on kompleksis valgulise ko-aktivaatoriga.
Väga hästi uuritud on imetaja CREB valgu happelise aktivatsioonidomääniga interakteeruvad valgud. CREB
fosoforüülitakse
rakulise cAMP taseme tõustes. Seda reguleeritud fosforüülimist on vaja, et CREB seonduks
oma koaktivaatori CBPga (CREB binding protein) ning aktiveeruksid nende geenide promootorid, mis
sisaldavad CREBi sidumissaite. Kui fosforüülitud juhusliku keerdkonformatsiooniga CREBi
aktivatsioonidomään integreerub CBPga, siis toimub konformatsiooniline muutus ja moodustuvad 2 heeliksit,
mis keerduvad ümber CBP interaktsioonidomääni.
Ligand-
siduvad domäänid funktsioneerivad tuuma-
retseptoritel kui aktivatsioonidomäänid, kui nad on
ligandseoselises vormis.
Ligandi sidumine indutseerib konformatsioonilise muutuse, mis lubab ligand-siduval
domäänil ligand-seoseliselt anda interaktsioone ka tuuma-retseptorite ko-aktivaatori lühikese -
heeliksi domääniga, mille tulemusena moodustub kompleks, mis aktiveerib nende geenide promootoreid, mis sisaldavad
tuuma-retseptori sidumissaiti.
23. Mis on
sidumissait ? ja mille poolest ta a)sarnaneb, b)erineb TATAbox-st?
Sidumissait on ala DNA-l, millele transkriptsioonifaktor seondub. Mõlemad on transkriptsioonifaktorite
sidumisalad. TATA-box esineb eukarüootides, sidumissait on ka prokarüootides; TATAbox-ile seondub TBP
(mõni TAF ka; TFIID kompleks), konkreetne järjestus; sidumissaitidel on erinevad järjestused, mistõttu võib
neile seonduda ka erinevaid transkriptsioonifaktoreid.
24. Nimeta peamised DNAd siduvad domäänid, mis on iseloomulikud transkriptsioonifaktoritele (vähemalt 3) ·
Tsink -sõrm-valgud. Paljudel eukarüootsetel valkudel on struktuursed alad, mis voltuvad ümber Zn2+ iooni, nii et suhteliselt lühikesest polüpeptiidahelast tekib kompaktne domään. Seda struktuuri nim. tsink-sõrmeks.
Esmalt kirjeldati seda kui DNAd-siduvat struktuuri, tänaseks on teada, et tsink-sõrmi võib esineda ka valkudel, mis otseselt DNAd ei seo.
C2H2 tsink-sõrm on kõige levinum DNAd-siduv struktuur, mida inimese ja teiste hulkraksete organismide genoomis esineb suurte kordustena. See on omane ka hulkraksetele taimedele. See
motiiv koosneb 23 kuni 26 AAjäägist, sisaldades kahte konserveerunud Cys ja kahte His, millede kõrvalahelad seovad Zn2+ iooni. Tsink-sõrme nimetus tuleb sellest, et struktuuri 2D kujutis meenutab sõrme. Paljud 6 transkriptsioonifaktorid sisaldavad palju tsink-sõrm motiive. Teist tüüpi tsink-sõrm motiivid on sellised, kus neli Cys koordineerivad Zn2+ ja neid nimetatatkse C4 tsinksõrmedeks. Selliseid motiive on tänaseks kirjeldatud ca 50l inimese transkriptsiooni faktoril. Esimesed valgud, millel see motiiv leiti, olid steroidhormooni
retseptorid , kust tuli ka nimetus
steroid -retseptorite superperekond. Kuna hiljem kirjeldati hulgaliselt sarnaseid rakusiseseid retseptoreid, aga mis ei seo steroide, siis seda perekonda hakati nimetama tuuma-reptseptorite superperekond. Need valgud sisaldavad 55 v6i 56 AAlist domääni, kus on 4 konserveerunud asetusega Cys. Peamine erinevus nende kahe tsink- sõrm valgu klassi vahel on see, et kui C2H2 valgud seovad DNAd monomeerina ning sisaldavad 2 ja enamat tsinksõrm-motiivi, siis C4 valgud on reeglina sidumiseks homo- või heterodimeerid ning ühes valgus on mitte enam kui 2 tsink-sõrm motiivi. Homodimeersed tsinksõrmed seovad DNA pööratud kordusega elemente (inverted
repeats ). ·
Leutsiin -lukud (lZip). DNAd-siduvate domäänide hulgas on suur hulk struktuure, milledes iga seitsmes aminohappejääk on Leu. Need valgud seovad DNAd kui dimeerid ning Leu-de mutageneesi katsed on näidanud, et
viimased on vajalikud just dimerisatsiooniks. Siit ka nimetus Leu-lukud. Tänaseks on leitud ka transkriptsiooni faktoreid, kus 7ndas positsioonis on teised AAd. Nüüdseks kasutatakse
nimetust aluselised-lukud (basic-zipper). Paljud aluselisedlukud moodustavad heterodimeere. · Aluselised
heeliks -ling-heeliks (bHLH) domäänid. Need
struktuurid on väga sarnased aluselise-luku struktuuriga, erinevuseks on nende mittehelikaalne ling, mis eraldab kummagi monomeeri kahte - heeliksit. Nende valkude struktuuri nimetus tuletati aminohappejärjestuse põhjal, kust selgus, et N- terminaalne -heeliks sisaldab DNA sidumiseks vajalikke aluselisi AAjääke, keskmises osas on
lingu ala ning Cterminuses on järgmine -heeliks. Sarnaselt aluselistele-lukkudele võivad ka bHLH valgud moodustada heterodimeere.
25. Mis roll on histoondeatsetülaasidel (
HDAC ) ja mis funktsioon on histoonatsetülaasidel (HAT)?
Histooni sabad on tavaliselt positiivse laenguga tänu lüsiini ja arginiini jääkidele, mistõttu
histoonid saavad hästi
siduda negatiivse laenguga DNA-d. Atsetüleerimine neutraliseerib laengud ja DNA keerdub lahti, saab toimuda
transkriptsioon. HDAC deatsetüleerib histoonid, mistõttu seonduvad need tagasi DNA fosfaatide külge ja tekib
kondenseerunud struktuur. HAT-i roll on täpselt
vastupidine viib atsetüül-koensüümA-lt atsetüüli positiivsete
aminohapete neutraliseerimiseks.
26. Histoonne kood.
Histoonide teatud AAjääkide modifitseerimise kaudu kontrollitakse
kromatiini kondensatsiooni. Kromatiini
histoonseid
sabasid kontrollitakse lisaks veel pöörduva fosforüülimise (Ser, Thr), pöörduva
monoubikvitineerimise (Lys H2A C-terminuses) ning mittepöörduva metüleerimisega (Lys). Viimasel ajal on
üha rohkem andmeid, et "histoonne kood" kontrollib kromatiini kondensatsiooni, mitte üksnes histoonide
atsetüleerituse astet. "Histoonne kood" tähistab kõiki histoonide N-terminuses toimunud modifikatsioone. Näit,
heterokromatiinses vormis on H3 Lys positsioonis 9 sageli metüleeritud. "Histoonset koodi" lugevad valgud 7
tunnevad neid spetsiifilisi modifikatsioone ära ja promoteerivad kromatiini kondensatsiooni (suletud) või
dekondensatsiooni (avatud struktuuri teket). Imetajatel on kirjeldatud palju valke, milledel on n.ö.
kromodomään, see seondub H3 N-terminaalsele sabale, kui viimane on Lys9 metüleeritud vormis.Arvatakse, et
need valgud kontrollivad kromatiini kõrgemat järku struktuuriks pakkumist. Alternatiivselt on leitud
bromodomääni sisaldavad valgud, mis tihti on seotud eukromatiiniga ning seonduvad histoonide N-
atsetüleeritud sabadele. TFIID suurim alaühik sisaldab kaht järjestikku asuvat bromodomääni, mis aitavad
moodustada aktiivse koodiga kromatiinil kompleksi, samas kui sellesama alaühiku atsetülaasne aktiivsus hoiab
kromatiini hüperatsetüleeritud vormis.
27. Pärmi rakus toimub vaigistamine (silencing) telomeersetes DNA alades. Missugused
biokeemilised protsessid on selle telomeerse vaigistamise taga?
Pärmi telomeeride vaigistamine (silencing). Mitu
koopiat Rap1
seob iga telomeerialas paiknevale korduvjärjestusele (
telomeerid on nukelosoomidevabad piirkonnad) (Ülal). See initseerib
multivalkse kompleksi kokkupaneku (all), kus osalevad Rap1,
Sir2, Sir3, Sir4, ja lähedusesasuvate histoonide H3 ja H4
hüpoatsetüleeritud N-terminused. * tähistab hüperatsetüleeritud
histooni aminoterminusi. Moodustuv heterokromatiinne struktuur
ulatub Rap1-siduvate saitidega külgneva DNA ~4 kb
alale ,
olenemata DNA järjestusest. DNA on selles konformatsioonis
välistele valkude toimele (nukelaasid, transkriptsiooni
regulaatorid, etc) ligipääsmatu. Kõrgemat järku heterokromatiini struktuurist palju ei teata.
RAP1 seondub DNA järjestustele, mida nimetatakse vaigistajaks (silencer). SIR aga tunneb ära histoonide H3 ja
H4 N-terminusi. H3 ja H4 N-terminused on SIR2 aktiivsusest tingitult deatsetüleeritud vormis. Antikehadega
värvimine andis tulemuseks, et SIR valgud ja RAP1 paiknevad telomeeride piirkonnas. Nende andmete põhjal
pandi kokku
kromatiin -vahendatud vaigistamise mudel. Paljude RAP1 valkude sidumine nukleosoomivabadele
telomeeri otsele indutseerib heterokromatiinse struktuuri tekke telomeeride piirkonnas. Valk-valk
interaktsioonide võrgustik, kus on telomeeriga seotud RAP1, kolm SIR valku (2, 3, 4) ja hüpoatsetüleeritud H3
ning H4 ning mis kokku loovad väga stabiilse kõrgemat järku DNA-valk kompleksi. Viimane sisaldub mitmeid
telomeere, kus DNA on välisvalkudele peaaegu kättesaamatu. Üks lisavalk, SIR1, on samuti vajalik piirkonna/geeni spetsiifiliseks vaigistamiseks, samas ei ole sellest valgust palju teada. On teada, et ta on vaigistatud alaga seotud ning arvatakse, et ta on vajalik selleks, et formeerunud multivalkset telomeerset vaigistamiskompleksi edasi struktureerida, s.t. et see struktuur laieneks väljapoole telomeerset piirkonda. 28. Mis on
nukleosoom ? Kirjelda eksperimenti, mis võimaldaks teha kindlaks, et transkriptsiooniliselt aktiivne DNA on teisti pakitud, kui transkriptsiooniliselt inaktiivne DNA. =
nukleosoomne tuum(147 8
bp)+
linker DNA(15-55 bp). Nukleosoom on DNA-valk(liitvalgu histooni, mis koosneb 8 valgust - H2A, H2B,
H3 ja H4, kahest koopiast) kompleks, kromatiini primaarstruktuuri elemendid. Aktiivne DNA on DNaasidele
kättesaadav, inaktiivne mitte. Aktiivse ja inaktiivse DNaasitud segud
Southern blottitakse, ning inaktiivses on
näha globiini kohal "auk".
29. Loetle DNA erineva struktureerituse
astmed , mis lõpevad kromosoomi tekkega.
DNA-ahel: Nukleosiid trifosfaatides on fosfaadid seotud C5'-ga. Suhkru aatomite järgi nimetatakse 5' ja 3'
otsteks.
Nukleiinhapped sünteesitakse 5' otsast alates 3' suunas nii, et järgmise nukleotiidi vahele tekib
fosfodiester side. Polükondensatsioonireaktsioon.
DNA sekundaarstruktuur: Antiparalleelne kaksikheeliks, Komplemetaarne
paardumine ; erinevad vormid: B
vorm (tavaline, 1 pööre 10
aluspaari , olulisim omadus on võime painduda pikki telge kui DNA komplekseerub
valkudega (kromatiin)), A vorm (kompaktsem, esineb DNA-RNA interaktsioonide korral, 11 aluspaari), Z vorm
(vasakukäeline heeliks, annavad lühikesed ahelad ja alternatiivsed
nukleotiidid ), Kolmikahelaline struktuur
(Hoogstein paardumine on komplementaarne)
Bakteritel veelgi enam
keerdunud (superspiraalid), seda aitavad teha topoisomeraasid.
Telomeersed struktuurid: Lineaarsete kromosomide
otstes telomeerid, mille funktsiooniks on lubada
replitseerida ka kromosoomide 3' otsi (
telomeraas ). Samuti
kaitsevad sellised struktuurid kromosoomide otsi nukleaasse lagundamise eest. Telomeerid- üksikahelalised DNA 3'
otsad , mis koosnevad tuhandetest TTAGGG kordustest. (G rikkad alad); Quadrupleks struktuurid- 4 G
tasapinnalised interaktsioonid alustega vesiniksidemete vahendusel, mis on tasakaalustatud metall-
iooniga - G quadrupleks struktuur; T-lingud- telomeerides, stabiliseeritud telomeere siduvate valkudega; D-lingud (displacement loops) , kolmikahelalised struktuurid, mis moodustuvad telomeersete alade kombineerumisel kaksikahelalise DNAga. Kõrgem struktuur:
kromosoom , aga ainult raku jagunemise hetkel ja
seostunud histooniga. Lämmastikalused paarduvad omavahel vesinisidemete
kaudu. Paarid moodustuvad puriinide ja pürimidiinide vahel komplementaarsusprintsiibi alusel (A=T, G=C)
30. Kuidas mõjutab kromatiini struktuur transkriptsiooni?
Heterokromatiin- 10% genoomist, telomeerse ja tsentromeerse ala moodustamine
Eukromatiin- vähem kondenseerunud, põhistruktuuriks 30nm fiibrid ja lingud (loops)
Tasakaal eu ja heterokromatiini vahel- geenide vaigistamine. Kokkupakitud alale ei ole võimalik
transkriptsioonifaktoreid siduda, mistõttu sealt ei toimu transkriptsiooni.
31. Kirjelda DNA struktuuri mõju geenide aktivatsioonile/inhibitsioonile.
Eukarüootides oleneb suurte DNA osade ligipääsetavus kromatiini struktuurist, mis võib olla vahelduv histooni
modifikatsioonide (DNA metülatsioon, ncRNA või DNA siduv valk) tulemusena.
Keemiline struktuur geenide vaigistamiseks kasutatakse DNA metülatsiooni, DNAd metüleerivad
metüültransferaaside ensümiid tsütosiinide kohalt CpG dinukleotiidide järjestuses (CpG saared). Metülatsiooni
mustri analüüs kindlas DNA osas saab teostada läbi bisulfiidse kaardistamise meetodi. Metüleeritud tsütosiini
jäägid jäävad muutumata,
kusjuures metüleerimata tsütosiinid muutuvad uratsiilideks.
Ebanormaalsed metülatsioonimustrid on seotud kartsinogeneesiga.
Struktuur mida
tihedamalt pakitud, seda vähem toimub transkriptsioon. Nukleosoomid vastutavad
superspiraliseerumise eest ja sellised kompleksid võivad ajutiselt moodustada fosforülatsioonil või püsivamalt
metülatsioonil. Need
variatsioonid on vastutavad suuremate ja väiksemate
muudatuste eest geeni ekspressiooni
tasemetes. Histooni atsetüleerimine on ka oluline protsess. HAT (ja näiteks ka CREB-siduv valk)
dissotsieerivad DNA histoonikompleksi küljest, aktiveerides nii transkiptsiooni. DNA metülatsioon ja HDAC
aga põhjustavad geenidde vaigistamist. Nende kahe kombinatsioon tundub olevat
signaal DNA-le, et see pakiks
end rohkem kokku ja inhibeeriks geenide ekspressiooni. 9
32. Kirjelda kromatiini immunosadestamismeetodit.
Kromatiini immunosadestamise meetod paljastab
atsetüleeritud staadiumi kromatiini histoonides.
Histoonid on in
vivo kergelt risti DNA-sse lülitatud
kasutades rakku läbistavat, pööravat, keemiliselt
ristilülitavat ainet. Nukleosoomid atsetüleeritud
histoonide sabadega on näidatud rohelisega. 1.
Ristilülituv kromatiin isoleeritakse ja lõigatakse
keskmise pikkusega kaks kuni kolm nukleosoomi.
2. Lisatakse
antikeha , mis on konkreetse
atsetüleeritud histooni saba järjestuse vastu ja (3.)
seondunud nukleosoomid immunosadestatakse. 4.
Immunosadestatud kromatiini lõikude DNA
vabastatakse pöörates ristlülitust ja siis
paljundatakse kasutades tundlikku PCR meetodit.
Immunosadestamismeetodit kasutatakse, et uurida
in vivo seostumist ükskõik, millisel
valgul kindla
järjestusega DNA-le, kasutades uuritava valgu
antikehasid (2)
33. Mis on kromatiini metüleerimise tähtsus?
Kromatiini metüleerides pakitakse see kokku ning selle lõigu pealt ei toimu geenide ekspressiooni. Aitab kaasa
rakkude diferentseerumisele ja kindlustab, et rakus toodetakse vajalikke valke. Kui metüleerimist ei toimuks,
ekspresseeritaks tõenäoliselt valku kõikjalt ja
rakk ei oma enam funktsiooni.
34. Milleks kasutatakse
FISH analüüsi?
Fluorescent In Situ Hybridisation-it kasutatakse
satelliit -DNA märgistamiseks, mis omakorda aitab
kromosoome identifitseerida, ja kromosoomi alasid täpsemalt kirjeldada; satelliit-DNA paikneb enamjaolt
telomeerides, telomeersetes osades ja ka kromosoomi õlgade teatud piirkondades.
35. Mis on
MARide funktsioon?
Matrix Attachment
Region - kromosoom, pakituna nukleosoomideks ja keeratud 30nm kiuks, kinnitub MARide abil valgumaatriksile, mis omakorda aitavad kromosoomi pakkida tihedaks- metafaasi aegseks X-struktuuriks; kutsutakse ka SAR-ideks (scaffold-associated regions); Üldreeglina asuvad SARsid transkriptsiooniühikute vahel, s.t.
geenid paiknevad peamiselt kromatiinsetel lingudel.
Transgeensetel loomadel on näidatud, et SARsid on vajalikud teatud naabergeenide ekspressiooniks.
Äädikakärbsel on demonstreeritud, et mõned SARsid eraldavad transkriptsiooniühikud teineteisest, nii et ühe
geeni transkriptsioonis osalevad valgud ei mõjuta naabergeene, mis on eraldatud SAR-iga.
36. Kuidas tuvastada transkriptsiooniregulaatorite märklaud geene?
Muteerides uuritavat transkriptsiooni regulaatorit (selle leidmiseks võib kasutada nt DNase I footprintingut, mis
näitab valgu
asukohta (valk DNA-l, hakatakse lõikama lühikesi juppe, märgistatud 32P-ga) ja EMSA-ga saab
tuvastada (tekitatakse radioaktiivne DNA ja seotakse sellega valgulüsaat, pärast lahustatakse geelis kompleks
liigub aeglasemalt)) nii, et see enam ei funktsioneeri, tuleb kaardistada muutus geeniekspressioonis (DNA 10
microarray analüüsi abil). Geenid, mille ekspressioon on selle tagajärjel muutunud, ongi antud
transkriptsiooniregulaatori märklaud geenid.
37. Miks kasutatakse "
heat -
shock " geenide promootoreid?
Heat-shock geenide (näit hsp70) transkriptsioonil, peatub RNAPolII pärast esimese 25bp sünteesi, aga ei
termineeri transkriptsiooni. Peatunud polümeraas jääb vastsünteesitud transkripti ja DNA matriitsiga seotuks
kuni tingimuste avaldumiseni, mis indutseeruvad HSTF (heat-shock transkriptsiooni faktoreid). Aktiveeritud
HTSFi sidumine proksimaalsepromootori teatud järjestustele stimuleerib peatunud RNAPolII-i jätkama ahela
elongatsiooni ning indutseerib kiiret re-initsiatsiooni uute RNAPolII molekulide seondumisega.
Transkriptsiooni pausimisemehanism heat-shock geenide regulatsioonis kirjeldati esmalt puuviljakärbsel. heat-
shock geenide aktiivsus indutseeritakse raku-siseste tingimuste muutumisel, mille tagajärjel valgud hakkaksid
denatureeruma (nagu temperatuur tõus). Mõned neist geenidest kodeerivad valke, mis on denatureerivatele
tingimustele erakordselt vastupidavad, teised jälle chaperone, mis
denatureeritud valke uuesti re-struktureerivad
(re-
fold ). Nende geenide ekspressiooni regulatsiooni mehanism on evolutsiooniliselt kujunenud
selliseks , et
ekstremaalsetes tingimustes on transkriptsiooniks kõik olemas, st pole vaja kromatiini lahti pakkida ega
transkriptsiooni preinitsiatsiooni kompleksi kokku panna.
38. Mida tähendab väljend "kasutades lacZ reporterina" ja mis analüüsimeetodiga on tegu?
Transgeenne analüüs.
Reporter geen on geen, mida lisatakse teise huvipakkuva geeni uurimiseks geeni sisse (rakukultuuris,
loomadesse või taimedesse). Neid lisatakse sellepärast, et tunnused, mida nad end ekspresseerivates
loomades tekitavad, on kergelt äratuntavad ja mõõdetavad. Üldjuhul kasutatakse, et uurida, kas huvipakkuv geen
ekspresseerub.
LacZ on geen, mida ekspresseerides hakkab organism
tootma laktoosi.
Rekombinantse DNA meetod DNA jupid saadakse restriktaasidega, mis
lõikavad tömbid otsad. · Kasutatakse saamaks teada teatud geenide funktsiooni. Võetakse uuritav geen ja hakatakse mõnes rakus (nt
bakter ) sellelt geenilt valku tootma ning vaadatakse, kuidas rakk reageerib · DNA kättesaamiseks lõhutakse rakk katki, lisatakse etanooli ja selle tulemusel sadenevad nukleiinhapped põhja; kromosoomi ja plasmiidi eraldamiseks töödeldakse sooladega, mille tagajärjel
plasmiid lahustub ja etanooliga saadakse pärast kätte. · Pöördtranskriptaasi käigus saab küpsest mRNA-st uue DNA, mida kutsutakse
cDNA -ks (complementary), nii saab bakter sealt valku tootma hakata (
bakterites puuduvad
intronid )
Transgeenne analüüs organismi viiakse sisse võõrgeen, mida seal hakatakse ekspresseerima, näiteks
viirusvektorite abil saab; kui on vähe rakke, siis rekombinantse DNA meetod.
39. Defineeri
enhancer ja loetle enhanceri omadused. Missuguseid analüüsimeetodeid kasutatakse enhanceralade tuvastamiseks?
Kauguste-tagant mõjuvad regulaatorelemendid on enhancer-id, mis on eukarüootsetele genoomidele väga
iseloomulikud, samas kui bakteris nad peaagu täielikult puuduvad. Asuvad sadade kuni tuhandeteid aluspaaride
kaugusel start-saidist, võivad
asuda nii ülavoolu, intronites kui ka
allavoolu .
Enhancer'id on üldreeglina 50bp kuni 200bp pikad DNA järjestused
sisaldades hulgaliselt transkriptsioonifaktorite sidumissaite. Arvatakse, et
need TFid, mis ühele enhacer'ile seovad, interakteeruvad ka omavahel. -
interferooni (valk, mis inimesel on viirusinfektsiooni blokkijaks) ca 70bp
enhancer'i
uurimine on näidanud, et see koosneb 4 regulaatorelemendist,
mis seovad samaaegselt nelja TFi. Väikeste HMG (kromatiini siduvad,
rakus väga kõrgelt ekspresseeritud valgud) valkude juuresolekul on see
TFite sidumine enhancer'ile tugevalt kooperatiivne protsess.
Kooperatiivse sidumise tulemusena moodustub -interferooni enhancer'il multivalk-kompleks. Termin
enhancesome (võimendi) tähistab suurt tuumset valk-kompleksi, mis assambleerub suurest hulgast
transkriptsiooni faktoritest nende kooperatiivse sidumise tulemusena enhancer'il asuvatele
regulaatorelementidele. 11
Heterodimeersed Jun/ATF-2, IRF-3, IRF-7, and NF-B (p50 ja
p65 heterodimeer) seovad ~70-bp enhancer's neljale
elemendile. TFite kooperatiivset sidumist võimendab HMGI,
mis on sidunud DNA väiksesse vakku. cJun, ATF-2, p50, and
p65 seovad kõik ka nende
naabruses istuvat HMGd. Enhancer'i
paindumine (bending), mis toimub HMGIga interaktsiooni
tulemusena, on enhancesome' moodustamise
eelduseks .
Erinevad DNAd-painutavad (bending) valgud käituvad teistel
enhancer'
itel sarnaselt MGI seondub DNA väiksesse vakku
sõltumata selle nukleotiidsest järjestusest ning selle tulemusena
painutab (bending) DNA molekuli. See DNA enhancer' ala
paindumine lubab seondunud TFidel omavahel optimaalselt
interakteeruda. Algselt nõrgad, mittekovalentsed valk-valk
interaktsioonid saavad kõrvalelementidele seondunud faktorite
poolt oluliselt tugevdatud, mis teeb ka TFide lokaalse
kontsentratsiooni hästi kõrgeks.
Valmistati
plasmiidid , mis koosnesid -globiini geenist koos ja
ilma 366-aluspaarise lõiguga SV40 DNAst. Need plasmiidid
transfekteeriti rakkude kultuuri ja iga tekkinud RNA
hübridiseeriti -globiini DNA prooviks (1, 2). Rakkude poolt
sünteesitud -globiini mRNA kogus transfekteeriti ühte
plasmiidi või teist töödeldi S1 nukleaasi kaitse meetodiga (3).
Restriktsiooni lõigu
proov , mis oli saadud -globiini cDNA
kloonist, oli komplementaarne 5' otsale -globiini mRNA-s.
5'ots proovis märgistati 32P (punane täpp). -globiini mRNA kaitstud proovi hübridiseeriti ~340-nukleotiidine
lõik, mida oli lagundatud S1 nukleaasiga, mis
lagundab üksikahelalise DNA, aga mitte RNA-DNA hübriidi.
Autoradiograafia elektroforeesitud S1-kaitstud lõikudest (4) näitas, et
rakud , mida oli transfekteeritud
plasmiid 1-ga (1. rida) tootsid palju rohkem -globiini mRNA-d kui need, mida oli transfekteeritud plasmiid 2-ga.
Tulemused näitaad, et SV40 DNA
fragment plasmiidis 1 koosneb elemendist, enhancerist, mis stimuleerib
-globiini mRNA sünteesi.
40. Nimeta vähemalt 3 üldist bioloogilist protsessi, kus enhancerite aktiivsusega kontrollitakse geeni ekspressiooni tasemeid?
Rasvlahustuvad steroidhormoonid (östrogeen,
testosteroon ) võivad difundeeruda läbi plasmamembraani ja
seonududa spetsiifilistele retseptoritele tsütoplasmas või tuumas. Hormooni sidumine muudab retseptori kuju
nii, et see seondub enhancerile, millega
retseptor muutub transkriptsioonid aktivaatoriks.
Paljud
enhancerid on rakutüüp spetsiifilised, näiteks geenid, mis toodavad antikehi, omavad teises intronis
enhancerit, mis soodustab kõikide promootorite transkriptsiooni, aga seda ainult B lümfotsüütides, mis tavaliselt
toodavad antikehi.
Enhanceri kaotamine ja muteerimine on näidanud, et need koosnevad mitmest elemendist, mis mõjutavad
koguaktiivsust.
HO geeni
aktivatsioon algab tema sidumisest SWI5 aktivaatorile ülavoolu enhanceril. Seondunud SWI5
interakteerub SWI/SNF kromatiini uuestiehitava kompleksiga ja GCN5-koosneva histooni atsetülaasi
kompleksiga. Kui kromatiin on dekondenseerunud ja hüperatsetüleeritud, saab seonduda SBF, mille
mediaatoriga seondumise tulemusena moodustub transkriptsiooni preinitsiatsiooni kompleks, milles on
polümeraas II ja generaalsed transkriptsioonifaktorid.
41. Kirjelda eukarüootse mRNA molekuliga toimuvad muutusi pärast transkriptsiooni lõppu ja translatsiooni algust.
3 peamist sündmust: 1) 5' capping 2) 3' lõikamine ja
polüadenüülimine 3) RNA splaisimine. pre-mRNA protsessitakse juba tuumas
transkriptsiooni ajal ning vaid funktsionaalne mRNA transporditakse tsütoplasmasse.
5' cap lisamine sünteesitud RNAdele toimub vahetult pärast transkriptsiooni initsiatsiooni 12
Kui RNAPolII on jõudnud sünteesida uuest
transkriptist 25-30 nukleotiidi, siis 7- metüülguanosiin
ja teised 5'cap komponendid on juba mRNAde 5' otsa
küljes. Seda algset staadiumi RNA protsessingus
katalüüsib dimeerne capping ensüüm, mis seostub
RNAPolII fosforüülitud CTDga. Kuna capping
ensüüm ei interakteeeru RNAPolI ja RNAPolIIIga, sest
neil pole CTDd, siis capping on omane vaid
mRNAdele. Capping
ensüümi üks alaühikutest eemaldab _-
fosfaadi sünteesitud RNA 5'otsast, s.o. RNAPolII kompleksist
väljaulatuvast alast. Selle alaühiku teise domeeni aktiivsus lisab transkripti 5' otsa jäänud difosfaatrühmale
GMP, nii et tekib unikaalne guanosiin 5'-5' trifosfaatne struktuur. Viimases
staadiumis kannavad erinevad
ensüümsed alaühikud üle metüülrühmad S-adenosüülmetioniinilt guaniini asendis N7 olevale lämmastikule ning
sünteesitud RNA 5'otsas paiknevate ribooside 2'oksügeenidele.
pre-mRNAde 3'lõikamine ja polüadenüülimine on omavahel tihedalt seotud protsessid
Eukarüoodis on kõigil mRNAdel, v.a. histoonide mRNAd, 3' polü(A) saba. Primaarse transkripti 3' lõikamine
eelneb polüadenüülimisele. Peaagu kõik mRNAd sisaldavad järjestust AUAAA, mis asub polü(A) sabast 10-
35bp ülavoolu. Kui see AUAAA ära muteerida (vaid AUUAAA puhul toimub ka), siis RNA transkriptide
polüadenüülimist peaagu ei esine. Muteerimisel need polü(A)-sabata transkriptid kogunevad (akumuleeruvad)
tuuma ning suunatakse lagundamisele. On leitud ka üks teine 3' lõikamiskohast allavoolu esinev signaal, mille
intaktsus on vajalik lõikamiseks ja polüadenüülimiseks. See signaal ei ole teatud kindel järjestus, vaid pigem
GU-rikas või U-rikas piirkond 3' lõikamiskohast allavoolu asuva 50bp pikkuse ala sees. Lõikamiseks ja
polüadenüülimiseks vajalike valkude identifitseerimine on viinud
järgmise mudeli väljatöötamiseni. 360kDa lõikamis-ja
polüadenüülimis-spetsiifiline faktor (CPSF), mis koosneb neljast
erinevast valgust, moodustab kõigepealt AAUAAA järjestusest
ülavoolu
seonduva ebastabiilse kompleksi. Seejärel seonduvad
sellele CPSF-RNA kompleksile vähemalt kolm valku: 1) 200 kDA
heterotrimeer, mida nim lõikamist-stimuleerivaks faktoriks
(CStF), mis interakteerub G/U rikka järjestusega; 2) 150kD
lõikamisfaktor I (CFI) ja halvasti kirjeldatud lõikamisfaktor II
(CFII); 3) polü(A) polümeraas (PAP), mis seondub kompleksile
vahetult enne lõikamist.
PAPi seondumine liidab omavahel 3'
lõikamise ja polüadenüülimisprotsessid, nii et lõigatud transkripti
vaba 3' saba koheselt polüadenüülitakse. Suure multivalkse
lõikamis/polüadenüülimiskompleksi moodustumine AU-rikka
polü(A) signaali ümbruses on väga sarnane
initsiatsioonikompleksi moodustumisele TATAboxil. Mõlemil
juhul moodustuvad kompleksid läbi valk-valk ja valk-NH
interkatsioonide järjestikuse jada. Polüadenüülimine toimub kahes
etapis . Esimese 12 A lisamine toimub väga
aeglaselt, sellele järgneb väga kiire 200 või enama A lisamine. Kiire faas eeldab mitme RRM motiivi omava
polü(A)ga-seonduva valgu sidumist. Seda valku nim. PABPII, et eristada seda polü(A)ga-seonduvast valgust,
mis asub tsütoplasmas. PABPII seondub lühikesele A sabale ning stimuleerib PAPi järgnevaid A jääke
polümeriseerima. PABPII on vajalik ka PAPile signaalimaks polümerisatsiooni terminatsiooni, kui polü(A) saba
pikkus on jõudnud 200-250 nukleiinhappejäägini.
Küpse, funktsionaalse mRNA tekkimine hõlmab veel protsessi, mida nim. splaisinguks, mille käigus intronid
lõigatakse välja ja eksonid liidetakse kokku.
mRNA-d ümbritsevad valgud ja ta toimetatakse rakutuumast välja.
Tsütoplasmas vahetuvad mRNAga seotud valgud (hnRNP-d tuuma tagasi). 13
Seejärel seondub ribosoomi väike subühik AUG järjestusele, liitub Met-tRNA, seejärel suur subühik ning
algab translatsioon
42. Nimeta keemilised reaktsioonid, mis on pre-mRNA splaisingu taga. Missugused on need 3 staadiumit, mida katalüüsib splaisosoom?
Splaisingut teostavad splaisosoomid, mis koosnevad snRNPdest ja pre-mRNAst. snRNPde (U1, U2, U4;
U5 ja U6) interakteeruvad pre-mRNAga ja omavahel hierarhilises järjekorras, moodustades splaisosoomi. See
suur nukleoproteiinne kompleks katalüüsib seejärel kahe transesterifikatsiooni
toimumist , mille tulemusena
eksonid splaisitakse ja intron lõigatakse lariaatstruktuurina välja. Kuigi ATP hüdrolüüsi transesterifikatsiooni-
reaktsioonide toimumiseks vaja ei ole, saadakse splaisosoomi struktuuri ümberkorraldamiseks vajalik energia
tsükli enda toimumisest. Splaisosoomi snRNP valgud on hnRNP valkudest erinevad. Kõrgematel eukarüootidel
U2 snRNPde assotsatsiooni premRNAga
vahendab valk U2AF, mis seondub 3'splaissaidi läheduses asuva
pürimidiin-rikka alaga. U2AF interakteerub tõenäoliselt teiste splasinguks vajalike valkudega domeeni kaudu,
mis sisaldab dipeptiidseid
seriin -arginiin (SR-motiiv) kordusi.
Katalüütiliselt aktiivne ümberstruktureeritud splaisosoom viib läbi transesterifikatsiooni reakstiooni nii, et
moodustub branchpoint A 2'hüdroksüülrühma ja introni 5' otsa fosfaatrühma vaheline 2'5'-fosfodiesterside.
Teise struktuurse muutuse tulemusena, teise transesterifikatsiooni reaktsiooni käigus, ligeeritakse kaks eksonit
omavahel 3'5'fosfodiestersideme moodustumise tulemusena, nii et intron vabaneb harulise lariaat--
struktuurina. Eemaldatud intron lagundatakse kiiresti tuumas olemasolevate Rnaaside poolt. Splaisosoom on
peaagu sama suur struktuur kui ribosoomi väike alaühik, koosnedes umbes 70 valgust, mis kõik osalevad
splaisingus. Mõned splaisingfaktorid (SF) on assotseeritud snRNPdega, teised mitte.
Selleks et
eksport tuumast välja toimuks, peavad mRNAd olema lõplikult protsessitud. mRNAd, mis on
splaisosoomi
koosseisus , on reeglina ekspordi vastu "kaitstud".
43. Missugused U snRNAd osalevad pre-mRNA splaisingu regulatsioonis?
Splaisingus osalevad viis väikest U-
rikast tuuma RNAd (
snRNA ), mida
tähistatakse U1, U2, U4, U5 ja U6.
Nad on 107 kuni 210 bp pikad ning
assotseeritud tuumas 6 kuni 10
väikse tuuma ribonukleopartikli
valguga (snRNP). U1 snRNA 5'
otsa ja pre-mRNA 5'splaissaidi
paardumine on splaisingu
toimumiseks hädavajalik. U2
snRNAde roll splaisingus on
branchpoint'i kõrval asuva
konserveerunud järjestusele komple-
mentaarse pinna tekitamine.
Branchpoint ise ei ole U2 snRNAga
paardunud , vaid lingub välja, mis võimaldab tema 2' hüdroksüülrühmal
osaleda RNA splaisingu esimeses transesterifikatsiooni reaktsioonis. Splaisosoomi kokku
panemine algab
snRNPde kompleksis olevate U1 ja U2 snRNAde paardumisest pre-mRNAga.
Ulatuslik U4 ja U6 snRNAde omavaheline paardumine vastavates snRNP kompleksides tekitab multivalk-RNA
kompleksi, mis interakteerub U5 snRNPga. U4/U5/U6 kompleks assotseerub seejärel eelnevalt moodustunud
U1/U2/premRNA kompleksiga moodustades splaisosoomi. Pärast splaisosoomi moodustamist viib snRNPde ja
pre-mRNA ulatuslik paardumine U1 ja U4 snRNPde vabastamiseni.
44. Mis on GT-AG reegel?
Intron algab GU-ga ja lõppeb AG-ga, need on ainsad peaagu invariantsed nukleotiidid intronis.
45. Kirjelda
mehanismi , mille abil tuumsest pre-mRNAst kõrvaldatakse intronid.
Splaisingu vaheühendite analüüsi tulemusena on jõutud avastuseni, et eksonite splaising toimub kahe
järjestikuse transesterifikatsiooni reaktsiooni tulemusena. Intronid eemaldatakse lariaadi (lasso,
silmus , ling)-
sarnase struktuurina, kus introni 5'otsa guanosiin (G) on ebatavalise 2'-5'fosfodiestersideme abil introni 3' otsa
adenosiini (A) küljes. A nukleiinhappejääki nimetatakse hargnemispunktiks (branchpoint), sest ta moodustab 14
lariaadistruktuuris RNA haru. Kummagi transesterifikatsiooni käigus
vahetatakse üks fosfodiesterside
teisega välja. Kuna fosfodiestersidemete arv molekulis ei muutu, siis energiat selles reaktsioonis ei kulu. Kahe
reaktsiooni summaarseks tulemiks on kahe
eksoni ligatsioon ning nende vahelise-introni vabastamine lariaat-
struktuurina.
46. Kirjelda protsesse, mille tulemusena tekib ühest pre-mRNAst hulgaliselt erinevaid mRNAsid.
Alternatiivne splaissing - mRNA protsessingu peamiseks regulatsiooni mehanismiks, selleks on vaja vähemalt
kaht intronit (kolme eksonit). Selle käigus võib eksoneid pikendada või vahele jätta,
saades nii palju erinevaid
mRNA-sid.
47. Too näiteid, kuidas alternatiivne splaising muudab bioloogilist funktsiooni. Äädikakärbse sugu, näit.
48. Inimese BDNFi geen on enam kui 80,000 aluspaari pikkune. BDNFi mazhoorsed transkriptid aga 1.8 ja 4.4 kb pikad. Millest selline geeni
versus mRNAde pikkuse erinevus on tingitud? Kirjelda molekulaarseid protsesse, mis on selle pikkuste erinevuse taga.
Mittekodeerivad RNA alad on väga suured.
Inimese genoomis on eksoni keskmiseks
pikkuseks 150bp, samas kui introni keskmiseks pikkuseks on ca
3500bp.
Pikimad intronid on kuni 500kb pikad. Kuna 5' ja 3' splaissaitide ja branchpointi järjestused on
suhteliselt kõdunud, siis pikkades intronites esineb neid järjestusi mitmete koopiatena. Järelikult on vaja veel
mingit lisainformatsiooni, et eksoni piire ära tunda. Eksoni rekognitsiooni kompleksne mehanism hõlmab SR
valkude aktiivsust. SR valgud interakteeruvad eksoni järjestustega, mida nim. eksoonseteks splaisingu
võimendajateks (exonic splicing enhancer). SR valkudel on üks või enam RRMi ning mitu valk-valk
interkatsiooni pinda, mis on suhteliselt arginiini ja seriniini rikkad. Kui SR valgud on exonic splicing
enhancer'ga seotud, siis nad vahendavad U1snRNPde kooperatiivset sidumist korrektsele 5' splaissaidile ja U2
snRNPde õige branchpointi äratundmist. SR valkude, snRNPde ja teiste splaisingfaktorite (U2AF) kompleks,
mis moodustub eksoni peale, kannab nimetust cross-exon recognition
complex .
49. Mida näitab rakule antud ajahetkes iseloomulik mRNA tase?
Rakk toodab kindlat mRNA-d ainult siis, kui tal läheb vaja selle poolt kodeerivat valku, seeläbi minimeerides
raisatavat energiat. Kogu antud ajahetkel iseloomulik mRNA tase näitab, milliseid geene on antud hetkel 15
transkribeeritud. Seda saab teada, kasutades DNA microarray-d, mis koosneb paljudest geenide järjestustest,
komplementaarne järjestus seostub ja saab teada, milliseid geene ekspresseeritakse.
50. Mis on RNA editing? Mis ensüümid katalüüsivad protsessi RNA editing? Kuidas nimetatud regulatsioonivorm muudab valku? Too näiteid.
RNA järjestuse redigeerimine (editing) on pre-mRNAde järjestuste muutmise alternatiivseks mooduseks. RNA
järjestuse redigeerimine (editing) avastati 1980ndate keskel. Editing on protsess, mille tagajärjel muudetakse
pre-mRNA järjestust, st küpse mRNA järjestus erineb vastavast genoomsest järjestusest. RNA editing on väga
levinud ainuraksete ja taimede mitokondrite ning ka kloroplastide mRNAde puhul. Neis
organellides on mõnede
mRNAde puhul ligi pooled järjestustest redigeeritud. Kõrgemates eukarüootides on RNA editing suhteliselt
harv nähtus ning on kirjeldatud vaid mõne üksiku nukleotiidi vahetus. Ehkki ka viimasel juhul võib sellega
kaasneda suuri funktsionaalseid muutusi.
Osaliselt puhastatud ensüümi, mis viib läbi C6666 deaminatsiooni U-ks, uuringud näitavad, et see tunneb ära ja
sisestab 26 nukleotiidise RNA apoB primaarse transkripti C6666 ümbritseva ala järjestusse.
Imetajatel esineva RNA editing'i üheks näiteks on apoB mRNA, mis kodeerib kahte alternatiivset seerumivalku
apoB-100, mida ekspresseerivad maksarakud, ja apoB48, mida ekspresseerib sooleepiteel. apoB-48 vastab
apoB-100 N-terminaalsele osale. Mõlemad valgud on suure lipiidse valkkompleksi komponendid ning osalevad
veres lipiidide transpordis. Samas vaid need kompleksid, mis sisaldavad apoB-100 valke, osalevad kolesterooli
transpordis teistesse keharakkudesse. Kahe apoB tüübi rakutüüpspetsiifiline ekspression on apoB pre-mRNA
editing'i tagajärg. Nukleotiid positsioonis 6666, s.o C konverteeritakse deamiinimise tagajärjel U-ks. See
muutus, mis toimub vaid soole rakkudes, muudab CAA koodoni UAA poolt kodeeritud Stop koodoniks ning
lõpeb poole lühema valgu apoB-48 sünteesiga.
51. RNA-sõltuv vaigistamine.
siRNA ,
miRNA .
RISC kompleks.
MikroRNAd (
miRNAd ) avastati nematoodi C.
elegans geenide lin-4 ja let-7 mutatsioonide analüüsil, kus
selgus, et lin-4 ja let-7 ei
kodeeri valke, vaid RNAsid, mis on
vaid 21 ja 22 bp pikad ning seostuvad märklaud-mRNAde 3'
UTRidele. Selle interaktsiooni tulemusena nad represseerivad
märklaudgeenide ekspressiooni. Arengu käigus lin-4 miRNA
ekspressioon väheneb, let-7 miRNA ekspressioon on samuti
arenguliselt reguleeritud. Tänaseks on leitud nematoodil ca
100 miRNAd ja vähemalt samapalju on leitud miRNAsid ka
inimesel. Kõik miRNAd näivad olevat pikema ca 70bp
prekursor RNA
produktid . Prekursor RNA moodustab stem-
struktuure, kus tüve osas on vaid üksikud mitte-paardumised.
Dicer on valk, mis lõikab kaksikahelalise RNA neist
struktuuridest välja. miRNA ja märklaud- mRNA 3' otsa vaheline paardumine ei ole korrektselt komplementaarne, vaid on ka üksikuid mittepaardunud
nukleotiide . See mittepaardunud nukleotiidide olemasolu eristab miRNAid siRNAdest. RNA
interferents (RNAi) avastati katsetustel, kus manipuleeriti teatud geenide ekspressiooni tasemega. Süstiti nematoodi 16
rakkudesse lühikesi RNA juppe, mis pidanuksid hübridiseeruma märklaudgeeni mRNAga ning pärssima selle
translatsiooni. Kontrollkatse aga, kus kasutati perfektselt paarunud mõnesaja aluspaarilist kaksikahelalist RNAd,
oli oluliselt edukam selle märklaua translatsiooni inhibitsioonis, kui spetsiifiliselt disanitud antisense RNAd.
Sellest tehnoloogiast on saanud geeni funktsiooni uurimise üks suuri läbimurdeid. Kaksikahelaline RNA
protsessitakse esmalt väikesteks interferents RNAdeks (siRNA). siRNA ahelad, mis on 21-23 bp pikad,
hübridiseeruvad üksteisega nii, et 3'otsmised 2 nukleotiidi on üksikahelalised. siRNAde tekkeks on vaja Dicer
kompleksi ribonukleaasset aktiivsust. Sellest järeldati, et miRNAde ja siRNAde vahendatud protsessid on väga
sarnased. Hiljutised uuringud näitasid, et kaksikahelalisi siRNAsid ja miRNAsid protsessitakse edasi
multivalkkompleksis, mis sisaldab vaid üht RNA ahelat. See RNA-indutseeritud vaigistav kompleks (RISC)
lõikab märklaud mRNA, mis on täpselt komplementaarne vastavale üksikahelalisele siRNAle,
ahela katki. RISC kompleks funktsioneerib ka kui translatsiooni inhibiitorid. Inimese let-7 miRNA, näiteks, on
leitud täiesti let-7 miRNAga komplementaarset sünteetilist märklaud RNAd lagundava RISC kompleksi
koosseisust. RISC kompleksil arvatakse olevat 2 funktsiooni : 1) siRNA funktsioon (ehk RNA interferents), 2)
miRNA funktsioon, mille tagajärjeks on üksikuid mitte-paardunud nukleotiide sisaldavate märklaud mRNAde
translatsiooniline repressioon.
RNA interferents arvatakse olevat iidne raku mehanism taimedel ja loomadel, millega rakk kaitseb end teatud
viiruste ja mobiilsete geneetiliste elementide vastu. Taimed, milledes on tuvastatud Diceri või RISC kompleksi
valkude mutatsioonid, on oluliselt tundlikumad RNA viiruste infektsioonile ja transposoonide liikumisele nende
genoomis. Taimede kaksikahelaline RNA indutseerib ka sama järjestust sisaldava genoomse DNA
metüleerimist. Viimase mehanism ei ole teada. Selline RNAivahendatud geenimetüleerimise mehanism ei esine
imetajatel.
RNAi tsütoplasmas toimuv posttranskriptsioonilise
geeniekspressiooni regulatsiooni mehhanism, mille käigus
kaheahelaline RNA viib komplementaarse mRNA vaigistamiseni
siRNA small interfering/inhibitory RNA väike kaheahelaline RNA (21-25 bp), mis saadase dsRNA
lõikamisel Dicer ensüümiga ning mis viib komplementaarse mRNA degradatsioonini siRISC kompleksis
miRNA microRNA genoomi poolt kodeeritud väikeste (21-23 bp) RNA-de tüüp, mis saadakse stem-
struktuuriga prekursos RNA lõikamisel Dicer ensüümiga ning mis viib komplementaarse mRNA translatsiooni
inhibitsioonini miRISC kompleksis
RISC RNA induced silencing compleks kompleks (160kDa 80S), mis viib läbi siRNA-le või miRNA-le
komplementaarse mRNA vaigistamist degradatsiooni või translatsiooni ihibitsiooni teel. Põhikomponendid
agronaudid (AGO), millel on 2 olulist domääni: PAZ RNA-d siduv domään ja PIWI Rnaas H-ga sarnanev
endonukleaasset aktiivsust omav domään.
Dicer RNaasIII perekonda kuuluv ensüüm, mis viib läbi dsRNA või pre-miRNA lõikamist lühikesteks
siRNA-
deks /miRNA-deks.
Drosha tuumne RNaasIII perekonna ensüüm, mis viib läbi pri-miRNA lõikamise pre-miRNA-ks
Pasha tuumne dsRNA-d siduv valk, mis interakteerub Droshaga, stabiliseerides kompleksi.
Drosha ja Pasha moodustavad mikroprotsessori
RLC RISC loading complex RISC-ile eelnev dsRNA valk kompleks üleminekul kaheahelalise siRNA
lahtikeeramine
52.
Nonsense -mRNA vahendatud lagundamine.
Nonsense mediated
decay ühe või enam eksoni vahele jätmine, mis põhjustab
ekson -intron
liidese vahetus 3'
läheduses Stop koodoni sissetuleku. Kõigi korrektselt splaisitud mRNade puhul Stop
koodon on viimases
eksonis. Nonsense mediated decay vahendab kiiret mRNAde lgundamist, kus Stop koodonid esinevad mRNAs
enne viimast splaisiliidest. Arvatakse, et osad ekson-intron liidese kompleksi valgud osalevad nonsense
mediated decay's. Ekson-intron liidese kompleksid interakteeruvad vahetult deadenülaasi kompleksiga.
Arvatakse, et Stop koodoni "sisse splaisimise" korral ekson-intron liidese kompleksid RNAlt lahti ei tule ning
seega ribosoomset skaneerimist ei alustata. Nonsense mediated decay toimub tsütoplasmas. Hulgaliselt on
tõendeid selle kohta, et pioneeriv
ribosoom transleerib mRNAd, kui tema 5' otsaga on assotseerunud cap-siduv
kompleks ning polü(A) saba on assotseerunud
tuumse PABPIIga. See on tekitanud oletusi, et esimene
translatsiooni tsükkel toimub tuumas kui nonsense-mediated decay järelvalve mehanismi üks komponente. 17
53. Mis vahe on inteiinil ja intronil?
Inteiin valgu osa, mis suudab ennast välja lõigata ja taasühendada järelejäävaid osasid peptiidsidemega (valgu
intronid). Enamik inteiine sisaldab ka endonukleaasset domääni, mis osaleb inteiinide levimises. Paljudes
geenides on inteiini kodeerivad osad erinevates asendites.
Intron - on eukarüootse geeni
mittekodeeriv piirkond, mis transkribeeritakse RNA molekuliks, ent seejärel
lõigatakse sealt splaisingu tulemusena välja.
54. Mis juhtub, kui polüadenülatsiooni motiivis (AAUAAA) või RNAP II CTD
alas on mutatsioon?
RNA Pol II Carboks Term
Domain mutatsioon võib lüüa välja sealt Ser ja Tyr jääke, mistõttu seda enam ei saa
fosforüülida, ning kaob transkriptsioonivõime.
Peaagu kõik mRNAd sisaldavad järjestust AUAAA, mis asub polü(A) sabast 10-35bp ülavoolu. Kui see
AUAAA ära muteerida (vaid AUUAAA puhul toimub ka), siis RNA transkriptide polüadenüülimist peaagu ei
esine. Muteerimisel need polü(A)-sabata transkriptid kogunevad (akumuleeruvad) tuuma ning suunatakse
lagundamisele.
55. Selgita lühidalt mRNA polüadenüleerimise tähtsust eukarüoodi rakus.
Poly -A Polüadenülatsioon on poly(A) saba süntees, RNA lõik, kus kõik alused on adeniinid RNA molekuli
lõpus (ca 200 tk). See on osa teest, mille vältel eukarüoodid toodavad küpse mRNA translatsiooni jaoks.
Transkriptsiooni lõpus viimane osa just tehtud
RNAst lõigatakse ära valgukompleksi poolt. See kompleks
sünteesib poly(A) saba RNA 3'otsa. See saba on oluline tuumaekspordi jaoks, translatsiooni ja mRNA
stabiilsuse jaoks. Kui saba lühendatakse, siis varsti mRNA degradeeritakse ensümaatiliselt.
56. Seleta lühidalt attenuatsiooni mehanismi põhimõtet. Kasuta oma selgituses jooniseid.
Attenuatisioon leiti kõigepealt E.coli trp operonis.
Uuring oli suunatud kahele faktile mutatsioonid, mis
lõid välja trp repressori ei olnudki täielikult
trüptofaaniga represseeritud, trp operoni alusjärjestus
oli
ebatavaline ORF, mis eelnes tuntud trüptofaani
biosünteesivate ensüümide struktuursete geenide ORF-
idele. Esmalt leiti, et ORF koosneb kahest
tandem Trp
koodonist, teiseks, et mRNA-l koosnes selles alas kahe
grupilisest sümmeetriast, mis võimaldas moodustada
tal eksklusiivseid sekundaarstruktuure, millest üks
meenudas rho-sõltuvat transkriptsiooni terminatsiooni
signaali ja teine kaitseks esimese formeerumise eest ja
seeläbi ka terminatsiooni eest (pre-emptor struktuur)
Attenuatsiooni mehhanism: · RNAP initsieerib trp promootori transkriptsiooni · RNAP teeb pausi 90nda nukleotiidi juures sekundaarstruktuuris ·
Ribosoomid haaravad selle
vastse mRNA kaasa ja initsieerivad põhivalgu translatsioni o RNAP vabastatakse pausist ja see jätkab transkriptsiooni. · Kui RNAP jõuab potentsiaalse terminatsiooni alani, sõltub ala jätkumine või lõppemine järelelohiseva ribosoomi asendist. o Kui ribosoom jääb seisma tandem Trp
koodonite juures
oodates sobivat tRNA-d, ala 1 võetakse ribosoomi sisse ja seetõttu ei saa see paarduda regiooniga kaks, mis tähendab, et alad 2 ja 3 paarduvad enne, kui ala 4 transkribeeritakse. See sunnib 4. ala jääma üksikahelaliseks takistades ala ¾ terminaatorstruktuuri tekkimist. Transkriptsioon jätkub. o Kui ribosooom transleerib juhtivpeptiidi kahtlemata, katab see suurt osa 2. alast, takistades sidumist alaga 3. Kui 4. ala transkribeeritakse, see moodustab tõve ja lingu
alga 3 ja transkriptsioon lõpetatakse luues umbes 140 aluseline baas-transkript. · See kontrollmehhanism mõõdab saadava laetud Trp-tRNA hulka. Ribosoomide asukoht määrab struktuuri, ka teisi operone kontrollitakse attenuatsiooni teel. Avastus viis paljude selliste operonide taasavastamiseni, mille repressoreid polnud kunagi leitud. 18
57. Seleta lühidalt antiterminatsiooni mehanismi põhimõtet. Kasuta oma selgituses jooniseid.
Antiterminatsioon on prokarüootsete rakkude
abistamine parandamaks varajast RNA sünteesi terminatsiooni
transkriptsiooni käigus. See toimub, kui RNA polümeraas ignoreerib terminatsiooni signaali ja tekitab
mehhanismi, kuis üks või enam geeni operoni lõpus võivad olla sisse või välja lülitatud, sõltuvalt sellest, kas
polümeraas tundis ära terminatsiooni signaali või mitte. Kui transkriptsioon lõpetatakse varajaselt (sageli 100-
1000 aluspaari järel), antiterminaator seondub transkribeeritud RNA külge või ülavoolu DNA terminaatorist ja
alustab transkriptsiooni ,,uuesti".
58. Post-transkriptsioonilise regulatsioon tsütoplasmas. miRNA-sõltuv translatsiooni repression. RNA interferents-põhjustatud mRNAde
degradatsioon . Tsütoplasmaatiline polüadenüülimine.
Kaks hiljuti avastatud regulatsiooni teed:
1) mikroRNAd (miRNAd) avastati nematoodi C. elegans geenide lin-4 ja let-7 mutatsioonide analüüsil, kus
selgus, et lin-4 ja let-7 ei kodeeri valke, vaid RNAsid, mis on vaid 21 ja 22 bp pikad ning seostuvad märklaud-
mRNAde 3' UTRidele. Selle interaktsiooni tulemusena nad represseerivad märklaudgeenide ekspressiooni.
Arengu käigus lin-4 miRNA ekspressioon väheneb, let-7 miRNA ekspressioon on samuti arenguliselt
reguleeritud. Tänaseks on leitud nematoodil ca 100 miRNAd ja vähemalt samapalju on leitud miRNAsid ka
inimesel. Kõik miRNAd näivad olevat pikema ca 70bp prekursor RNA produktid. Prekursor RNA moodustab
stem-struktuure, kus tüve osas on vaid üksikud mitte-paardumised. Dicer on valk, mis lõikab kaksikahelalise
RNA neist struktuuridest välja. miRNA ja märklaud-mRNA 3' otsa vaheline paardumine ei ole korrektselt
komplementaarne, vaid on ka üksikuid mittepaardunud nukleotiide. See mittepaardunud nukleotiidide
olemasolu eristab miRNAid siRNAdest.
RNA interferents (RNAi) avastati katsetustel, kus manipuleeriti teatud geenide ekspressiooni tasemega. Süstiti
nematoodi rakkudesse lühikesi RNA juppe, mis pidanuksid hübridiseeruma märklaudgeeni mRNAga ning
pärssima selle translatsiooni. Kontrollkatse aga, kus kasutati perfektselt paarunud mõnesaja aluspaarilist
kaksikahelalist RNAd, oli oluliselt edukam selle märklaua translatsiooni inhibitsioonis, kui spetsiifiliselt
disanitud antisense RNAd. Sellest tehnoloogiast on saanud geeni funktsiooni uurimise üks suuri läbimurdeid.
Kaksikahelaline RNA protsessitakse esmalt väikesteks interferents RNAdeks (siRNA). siRNA ahelad, mis on
21-23 bp pikad, hübridiseeruvad üksteisega nii, et 3'otsmised 2 nukleotiidi on üksikahelalised. siRNAde
tekkeks on vaja Dicer kompleksi ribonukleaasset aktiivsust. Sellest järeldati, et miRNAde ja siRNAde
vahendatud protsessid on väga sarnased. Hiljutised uuringud näitasid, et kaksikahelalisi siRNAsid ja miRNAsid
protsessitakse edasi multivalkkompleksis, mis sisaldab vaid üht RNA ahelat. See RNA-indutseeritud vaigistav
kompleks (RISC) lõikab märklaud mRNA, mis on täpselt komplementaarne vastavale üksikahelalisele siRNAle,
ahela katki. RISC kompleks funktsioneerib ka kui translatsiooni inhibiitorid. Inimese let-7 miRNA, näiteks, on
leitud täiesti let-7 miRNAga komplementaarset sünteetilist märklaud RNAd lagundava RISC kompleksi
koosseisust. RISC kompleksil arvatakse olevat 2 funktsiooni : 1) siRNA funktsioon (ehk RNA interferents), 2)
miRNA funktsioon, mille tagajärjeks on üksikuid mitte-paardunud nukleotiide sisaldavate märklaud mRNAde 19
translatsiooniline repressioon.
RNA interferents arvatakse olevat iidne raku mehanism taimedel ja loomadel, millega rakk kaitseb end teatud
viiruste ja mobiilsete geneetiliste elementide vastu. Taimed, milledes on tuvastatud Diceri või RISC kompleksi
valkude mutatsioonid, on oluliselt tundlikumad RNA viiruste infektsioonile ja transposoonide liikumisele nende
genoomis. Taimede kaksikahelaline RNA indutseerib ka sama järjestust sisaldava genoomse DNA
metüleerimist. Viimase mehanism ei ole teada. Selline RNAivahendatud geenimetüleerimise mehanism ei esine
imetajatel.
2) tsütoplasmaatiline polüadenüülimine - Lisaks miRNA-vahendatud translatsiooni repressioonile esineb ka
valkvahendatud translatsiooni regulatsiooni, mis aitab teatud geenide ekspressiooni kontrollida. mRNAdes on
regulatoorsed elemendid või järjestused, mida tunnevad ära teatud valgud, mis osalevad translatsiooni
regulatsioonis. Üldreeglina asuvad need järjestused UTRides. Järjestus-spetsiifilised regulaatorvalgud võivad
kooperatiivselt siduda 3'UTRis asuvatele naabersaitidele, sarnaselt transkriptsiooni faktorite kooperatiivse
sidumisega enhanceritele. Enamusel juhtudest see sidumine represseerib translatsiooni. Valkudel, millel selline
repressioon esineb, on kirjeldatud tsütoplasmaatilist polüadenüülimist, mis peab sellisele repressiooni
mehhanismile eelnema. Tsütoplasmaatiline polüadenüülimine on varases embrüos üks olulisimaid
geeniregulatsiooni teid. Hulkraksete
munarakud sisaldab paljude valkude mRNAsid, mida enne ei transleerita,
kui alles pärast munaraku viljastamist. Mõnedel neist "ladustatud" mRNAdest on lühike polü(A) saba,
koosnedes vaid 20-40 A jäägist. See tähendab, et vaid üksikud polü(A)ga-seonduvad PABPI valgud savad
selliste sabadega seostuda ja neid stabiliseerida. Kuna lühikese polü(A) saba stabilisatsiooni ei toimu, siis neid
mRNAsid efektiivselt ei transleerita. Teatud munaraku arengustaadiumis või koheselt pärast viljastumist,
tavaliselt vastusena teatud välissignaalile, hakatakse nende mRNAde sabasid pikendama, lisades sinna kuni 150
A-d ja stimuleerides nende mRNAde translatsiooni. Sarnased mehanismid
toimivad ka õppimise ja mälu
protsessides.
mRNAsid lagundatakse tsütoplasmas teatud mehanismide abil:
mRNAde kontsentratsioon on funktsioon tema sünteesi kiirusest ja tema lagundamise kiirusest. mRNA
stabiilsus määrab ka ära, kui kiiresti temalt sünteesitud valgu süntees termineeritakse. Enamus bakteriaalseid
mRNAsid on väga ebastabiilsed (lagundatakse minutite jooksul), nii et nende poolt kodeeritud valkude süntees
kestab oluliselt kauem pärast vastavate geenide transkriptsiooni repressiooni. See võimaldab bakteri rakul kiirelt
kohaneda keskkonna
muutustega . Enamus hulkraksete organismide puhul aga on mRNAde eluiga mõõdetav
tundidega. Mõnede geenide regulatsioon toimub hüppeliselt, näit tsütokiinide (immuunvastusega seotud valgud)
puhul on see nii. Selline regulatsioon on võimalik tänu sellele, et nende geenide transkriptsiooni reguleeritakse
kiire vastustena vastavate mRNAde
eluea kaudu. Tsütoplasmaatiliste mRNAde lagundamine algab polü(A)
sabast, kuni see on nii lühike, et PABPi ei saa enam siduda ega stabiliseerida 5'cap-i ja initsiatsioonifaktoritega
moodustunud kompleksi. Eksponeeritud 5' cap struktuuri eemaldab decapping ensüüm ning kaitsmata mRNA
lagundatakse 5'3' eksonukleaaside poolt. Polü(A) saba kõrvaldamine muudab mRNA 3' otsa tundlikuks ka
eksosoomide 3'5 eksonukleaaside aktiivsuse suhtes. Deadenülatsioonikiirus on pöördvõrdeline sellelt mRNAlt
translatsiooni initsiatsiooni sagedusega: mida kõrgem initsiatsiooni sagedus, seda aeglasem deadenüülimise
kiirus. Mõnede mRNAde lagundamine ei sõltu deadenüülimisest. Mõnikord alustatakse cap struktuuri
eemaldamisest või siis mRNA sisesest lõikamisest, mida tõenäoliselt viib läbi RISC kompleksi eksonukleaas.
TRANSLATSIOON JA VALKUDE SORTEERIMINE
1. Valkude ja nukleiinhapete sünteesi sarnasused
Nii valgud kui nukleiinhapped on koostatud piiratud arvust monomeeridest · Monomeere lisatakse ahelasse ükshaaval · Iga polünukleotiid ja polüpeptiid sünteesitakse kindla orientatsiooniga ja see lõpeb kindlaksmääratud kohas · Primaarne
produkt modifitseeritakse
2. RNA funktsioonid valkude sünteesil
RNA-l on kolm funktsiooni valgusünteesis
Valgusünteesiks (translatsiooniks) vajatakse kolme komplementaarse funktsiooniga nukleiinhapet: 20 · mRNA kannab koodonitena informatsiooni valgu primaarjärjestuse kohta · tRNA identifitseerib koodi ja toob ribosoomi koodile vastava aminohappe · rRNA assotsieerub ribosoomi valkudega moodustades ensüümkompleksi, mis otseselt sünteesib peptiidsideme
3. Translatsiooniprotsess,
presibosomaalne ja ribosomaalne etapp
Translatsioon on kaheastmeline dekodeerimisprotsess 1. Preribosomaalne etapp: aminoatsüül tRNA süntees (ligaasid, ARS või ARL) 2. Ribosomaalne etapp: koodon-
antikoodon translatsioon ja peptiidsideme süntees ribosoomil
4. tRNA struktuur,
funktsionaalsed osad
mRNA kood transleeritakse valguks läbi adaptermolekulide-tRNA · Sekundaarstruktuur ristikheinalehe kujuline: · Pikkus 72-94 nukleotiidi, loetakse 5' otsast · Antikoodon- 34-36 · Kolm lingu +
paardumata 3'osa · 3' järjestus CCA 74-76, lisalingu nukleotiidid 47, 47:1 jne. · tRNA 3' paardumata osale järgneb aktseptoorne õlg, 7 aluspaari · Pseudouridiini õlg (T õlg) · Antikoodon õlg- 5 aluspaari,
lingus alati 7 nukleotiidi · D-õlg, 4 aluspaari, dihüdrouridiin, lingu pikkus varieeruv
tRNA kolmemõõtmeline struktuur on L kujuline, moodustub heeliksite
liitumise teel- coaxial stacking
Struktuurid sarnased, kuna liituvad ribosoomi
samasse saiti- erinevused antikoodoni ja aktseptorõla osas
(ka teised nukleotiidid võivad olla erinevad) 5. Mittestandartne paardumine (Wobble
base pairing), milleks vajalik?
Uridiin -
Guaniin ; Adenosiin-Inosiin; Tsütidiin-Inosiin; Uridiin-Inosiin Vajalik selleks, et erinevaid koodoneid on 64 (aminohapetele 61) ja siis pole nii palju erinevaid tRNA-sid vaja. Põhilised ensüümid, mis peavad tRNA molekuli identifitseerima on aminoatsüül-tRNA süntetaasid e. ligaasid (ARL või ARS või ka Ala- RS, Phe-RS).tRNA identsuse elemendid- tRNA struktuuri elemendid, mis määravad ära millise aminohappega tRNA aminoatsüleeritakse.
Igat aminohapet kodeerib 1-6 koodonit- mitu erinevat tRNA molekuli (isoaktseptoorset tRNA-d)- sünonüümsed koodonid. Geneetiline kood on degenerantne Igas rakus on 20 erinevat süntetaasi ja üks süntetaas peab ära tundma ja ühe ning sama spetsiifilise aminohappega aminoatsüleerima mitut erineva antikoodoniga tRNA molekuli. 21
Kõik süntetaasid tunnevad ära 4. nukleotiidi 3' otsast- isoaktseptooretel tRNA'del identsed nn.
diskriminaator-aluseks. Samuti on identsuse elemente ka antikoodon lingus, nende nukleotiidide osas, mis on
identsed. 6. Aminoatsüül-tRNA süntees, kirjelda protsessi ja ensüüme Aminoatsüül-tRNA süntetaas aktiveerib aminohapped ja liidab need tRNAle Iga aminohappe jaoks on oma, spetsiifiline aminoatsüül-tRNA süntetaas Need jagatakse kahte klassi (klass I ja II) kummassegi klassi kuulub 10 liiget. Jaotus põhineb ensüümide aminohappelisel järjestusel ja katalüütilise saidi ehitusel Aminohappe
liitmine on kaheastmeline: 1. Liituvad süntetaasiga AH ja ATP- moodustub aminoatsüül adenülaat 2. Estersideme süntees COOH ja
riboosi 2' või 3' C vahel Vigade sagedus 10-5, -spetsiifiline hüdrolüütiline aktiivtsenter · Aminoatsüleerimise suur täpsus tagatakse editeerimisaktiivsusega- hüdrolüütilist aktiivsust, mille abil välditakse vale aminohappega laetud tRNA vabanemist. · Ülekande-eelne editeerimine: kontrollitakse aminoatsüleerimisreakatsiooni esimest
etappi , st välditakse vale aminoatsüüladenülaadi moodustumist (vale aminoatsüül-adenülaat lagundatakse enne selle ülekannet tRNA külge. · Ülekande-järgne editeerimine: kontrollitakse aminoatsüüladenülaadi ülekandmist tRNA-le. (hüdrolüüsitakse esterside aminoatsüül-adenülaadi ja vale tRNA vahel).
7. tRNA identsuse elemendid, diskriminaatoralused
Kõik süntetaasid tunnevad ära 4. nukleotiidi 3' otsast- isoaktseptooretel tRNA'del identsed nn.
diskriminaatoralused. Samuti on identsuse elemente ka antikoodon lingus, nende nukleotiidide osas, mis on
identsed.
8. Ribosoomide struktuur, eukrüootne ja prokarüootne ribosoom
A, P ja E
sait ribosoomil
RIBOSOOMIDE FUNKTSIOON- viivad kõikides organismides läbi programmeeritud valgusünteesi
kasutades aminoatsüül-tRNA'd (aa-tRNA) substraadina. Ribosomaalne RNA (rRNA) moodustab bakteriaalsetes 22
ribosoomides 66% massist ja eukarüootsetes ribosoomides 60% massist. Ribosoomid moodustavad bakterites
20-40% kuivmassist, eukarüootides tunduvalt väiksema osa
Ribosoomi subühikute vahele jääb põhiline aktiivtsenter, mis moodustab tRNA'de sidumiskohad: · A-saiti seondub aminoatsüül-tRNA - paikneb nii väiksemal kui
suuremal subühikul. · P-saiti seondub peptidüül-tRNA. Peale kasvava peptiidahela kandmist ribosoomis peptidüül-tRNA'lt A- saidis asuvale aa-tRNA'le jääb P-saiti deatsüleeritud tRNA · E-sait on deatsüleeritud tRNA spetsiifiline. Peale seda kui P-saidis tekkis deatsüül- tRNA liiguvad tRNA'd koos mRNA'ga ribosoomis ühe koodoni võrra edasi -asub põhiliselt ribosoomi suuremal subühikul.
9. Ribosoomi tsükkel
Translatsioonil on kolm etappi: initsiatsioon,
elongatsioon ja terminatsioon. Ribosoom läbib selle käigus
valgusünteesi ribosoomi tsükli
Bakteriaalne mRNA on polütsistoorne. Valku kodeerivat järjestust nii mRNA'l kui DNA'l nimetatakse avatud
lugemisraamiks(ORF).
Avatud lugemisraam -nukleiinhappe järjestus, mis sisaldab järjestikuseid aminohappeid kodeerivaid
koodoneid ja mis algab initsiaator-koodoniga ning lõpeb stop-koodoniga
Initsiaator-koodonile eelneb ribosoomi sidumispiirkond RBS (ribosome
binding site) ehk
Shine -
Dalgarno järjestus. RBS (4-7 NA) on
komplementaarne ribosoomi väiksema subühiku RNA (16S rRNA) 3' otsaga.
Paikenb AUG koodonist 5-7 nukleotiidi
eespool 10. RBS (Shine Dalgarno järjestus vs Kozaki järjestus), translatsiooni initsiatsioon, eukarüoodi ja prokarüoodi võrdlus
Initsiatsioon:
Valgusünteesi initsiatsioonil osalevad lisaks ribosoomidele
initsiatsioonifaktorid, initsiaator-tRNA ja GTP.
Bakteriaalne initsiatsioon: · ·IF1 ja IF 3 koos ribosoomi väikese subühikuga ja fMet-tRNAi seondub mRNAle-RBSile (Shine-Dalgarno sequence) 70S initsiatsioonikompleks saavutatakse kui IF2 vahendatud GTP hüdrolüüsi energia arvelt viiakse kokku väike ja suur alaühik.
Shine-Dalgarno järjestus ( Shine-Dalgarno box), on ribosomaalne
seondumissait, mis paikneb 6-7 nukleotiidi ülevalpool start koodonist. See on
kuue aluseline konsensusjärjestus mis aitab ribosoomi väikesel subühikul
õigesti positsioneeruda, kusjuures se toimub üle anti-Shine-
Dalgarno järjestuse (CCUCCU) 16S rRNA-l
Eukarüootne initsiatsioon toimub mRNA 5' otsa juures: · eIF3 ja eIF6 hoiavad ribosoomi dissotsieerunult. eIF 2, GTP ja Met-tRNA moodustava aktiivse ternaarkompleksi, selle ühinemisel 40S subühikuga (+eIF1A ja eIF3 moodustub preinitsiatsioonikompleks. · eIF3- fosforüleerimise kaudu reguleeritakse initsiatsiooni · 40S subühiku interaktsiooni mRNA 5' cap osaga vahendab 23 eIF4, mRNA-d skaneeritakse kuni AUG koodonini · Kozaki järjestus- spetsiifilised järjestuse AUG ümber, mis
kergendavad äratundmist ACCAUGG ·
IRES -
internal ribosome
entry sites
Eukarüootne initsiatsioon algab reeglina mRNA 5' otsa juures, osadel juhtudel aga ka sisemistel saitidel
11. Elongatsioon ja terminatsioon
Elongatsiooni käigus liigub aminoatsüül-tRNA läbi kolme ribosoomaalse saidi
Valgusüntees termineeritakse mRNA stopkoodoni jõudmisel ribosoomi vastavate faktorite poolt (
release factors)
12. Transkriptsiooniline regulatsioon eukarüoodis ja prokarüoodis.
Ribosoomid on võimelised samaaegselt transleerima ühelt mRNA-lt ja moodustama uusi komplekse- kiirendab
valgusünteesi
efektiivsust - polüsoomid
Translatsiooni initsiatsiooni tasemel: negatiivne translatsiooniline kontroll · Shine Dalgarno järjestuse
blokaad valguliste regulaatoritega-repressorid · Shine Dalgarno järjestuse paardumine mRNA komplementaarse osaga
Nt. ribosoomaalsed valgud võimelised siduma 5' UTR juhul, kui neid ei inkorporeerita ribosoomidesse · Eukarüootidel- translatsioonilised repressorid
Akonitaas- blokeerib ferritiini (rauda deponeeriv valk) biosünteesi. Vastav järjestus 5' UTR 30bp.(iron
response element). · Initsiatsioonifaktorite
modifikatsioon : eIF2
eIF2B- guanosiinnukleotiidi vahetusfaktor, vabastab GDP eIF2,
eIF2 fosforülatsioon deaktiveerib eIF2-G0 · Eukarüootidel- AUG kontekstist sõltuv regulatsioon- lekkiv
skaneerimine (leaky scanning) · Rakutüübispetsiifiline eIF4F soodustab 5' otsale lähima AUG initsiatsiooni · uORF- ribosoomkompleks dissotsieerub enne ORF jõudmist- frosforuleeritud eIF2 · IRES- ei vaja
eIF4E (cap binding protein), eIF4G proteolüüsi tagajärjel ei teki kompleksi 5' capil, eIF4G lühendatud variant ei vaja cap-i · eIF4E defosforülatsioon- inhibeerib 5' sõltuva translatsiooni- M faasis, apoptoosivalgud aga IRES-idelt (apoptootilised eIF4G fragmendid suudavad IRES-del initsieerida translatsiooni)
13. Valkude topoloogia, signaaljärjestused
Valkude bioloogilise aktiivsuse avaldumine
toimub kindlates kohtades, selle
realiseerumiseks kasutatakse erinevaid
signaaljärjestusi:
2 tüüpi: - võivad olla järjestikused 24 -
moodustuda valgu eri osadest selle pakkimise tulemusena
20% juhuslikest järjestustest moodustavad erinevaid topoloogilisi signaale
Organell -spetsiifilised signaaljärjestused
14. Sekretoorne rada, eksotsütoos ja endotsütoos · Valkude süntees toimub karedapinnalisel ER-l, kust vastsünteesitud valgud suunatakse läbi Golgi kompleksi erinevatesse lokalisatsioonidesse rakus või sellest väljaspool- sekretoorne rada- lüsosomaalsed valgud, Golgi valgud, integraalsed, membraansed ja sekreteeritavad valgud · Ülejäänud valgud sünteesitakse vabadel, tsütosoolsetel ribosoomidel- jäävadki sinna, kui ei sisalda signaaljärjestusi (
mitokondriaalne , peroksüsomaalsed, tuumavalgud · SRP- Signaali äratundev kompleks (
signal recognition particle) interakteerub N terminaalse signaaliga · Valkude suunamine ER toimub sünteesi käigus- kotranslatsiooniline translokalisatsioon
Selle kaudu sorteeritakse ER, Golgi kompleksi, lüsosomaalsed ja
sekreteeritavad valgud
Cis-ternaalne progressioon - Golgi kompleksis viiakse läbi valkude
protsessimine: Erinevad glükosüülimisreaktsioonid valkude
liikumise vältel Cis-golgist trans-Golgisse
Konstitutiivne
sekretsioon , väikesed, vähem pakitud vesiiklid
Reguleeritud sekretsioon- tihedamini pakitud, suuremad vesiiklid-
vabanevad mingi signaali tulemusena
Endotsütoos- retseptorvahendatud vs. Pinotsütoos- vesiiklid
liiguvad lüsosoomi, ained rakku sisse
Eksotsütoos ained rakust välja
15. Kaetud
vesiiklite tüübid
Erinevat tüüpi vesiiklite tekke mehhanism sarnane · Initsieeritakse GTP-d siduvate valkude
viimisega tekkiva vesiikli juurde · Kattevalkude sidumine transmembraansete retseptorite tsütosoolsesse ossa · Osa
sellistest valkudest on ka retseptorid,mis sorteerivad valke (ligande), mida moodustuv vesiikel transpordib
Valkude transpordil läbi sekretoorse raja osalevad kolme põhitüüpi vesiiklid · COPII- vesiiklid transpordivad valke ER-st Golgisse 25 · COPII vesiiklite teket intsieerib Sar1 · GTP seondumine võimaldab kattevalkude soendumise tekkivale vesiiklile · COPI - vesiiklid viivad läbi retrograadset transporti Golgis · COPI kaetud vesiiklid transpordivad valke retrograadselt, vesiiklite teke initsieeritakse ARF valkude (GTP-d siduv valk) seondumisega retseptoritele · Transporditavad valgud interakteeruvad samuti kattevalkudega · Vesiikli moodustumine põhjustab ARF-l GTP hüdrolüüsi · Kuidas sorteeritakse transporditavad valgud? - Sorteerimissignaalid transmembraansetel retseptoritel · Klatriinkaetud vesiiklid transpordivad valke trans-Golgist ja membraanist endosoomidesse · Koosneb kolmest
raskest (Mw 180 kDa) ja kolmest kergest 35-40 kDa)
ahelast , fibroosne triskelion on 36 korduses, sisekiht sisaldab adaptervalgu komplekse (AP, 350 kDa) · AP valgud on transporditava retseptori spetsiifilised, see on tagatud erineva valgulise koostisega · AP kompleksi teke trasporditava valguga kutsub esile ARF seondumise · GTP hüdrolüüs muudab dünamiini konformatsiooni ja on vajalik klatriin-kaetud vesiiklite eraldumiseks membraanist · COP tüüpi vesiiklid ei vaja sellist GTPaasset energiat
· Ilmselt olemas ka spetsiifilised kattevalgud, mis moodustavad eksotsütootilise ja sekretoorseid vesiikleid
Konstitutiivne sekretsioon, väikesed, vähem pakitud vesiiklid
Reguleeritud sekretsioon- tihedamini pakitud, suuremad vesiiklid- vabanevad mingi signaali tulemusena
16. ER signaaljärjestus, kus paikneb ja millised seaduspärasused
Laetud N terminaalne signaal (Met + 2-5 laetud AHd)
-6-12 hüdrofoobset Ah-d sisaldav regioon
-5 polaarset Ah-t sisaldav N-terminus
-Preproteiin ja preproproteiin
Sellise struktuuriga järjestus määrab ära tema äratundmise SRPde poolt ja selline polüpeptiid suunatakse ERi.
Pärast seda lõigatakse signaal ära vastavate signaalpeptidaaaside poolt.
SRPga inteaktsioon on adaperiks ribosoomide ja ER vahel (1000x afiinsuse erinevus)
Erandjuhtudel võib selline signaal olla ka polüpeptiidi sees nt ovalbumiin (22-41)
Polüpeptiidid, millel selline signaal puudub suunatakse tsütosooli
17. Valkude suunamine ER-I, SRP ja translokoon,
energeetilised aspektid
Sekretoorsete valkude signaaljärjestus suunab need ER-i, seejärel lõigatakse selline järjestus kui funktsionaalselt
mittevajalik 26
Sekretoorsed valgud sisenevad ER luumenisse Signaali äratundmise partikli (SRP) struktuur
vahetult peale sünteesi algust- kotranslatoorselt
Signaaljärjestuse interaktsiooniks ER membraaniga on vaja kahte valku: SRP ja SRP retseptor
Polüpeptiid liigub ER luumenisse läbi translokoni
TRAM Transport Associated Membrane kui seal on mutatsioon, ei suunata ER-i GTP hüdrolüüsil saadav energia tagab valkude liikumiseks ER-i vajaliku energia 27
18. Signaaljärjestuse lõikamine ja valkude pakkimine ER valkudel
Karedapinnalisel ER-l sünteesitud integraalsete membraanivalkude topoloogia
Tüüp I - N terminaalne signaaljärjestus, N terminus eksoplasmaatiline
Tüüp II - signaaljärjestus puudub, N terminus tsütosoolis
Tüüp III - signaaljärjestus puudub, N terminus eksoplasmaatiline
Tüüp IV - mitu korda membraani läbivad. A klass- N terminus tsütosoolne, B klass- eksoplamaatiline
19. Transmembraansete valkude membraani suunamine signaalankurjärjestus ja "stop-transfer" ankurjärjestus.
Tüüp I ,,stop transfer" ankurjärjestus
Tüübid II ja III sisemine signaaljärjestus
Tüüp IV mitu sisemist signaali
Üks kord membraani läbivad valgud sisaldavad ühte sisemist topogeenset järjestust
Mitu korda membraani läbivad (multipass transmembrane) valgud sisaldavad vastava arvu topogeenseid
järjestusi 28
20. Kuidas viiakse membraani
intra - ja ekstratsellulaarse N terminusega valke?
Signaal-
ankur järjestused: seondub translokoniga, kuid süntees jätkub.
Stop-transfer-ankur järjestused: seondub translokoniga, aga inhibeerivad sünteesi (arestitakse valgu süntees)
Vt ka 20ndat!
21. Mitokondriaalsed signaaljärjestused, kuidas toimub valkude suunamine erinevatesse lokalisatsioonidesse mitokondris(
matriks ,
membraan , membraanidevaheline ruum)?
Mitokondriaalsed valgud sünteesitakse eelasvalkudena, mis sisaldavad signaaljärjestusi. Sellised järjestused
vastutavad nii suunamise kui organell-
spetsiifilisuse signaali (uptake-
targeting sequences)
Mitokondriaalsete valkude suunamine · Mitokondriaalse ja kloroplastide DNA poolt kodeeritud valgud sünteesitakse nendes organellides ja inkorporeeritakse otseselt nendesse · Suurem osa nende organellide valkudest sünteesitakse tsütosoolis ribosoomidel · Mitokondriaalsed valgud sisaldavad spetsiifilisi järjestusi, nn "uptake-targeting" järjestusi · Sellised järjestused on signaaliks vastavatele retseptorvalkudele, mis translokeerivad vastavad valgud läbi
organelli membraani · Mitokondriaalne signaal sisaldab 3-5 positiivselt laetud AH-t, mis on
vaheldumisi hüdroksüleeritud AH- ga (Ser või Thr) 29
22. Mitokondriaalne membraanitransport
Transporditavad valgud peavad
olema lahti pakitud.
Tsütosoolsed chaperonid
transpordivad mitokondriaalsed
valgud
kanal -seotud retseptoriteni.
Mitokondriaalsed valgud seotakse
kas mitokondriaalset importi
stimuleeriva faktori (MSF) või
Hsc70 valkudega- hoiab ära
agregatsiooni
Mitokondriaalsete valkude
membraanist läbiviimine kasutab
energiat · ATP energiat tsütosoolis · Prootonite gradienti (proton- motive
force ) sisemembraanist läbiviimisel · ATP energiat maatriksis
Erinevad mitokondriaalsed valgud kasutavad transprodiks submitokondriaalsetesse struktuuridesse erinevaid
radu . Selleks on neil erinevad
signaalid Membraanidevahelised valgud: · konservatiivne mehanism- viiakse maatriksisse ja seejärel seob sekundaarne signaal vastavat valku sisemembraanis (Cyt c1) · mittekonservatiivne mehanism- valk sisaldab nn "stop-transfer" järjestust (Cyt b2) · Cyt C liigub vabalt läbi välismembraani läbi P70
poori , kui lisatakse heem, ei saa enam välja
Membraanivalgud- sisaldavad ka hüdrofoobset ala, mis jääb membraani kinni- stop transfer signaal
23. Mitokondriaalsete valkude pakkimine.
Kui valgud sisenevad mitokondri maatriksisse,
seondub nende külge
maatriks Hsc70, mis on
tsütosoolse Hsc 70 tsaperoni analoog (
struktuurilt ja funktsioonilt). Maatriks Hsc70 on seotud
maatriksis mitokondri sisemembraani pinnale,
transpordikanalite lähedusse. See seondub
pakkimata valgu külge kohe siis, kui see
maatriksisse siseneb, kaitstes valku
liitmise või
sadestamise ja varajase pakkimise eest. See on eriti
oluline valkkomplekside alaühikute korral, kuna
ükskõik millise alaühiku korrektne pakkimine
nõuab kõikide teiste alaühikute olemasolu. Peale
seda, kui valk jõuab mitokondri maatriksisse,
proteaas eemaldab selle N-terminaalse maatriksi
märklauaga järjestus.
Mõned imporditud valgud võivad oma lõplikkusse aktiivsesse konformatsiooni pakkuda kõrvalise abita. Siiski
paljude maatriksi valkude puhul on vaja kasutada Hsc60 (suguluses bakteriaalse
GroEL -iga). Kui selles
tsaperonis on mutatsioon, siis pärmi puhul näidati, et
import ja lõikamine toimuvad normaalselt, aga see ei
suuda täita oma rolli multivalk kompleksis (pakkimises vead). 30
24. Peroksüsomaalne transport, erinevused ja sarnasused mitokondriaalsest transpordiga
C- and N-terminaalsed signaaljärjestused suunavad valke
peroksüsoomiaalsesse maatriksisse
Peroksüsomaalsed valgud pakitakse tsütosoolis
Signaaljärjestus katalaasil: SKL või sarnane signaal C terminuses
Transport nõuab ATP energiat kuid mitte elektrokeemilist gradienti
Katalaas seob tsütosoolset retseptorit PTS1R, see kompleks omakorda
seondub Pex14P retseptorile
N-terminaalne signaal tiolaasil: 26 aminohapet, seda tunneb ära
PTS2R, pärast internalisatsiooni see signaal lõigatakse ära
25. Millised valgud on võimelised tuuma
sisenema passiivse transpordiga?
Tuumapoorikompleks- koosneb rohkem kui 50 valgust
nukeloporiinidest- 3000-4000 poori
Madalmolekulaarsed lähevad läbi
Kuni 17 000Da suhteliselt kiire diffusioon
26. NLS, ja NES, tuumatranspordi mehhanism ja energeetilised aspektid.
Suuremad kui 60 000Da ei läbi- peab olema aktiivne transport-NLS
NLS- 4-8 AH järjestused, Lys Arg rikkad
NLS tunneb ära tuumaimpordi retseptori
NLS nuclear localisation signal , NES nuclear export signal. Energeetiline aspekt on see, et nad on
GTPaasse aktiivsusega. Ran-GEF interakteerub ja fosforüleerib Ran-GDP-d, tõrjub valgu küljest ära, aitab
valku vabastada. Ran-GAP interakteerub Ran-GDP-ga ehk siis defosforüleerib.
27. Ran valkude tsükkel, nende osa transpordil.
Ran'id seonduvad valkudega, millel on tuumalokalisatsiooni signaal ning aitavad nad tuuma transportida ning
tuumast välja transportida. 31
SIGNAALIÜLEKANNE
1. Rakkudevaheline kommunikatsioon, milliseid protsesse sisaldab?
Ekstratselulaarne signaaliülekanne e. transmissioon sisaldab signaali teket ja selle liikumist signaali vastuvõtva
rakuni
(1) signaalmolekuli sünteesimine signaliseerivas rakus
(2) signaalmolekuli vabanemine signaliseerivast rakust
(3) signaalmolekuli transport märklaudrakuni
Rakusisene signaaliülekanne e. transduktsioon on protsess mille käigus toimub signaali vastuvõtmine,
rakusisene edaskandumine ja primaarne vastus sellele signaalile (signaali
konversioon rakuliseks vastuseks)
Tinglikult võib jagada kolme etappi: 1. Signaali vastuvõtmine raku pinnal
spetsiifiliste retseptorite poolt; 2. Signaali edasikandumine tsütoplasmas, mille käigustoimuvad rea valkude aktiivsuse muutused ja/või madalamolekulaarsete
sekundaarsete vahendusmolekulide (second messengers) kontsentratsiooni muutused; muutus raku metabolismis, funktsioonis või arengus, mille on esile kutsunud signaalmolekuli ja retseptori komplekseerumine; 3. tuumasisene etapp, mis sisuliselt seisneb geeniekspresioonimuutmises transkriptsiooni faktorite aktivatsiooni või inaktivatsiooni kaudu
Signaali eemaldamine ja rakulise vastuse kadumine
Kõik kirjeldatud etapid pole alati vajalikud!
2. Signaalmolekulide
klassifikatsioon vastavalt toime ulatusele, endokriinsed, parakriinsed ja autokriinnsed signaalmolekulid
Oma
olemuselt võib signaalikandjaid jagada kolme rühma: Füüsikalised signaali
kandjad . 9 Valguskiirgus 9 Elektriline pinge ja membraanipotsentsiaal ja selle muutus ioonide kontsentratsiooni muutuste kaudu Keemilised signaali kandjad. 9
feromoonid . 9
Neurotransmitterid , klassikalised neurotransmitterid neuropeptiidid 9 steroidhormoonid ja muud rasvlahustuvad molekulid (östradiool, testosteroon, progesteroon,
kortisool , A ja D
vitamiinid ), 9 Gaasid (NO, CO) 9
Peptiid - ja valgulised hormoonid (nt.
insuliin ja kasvufaktorid). Bioloogilised signaalikandjad 9
Viirused ,
bakterid 9 Rakk-rakk interaktsioonid 1. Endokriinne- signaalmolekulid(hormoonid) toimivad oma sünteesikohast eemal. Tavaliselt imetajates kantakse hormoone edasi vereringe kaudu 2. Parakriinne- signaalmolekulid (neurotransmitterid) toimivad sünteesikoha vahetus läheduses 3. Autokiinne- signaalmolekul toimib samale rakule, kus sünteesiti, parakriinse
erijuht 4. Signalisatsioon valk-valk kontaktide kaudu (adesiooon, k.a. Jukstakriinne toime) ~parakriinne, aga veidi pikema toimega 32
3. Hormoonide klassifikatsioon
Klassifitseeritakse vastavalt nende lahustuvusele ja retseptori lokalisatsioonile: · Väikesed lipofiilsed molekulid- võimelised läbima plasmamembraane (
steroidid , türoksiin, retionoolhape) seonduvad tsütosoolsetele retseptoritele · Hüdrofiilsed molekulid, mis seonduvad rakupinna retseptoritele (peptiidhormoonid) · Lipofiilsed molekulid, mis seonduvad rakupinna retseptoritele (prostaglandiinid)
Rakupinna retseptorid võib jaotada nelja suurde klassi 33
4. Sekundaarsed vahendajad
Suurema osa hormoonide efektid vahendatakse efektormolekulideni sekundaarsete vahendajate (second
messenger) abil
Adaptervalgud, mis osalevad signaali transduktsioonil, on konserveerunud molekulid
GTPaasse aktiivsusega lülitid ja proteiinkinaasid
Erinevate hormoonide süntees, vabanemine depoodest ja degradatsioon toimub erineva kiirusega ja on täpselt
reguleeritud
Rakupinna retseptorite KD (dissotsiatsiooni konstant) väärtused näitavad ligandide ligikaudset kontsentratsiooni
mida väiksem on KD, seda suurem on tasakaal retseptor-ligand kompleksi suhtes. KD umbes 50% ligandist
on valgule seostunud.
Agonistid- molekulid, mis seonduvad retseptorile ja aktiveerivad signaaliülekande ka ligandid ise
Antagonistid- molekulid, mis seonduvad, kuid signaaliülekannet ei tekita, blokeerivad retseptori
G-valkude korral: Retseptorvahendatud Gs- valkude stimulatsioon viib adenülattsüklaasi aktivatsioonini ja sekundaarse
vahendaja cAMP sünteesini cAMP ei funktsioneeri RTKde vahendatud signaaliülekandes, nii
RTKd kui GPRCd võivad initsieerida teiste sekundaarsete vahendajate sünteesi cAMP ja teised sekundaarsed vahendajad aktiveerivad spetsiifilisi proteiinkinaase cAMP aktiveerib spetsiifiliselt cAMP sõltuvaid (cAMP-
dependent ) proteiinkinaase (cAPKs)
Fosfolipiidide ensümaatilisel modifitseerimisel moodustuvad erinevad sekundaarsed vahendajad:
diatsüülglütserooli (DAG) ja inositooltrifosfaadi (IP3) süntees 34
IP3 vahendab hormoonide poolt indutseeritud Ca2+
cGMP vahendab NO signaliseerimist
vabanemist ERist
5. G-valk seoselised retseptorid ja nende effektorid. Suurte G valkude klassifikatsioon · Suurem osa imetajate rakupinnaretseptoritest on G-valguga seotud retseptorid (GPRCd). Sellised retseptorid on funktsionaalselt kompleksis trimeersete GTPaasse aktiivsusega valkudega · Ligandi seondumine retseptoriga aktiveerib G valgu, see omakorda aktiveerib effektorensüümi, mis sünteesib sekundaarse vahendaja · Kõik GPCRd on väga sarnase struktuuriga, koosnedes 7 transmembraanses domääniga monomeersest valgust, kusjuures N- terminus on rakust väljas ja C-terminus raku sees · GPCRd vahendavad väga erinevaid signaale, nt. nägemine, lõhnad, (peptiid)hormoonid, neurotransmitterid jne.
GPRC stimuleerimine spetsiifilise ligandiga aktiveerib
adenülaaditsüklaasi aktivatsiooni (-andrenergilise retseptori
näide)
Trimeersed Gs- valgud seovad -adrenergilised retseptorid ja
adenülaattsüklaasi
Läkaköhatoksiin ja kooleratoksiin modifitseerivad pöördumatult G-valke
CREB valgud seovad cAMP signaali transkriptsiooniga 35
GPRC signaali kustutamine: · Ga eemaldumine kompleksilt kutsub esile retseptori afiinsuse languse · cAMP eluiga reguleeritud cAMP fosfodiesteraasse aktiivsusega- AMP
Pikemaajalisel stimulatsioonil toimub retseptorite desentsitiseerimine (positivne tagasiside): · Homoloogne desentsitiseerimne: retseptori tsütosoolse osa
forforüülimine spetsiifiliste kinaasidega (
BARK adrenergic receptor kinase) · Heteroloogne desentsitiseerimine: Retseptori tsütosoolse osa fosforüülimine PKA poolt, fosforüülib kõik sobivad retseptorid
Arrestiinid: - blokeerivad spetsiifiliselt fosforüleeritud retseptorid - takistavad G seondumist - vahendavad AP2 ja klatriini seondumist- coated pits
6. Retseptor türosiinkinaasid (RTK) ja Ras valgud · RTKd seovad lahustunud või membraaniseoselisi peptiidhormoone, mis toivad tavaliselt kasvufaktoritena · Ligandi seondumine kutsub esile retseptori türosiinkinaasse aktiivsuse, fosforülatsiooni tagajärjel retseptor aktiveerub ja see omakorda käivitab signaaliülekande rakus · RTKd on tavaliselt aktiveeritud olekus dimeersed, kusjuures alati ei ole tegemist homodimeerse interaktsiooniga · RTK
rajad on olulised rakkude prolifereerumise 36 regulatsioonis, rakkude vananemise signaalide vahendamisel ja metabolismi moduleerimisel · RTKd transmiteerivad signaali Ras-valkudeni. · Ras valgud on GTPaasse aktiivsusega lülitid,
Ligandi seondumine viib RTKde autofosforülatsioonile ja nende aktiveerumisele
Ras-valgu tsükkel: toimub üleminek GEF-valk on adaptermolekuliks, mis ühendab aktiveeritud RTK
aktiivse ja inaktiivse vormi vahel Ras- valkudega
7. MAP kinaaside rada aktiveeritud Ras indutseerib kinaasse signaalikaskaadi, mis omakorda kulmineerub MAP kinaasi aktivatsiooniga MAP
kinaas on
seriini /treoniini kinaas, mida iseloomustab võime translokeeruda tuuma ja fosforüleerida erinevaid valke (nt. transkriptsioonifaktorid)
Aktiveeritud Ras signaal liigub edasi läbi proteiinkinaaside Eukarüootsetes rakkudes on rida erinevaid
kaskaadi MAP kinaaside radasid
GPCRs ja RTKs aktiveerumine on keerulisem protsess kui lihtne järkjärguliste interaktsioonide jada GPCRs või RTKs aktivatsioon viib sageli paljude erinevate sekundaarsete messengeride tekkeni Sarnane
rakuline vastus võib tekkida erinevate signaalradade kaudu Erinevate signaalradade
omavahelised interaktsioonid lubavad äärmise täpsusega reguleerida rakulist aktiivsust Suur osa rakulistest vastustest hormoonidele, kasvufaktoritele ja neurotransmitteritele realiseeruvad effektides geeniekspressioonile Selline efekt saavutatakse läbi proteiinkinaaside, mis otseselt või kaudselt fosforüleerivad spetsiifilisi transkriptsioonifaktoreid Heteroloogne ja homoloogne desensitiseerimine, PKA, BARK ja arrestiinid 37
8. Retseptorite desentsitiseerimine ja retsükleerimine
Retseptorite tundlikkuse vähendamine ja taaskasutamine.
Tähtis mehhanism kindlate rakuliste
vastuste saamiseks on
retseptori tundlikkuse vähendamine kõrgete signaali
kontsentratsioonide või peale signaali pikendatud mõju. Raku
tundlikkust kindlale signaali molekulile saab alla reguleerida
molekuli retseptorite endotsütoosi teel, seeläbi vähendades nende
arvu raku pinnal või muutes nende aktiivsust nii, et retseptorid kas
ei saa ligandile seonduda või moodustavad retseptor-ligand
kompleksi, mis ei anna normaalset rakulist vastust. Sellist
retseptorite aktiivsuse moduleerimist viiakse läbi retseptorid
fosforüleerides, sidudes selle külge teisi valke või tehakse
mõlemat korraga.
Homoloogiline desnsitiseerimine täiendavad jäägid -
adrenergilise retseptori tsütosoolses domäänis fosforüleeritakse
retseptorspetsiifilise ensüümiga (-adrenergilise retseptori kinaas
BARK), aga ainult siis, kui
epinefriin või agonist seondub retseptorile. Kuna BARK fosofrüleerib ainult
aktiivseid -adrenergilisi retseptoreid, ongi see homoloogiline.
Heteroloogiline desnsitiseerimine risti reguleerimine; PKA aktiivsust võimendab kõrge cAMP-i tase, mida
toodab ükskõik milline hormoon, mis aktiveerib Gs, sellise hormooni (nt epinefriini) pikendatud mõju põhjustab
tundlikkuse vähenemist nii -adrenergilisel retseptoril kui ka Gs valguga seondunud retseptoritel, mis seovad
erinevaid ligande (nt glükagoon).
Kui endotsütoosi käigus viidi retseptor vesiikuliga rakku sisse, siis sama
vesiikul võib eksotsütoosi teel jälle
raku pinnale minna.
RAKUBIOLOOGIA.
1. Nimeta rakuorganellid ja kirjelda lühidalt nende olulisi funktsioone. mitokonder 2 membraani (sisemembraani sopistised kristad), oma DNA (väike, sageli muutuv, pärandub emaliini pidi) Seal toimuvad tsitraadi tsükkel, elektrontransport, oksüdeeriv fosforüülimine, rasvhapete ja püruvaadi oksüdatsioon ja aminohapete
katabolism (kogu
aeroobne energiavarustus). Raku hingamine, toitainete lagundamine (ensüümide ja hapniku abil), moodustub vesi, süsihappegaas, energia). ATP süntees. rakusein (
rakukest ) eraldab väliskeskkonnast endoplasmaatiline võrgustik Karedapinnaline sellel paiknevad ribosoomid, Valkude süntees, talletamine ja bioühendite suunamine ,,koju". Siledapinnaline süsivesikute ja lipiidide süntees (lihasrakkudes kaltsiumi säilitamine); osade mürgiste ühendite lagundamine. Siledapinnalisest ER-ist tekib karedapinnaline kui sellele moodustuvad ribosoomid. Raku kuju säilitamine.
vakuool - vee
reservuaar , kindlustab raku turgori, toitainete varu/jääkinete varu ja kus toimuvad lõhustumisprotsessid. plasmamembraan eraldab väliskeskkonnast Golgi kompleks Glükoproteiinide jm membraanikomponentide valmimine, uute valgumolekulide süntees. Materjal tuleb ER-ist, see sorteeritakse, muudetakse (
lihtvalk -> liitvalk), tihendatakse, pakendatakse membraanstruktuuridesse. Seal on membraanstruktuuride varu, taimedes toodab lima ja seedeensüüme,
loomarakkudes on spermide puhul seal lõhustava toimega ensüümid, mis aitavad läbi munaraku kesta tungida. tuum DNA replikatsioon, tRNA, mRNA ja tuumavalkude süntees kloroplast - 2 membraani, sisemembraan hästi liigendatud. Sisemust täidab valguline
vesilahus strooma; oma DNA (määrab teatud kloroplasti valke); neil on ka tülakoid, kus paiknevad fotosünteetiline valguse absorbeerimise süsteem, elektrontransportahel ja ATP süntetaasi kompleks. Fotosüntees (
valgusstaadium sisemembraanistikul, kus asub ka klorofüll,
pimedusstaadium ensüümide vahendusel
stroomas ATP energia abil). 38 lüsosoom Sisaldavad lõhustava toimega ensüüme ja lagundavat materjali. Hüdrolaaside eraldamine, hüdrolüütiline lõhustamine. Autofaagia
iseenese seedimine (lagundatakse rasv- ja lihaskudet). Heterofaagia kehavõõrate ühendite lagundamine (ainuraksete toitumine). · tsütoskelett Tsütoskelett on eukarüootsetel rakkudel, see koosneb tuubulitest (
mikro ) ja filamentidest (vahe, mikro), valgud:
aktiin , tubuliin, müosiin. Funktsioonid:
struktuurne raku kuju formeerimine, säilitamine, muutmine; sideme loomine rakuorganellide vahel; transport. Tagab raku ja tsütoplasma liikumisvõime. · peroksüsoom aminohapete oksüdeerimine, Peroksisoomid sisaldavad ensüüme, mis kasutavad orgaaniliste molekulide oksüdeerimiseks molekulaarset hapnikku. Osa neist ensüümidest toodavad keemiliste reaktsioonidega vesinikperoksiidi, mis on väga tugev oksüdeerija, teised jälle lagundavad seda. · ribosoom Koosnevad kahest rRNA molekulist ja 34 valgumolekulist (rRNA-d rohkem kui valku). Valgumolekulid asuvad pinnal. Jagunevad suureks (50S/60S) ja väikseks (30S/40S) subühikuks. Ribosoomid (70S/80S) moodustavad tuumakestes. Seal toimub valkude süntees (asuvad tsütoplasmas, karedapinnalisel ER-il, mitokondrites, kloroplastides) Kristalliseeriti esimest korda 1999/2000, elektronmikroskoobiga nähti, et valkude sünteesi katalüüsivad rRNA-d
2. Missugused
organellid on omased nii bakteritele kui eukarüootidele? Missugused vaid bakteritele? Eukarüootidele?
Bakteritel pole organelle, struktuurilt on mõlemal olemas tsütoplasma, ribosoomid, plasmamembraan ja
kromosoom(id). Bakteritel on olemas veel
vibur (mõnel),
piil , võrreldes
loomarakuga ka rakukest.
Eukarüootidel on palju erinevaid organeid.
3. Nimeta erinevaid raku transpordi tüüpe ja kirjelda, kuidas nad erinevad, on sarnased ning missugused protsessid nendega kaasuvad ja miks.
Passiivne transport
Difusioon Saab toimuda, kui kontsentratsiooni gradient on erinev, osakesed liiguvad tihedamast keskkonnast hõredamasse. Saavad: hüdrofoobsed ja mittepolaarsed molekulid (O2, CO2, N2,
benseen ), väiksed laenguta polaarsed molekulid (vesi,
uurea , glütserool) o Osaliselt läbilaskev membraan o Osmoos o Kaasaaidatud osmoos
Aktiivne transport Membraanipumbad - Transmembraansed valgud, mis viivad erinevaid ioone vastu nende kontsentratsioonigradiente; ATP-
aasid - kasutavad
transpordiks ATP energiat; Ei moodusta poore vaid ioonide liikumine läbi konfomatsiooniliste muutuste; 4 klassi; Vastutavad membraanipotentsiaali tekke eest koos ioonkanalitega.
Klassid : P loomarakkudest, vastu gradienti (2x, 2x); F keerulisem, asub mitokondris, V keerulisem, levinud prokarüootides,
prootonid vastu gradienti, ABC perekond muud molekulid nt flipaase (2x transmembraanne, 2x ATP siduv) Endotsütoos o Fagotsütoos -suurte partiklite (mikroorganismid, surnud rakkude osad jne.) sissevõtmine (·makrofaagid ja neutrofiilid ·amööb) o Pinotsütoos (= endotsütoos) - lahustunud makromolekulide sissevõtmine väikeste (kanaleid . Niisugused ained võivad plasmamembraane
läbida vesiikulite 39
sisaldisena endo- või eksotsütoosi teel.
·
Ekso - ja endotsütoos tähendab membraaniga ümbritsetud transportvesiikulite teket
ja nende ühinemist kas välismembraaniga (eksotsütoos) või endosoomi
membraaniga (endotsütoos). Ühinemine toimub kahes etapis:
1. bilipiidkihid lähenevad üksteisele
2. seejärel toimub ühinemine.
·Mõlemat etappi aitavad läbida teatud membraanvalgud, nn. fusogeensed valgud,
milliseid pole õnnestunud identifitseerida.
·Ekso- ja endotsütoosi puhul vastavad makromolekulid eraldatakse vesiikulisse
ning nad ei segune teiste tsütoplasmas olevate makromolekulidega.
·Iga vesiikul on määratud ühinema ainult kindla membraaniga.Tulemuseks on
makromolekulide suunatud liikumine raku sise- ja väliskeskkonna vahel.
4. Kuidas rakk reguleerib valgu funktsiooni? Too 3 näidet ja kirjelda neid põhjalikumalt.
a) Rakk hakkab valku tootma alles siis, kui selleks tekib vajadus. Näiteks kui E.Coli keskkonda ilmub
laktoos ,
hakkab ta tootma laktoosi lõhustavaid valke.
b) Rakk
jaotab erinevate ülesannetega valgud eri organellide vahel.
Niimoodi ei lähe substraadid ja produktid
segi, ning vastupidised protsessid saavad korraga toimuda raku eri osades.
c) Rakk saab kontrollida valkude aktiivsust erinevate lülititega:
1) Allosteeria ehk ligandi sidumisest põhjustatud tertsiaar-või kvarternaarstruktuuri muutus. Näiteks Proteiin
Kinaas A cAMP puudumisel keskkonnast ei toimi. cAMP-i
ilmumisel PKA tetrameer laguneb kaheks
monomeerseks katalüütiliseks ja üheks dimeerseks regulatoorseks alaühikuks. Lisanäitena
hemoglobiini ligand
on O2, mille esialgne vähene sidumine viib hemoglobiini konformatsiooni, kus ta saab rohkem siduda.
2) GTPaaside superperekond. Siia alla kuulub näiteks RAS. GTPaas on aktiivne, kui talle on seotud GTP,
inaktiivne, kui on seotud GDP.
3) Seriini, Treoniini, Türosiini fosforüleerimine mõjutab valgu aktiivsust.
Kinaasid fosforüleerivad, fosfataasid
defosforüleerivad. Need mõlemad muudavad valgu laengut ja seeläbi ka konformatsiooni, ning aktiivsust.
4) Proteolüütiline lõikamine in/aktiveerib pöördumatult. Pankreasest tulebad trüpsinogeen ja kümotrüpsinogeen.
Peensool
eritab Enterokinaasi, mis lõikab trüpsinogeeni trüpsiiniks, mis omakorda kümotrüpsinogeeni
kümotrüpsiiniks.
Hiline aktiveerumine on vajalik, et keharakke ei hävitataks.
fosforüleerimine - valkude pööratav fosforüleerimine on tähtis regulatoorne mehhanism, mis toimib nii
prokarüootsetes kui ka eukarüootsetes organismides. Ensüümid, mida kutsutakse kinaasideks (+P) ja
fosfataasideks (-P) on sellesse protsessi haaratud. Paljud ensüümid ja retseptorid lülitatakse "sisse" ja "välja"
fosforüleerimise ja defosforüleerimise kaudu. Pööratava fosforüleerimise tulemus on konformatsiooni muutus
paljude ensüümide ja retseptorite struktuurides, mis põjustab nende aktiveerumist või deaktiveerumist.
Fosforüleeritakse tavaliselt seriini, treoniini ja türosiini jääke eukarüootsetes valkudes. Lisaks toimub
fosforüleerimine põhilistes aminohappe jääkides histidiinis, arginiis või lüsiinis prokarüootsetes valkudes.
Fosfaadi molekuli lisamine polaarsele R rühmale aminohappes võib muuta hüdrofoobse osa polaarseks ja väga
hüdrofiilseks, mistõttu toimubki konformatsiooni muutus. Nt p53 tuumorsuppressor valk aktiveerides võib
viia apoptoosini.
metüleerimine - valgu metüleerimine toimub tavaliselt arginiini või lüsiini jäägi juures. Arginiini saab
metüleerida 1 või 2 korda või ühe metüülrühmaga mõlemasse lämmastikku, läbi viivad peptidüülarginiin
metüültransferaasid. Lüsiini saab metüleerida 1-3 korda lüsiini metüültransferaaside poolt. Valkude
metüleerimist on enim uuritud histoonides, kus seda viivad läbi
histoon metüültransferaasid (Met kohal?). 40
Metüleeritud histoonid võivad epigeneetiliselt represseerida või aktiveerida geeni ekspressiooni teatud
tingimustel.
ubikvitineerimine ubikvitiini
aktiveerimine (2-astmeline
reaktsioon E1 ubikvitiin-aktiveeriv ensüüm viib
läbi, vajab ATP energiat; 1 samm hõlmab ubikvitiin-adenülaadi intermediaadi tootmist ja teine samm viib
ubikvitiini E1 aktiivtsentri tsüsteiini jääki, vabastades AMP, tekib tioester side C-terminaalse ubikvitiini
karboksüülrühma ja E1 tsüsteiini sulfhüdrüülrühma vahel).
Ubikvitiini
viimine E1-st ubikvitiini konjugeeriva ensüümi E2 aktiivtsentrisse käi trans(tio)esterifikatsiooni
reaktsiooni kaudu.
Viimane samm ubikvitülatsiooni kaskaadis loob isopeptiidi silla sihtmärkvalgu lüsiini ja C-terminaalse
ubikvitiini glütsiini vahel. See vajab sadade E3 ubikvitiin-valk ligaaside aktiivsust. E3 ensüümid suudavad
seonduda nii E2-le kui ka substraadile.
5. Mis on
homeostaas ,
diferentseerumine ,
apoptoos ? Homeostaas kui organismi omadus luua sisekeskkonna püsiv tasakaal. Homeostaas saavutatakse koordineerides füsioloogiliste reaktsioonide kompleksi kudede vahel keemiliste või elektriliste signaalide abil. Selle kommunikatsiooni juures mängivad keskset osa hormoonid ja on seetõttu tähtsad homeostaasi säilitamisel Diferentseerumine protsess, mille jooksul rakk kujuneb spetsialiseeritud rakuks Apoptoos raku programmeeritud surm, ebavajalike rakkude eemaldamine
Apoptoosi kutsuvad esile füsioloogilised (surmaretseptorite aktiveerimine) ja patoloogilised stiimulid (mutatsioonid, nakkused). Apoptoosi teel surevad
tavalised ,
terved rakud, mida pole enam vaja või siis haiged ja
stressis rakud, mida on küll vaja, aga mis muutuvad kahjulikuks või mittefunktsionaalseks. Apoptoosil on kaks varianti (rohkem uuritud) sisemine (intrinsic - otsus surra tuleb raku seest, mitokondrid asjaga seotud) ja välimine (extrinsic -
rakusurm aktiveeritakse rakupinnal teiste rakkude poolt)
6. Mis on kaspaasi funktsioon ja mis juhtub pro-kaspaasi aktiveerumisel?
Kaspaas: tsüsteinüül
aspartaat -spetsiifilised proteaasid, mis aktiveeritakse ainult apoptootilistes rakkudes.
Aktiivtsentris on kriitiline tsüsteiini jääk konserveerunud pentapeptiidis (QACXG), lõikab substraati
aspartaadi järel peaaegu absoluutse spetsiifilisusega. Apoptoos on pöördumatu, kui kaspaase on aktiveeritud üle
kriitilise taseme. Kaspaaside blokeerimine peatab apoptoosi. Inimesel on 11 kaspaasi, osad osalevad põletikes, mõned
alustajad, mõned
teostajad .
Kaspaasi aktiveerimiseks tuleb esmalt pro-kaspaas katki lõigata esimeste aktiveerimiseks kogutakse need hästi
lähedale, edasi aktiveerivad üksteist. Aktiivne kaspaas koosneb kahest lõigatud prokaspaasist (2 suurt + 2
väikest alaühikut)
Kaspaasidel on 2 rada, surma retseptori rada
DISC (koosseisus surmadomään + adaptorid + pro-kaspaas-8) ja
mitokondriline rada apoptosoom (koosseisus Apaf1 (adaptervalk) + dATP + tsütokroom c + pro-kaspaas-9).
Mõlemad aktiveerivad kaspaas-3-e, mis hakkab teostama lagundamist.
7. Nimeta apoptoosiga kaasnevad kolm degradatsiooni protsessi.
Apoptoosiga kaasneb tsütoskeleti
purunemine , mille tõttu rakk omandab ümara kuju, membraani asümmeetria
ja lisandid kaovad, rakk tõmbub kokku, tuum laguneb, kromatiin kondenseerub ja kromosomaalne DNA
lõhutakse. 1. Rakk tõmbub kokku ja ümbrust on näha, sest kaspaaside poolt on lõhutud valguline tsütoskelett. 2. Tsütoplasma on tihe ja organellid kõvasti pakitud. 3. Kromatiin kondenseerub kompaktsetesse kogumitesse (püknoos) tuuma ümbrikusse 4. Tuuma ümbrik muutub katkendlikuks ja selles olev DNA tükeldatakse, tuum puruneb mitmeks eraldi
kromatiini kehaks või nukleosomaalseks üksuseks. 5. Membraanile tekivad kühmud 6. Rakk puruneb mitmeks vesiikliks (apoptootilised kehad), mida fagotsüüdid söövad.
8. Mida reguleerib "nonsense-mediated decay pathway"?
Nonsense-mediated decay ühe või enam eksoni vahele jätmine, mis põhjustab ekson-intron liidese vahetus 3'
läheduses Stop koodoni sissetuleku. Nonsense mediated decay vahendab kiiret mRNAde lgundamist, kus Stop
koodonid esinevad mRNAs enne viimast splaisiliidest. Arvatakse, et osad ekson-intron liidese kompleksi valgud
osalevad nonsense mediated decay's. Ekson-intron liidese kompleksid interakteeruvad vahetult deadenülaasi 41
kompleksiga. Arvatakse, et Stop koodoni "sisse splaisimise" korral ekson-intron liidese kompleksid RNAlt
lahti ei tule ning seega ribosoomset skaneerimist ei alustata. Nonsense mediated decay toimub tsütoplasmas.
Hulgaliselt on tõendeid selle kohta, et pioneeriv ribosoom transleerib mRNAd, kui tema 5' otsaga on
assotseerunud cap-siduv kompleks ning polü(A) saba on assotseerunud tuumse PABPIIga. See on tekitanud
oletusi, et esimene translatsiooni tsükkel toimub tuumas kui nonsense-mediated decay järelvalve
mehanismi üks komponente.
9. Mis on tagasisidestusmehanism? ja too mõni näide.
See on mehhanism, mille alusel rakk ,,otsustab", kas
ekspresseerida mingit geeni või mitte. Sõltub ümbritsevatest
tingimustest nt kui valku palju, siis ei sünteesita, kui vähe, siis
sünteesitakse.
10. Too näiteid positiivsest ja negatiivsest geeni ekspressiooni regulatsioonist.
Positiivseid:
Histoonide atsetüleerimine
Aktivaatori seondumine
Negatiivseid:
Histoonide deatsetüleerimine
Repressori seondumine
DNA metüleerimine
Up-
regulation -protsess mis toimub raku sees, mis on
signaalipoolt esile kutsutud, resulteerub ühe või rohkema
geeni suurenenud ekspressioonis, seega ka valgu tootmises.
Down-regulation- protsess, mille tulemusena geen on
represseeritud ja toodetakse vähem valku.
Up-regulation toimub näiteks kui rakul on mingit tüüpi
retseptorist puudus, sellisel juhul sünteesitakse rohkem retseptor-valku ja transporditakse raku membraani ja
seega raku homeostaas taastatakse.
down-regulation toimub näiteks, kui rakk on neurotransmitteri hormooni või ravimi poolt üle-stimuleeritud
pikka aega ning retseptorvalgu ekspressiooni vähendatakse et rakku kaitsta.
Nt. seda protsessi illustreerivad insuliini retseptor saidid tüüp2 diabeediga inimese rakkudes.Ülekaalulise
inimese veresuhkru kõrgendatud taseme tõttu peavad beeta lehed pankreases vabastama rohkem insuliini kui on
normaalne, et tagada piisavalt insuliini ja seeläbi viia
veri homeostaatilistele tasemetele tagasi. see pidev vere
insuliini tasemete tõus resulteerub katsega suurendada veresuhkurt mis põhjustab retseptor saitide ala-
regulatsiooni inimese rakus. See vähendab insuliini retseptorite arvu. Inimene muutub resistentsemaks seetõttu,
et hormooni tundlikkust vähendatakse.
Nt2. ala-regulatsioon samal juhtumil. Tüüp2 diabeediga inimene saab suurendada oma tundlikkust insuliinile
läbi korraliku dieedi ja regulaarse trenniga, seega kaotades kaalu; mõned võivad isegi naaseda oma diabeedi
eelsesse olekusse jälgides sellist reziimi.
11. Kuidas väliskeskkonnast tulnud signaal amplifitseeritakse rakus vastusena üles?
Väliskeskkonnast tulnud signaal käivitab rakus esialgse vastuse- geeni ekspressiooni. Ekspresseeritud geenide
hulgas aga võib olla ensüüme ja transkriptsioonifaktoreid, mis omakorda aktiveerivad teisi geene. Tekib
aktivatsiooni-
kaskaad , mille tagajärjel hakatakse mingit valku tootma (vastuseks väliskeskkonna signaalile).
12. Miks on vaja rakusisest kompartmentaliseeritust ja signaali ülekannet? Too näiteid, mis rakuga juhtub vastavate protsesside/struktuuride puudumisel või vigasuse korral.
Kompartmentaliseeritus on vajalik, et eri protsessid ei hakkaks üksteist segama. Et rakust välja eritamiseks
mõeldud ainet ei satuks raku sisse, ning ei looks seal metaboolset vastust. Ilma membraanideta ei saa luua
membraanipotentsiaali. 42
13. Aktiivsed filamendid ja
mikrotorukesed osalevad eukarüootsete rakkudele iseloomuliku kuju tekitamises ja hoidmises, ometi ei ole need permanentsed struktuurid. Põhjenda, miks dünaamiline tsütoskelett on raku normaalseks funktsioneerimiseks oluline. Oma argumentide tugevdamiseks, too näiteid (kokku 4 näidet)olukordadest, kus on oluline eri tüüpi filamentide stabiilsuse säilitamine ja vastupidi.
Tsütoskelett osaleb mitmetes erinevates rakulistes protsessides: diferentseerumine, signaalülekanne, muude
ainete transport, raku kinnitumine(
adhesioon ), raku liikumine, apoptoos. Häired tsütoskeleti dünaamikas võivad
viia raku liigse liikuvuseni(vähk), liiga madala liikuvuseni (inaktiivsed leukotsüüdid-makrofaagid), probleemid
raku jagunemisel, rakusiseste signaalide ja muude ainete ringluse häired. Eukarüoodi suhteliselt suures rakus
oleks keemiliste reaktsioonide kiirus limiteeritud substraatide difusiooni kiirusega.
14. Organelli membraani koostis on selle organelli ja raku kui terviku normaalseks funktsioneerimiseks elulise tähtsusega. Loetle organelli membraanide üldised omadused ja kirjelda, kuidas need omadused võimaldavad antud organellil täita oma funktsiooni. Sealhulgas too näiteid organelli membraani integraalsetest komponentidest, nende paiknemisest ja kuidas see (asukoht) on seotud antud organelli funktsiooniga.
Endosoom- endotsütoosi käigus rakumembraani sissepungumisest tekkinud organell. Membraanis
väliskeskkonna molekulide retseptorid ja teised rakumembraani osad. Funktsiooniks ainete toimetamine
lüsosoomi lagundamisele.
Lüsosoom- loomaraku organell (jääk)ainete lagundamiseks. Membraan väga tihe, ja omab H+ pumpasid, et
organelli sise pH saaks olla palju happelisem kui tsütosoolil.
Peroksüsoom- fosfolipiidne
kaksikkiht , milles on mõned valgulised retseptorid importimaks aineid sisse
lagundamisele.
Sile ER- fosfolipiidne kaksikkiht, milles on mõned valgulised ensüümid lipiidide tootmiseks ja glükoos-6-
fosfatass glükoneogeneesiks
Kare Er- membraanile kinnituvad ribosoomid, mis toodavad membraanseid valke. Et toodetav valk saaks läbi
membraani, on vaja membraanis ees olevaid abivalke.
Golgi kompleks-
Vakuool- Membraanis erinevad valgud ainete transpordiks, samuti H+ pumbad madala pH hoidmiseks.
Tuum- kaks lipiidset kaksikkihti, välimist jagab ta kareda ERiga, sisemine defineerib tuuma ennast. Mõlemal
kihil on erinev valguline koostis. Tuuma
poorid on vallgud, mis läbivad mõlemat kihti korraga, ühendades nii
tsütosooli otse
tuumaga . Sisemembraani siseküljel on lamiinide võrgustik, mis annab tuumale tema kuju, ja
ühtlasi seob ka DNA-d
Tuumake - membraan puudub
Mitokonder- kaks membraani. Välimine 50% lipiidne, 50% valguline. Välimine sisaldab Poriine- kanaleid, mis
lubavad ka suurtel(kaal kuni 10'000) molekulidel vabalt
liikuda läbi membraani. Sisemine membraan on palju
tihedam(ei lase isegi H+ läbi), ja sisaldab u. 20%
lipiide , 80% valke. Valgud seal on näiteks ATP-süntetaas, H+
pumbad, lipiidide ja
suhkrute lõplikud oksüdeerijad. . Osad vajalikud valgud on sünteeritud mitokondri enda
DNA-st.
Kloroplastid - kaks membraani. Sisemisele tülakodi-membraanile on kinnitunud kloroplastid. Osad vajalikud
valgud on sünteeritud kloroplasti enda DNA-st.
15. Nimeta lisaks fosfolipiididele veel vähemalt 3 membraani komponenti.
Glükolipiidid, steroidid (
kolesterool ) (Membraanide assümeetria realiseerub mikrodomäänide moodustumisega-
lipiidide
parved (lipid rafts)- suure kolesterooli , sfingolipiidide sisaldusega piirkonnad. Lipiidide parved
moodustavad jäike struktuure, millele kinnituvad valgud); membraanivalgud (25-75% membraani massist (hästi
palju mitokondrites, sest seal on ATP süsteem ja palju pumpasid, hästi vähe müeliinis, sest see on metaboolselt
väheaktiivne ning isoleerib närvirakku)).
16. Valgu struktuurne intaktsus on vajalik selle valgu normaalseks funktsioneerimiseks. Too 2 näidet, kuidas valgu struktuuri muutus põhjustab muutuse selle valgu rakulises funktsioonis.
Kui hemoglobiinile seondub -subühikule hapnik, toimub konformatsiooni muutus ja seejärel saab hapniku liita
ka -subühikule ning seejärel kannab seda kopsudest teistesse kudedesse. 43
Trüpsinogeen on tavaolukorras pankrease ensüümi trüpsiini prekursor, millel on leitud madal aktiivsus ja
mille taset mõõdetakse vereproovides. Kui enteropeptidaas seda lõikab, muutub see aktiivseks trüpsiiniks,
trüpsiin omakorda hakkab ise trüpsinogeene lõikama.
17. Neuronil on 4 morfoloogiliselt eristatavat piirkonda (dendriidid, rakukeha,
akson , sünaps). Missuguste protsesside abil tagatakse valkude õigeaegne olemasolu "õiges piirkonas" ja miks see on vajalik? Valdav osa valke sünteesitakse rakukehas, väike hulk ka dendriidides. Signaaliülekandega määratakse ära, millal millist valku toota ning kuhu see transportida. Selle tulemusena saabki õigesse piirkonda õigeaegselt õige valk. 18. Mis molekul on signaaliks, mis suunab valgu proteosoomsele lagundamisele? Ubikvitiin 19. Fagotsütoos? Pinotsütoos? Endotsütoos? Eksotsütoos? Endotsütoos aubete transport rakku sel teel, et rakumembraan koos ainega sopistub sisse ning moodustab seal membraaniga ümbritsetud vesiikuli Retseptorvahendatud endotsütoos- spetsiifiline · Toimub AP2/klatriin seoseliste vesiiklite kaudu · Alati viiakse transporditav molekul endosoomidesse · Initsieeritakse klatriin-kaetud aladel,kuhu on juba
viidud retseptorid Fagotsütoos tahke aine sisse Pinotsütoos vedelik sisse Eksotsütoos põhiline eukarüootse raku sekretsioonimehhanism. Molekulid pakitakse membraaniga ümbritsetud vesiikulitesse, mis sulavad ühte raku välismembraaniga, vabastades oma sisu väliskeskkonda. Kaks teed: Pidev eksotsütoos toimub kõigis eukarüootsetes rakkudes transportvesiikulid kannavad pidevalt uusi membraanikomponente Golgi kompleksist välismembraani. Eksotsütoosi teel toimub pidev plasmamembraani uuendamine. Pidevalt eksotsüteeritakse valke, mida antud rakk ise ei vaja, kuid mida on organismil kui tervikul tarvis.
Reguleeeritud eksotsütoosi puhul kogutakse vastavad ained sekretoorsetesse vesiikulitesse, mis ühinevad raku
välismembraaniga pärast keskkonnast tulevat kindlat signaali. Reguleeritud eksotsütoos esineb neis rakkudes,
mis on spetsialiseerunud oma
produkti kiirele ja vastavalt vajadusele sekreteerimisele. Sekretoorsetes
vesiikulites ei ole alati valgumolekulid, vaid võivad olla ka madalmolekulaarsed ühendid. Madalmolekulaarsed
(mittevalgulised) ühendid satuvad sekretoorsetesse vesiikulitesse tsütoplasmast, mitte Golgi kompleksist.
Madalmolekulaarsed ühendid seotakse vesiikulis kindlate kõrgmolekulaarsete ühenditega, et vältida osmootse
gradiendi teket. Signaaliks reguleeritud eksotsütoosile on kas mingi keemilise aine (hormoon, glükoos,
elektriline signaal ehk aktsiooni potentsiaal). Signaali äratundmise tagajärjel genereeritakse membraani
retseptorite poolt üks või mitu rakusisest signaali, millega kaasneb rakusisese Ca2+-ioonide kontsentratsiooni
ajutine tõus, see käivitabki reguleeritud eksotsütoosi.
20. Kirjelda tuuma lamiine. Mille poolest erinevad tuuma
lamiinid teistest intermediaarsetest filamentidest? Kuidas mõjutab fosforüleermine tuuma lamiinide assotsatsiooni?
Lamiinid moodustavad tuumaümbrise sisemisel pinnal
filamentse võrgustiku. Tsütoplasmaatilised filamendid
ulatuvad NPCde tsütoplasmaatilisse ossa.
Ioonid ,
väiksemad metaboliidid ja globulaarsed valgud
suurusega kuni 60kDa võivad vabalt läbi veega-
täidetud tuumapoorikompleksi kanali difundeeruda.
See kanal käitub nagu ta oleks 0.9 m lai. Tegelikult 44
suured valgud ja RNP kompleksid tuuma ja tuumast välja difundeeruda niisama lihtsalt ei saa. Neid aitavad
transportida läbi tuumapooride lahustuvad transportervalgud, mis seovad transporditavaid valke ja ühtlasi
interakteeruvad nukleoporiinidega.
Koosneb intermediaarsetest filamentidest ja nendega seostunud valkudest, annavad tuumale tugevuse
Kui neid fosforüleerida (tsükliin B/Cdk1-ga) tuuma lamiinid depolümeriseeruvad ja B-tüüpi lamiinid jäävad
seotuks tuuma ümbriku fragmentidega, A-tüüpi lamiinid on mitoosis lahustunud.
21. Mis on iseloomulik intermediaarsetele filamentidele?
Läbimõõt on 10-12 nm, koespetsiifilised, loomarakkudes nt
keratiin . Need annavad mehaanilise tugevuse.
Asetsevad enamast tsütoplasmas. Peamine struktuur on konserveerunud, igal valgul on globulaarsed
tuumad N-
ja C-terminuses, mille vahele jääb alfa-helikaalne
varras . Peamine ehitus on anti-paralleelne (pole laengut) ja
dimeerne. Dimeerid tulevad varda osa interaktsioonidest, mis moodustavad keritud
spiraali . Neil pole
sidumissaiti NTP-dele.
22. Kuidas mõjutab fosforüleerimine intermediaarsete filamentide struktuurseid omadusi?
Toimuvad sarnased asjad nagu tuuma lamiinidega fosforüleerides laguneb
osadeks , fosforit eemaldades saab
jälle kokku.
23. Millel põhineb mikrotorukeste stabiilse struktuuri säilimine? Treadmill efekti olemus.
Stabiilne struktuur koosneb tsentraalsest mikrotuubulite kimbust ripsmetes ja viburites, mis on
plasmamembraanist väljaulatuvad osad, mis töötavad rütmiliselt, et liikuda (spermi
ujumine , munaraku
liikumine). Nad moodustavad sisemise tuuma näiteks ka närvirakkudes.
Mikrotuubulid on polaarsed liitumine
toimub ka alati + otsa ja tubuliinid tulevad ära otsast.
Treadmill ku tubuliini kontsentratsioon on kõrgem kui Cc + otsas, aga madalam kui Cc otsas, saavad
mikrotuubulid kohapeal ,,liikuda" lisades alaühikuid ühte otsa ja dissotsieerides neid teisest otsast.
24. Joonista mikrotorukese struktuur ja tähista valgulised alaühikud.
Läbimõõt 25nm, koosneb tubuliinist. Tubuliinide sõlm on MTOC- microtubul organizing center. Moodustab ja -dimeere; duplette, triplette.
25. Loetle keemilisi ühendeid, mis inhibeerivad mikrotorukeste struktuurset dünaamilisust. Too välja ka nende ühendite toimekoht.
Mikrotuubulite dünaamikat saab muuta ravimitega. Näiteks taksaanide (C47H51NO14, C43H53NO14) ravimite
klass, mida kasutatakse vähi ravimiseks, blokeerib dünaamilist aktiivsust stabiliseerides GDP seondunud
tubuliini mikrotuubulis. Seetõttu isegi kui GTP hüdrolüüs jõuab lõppu, ei toimu depolümerisatsiooni ja
mikrotuubul ei tõmbu tagasi. Nocodazole ja Colchicine omavad vastupidist efekti blokeerides tubuliini
polümerisatsiooni mikrotuubuliteks.
26. Missugustes pankrease B-raku organellis toimub pro-insuliinist insuliini tootmine?
Pro-insuliini sünteesitakse ER-is, siis saadetakse Golgoisse. Pro-insuliinist tehakse lõplik insuliin sekretoorses
vesiikulis proteaaside abil, pärast Golgi kompleksist lahkumist.
27. Mis on SNARE valkude funktsioon?
(SNARE valkude perekond koosneb üle 60 liikmest) Peamine roll on transportvesiiklite fusiooni
vahendamine läbi rakumembraani porosoomi või suure vesiikli korral. Jaotatakse kaheks: v-SNARE-d on transpordivesiiklite
hulgas ja t-SNARE-d asuvad sihtmärkide membraanides.
NSF- heksameerne valk, mis seotuna SNARE-le lõhub viimatinimetatud kompleksi
28. Millised valgud osalevad RNA ekspordis?
RNA-l puudub valguline Nuclear Export Signal (NES), seega ta ei saa iseseisvalt tuumast välja. Tuumast
väljumiseks peab ta seonduma valguga, mis omab NES-i. Näiteks Ran-GTP ja eksportiin. 45
29. Loetle kõik tuumakeses sünteesitavad RNA tüübid. Missugused ensüümid neid RNAsid transkribeerivad?
Tuumakeses sünteesitakse rRNA-sid- RNA pol 1; snRNA- RNA pol 3, osad ka pol 2 poolt
30. Mis vahe on miRNAl ja siRNAl?
MikroRNAd (miRNA; 21-22 nukleotiidi) on leitud eukarüootidest ja on aktiivsed RNA interferentsis, kus
efektor kompleks miRNA-st ja RNA ensüümidest võib lõhkuda mRNA, millele miRNA on komplementaarne,
takistada mRNA transleerimist või kiirendada selle lõhkumist. Small interfering RNAd (siRNA; 20-25
nukleotiidi) saadakse sageli viiruslike RNA-de lõhkumisest. siRNA käitub RNAi-s sarnaselt miRNA-le.
31. Kirjelda hnRNP A1 tuumast ekspordi protsessi. Missugused valgud selles osalevad? Kuidas rakk kontrollib, et see protsess on ühesuunaline? Missugune tsütoplasmaatiline valk on selle protsessi toimumiseks oluline ja miks?
Heterogeensete ribonukleovalkpartiklite (hnRNP), mis sisaldavad heterogeenset tuuma RNAd (hnRNA), mazhoorsed
valgulised alaühikud on pre-mRNA-d, mis ei ole rakus vaba nukleiinhappena, vaid on kaetud tuuma valkudega. hnRNP-d
kasutatakse tähistamaks pre-mRNA ja teiste, erineva suurusega tuuma RNAde komplekse. hnRNP valgud on väga
heterogeensed suurusega 34 kuni 120 kD. Nagu TFid, on ka enamus hnRNP valke modulaarse struktuuriga. Neil on üks
või enam RNAgaseostumise domeeni ning üks valkudega interakteerumise
domeen . Tänaseks on teada palju erinevaid
RNAga-seostumise motiive.
pre-mRNAde kompleks hnRNP valkudega takistab lühikeste sekundaarstruktuursete alade moodustumist, mis teeksid pre-
RNA teistele RNAdele või valkudele äratundmismärklauaks. Erinevad hnRNP valgud (A1, C, D)
tunnevad ära erinevaid järjestusi, näit. hnRNP valgud A1, C, ja D tunnevad ära RNA pürimidiinirikkaid alasid.
Lõplikult protsessitud mRNA-5' ots ning hnRNP valk-kompleks (mRNP) assotseerub "cap"-siduva
kompleksiga (CBC), mis läbib esmalt NPC. Kui mRNPsid transporditakse läbi NPC, siis tuumas lokaliseeruvad
hnRNPd eemaldatakse; need valgud, milledel pole NESi (nuclear export signal, tuumaekspordi signaali),
hoiavad mRNAd tuumas. NES-sisaldavad hnRNPd, nagu hnRNP A1, transporditakse läbi NPC ning nad
kannavad assotseeritud RNA tsütoplasmasse. Tsütoplasmaatiline RanGAP stimuleerib Rani GTPd
hüdrolüüsima.
Shuttle 'vad hnRNP valgud dissotseeruvad retseptorvalkudelt (leutsiin-rikaste NESide puhul
eksportiinilt) ning nad transporditakse tagasi tuuma. mRNA seondub nüüd tsütosoolis asuvate mRNP
valkudega, nende hulgas polü(A)ga-seonduva valgu PABPga, mis interakteerub mRNA 3' otsaga.
32. Mis on rakulise diferentseerumise molekulaarseks aluseks?
Ulatuslik rakkude eristumine sõltub geeni ekspressiooni kvantitatiivsetest ja kvalitatiivsetest muutustest, mida
reguleeritakse suures osas transkriptsiooni tasemel mõnes organism muutub transkriptsioon iga vähimagi
muutuse peale.
Seda tehakse näiteks erinevate signaaliradadega (põhilised on Notch, Wnt), mis määravad transkriptsiooni-
faktorite tootmise. Differentseerumine toimub raku sugupuu alusel.
Transkriptsiooni faktorid koordineerivad diferentseerumist aktiveerides või represseerides transkriptsiooni.
Rakuvälised kasvufaktorid tsütokiinid reguleerivad diferentseerumist
andes signaalirajale signaali.
Lihasraku moodustamises on väga oluline roll HDAC-itel (inhibeerib) ja HAT-idel (aktiveerib). Kui HDAC on
fosforüleeritud, asetseb see tsütoplasmas ja ei saa inhibeerida.
Kui DNA-d metüleerida, siis ei saa sealt ka transkriptsiooni teostada.
Põhimehhanism ühe raku poolt toodetud ligand seondub teise raku ekstratsellulaarses regioonis asuva
retseptori külge, muutes retseptori struktuuri. Tsütoplasmaatilise domääni kuju muutub ja retseptor omandab
ensümaatilise aktiivsuse ja katalüüsib reaktsioone, mis fosforüleerivad teisi valke, aktiveerides neid.
Fosforüleerimisreaktsioonide järgnevus aktiveerib transkriptsioonifaktori või tsütoskeleti valgu, mis viib raku
diferentseerumiseni.
33. Kas diferentseerunud rakk võib valida mõne muu arengusuuna?
Üldjuhul seda enamasti ei toimu, aga vahel võib. Varem arvati, et raku diferentseerumine on pöördumatu
protsess. Katseliselt näidati esmalt, et
hiire luuüdirakkudest võivad areneda skeletilihased. Ajaliselt võttis see
nädalaid, seega võib eeldada, et metaplaasia on
mitmeastmeline protsess. Näiteks luuüdi päritoluga rakust on
siiani saadud vöötlihas, endoteeli, südamelihas, purkinje
neuron (suurimad
neuronid inimese ajus=,
hepatotsüüte. Vöötlihasest aga luuüdi. Närvkoest vererakke, vöötlihasrakke, lootekudesid.
Vähirakk võib diferentseerunud rakust tagasi vähitüvirakuks muutuda. 46
34. Miks erinevate regulaatorvalkude ekspressioon viib erinevate rakutüüpide tekkele?
Sellepärast, et erinevad regulaatorvalgud on omased erinevatele rakkudele.
Regulaatorvalgud on ka transkriptsioonifaktorid ning need hakkavad läbi viima edasist geeniekspressiooni.
Aktivaatorid ja repressorid seonduvad spetsiifilistesse
kohtadesse DNA-l ja reguleeruvad sarnaste valku
kodeerivate geenide ekspressioni. Nad toimivad kas koos teise valkudega muutes kromatiini struktuuri ja
mõjutades generaalsete transkriptsioonifaktorite seondumist või toimivad ise transkriptsioonifaktoritena läbi
mediaatori.
35. Miks on geeni ekspressiooni
arenguline regulatsioon oluline? Too üks näide, kuidas see toimub (väliskeskkonnast tuumani). Hulkraksetel peab arenevas lootes olemasolevast mitmesajast erinevast rakutüübist igat tüüpi loodama õigel hulgal õiges ajaaknas/
hetkes ja asukohas nii, et see integreeruks ümbritsevate rakkude ja kudedega. Eristunud/diferentseerunud rakkudel on tihti oma kindel/äratuntav
morfoloogia ja neis ekspresseeruvad valgud on spetsifitseerunud teatud kindlate
bioloogiliste funktsioonide täitmisele, mis on siis omased vaid sellele rakutüübile. Loomade ja taimede arengus toimuv ulatuslik rakkude eristumine sõltub geeni ekspressiooni kvantitatiivsetest ja kvalitatiivsetest muutustest, mida reguleeritakse suures osas transkriptsiooni tasemel. Sellise
rajaga tekivad näiteks vere-,
epiteel - ja soolerakud.
36. Loetle vähemalt 4 erinevat rakupinna retseptori perekonda, mis on olulised kõigi hulkraksete varaseks arenguks, ja kirjelda nendega seotud signaaliülekande raja komponente?
Wnt rada Wnt, retseptor -Frizzled 1 -10 (vere, epiteeli ja soole rakud)
SHH rada SHH, IHH, DHH, retseptor - patched1, 2 (vere, närvi ja
sugurakud )
Notch rada
Delta , Jagged, Serrate, retseptor - Notch 1,2,3 (vere, närvi ja sugurakud)
RTK EGF, retseptor EGFR; FGF retseptor FGFR, NTs retseptor TRKs
Mehhanismid allpool.
37.
Tuues näiteid, kirjelda, kuidas polüpeptiidsed hormoonid (k.a. neurotrofiinid) võivad reguleerida geeni ekspressiooni.
Nerve
growth factor (NGF) kuulub neurotrofiinide perekonda, mille liikmed osalevad rakkude elus
hoidmises ja jagunemises. NGF on oluline ka neuronite regeneratsioonil ning on potentsiaalseks Alzheimer'i
tõve ravimiks. NGF arvatakse olevat ka teisi füsioloogilisi funktsioone, eeskätt immuunsüsteemis. NGFil on
kaks retseptorit TrkA ja p75(NTR). NGFi signaali edastav retseptor on türosiinkinaasne TrkA ja mõnedes
rakkudes indutseerib p75 programeeritud rakusurma (apoptoosi). NGFi kõrge afiinsusega sidumiseks on vaja
mõlemat retseptorit.Sidumise tulemusena, retseptori türosiinkinaas aktiveerub ja indutseerib signaalsete
kaskaadide tekke. Üheks signaali
rajaks on, phospholipase C aktiveerumine, kusjuures vabaneb DAG ja IP3ning
aktiveeritakse assotseeritud rajad, nagu näiteks protein kinase C rada. Teiseks NGF-aktiveeritud rajaks on
rasseoseline MAP kinaaside kaskaad. See rada aktiveerib TFi AP-1. Lisaks sellele TFile i on NGFi
radade märklaudadeks ka Egr ja CREB. Egr perekonnaliikmed nagu ka Mek/Erk rada osalevad NGFseoselises
neuriitide väljakasvus. CREB perekonnaliikmed on olulised sünpaatiliste neuronite elus hoidmises.
Maksarakkudes , hormoon-
ajendatud fosfolipaas C aktivatsioon reguleerib glükogeeni lagundamist ning sünteesi
inositol-lipiidi kaheharulist signaalrada pidi. Fosfolipaas C genereerib kaht second messenger'i,
diatsüülglütserooli (DAG) ning inositol-1,4,5-tri-
fosfaat 'i (IP3). DAG aktiveerib proteiin kinaas C, mis
fosforüülib glükogeeni süntetaasi andes fosforüülitud inaktiivse vormi, seega inhibeerides glükogeeni sünteesi.
IP3 põhjustab tsütosoolsete Ca2+ ioonide tõusu, mis aktiveerib glükogeen fosforülaas kinaasi, just nagu
lihasrakkudeski, alandades glükogeeni. Antud juhul on erinevate rakusiseste signaal-transduction radade
aktiveerijaks üks ja sama signaal.
Maksarakkudes epinefriini poolt ajendatud glükogenolüüsi suurenemine stressiperioodil viib veresuhkru taseme
kõrgenemiseni. Argise elurütmi ajal on veresuhkru tase insuliini ning glukagooni dünaamilise kontrolli all. 47
38. TGFb/BMP signaalirada, Notchi signaalirada, WNT signaalirada, türosiinkinaaside signaalirada, SHH signaalirada. Põhilised aktiivsusekandjad väliskeskkonnast tuuma ja märklaud geenid.
TGFb/BMP signaalirada
Notchi signaalirada
türosiinkinaaside signaalirada 48
WNT signaalirada SHH signaalirada
peale SHH veel IHH ja DHH
39. Kirjelda neurogeneesi ja müogeneesi protsesside molekulaarset sarnasust bHLHde võrgustiku (signaalikaskaadide) näitel.
Neurogeneesi regulaatorite sarnasus müogeensete bHLH valkudega
· Selgroogstetel on kirjeldatud valkude olemasolu, mis on
Drosophila bHLH valkude Achaete ja Scute
sugulased. Nendeks on neurogeniinid ja NeuroDd. neurogeniin ekspresseerub väga varases arengus, on vajalik
NeuroD indukstiooniks ning funktsioneerib kui neuraalsete prekursorrakkude
determinatsiooni faktor.
· neurogeniin-1 deletsiooni kandvatel hiirtel puuduvad trigemiinsed ganglionid, muud närvisüsteemi struktuurid
on aga normaalsed, mis
viitab , et neurogeniin-1 ja neurogeniin-2 on vastastikku funktsionaalselt kõdunud.
Imetajate skeletilihaste müogenees jaguneb kolmeks põhistaadiumiks:
· lihasrakkude prekursorite (eelasrakkude) e. müoblastide determinatsioon
· müoblastide aktiivne jagunemine (
proliferatsioon ) ja mõnigail juhtudel ka migratsioon
· ja sellele järgnev müoblastide eristumine (diferentseerumine) küpseteks lihasrakkudeks. 49
40. Mis vahe on totipotentsel, pluripotentsel ja multipotentsel rakul?
Totipotentne võib olla eellaseks kõikidele rakkudele. Viljastatud
munarakk on totipotentne, st ta sisaldab
geneetilist infot, et areneda igaks keharakuks ja ka platsentaks ning embrüovälisteks rakkudeks, seega arenema
terviklikuks inimorganismiks. Inimese arengu 4.-5. päevaks on see totipotentne rakk 3-4 jagunemise tulemusena
andnud järglastena hulgaliselt omasuguseid.
Pluripotentne on arenenud totipotentsest rakust, võib olla eellaseks peaaegu kõikidele rakkudele, määravaks
on
looteleht . (Embrüonaalne tüvirakk (ES)): Umbes 5ndal arengupäeval hakkavad need totipotentsed rakud
diferensteeruma (e. spetsialiseeruma) ja moodustavad seest tühja rakukogumi, mida nim. blastotsüstiks.
Blastotsüsti välimine rakukiht areneb platsentaks ja
sisemisest rakukihist arenevad edasise arengu käigu kõik
keha rakud. Sisemine rakukiht on pluripotentsete rakkude kogum, st et neil rakkudel on võime areneda igaks
keha rakutüübiks va platsenta ja looteväliste kudede rakud ega seega ole võimelised arenema terviklikuks
organismiks. Pluripotentseid rakke saab eraldada blastostüstist ja loote sugurakkudest
Multipotentne võib areneda mitmeks rakutüübiks, tingimuseks on sama kude (sama rakutüüp).
Pluripotentsed rakud spetsialiseeruvad/ diferentseeruvad edasi multipotentseteks rakkudeks, mida tihti nim. ka somaatilisteks
tüvirakkudeks. Multipotentsiaalsed tüvirakud on eritüübilised tüvirakud, mis on võimelised arenema vaid
kindlateks
kudedeks : veretüvirakud vereks, nahatüvirakud nahaks jne. Multipotentsed rakud on olemas nii
varases arengus ka täiskasvanud organismis.
Senini on suudetud identifitseerida umbes 60 erinevat tüvirakku, st
ei ole teada, kas kõigi erinevate rakutüüpide jaoks on oma tüvirakud.
41. Loetle tüviraku omadusi.
Tüvirakk on selline jagunemisvõimeline rakk, mille järglasteks võivad olla kas teda ennast asendavad tüvirakud
(taastootmine) või diferentseeruvad rakud (naha tüvirakud jms)
42. Mis on
somaatilised ja mis on embrüonaalsed tüvirakud?
Somaatilised on multipotentsed rakud (keha kudedest) ja embrüonaalsed on pluripotentsed rakud (embrüotest).
43. Millised on tüvirakkudega seotud rakuteraapia peamised probleemid (takistused)?
Tüvirakkude
viimisel inimkehasse võib alguses saada küllalt häid tulemusi, aga paari kuu mööduddes on ~90%
sisse viidud rakkudest surnud. Samuti tihti ei hakka tüvirakud täitma rolli, mida me
soovime , et nad täidaks,
näiteks ei taasta funktsioneerivat närvkudet. Lisaks suremusele kipuvad sisse toodud tüvirakud ka rändama
minema, võib tekkida vähk.
44. Missugused on inimteraapias kasutatavad potentsiaalsed tüvirakkude allikad?
1) võtta
inimeselt endalt tüvirakke, näiteks luuüdist, ning anda neid talle mujale tagasi. 2) Võtta lootelt. 3) võtta
viljastatud munaraku blastotsüstist. 4) Võtta kloneeritud munaraku blastotsüstist 5) võtta lähedaselt sugulaselt?
* loomadelt, hiired, sead aga see pole siis tüviraku tasemel, see on see, et võtad hiirelt tüviraku (või võtad
hoopis koe), kasvatad neeru ning siis siirdad inimesele tegelikult siiski teed temast veidi GMO,
lased tal enda
sees kasvatada organi, opereerid välja ja siirdad inimesele (loom võib surra!)
45. Misugused onkogeeni kromosomaalse translokatsiooni molekulaarsed tagajärjed?
Onkogeen - Valku kodeeriv geen, mille regulatsiooni vead tekitavad vähi. Kromosomaalse translokatsiooni
tagajärjel suureneb geeni ekspressioon valel ajal ja
vales rakutüübis; võivad tekkida hübriidsed valgud, mis on
kogu aeg aktiivsed.
46. Selgita vähi tekkes tüviraku-
kontseptsiooni .
Mutatsioonid rakkudes on küllalt tavalised, aga igast
muteerunud rakust ei tule veel vähk. Vähi risk on palju
suurem, kui muteerub tüvirakk, kuna tüvirakk on oma olemuselt paljunemisvõimeline, seega kui tema puhul on
muteeritud nii onkogeen, tuumporsupressor (apoptoosi rada ei käivitu) kui ka geen, mis koordineerib liikumist,
saab temast vähirakk, mis on samas väga jagunemisvõimeline. Vähi tüvirakk võib tekkida samas ka juba
osaliselt diferentseerunud rakust. Täiskasvanutel on
haruldased lihas- ja närvikoekasvajad, sest seal enamasti
lõplikult diferentseerunud rakud.
47. Mis on
proto -onkogeeni funktsioon rakus ja kuidas selle muutumine onkogeeniks mõjub raku jagunemisele? eluspüsimisele? diferentseerumisele? surmale? genoomile?
Proto-
Onkogeenid kodeerivad valke, mis reguleerivad raku kasvu ja diferentseerumist. Onkogeeniks muutudes
hakkab rakk rohkem paljunema, vältima surma ja diferentseerumist, raku
genoom võib hakata suure kiirusega
muteeruma. 50
48. Mis on genoom? proteoom? transkriptoom?
reguloom ?
tselloom (cellome)?
metaboloom ? interaktoom? metüloom? morfoom? nukleoom? operoom? ORFeoom? translatoom? transportoom? unknome? Selle küsimuse vastuse leiad
http://www.geocities.com/pribond/bioinfo/glossary/omes.ht m
Genoom on ühes liigiomases kromosoomikomplektis (haploidne kromosoomistik) sisalduv geneetiline materjal.
Proteoom on organismis sisalduvate valkude kogum, mis on erinev organismi erinevates rakkudes ja muutub
pidevalt
Transkriptoom kõikide mRNA molekulide või transkriptide kogum, mis on olemas ühes rakus või rakkude
kogumis. Seda võib kasutada kõikide transkriptide kohta organismis või ka kindlas rakutüübis.
Reguloom regulatoorsete komponentide (geenid, mRNAd, valgud, metaboliidid) kogum rakus. See hõlmab ka
regulatoorseid
efekte nende komponentide vahel ja nende sõltuvust muutujatest.
Tselloom rakkude ja nende interaktsioonide bioloogiliste olemuste ning bioloogiliste rakkude terviklikkuse
kogum, mille põhiliseks moodustajaks on valgud.
Metaboloom on inimese ainevahetusprotsessi ehk metabolismi väikesemolekuliliste vahe- või lõppsaaduste
metaboliitide - kogum. Metaboliitide hulka kuuluvad nt aminohapped, suhkrud, rasvad ja kolesterool.
Interaktoom molekulaarsete vastasmõjude kogum, mida enamasti näidatakse graafikuga.
Metüloom metüleeritud DNA saitide ja genoomi mustrite kogum.
Morfoom morfoloogiliste tunnuste või liikide kaart ja klassifikatsioon.
Nukleoom kõikide nukleiinhapete kogum rakus
Operoom bakteris sisalduvate operonide kogum
ORFeoom is the
totality of open
reading frames(ORF) in biology
Translatoom kõikide valkude või nende järjestuste kogum, mis on ühes rakus või rakkude kogumis
Transportoom rakus sisalduvate transportvalkude ja süsteemide kogum (peamiselt membraanikanalid)
Unknome suur geenide hulk, mille kohta pole hetkel funktsionaalset informatsiooni
RAKKUDE
MANIPULEERIMINE JA KASUTAMINE EKSPERIMENTAALSES TÖÖS
1. Rakukultuuride eelised ja puudused võrreldes in vivo mudelitega.
Eelised: · Homogeensed võrreldes koerakkudega · Eksperimenditingimused on selgelt defineeritud · Rakukultuuris saab
isoleerida huvipakkuva raku ja sellest kasvatada geneetiliselt homogeense koloonia- rakkude
kloonimine · Geneetiliselt homogeenne rakutüvi - kloon
Puudused: ei saa jälgida organismi kui
tervikut .
Eritüübilised rakud on erinevad: · väljanägemiselt ja suuruselt · kaalult · laengult · tiheduselt · antigeensuselt jne. Osa erinevusi on võimalik kasutada teatud tüüpi rakkude eraldamiseks teistest rakkudest.
Rakkude eraldamise
enamlevinud meetodid: ·
tsentrifuugimine ; tsentrifuugimine gradiendis · afiinsusel põhinevad meetodid; magnet · FACS
2. Mikroorganismide kasvatamine kultuuris. · Escherichea coli, Saccaromyces cerevisiae · Kiire kasv ja väga lihtne
kasvukeskkond · süsiniku allikas: glükoos või glütserool · lämmastiku allikas: NH4+ ·
soolad : Na+, K+, Mg2+, Ca2+,
SO42 -, Cl-, PO43- 51
3. Mikroorganismide kasvukeskkond kultuuris. · Rikkamas söötmes ka osaliselt hüdrolüüsitud loom- või taimkudet, pärmiekstrakti. · Pooltahke keskkond - agar vs.
suspensioon .
Tingimused: · Optimaalne temperatuur · Niiskus · Õige pH · Lahustunud toitained · Hapniku olemasolu/puudumine · Fotosünteesivatel valgus · Vähe jääkaineid
4. Auksotrioofid, kuidas neid selekteerida? Replikajäljendite meetod.
Auksotroofid - mikroorganismide tüvi, mis kasvavad mingi
spetsiifilise toitaine olemasolu korral, selekteerida saab neid tõstes
teise toitainesse replikajäljendite meetodit kasutades.
Replikate/jäljendite tegemist kasutatakse muteerunud tüvede
tuvastamiseks (tõstetakse templiga). Selliste auksotroofsete tüvede
hoidmiseks tuleb nad pidada selektsiooni all! Minimalsele
söötmele tuleb lisada minimeerivat ainet.
E.Colil tekib mutatsioon106 -107 aluspaari kohta
5.
Loomsed rakukultuurid, nende erinevus mikroorganismide kultuurist. · Loomsed rakud vajavad rikastatud söödet. · AA: his, leu, ile, lys, met, phe, thr, trp, val + cys, glu, tyr ·
Seerum on vere mitterakuline osa. · Vitamiinid, kasvufaktorid (nt NGF, EGF, IL) insuliini, transferriini · Tavaliselt kasvavad
tasapinnal , mis on kaetud spetsiaalse maatriksiga. Selline maatriks on kinnituskohaks ECMile-hüaluroonhape,
kollageen ja teised ECM valgud · antibiootikumid · Konfluentne (samasuunaline, koos kulgev) kultuur · Osad kasvavad ka suspensioonis ja 3D-geelis
Rakkude kasvatamiseks on vaja: · Tagada kontrollitud keskkond (inkubaatorkapid) · Tagada steriilsus (vertikaalne laminaarne õhuvool) · Tagada töötaja turvalisus patogeensete mikroorganismide või rakukultuuride korral (horisontaalne õhukardin) ·
Steriilsed töövahendid
6. Defineeri primaarne rakukultuur ja rakuliin. · Primaarsed rakukultuurid on saadud loomsetest kudedest · Suurem osa loomsest koest eraldatud rakke jaguneb piiratud
kordi piiratud aja jooksul (keskmiselt 50 jagunemist), pärast seda rakud surevad · Nn. "transformeeritud rakud" jagunevad lõpmatult ja selliseid ühest rakust alguse saanud immortaliseeritud kloone nimetatakse rakuliiniks · Rakuliinid sageli aneuploidsed- kromosoomide arv erinev normaalsest · Rakuliinid pole tavaliselt differentsieerunud, osadel siiski algse koe funktsioonid 52
Jagunemised päevad
7. Mis on rakkude transformatsioon, transformatsiooni erinevad viisid? · Nn. "transformeeritud rakud" jagunevad lõpmatult ja selliseid ühest rakust alguse saanud immortaliseeritud kloone nimetatakse rakuliiniks · Rakkude transformatsioon võib tekkida iseeneslikult mutatsioonide tagajärjel, selliseid rakke saab eraldada ka nt kasvajakoest või tekitada nt. telomeraasi kodeeriva geeni sisseviimisega rakku · Erinevad rakutüübid transformeeruvad erineva sagedusega, väga suured on ka liikidevahelised erinevused
Mõned kultuuri viidud rakud differentsieeruvad ja võivad moodustada koesarnaseid struktuure
8. Rakkude fusioonid, milleks kasutatakse. · Rakud võivad teatud tingimustel liituda- rakkude
fusioon . Kui liituvad kaks erinevat rakku, tekib hübriidrakk, mida nimetatakse heterokarüooniks (heterokaryon). Selliste rakkude tuumad ühinevad rakkude jagunemisel. · Fusioonid tekivad iseeneslikult väga harva, selle nähtuse suurendamiseks: PEG,
elektriimpulss · Eri liikide rakkude fusioonid- geenide kaardistamine · Monokloonsete antikehade saamiseks tekitatakse rakkude fusiooni teel hübriidrakud (hübridoomid) · Monokloonne üks
primaarstruktuur IG nt hiired · Polükloonne primaarjärjestus erinev (ükskõik, mis loom nt küülik/lammas) 53
9. Mis on hübridoom, millised on selektsiooni tingimused?
Hübridoom on hübriidrakk, mis on saadud rakkude fusioonist (monokloonsete antikehade saamiseks)
10. Viiruste kasutamine
rakubioloogias . ·
Viirus on väike parasiitorganism, kes ei reprodutseeru ise, vaid kasutab selleks
peremees -rakku. Bakteriofaag/
faag . Looma- ja
taimeviirused . · Liigispetsiifilisus · Viiruste pärilikkuse
materjaliks on kas DNA või RNA, mis võib olla nii ühe kui ka kaheahelaline. · Virion on viiruspartikkel tervikuna, st. et ta koosneb nukleiinhappest ja välimisest valgukoorest ja on võimeline infitseerima rakku. (Lihtsaimad viirused kodeerivad 4 valku, keerulisemad umbes 200.) · Viraalne transduktsioon- võimaldab viia peremeesrakku uuritavaid geene · Viiruseid kasutatakse transkriptsiooni, rekombinatsiooni uurimiseks ·
Retroviirused - onkogeenide uurimiseks
11. Läbivoolu tsütomeetria ja FACS, põhimõte ja kasutusvaldkonnad.
Läbivoolu tsütomeetria - Flow cytometry; See identifitseerib rakud, mõõtes valgust, mida need hajutavad ja
fluoressentsi, mida need eritavad, kui nad läbivad laseri
kiirt . Sellega saab sorteerida rakke välja
kindlast segu
tüübist.
FACS - Fluorescence activated cell sorting= rakkude sorteerimine; Seade, mis põhineb läbivoolu tsütomeetrial
ja suudab leida ühe raku tuhandete teiste seast. Näiteks
kui antikeha spetsiifilisele kindlale rakupinna
molekulile ühendatakse fluorestsents värv, siis iga
rakk, mis seda molekuli kannab, seondub antikehale ja
eraldatakse teistest rakkudest, kui see fluorestseerub.
Peale sorteerimist saab eraldatud rakku kasvatada
kultuuris.
FACS-i kasutatakse tavaliselt, et puhastada erinevat
tüüpi valgeid vererakke, millest igaüks kannab oma
pinnal üht või enamat eristuvad valku ja seetõttu seob
monoklonaalseid antikehasid, mis on sellele
spetsiifiliselt. Ainult T rakud immuunnsüsteemis
omavad nii CD3 kui ka Thyl2 valke. Nende olemasolu
muudab T-rakud teistest vererakkudest või
põrnarakkudest kergesti eristatavaks.
Varieerides monoklonaalsete antikehade kasutamist rakkude
eristamisel, kaetakse väikesed magnetilised
tilgad 54
monoklonaalsete antikehadega, mis on spetsiifilised pinnavalkudele nagu CD3 ja Thyl 2. Nii kinnituvad sinna
ainult need rakud, millel on mõlemad valgud olemas ja neid saab taastada ettevalmistamisest adhesiooniga
väikesele magnetile katseklaasi küljel.
Veel saab läbivoolu tsütomeetriaga mõõta raku DNA ja RNA sisu ja määratleda selle üldist kuju ja suurust.
FACS-iga saab samaaegselt teha raku suuruse (hajuv valgus) ja DNA koguse (fluorestsentsi eraldamine DNA-
siduva värvi korral) mõõtmisi.
Analüüsi käik kontsentreeritud märgistatud rakkude lahus segatakse puhvriga nii, et rakud läbiksid laserkiirt
ükshaaval. Mõõdetakse nii fluorestseeruva valguse eraldamist kui valguse hajutamist (suurus ja kuju). Lahus
viiakse läbi
tila , kus moodustavad pisikesed tilgad, kus on enamasti ainult 1 rakk. Moodustumise ajal antakse
igale tilgale negatiivne elektriline laeng, mis on võrdeline selle eraldatud fluorestsentsiga. Ilma laenguta tilgad
ja erinevate laengutega tilgad eraldatakse elektrivälja abil ja kogutakse. (ühe tilga sorteerimine toimub
millisekunditega). Tunnis läbib masinat kuni 10 miljonit rakku, mistõttu saabki erinevate omadustega rakke
eraldada ja siis kasvatada.



































































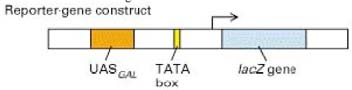
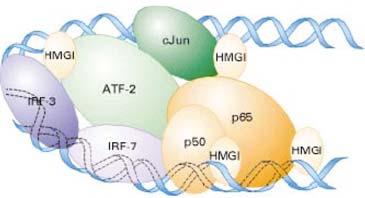
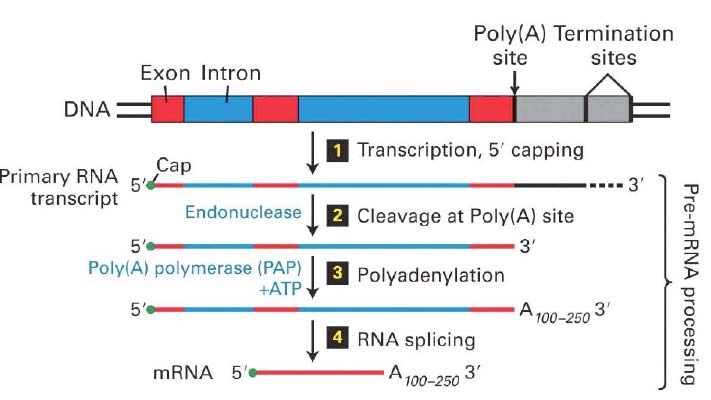
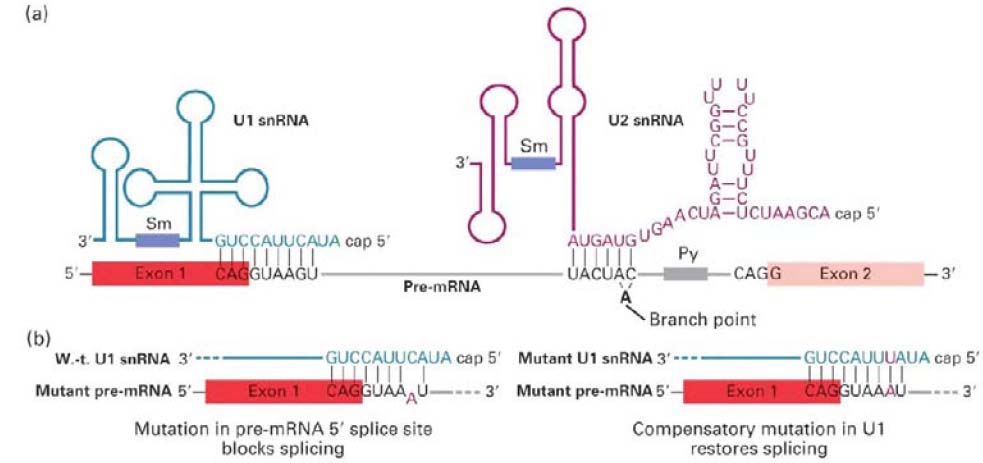
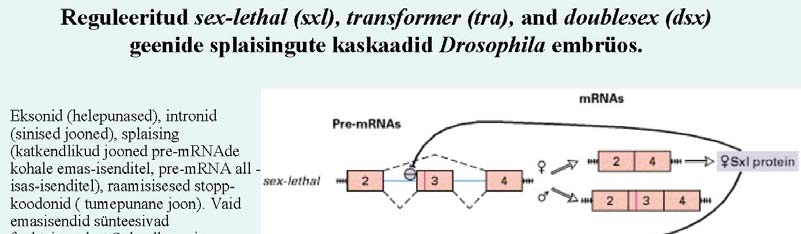
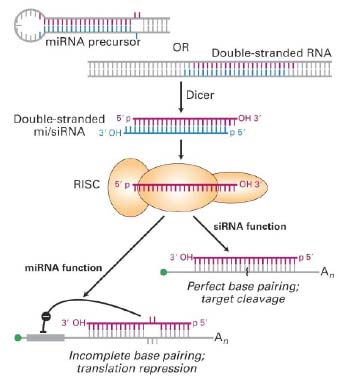

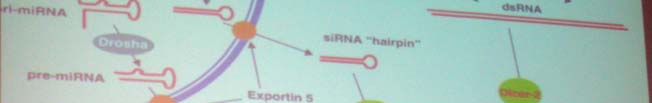




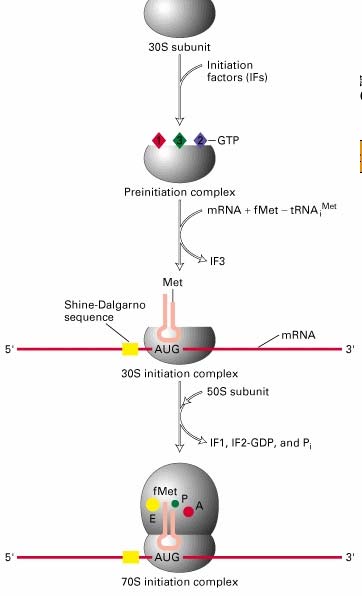
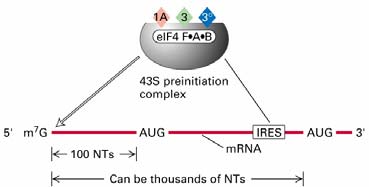
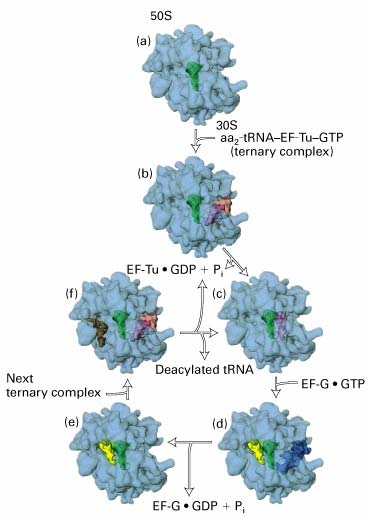
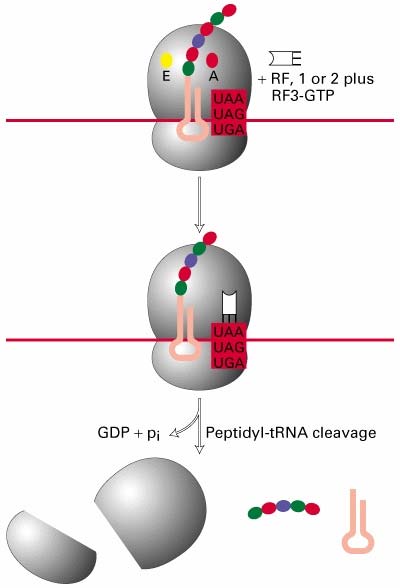
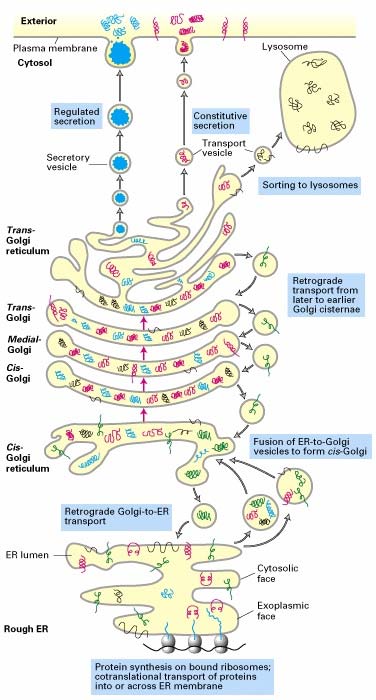
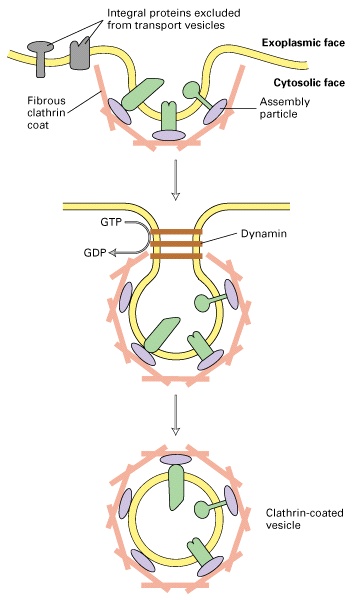
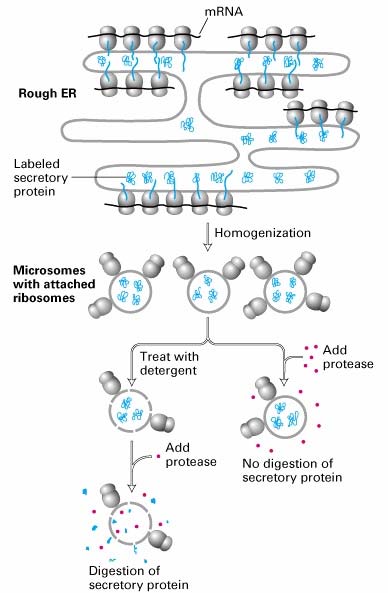
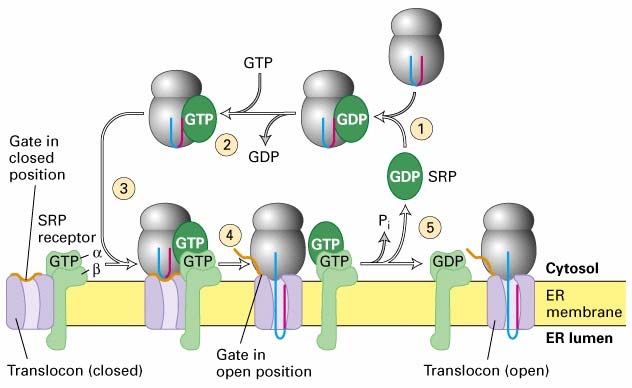
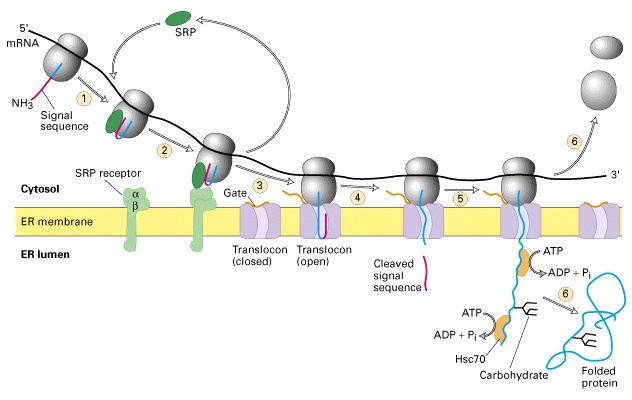

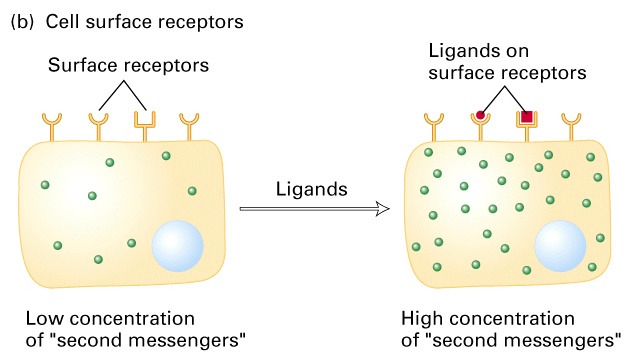
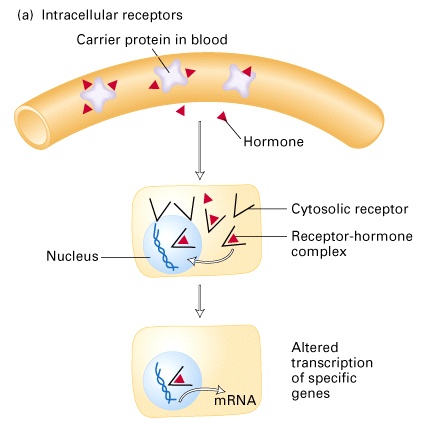
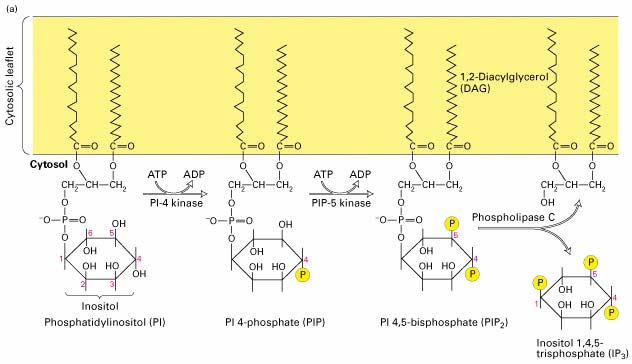
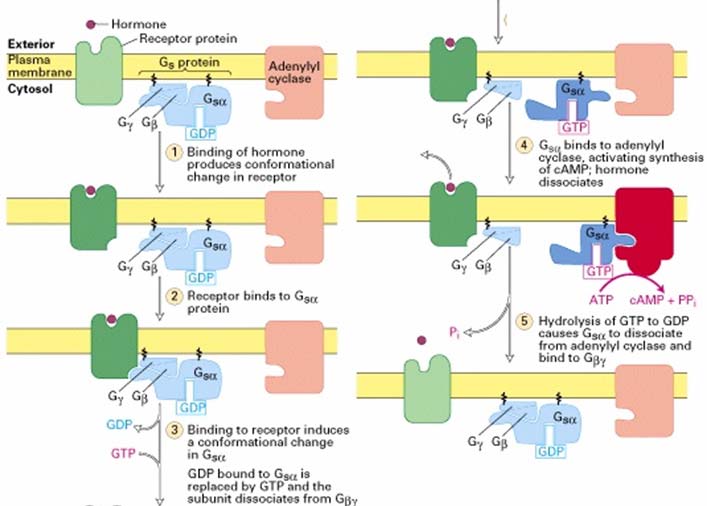
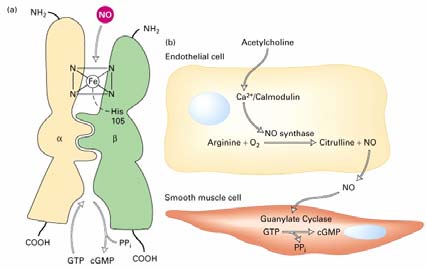
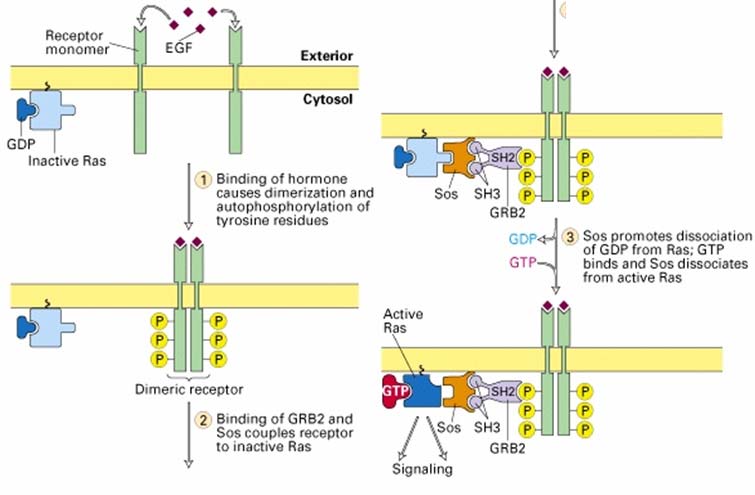
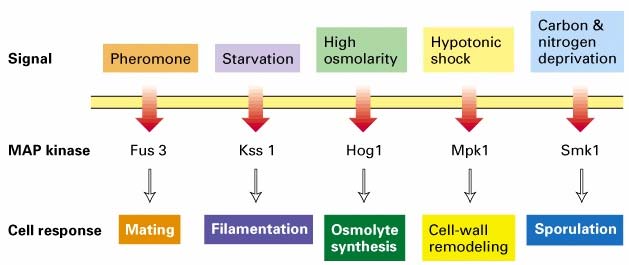
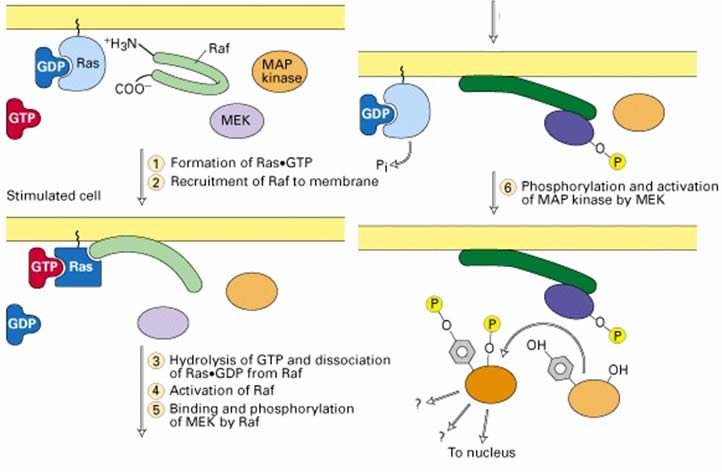
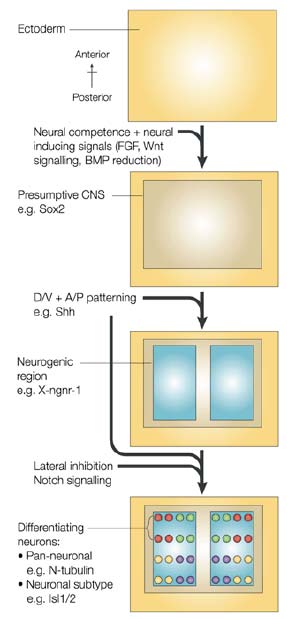
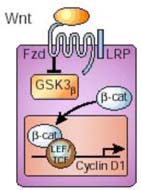
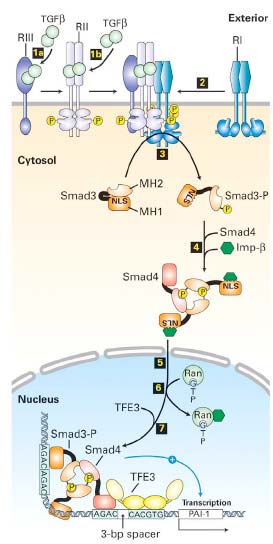
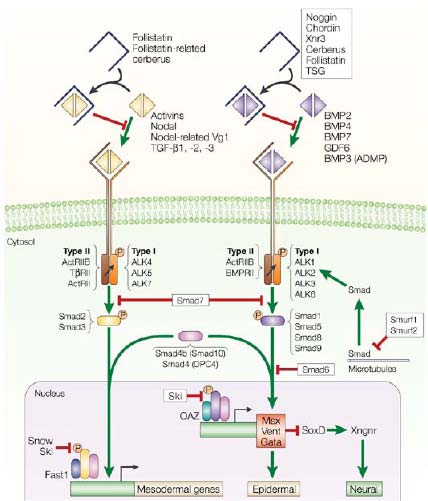
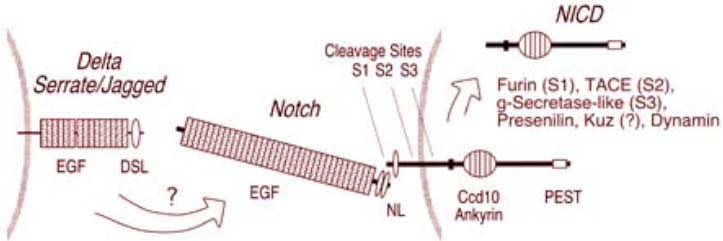
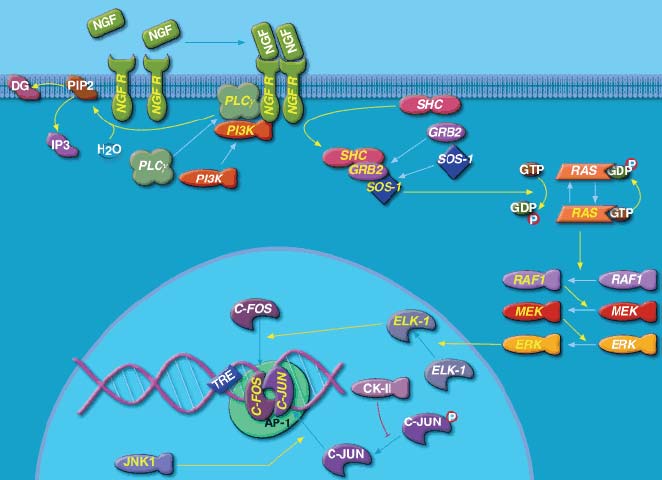
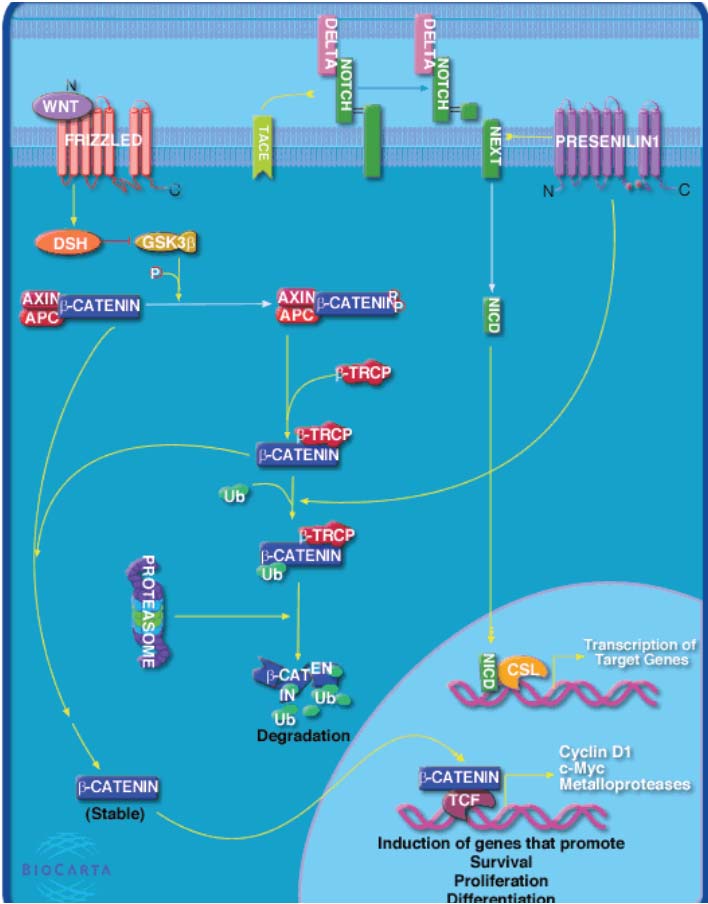
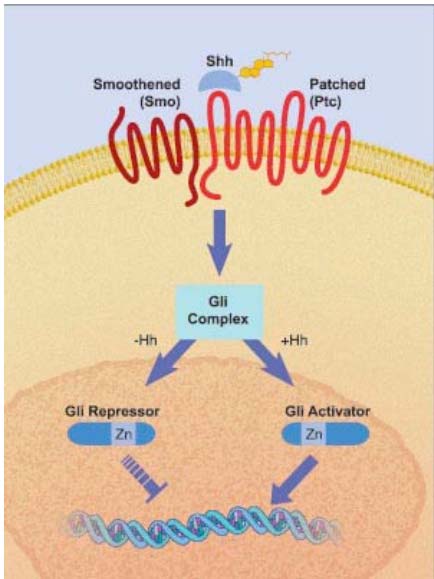
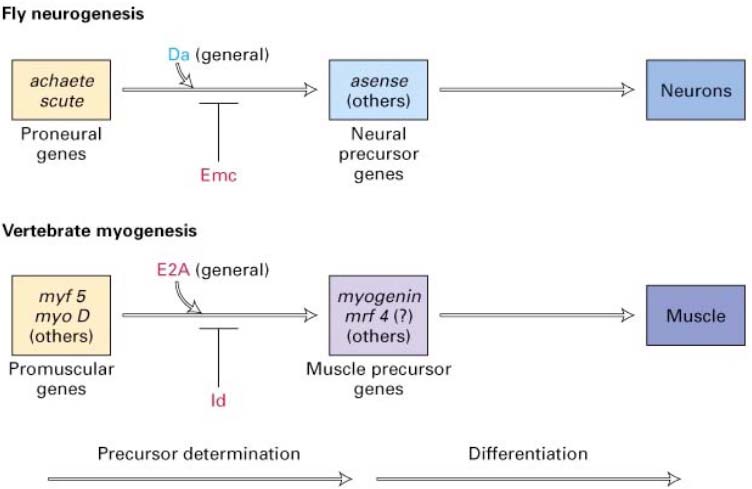
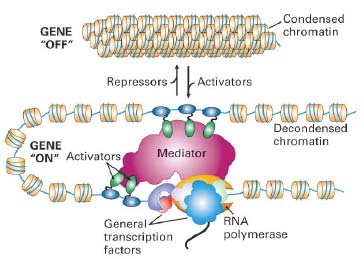
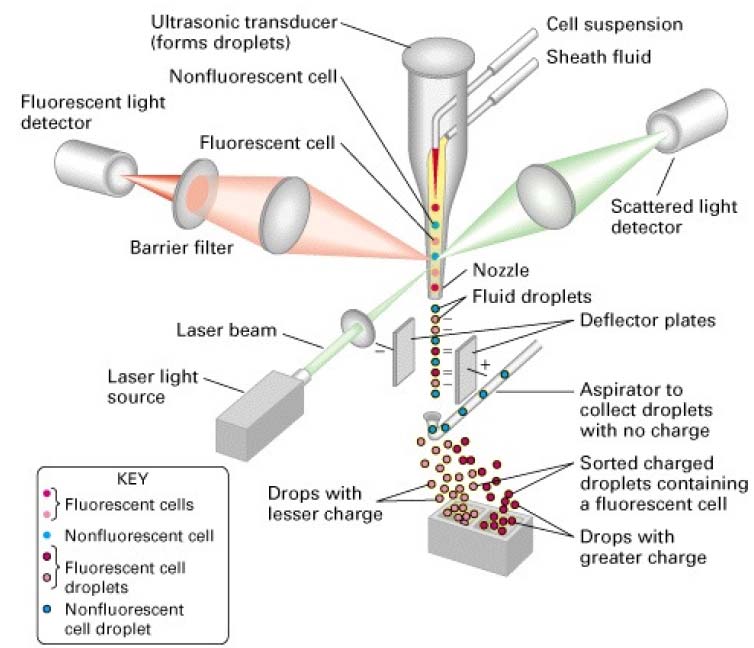
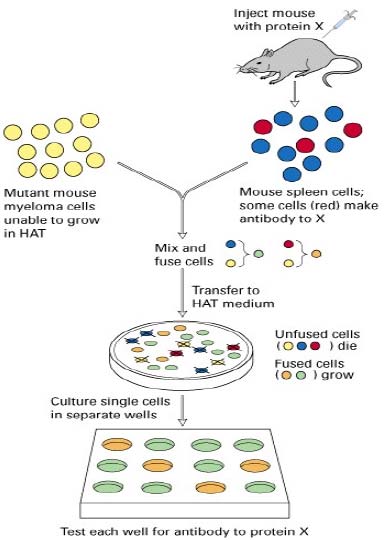
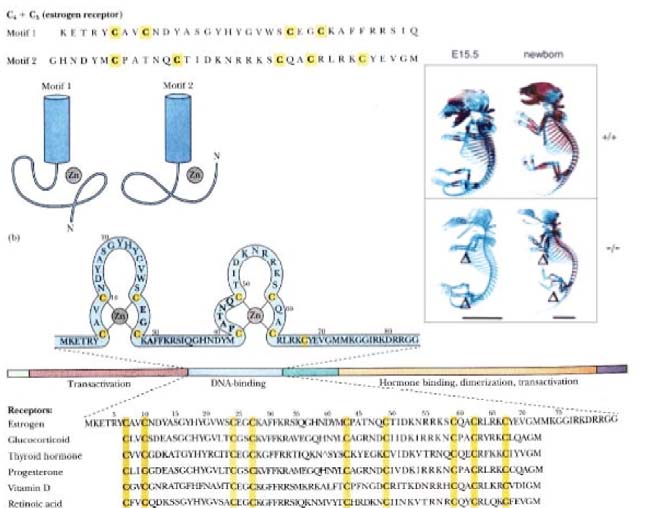
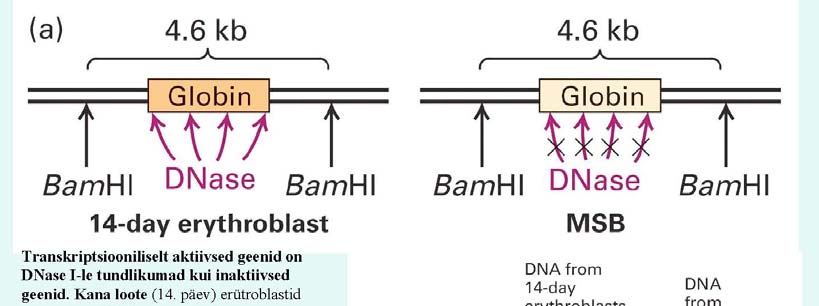
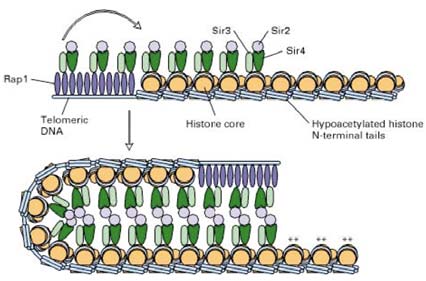
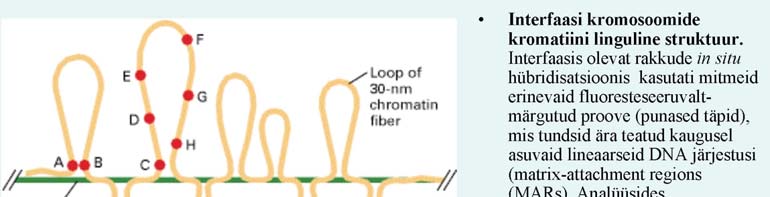

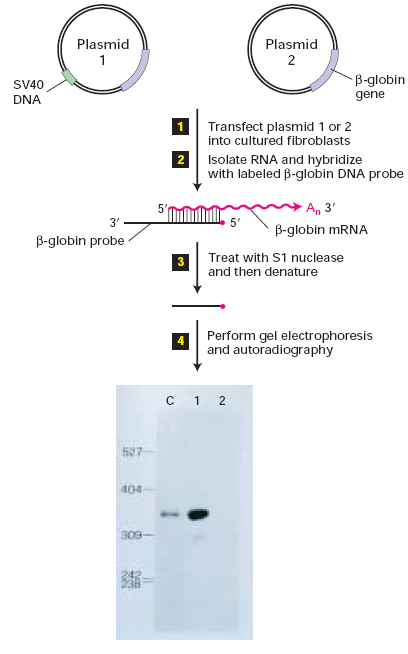
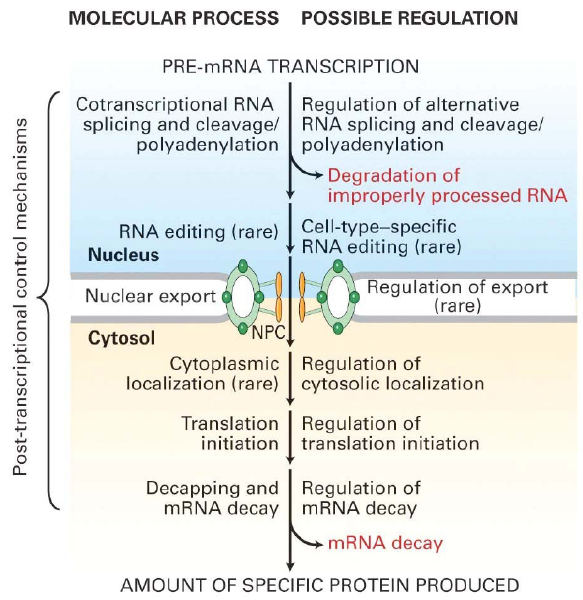
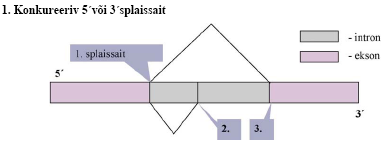
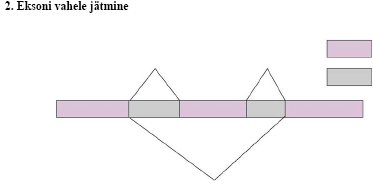
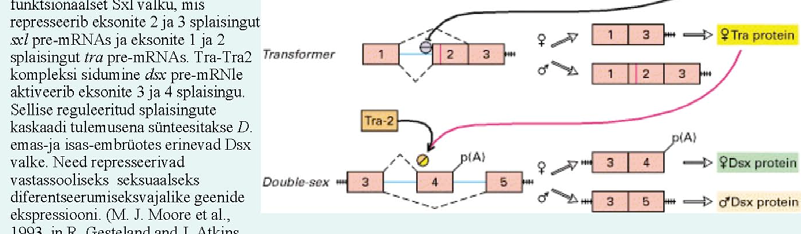
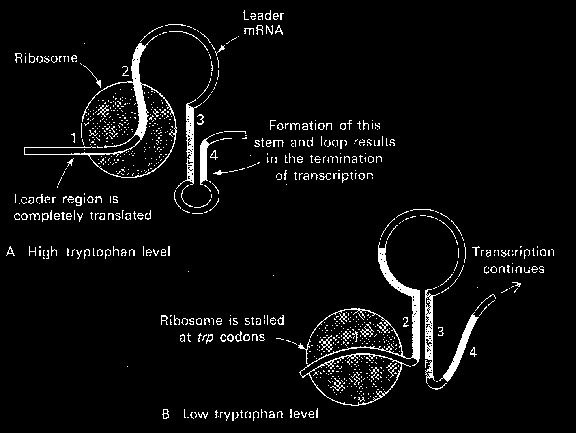
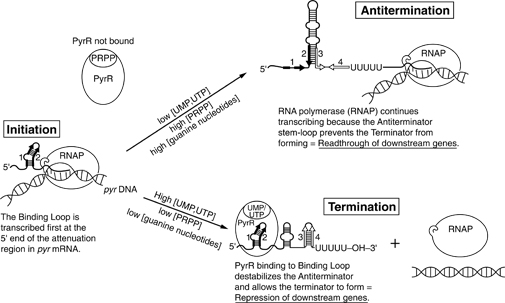
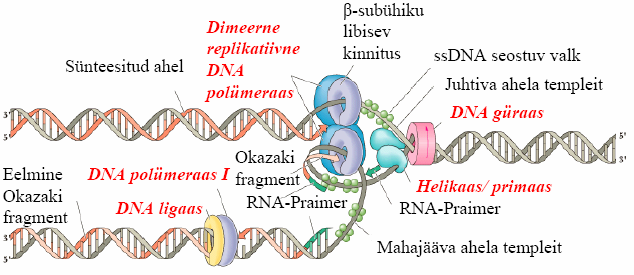
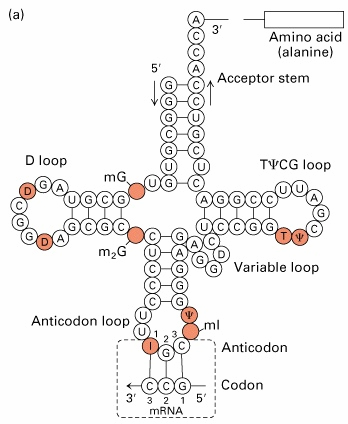
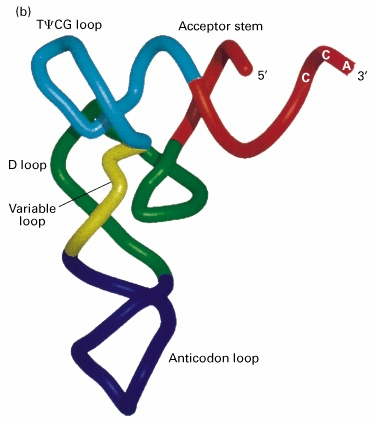
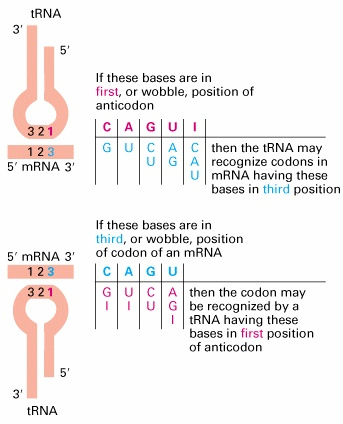
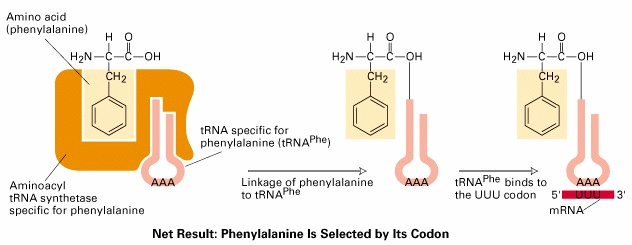
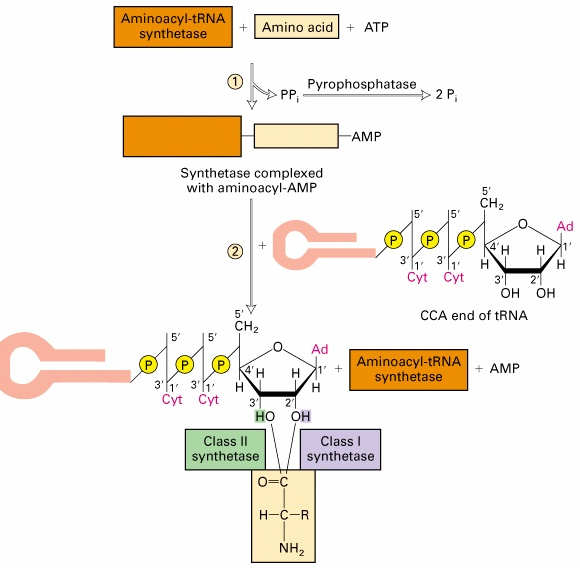
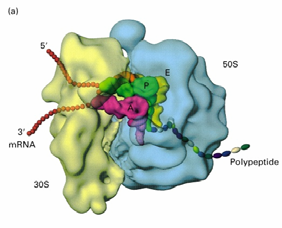
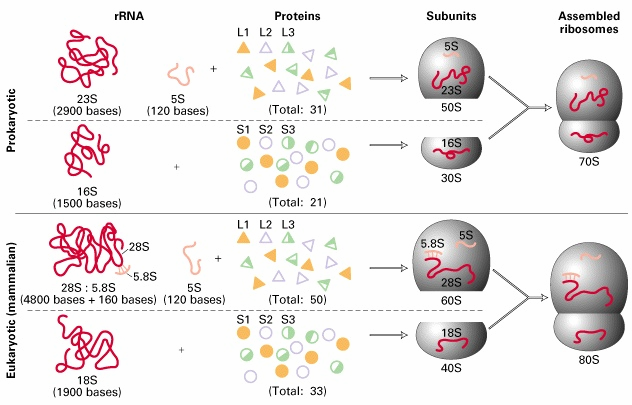
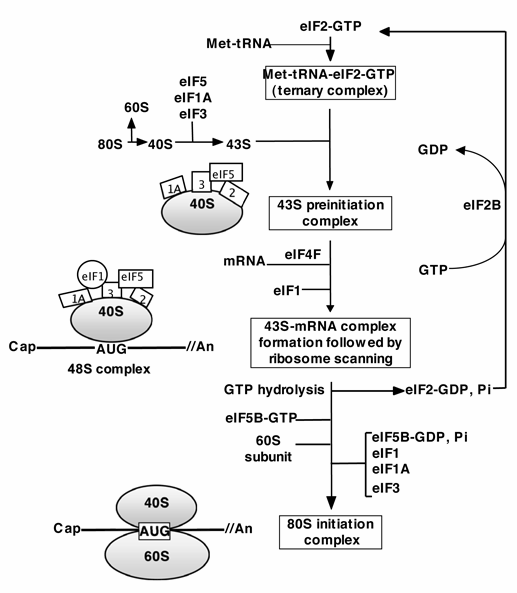
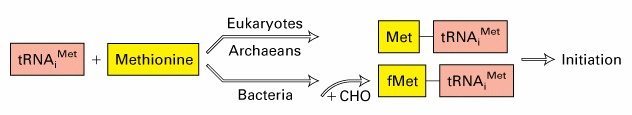
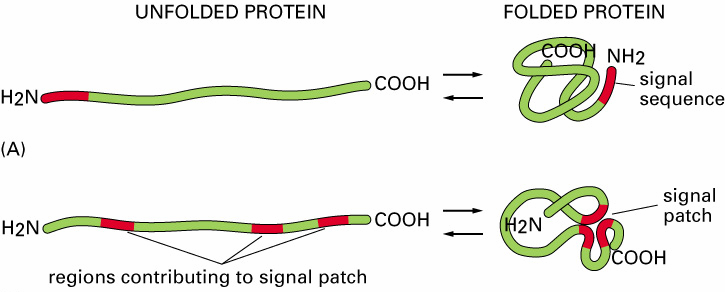
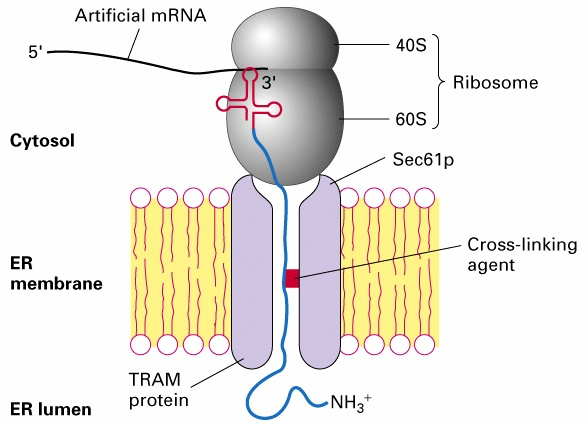
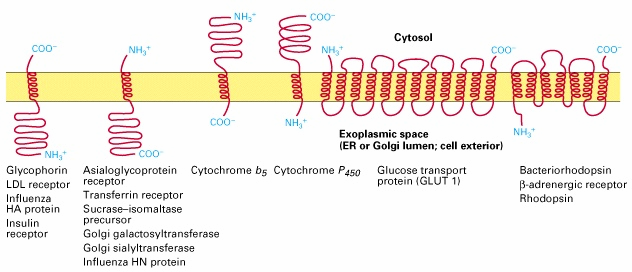
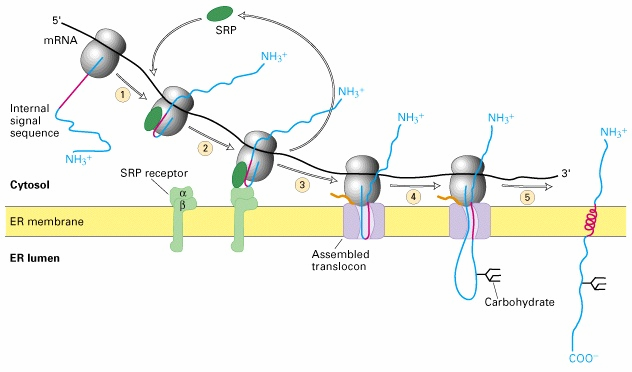
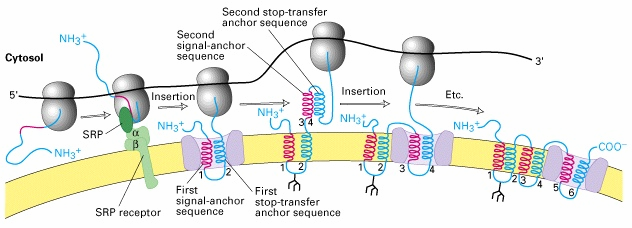
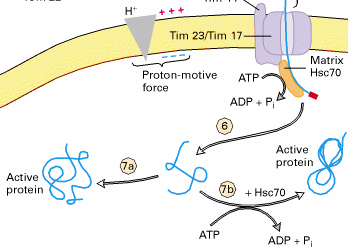
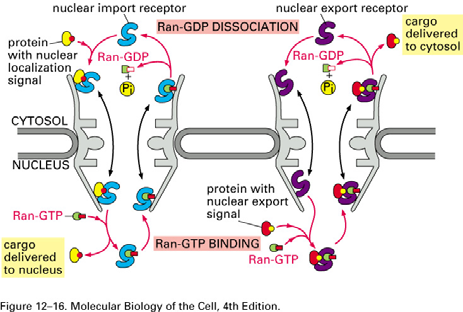
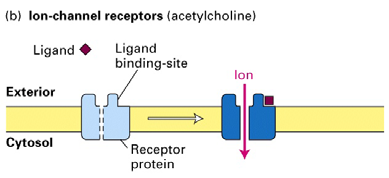
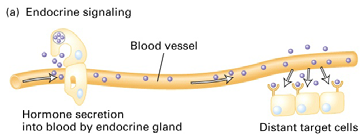
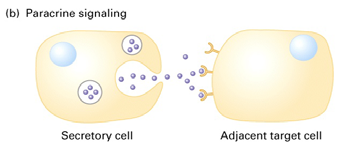
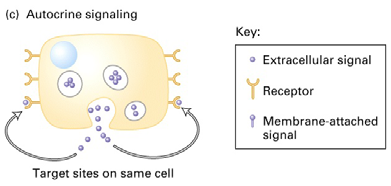
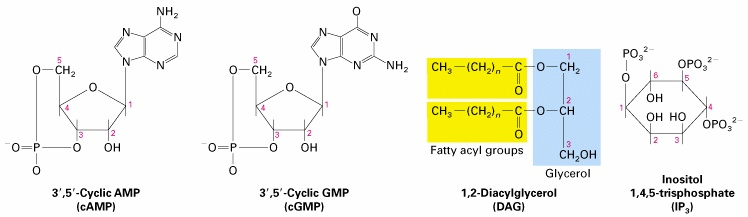
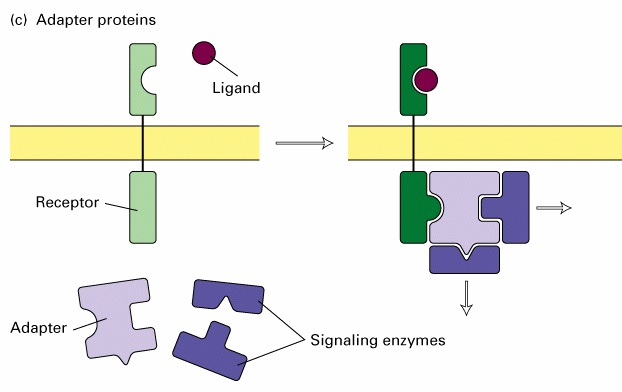
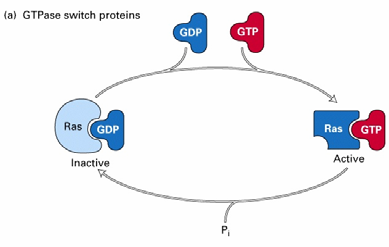
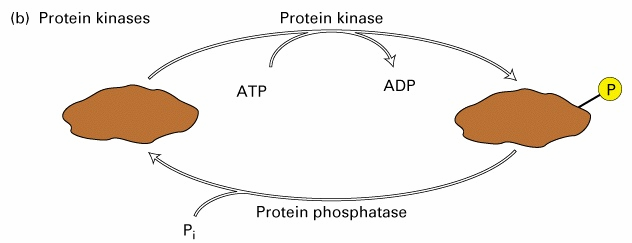
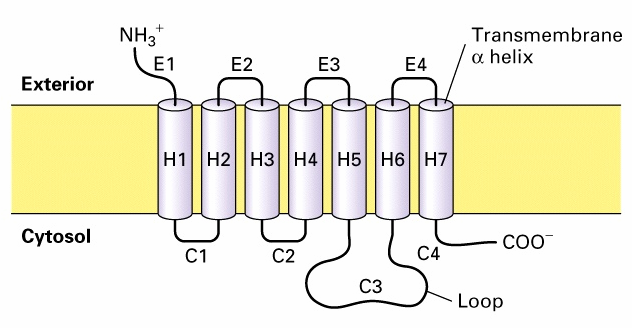
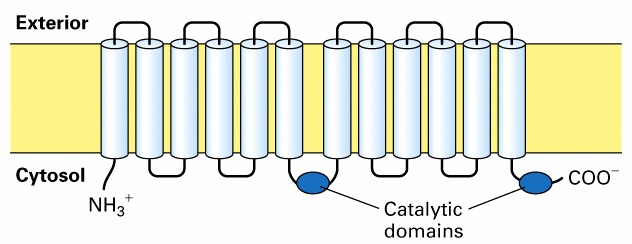
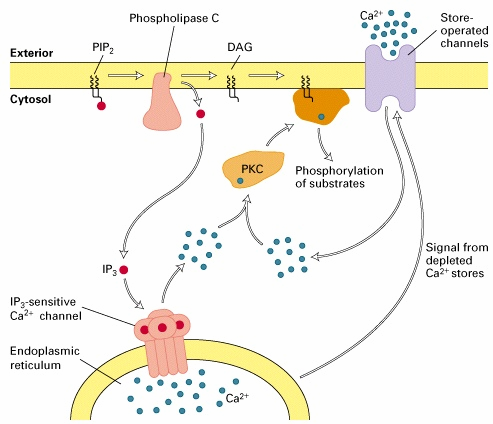
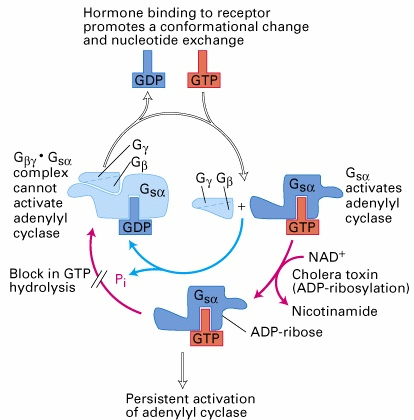
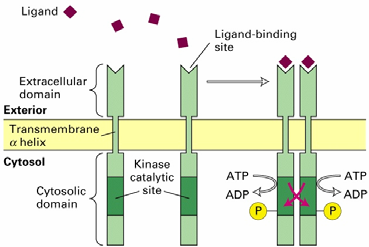
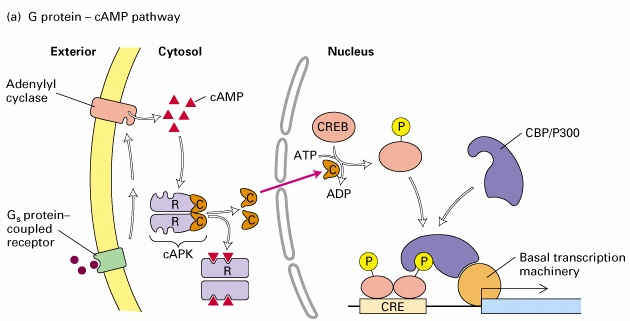
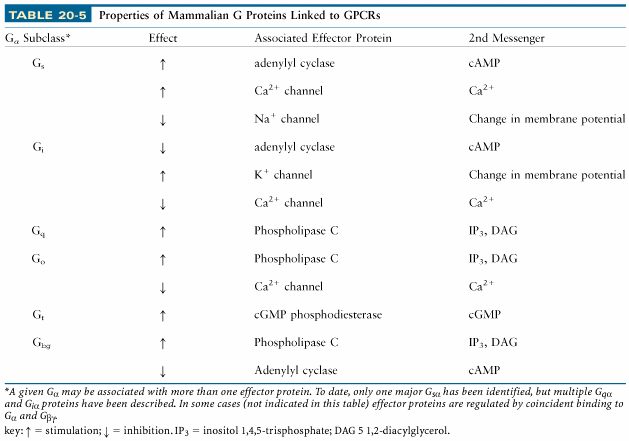
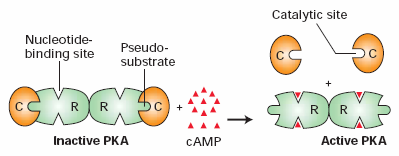
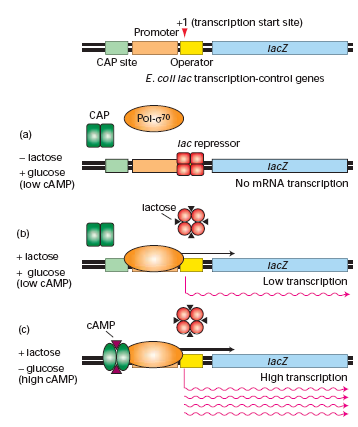
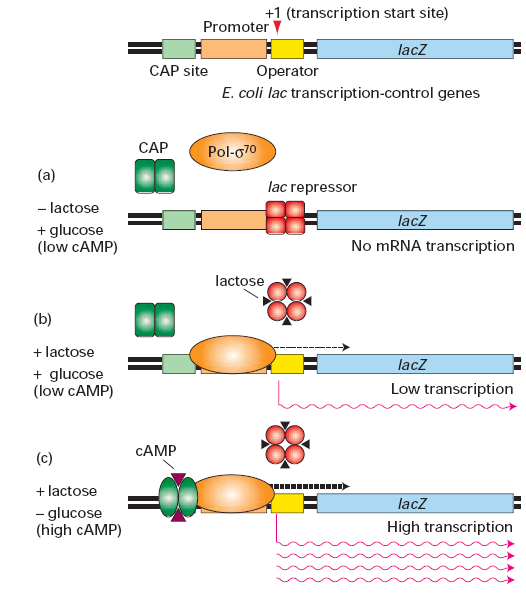
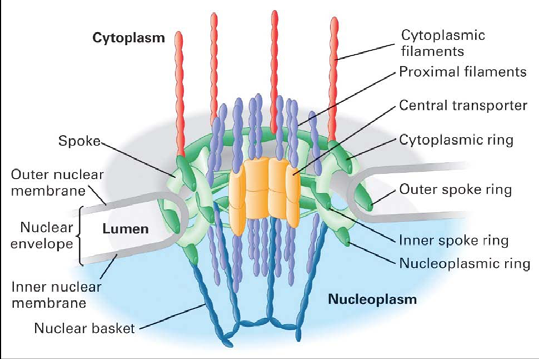
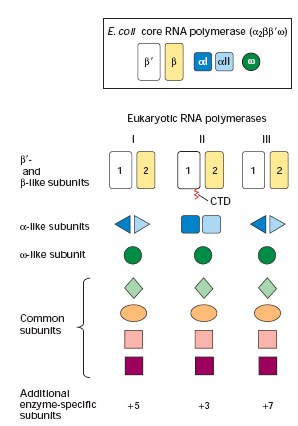
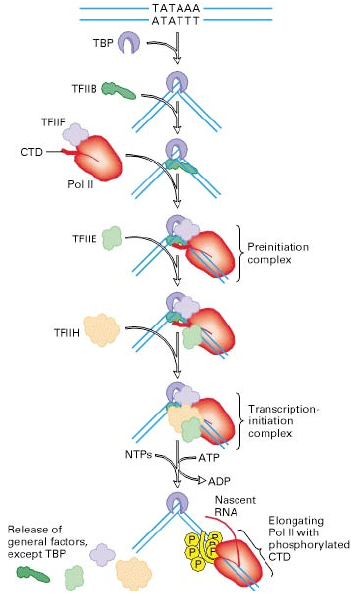
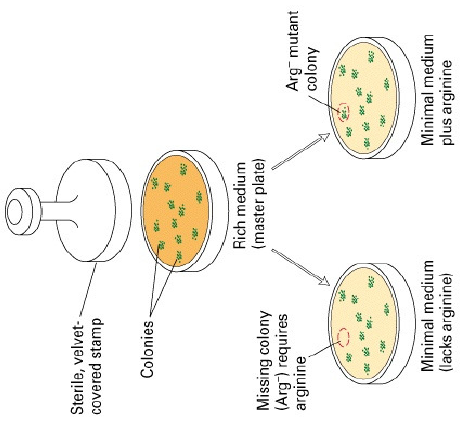
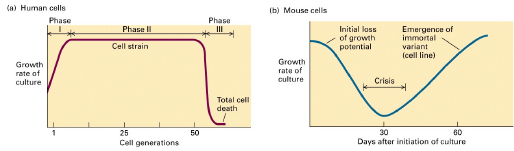
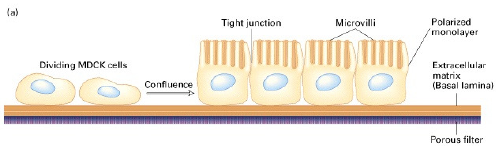
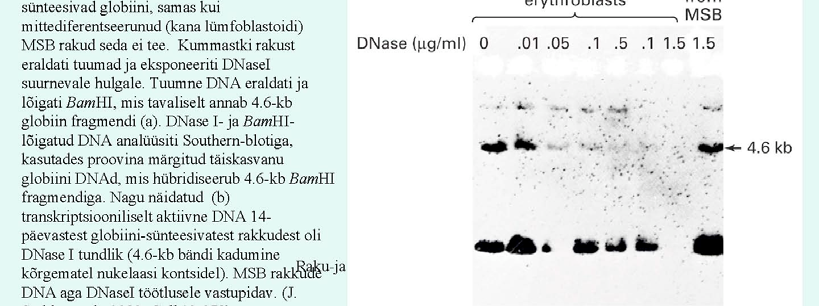
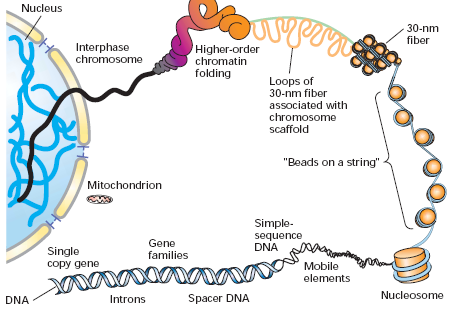
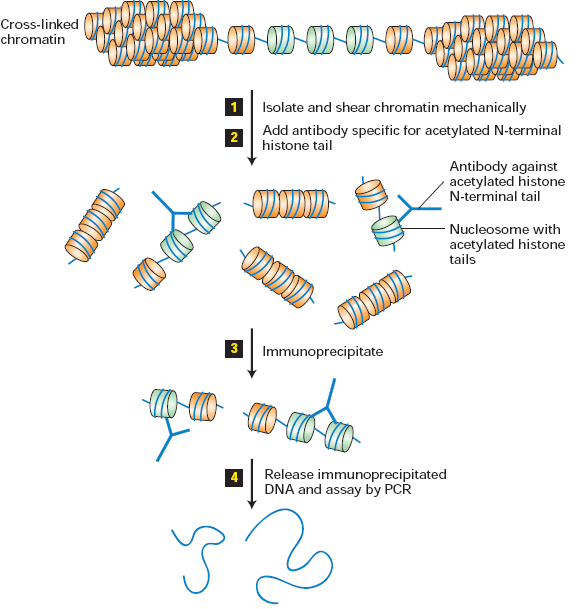




Kõik kommentaarid