1.
Transformatsioon - geneetilise informatsiooni ülekandumine
ühest bakterirakust teise rakku isoleeritud DNA abil.
Transformatsioon võib toimuda ka looduslikes tingimustes. Sel juhul
kandub elusrakkudesse surnud rakkudest vabanenud DNA.
Transkriptsioon – ümberkirjutamine, DNA ühe ahela alusel
komplementaarse RNA molekuli süntees.
Translatsioon – mRNA põhjal ribosoomides valguahela
sünteesimine ehk lihtsamalt öeldes valgu süntees. (RNA alusel
valgu süntees tsütoplasmas paiknevatel ribosoomidel.)
Translatsiooniprotsess loob geneetilise koodi ehk vastavuse mRNAde
nukleotiidahelate ja valkude polüpeptiidahelate vahel.
2.
Initsiatsioon prokarüootidel: transkriptsioon algab
sellega, et protsessi läbiviiv ensüüm RNA polümeraas kinnitub
struktuurgeenide ees
asuvas promootoris sisalduvatele
transkriptsiooni algussignaalidele. RNA polümeraas ei vaja
transkriptsiooni initsiatsiooniks praimerjärjestust nagu seda vajas
DNA polümeraas. Promootori piirkonnas, 10np transkriptsiooni
alguspunktist
eespool , on kindel järjestus – TATAAT -, mis on
tuntud Pribnow box’i nime all. Sellel järjestusel kinnitub RNA
polümeraas DNA-le ja oletatakse, et selles kohas avatakse DNA
kaksikahel nii, et DNA matriitsahelal oleks võimalik seostuda RNA
nukleotiidiga. Kuna pribnow box’i ala on A : T paaride rikas, on
seal kergem ahelaid denatureerida. Transkriptsiooni alguspunkt on
tähistatud ühega pürimidiinalustest, kas T või C-ga.
Initsiatsioon eukarüootidel: siin osalevad teatavad
promootorpiirkonna elemendid, kuhu seonduvad transkriptsioonifaktorid
ja mis osalevad ka regulatsioonil. Promootori elemendid: TATA, CAAT
ja GC box’id. Peale transkriptsioonifaktorite seondumist
promootorile, millest tähtsaim on TATA-box’i
seonduv valk TFIID
ehk TATA-seonduv valk,
seondub trans-faktoritele omakorda RNA
polümeraas ja initsieerib RNA sünteesi. Milline RNA polümeraas
tööle hakkab, sõltub sellest, mida transkribeeritakse. „
Enhancer “
– võimendavad järjestused, mis stimuleerivad transkriptsiooni ja
mida vajatakse geeni maksimaalseks transkriptsiooniks. Initsiatsiooni
lõpuks moodustavad trans-
faktorid , TATA-box, „enhancer“
seondunud valk ja RNA polümeraas koos transkriptsiooni
initsiatsiooni kompleksi.
3.
Sigma faktori (σ) kuulumine RNA polümeraasi koosseisu
võimaldab RNA polümeraasil spetsiifiliselt „ära tunda“ ja
seonduda promootoralale. Bakterirakus on mitmeid erinevaid sigma
faktoreid, mis võimaldavad RNA polümeraasil seonduda erinevatele
promootorjärjestustele. Transkriptsiooni initsiatsiooni lõppedes
vabaneb sigma faktor RNA polümeraasi koosseisust.
4.
Prokarüootne operoon – regulatoorne üksus, milles
asuvad struktuurgeenid,
operaator ja promootor. Operooni mudeli järgi
kontrollivad struktuurgeenide transkriptriooni regulaatorgeen ja
operaatorgeen. Laktoosi (lac) operoni
regulatsioon E.Coli rakkudes.
Lac operon sisaldab promootorit P, operaatorit O ja 3.
struktuurgeeni – lacZ (-galalaktosidaasi geen), lacY (-galalaktosidaasi
permeaasi geen) ja lacA (transatsetülaas). Operoni geenide
negatiivne kontroll toimub läbi repressorvalgu Lac I seondumise
operaatorile ja positiivne kontroll läbi aktivaatorvalgu CAP
seondumise operaatoralale.
5.
Eukarüootse mRNA kodeeriv osa erineb prokarüootsest selle
poolest, et sisaldab tüüpiliselt vaid üht tsistronit e. ORF’i.
Erineva struktuuri tõttu ei ole mRNA eukarüootide ja prokarüootide
vahel vahetatav, st. prokarüootne mRNA ei tööta eukarüootses
valgusünteesis ja vastupidi, eukarüootne mRNA ei ole
substraadiks prokarüootsetele ribosoomidele.
6.
Eukarüootides: RNA pI – koosneb 14-st subühikust ning
asub tuumakeses, vastutab rRNA-de sünteesi eest. RNA pII – koosneb
12-st subühikust ning viib läbi mRNA ja osade
snRNA -de sünteesi.
RNA pIII – koosneb 17-st subühikust ning katalüüsib tRNA, osade
snRNA-de ja 5SrRNA sünteesi.
Prokarüootides: RNA polümeraas – ensüüm, mis sünteesib
DNA
ahelale komplementaarse RNA ahela. Toimub transkriptsioon.
Transkriptsiooni initsiatsiooniks seondub RNA polümeraas
spetsiifiliselt promootorjärjestusega ning seejärel
katkevad promootrorpiirkonnas DNA
ahelate vahelised
vesiniksidemed . Ei vaja
initsiatsiooniks praimerit.
7.
TBP on esimene valk, mis ”istub” TATAbox promootorile,
TATA-siduv
proteiin .
Temal on oluline roll RNAPolII-katalüüsitud
snRNAde geenide transkriptsioonis. TBP seondub DNA väiksese valku,
rikub DNA normaalse dupleksi struktuuri, koolutades/väänates DNAd
oluliselt.
9.
eIF1 (15 kDa) koos eIF1A-ga (16 kDa) on vajalik 43S-i
skaneerimiseks mRNA-l.
eIF2 (koosneb 3 valgust: a - 35 kDa, b - 38 kDa ja g -52 kDa).
a alaühiku fosforüleerimine inhibeerib translatsiooni initsatsiooni
peatades eIF2-GTP kompleksi tekke. See fosforüleerimine kutsutakse
esile näiteks viirusinfektsiooni korral, et pidurdada viiruse
elutsüklit rakus. b ja g alaühik osalevad Met-tRNAi ja GTP
sidumisel. Seega on eIF2 bakteri IF2
analoog .
eIF2B (5 alaühikut). Katalüüsib G nukleotiidi vahetust
eIF2-l.
eIF3 (8-10 alaühikut, 650 kDa). Seob ribosoomi 40S alaühikut,
ei lase sel seonduda 60S-ga. Seob ka eIF4G-d.
eIF4A (25 kDa). RNA
helikaas , nõrgendab mRNA
sekundaarstruktuuri ATP hüdrolüüsist sõltuvalt.
eIF4B (80 kDa). Stimuleerib eIF4A helikaasset aktiivsust,
seostub mRNA-ga.
eIF4E (24 kDa). Seob mRNA 5´cap struktuuri. eIF4E on
translatsiooni initsatsioonil limiteeriv faktor. Üle selle valgu
toimub translatsiooni
aktiveerimine vastusena insuliinile või
kasvufaktoritele, viirusinfektsioonil aga võib üle eIF4E toimuda ka
cap-sõltuva valgusünteesi inhibitsioon.
eIF4G (220 kDa).
Molekulaarne adaptor, seob eIF4E, eIF4A,
eIF3, Pab1p.
eIF5 (125 kDa). Ribosoomist sõltuv GTPaas, soodustab
ribosoomi alaühikute ühinemist.
eIF6 (25 kDa) Seob 60S alaühikut, põhiline
antiassotsiatsioonifaktor.
10. TFIIH helikaasne aktiivsus saab oma energia
ATP hüdrolüüsilt.
Transkriptsiooni-initsiatsiooni kompleksi moodustumine.
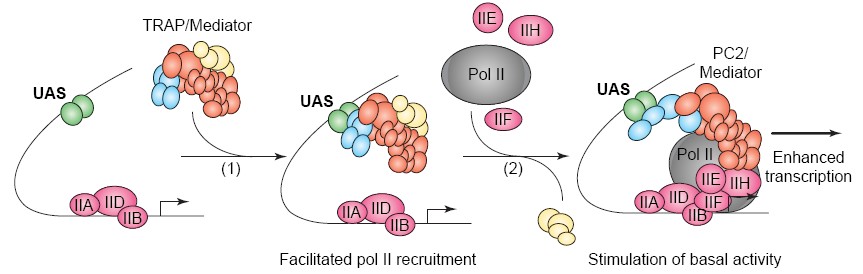
11.
Mediaator PILT on suur valgukompleks, koosneb 20-30
subühikust. Mediaator on samuti väga sageli transkriptsiooni
aktivaatorite sihtmärgiks: püüdes kinni Mediaatori, saab
promootorile tuua ka RNA polümeraasi. Mediaator ongi vahelüli
transkriptsiooni aktivaatorite ja RNA polümeraasi vahel.
PolII -
koosneb 12-st subühikust (Rpb1-12), millest suurimad on Rpb1 ja
Rpb2.
12
. PolI -sõltuv transkriptsioon: koos seonduvad UBF (
upstream binding factor ) ja SL1 (
promoter selectivity factor). SL1
koosneb TBP-st ja kolmest TAF-ist. TIFIA (
transcription
intermediary factor) seondub SL1 ja Pol I-ga. Pol I seondub ka
veel vähemalt kahe faktoriga TIFIC ja TFIIH.
PolII-sõltuv transkriptsioon: vajalik fakrot - TFIID. Pol
I ja Pol III promootoritel moodustub PIC üldjoontes sarnaselt Pol II
promootoritega, kuid erinevaid faktoreid on vähem.
PolIII -sõltuv transkriptsioon: vajalik fakrot - TFIIIC.
promootorid on reeglina transkribeeritava ala sees. Promootorit
tunneb ära TFIIIC (5 subühikut), mis toob kohale TFIIIB (koosneb
TBP-st ja kahest TAF-ist) TFIIIB toob kohale Pol III.
13.
Transkriptsioon - on RNA molekuli süntees, mis toimub
rakutuumas interfaasi ajal + tsütoplasmas. Transkriptsioonifaktorid
paiknevad tuumas.
14.
Sidumissait – on piirkond proteiinil, DNAl või RNAl,
mille külge teised
spetsiifilised molekulid ja
ioonid moodustavad
keemilist sidet.
TATA box - TATA järjestus, paljude
eukarüootsete geenide promootoralas olev konsensusjärjestus, mis
seob üldist transkriptsioonifaktorit ja määrab seega ära
transkriptsiooni alguskoha.
15. „
Enhancer“ ehk võimendav järjestus, mis stimuleerib
transkriptsiooni ja mida vajatakse geeni maksimaalseks
transkriptsiooniks. Paljude eukarüootsete
promootorite aktiivsust mõjutavad need oluliselt järjestused, mis
asuvad nende geenide proksimaalsest promootorist tuhandete
aluspaaride kaugusel. Nende piirkondade asukoht struktuurgeeni
suhtes pole tähtis ja nende funktsioon ei sõltu nende
orientatsioonist geeni suhtes. Üldiselt asuvad promootori ees,
pikkuseks on 50-150 np, mis omakorda koosnevad väiksematest
alaosadest 18-20 np. Neid esineb sagedamini kui vaigistavaid
piirkondi. Omadused: orienteeritud DNA
molekulil igas suunas
(5’-3’ või vastupidi); seostub ühe või mitu regulatoorste
valkudega; paiknevad enne või pärast geeni, mille ekspressiooni nad
reguleerivad.
16.
„Enhancer“ite aktiivsusega kontrollitakse geen
ekspressiooni tasemeid:fosfolüürimine, ubikvineerimine,
metüleerimine.
18.
heat - shock geenide
(näit hsp70) transkriptsioonil, peatub RNAPolII pärast esimese 25bp
sünteesi, aga ei termineeri transkriptsiooni. Peatunud polümeraas
jääb vastsünteesitud transkripti ja DNA matriitsiga seotuks
kuni tingimuste avaldumiseni, mis indutseeruvad HSTF (
heat-shock
transkriptsiooni faktoreid).
Aktiveeritud HTSFi sidumine proksimaalsepromootori teatud
järjestustele stimuleerib peatunud RNAPolII-i jätkama ahela
elongatsiooni ning indutseerib kiiret re-initsiatsiooni uute RNAPolII
molekulide seondumisega.
19.
Kasutades LacZ
reporterit pärmides
S.cerevisiae
ja
Z. bailii
selgus, et näiteks nende kahe erineva pärmi
TPI
geeni (
kodeerib glükolüüsivalku trioosfosfaadi isomeraasi)
promootorid olid
funktsionaalsed mõlemas pärmis, kuid reportergeeni
ekspressioonitase neilt oli mõnevõrra erinev.
Transgeenne analüüs – analüüs, mille jooksul
geenide rühma viiakse üle ühe organismi
DNAst teisse.
20.
Transkriptsioonilise
aktivaatori funktsionaalsed domäänid:
N-
terminaalne DNAd siduv domään,
mis seondub teatud DNA järjestustele,
ja C-terminaalne
aktivatsiooni domään,
mis interaktsioonis teiste valkudega
aktiveerib transkriptsiooni.
21.
Peamised DNAd siduvad domäänid
heeliks -pööre-heeliks (
helix -turn-helix),
tsink -sõrm-
valgud (zinc
finger ),
leutsiin -lukud (leucine zipper),
aluselised heeliks-
ling -heeliks (
bHLH ) domäänid, homeodomään
valgud .
23.
Histoondeatsetülaas
( HDAC ) – ensüümide klass, mis
eemaldavad atsetüül rühmasid -N-atsetüüli lüsiini
aminohappest histoonil.
Histoonatsetülaas (HAT)
– ensüümid, mis atsetüleerivad kaitstud lüsiini
aminohappeid histooni proteiinidel transfeerides atsetüül rühmad astüül
CoA-st lüsiini -N-atsetüüli lüsiinist, funktsioneerib üle
terve ensüümi. Histoonide atsetüülimine on seotud DNA
transkriptsiooni aktivatsiooniga. HAT seostub eukromatiiniga siis,
kui
viimasele seostub transkriptsioonifaktor. Järgmine histoonide
atsetüülimine toob nende peale negatiivse laengu, mis tekitab
kõrvaletõukamist histoonide vahel. Selle tulemusena kinnine enne
seda DNA muutub liigipäästvaks ensüümide jaoks.
24. Viimasel ajal on üha rohkem andmeid, et
“histoonne kood”
kontrollib
kromatiini kondensatsiooni, mitte üksnes histoonide
atsetüleerituse aste. “Histoonne kood” tähistab kõiki
histoonide N-terminuses toimunud modifikatsioone. Näit,
heterokromatiinses vormis on H3 Lys positsioonis 9 sageli
metüleeritud. “Histoonset koodi”
lugevad valgud
tunnevad neid spetsiifilisi modifikatsioone ära ja
promoteerivad kromatiini kondensatsiooni (suletud) või
dekondensatsiooni (avatud struktuuri teket).
26.
Nukleosoom
- DNA pakkimise väikseim „ühik“, mis koosneb 8-st histooni
molekulist ja 147-st aluspaarist DNA-st. Nukleosoomi tuumiku
moodustavad
histoonid H3, H4, H2A ja H2B, mida kõiki on kahes
korduses.
Nendest H3 ja H4 moodustavad tetrameeri (2 x H3-H4 dimeer),
millele omakorda kinnituvad kaks H2A-H2B dimeeri. Oli teada, et
selgroogsetes sisaldab transkriptsiooniliselt inaktiivne DNA
tihtipeale metüleeritud tsütidiinijääke (
5’metüültsütidiin,
mC), millele vahetult järgneb G.
Transkriptsiooniliselt aktiivses DNAs mC jäägid puuduvad. On
näidatud, et DNA mC vormis seob spetsiifiliselt valku, mis
interakteerub mSin3ga. See avastus
viitab võimalusele, et mSin3e
sisaldavad ko-repressorid on kompleksis mC valkudega.
28.
Kuidas kromatiin mõjub transkriptsioonile?
Elektronmikroskoopiliselt on näidatud, et
nukleosoomne struktuur on DNA-l nii vahetult enne kui ka kohe pärast mingi lõigu
transkribeerimist. Tõenäoliselt toimub mingi ajutine
konformatsiooniline muutus, mille tulemusel side histoonide ja DNA
vahel nõrgeneb ning transkriptsioonifaktorite seondumine saab
võimalikuks. Lihtsustatud mudeli järgi struktuursed valgud ja
transkriptsioonifaktorid konkureerivad DNA-ga seostumise suhtes,
kusjuures struktuursed valgud takistavad nendega seoses oleva DNA
kättesaadavust transkriptsiooni masinavärgile. Katsed näitavad, et
kui näit. geeni promootorpiirkond on nukleosoomi koostises, siis on
transkriptsioon kõvasti alla surutud. Kui in
vitro tingimustes
lisada DNA-le histoone, et moodustuks nukleosoomne struktuur,
surutakse transkriptsioon alla. Eriti tugevalt mõjub H1 lisamine.
Aktiivselt transkribeeritavad
geenid on nõrgalt seotud H1-ga või
see puudub hoopis.
30.
ChIP – immunosadestamismeetod -
usutav meetod kui
proteiin seob spetsiifilisse genoomi piirkonda
in vivo .
Elurakud on töödeldud sillutamise agendiga, et formeeruda
mittespetsiifilisi sillutamisi DNA ja seostavate proteiinide vahel.
Peale kromatiini pügamise väiksteseks fragmentideks, eelis proteiin
on imunosadestav. Iga proteiin-seov DNA on pärast analüüsitav
kvantitatiivse PCR, kloneerimise, järjestuse või hübridisatsiooniga
mikroridadel.
31.
Kromatiin on kromosoomi
koostisaine , mis koosneb peamiselt
DNA-st ja valkudest (RNA’st ka). Valgud võivad olla aluselised
(histoonid -H1, H2a, H2b, H3, H4- ja protamiinid spermides) või
happelised (mittehistoonsed valgud).
32.
FISH analüüs (Fluorescent In Situ Hybridisation
)
- kromosoomiaberratsiooni määramine. Interfaasi FISH analüüs on
kiirmeetod, millega on võimalik diagnoosida 13., 18., 21., X ja Y
kromosoomi arvu anomaaliat või uurida kromosomaalseid
ümberpaigutamisi. Omadused: Inimese
telomeerid ; In situ
hybridization; Fluorescen probes; TTAGGG lokaliseerimine.
33.
Matriksi kinnitumise piirkond (MAR) – immunoglobuliini
sõltuv funktsioon, alad genoomsel DNAl, mis interakteerub tuuma
matriksiga. Funktsioonid: transkriptsiooni/replikatsiooni domäänide
definitsioon; ex. betaglobiini (LCR) SCS/SCS’ mudeli süsteemid
Drosophila Heat Shock Locus-s; reguleerib transkriptsiooni,
kromatiini struktuuri, geeni ekspressiooni. Need elemendid
moodustavad DNA pidepunkti kromatiini tellingute jaoks ja sobivad,
selleks et oragniseerida kromatiin struktuurseteks domäänideks.
34.
Eukarüootse mRNA molekuliga toimuvad
muutusi pärast transkriptsiooni lõppu.
Eukarüootides sünteesitakse mRNA tuumas, translatsioon
toimub aga tsütoplasmas, seega on transkriptsioon ja translatsioon
ruumiliselt lahutatud. Eukarüootne mRNA läbib reeglina enne
tsütoplasmasse jõudmist “protsessingu”, mille käigus
primaarsest transkriptist eraldatakse
intronid ja lisatakse 5’ otsa
cap struktuur ning 3’ otsa lisatakse
poly (A) järjestus. Peale
protsessingut transporditakse küps mRNA läbi tuumapooride
tsütoplasmasse. Nii “cap” kui polü(A) on lisaks mRNA
stabiilsusele ja transpordile vajalikud ka mRNA seondumiseks
eukarüootsete intisiatsioonifaktoritega ja soodustavad väga
tugevalt translatsiooni initsatsiooni.
35. Introneid lõigatakse välja
pre-mRNA transkriptidest
kolmeetapiliselt splaisosoomide abil.
Splaisosoomid –
organellid raku tuumas, mis koosnevad valkudest ja snRNA-st. 1
etap –
introni 5’ otsa lahtilõikamine, mille tagajärjel introni ees olev
ekson lahutatakse ülejäänud mRNA järjestusest. Introni 5’ ots
keeratakse aasana tagasi ja ühendatakse introni sees olevasse
kindlasse järjestusse nukleotiidiga A, mis asub 18-38 nukleotiidi
introni algusest 3’ suunas. 2 etap – introni 3’ otsa
lahtilõikamine, mille tulemusena
intron vabaneb pre mRNA
koosseisust. 3 etap – allesjäänud eksonite alad ühendatakse
ligaaside abil ning tekib ainult eksonitest koosnev mRNA
molekul .
Intronid degradeeritakse.
36.
U snRNAd osalevad pre-mRNA splaisingu regulatsioonis: 5
snRNPd: U1, U2, U4, U5, U6.
37.
GT-AT reegel –
on selline
vaatlus , et kõik intronid DNAs algavad GT nukleotiitudega
(
guaniin , tümiin) ja lõppevad AG nukleotiitidega (
adeniin ,
guaniin). Kui DNA on transkribeeritud RNAsse, intronid on eemaldatud
RNAst mehaniismi abil, mis tunneb ära neid
nukleotiitide alguseid ja
lõppe – RNAs nad oleksid CU (tsütosiin,
uratsiil ) ja AC (adeniin,
tsütosiin).
39.
RNA
protsessimine. Protsess nimega „RNA
editing“ muudab transkriptis sisalduva info hulka, muutes
aluspaaride järjestust asenduste teel. Algset staadiumi
RNA
protsessingus katalüüsib dimeerne
capping ensüüm, mis seostub RNAPolII fosforüülitud CTDga.
Viimases
staadiumis kannavad erinevad ensüümsed alaühikud üle
metüülrühmad S¬-adenosüülmetioniinilt guaniini asendis N7
olevale lämmastikule ning sünteesitud RNA 5’otsas paiknevate
ribooside 2’oksügeenidele.
Muudab
CAA koodoni UAA poolt kodeeritud Stop koodoniks ning lõpeb poole
lühema valgu apoB-48 sünteesiga.
40. Vaid
puuviljakärbse
emasisendid sünteesivad
funktsionaalset SXL (sex lethal) valku, mis represseerib eksonite 2
ja 3 splaisingut sxl pre-
mRNAs ja eksonite 1 ja 2 splaisingut tra
(transformer) pre-mRNAs. TRA-
TRA2 kompleksi sidumine dsx (
double -sex)
pre-mRNAle aktiveerib eksonite 3 ja 4 splaisingu. Sellise
reguleeritud splaisingute kaskaadi tulemusena sünteesitakse
puuviljakärbse emas-ja
isas -embrüotes erinevad DSX valke. Need
represseerivad vastassooliseks seksuaalseks diferentseerumiseks
vajalike geenide ekspressiooni. DSX isas-vorm represseerib
naissooliseks diferentseerumiseks vajalike geenide ekspressiooni ja
vastupidi.
44.
RNA-sõltuv
vaigistamine on loomupäraselt
lühiaegne (transient).
miRNA
(ehk microRNA) on RNA tüüp, mis on seotud geenide aktiivsuse
reguleerimisega. Tema omadused: mRNA molekuliga ühinevad
komplementaarselt; ei vaja vahendajaks regulaatorvalku; väiksed.
siRNA –
väikene
interferents RNA, siRNA
ahelad , mis on 21-23 bp pikad,
hübridiseeruvad üksteisega nii, et 3’otsmised 2 nukleotiidi on
üksikahelalised. siRNAde tekkeks on vaja Dicer kompleksi
ribonukleaasset aktiivsust. Sellest järeldati, et miRNAde ja siRNAde
vahendatud protsessid on väga sarnased.
RNA-indutseeritud
vaigistav kompleks ( RISC ) lõikab
märklaud mRNA, mis on täpselt komplementaarne vastavale
üksikahelalisele siRNAle, ahela katki. RISC kompleks funktsioneerib
ka kui translatsiooni inhibiitorid. Dicer samuti funktsioneerib
Droshast
allavoolu , et produtseerida täiskasvanud miRNA-d, mis nagu
siRNA, on sulatatud geeni vaigistavatesse kompleksidesse nimega
RNA-indutseeritud vaigistavad
kompleksid .
45.
Nonsense mediated decay – ühe või enam
eksoni vahele
jätmine, mis põhjustab ekson-intron
liidese vahetus 3’ läheduses
Stop
koodoni sissetuleku, mis kuulub mRNA järelvalve mehanismide hulka.
Ta
vahendab kiiret mRNAde lgundamist, kus Stop
koodonid esinevad
mRNAs enne viimast splaisiliidest.
46.
Inteiin
–
proteiini segment, mis on võimeline eemaldama ennast ja
taasühenema eksteiini peptiidsidemega. Nad on väga võimekas
proteiini splisingus.
Intron
on eukarüootse geeni
mittekodeeriv piirkond, mis transkribeeritakse
RNA molekuliks, ent seejärel lõigatakse sealt splaisingu tulemusena
välja.Vahe on selles, et intronid lõigatakse välja ja
eksonid (inteiinid) liidetakse kokku.
47. Peaagu kõik mRNAd sisaldavad
järjestust
AUAAA, mis asub polü(A) sabast 10-35bp
ülavoolu. Kui see AUAAA ära muteerida, siis RNA transkriptide
polüadenüülimist peaagu ei esine. Muteerimisel need polü(A)-sabata
transkriptid kogunevad tuuma ning suunatakse lagundamisele. 360kDa
lõikamis-ja polüadenüülimis-spetsiifiline faktor (CPSF), mis
koosneb neljast
erinevast valgust, moodustab kõigepealt AAUAAA
järjestusest ülavoolu
seonduva ebastabiilse kompleksi. Seejärel
seonduvad sellele CPSF-RNA kompleksile vähemalt kolm valku: 1) 200
kDA heterotrimeer, mida nim lõikamist-stimuleerivaks faktoriks
(CStF), mis interakteerub G/U rikka järjestusega; 2) 150kD
lõikamisfaktor I (CFI) ja halvasti kirjeldatud lõikamisfaktor II
(CFII); 3) polü(A) polümeraas (PAP), mis seondub kompleksile
vahetult enne lõikamist. Tehtud on
suuremaid uuringuid, kus CTD-s
saavutati konkreetse aminohappe väljalöömine. Tulemused näitavad,
et RNA plümeraas II CTD kärpimise
mutatsioonid mõjutavad võimet
indutseerida in vivos geenide alamrühma transkriptsiooni ja nende
geenide tundlikkuse
puudulikkust induktsiooni ühendustele(kaartidele)
ülesvoolu aktiveerivatele järjestustele.
48.
mRNA
polüadenüleerimise on oluline
3'-
terminal exoni määramiseks
imetaja geenis, on
soovitatud ,
selleks et
virtuaalselt mõjutada kõike mRNA metabolismi
aspekte .
Tähtis sellepärast et mängib rolli raku-tsükli-reguleeritud
histoonide mRNA, snRNA, tRNA küpsemises; transpordib mRNA tuumast
tsütoplasmasse; parandab DNAd ja kromosoomi eraldamist.
49.
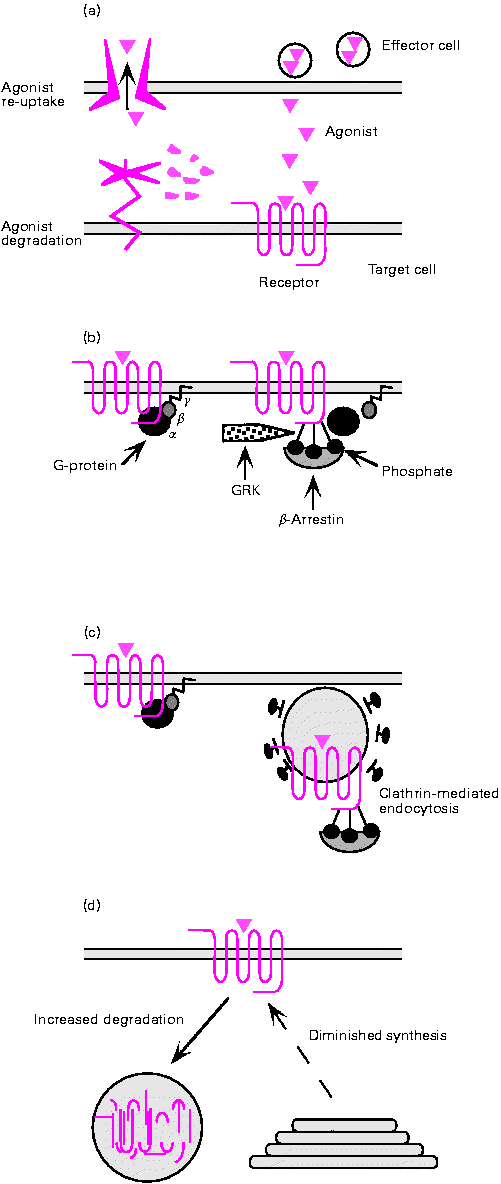
Mechanism of attenuation PILT
1. Agonistide eemaldamine lahjendamise teel, neelamine transporter
proteiinide ensüümilise degradeerimise poolt on esimene samm
signaali lõpetamises. 2. Aktiveeritud retseptorite fosforülatsioon
GRK-de poolt ja järgnev retseptori interaktsioon "arrestiinidega"
keelab
interaktsiooni G-proteiinidega ja efektiivselt lõpetab
signaali põhjustades kiire tundetuks muutumise. 3.
Agonist-indutseeritud endotsütoos vähendab kõrg-vastastikuse
tõmbega retseptorite plasma membraane, ja võib esindada teist
tundetuks muutumise faasi. 4. Alla-regulatsioon esineb rakkude
pikendatud ekspositsiooni jooksul agonistidele, ja võib olla
vahendatud
suurendatud degradeerimise ja retseptorite kahanenud
sünteesi poolt.
50.
Mehanismi , mille abil tuumsest pre-mRNAst kõrvaldatakse
intronid. Intronite väljalõikamine võib toimuda kolme erineva
mehhanismi alusel:
(1)
tRNA prekursorite puhul teeb katked RNA ahelasse
spetsiifiline splaissingu endonukleaas ning eksoneid sisaldavad RNA
segmendid ühendatakse splaissingu
ligaasi abil. Need ensüümid
tunnevad spetsiifiliselt ära tRNA prekursormolekuli kõrgemat järku
struktuuri, mitte aga spetsiifilist nukleotiidset järjestust.
(2)
Osade rRNA prekursorite puhul (paljudes madalamates
eukaüootides, näit. Tetrahymena thermophila samuti ka
rakuorganellides) kõrvaldatakse intronid autokatalüütiliselt, RNA
molekuli enda poolt. Splaissingureaktsioon ei vaja välist
energiaallikat ega valkude aktiivsust. Kofaktorina on vaja vaba 3´-OH
rühma kas GTP-lt, GDP-lt, GMP-lt või guanosiinilt (G-3´-OH) ning
monovalentset ja divalentset
katiooni .
Splaissing toimub
etapiviisiliselt: kõigepealt toimub fosfodiestersideme ülekanne
ekson-intron ühendusalalt G-OH-le (RNA ahela katkeb eksoni ja
introni ühendusalas), seejärel katkeb fosfodiesterside järgmise
eksoni ja introni 3´-otsa vahel ning fosfodiesterside moodustub
eksonite vahel. Väljälõigatud intron tsirkulariseerub
molekulisiseselt (toimub veel üks fosfodiestersideme ülekanne).
(3)
Rakutuumas asuvate pre-mRNA molekulide splaissing
toimub kahe-etapiliselt ribonukleoproteiin-partiklites –
splaissosoomides. Splaissosoomid sisaldavad snRNA molekule U1-U6 (U3
asub tuumakeses) ja üle 40 erineva valgu. snRNA-d ei ole tumas
vabalt, vaid kuuluvad väikestesse RNA-valk kompleksidesse, mida
nimetatakse snRNP-
deks (small
nuclear ribonucleoproteins). Splaissing
toimub etapiviisiliselt.
Esmalt katkestatakse fosfodiesterside
introni 5´-splaissingu saidis introni GU järjestusest 5´-suunas.
Protsessis osaleb kogu splaissosoom. Splaissingu saiti seondub
otseselt U1 snRNP. Fosfodiesterside moodustub introni 5´-otsa ja
konserveerunud A nukleotiidi vahel introni 3´-otsa lähedal.
Seejärel seondub U2 snRNP introni splaissingu 3´-saiti, kus
konserveerunud A
nukleotiid on ühenduses introni 5´-otsaga
(selliseid struktuure nimetatakse lariaatideks). Kompleksile
lisanduvad ka teised snRNP-d, et moodustuks täielik splaissosoom
ning seejärel lõigatakse intron 3´- splaissingusaidist välja
(fosfodiestersideme lõhkumine) ja eksonite vahel moodustub
fosfodiesterside.
51.
Post-transkriptsioonilise regulatsioon tsütoplasmas.
Eukarüootsete geenide esmased primaarsed transkriptid
protsessitakse, et tekiks funktsionaalne RNA ning see transporditakse
ribonukleopartiklitena tuumast tsütoplasmasse. Nii RNA protsessimine
kui nende
eksport tuumast on veel omakorda allutatud
regulatsioonilisele kontrollile. Lisaks toimuvad kontrollimehanismid
ka tsütoplasmas. Nii näiteks sõltub valgu hulk rakus suuresti
tsütoplasmaatilise RNA stabiilsusest (lagunemiskiirusest). Samas on
mõnede rakuliste mRNAde asukoht tsütoplasmas reguleeritud, nii et
uut valku sünteesitakse just selles raku osas (näit. närviraku
dendriitides).
52.
Kõik tuumakeses sünteesitavad RNA tüübid. Tuumake on
osa rakutuumast, mis on spetsialiseerunud rRNA sünteesiks ja rRNA
assambleerimiseks ribosoomidesse. Eukarüootides sünteesitakse rRNA
tuumakeses RNA polümeraasi I poolt. Valgusünteesiks
(translatsiooniks) vajatakse kolme komplementaarse funktsiooniga
nukleiinhapet: mRNA kannab koodonitena informatsiooni valgu
primaarjärjestuse kohta; tRNA identifitseerib koodi ja toob
ribosoomi koodile vastava aminohappe ; rRNA assotsieerub ribosoomi
valkudega moodustades ensüümkompleksi, mis otseselt sünteesib
peptiidsideme.
53.
miRNA ja siRNA vahe tuleneb nende päritolust: siRNA
pärineb dsRNA-st; siRNA on kõige tavalisemalt vastus võõrale
RNA-le ja on tihti 100% täiendav(komplementaarne) sihtmärgile.
miRNA pärineb
ssRNA -st, mis moodustab U-kujulise sekundaarse
struktuuri; miRNA reguleerib post-transkriptsionaalset geeni
väljendust ja pole sageli 100 täiendav(komplementaarne)
sihtmärgile.
54.
hnRNP A1 tuumast ekspordi protsess: Lõplikult
protsessitud mRNA-5’ otsa ning hnRNP valk-kompleks (
mRNP)
assotseerub “
cap”-siduva
kompleksiga (
CBC),
mis läbib esmalt NPC (
tuumapoori-kompleks).
Kui mRNPsid transporditakse läbi NPC, siis tuumas lokaliseeruvad
hnRNPd eemaldatakse; need valgud, milledel pole NESi (
nuclear
export signal , tuumaekspordi signaali),
hoiavad mRNAd tuumas. hnRNP A1 transporditakse läbi NPC ning nad
kannavad assotseeritud RNA tsütoplasmasse.
Tsütoplasmaatiline
RanGAP stimuleerib Rani GTPd hüdrolüüsima.
Shuttle ’vad
hnRNP valgud dissotseeruvad retseptorvalkudelt (leutsiin-rikaste
NESide puhul eksportiinilt) ning nad transporditakse tagasi tuuma.
mRNA seondub nüüd tsütosoolis asuvate mRNP valkudega, nende hulgas
polü(A)ga-seonduva valgu PABPga, mis interakteerub mRNA 3’ otsaga.
55.
Genoom
– on ühes liigiomases kromosoomikomplektis (haploidne
kromosoomistik) sisalduv geneetiline materjal. Kõikide
geenide kogum.
Proteoom –
organismis sisalduvate valkude kogum, kodeeritud raku, koe,
organismi genoomi poolt. Erinevalt genoomist: proteoom on erinev
organismi erinevates rakkudes; proteoom muutub pidevalt
Transkriptoom –
kõigi mRNA molekulide või "transkriptide" kogum, mis on
toodetud ühes rakus või terves populatsioonis rakkudes.
Reguloom –
geenide kogum, mida reguleeritakse sama regulaatorproteiini poolt.
See mõiste on üldiselt kasutatav prokarüootsete süsteemide jaoks,
näiteks „quorum“ tundlik bakteris.

























Kõik kommentaarid