BIOFÜÜSIKA ERIOSA Konspekti koostamisel on kasutatud loengumaterjale, Silverthorni „Human physiology“,
Sartoriuse „Biofüüsika“, mõmmi konspekti ja internetis leiduvat materjali.s 24) Bioloogiliste membraanide struktuur.
Membraanid moodustavad 80% loomsete rakkude kuivkaalust. Rakumembraani paksus on
umbes 8nm.
1972 Singer-Nicolsoni mudel, mille kohaselt
fosfolipiidid on kaksikkihis(seda teati juba
varem) ning lisaks on nende vahel
valgud , mis on võimelised ringi liikuma.
Demonstreerimiseks liideti inimese ja
hiire rakud - algul olid hiire valgud ühel pool
rakku ja inimese omad teisel pool, kuid 40 min pärast olid valgud ühtlaselt jaotunud.
Ka
lipiidid saavad ühe lipiidikihi piires üsna vabalt
liikuda , kuid vertikaalne „flip-
flop “ liikumine on väga aeglane.Valgud võivad
ulatuda läbi kogu membraani või
kinnitada sisse- või väljapoole. Funktsioonid on
struktuuri andmine-
ühendavad membraani tsütoskeletiga
moodustavad rakuliiduseid
kinnitavad rakud ekstratsellulaarse maatriksi külge
retseptoriks olemine .2 tüüpi
teiste rakkude ära
tundmine keemiliste signaalide äratundmine
transporteriks olemine carriers-
glükoos ja
aminohapped channels-vesi ja
ioonid . Na+ ja K+ läbi mõlema
ensümaatiline funktsioon- nt
peensooles peptiidide ja süsivesikute lagundamine.
Fosfolipiidide hüdrofiilsed pead on suunatud väljapoole ning
hüdrofoobsed sabad
sissepoole.Sellised ained on AMFIFIILID. Neil on glütseroolist „
selgroog “, mille
küljes kaks rasvhappe molekuli ning üks
fosfaatrühm . Pea- fostaatrühm+glütserool.
Saba-
rasvhapped .
Lisaks
fosfolipiididele leidub membraanis teisigi lipiide- glükolipiidid nt TSERAMIID ja
ka suuremas osas
hüdrofoobne kolesterool, mis aitab teha membraani läbipääsmatuks
väikestele veeslahustuvatele molekulidele, ka veele, ning hoiab membraani
painduvana. Valkude ja lipiidide suhe on eri membraanides erineb.
Plasmamembraanides 1, müeliinis 0,32 ja mitkondrite sisemembraanides 3,2. Mida
rohkem valku, seda aktiivsema metabolismiga
rakk .
2
Membraanide ül: Tänu lipiidsele kaksikkihile saavad membraanid eristada erineva
1
0
keemilise koostisega regioone. Membraani läbivad
rasvlahustuvad molekulid.
ar 2
Membraanides endis toimuvad keemilised reaktsioonid. Membraanides olevad
u
TRANSPORTVALGUD ja
RETSEPTORVALGUD . Vahendavad suhtlemist eri
aan
rakkude vahel, ka kudede moodustumist.
a, j
n
iiLi-
aal
M
1
Ainete transport läbi bioloogiliste membraanide. Erinevate molekulide erinev võime membraani läbida oleneb nende keemilisest
struktuurist. Lipiidset kaksikkihti suudavad läbida vaid LIPIIDLAHUSTUVAD ained
ja GAASID.Paljud ioonid ei suuda praktiliselt üldse läbida. Ka polaarsed läbivad
halvasti, kuid vesi erandinda difundeerub hästi. Permeaablus vee suhtes o n 10-2 cm/s
ning Na+ suhtes kõigest 10-12 cm/s.
Membraan kui selektiivne barjäär ainete liikumisele. Erinevate molekulide erinev võime membraani läbida oleneb nende keemilisest
struktuurist- nende
suurusest ning
polaarsusest. Lipiidset kaksikkihti suudavad
läbida vaid LIPIIDLAHUSTUVAD ained ja GAASID.Paljud ioonid ei suuda
praktiliselt üldse läbida. Väikesed polaarsed molekulid saavad läbi?
Passiivne ja aktiivne ainete transport läbi bioloogilise membraani. Passiivse transpordi
põhivorm on
difusioon , passiivne transport
ei vaja lisaenergiat
kulgeb pärisuunas mööda kontsentratsioonigradienti (
gradient on suuruse muutumise
määr pikkusühiku kohta).
Passiivne transport võib olla lihtne või abistatud valkude poolt (nt glükoosi ja
aminohapete transport)- nimetatakse hõlbustatud
difusiooniks .
Aktiivne transport
vajab lisaenergiat ATP vormis
toimub vastu kontsentratsioonigradienti
põhiliselt toimub ioonpumpade vahendusel. (nt Ca2+
pump või Na+-K+ pump või
Na+-K+- ATP-aas,mis on antiport pump ja mille toimimise tagamisele läheb 30%
kogu raku energiast. 1 ATP molekuli energiast
piisab , et viia sisse 2 K+ ja välja 3
Na+).
PRIMAARSE AKTIIVSE TRANSPORDi jaoks vajalik energia tuleb kõrge energiaga
fosfaatsidemest ATP-st.
SEKUNDAARSES (e KOTRANSPORDIS) kasutatakse kontsentratsioonigradiendi
potentsiaalst energiat. See gradient tekkis primaarse aktiivse transpordi abil.
Liigutades üht molekuli päri tema kontsentratsioonigradienti, liigub teine vastu oma
kontsentratsioonigradienti. Eriti palju kasutatakse Na+ gradienti. Sümport-
transporditav
molekul (nt glükoos) ning kotranspordi
molekul (nt Na+) liiguvad samas
suunas, Antiport- molekulid liiguvad vastassuunas. Membraanis on kotranspordi jaoks
vastavad molekulid.
2
1
Ka endotsütoos, eksotsütoos ja fagotsütoos nõuavad energiat, kuid ei klassifitseeru
0
klassikalise aktiivse transpordi alla (see on vähemalt kolloidkeemia õppejõu väide).
ar 2
u
aan
Membraani valkudega seotud transpordimehhanismid: ioonkanalid ja a, j
transportervalgud. niiLi-
aal
M
2
Ioonkanalid on membraanivalgud, mille avatud olek võimaldab
ioonidel liikuda läbi
membraani. Kui
kanal on avatud, toimub liikumine mööda
elektrokeemilist gradienti.
On kaht tüüpi
kanaleid -
reguleeritud kanalid
o VOLTAAŽTUNDLIKUD, mis
avanevad , kui membraani polariseeritus e.
membraanipotentsiaal muutub o LIGANDTUNDLIKUD, mis avanevad, kui ligand seostub e. reguleeritakse
regulaatorainete e ligandite abil. o MEHAANILISELT REGULEERITUD
pidevalt avatud e. lekkekanalid.
Ioonkanalid on
suuremal või vähemal määral
spetsiifilised erinevatele ioonidele. Selektiivsuse
annavad kanalitele nende seinte
LAENGUD ja SIDUMISKOHAD.
Erand on nt
katioonikanalid, mis lasevad läbi kõik positiivsed ioonid.
Transportervalgud on ilmselt ioonpumbad, transpordivad osakesi vastu kontsentratsiooni
ja/või
elektrilist gradienti.
25) Difusioon, Ficki difusiooniseadus. Difusioon on molekulide liikumine kõrge
kontsentratsiooniga alalt madala
kontsentratsiooniga piirkonda soojusliikumise tõttu, kestab kontsentratsioonide
ühtlustumiseni. Difusioonil põhineb aine passiivne transport läbi membraani. Lahustes ja
gaasides liiguvad
aatomid ja molekulid vabalt, kontsentratsiooni erinevused tasandatakse
difusiooni teel. Vesilahustes on difusioon väikeste vahemaade korral enamike molekulide
puhul tähtsaim vahetusprotsess. Sama kehtib ka raku kohta seni, kuni difusiooni ei takista
membraanid.
Vesi ja lahustunud gaasid, samuti lipiidlahustuvad ained difundeeruvad läbi membraani
vabalt. Siiski ei ole plasmamembraan permeaabel mitte ainult nendele ainetele, vaid läbi selle
difundeeruvad ka ioonid, aminohapped,
suhkrud , nukleotiidid. Need ained läbivad membraani
pooride kaudu, mille moodustavad membraanis paiknevad transportvalgud.
Difundeeruva aine
mass ajas on võrdeline kihi pindala ning ainete sisalduste vahega lähte- ja
lõppkohas ning pöördvõrdeline kihi paksusega.
2
1
0
D-difusioonikonstant, mille väärtus on antud
aine,
ar 2
u
lahusti ja
temperatuuri puhul
konstantne .
aan
a, j
niiLi-
aal
M
3
C1-C2 tuleb negatiivne, sest vool lähtekoha suhtes ongi negatiivne.
Osmoos . Hüpo -, iso- ja hüpertoonilised lahused, rakkude käitumine nendes. Osmoos on difusiooni erijuht, kus
läbi poolläbilaskva membraani difundeeruvad lahusti
molekulid. Osmoos põhjustab lisarõhku piirkonnas,
kuhi molekulid hakkavad liikuma-
osmootne rõhk, tähis π.
Osmootne rõhk väljendatakse
van´t Hoffi valemiga = iRTC, kus i on
isotooniline koefitsent e siis ioonide arv, milleks aine dissotsieerub. R on universaalne
gaasikonstant , T on temperatuur kelvinites, C on
osmootselt aktiivse aine
kontsentratsioon.
Vereplasma osmootne rõhk on u 7,4 atm. 0,9% NaCl lahus on vereplasmaga isotooniline,
samuti 4-4,5% glükoosilahus.Vereplasma valkude kolloidosmootne e onkootne rõhk
on 20-30 mm Hg, onkootne rõhk aitab kaasa vee filtratsioonile kapillaarides.
Põhjustab vee liikumist vereringesüsteemi.
Toonilisus on suhteline suurus ning ühikuta, alati mingi konkreetse raku suhtes.
Hüpertoonilines lahuses on kõrgem osmootne rõhk, lahustunud aine
kontsentratsioon rakku ümbritsevas lahuses on suurem ning vesi liigub rakust
välja, et väliskeskkonda lahjendada, rakk kortsub (plasmolüüs).
Hüpotoonilises lahuses on osmootne rõhk väiksem ning lahustunud ainet vähem
kui rakus ning vesi liigub rakku- rakk
paisub ja võib lüüsuda.
Isotoonilises lahuses osmootsed
rõhud on võrdsed, raku ruumala ei muutu,
ioonide liikumine kahe keskkonna vahel on võrdne.
26) Membraaniga seotud bioelektrilised protsessid. Membraani aktiivsed ja passiivsed
elektrilised omadused. Membraani elektriline skeem. Ioonid kui bioloogilise elektri
materiaalsed kandjad. Elusat rakku iseloomustab
polariseeritud plasmamembraan, mille
sisepind on välispinna
suhtes negatiivselt laetud.
Lipiidne
kaksikkiht on
dielektriliste omadustega, mis tähendab, et teda saab välise välja
mõjul polariseerida. Membraanis esineb elektrivool ja takistus.
Ioonkanaleid läbivad ioonid genereerivad elektrivoolu.
Takistus muutub vastavalt
kanalite avatusele- mida rohkem avatud , seda väiksem
2
1
0
takistus.
ar 2
u
aan
a, j
niiLi-
aal
M
4
Membraanil on
Takistus harilikult umbes 1000
oomi /cm2 , kuid võib ulatuda 10 000 oom/cm2, kui enamik
ioonkanaleid on suletud. Osad kanalid käituvad takistitena, teised
alalditena(soodustavad ühesuunalist voolu)
Mahtuvus Mahtuvus tekib mittejuhtiva (
lipiid ) ja juhtiva kihi olemasolust. Membraani
elektriline mahtuvus on 1 mikrofarad/cm2
Pinge(potentsiaalide vahe). Jõud, mis tekib selle tõttu, et
laengud on ruumiliselt eraldatud.
Membraanipotentsiaali tekkel on oluline roll lekkekanalitel.
Membraani elektrilisest skeemist-
http://faculty.plattsburgh.edu/donald.slish/Basic.html 2
1
0
Aktiivsed nähtused membraanis on
ar 2
u
aktsioonipotentsiaal=tegevuspotentsiaal=
erutus =
närviimpulss =AP.
aan
Aktsioonipotentsiaal tekib närvi- ja lihasrakkudes, kui nende membraanipotentsiaal
a, j
häirub lühiajaliselt positiivses suunas.
niiLi-
aal
M
5
Passiivsed e ELEKTROTOONILISED nähtused (NB vaata edasi konspektis, eraldi
punkt) ilmnevad siis ,kui rakule toimitakse kestvama elektrivooluga, närvi mõjutatakse
väikese potentsiaalimuutusega. Kui membraani ainult veidi polariseerida, nii et see ei
ulatu läveni ja
pingest sõltuvad kanalid veel ei avane, tekib astmeline potentsiaal, mis
liigub küll mööda membraani edasi, kuid hakkab kustuma e. kaugemale liikudes
väheneb.
Iseloomustatakse kolme suurusega :
Sisendtakistus - määrab, kui palju potentsiaal muutub.
rakusisene
aksiaalne takistus- määrab, kui kaugele ja kui kiiresti impulss levib.
Tsütoplasma takistuse ja närvi diameetriga määratud.
mahtuvus- määrab, kui kiiresti pärast ärritust muutus tekib.
Membraanipotentsiaal ja AP levik omavad nii passiivsete kui aktiivsete nähtuste tunnuseid.
(NB!!AP puhul Na K liikumised alati päri kontsentratsioonigradienti)
27) Iooni tasakaalupotentsiaal, Nernsti võrrand. Iooni
tasakaalupotentsiaal on selline
membraani puhkepotentsiaal , mille tekitaks üht
tüüpi
ioon , kui membraan oleks läbilaskev vaid sellele
Lipiidne kaksikkiht e fosfolipiidne membraan on hea elektriinsulaator. Elektri- ja
kontsentratsioonigradient kokku on elektrokeemiline gradient. Rakk võib olla elektrilises
tasakaalus ,aga ei pruugi olla keemilises tasakaalus.
Iooni tasakaalupotentsiaali korra on iooni liikumine ühes suunas kontsentratsiooni
gradiendi tõttu täpselt sama suur sama iooni liikumisega vastupidises suunas elektrilise gradiendi tõttu.
Ioonide NETOliikumine saab nulliks. Kehtib vaid ioonide puhul- laenguta osakeste puhul
lõpeb netoliikumine (aine sisse- ja väljavoolu vahe), kui kontsentratsioonid võrdsustuvad.
Näide: Oletame, et K+ liigub rakust kontsentratsioonigradiendi tõttu välja. Ta viib kaasa ühe
postiivse laengu. Nüüd tekib raku
sisemusse negatiivne laeng, mis raskendab järgmiste K+
välja liikumist, see tahab hoopis sisse tagasi liikuda. Lõpuks tekib olukord, kus areneva
elektrivälja mõju kompenseerib kontsentratsioonide erinevuse poolt tekitatud
DIFUSIOONIRÕHU.
2
1
0
ar 2
u
aan
a, j
niiL
i-
aal
M
6
F- Faraday konstant. Z- reaktsioonis ülekantavate elektronide moolide arv
Ce-iooni ekstratsellulaarne kontsentratsioon. Ci- iooni
intratsellulaarne kontsentratsioon.
Membraani puhkepotentsiaal: olemus, suurus, tekkimise tingimused, Goldmann -
Hodgkin-Katzi võrrand. Gibbs -Donnani tasakaal. Olemus: Puhkepotentsiaal Vm on
elektrigradient ekstra - ja intratsellulaarse vedeliku
vahel. Tekib membraani polariseerituse tõttu.
Puhkepotentsiaali nimetus tuleneb sellest, et juhul, kui mingi mõjutus rakku ei aktiveeri,
püsib (närvi-ja lihas)rakkude membraanipotentsiaal pikka aega konstantsena.
Suurus: -100 kuni -30 mV ,määratud suures osas K+ ioonide poolt. Stabiilne seisund,
kus membraanipotentsiaal ei muutu. Esineb kõigis rakkudes. Intratsellulaarne vedelik
on negatiivselt laetud ja ekstratsellulaarne positiivselt, kuid ekstratsellulaarne
pannakse 0-punktiks ja intratsellulaarne on siis selle võrra negatiivsem.
Kui Na+ ja K+ puhul liiguks ainult üks ioon, siis ENa on +60mv, EK on -90 mv (e nende
ioonide tasakaalupotentsiaalid) ja Em e puhkepotentsiaal on -70mv, mis on lähemal
EK-le, sest membraan on K+ suhtes 40 korda läbilaskvam. Et EK ja Em ei kattu, on
tingitud Na+, mis teeb raku positiivsemaks.
Tekkimise tingimused:
Ebaühtlane laengute jaotus tuleneb kahest tegurist:
membraanide erinev läbilaskvus erinevatele ioonidele
ioonide aktiivne transport vastu kontsentratsioonigradienti. K+ liigub välja, aga Na+ ei
jõua tasakaalustada, sest membraan pole talle nii läbilaskev, tekib negatiivne laeng
raku sees ning membraanipotentsiaal.
Membraanipotentsiaali säilitavad:
membraani suur läbilaskvus K+ suhtes
ioonide ebaühtlane jaotus kahel pool membraani, mida hoitakse Na-Ka-pumbaga (Na-
K pump on elektrogeenne - 3Na välja ja 2 K sisse.
Gibbs-Donnani
efektid , mis tekivad rakusiseste suurte anioonide olemasolust
2
1
0
ar 2
u
aan
a, j
niiLi-
aal
M
7
Goldmann-Hodgkin-Katzi võrrand kirjeldab olukorda, kus membraan on läbilaskev
mitmetele ioonidele, mitte vaid ühele. Korraga liigub mitmeid ioone.
Gibbs-Donnani tasakaal- kui ühel pool mingit lahutuskihti nt rakumembraani on
mittedifundeeruvaid ioone, jaotuvad ka difendeeruvad ja membraani hästi läbivad
ioonid mõlemal pool lahutuspinda erinevates kontsentratsioonides. Näiteks kui ühel
pool on suured negatiivselt laetud osakesed (nt anioonsed proteiinid), mis ei saa
membraani läbida, siis sealt
piirkonnast difundeeruvad väikesed negatiivselt laetud
ioonid kergemini ära kui katioonid.Kirjuta näide- Nt läbivad ioonid A+ ja B- ning R-,
mis ei suuda membraani läbida. A ja B ebaühtlane jaotus kahel pool membraani ning
täiendav osmootne rõhk R- poolel.
28) Membraani elektrotoonilised omadused e kaabliomadused. Membraani aja- ja
pikkuskonstant. Elektrotooniliste e passiivsete nähtuste sõltuvus närviraku omadustest. https://wiki.brown.edu/confluence/download/attachments/13802/BN102+Passive+2007.pdf?v ersion=1
Elektrotoonilised ehk passiivsed omadused on need omadused, mis võimaldavad (närvi)rakul
impulssi edasi kanda ilma voltaažtundlike kanalite avamiseta ehk ilma membraani
elektrijuhtivuse muutuseta. Elektrotooniliste potentsiaalide tekke eelduseks on ,et membraan
käitub passiivsena.
2
1
0
ar 2
u
Kolm omadust :
aan
a, j
Sisendtakistus- määrab, kui palju potentsiaal muutub.
niiL
rakusisene aksiaalne takistus- määrab, kui kaugele ja kui kiiresti impulss levib.
i-
Tsütoplasma takistuse ja närvi diameetriga määratud.
aal
M
8
mahtuvus- määrab, kui kiiresti pärast ärritust muutus tekib.
Elektrivoolu toimel tekkinud potentsiaalimuutus on elektrotooniline potentsiaal ehk
elektrotoonus. Elektirvooluga mõjutades määrab elektrotoonilise potentsiaali tõusu kalde
ainult membraani mahtuvus (vt joonist Schmidtist lk 35) , membraani läbib ainult
mahtuvusvool. Kuid
depolarisatsioon hakkab aeglustuma (sest K+ hakkab väljuma), lõpuks
saavutab see kindla lõppväärtuse, kus ioonide vool läbi membraani võrdsustub aplitseeritud
elektrivooluga ja membraanikondensaator edasi ei tühjene.
Membraani ajakonstant τ on aeg, mille jooksul elektrotooniline potentsiaal (elektrivoolu
toimel tekkinud potentsiaalimuutus) on lähenenud 37% oma täisamplituudist e
lõppväärtusest. Ajakonstant iseloomustab potentsiaali muutumise kiirust ,näitab, kui kiiresti
potentsiaal jõuab oma max väärtuseni, sõltub mahtuvusest. On ka membraani takistuse ja
mahtuvuse korrutis. Ajakonstant on eri rakkudel 5-50 ms.
Elektrotoonilised omadused mõjutavad
potentsiaalimuutuse ulatust
potentsiaali muutuseni kuluv aeg mõõdetuna ärrituse algusest
kui kaugele potentsiaalimuutus levib
kui kiiresti AP-sid genereeritakse
Elektrotoonilised nähtused sõltuvad membraani takistusest, mahtuvusest ja aksiaalsest
takistusest ehk siis kiu läbimõõdust , kuid esimesed kaks on erututvatel rakkudel sarnased, siis
põhiliselt mõjutab ikkagi kiu läbimõõt ning ka tsütoplasma takistus. Mida suurem
diameeter ,
seda rohkem membraani, seda rohkem kanaleid ja seda kiirem levik. Samas membraani
pindala suurenemisega suureneb mahtuvus ja see vähendab erutuse levikut. Isolatsioon
suurendab samuti levikut.
Pikkades
elongeerunud rakkudes tuleb arvestada ka pikkuskonstanti, sest pikkades rakkudes
voolab laeng välja väga ebaühtlaselt.
Pikkuskonstant λ näitab, kui kaugele elektrotoonilised
potentsiaalid elongeerunud rakkudes
levivad. võrdub vahemaaga, millel potentsiaal Emax on kahanenud 37%-le amplituudist voolu
aplitseerimise kohal. Närvirakkudel 0,5-1.
2
1
0
ar 2
u
aan
a, j
niiLi-
aal
M
9
29) Aktsioonipotentsiaal: olemus, ajaline kulg ja põhiomadused. Aktsioonipotentsiaal on erutus ,mis võib tekkida närvi- ja lihasrakkudes, kus
aktsioonipotentsiaal on
tsükkel , mis koosneb membraani
depolarisatsioonist
hüperpolarisatsioonist
repolarisatsioonist
(Na+ sissevool rakku ja sellele järgnev K+
väljavool )
Voltaažitundlikud ioonkanalid membraanis avanevad, tõstavad raku läbilaskvust
kõigepealt Na+ suhtes, siis K+ suhtes.
Na+ sissevool depolariseerib rakku, sellele järgneb K+ väljavool mis muudab raku
potentsiaali tagasi puhkepotentsiaalile. Kestab 1-2 ms, võib toimuda sadu kordi
sekundis.
Aktsioonipotentsiaal on
positiivse tagasiside mehhanism ,kus avanevad Na+ kanalid
põhjustavad järgmiste
avanemise . „Surnud ringi“ katkestab väline faktor, milleks on
Na+ kanali inaktivatsiooni värat. Kõik väratid reageerivad depolarisatsiooniga, aga
inaktivatsiooni oma 0,5 ms hilinemisega.
Kõik või mitte midagi seadus – iga
signaal , mis on üle
lävi (teatud potentsiaal, mille juures
depolariseerumine vallandab aktsioonipotentsiaali), põhjustab sama kuju ja suurusega
aktsioonipotentsiaali, iga signaal ,mis on alla lävi, ei põhjusta üldse aktsioonipotentsiaali.
AP levimisel tema
amplituud (u 100 mV) ei muutu.
AP levikukiirus
varieerub erinevates närvikiududes 0,5-100 m/s, esinevad
elektrotooniline
saltatoorne levik.
Viimane on kiirem ning energeetiliselt ökonoomsem.
AP-d iseloomustab
210
ar 2
Refraktaarsuse põhjustab Na+ süsteemi inaktiverumine
eelnenud aktsioonipotentsiaali ajal.
u
Refraktaarsus tagab ühesuunalise impulsi leviku.
aan
a, j
n
Suur muutus membraanipotentsiaalis leiab aset juba vaid mõne molekuli liikumisega.
iiLi-
aal
Erinevatel rakkudel on erineva kuju ja kestusega aktsioonipotentsiaalid.
M
1
0
Aktsioonipotentsiaali ülekande sünapsites tagavad neurotransmitterid.
Sel ajal kui Na+ kanalid hakkavad juba sulguma, hakkavad avanema K+ kanalid ning
algab hüperpolarisatsioon. K+ kanalid jäävad avatuks nii
kauaks , kui membraan on
depolariseeritud ja sulguvad alles siis, kui membraanipotentsiaal on jõudnud tagasi
algväärtuseni. Lõpuks voltaažtundlikud K+ ja Na+ kanalid sulguvad, ainsana on
avatud mittevoltaažtundlikud K+ kanalid (lekkekanalid), mis genereerivad
membraanipotentsiaali negatiivse puhkeväärtuse.
http://www.ncbi.nlm.nih.gov/books/NBK21668/-hea seletus Kui membraani iseloomustab potentsiaalide erinevus, on ta polariseeritud.
DEPOLARISATSIOON =HÜPOPOLARISATSIOON- polariseerituse vähenemine
HÜPERPOLARISATSIOON- polariseerituse suurenemine.
REPOLARISATSIOON on
polarisatsiooni
taastumine .
Muutes rakumembraani läbilaskvust eri ioonidele (e Px väärtust Goldmanni valemis),
muudame potentsiaali. Põhilised ioonid, millest oleneb on K+, Na+, Cl-, Ca2+.
21
Aktsioonipotentsiaalide ioonmehhanismid, 0
ar 2
u
Nii Na+ kui K+ kanalid on voltaažtundlikud ning
reageerivad membraani kriitilisele
depolarisatsioonile. Na+ kanalid avanevad kiiresti lühikeseks ajaks ning sulguvad uuesti.
aan
Eristatakse kolme olekut: suletud aktiveeritav, avatud aktiveeritud ja suletud inaktiveeritud (ja
a, j
nii
aktiveerimatu- jääb inaktiveerituks seni, kuni ta membraani re- või hüperpolarisatsiooni mõjul
L
i-
uuesti tagasi pöördub suletud , aga aktiveeritavasse olekusse). K+ kanalid avanevad
aal
M
1
1
aeglasemalt, veidi hiljem(u millisekund) kui Na+ kanalid , ei inaktiveeru depolarisatsiooni
jooksul.K+ Kanalil on kaks olekut: avatud ja suletud.
nende uurimise meetodid ( patch clamp meetod). Patch clamp meetod e
membraanilapikese kinnistamine.Kasutatakse mõõtepipetti, mille
umbes 1 mikromeetrise läbimõõduga ava on kinnistunud membraanile. Alarõhu rakendamisel
võib tekkida sulustus (kui elektroodi ots on absl. tolmuvaba ning raku
pealispind on
puhastatud sidekoefibrillidest ning
muust sarnasest. Sulustus isoleerib elektroodi otsal
olevas membraanilapikeses olevad kanalid ülejäänud membraanist elektriliselt (suure takistuse
tekitamise abil pipeti ja välislahuse vahel).
Pingekinnistamise võimendiga (hoiab pinge nõutav-pinge tasemel) , mis on ühendatud pipetti
täitva soolalahusega, on võimalik mõõta
kanalivoolusid. Saame üksikkanalivoolud ning neid
summeerides saame summaarsed
voolud , mille põhjal näeme, et Na+ kanalite avanemine on
kõige tõenäolisem üsna kohe pärast potentsiaali muutust, umbes 1ms pärast jäävad
avanemised harvemaks ja lõpuks lakkavad täiesti. K+ kanalid avanevad viivitusega, kuid
kujuneb välja keskmiste avanemiste sagedus, mis püsib konstantsena nii kaua, kui kestab
depolarisatsioon. Potentsiaali püsivana hoidmiseks vajaliku elektrivoolu muutsi mõõtes saame
iseloomustada membraani läbivaid ioonivoole.
210
ar 2
u
aan
a, j
niiLi-
aal
M
1
2
210
ar 2
u
aan
Ülemine
graafik näitab seda, et fikseeritud membraanilapikesel muudetakse
a, j
n
membraanipotentsiaal -80 mv -40mv-le, sellist pingemuutust korratakse sageli. Alumised
iiL
graafikud on saadud, kui on summeeritud palju registreeritud üksikkanalivoole, Na vool
i-
peatub teatud hetkel, K omadel kujuneb välja keskmiste avanemiste sagedus
aal
M
1
3
Aktsioonipotentsiaalid erinevates erutuvates kudedes. Schmidti õpikus lk 23 joonis 2-4
Närvikoes kestus u 1 ms, skeletilihases 10 ms (lihase
kontraktsioon 100 ms), südamelihases
200 ms.
Skeletilihases 10 ms. AP toimimise ajal on koes refraktsiooniperiood.
Südamelihases 200 ms.
Südamelihases on
TÜÜPILINE e TÖÖMÜOKARD (pumbafunktsioon)
ATÜÜPILINE e ERUTUSTEKKE- ja
JUHTESÜSTEEM (genereerib südamerütmi
ning korraldab eri südame osade koostöö)
Eri tüüpi müokardis on eri kujuga AP-d.
210
ar 2
u
aan
a, j
niiLi-
aal
M
1
4
21
0
ar 2
u
aan
Aktsioonipotentsiaal kui universaalne info ülekandmise viis närvisüsteemis. a, j
niiLi-
aal
M
1
5
Universaalne info ülekandmise viis- infot kannab edasi mitte signaalide amplituud (see on AP
puhul alati sama) ,vaid sagedus,millega AP-sid initseeritakse. See oleneb neurotransmitteri
hulgast.
Aktsioonipotentsiaali elektrotoonilise ja saltatoorse levimise mehhanismid. Elektrotooniline levik e
pidev levik- müeliinita kiududes. Järk- järguline membraani
depolariseerimine, kus aina uued alad jõuavad läveni, aina uued Na+ kanalid avanevad.
Depolarisatsioon ulatub läveni ning „päästiktsooni“(
trigger zone-i), siis voltaažtundlikud Na+
kanalid avanevad ja Na+ siseneb aksonisse. Na+
sisenemine depolariseerib membraani , see
avab uusi Na+ kanaleid
. Positiivne laeng liigub mööda aksonit edasi. Juba läbitud osa
(trigger zone) on refraktoorne osa, refraktoorses perioodis , K+ kanalid on seal avatud ning
Na+ inaktivatsiooniväratid sulgunud. See kindlustab aktsioonipotentsiaali ühesuunalise
leviku. K+ väljumine repolariseerib membraani. Aksoni distaalsemates osades aga
depolariseeruvad uued membraanipiirkonnad.
Saltatoorne levik e
hüppeline levik Leiab aset müeliiniga närvikius. See on hea
alternatiiv väga jämedatele kiududele.
Müeliin on isolaator, on vähem kontakti ekstratsellulaarse maatriksiga sp elektrilaine ei
„leki“.. Saltatoorne levik kasutab ainult müeliinivahelist vaba pinda. Kui AP tekib
päästikupiirkonnas, siis
AP laine liigub mööda müeliiniga tsooni müeliinvabasse tsooni e
Ranvieri soonisesse, mille membraanis on palju voltaažtundlikke Na+ kanaleid, tekib
depolarisatsioon Na+ sisenemise abil ning AP potentsiaal hoitakse konstantne, kui ta liigub
ühest soonisest teiseni. Levik on kiirem kui elektrotooniline, sest pideva kanalite
avamine aeglustab AP laine liikumist.
E levikukiirus oleneb kiu diameetrist ja müeliniseeritusest.
210
ar 2
u
aan
a, j
niiLi-
aal
M
1
6
Na+-K+- pump tekitab pidevalt vastukon tsentratsiooni.
30) Südame-vereringesüsteemi üldiseloomustus ja peaülesanded. Tegemist on peaaegu suletud hargneva ja seejärel
koonduva torustikuga (
veresooned -
arterid,
arterioolid ,kapillaarid,
veenulid ,veenid), milles on kaks
pumpa - südame
vatsakesed .
Vasak on ühenduses
suure vereringega ning parem on ühenduses
väikese vereringega.
Vasakusse kotta tuleb kopsuveenist hapnikurikas
veri , läheb
aordi kaudu kehasse,
paremast aordist läheb õõnesveenidest tulev hapnikuvaene veri pulmonaalarteri kaudu kopsudesse.
Paremasse kotta tuleb kehast hapnikuvaene veri, mis viiakse kopsudesse, et see seal
hapnikuga rikastada. Torustik ei ole päris suletud, sest kapillaarides on võimalik ainete
vahetus interstitsiaalse ruumi e rakkudevahelise ruumi ja rakkude vahel.
Bioloogiliste funktsioonide täitmiseks peab veri olema ühesuunalises ringluses, selle tagavad
rõhkude
vahed vereringesüsteemis ning
klappide olemasolu. Suurel vereringel on
paralleelringed, viivad nt ajju, jäsemetesse, neerudesse, maksa jne. Kuna organismi eri
piirkondade ainevahetuslikud vajadused on erinevad, peab ka eri piirkondade verevarustus
olema paindlikult reguleeritav. Vereringes voolab veri, mida on täiskasvanul umbes 5 l, 55-
60% on vereplasma ja 40-45% on
vererakud , seda protsenti nimetatakse HEMATOKRITIKS.
Vere
viskoossus on 3-5 korda suurem kui veel.
Ülesanded: Veri kannab hapniku kopsudest ning toitained seedeelundkonnast ning kannab
need kõigi rakkudeni, samaaegselt jääkaineid ära kandes. Lisaks on kardiovaskulaarsel
süsteemil tähtis roll rakkudevahelises kommunikatsioonis hormoonide jm signaalmolekulide
edasikandjana, samuti on vererakkudel kaitsefunktsioon, nad osalevad immuunsüsteemi töös.
Vereringe talitluse aluseks olevad füüsikalised protsessid. Vere vool teatud kehapiirkonda on kõige enam määratud
rõhkude vahe
soonte diameetri abil
Rõhkude vahe tekitamise üks viise on südamelihase kontraktsioon ning lõõgastumine.
Veresooned ei ole jäigad torud, nende diameeter on
muudetav seinte elastsuse ning
seinas oleva silelihaskihi olemasolu abil.Samuti muutub keha asend ruumis. Seega on
21
vere
voolamine väga
komplitseeritud ning kasutatakse lihtsustatud
mudeleid .
0
Veri peab voolama ühes suunas-
klapid .
ar 2
u
Vere
filtratsioon kapillaarides toimub filtratsiooni ja difusiooni teel, mille suuna
määrab ära keskkondade hüdrostaatilise ja
osmootse rõhu summa.O
aan
nkootse ja
a, j
hüdrostaatilise rõhu vahe määrab ära, kas vesi liigub kudedesse või kudedest ära.
nii
Südame pumbafunktsiooni aitab täita südamelihasrakkude poolt
tehtav töö, mida saab
L
i-
arvutada A=pv,lisandub veel kiirendustöö, mis kulub verele kiirenduse andmiseks.
aal
M
1
7
31) Südame pumbafunktsioon, olulisemad seda iseloomustavad parameetrid . Südameõõnte, klappide ja müokardi roll südame pumbafunktsiooni
realiseerumisel. Südametoonid, nende tekkimise põhjused. Südame-vereringesüsteemis on kaks pumpa- südame vatsakesed. Vasak on ühenduses
suure vereringega ning parem on ühenduses väikese vereringega. Vasakusse aorti
tuleb kopsuveenist hapnikurikas veri, läheb aordi kaudu kehasse, paremast aordist
läheb õõnesveenidest tulev hapnikuvaene veri pulmonaalarteri kaudu kopsudesse.
Koja ja vatsakese vahel on HÕLMASED e ATRIO-VENTRIKULAARklapid.
Vasaku vatsakese ja aordi vahel ning parema vatsakese ja kopsuarteri vahel on
POOLKUUKLAPID ehk semilunaarklapid.
Klapid sulgevad ja avanevad vererõhu erinevuste tõttu mõlemal pool klapi tööpinda.
Südametoonid on klappide sulgemise helid, 1. Südametoon on atrioventrikulaarklapi
sulgumine (süstoli algus) ja 2. Toon on poolkuklapi sulgumine (
diastoli algus) Klapid
tagavad ühesuunalise verevoolu nii südames kui kogu vereringes. Südametoonide
tekitatavad
võnked kanduvad rindkerele ja on kuulmispiirkonnas.
Pumbafunktsioon realiseerub järjest korduvate tsüklite käigus. Tsükkel koosneb
diastolist, mille käigus
müokard lõõgastub ning südameõõn täitub ja süstolist, mille
käigus müokard kontrakteerub ja veri väljub õõnest.
Iseloomustavad parameetrid:
-
FREKVENTS ehk südametsükli sagedus. Normaalses rahulolekus 70 tsüklit
minutis .
-
LÖÖGIMAHT ehk vatsakesest süstoli käigus väljutatud vere maht. Normaalselt 70 ml.
-vatsakese
MINUTIMAHT ehk
MAHTKIIRUS Q=V*fc, kus V on löögimaht ja fc on
südametsükli sagedus e
löögisagedus minutis. Normaalselt 5 l/min (70*70=4900ml)
Kumbki
vatsake pumpab ajaühikus sama koguse verd.
Pumbad on järjestikku, seega
mahtkiirus on ühest vatsakesest minutis väljutatud vere ruumala.
Müokardi roll: MÜOKARDIOTSÜÜDID on erutumisvõimelised, võimelised
genereerima aktsioonipotentsiaali.
Tüüpiline müokard, mille kontraktsioon aitab täita pumbafunktsiooni
atüüpiline müokard, mis on südame erutustekke- ja juhtesüsteem, kontrollib
südamerütmi ja eri osade koostööd.
21
Südame rütmiliste pulsatsioonide vallandajateks on
südames endas tekkivad erutused.
0
Erutus levib, kuni haaratud on kõik südamelihaskiud.
ar 2
u
aan
32) Südametsükkel, selle põhifaasid . Vererõhu ja mahu muutused tsükli eri faasides . a, j
nii
Kodade ja vatsakeste tsüklid toimuvad mõlemal poolel sünkroonselt, kuid
L
paremal pool
i-
arendatavad rõhud on palju väiksemad. Vatsakese tsükkel on 3 korda pikem. Kodadel
aal
M
1
8
ja vatsakestel mõlemal on eristatavad
diastol ja
süstol . Kui löögisagedus on 60 l min,
siis ühe tsükkli kestus on 1 sek.
Südametsükkel:
Hiline diastol, kus
kojad ja vatsakesed on lõõgastunud ja vatsakesed täituvad verega.
Atrioventrikulaarklapid avanevad ,sest vatsakeses on madalam rõhk, ta on
lõõgastunud.
Kodade süstol- kodade kontraktsioon
surub väikese lisahulga (viimase 20%) verd
vatsakesse. Samal ajal vatsakesed on diastolis.
Vatsakese süstol:
1) Isovolumeetriline kontraktsioon- Atrioventrikulaarklapid surutakse kinni (vere poolt,
mis surub nende vastu vatsakese poolt), kuid rõhk pole veel piisavalt suur, et
poolkuuklapid avaneksid.
2) Väljutusfaas- vatsakestes on suurem rõhk kui
arterites , poolkuuklapid avanevad ja veri
väljub
Samal ajal kojad diastolis ja täituvad. AV
klapp on kinni.
Vatsakese diastol 3) Isovolumeetriline lõõgastus
4) Vatsakesed lõõgastuvad, rõhk väheneb, poolkuuklapid sulguvad, sest veri hakkab
hetkeks tagasi voolama, surub klapid kinni arterite poolsest küljest.
5) Täitumisfaas e hiline diastol vt eespool. Vatsakeste lõõgastumine põhjustab AV
klappide avanemise.
6) Samal ajal kojad diastolis.
Diastol on pikem kui süstol.
VATSAKESTE Süstol jaguneb - Isovolumeetriline kontraktsioon ja väljutusfaas. Diastol -
isovolumeetriline lõõgastumine ja täitumisfaas.
Rõhu-mahu diagramm
210
ar 2
u
aan
a, j
niiLi-
aal
M
1
9
Südametsükkel algab punktis A. Vatsake on lõõgastunud, veri on voolanud kotta,on
jõudnud kätte hetk, mil
kojas on kõrgem rõhk kui vatsakeses ning atrioventrikulaarklapp
avaneb punktis A, jätkub vatsakese
täitumine , kuid rõhk suureneb väga vähe. Nüüd algab
koja süstol e kontraktsioon ,mis täidab viimase 20% EDV-st (end-diastolic volume e
lõppdiastoolne maht, mis täiskasvanud mehel on umbes 135 ml). Algab vatsakese
kontraktsioon- AV klapp sulgub, poolkuuklapp ei ole veel avanenud- isovolumeetriline
kontraktsioon, kus rõhk kasvab,aga ruumala püsib sama. Kui rõhk on piisavalt suur, siis
semilunaarklapp avaneb ning algab väljutusfaas, kus rõhk kasvab jätkuva kontraktsiooni
tõttu, veri väljutatakse südamest, osa verd jääb sisse- EDV (end-diastolic volume e
lõppsüstoolne maht) Kontraktsiooni lõpus hakkab rõhk vatsakeses vähenema,
poolkuuklapid sulguvad. Vatsake on nüüd suletud kamber ja ülejäänud lõõgastumine
toimub sellises olekus, lõpuks, kui rõhk on madalam kui kojas, avaneb
mitraalklapp e üks
hõlmalistest klappidest ning tsükkel algab otsast. Lõppdiastoolse ja lõppsüstoolse mahu
vahe on väjutusfraktsioon. Rõhkude muutused südametsükli vältel vasakus vatsakeses 0-
115 mm/Hg,
aordis 76-115 mm/Hg, vasakus kojas 0-10 mm Hg
33) Vere voolamise seaduspärasused veresoonkonnas. Vere voolamise maht- ja joonkiirused ning takistused veresoonkonna eri osades. Veri voolab suurema rõhuga piirkonnast madalama rõhuga piirkonda. Kirjeldatakse
Poiseuille võrrandiga. Tegurid on
soone pikkus, diameeter, vedeliku viskoossus ja muidugi rõhkude
210
vahe.
ar 2
u
Vere
voolutugevus on tugevasti lihtsustatult analoogne Ohmi seadusega V=ΔP/R=Q.
aan
a, j
Vere mahtkiirus on ajaühikus ristlõikepindala läbiva vere ruumala.
niiLi-
aal
M
2
0
Mida suurem on vere viskoossus, seda suurem takistus, mida suurem
veresoone pikkus,
seda suurem takistus, mida suurem diameeter, seda väiksem takistus, sest vähem
vedelikku puutub kokku soone seintega ja seda vähem hõõrdumist.
Laminaarse voolamise korral osakeste liikumise (voolamise) trajektoorid ei ristu.
Turbulentsi korral suureneb energia dissipeerimine (
hajumine , mõnes mõttes
raiskamine)
Voolamist saab kirjeldada kaht erinevat viisi- 1)
Bernoulli võrrand, mis
arvestab inertsi,
aga mitte sisehõõrdumist ning 2) Hagen-Poiseuille seadus, mis arvestab
sisehõõrdumist ja mitte inertsi.
210
ar 2
u
aan
a, j
nii
Tegemist on vaid kaudse kirjeldusega, sest vereringesüsteem ei täida täielikult järgmisi
L
i-
nõudeid:
aal
M
Torud on jäigad ,mittehargnevad, ümara
ristlõikega .
2
1
Vedelik on
homogeenne Voolamine on laminaarne ja
statsionaarne (paikne).
Kõige rohkem saab modifitseerida veresoone diameetrit.
Silelihaskihi abil!
Voolutugevus sõltub võrdeliselt r4 ja takistus samast
asjast pöördvõrdeliselt.
Joonkiirus = mahtkiirus/ristlõike pindala= Q/A
Mahtkiirus on kõigis veresoonkonna osades sama, kuid joonkiirus on kapillaarides
väiksem (nende kogu ristlõikepindala on suurem) ning suurtes veresoontes sellega
võrreldes kiirem, kuid üldiselt, võrreldes üht suurt ja üht väikest soont, on suures
voolamine
aeglasem .
Takistus aordis, suurtes arterites ja arteriharudes ei moodusta üldisest takistusest üle 1/5,
sest neil on suur diameeter(?). Terminaalsetes arterites ja arterioolides on takistus
peaaegu pool kogutakistusest, tingitud nende diameetri vähenemisest, mida
paralleelselt lülitatud sooned kompenseerida ei suuda. ¼ takistusest on põhjustatud
kapillaaride poolt. Veenulite takistus on 4 % ja ülejäänud veenide oma 3%.
34) Vererõhk veresoonkonna eri osades, vererõhu mõõtmise meetodid. Vererõhk on jõud, mida veri avaldab veresoone seinale pinnaühiku kohta.
210
Arterites on rõhk suurim, kuid südametegevuse
faaside tõttu perioodiliselt muutuv.
ar 2
u
Aordis on rõhk max 120 mm Hg vatsakese süstoli ajal(
süstoolne rõhk, normväärtus
aan
110-140) ning min 80 mm Hg (60-90)vatsakese diastoli ajal.(diastoolne rõhk) Kuigi
a, j
rõhk vatsakeses langeb ligi 0, on aordis rõhk
püsivalt üsna kõrge. Antud nähtus on
niiL
põhjustatud sellest, et
arterite seinad on elastsed ning venides võimelised energiat i-
säilitama. aal
M
2
2
Pulsirõhk on süstoolne rõhk- diastoolne rõhk. Vererõhk väheneb teekonnal läbi keha
tagasi südamesse, sest energia väheneb takistuse tõttu. Arterioolidest alates vererõhu
kõikumised kaovad (ning
pulssi ei saa
katsuda ).Veenides on rõhk väga madal ning
tagurpidi voolamist takistavad veeniklapid.
Keskimne arteriaalne rõhk on diastoolne P+ 1/3(süstoolne P-diastoolne P). Diastol on
2x pikem kui süstol, sp on keskmine arteriaalne rõhk lähemal diastoolsele.
Keskmine arteriaalne rõhk näitab
vere voolamist tekitavat jõudu. Vererõhu
hetkväärtuste keskmine tase teatud lõigus. Arterites muutub üsna vähe.
Ning 1mm Hg = 13,6 mm H2O
Mõõtmine
Vererõhu mõõtmised on OTSESED ja
KAUDSED .
Otseste korral viiakse
arterisse nõela või kanüüliga ühendatud
rõhuandur.
Kaudsete puhul kasutatakse sageli rõhumansetti. Eeldatakse, et rõhk kandub sügavamal
asuvatele kudedele ilma rõhukadudeta. Arteriseina käitumine sõltub arteri rõhu ja
mansetirõhu vahest, mida nimetatakse TRANSMURAALSEKS rõhuks.
Riva -Rocci e Korotkovi meetod- õlavarremanseti rõhk viiakse kiiresti süstoolsest rõhust
kõrgemale ja hakatakse seejärel aeglaselt
langetama , kuulates samaaegselt arteria
brachialise kohal
küünarliigese sisepinnal stetoskoobiga Korotkovi toone, nende
põhjuseks on turbulentne voolamine, mis tekib arteri mansetialuses ahenenud osas.
Kui manseti rõhk on kõrgem süstoolsest rõhust, on
arter kokkusurutud, verevool
peatub ja toonid puuduvad. Kui manseti rõhk langeb madalamale süstoolsest rõhust,
tekib iga pulsilöögi ajal lühike
terav kahin. Kui mansetirõhk on süstoolse ja
diastoolse rõhu vahel, on kahinate käitumine küllalt individuaalne. Kui mansetirõhk langeb
diastoolse rõhuni, siis kahinad äkki tumenevad ja nõrgenevad kiiresti.
210
Automaatsetes vererõhumõõturites on kasutusel
ostsillograafiline meetod. Selle puhul
ar 2
u
määratakse vererõhk süsteemis arter+rõhumansett tekkivate pulsituigete e
aan
ostsillatsioonide põhjal. Muudetakse vasturõhku, ning sellega muutub arteri
a, j
transmuraalne rõhk ning ostsillatsioonide kuju ja amplituud. Kui arteri sein on
niiL
pingevaba , on ostsillatsioonide amplituudi max. See vastab
keskmise arterisisese i-
rõhu tasemele . Süstoolne ja diastoolne rõhk leitakse vastavalt algoritmile arvestades
aal
ostsillatsioonide KUJU ja AMPLTUUDI muutusi mansetirõhu langetamisel.
M
2
3
35) Hingamise põhietapid, nende realiseerumise aluseks olevad füüsikalised protsessid. Hingamine on protsesside kogum , mille käigus varustatakse rakud hapnikuga ja
eemaldatakse liigne
süsihappegaas . Täiskasvanu tarvitab min hapnikku 0,25-3 l ja
eraldab süsihappegaasi 0,2-0,25 l.
Hingamise põhietapid e hapniku tee:
Kopsuhingamine ehk väline hingamine 1)
Konvektiivne transport õhust kopsualveoolidesse e
ventilatsioon . (
Boyle seadus)
2) Difusioon kopsualveoolidest verekapillaaridesse.
Ficki difusiooniseaduse põhjal. Hingamisgaaside transport verega 3)Konvektiivne transport vereringe vahendusel koekapillaaridesse.
Koehingamine ehk sisemine hingamine 4) Difusioon koekapillaaridest neid ümbritsevatesse kudedesse
5) Hapniku kasutamine rakkude ainevahetuses
!CO2 eemaldamine vastupidises järjekorras.
Rindkere seina elastsed jõud püüavad rindkere mahtu suurendada, aga kopsude elastsed
jõud ja alveoolide
pindpinevus (tekib, sest alveoolide sisepind on kaetud vedelikuga)
püüavad ruumala vähendada, tekib negatiivne intrapleuraalne rõhk (negatiivne
atmosfäärirõhu suhtes).
Elastne takistus on venitatavuse pöördväärtus.
Üldine gaasiseadus
PV=
RT. Kus P on rõhk, V on ruumala, T on temperatuur, R on
universaalne gaasikonstant. Võib
väljendad ka nii, et
PV/T on konstantne.
Mõned tähtsamad gaasiseadused on: DALTONI SEADUS: gaasisegu kogurõhk on tema komponentideks olevate gaaside
osarõhkude summa.
210
ar 2
u
aan
a, j
niiLi-
aal
M
2
4
Daltoni seadusel põhineb hingamisgaaside vahetus verega. Kopsudesse tulevas venoosses
veres on hapniku osarõhk madalam kui
alveoolides ning süsihappegaasi osarõhk
suurem. Järelikult hapnik verre (kõrgemalt osarõhult madalamale) ning CO2
alveoolidesse.
BOYLE SEADUS: kui gaasianuma ruumala muutub, muutub rõhk pöörvõrdeliselt. Kui
vähendame ruumala, siis nt ½ võrra, siis rõhk kahekordistub, sest kokkupõrked
sagenevad. PV=P1V1, KUI T on
const !.
210
CHARLES´i SEADUS: V/V1=T/T1 kui P on const! St et kui temperatuur tõuseb, siis
ar 2
u
ruumala suureneb ka!
aan
a, j
Henry seadus: Tasakaaluseisundis on
vedelikus lahustunud gaasi kontsentratsioon
nii
võrdeline gaasi osarõhuga vedeliku kohal.
Seda seadust järgib hapniku lahustumine L
i-
kopsu ümbritsevas vedelikus. Lahustumine on vajalik difusiooniks rakkudeni, aal
kus hapnikku vajatakse. M
2
5
Rinnaõõne e thoraxi ruumala muutmisega muudetakse rõhkude vahet ning
saavutatakse ventilatsioon.
Gaas või gaaside segu voolab kõrgema rõhuga piirkonnast madalama rõhuga piirkonda.
Difusiooni soodustab lühike difusioonitee ja suur difusioonipindala.
36) Hingamistsükkel, selle osad. Sisse- ja väljahingamise mehhanism, nendega kaasnevad rõhu ja mahu muutused rindkeres . Kopsukoe elastsus , selle tähtsus
hingamismehhaanikas. Venitatavus hingamissüsteemis, selle mõõtmine. Hingamismehaanika all mõistetakse hingamistsükli jooksul esinevate rõhu-mahu suhete ja
rõhu-voolu suhete analüüsi.
Hingamistsükkel koosneb sisse ja väljahingamisest. Sisse- ja väljahingamise aluseks on
intraalveolaar- ja atmosfäärirõhu erinevus.
Sissehingamisel (
inspiratsioon ) peab
intraalveolaarne rõhk olema väiksem ja
väljahingamisel (ekspiratsioon) suurem kui
atmosfäärirõhk. Hingamine saab alguse hingamislihastest (m.
Intercostales interni et
externi et m
scalenus et
diaphragma), mis saavad muuta rindkere mahtu
kolmes
projektsioonis. Hingamistsüklite arv on täiskasvanul
12-16 korda minutis. Hingetõmbe sügavus on 0,5 l.
Minutiventilatsioon on siis u 16*0,5=8 l
Sissehingamisel ei saa hingamisteede takistuse tõttu õhk küllalt kiiresti
tungida suurenenud alveolaarruumi. Seetõttu peab rõhk alveoolides langema st võrreldes
välisrõhuga negatiivseks muutuma. Intrapleuraalne rõhk muutub tulemusena veel
negatiivsemaks.
Kui rindkere on lühiajaliselt puhkeseisundis,nagu
sissehingamise üleminekul
väljahingamiseks, avaldab pleuraõõnele mõju ainult mõju ainult
kopsude elastne
retraktsioon (kokkutõmbumine) ja see põhjustab negatiivse rõhu. Pidev negatiivne
2
inrapleuraalne rõhk väldib kopsude kokkuvajumist. Intrapulmonaarne rõhk on
1
0
puhkeoleku ajal võrdne nulliga, sest suuõõs ja
alveoolid on omavahel ühenduses.
ar 2
u
Väljahingamisel rindkere maht väheneb, intrapleuraalne rõhk muutub positiivsemaks ning
aan
intraalveolaarne rõhk suureneb.(positiivseks!)
a, j
niiLi-
Sissehingamine: Hingamislihaste kontraktsioonil (intercostalestest externused) suureneb
aal
rindkere maht ning rõhk intrapleuraalruumis muutub veel negatiivsemaks. Tulemusena
M
2
6
väheneb intraalveolaarne rõhk ning õhk voolab kopsudesse. Intrapleuraalne rõhk on u
-0,5 kPa tavaolekus, sissehingamisel kuni
-1 kPa (-3 mm Hg ja -6 mm Hg resp.) LaPlace jõud väljendab rõhku. Vedelikumullisisene rõhk P= 2x T/r, kus T on pindpinevus
ja r on mulli raadius. Väikestes mullides ja ka väikestes alveoolides seega peaks olema
suurem rõhk, kui vedelik on sama pindpinevusega. Vedelik alveoolide seintes
põhjustab pindpinevust,mis vähendab kopsude venimisvõimet. Surfaktant vähendab
kohesiivseid jõude veemolekulide vahel. Väiksematel alveoolidel on rohkem
surfaktanti, nii et nende pindpinevus on väiksem kui suurtes alveoolides- see aitab eri
suurusega alveoolides rõhku ühtlustada.
Kopsude venitatavus e compliance- ruumala muut rõhu muudu kohta .
Venitatavus on elastse takistuse pöördvärtus, mis kujuneb kopsude ja rindkere
venitatavuse koosmõjul. Sügaval sissehingamisel piiravad venitatavust
kopsud ,
sügaval väljahingamisel
rinnakorv .
1/(Cth+l)=1/Cth+ 1/Cl
Praktikumis mõõtsime venitatavust DONDERSI
mudelil , kus kaks õhupalli on analoogid
pleuraruumile ning kopsuruumile. Sisse-ja väljahingamist imiteeritakse süstla abil.
Määratakse erinevatele mahtudele vastavad rõhud. Andmed salvestatakse rõhu-
ruumala graafikutena.
210
Saadud kõverate (rõhu-mahu graafikute) tõus on venitatavus. Rindkere venituskõvera
alumine osa on
lamenenud ja kopsude venituskõvera ülemine osa on lamenenud, seal
ar 2
u
on tõusud väiksemad ehk venitatavus vähenenud. Seega sügaval sissehingamisel ehk
aan
kopsude mahu
suurenemisel on takistavaks kopsud, väljahingamisel ehk mahu
a, j
vähenemisel on takistuseks rindkere.
niiLi-
Venitatavus on suurim kui intrapleuraal- ja intraalveolaarrõhkude erinevus on 0.
aal
M
2
7
Rõhu-mahu graafiku puhul tuleb arvestada ka sellega, et toimub
hüsterees- mitteelastse
deformatsiooni tõttu sõltub mahu väärtus eelmisest mahu väärtusest.
Lisaks elastsele takistusele tuleb ületad
mitteelastne takistus, mille peamine vorm on
voolutakistus
hingamisteedes lisaks koetakistus kopsude ja rinnaõõne seina poolt
õhu ja kudede inerts.
37) Organismi tugiaparaadi ( skelett , liigesed, lihased) biomehhaanika, selle
koordineerituse vajalikkus stabiilseks kehaasendiks ja liikumiseks ruumis. Biomehaanika on teadus, mis käsitleb elusolendite liikumist ja liigutusi ning
nende põhjuseid. Organismi tugiaparaati kuuluvad
luud , liigesed ja lihased.
Lihased moodustavad umbes 40-50% kehakaalust. Lihased panevad liigesed (ja selle
kaudu luud ning kogu skeleti) liikuma.
Lihasrakkudel on võime kontraheeruda e aktiivselt lühendada oma pikkust.
Lihaskontraktsioon on keemilise energia muutmine
mehaaniliseks energiaks ja
soojuseks.
Vajalik on tugiaparaadi koordineeritus ,et säilitada stabiilne kehaasend ja liikumine.
210
Inimese liigutustegevuse aluseks on
kehahoid ehk
rüht . Korrektne kehahoid tagab
ar 2
u
liikumistegevuse korral koormuse õige
jaotumise kehaosade vahel.
aan
a, j
niiLi-
Rühti ehk kehahoidu mõjutavad:
aal
M
kehapoolte sümmeetrilisus
2
8
agonist - ja antagonistlihaste tasakaalustatud areng
liigeste liikuvuse ja jõu tasakaalustatud areng
38) Lihaskontraktsiooni libisevate filamentide teooria. Lihase mudel. Libisevate filamentide teooria kohaselt on lihase arendatud pinge otseselt proportsionaalne
jämedate ja peente filamentide vastasmõjuga.
Lihase kontraktsioon e lühenemine on müofibrillis(sama pikk kui
lihaskiud ) jadamisi
ühendatud sakromeeride ( piirkond ühest Z vöödist järgmiseni) lühenemise resultaat.
Lühenemisel libisevad
peened aktiinifilamendid mööda jämedaid müosiinifilamente, millega
nad on
vaheliti ja lükkuvad sügavale jämedate filamentide kimpu, kuni jõuavad lõpuks
sarkomeeri keskpaika. Müosiini- ja aktiinifilamendid ise ei lühene. A-vöötide pikkus jääb
konstantseks, aga I-vööt ja H-tsoon
ahenevad . Venitamisel tõmmatakse peenikeste
filamentide kimpu jämedate filamentide vahelt vähemas või suuremas ulatuses välja.
Müosiinifilamendi ristijätketeks on umbes 150 müosiinimolekuli PEAD. Igal peal on 2
seostumiskohta - üks aktiini jaoks ja üks ATP jaoks. Iga müosiinipea võib kontraktsioonil
müosiinifilamendi ühendada aktiini naaberiflamendiga
ristsillana.Kallutusliigutustega
sõuavad müosiinipead ühisel jõul sarkomeeri keskpaiga poole. Müosiinimolekulide
bipolaarne paigutus põhjustab aktiininiitide liikumise teineteisele vastu.
Kontraktsiooni alguses on müosiinipead seotud G-aktiini
molekuliga , väga lühike periood,
järgnevalt seostub ATP ning müosiinipea vabastatakse aktiinist. ATP- ADP+ P- mõlemad
jäävad seotuks müosiiniga. ATP-st vabanenud energia muudab müosiini kuju, nii et see
seostub uue G-aktiiniga, üks või kaks kohta kaugemal algsest, nüüd on müosiinil
potentsiaalne energia. P-rühm vabastatakse – power
stroke e AERULÖÖK.
Aktiin liigub
sakromeeri keskpaiga poole. Kui ADP ka vabastatakse, on müosiin jälle tihedalt aktiiniga
seotud. Müofibrilli järjestikku ühendatud üksiksakromeeride lühenemised liituvad. Ristsildade
korduvad
molekulaarliigutused muudetakse
makroskoopiliseks liikumiseks.
Müosiinipeadel on ka ATP-aasne aktiivsus e nad käituvad ensüümina.
TROPONIIN ja
TROPOMÜOSIIN reguleerivad aktiini seostumist müosiiniga. Realiseerub trossi tirimise
printsiip.
Ühe ristsillakeste tsükliga lüheneb
sarkomeer 1% oma pikkusest, lihas aga
üksikkontraktsioonil 20%, seega toimub hulk tsükleid.
Lihase mudel koosneb
kolmest komponendist .
1. Kontraktiilsed elemendid, mis tagavad lihase lühenemise. Müosiini- ja
aktiinifilamentide moodustatud ristsildade kompleks
2. Jada- e järjestikühenduses elastsed elemendid.
KÕÕLUSED 21
3. Rööp- e paralleelühenduses elastsed elemendid. SARKOLEMM ja
0
SARKOPLASMAATILINE RETIIKULUM ja muud sarkomeeri
struktuurid .
ar 2
u
aan
a, j
niiLi-
aal
M
2
9
Lihas võib kokku tõmbuda ja arendada jõudu ilma lühenemata.
Lihaskontraktsiooni põhiviisid, neid iseloomustavad füüsikalised näitajad Kontraktsioonijõud oleneb sarkomeeri pikkusest puhkeolekus.
Liiga lühikeste sakromeeride puhul kattuvad aktiini-ja müosiinifilamendid nii palju, et
libisemisel hakkavad aktiinimolekulid kiiresti kattuma ning see takistab ristsildade
teket ja kontraktsiooni. Liiga pikkade sakromeeride puhul kattuvad müosiin ja aktiin
liiga vähe.
Lihase arendatavat pinget ehk jõudu saab suurendada ,suurendades sagedust, millega
lihase aktsioonipotentsiaalid lihaskiude stimuleerivad. Aktsioonipotentsiaale
genereerib lihast
kontrolliv motoorne neuron .
Leiab aset ÜKSIKKONTRAKTSIOONIDE
summatsioon - kui lihas ei ole enne uut
stiimulit täielikult lõõgastunud, siis
kontraktsioonid liituvad.
Lõpuks jõuab lihas
maksimaalsesse kontraktsiooni- teetanus .
Mittetäielik ehk hambuline teetanus- lihaskiud lõõgastub veidi kontraktsioonide
vahel. Täielik teetanus e sile teetanus- lõõgastumist ei toimu üldse. Kui lihas väsib,
siis pinge hakkab vähenema, hoolimata jätkuvatest stiimulitest.
Lihase
kontraktsiooniühik on
motoorne ühik, mis koosneb ühest somaatilisest
motoorsest neuronist ning lihaskimbust, mida see inerveerib.
Ühe
motoorse ühiku kontraktsioon allub põhimõttele „kõik või mitte midagi“, kuid
varieeruvus lihaskontraktsioonitugevuses tuleneb sellest, et erineva
tugevusega stiimulus põhjustab eri
motoorsete ühikute kontraktsiooni.
210
ar 2
u
aan
a, j
niiLi-
aal
M
3
0
ISOMEETRILINE KONTRAKTSIOON
Jõud ilma liikumiseta. Nt
hoiad kangi enda ees õhus paigal või lükkad liikumatut
objekti. Elastsed elemendid (kõõlused, sidekude lihaskiudude vahel, tsütoskeleti
valgud müofibrillide vahel) venivad- tänu sellele saab üldse toimuda isomeetriline
kontraktsioon, see võimaldab lihaskiududel jääda sama pikaks, kuigi sarkomeerid
lühenevad. Kui elastsed elemendid on välja venitatud ja
jõud vastab raskusele,
toimub isotooniline kontraktsioon ja raskus tõstetakse üles.
Kui koormus on võrdne maksimaalse isomeetrilise jõuga või lihas lüheneb
koormuseta- töö on 0, sest lihastöö A= f * d ja sellistel juhtudel on üks
komponentidest 0, seega kogu töö on 0.
ISOTOONILINE KONTRAKTSIOON
Lihase lühenemine konstantse pinge või koormuse juures. Nt raskuse tõstmine
konstantse kiirusega. Kontraktsioon, mis tekitab jõudu ja liigutab raskust.
Jaguneb kaheks-
kontsentriline lihaspinge kasvab, et vastata koormusele ning siis on konstantne ,kui lihas
lüheneb
ekstsentriline lihas pikeneb, sest koormus on suurem, kui lihas tõsta jõuab.
AUKSOTOONILINE KONTRAKTSIOON
Nt kummipaela
venitamine .
21
Kontraktsioon, kus üheagselt lihase lühenemisega suureneb tema poolt arendatav
0
jõud. Auksotoonilise kontraktsiooni maksimum on palju väiksem kui isomeetrilise
ar 2
u
kontraktsiooni
maksimumi korral.
Peaaegu isotooniline kontraktsioon (
vist seepärast, et isotoonilisel kontsentrilisel
aan
kontraktsioonil algul lihaspinge kasvab ja siis lihas hakkab lühenema, kuid
a, j
nii
isotoonilise puhul on selles faasis pinge konstantne)
L
i-
aal
Mida väiksem on koormus, seda suurem on lühenemine ajaühikus.
M
3
1
Lihastöö on veel kord jõud *
teepikkus . A= f*d Teepikkus on kangi kinnituskohast
punktini, millele avaldatakse jõudu e jõu mõjusirge kaugus pöörlemisteljest.
Keskmiste koormuste seadus- keskmiste koormuste korral on töö suurem kui väga
väikese või väga suure koormuse korral. Töö on 0, kui lihas lüheneb koormuseta või
koormus on võrdne maksimaalse isomeetrilise jõuga.
Biitsepsi
kinnituskoht luule annab võimaluse 1cm lihaskontraktsiooniga liigutada kätt
5 cm.
39) Shannoni infoteooria põhiskeem. Informatsioonilised protsessid elussüsteemides: nende tähtsus organismi talitluses ja informatsiooni ülekande
olulisemad viisid. Infohulga mõõtmine. Informatsiooniteooria ja reguleerimisteooria on mõlemad küberneetika (juht,
tüürimees )
osad.
Informatsiooniteooria
mõõdab sõnumite infosisaldust
kirjeldab infoülekandesüsteeme
Informatsioon ei ole ühe süsteemi absoluutne omadus, tähtis on see, kuidas ta mõjub
teisele süsteemile, Informatsioon on stiimul/suurus, mis muudab midagi selles, kes
infot vastu võtab, muudab tema olukorda ehk siis kõige tähtsam on uudsus. See, et
täna on näiteks reede ei ole info, kui ma seda juba tean. Info puudub, kui ei teki
muutust või kui tal puudub tähendus
vastuvõtja jaoks.
Shannoni järgi on informatsioon MÄÄRAMATUSE VÄHENEMISE MÕÕT PÄRAST
SÕNUMI VASTUVÕTMIST.
21
Sündmuse infosisaldus on seda suurem, mida väiksem on selle sündmuse ilmumise
0
tõenäosus. Täringunäited.
ar 2
u
aan
Kui tõenäosus mingi sündmuse jaoks on p=1, siis on infosisaldus 0.
a, j
nii
Infosisaldus
(logL
2(1/p) või log2n, sest p on 1/n.
i-
aal
M
3
2
Infosisalduse ühik on
BITT (
binary digit) e infokogus, mis antakse edasi üheainsa
kahendmärgiga.
Kahendsüsteem – on/ei ole e AP/ ei ole AP
BAIT –infokogus, mis antakse edasi 8 bitiga. Pmt 8 ruudukest ,kus 0 või 1.
Aktsioonipotentsiaal kasutab kahendsüsteemi!
Kõik meeled koos 1010 bit/s aga teadvustatud taju on 100 bit/s (infovool ehk kanali
mahtuvus) .NB! 1ms üks AP
Kommunikatsioon – info ülekande protsess ja selle osad.
Müra - informatsiooni kulgemise häired infokanalis
Redundants ehk liiasus- see osa sõnumi infohulgast, mille võib eemaldada, ilma ,et
läheks kaduma sõnumi sisu e essentsiaalne informatsioon. Redundants on vajalik
ülekandmisel tekkivate häirete tõttu info kadumamineku vältimiseks.Keele ja
210
inimkõne puhul 50%. Geneetilise koodi liiasus- 64 võimalikku koodi 20 aminohappe
jaoks. Redundants närvisüsteemis avaldub nii, et sama sõnum
saadetakse edasi mitme
ar 2
u
närvi kaudu.
aan
a, j
niiLi-
aal
M
3
3
Näide telefoni põhjal: Info allikas on kõne , kodeeriv
vahendaja on telefoni mikrofon,
ülekandekanal on
kaabel või lained, vastuvõtja on teise telefoni
valjuhääldi ning info
kasutaja on
kuulaja . Häireallikas- mobiiltelefonil levi puudumine, taustamüra.
Närvisüsteemis on infoallikas
ärriti nt nõel. Vahendaja on aktsioonipotentsiaalide
sensoorne muster, mille tekitab
sensor . Ülekandekanaliks on närvikiud (aferentne
närv). Vastuvõtja on
sünapsid kesknärvisüsteemis ja kasutaja on tsentraalne sensoorne
süsteem.
Info ülekandekanalid:
Maksimaalne infovool
kanalis e kanali mahtuvus C=Imax/t. Bitt/s on ühik.
On NEURONAALSED INFORMATSIOONIVOOD(objektiivne mõõtmine nt AP tekke
mõõtmise põhjal), sõltub aferentse närvi võimest erutust edasi kanda ja
PSÜHHOFÜÜSIKALISED INFORMATSIOONIVOOD.(kas KNS teadvustati muutust),
info jõudis teadvustatud tasemele.
21
Ülekande olulisemad viisid : Nägemine,
kuulmine , naha meeled.
0
ar 2
u
Informatsioonteooria meelefüsioloogias. Informatsioon
kodeeritakse retseptoris
aan
närviimpulsside jadaks . Füüsikaliselt mõõdetavad ärritite parameetrid näiteks
a, j
nii
nahale avaldatava rõhu tugevus
L
i-
ärriti asukoht keha perifeerias
aal
valguse ja heli lainepikkus
M
3
4
muudetakse selliselt, et informatsioon „
ärriti tugevus“ kodeeritakse
informatsiooniks „
Närviimpulsside keskmine sagedus.“
Esineb kõigil modaalsusega sensoritel:
lihasekäävidel
naha rõhuretseptoritel
keele kemoretseptoritel
võrkkesta fotoretseptoritel.
Närvisüsteemis edastatakse sama sõnum mitme närvi kaudu- saavutatakse
redundants, mis kompenseerib kodeerimise häired sensoris.
40) Meele- ehk sensoorsete süsteemide vahendusel toimuva infoülekande üldised seaduspärasused ja põhietapid. J. Mülleri „meeleelundite spetsiifilise energia
seadus“. Sensoorsetes süsteemides ülekantavad infohulgad. Meelesüsteemi osad on
SENSOR
JUHTETEE KESKUS KESKNÄRVISÜSTEEMIS
Meelesüsteem on võimeline
infot vastu võtma
infot
töötlema infot
edastama Kuigi meelesüsteemid on spetsiifilised, kuid ülekande põhimõtted on sarnased.
Infoülekande põhietapid:
Kõigepealt sensor saab stiimuli. Närvi aferentse otsa rakkude membraanide läbilaskvus
ioonidele muutub, tulemusena närvi lõpposa membraanipotentsiaal muutub
(TRANSDUKTSIOON!), genereeritakse aktsioonipotentsiaalid
(
TRANSFORMATSIOON ). See toimub juba
juhtetees . AP-d saadetakse
kesknärvisüsteemi ja integreeritakse seal (KESKUSes).
210
ar 2
u
aan
a, j
niiLi-
aal
M
3
5
210
ar 2
u
aan
a, j
n
Meelesüsteem ehk sensoorne süsteem võtab vastu, edastab ja töötleb teatud liiki
iiLi-
informatsiooni organismi välis- või sisekeskkonnast. Meelesüsteemiosad on SENSOR,
aal
JUHTETEE ja KESKUS KNS-s.
M
3
6
Mülleri meeleelundite spetsiifilise energia seadus- aistingu
laadi määrab ära
meeleelund, mida ärritatakse , mitte ärrituse iseloom. Adekvaatne ärriti tekitab vastuse
kõige väiksema energiakuluga. Nt silma puhul valgus. Kuigi vastuse tekitab ka löök,
hõõrumine,
elektriimpulss . Nt mentool mõjub keele külmaretseptoritele ja on
ebaadekvaatne signaal.
Kõik meeled koos 1010 bit/s. Üks närvikiud kuni 1000 bit/s.(1ms üks AP!!). Teadvusega
tajutud u 100 bit/s.Psühhofüüsikalised infovood : nägemine 40, kuulmine 30, nahk 5.
41) Sensorite talitluse üldpõhimõtted. Transduktsioon sensorites. Sensoripotentsiaali omadused. Sensoripotentsiaali transformeerumine
aktsioonipotentsiaalide jadaks. Ärriti tugevuse kodeerimine meelesüsteemides.
Adaptatsioon sensorites, selle biofüüsikalised mehhanismid ja bioloogiline
tähendus. Sensorite jaotus adaptatsiooni kiiruse alusel. Sensor on raku või rakumembraani osa, kus tekib sensoripotentsiaal, mis
juurdekuuluvates aferentides aktsioonipotentsiaalideks ümberkodeeritakse.
On
kesksed ja
perifeersed sensorid . Kesksed sensorid on
kõrvad , nina,silmad,keel .
Perifeersed:
kemoretseptorid
termoretseptorid Osmoretseptorid
Baroretseptorid propriotseptorid (
kehahoiak )
muud mehhanoretseptroid.
asukoha järgi enterosensorid
eksterosensorid
stiimuli olemuse põhjal nt kemoretseptorid jms,
stiimuli kauguse põhjal distantssensorid
kontaktsensorid jne.
On
vabad närvilõpmed (müeliiniga ja ilma, inkapsuleeritud on ka olemas nt
Vater -Pacini
21
kehake ) ning
spetsiaalsed retseptrorrakud nt kõrvas
0
ar 2
u
Retseptorrakud, kui neid aktiveeritakse, vabastavad neurotransmitterit, mis initsieerib
aan
aktsioonipotentsiaali.
a, j
niiL
i-
aal
M
3
7
Sensorite põhilised omadused:
sensorit aktiveerivad vaid teatud laadi
stiimulid , see on kvalitatiivne aspekt. On
adekvaatsed ja mitteadekvaatsed stiimulid. kui sensor on tundlik teatud stiimulile,
siis on tal selle suhtes mada lävi. Mitteadekvaatse puhul on lävi kõrge.
Iseloomustab ka lävi ehk
kvantitatiivne aspekt- stiimuli tugevus peab
ületama teatud kriitilise väärtuse ehk lävi, et aktiveerida retseptorit.
Sensoritele on omane TRANSDUKTSIOON- stiimuli energia
konverteerimine bioelektriliseks
protsessiks , mida KNS saab töödelda.(ehk pmt stiimuli energia
põhjustab membraani mehaanilise muutuse).
Sensoripotentsiaal levib ja hüpopolarisatsioonienergia lükkab kanali lahti.
Lisaks on iseloomustav omadus ka
ADAPTSIOON e
muudetav tundlikkus e lävi
on muudetav.
Erutuse vähenemine aja jooksul muutumatu tugevusega ärriti
korral.
Retseptrorrakkude puhul räägitakse
retseptoripotentsiaalist e muutustest
membraanipotentsiaalist, mis mõjutavad neurotransmitteri sekretsiooni retseptorraku
poolt ning see mõjutab seotud
sensoorse neuroni elektrilist aktiivsust.
Adaptsioon- sensorid ja
sensoorsed süsteemid on
muudetava tundlikkusega.
Stiimul e ärritaja kutsub sensoris esile membraani läbilaskvuse muutuse(nt
mehaaniline kanalite avamine) ja sellega SENSORIPOTENTSIAALI. Selline membraani
läbilaskvuse muutus on transduktsioon.
Sensoripotentsiaal levib elektrotooniliselt
mööda membraani ja hüpopolarisatsioonienergia lükkab kanali lahti.
Sensoripotentsiaal peab jõudma
päästikutsiooni. Sensoripotentsiaali teke e
transduktsioon (e stiimuli energia konverteerimine bioelektriliseks protsessiks).
Sensoripotentsiaal-aktsioonipotentsiaal transformatsioon toimub tavaliselt aferentse
närvikiu esimesel Ranvier´i soonisel.
Sensoripotentsiaal on
astmeline.
Amplituud ning
kestus olenevad stiimuli amplituudist
ja kestusest, ei kehti seadus „kõik-või-mitte-midagi.“ SP-d võivad ajaliselt ja
ruumiliselt summeeruda. Järgnevalt toimub TRANSFORMATSIOON, kus
sensoripotentsiaal muudetakse aktsioonipotentsiaalide jadaks. Suurema amplituudiga
sensoripotentsiaal põhjustab suurema sagedusega aktsioonipotentsiaali ning sünapsis
vabaneb rohkem mediaatorainet. Sensoripotentsiaal võib olla nii HÜPO-kui
HÜPERPOLARISEERIV.Sellest sõltub impulsside sageduse muutus sensoorses
närvikius. Hüpopolarisatsioon- suurem osa signaale. Sensoripotentsiaalid tekitatakse
närvilõpmetes enestes, mitte närvilõpmeid ümbritsevates ja
meeleelundi koostisesse
kuuluvates rakkudes. Sensoripotentsiaal on
lokaalne potentsiaal,mis levib mööda
membraani elektrotooniliselt ja mida
aktiivselt edasi ei juhita.
210
Adaptsioon sensorites- erutuse vähenemine aja jooksul muutumatu tugevusega ärriti
ar 2
u
korral.On kiiresti ja aglaselt adapteeruvad sensorid.
aan
a, j
Toonilised
retseptorid adapteeruvad aeglaselt. Saadavad KNS-i signaale nii kaua,
nii
kui stiimul kestab. Tavaliselt retseptorid, mis jälgivad parameetreid, mis peavad
L
i-
keha poolt püsivalt kontrollitud olema. Nt mõned proprioretseptorid,
aal
rõhutundlikud baroretseptorid.
M
3
8
Faasilised adapteeruvad kiiresti. Kui stiimul jõuab stabiilse väärtuseni, retseptorid
signaali enam ei saada. Nt
regio olfactiora retseptorid. kontsentreerumine uuele,
organism ignoreerib infot, mis on juba hinnatud ja leitud mitte ohustamas
homeoöstaasi või heaolu.
Mis juhtub adaptsiooni ajal?
Olenevalt retseptori tüübist on erinevaid variante. Mõnel juhul K+ kanalid retseptrori
membraanis avanevad ning seeläbi membraan repolariseerub. Mõnel teisel juhul
lisastruktuurid vähendavad stiimuli intensiivsust. Näiteks kõrvas lihased
kontraheeruvad ja vähendavad väikeste luude vibratsiooni vastuseks valjudele
helidele. Ainuke viis uut signaali luua, on muuta ärritaja intensiivsust.
Adaptsioon võimaldab ka kanda
riideid - nahal olevad retseptorid adapteeruvad riietele, ka
silmad adapteeruvad
valgusele üsna kiiresti.
2
1
0
ar 2
u
aan
a, j
niiLi-
aal
M
3
9
42) Silma optilise süsteemi põhiosad ja omadused. Kujutise teke silma võrkkestal. Silma optiline süsteem on läätsede süsteem, mis projitseerib võrkkestale ümbruse vähendatud
ja
ümberpööratud kujutise. Silma dioptriline
aparaat koosneb läbipaistvast sarvkestast,
vesivedelikuga täidetud ees- ja tagakambrist, silmaava e
pupilli moodustavast vikerkestast,
läätsest ja klaaskehast (läbipaistev
geel ). Lisaks on kujutise tekkel oluline
silmamuna tagumist
sisepinda
kattev võrkkest ehk
reetina , kus on sensorrakud kepikesed ja
kolvikesed .
(võrkkesta ümber on soonkest, selle ümber
kõvakest )
Optiline tugevus D=1/f dioptriat. Silma keskmine optiline tugevus on
58,6 dioptriat.
Silma tööpõhimõte on sarnane kaamerale. Silm koondab valgust läätse abil (see lääts on
muudetava fookustugevusega)
valgustundlikule pinnale reetinale, samuti on olemas lääts
ning avaus e
pupill , millega saab reguleerida valguse hulka.
Nagu igal meelesüsteemil, on ka optilisel süsteemil
sensorid (sensorid silmapõhjas e reetinal)
21
juhteteed (aferentne närv nervus opticus)
0
keskus (aju kuklasagar, enne toimub veel ümberlülitamine taalamuses).
ar 2
u
aan
a, j
Tegelikult on silm liitläätsede süsteem. On palju erinevaid silmamudeleid, võib vaadelda
nii
nii, et valgus murdub kahes kohas : õhk-sarvkest ja lääts-klaaskeha (
Gullstrandi mudel)
L
i-
,meie kasutame
REDUTSEERITUD SILMA mudelit, kus toimub üks
murdumine , murdja
aal
on lääts, arvestatakse üht PEATASANDIT ja
KESKPUNKTI .
M
4
0
Omadused:
Adektvaatne ärriti on valgus.
Silm kui optiline süsteem registreerib valgust lainepikkuse vahemikus u 400 ja 760 nm
summaarne optiline tugevus 58,6 dioptriat (lõpmata kaugel asuva objekti
vaatamisel tekib terav kujutis võrkkestale, vahemaa
sarvkesta pooluse ja
fovea centralise(teravaima nägemise punkt) vahel on
24,4 cm,
kui silmamuna on pikem,
tekib kujutis võrkkesta ette- lühinägelikkus)
reguleeritav, liitläätsede süsteem.
Silm suudab eristada objektide
o Suurus
o Kuju
o
Heledust o värvust
o ruumilist paiknevust
o objektide liikumist
Fototoopilisel nägemisel nähakse kõiki
5 komponenti,
skotoopilisel värvusi ei eristata.
Ruumiline nägemine eeldab kahe silma koostööd.
Nägemisel on kolm
etappi -
1) valgus siseneb silma ning fokuseeritakse läätsega reetinale e VÕRKKESTALE.
2
2) Reetina fotoretseptorid muudavad
valgusenergia eletkrisignaaliks
1
0
ar 2
3)Elektriliste signaalide töötlemine ja transport närvisüsteemis
u
aan
Kujutis tekib vähendatud ja ümberpööratud. 2 tähtsat etappi: pupill muutub, et reguleerida
a, j
n
sissetuleva valguse hulka. Pimedas laiem ,valges kitsam,
lähedale vaadates kitsam, kaugele iiLi-
laiem. ning lääts muutub (AKOMMODATSIOON) ,et muuta fookuskaugust. Lähedal asuva
aal
objekti korral lääts muutub kumeramaks.
M
4
1
Kujutis on vähendatud ja ümberpööratud. 1/f= 1/g+1/b. Läätse fookuskaugus on f, g meetri
kaugusel on objekt, saame kujutise b meetrit läätsest tagapool.
Kujutis tekib võrkkestale e reetinale, kus asuvad sensorrakud e fotoretseptorid e
KEPIKESED ja KOLVIKESED. Suhe 20:1 .Nägemisel on kaks tüüpi: SKOTOOPILINE ja
FOTOOPILINE. Skotoopilise abil saab näha hämaras, vahendatud
kepikeste poolt.
Fototoopilise abil saab näha suurema
valgusega tingimustes, vahendavad kolvikesed. On
eri
tüüpi kolvikesed, registreerivad sinist, rohelist, punast. Kepikeste
sensoripotentsiaal on
hüperpolariseeriv. Pimeduses on Na-kanalid avatud, puhkepotentsiaal
-40 mV, valges Na-
kanalid suletakse, potentsiaal muutub
-70mV. Sensorrakud on reetina viimane
rakukiht .
FOVEA
CENTRALIS on punkt, mis näeb kõige teravamalt- seal valgus langeb
otse
kepikestele ja kolvikestele ja PIMETÄHNi langevat valgust ei detekteerita üldse- sealt
väljub
nägemisnärv .
43) Silma optilise süsteemi regeleeritavus. Akommodatsioon. Silma optiline süsteem lühinägevuse ja kaugelenägevuse korral. Lühinägevuse ja kaugelenägevuse
korrigeerimise võimalused. On kaks regulatsiooni
varianti :
pupillireaktsioonid, kus lähedale vaatamisel ning suure valgushulga korral on pupill
kitsam. Pupilli suurust reguleeritakse
iirise suuruse muutmise abil seal leiduvate
lihaste abil. Pupilli ahenemisel suureneb teravussügavus.
akommodatsioon, kus muudetakse läätse
kumerust - lähedale vaatamiseks peab lääts
olema kumeram, kui ei oleks akommodatsiooni, tekiks kujutis lähedale vaatamisel
võrkkesta taha. Lääts muutub kumeramaks, kui
tsiliaarlihased kokkutõmbuvad, sest
siis läätse ümbritsev tsiliaarlihaste ringkiht kitseneb ning läätse ümbritsevad
ligamendid pikenevad ja lääts on ümaram (pole nii palju välja venitatud) Normaalne e
EMMETROOPNE silm.
MÜOOPIA E LÜHINÄGELIKKUS. Kujutis tekib võrkkesta ette, (silmamuna liiga pikk)
korrigeeritakse hajutava e negatiivse e nõgusläätsega.
HÜPEROOPIA E KAUGNÄGELIKKUS .Kujutis tekib võrkkesta taha (silmamuna liiga
lühike), korrigeeritakse koondava e postiivse e kumerläätsega.
Kumerlääts teeb silma
dioptriad
suuremaks .
21
44) Heliärritaja füüsikalised omadused ja põhimõisted: helisagedus, helirõhk, toon, 0
kõla, müra. ar 2
u
aan
Kuulmisel tajume
energiat, mida kannavad
helilained . Heli on elastse aine nt õhu
a, j
molekulide lainena edasikanduv
võnkumine ehk siis keskkonna võnkumine, kus
laineharjad nii
on keskkonna kokkusurutud piirkonnad.
L
i-
aal
M
4
2
Heli on meie interpretatsioon nende lainete
sagedusest (mitu lainet sekundis),
amplituudist ja
kestusest. Helilainete levimiskiirus õhus on u 300 m/s ja inimene kuuleb
helisid 20Hz kuni
20 kHz.
Heli tekitavad keskkonnas võnkuma pandud
kehad, mis panevad võnkuma nende vahetus
ümbruses olevad õhumolekulid, millelt võnkumine kandub naabermolekulidele jne.
Helisagedus- võnkumise kiirus.
Kiiremal võnkumisel tekib
kõrgem heli ja
aeglasemal ehk
väiksemal sagedusel
madalam heli.
Helirõhk, mida heli avaldab pinnaühiku kohta, tihedamad piirkonnad e laineharjad
põhjustavad suurema rõhu.
Mõõtühik on Pa (N/m2). HELIRÕHU TASET
väljendatakse
detsibellides(dB), mille saamiseks võrreldakse mõõdetud helirõhku Px
kokkuleppelise helirõhuga P0. Kokkuleppeline rõhutase on
kuulmislävi 1000 Hz
sageduse korral. Tavaline vestlus on u 60 dB. 80dB võib juba tundlikumaid
retseptoreid kahjustada.
Toon-ainult ühest sagedusest koosnev heli.
Kõla- põhitoon ja mitu ülemtooni, mille sagedused on põhitooni täisarvkordsed.
Müra- heli, mis sisaldab
peaaegu kõiki kuulmispiiri sagedusi.
210
ar 2
u
aan
a, j
niiLi-
aal
M
4
3
Helitugevus on ajaühikus pindalaühikut läbiv helienergia.
Kuulmisaistingu psühhofüüsika põhimõisted: kuulmislävi, helivaljus, helitugevuste eristamislävi, sageduste eristamislävi Kuulmislävi- HELIRÕHU tase ,millest alates helid on kuuldavad. Sõltub helisagedusest.
210
Kõige tundlikum on
kõrv 2000-5000 Hz juures. Kuulmisläve määramise meetod on
AUDIOMEETRIA.
ar 2
u
aan
Helivaljus- subjektiivne, kuid võimalik kvantitatiivselt kirjeldada. Väljendatakse
a, j
foonides
. Põhineb helirõhutasemel. Mõõtühikuks kasutatakse 1000 Hz juures sama
niiL
valjusega tajutud heli helirõhu TASET. Nt kui
katseisik reguleerib 1000Hz tooni
i-
reguleerinud 70 dB peale, on heli 70 fooni.
aal
M
4
4
Helitugevuste eristamislävi- väikseim subjektiivselt
eristatav helirõhu taseme erinevus.
on väga madal. Sama sagedusega toone kuuldakse eri valjusega juba siis, kui
helirõhutasemete erinevus kuulmisläve piirkonnas on 3-5 dB. Valjemate helide
puhul(+40dB) 1 dB.
Sagedust eristamislävi- Optimaalpiirkonnas 1000 Hz lähedal on eristuslävi u 3 Hz e
0,3%.
45) Heli ülekanne väliskeskkonnast sisekõrva ja selle muundumine sensoripotentsiaaliks. Kuulmine koosneb
energia muundumisest
õhulainetest mehaaniliseks vibratsiooniks
vedelikuvibratsioonideks
keemilisteks signaalideks
lõpuks aktsioonipotentsiaalideks Helilained levivad väliskeskkonnast mööda kuulmekanalit kuni nad jõuavad membrana
tympanica ehk kuulmekileni ning panevad selle võnkuma. Mehaaniline võnkumine
kandub vasarale, alasile, jalusele. (malleus,
incus , stapes) Need kolm luukest
võimendavad vibratsioonijõudu, nii et helienergiat vähem kaotsi läheks. Väga tugevate
helide puhul võivad keskkõrva väikesed lihased
luid rohkem pingule tõmmata, nii et
nende võnkumine väheneb. Kui järjekorras viimane kuulmisluuke e jalus vibreerib,
muutuvad
ovaalakna vibratsioonid vedelikuvibratsioonideks
cochlea ehk teo kanalites. Sellel üleminekul tekivad helienergia peegeldumiskaod, nende
vähendamiseks –
kuulmekile ja jaluse plaadi pindalade erinevus suurendab rõhku
umbes 17 korda. Rõhk suureneb lisaks tänu kangiõlgade pikkustele kuulmeluukeste
ahelas. Vedelik, liikudes mööda cochlea kanaleid, aktiveerib
karvarakud e
sensorrakud. Aktiveeritud karvarakud vabastavad neurotransmitterainet primaarsetele
cochleas olevatele neuronitele, kus
neurotransmitter põhjustab aktsioonipotentsiaalide
tekke. Cochlea e
tigu koosneb kolmest kanalist- ESIKUASTRIK ,TRUMMIASTRIK,
TEOJUHA . Trummiastrik ja esikuastrik on omavahel ühendatud
teomulgu kaudu.
Esikuastrikku ja trummiastrikku täidab PERILÜMF.- sarnaneb rakuvälisvedelikuga,
palju Na+. TEOJUHA täidab ENDOLÜMF, kus on palju K+ TEOJUHAS on
CORTI organ (teojuha ja trummiastriku vahel), mis asub basilaarmembraanil ning on kaetud
kattemembraaniga, Corti
organis asuvadki karvarakud. Kui kattemembraan liigub
vedeliku liikumise tõttu, liiguvad ka karvarakkude
karvakesed , mis on omavahel
ühendatud
proteiinisildadega, mis avavad ja sulgevad ioonkanaleid ja sellega
muudavad membraanipotentsiaali. Kui karvakesed painudvad ühes suunas,
21
avanevad kanalid,
0
katioonid tulevad rakku ,
ar 2
u
neurotransmitterit vabaneb ning
tekib AP närvirakkudes.
aan
a, j
nii
Mida rohkem kanaleid avaneb, seda rohkem katioone rakku, seda suurema
L
i-
amplituudiga sensoripotentsiaal ja seda suurema sagedusega aktsioonipotentsiaalid.
aal
M
4
5
Teises suunas paindumine
suleb ioonkanalid ja
rakk hüperpolariseeritakse. Ovaalaken→ümaraken (E
kandumine )
Mida rohkem rakk
paindub , seda rohkem kanaleid avaneb, seda suurem sensonripotentsiaal,
seda sagedasemad aktsioonipotentsiaalid.
NB.Tigu paikneb sisekõrvas, kus asub ka tasakaaluorgan.
Kuulmismeele keskus on OIMUSAGARAS.
46) Homöostaas : mõiste, kontseptsiooni kujunemise ajalugu (C. Bernardi ja W. Cannoni tööd), olulisemad homöostaatilised parameetrid. Homöostaasi printsiibi
tähtsus bioloogias, meditsiinis, psühholoogias jm. Homöostaas on bioloogiliste ja küberneetiliste süsteemide
võime säilitada neis toimuvate
protsesside tasakaalu ning vältida süsteemi ohtlikke kõrvalekaldeid.
Organisimi ekstratsellulaarse vedeliku teatud füüsikaliste ja keemiliste omaduste püsivus-
suhteliselt stabiilse sisekeskkonna hoidmine.
Sisekeskkonna ideed arendas
füüsik Claude
BERNARD 19.sajandi keskpaigas: oma
uuringute käigus märkas ta, et mitmed füsioloogilised parameetrid on stabiilsed-
keha
temperatuur,
südame löögisagedus,
vererõhk.
Tema teoorias oli stabiilsusel nii struktuurihoidmise kui
kohastumuslik tähtsus. Kirjutas
sisekeskkonnast raamatu.
HOMÖOSTAASi mõiste võttis kasutusele W.B.
Cannon aastal
1929,
et kirjeldada stabiilse sisekeskkonna regulatsiooni.
Homeo- sarnane,
stasis - seisund. Cannoni seisukoht oli see, et füsioloogilised protsessid
,mis tagavad
stabiilsuse on nii komplitseeritud ,et on vaja eraldi terminit- homöostaas.
Olulisemad homöostaatilised parameetrid (jaotuse tegi Cannon):
A) Rakkudele vajalikud ained:
-vesi
-hapnik
210
-glükoos, proteiinid, lipiidid
-Naatrium,
kaalium , kaltsium ja teised rakule vajalikud anorgaanilised ained
ar 2
u
Jne
aan
a, j
B) Rakke ümbritseva keskkonna omadused
niiL
-pH e vesinikioonide kontsentratsioon
i-
-osmootne rõhk
aal
M
-temperatuur
4
6
Kõik piirkonnad eraldi
võetuna ei ole stabiilse koostisega, küll aga
tervikuna .
Homöostaatiliste näitajate tasakaalu hoidmisel on väga tähtis
tasakaalustatud bilansi
põhimõte, mille kohaselt on parameetrit võimalik hoida samal tasemel vaid siis, kui
selle suurenemist ja vähenemist põhjustavad mõjud on sama suured. Hulkrakses
organismis on parameetri väärtuse suurendajad ja vähendajad ruumiliselt lahutatud,
seega on vajalik
regulatsioon üle terve organismi.
Näiteks veresuhkru regulatsioon. Pärast söömist veresuhkru tase vereplasmas tõuseb-
osa läheb maksa hoiule glükogeenina. Kui sinna ei mahu,siis muudetakse
rasvamolekulideks. Ka vesi kehas- lui on liiga palju vett siis vähendatakse
ultrafiltratsiooil tagasiimetava vee hulka, suurendatakse higistamist. Keha ütleb ajule,
et pole rohkem vaja juua. Vastupidi näide ka.
Pool- ekstratsellulaarne
maatriks . Skeemi tähtsus seisneb selles, et kui kehas on mingit
ainet liiga palju, siis selle aine
2
sissetulekut vähendatakse ja
1
0
väljutamist suurendatakse. Nt
glükagoon ja insluiin.
Glükagoon suurendab
ar 2
veresuhkru sisaldust ja
insuliin vähendab.
u
aan
Homöostaasi printsiip on tähtis
a, j
n
bioloogias- aitab hoida
Gibbsi vabaenergia kõrge ning annab nii eluks
iiLi-
vajalikku energiat. Näiteks kui kehatemperatuuri ei reguleeritaks, sureksid
aal
kõrbeloomad valkude denatureerumise tõttu.
M
Meditsiinis saab regulaatortoimet
asendada või muuta, kui teame, kuidas keha
ise reguleerimishäireid muudab.
4
7
Psühholoogias nt „obsession-fixation“- juhtsuurus on teatud asja liigsus, kui
asendame selle
teisega kujuneb välja ahnimismaania
aseaine suhtes.
47) Organismi funktsioonide reguleerimine. Regulatsiooni vajadus, tasemed ja eesmärgid. Organismi funktsioonid
teatud seisundi hoidmiseks või
muutustega kohanemiseks.
kehatemperatuur, vererõhk, kehaasend raskusjõuväljas.
Hulkrakses organismis on parameetri väärtuse suurendajad ja vähendajad ruumiliselt
lahutatud, seega on vajalik
regulatsioon üle terve organismi. Reguleerimisülesandeid täidavad peamiselt
Närvisüsteem
Hormoonid nt insuliin, mida toodab pankreas, reguleerib veresuhkru taset, sellel
regulatsioonil on organismis kriitiline tähtsus.
Regulatsiooni alla kuuluvad juhtimisprotsessid (teatud muutujate muutmine kindlale
eesmärgile orienteeritud viisil), mis aitavad teatud parameetreid kehas hoida vajaliku
kitsa väärtustevahemiku juures.
Ajalise
dimensiooni alusel on
Kiired nt oodatud teate saabumine- südame löögisageduse kasv
Aeglased regulatsioonimehhanismid
Ruumilise dimensiooni alusel on
lokaalsed (nt ühel jalal hüppamisel teatud lihased)
üldised(kogu keha
haaravad nt vererõhu muutus),
Vahendilise dimensiooi alusel on
neuraalsed(närvid ja AP)- kiired ja hästi lokaliseeritud.
humoraalsed (hormoonide abil)
regulatsioonid - aeglased, kogu organismi mõjutavad.
Reguleerimisel on alati kolm komponenti:
21
1. stiimul või muutus reguleeritavas muutujas
0
2. rakk või kude, mis
hindab stiimulit ja algatab vastuse ar 2
u
3. rakud või
koed , mis viivad läbi vastuse
aan
a, j
Regulatsiooni on vaja, et
niiLi-
tagada vajalikud tingimused ettekodeeritud arenguks
aal
M
adaptsiooniks muutuvatele keskkonnatingimustele
4
8
tagada homöostaas ja kõrge Gibbsi vabaenergia , madal
entroopia Tasemed:
-membraan- nt pH mitokondri membraanis. Õige tase vajalik ATP sünteesiks
-organell- nt tärklisesisaldus taimeraku vakuoolis
-rakus- sünteesitud valkude eksotstütoos, pH
-kudedes- Ca2+ kontraheeruvas lihases.
-organis
- pH ja
ensüümide eritus (
maks ja pankreas)
-organsüsteemis-
seedeelundkonna verevarustuse kasv pärast
sööki , suguhormoonide mõju
-organism- pH ,osmootne rõhk, temperatuur
Veelkord eesmärkidest : hoida Gibbsi vabaenergia kõrgel, kui ΔG=0 ,siis on surm., arenguks
vajalike tingimuste tagamine ja kohanemiseks vajalike tingimuste tagamine.
48) Reguleerimiskontuur, selle põhiosad ja põhimõisted. Negatiivse ja positiivse tagasisidega reguleerimiskontuurid, nende põhiomadused ja regulatsiooni
iseloom. Reguleerimiskontuuri talitluse stabiilsus ja ostsillatsioonid. 210
ar 2
u
aan
a, j
niiLi-
aalM
Reguleeritav suurus on seisund, mida tuleb hoida konstantsena. Reguleeritav süsteem on
süsteem, mida iseloomustab seisund, mida tuleb hoida konstantsena.
Andur mõõdab
4
9
tegelikke väärtusi.
Regulaator võrdleb
tegelikku väärtust
juhtsuurusega, mis on
reguleeritava muutuja nõutav väärtus ehk seadepunkt ehk set point. Reguleertoime
leiab aset nii kaua, kuni juhtsuurus ja tegelik suurus üksteisest enam ei erine. Häiringu
suurus on tegur, mis põhjustab reguleeritava suuruse kõrvalekalde
juhtsuurusest.Reguleerimishälve on nõutava ja tegeliku suuruse vahe.
Näide kehas: temperatuuri regulatsiooni puhul sensorid on termosensorid . Keskus on
termoregulatsioonikeskus
hüpotaalamuses. Efektorid- veresoonte silelihased,
vöötlihased, higinäärmed. Häiring võib olla nt akna avamine.
Mittebioloogiline (tehniline) näide: Reguleeritav süsteem on
akvaarium , täpsemalt
21
akvaariumi vesi. Regulaator hoiab temperatuuri 30 C juures toas, kus temperatuur on
0
25 C. Juhtsuurus e nõutav suurus on 30C. Oletame, et alguses on vee temperatuur ka
ar 2
u
25 C.
See madal veetemperatuur on häiring. Lülitame
kontrollkarbi e tsentraalse aan
regulaatori sisse.
Juhet mööda tuleb talle signaal, et vesi on liiga külm- juhe on
a, j
aferentse tee
analoog . Tsentraalne reguleerija otsustab, et on vaja veetemperatuuri
nii
tõstvat vastust. Kontrollkarbi ja kuumutaja vahel on juhe –
eferentne tee. Kuumutaja on
L
i-
efektor.
aal
M
5
0
Nõutav väärtus e set point e SEADEPUNKT võib olla konstantse või muutuva
väärtusega.Näiteks palaviku puhul on temperatuuri nõutav väärtus kõrgem kui
normaalselt. pH ja osmootne rõhk on üsna püsivate nõutavate väärtustega.
Vt eespool regulatoorsete mehhanismide jaotus.
On kaks süsteemide klassi:
Avatud süsteem e lihtne süsteem– väljund ei mõjuta otseselt süsteemi ennast või
sisendit sisend→süsteem→väljund
Tagasisidesüsteem sisend→süsteem→väljund
on positiivse ja negatiivse tagasisidega süsteemid.
Negatiivne tagasiside on
homöostaatiline.Sellega hoitakse parameetrit üsna stabiilsel
tasemel. Vastus eemaldab signaali või vastandub signaalile. Nt kui glükoosi tase
kasvab, siis vastus on insuliini tootmine pankreases, mis muudab glükoosi
glükogeeniks ning glükoosi tase langeb.
Positiivne tagasiside-
vastus suurendab
vastust algsele signaalile
eksponentsiaalselt.Süsteem on
ajutiselt kontrolli alt väljas. Süsteem viiakse
maksimaalse kiirusega maksimaalse aktiivsuse seisundisse. On vaja välist mõjutust, et
sellist „surnud ringi“ peatada. Nt aktsioonipotentsiaali ajal Na+ rakku saabumine.
Väike hüpopolarisatsioon avab pingest sõltuvad Na kanalid, see avab üha uusi
kanaleid. Väline sekkumine on teiste väratite
sulgemine . Samuti ka
sünnitus , mis
initsiateeritakse sellega, et
beebi on raske ja vajub alla.
Emakakael venib -
oksütotsiin vabaneb- kontraktsioon- beebi allapoole- emakakael venib- rohkem oksütoksiini.
Regulaatorkeskuse tegevuse
edu sõltub tema
tundlikkusest. Enamasti parameetri väärtus
kõigub nõutava väärtuse ümber. Tundlikkus on erinev, nt osmootilise rõhu sensorid
tajuvad juba 3% vere kontsentreerituse tõusu e hakatakse vett kokku hoidma. Samal
ajal hapnikutaseme sensorid ei vasta enne, kui
hapnikusisaldus on vähenenud 40%
210
ar 2
u
aan
a, j
niiLi-
aal
M
5
1
Stabiilsus ja ostsillatsioonid On olemas püsi- ja järgivregulaatorid. Järgivregulaatorite set point ehk seadepunkt on
muudetav. pH ja osmootne rõhk on üsna püsivate väärtustega, kuid temperatuuri set
point võib muutuda.
Kui tekib reguleeritava suuruse ülereguleerimine suletud reguleerimiskontuuris, võivad
reguleerimise ebastabiilseks muutumisel (nt liiga suure reguleerimisvõime korral)
tekkida isegi reguleeritava suuruse sumbumatud võnkumised, kuid ka siis, kui
võnkumisi ei teki, säilib reguleeritava suuruse reguleerimishäire.
Tagasiside võib olla stabiilne või kaootilistes/regulaarsetes ostsillatsioonides.
Elussüsteemidele on omased mitmesugused
regulaarsed ostsilaatsioonid ehk
rütmid.
Iseloomustavad pea kõiki elusolendeid.
Rütmid võimaldavad:
sünkronisatsiooni
ennetavaid reaktsioone
2
ruumilist orientatsiooni
1
0
efektiivsem homöostaasi tagamine
ar 2
informatsiooni kodeerimist
u
aan
Kõikumisi tingivad:
a, j
niiLi-
1. põhjalikud muutused
elukeskkonnas aal
a. öö ja päeva vaheldumine ehk tsirkadiaansed ostsillatsioonid. On
M
arvatud et selline
perioodika on organismi passiivne
reaktsioon öö-
päeva vaheldumisele ,samas säilub perioodika ka pärast väliskeskkonna
5
2
tegurite väljalülitamist. Perioodikat põhjustab
ENDOGEENNE protsess- bioloogiline kell. Sp vastab periood ainult umbkaudu päeva
loomulikule kestusele.(TSIRKAdiaanne).
Iseerutuv ostsillatsioon.
Välised tegurid sünkroniseerivad perioodika 24h perioodikaga.
Inimesel üle 100 parameetri ööpäevased kõikumised. Kehatemperatuur
on min
hommikul ja max õhtul, nende vahe 1-1,5 C.
b. Naiste
reproduktiivne tsükkel-
lunaarkuu 2. muutused organismis endas
49) Elussüsteemide termodünaamika , termodünaamika I ja II seadus. Termodünaamika on füüsika haru, mis käsitleb soojusnähtusi,nende seost füüsikalis-
keemiliste omadustega, füüsikaliste ja keemiliste protsesside energeetilisi aspekte,
samuti määrab protsessi kulgemise suuna ja energeetilise võimalikkuse. Süsteem- ühendus, tervik.
Termodünaamiliste süsteemide jaotus:
Isoleeritud- ei vaheta energiat ega ainet
Suletud-
vahetab ainult energiat
Avatud- vahetab energiat ja ainet.
Kaks
põhilist reeglit määravad ära energiaülekanded bioloogilistes süsteemides.
Universum- lokaalne süsteem ja ümbritsev keskkond.
Termodünaamika esimene seadus- energia hulk universumis on püsiv. Energia ei teki ega
kao, vaid ta muundub ühest liigist teise. Kehtib isoleeritud süsteemis
. Inimene ei ole
isoleeritud süsteem, ta saab energiat väljast toiduna ning kaotab energiat soojusenergia
näol keskkonda. Termodünaamika teine seadus- loomulikud spontaansed protsessid kulgevad alati
entroopia e korratuse suurenemise suunas.
Korrastatuse hoidmine nõuab energiat.
Korrastatus tekib, kui avatud süsteemid
kaotavad energiat ilma seda tagasi saamata.
Soojus ei saa minna külmemalt kehalt soojemale. Entroopia on korrastamatuse mõõt.
Organism 2
tahab, et ained oleks suurte molekulidena- korrastatus. 1
0
ar 2
Inimorganismis kulub energia
u
aan
keeruliste ainete sünteesiks
a, j
n
mehaaniliseks tööks
iiLi-
ainete transpordikskonstantse
aal
kehatemperatuuri hoidmine
M
5
3
osa energiast hajub soojusena keskkonda.
Keerulised ained, millel on
kõrge Gibbsi vabaenergia ning madal entroopia, muudetakse
elutegevuse käigus lihtsamateks (C02,H20,NH3,CO(NH2)2), süsteem liigub tasakaaluoleku
poole, kuid see tähendab surma, sest siis ei saa organism enam tööd teha. JÄRELIKULT:
Organism töötab tasakaaluoleku vastu.
Organismi puhul ei saa me kasutada klassikalist mehaanikat, tuleb kasutada avatud süsteemi
termodünaamikat, kus tasakaaluoleku asemel on statsionaarne olek. Gibbsi energia
kompenseeritakse (Gibbsi energia väheneb, sest elutegevuse käigus organism teeb tööd)
väljast tuleva Gibbsi energiaga, toidu G on kõrge. Entroopia kasvab pidevalt, kuid see
kompenseeritakse nii, et
väljast sisse tulevad keerulise struktuuriga ained, mille entroopia on madal
organism sünteesib ise selliseid madala entroopiaga aineid
kõrge entroopiaga elutegevuse produktid eemaldatakse organismist
Organismi madal entroopia tuleb keskkonna entroopia kasvu arvelt.
Summaarselt toimub Gibbsi energia vähenemine. (sest iga reaktsiooniga, kus G on positiivne,
on seostatud mõni teine reaktsioon, kus G on negatiivne)
Üldine Gibbsi energia ja entroopia tase organismis on püsiv.
Tasakaal soojuse tekke ja äraandmise vahel-
termoregulatsioon.
Suurem inimene toodab rohkem soojust. Keha pindalast oleneb soojuse äraandmine. Kui
võtame nn lootepoosi, siis on väiksem soojuse kadu.
Inimkehas mehaanilise e kasuliku töö osakaal on 25%. Jõudsidki lõpuni! Siiralt loodan, et eksam läheb edukalt!
Aga enne kui selle faili kinni paned, tahan jagada Sinuga midagi, mis olen oma elus
õppinud ,
ja mis usun, on veel vähemalt mitu korda tähtsam kui biofüüsika või ükskõik mis muu aine,
mida oma armsas arstiteaduskonnas õpime!
210
Kas Sa oled kunagi mõelnud, KES Sa oled?
ar 2
u
aan
Kust tuleb Sinu identieet või mis teeb Sind väärtuslikuks? Võib-olla püüd olla hea sõber oma
a, j
n
sõpradele? Või on sinu elu keskmes Sinu poiss-või tüdruksõber? Vaevanägemine, et olla tubli
iiLi-
tudeng ? Tark
tulevane arst? Soov olla ilus, stiilne, sõbralik, tore, naljakas, huvitav, erinev?
aal
Või tahaksid olla nii sõltumatu, et Sinu
identiteet ei sõltu
millestki ?
M
5
4
Tihti on soov olla KEEGI, olla midagi väärt peamine motiiv, ükskõik mida ette võtame. Samal
ajal on tõsi, et mida iganes hakkame
pidama oma tähenduslikkuse allikaks, pole ükski neist
kaev, kust vesi otsa ei saaks- isegi kui oled korra tõestanud, et oled midagi väärt, kestab see
vaid hetke ja kohe tuleb hakata jälle samas suunas rühkima, niimoodi 70 või heal juhul
natuke rohkem aastat.
Oled ehk vahel mõelnud,
kellele Sa end siis lõpuks tõestad? Iseendale? Inimestele, kes
lõpuks on täpselt sama
huviga ja kellele t e g e l i k u l t ei lähe korda muu kui tunda
iseennast väärtuslikuna? Ilmselt on see juba vana uudis, et oleme isekad.
Mõtle korraks ükskõik mis probleemi peale enda elus või laiemalt ühiskonnas ja maailmas.
Nälgivad Aafrika
beebid ?
Kehv läbisaamine vanematega?
Üksindus , kadedus, viha? Kui
hakata mõtlema, siis kas pole nii, et pea kõigel, mis tundub valesti olevat, on algpõhjuseks
kellegi (tihti minu enda) isekad valikud või käitumine?
Kui vaatad enda ümber ringi ja oled aus, siis pole raske märgata, kuidas inimesed teevad
üksteisele haiget, on üksikud, pingutavad väikeste eesmärkide poole teadmiseta suuremast
või kestvamast.
Mõnikord mõtleme, kuidas olime ikka naiivsed, kui
lastena uskusime, et meil igaühel saab
olema
veatu täisväärtuslik elu, ideaalsete suhete ja karjääriga, mõtestatud ja muredeta-
tundus reaalne, et just minu elus selline unistus täitub (mõnikord
salaja loodame seda ikka
veel).
Võib-olla leiad, et miks peaks end vaevama ja üldse mõtlema sellistele asjadele.
Aga mis siis ,kui see on kõige tähtsam asi, mille peale mõelda?
Minu jaoks on hirmutav mõte, et ühel päeval oma elu lõpusirgel
vaatan tagasi ja tõden, et
kuigi aastate jooksul järjest vallutasin erinevaid mäetippe ja lootsin, et iga järgmine eluetapp
on just see, mis mind tõeliselt õnnelikuks teeb, just see, mis mu elu täisväärtuslikuks
muudab, siis tegelikkuses nii ei läinud ja
tunnen ikka veel, et olen poolel teel.
Kas sulle ei tundu mõnikord, et maailmas on midagi põhimõttelist ja põhjapanevat paigast
ära? Miks on nii, et mul on võime mõelda, milline oleks mu ideaalne elu, olla ideaalne
inimene, aga selle realiseerimise võime on nii kehv?
2
Kui keegi küsiks sinult sellise huvitava küsimuse, et mis asi on „patt“, siis ilmselt enamik
1
0
vastuseid oleks stiilis, et see on midagi, mida on „vale“ teha,
kristlaste arvates. Aga mis siis,
ar 2
u
kui selle sõna tähendus on midagi
enamat ?
aan
Võib-olla
patu tähendus ei ole ennekõike mitte halvad asjad, mida teeme, vaid see, et
a, j
võtame h e a d asjad ja hakkame uskuma, et need ongi parim võimalik, mida võime elus
niiLi-
saavutada?
aal
Hakkame uskuma, et need samad ebapüsivad väärtused, kust otsime oma identiteeti, ongi
M
need, mis peavad tegema õnnelikuks ja andma tähenduse; ja isegi kui see hästi ei toimi, siis
5
5
midagi paremat ei ole ju olemas?
Võib ju olla, et selle sama „patu“ pärast ei täida me kunagi kogu oma potentsiaali. Võib-olla
selle sama „patu“ pärast olengi
isekas , sest pean iga hinna eest pingutama, et saada neid
„häid asju“, pean pingutama, et tõestada, et olen keegi!
Kas ei või olla, et lõpuks on kõigi probleemide põhjus selles samas isekuses, selles samas
„patus“? Võib-olla see, mis on maailmas paigast ära, ongi „patu“ põhjustatud?
Tean, et võib vabalt olla, et
sinus tekitab väikest või mitte nii väikest vastumeelsust kuulda
sõnu nagu „kirik“ või „
Jeesus “ või isegi seesama „patt“, aga kas oled kunagi proovinud
vaadata eelarvamusteta seda sõnumit, mida
kristlus kõneleb?
Sõnumit sellest, et on olemas intelligentne looja, Jumal, kes on algusest peale teinud meid
nii, et otsiksime kogemust väärtuslikkusest ja tähendusrikkusest, otsiksime teadmist, et
oleme
armastatud , kuid ainuke, mis meile seda tõeliselt suudab pakkuda on suhe selle sama
meie Loojaga.
Ometi, pole raske aru saada, et iga päev me valime miljon muud asja meile rahuldust
pakkuma enne Jumalat. Püüdlus olla iseenda Jumal ja ise ennast õnnelikuks teha ongi see,
mida nimetame
patuks . Olla iseenda Jumal- võib-olla kõlab sulle väga normaalselt, aga
samas usun, et oled oma elus näinud, kuidas, tee mis
tahad , püüded iseennast väärtuslikuks
teha ja „lunastada“ lõpuks lihtsalt ei tööta.
Kirjutasin, et olen elus õppinud midagi, mida tõesti pean hindamatuks- see miski on
teadmine tõest ja sellest, kus on minu väärtus ja identiteet.
Teadmine, et kumbki neist ei sõltu mitte mingil moel minu pingutusest, saavutusest ega ka
inimsuhetest ja kuidas samal võin kõiki neid nautida ilma hirmuta, et kui nad kaovad, kaotan
osa iseendast.
Usun siiralt, et on olemas Jumal, kes mind armastab ja kelle armastus on nii suur, et
hoolimata minu isekusest ja puudujääkidest on see sama Jumal ise tulnud inimesena-
Jeesusena- maailma ja näidanud, et armastab mind nii väga, et on valmis tegema
ü k s k õ i k mida, et olla jälle suhtes minuga, parandada see, mis on olnud patu poolt rikutud.
Jumal ütleb, et patu tagajärg on surm, nii vaimses kui igas muus mõttes, see on paratamatu,
samamoodi nagu kehtivad füüsikaseadused. Ka oma ühiskonnas väärtustame õiglust, kui
keegi rikub reegleid, seadusi, siis on sellel mingisugune tagajärg,
karistus . Jumal on
samaaegselt õiglane ja armastav- Ta on kandnud patu tagajärje, surma, Kristusena
ristil , mis
2
teeb ta ühest küljest õiglaseks ja
teisalt , see sama fakt teeb Ta ka armastavaks- Jumal on
1
0
valmis ohverdama ükskõik mida, et olla jälle meiega sellises lähedases suhtes.
ar 2
u
See ongi lunastus- Jumal annab meile uue elu, mis on vaba isekuse ja patu vangist.
aan
a, j
n
Ei ole suuremat rõõmu ja vabadust kui teadmine, et olen armastatud püsiva,
kõikumatu iiLi-
armastusega, mis ei sõltu minu saavutustest. Veel enam, Jumal ütleb, et kui me võtame
aal
vastu selle lunastuse, mida ta pakub, siis ta paneb
omaenda vaimu meie sisse. Ja ainult
M
sellesama Jumala vaimu abil võid sa õppida elama seda täisväärtuslikku ja mõtestatud (kuigi
5
6
kindlasti mitte lihtsat) elu, mida sa lapsena veel uskusid võimalik olevat.
Jeesus suri Sinu asemel, et Sina võiksid elada koos
Jumalaga edasi ka siis, kui su füüsiline
keha
sureb ja nii terve igaviku. Ainuke, mis selleks tegema pead, on uskuda, et see on nii ja
lasta selle usu kaudu Jumalal end muuta, järgneda oma eluga Jeesusele.
Sul on vaba valik- Jumal ei sunni end kellelegi peale ja samamoodi ei ole selle kirja eesmärk
sulle „
ajupesu “ teha, vaid panna Sind mõtlema, mis on sinu eesmärk või mõte.
Mõned aastad tagasi olin väga skeptiline kogu selle „Jeesuse jutu“ osas, kuid samal ajal olin
ka uudishimulik ning hakkasin usku uurima, panemata endale
kindlaid piire, kuhu tahan välja
jõuda.
Igatsesin lihtsalt rohkem teada saada, mida
kristlased usuvad, tundusid nad ju
keskmisest rõõmsamad inimesed. Lugesin erinevaid raamatuid, nii kristlikke kui muid
maailmavaateid tutvustavaid. Varem olin alati arvanud, et kui peaksin uskuma
piiblit , siis
oleks see mu kaalutletud mõistuse ja
loogika surm, kuid endalegi üllatuseks hakkasin
nägema, kuidas see sama sõnum Jeesusest tegelikult võib olla nii loogiline, universaalne ja
kõikehõlmav, mitte ainult pühapäeval vanade tädikeste seas unisel jumalateenistusel!
Ometi tean ka, et kunagi ei saa ma kogu tõde oma mõistusega täielikult haarata- ma ei
tahaks järgida Jumalat, kelle mõistus ei ole minu omast suurem!
Mis pani mind
pöörama oma elus uue lehekülje ja tahtma järgneda Jeesusele, ei olnud
esmajoones intellektuaalsed kaalutlused, vaid minu enda kogemus, kuidas ükskõik, mis ma
teen- isegi, kui mul on tore perekond, parimad sõbrad, isegi kui lõpetan gümnaasiumi
parimate eksamitulemustega, isegi kui mul on
imeline poiss-sõber, kellega varsti abiellun,
jääb minu ellu üks Jumala-kujuline auk, mida ei täida ära mitte midagi muud kui suhe sellega,
kes on mind eriliselt kavandanud ja loonud, elu mis on suurem kui vaid mina ise.
Tegelikult kogu selle kirja eesmärk on küsida Sinult- kes on Jeesus Sinu jaoks?
Olen veendunud, et see on kõige tähtsam küsimus, mis määrab kõik muu! On väärt otsida
sellele vastust! Sul ei ole midagi kaotada, sest isegi kui päeva lõpuks otsustad elada ilma
Jumalata (see on Sinu vaba valik), siis oled vähemalt natuke haritum ja teadlikum kui enne,
kuid siiralt usun, et Jeesusel on Sulle rohkem öelda, kui arvata oskad ja see rohkem võib
muuta Sinu elu
parimas mõttes!
Kõige paremaga,
Maali-Liina, stud med II 210
([email protected]) ar 2
u
aan
a, j
Veel Jumalast- www.jumal.ee
niiLi-
minu
kogudus - www.3dkogudus.ee
aal
M
5
7
minu lugu-
http://vimeo.com/15127704 Raamatuid:
„The
language of God“- Francis Collins (Humane Genome
Project ´i juht)
„Lihtsalt kristlus“- C.S.
Lewis „Kaalukad tõendid“ ja „Rohkem kui
puusepp “ Josh
Mcdowell Piibel
210
ar 2
u
aan
a, j
niiLi-
aal
M
5
8










































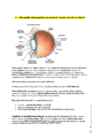























































































Kõik kommentaarid