1. Bioeemia
areng ja seos teiste teadusharudegaEsimesed sammud
biokeemias tegi
Scheele aastatel 1770.....
1786 eraldades orgaanilisi
happeid ja glütserooli. Aastatel 1770...1774 avastas Priestley
hapniku- keemilise ühendi, mida loomad
neelavad aga taimed toodavad.
Olenevalt uurimisobjektist eristatakse biokeemias kolme erinevat
suunda: staatiline, dünaamiline ja funktsionaalne
biokeemia .
Varasem biokeemia
areng oli seotud 19. sajandi keskpaiku, kui hakkas tunnustust võitma
seisukoht, et
elusorganismide keemia ei ole põhimõtteliselt erinev
eluta aine
keemiast 20. sajandi
esimesel poolel algas biokeemia kiirem areng. Võeti kasutusele
kaasaegsed analüüsimeetodid, tehti kindlaks peamised
ainevahetusrajad (O. Warburg, O. F. Meyerhof, H. A. Krebs, M. Calvin
jpt). 1944 tõestasid
Oswald Avery ja
Colin MacLeod lõplikult
nukleiinhapete seose geenidega. Järgnev biokeemia areng on toimunud
tihedas seoses
molekulaarbioloogia arenguga, olulisemateks
sündmusteks näiteks valkude struktuuri avastamine 1951 Linus
Paulingi poolt ning DNA struktuuri avastamine 1953 James Watsoni ja
Francis Cricki poolt.
Meditsiinilise
biokeemia
baasteadmised on aluseks füsioloogiale, immunoloogiale,
farmakoloogiale, farmaatsiale, endokrinoloogiale,
molekulaarbioloogia,
molekulaargeneetika ,
geenitehnoloogia ,
bioinformaatika , molekulaarmeditsiin jt uutele spetsiifilistele
arengutrendidele.
2. Keemilised
elemendid ja ühendid loomorganismisPõhibioelemendid
– põhibioelementideks on H, C, O, N, P, S (moodustavad 96...98%
elusorganismide elementaarkoostisest ja nende baasil formeeruvad
biomolekulid , raku orgaaniline aine). Nende ainete evolutsiooniline
„eelistatus“ tuleneb sellest, et nad annavad kergesti
kovalentseid
sidemeid (tugevad sidemed tagavad biomolekulide
stabiilse ehituse) ning kaksik ja kolmiksidemete võimalus on alus
mitmekesisusele ja reaktsioonivõimele;
nendest moodustuvad
organismis
vesilahustuvad anorgaanilised ühendid on kergesti
kasutaavad/väljutatavad.
Süsinik – Juhtroll
bioelementide seas tuleneb sellest, et: C-
aatom võib anda neli
ensümaatiliselt sünteesitavat/lõhustuvat kovalentset sidet kas
teiste aatomite või C-aatomiga; moodustab üksik-, kaksik- ja
kolmiksidemeid (biomolekulide
mitmekesisus !); moodustavad
lineaarseid, hargnevaid ja tsüklilisi struktuure
Hapnik –
Kudedesse jõudnud hapnikust umbes95% kasutub biomolekulide
lõhustumiseks, et
salvestada nende energiat organismi poolt
kasutatava metaboolse energia (peamiselt ATP) vormis. Umbes 2...5%
hapnikust kulub biofunktsioonideks vajalike hapniku reaktiivsete
vormide tekkeks
Vesinik – tähtsus seisneb
vesiniksidemete andmises biomolekulides.
Vesiniksidemed kindlustavad
biopolümeeride (
valgud ,
nukleiinhapped , polüoosid) kõrgemate
struktuuritasemete stabiilsuse.
Lämmastik – Esineb
aminohapetes, nukleiinhapetes ja heterotsüklilistes
lämmastikuühendites. Biomolekulised on lämmastik süsiniku-skeletti
täiendav, mitmekesistav ja reaktiivsust tõstev element.
Fosfor
– Fosfor osaleb makroergiliste sidemete moodustamises, teda leidub
nukleiinhapetes, fosfolipiidides, mitmetes koensüümides.
Väävel
– Rohkesti naha, küünte ja juuste valkudes. Biomolekulides leidub
ta aminohapete, glutatiooni, koensüüm A, vitamiinide B1
ja H, hepariini koostises. –SH rühm on tihti ensüümide
aktiivtsentris.
MakrobioelemendidKaltsium
– levinuim makrobiogeenne element kehas, ligikaudu 99% asub
luudes ja hammastes. Osaleb vere hüübimisprotsessis, lihaskontraktsioonis,
neurotransmissioonis, ensüümide aktiveerimises, vitamiini
D-metabolismis,
signaal -ülekandes, v.ere
osmootse rõhu tagamises.
Naatrium ja kaalium – Naatrium asub valdavalt
rakuväliselt – Na-
pump , mis eemaldab rakutalitluses rakku toodud
liigse naatriumi rakust. Kaalium on
rakulise lokalisatsiooniga –
liigne rakust väljuv kaalium viiakse raku tagasi Na-pumba
abil.
Naatriumi ja kaaliumine funktsionaalses koostöös
täidetavad ülesanded on: a) Na-pumba poolt loodud naatriumi ja
kaaliumi erinev
jaotumine raku ja tema väliskeskkonna vahekl on
rakkude normaalse membraanipotensiaali
tekitamise kaudu närvikoe ja
lihaskoe talitluse aluseks, b) vere osmolaalsuse
regulatsioon , c)
hape -alustsakaalu hoidmine, d) normaalne veevahetus, e)
membraanitranspordi tagamine, f) mitmete ensüümide aktivatsioon.
Magneesium – rakus 10 korda rohkem kui rakuvälises
vedelikus . Rohkesti luudes ja lihastes. Ta on kofaktoriks rohkem kui
300 ensüümis. Tagab ribosoomide ja mitokondrite tervislikkuse ja
osaleb nukleiinhapete ning valkude sünteesil. Teda vajab
rakuenergeetika, ta stabiliseerib biomembraane. Magneesiumit vajab
närvitalitlus ja lihaskoe lõõgastus, reguleerib ka südamelihase
tööd.
Kloor – Rakuvälise lokalisatsiooniga.
Ininmorganismi
keskne anioon . Tema biofunktsioonid haakuvad
naatriumi ja kaaliumi omadega. Kloori
ioonid on hädavajalikud
soolhappe sünteesiks maos.
MikrobioelemendidRaud
– rauda vajab hapnikku transportiva
hemoglobiini ja lapnikku
lihaskoes salvestava müoglobiini süntees. Esineb inimorganismis
Fe2+ ja Fe3+ vormis. Paljude raudasisaldavate
biomolekulide tegevuse alus ongi raua oa muutus. Raud on organismis
kasulik vaid seotuna! Vabanenud raud oksüdeerub prganismis koheselt
raskestilahustuvateks toksilisteks
produktideks .
Tsink –
luudes, skeletilihastes, eesnäärmes, neerudes,
maksas , hüpofüüsis.
Umbes 300 ensüümi
kofaktor , osaleb valkude ning nukleiinhapete
sünteesis. Tsingita häirub normaalne areng, kasv ja paljunemine
samuti immuunsüsteemi,
epidermise ning maitseretseptorite normaalne
areng ja insuliini toime. Soodustab B-kompleksi vitamiinide
imendumist/omastamist. Osaleb alkoholi metabolismis.
Fluor
– hammastes, luudes. Vajalik hammaste arenguks, suurendab kaltsiumi
deponeerumist hambakudedes, on oluline vereloomes. Pidurdab
suhkrute muutumist
suus orgaanilisteks hapeteks.
Räni –
kõhredes, kõõlustes, luudes, veresoonte seintes.
Vask –
Vaske vajab hemoglobiini süntees, kofaktorina aminohapete
metabolismi ja fosfolipiidide sünteesi ensüümid. Vajalik
rakuhingamises, soodustab raua omastamist.
Mangaan –
osaleb kilpnäärme hormoonide, rasvhapete, kolesterooli sünteesil.
Soodustav C-vitamiini bioaktiivsust. Vajalik
rinnapiima produtseerimiseks(inimestel), vereloome
soodustamiseks ning side- ja
luukoe moodustumiseks.
Jood – kilpnäärme hormoonide
süntees, kilpnäärme töö ja valkude süntees, millest sõltub
järglaste kasv, areng; metabolismi kiirus; termogenees; juuste,
küünte ja naha seisund.
Organismis veel leiduvaid
mikrobioelemente :
Seleen , Tina,
Koobalt , Molübdeen,
Nikkel ,
Kroom ,
Arseen,
Vanaadium , Boor
3. Aminohapped : Omadused, klassifikatsioon Aminohapped on
karbksüülhapete
derivaadid .
Inimkeha valgud ja peptiidid koosnevad
aminohapetest.
Aminohappeid kasutab inimkeha: ehitusüksustena;
ensüümide, valkude, hormoonide süntees; energiamaterjalina
süsinikskeleti lammutamisel; teiste biomolekulide sünteesil.
Aminohappeid kui
lihtbiomolekule kasutatakse inimorganismis :
* Ehitusüksustena –
valkude, ensüümide, hormoonide, jne sünteesiks
*Energeetiliste
materjalidena (metaboolse kütusena) – aminohapete süsinikskeleti
lammutamisel saab salvestada metaboolset energiat.
*Eelühenditena
– paljude signaalmolekulide (hormoonid,
neurotransmitterid ) ja
lihtbiomolekulide (süsivesikud, sukleotiidid) sünteesil.
Omadused:
Amino-
ja karboksüülrühma tõttu on aminohapped vesilahustes bipolaarse
ioonina
Sõltuvalt keskkonna pH-st käituvad aminohapped prootoni
doonoritena (lahus on nõrk -hape) või prootoni aksteptoritena
(lahus on nõrk -alus).
Füsioloogilise pH (7-7,4) juures on
aminorühm protoneeritud ja karboksüülrühm karboksülaataniooni
vormis. (need on laenguga, st molekulid on bipolaarsed
ioonid )
Isoeletriline punkt - pH väärtus, mille juures ta on
elektriliselt
neutraalne , s.t. anioonsed ja katioonsete
laengud on
võrdsed. pI juures aminohape elektriväljas ei liigu.
Aminohapped
on optiliselt aktiivsed –
polariseeritud valguse
tasapinna pööramine. Molekulid on asümmeetrilised. Omavad (k)hiraalset
tsentrit, v.a glütsiin (Gly). L- ja D-isomeersus.
Lahustuvad
polaarsetes
lahustites (vees, etanoolis), ei lahustu apolaarsetes
lahustites (
benseen jt).Kõrge
sulamistemperatuur –
kristallstruktuuri lõhkumiseks.
Klassifikatsioon:
*
Proteinogeensed aminohapped – valkude ehitusüksused
*
Aproteinogeensed aminohapped – valkudes mitteesinevad
aminohapped
Asendamatud aminohapped on aminohapped, mida
inimese organism ise kas üldse ei tooda või toodab vähesel määral,
nii et nende
omastamine toidust on möödapääsmatult
vajalik.(
Histidiin , Isoleutsiin,
leutsiin , lüsiin,
metioniin ,
fenüülalaniin, treoniin, trüptofanaan, valiin))
Asendatavaid
aminohappeid suudab organism ise toota asendamatute aminohapete ja
muude ainete baasil. (
alaniin )
Happelised –
aspartaadi ja lutamaadi R-grupid sisaldavad krboksüülrühma, mis on
füsioloogilise pH juures negatiivselt laetud.
Aluselised –
Lüsiini ja arginiini r-grupid on füsioloogilise pH juures
positiivselt laetud. Histidiini R-grupp võib sõltuvalt ümbritsevast
keskkonnast olla laenguta või positiivselt laetud – esineb tihti
ensüümide aktiivtsentris.
Aromaatsed – Fenüülalaniin ja
trüpofaan on väga aromaatsed.
Hüdroksüaminohapped – Seriini
ja treoniini hüdroksüülrühmad teevad nad palju hüdrofiilsemaks
ja reaktiivsemateks
Väävlit sisaldavad aminohapped – Tsüseiini
tioolrühm –SH on väga
reaktiivne . Esineb tihti ensüümide
aktiivtsentris
Aminohapete
amiidid –
aspargiin on aspartaadi
amiid ja glutamiin on glutamaadi amiid
4. Valgud:
üldiseloomustus, funktsioonid loomaorganismidesMille poolest
erineb valk peptiidist?
Oligopeptiid – 2-20
aminohappejääki
Polüpeptiid – 20-50 aminohappejääki
Valk
– üle 50 aminohappejäägiValkude
üldtunnusjooned on:
Nad on biomakromolekulid, mis koosnevad
ühest või mitmest polüpeptiidiahelast
Nende aminohappelise
koostise erinevus, mis tingib nende
individuaalsuse/rohkuse
Peptiidside aminohappejääkide
vahel
Mitmetasemeline
struktuurne organisatsioon Omavad
aktiivalasid ligandide sidumiseks
Kuna valgud on
geneetilise info realiseerimisvahendid, siis on nad inimeha
arvukaimad biomakromolekulid. Suur arv tuleneb antud struktuurvalkule
omasest aminohappejääkide
kindlast järjestusest ja koosseisust.
Kudede/organite valgusisaldus sõltub nende ülessannetest ja
valgusisaldus muutub organismi individuaalse arengu jooksul ja
haiguse korral. Inimkehas on valke umbes 40...46%
kuivkaalust.
Amfoteersus ( happelisus /aluselisus)- Valgud on
amforteersed polüelektrolüüdid (neil on happe ja aluse loomus).
Selle määravad aminohappejääkide ioniseeruvad
R-grupid.
Puhverdusvõime-seovad pöörduvalt H+
või OH-
Valkude adsorptsioonivõime-võivad
absirbeerida mitmesuguseid aineid ja ioone (hormoone, vitamiine,
rasvhappeid , rauda, vaske, ravimeid). See muudab need ained/ühendid
lahustuvateks või blokeerib nende
toimet/
toksilisust Makromolekulaarsus- valkudel, mille
primaarstruktuur on teada, sab Mrarvutada. Kui koostis
pole teada, määratakse Mr eksperimentaalselt(mass-spektromeetriga)
Valgud on keha
peamised ehitusmakterjalid ( lihastes valgu osakaal 80%)
Ainevahetus (vitamiinide ja teiste ainete
trantsport )
Aitavad vere pH
säilitada (puhversüsteemid)
Vere hüübimist
teostab vereplasma
valk-
fibrinogeen Trantsport
hemoglobiin , mis
varustab kogu keha
hapnikuga
Biokatalüsaatoriteks-> fermentideks(
ensüümid)
Valgustruktuurid kindlustavad kudedes erituse tekke ja
erutuse levimise
Funktsioonid:
*ensümaatiline katalüüs (CO2 hüdraatimine, RNA).
Peaaegu kõik ensüümid on valgud.
* Transport ja säilitus
funktsioon. Ainete transport biovedelie kaudu, transport läbi
biomembraanide
* Koordineeriutd liikumine (lihaste kontraktsioon,
kromosoomide liikumine mitoosis)
* strukturaalne (nahk, kondid)
*
immuunvastutus
* närviimpulsside teke ja ülesanne
(
retseptorvalgud meeleelundites, sünapsis)
* rakkude, kudede,
organismi kasv ja diferentseerumine, jagunemine
Denaturatsioon-valgu
bioaktiivuse kadumine ( temperatuur,
vibratisoon , ultraheli,
keskkonna pH, ioniseeriv kiirgus)
Renaturatsioon -fibrille rohkem,
kui vanemal loomal. Valgu bioaktiivuse taastamine
5. Valkude
primaarstruktuurPrimaarstruktuur
on
kovalentne peptiidsidemetega seotud aminohappejääkide kindel järjestus antud valgu polüpetiidahelas(-tes) . Aminohappejääke
seob polüpeptiidahelaks kovalentne peptiidside. Paljudes valkudes
esineb ka kovalentseid disulfiidsidemeid (S-S sidemeid). Need tekivad
endoplasmaatilises retiikulumis, esinedes nii
enamikes sekreteerivates ja membraansetes valkudes. Disulfiidsidemed
luuakse kahest tsüsteiinjäägist - ahelasisesed (
tugevdavad primaarstruktuuri) või ahelatevahelised (aitavad siduda
polüpeptiidahelaid).Peptiidsidemed ja disulfiidsidemed on
kovalentsed.
Valkude primaarstruktuur on molekulaaraluseks:
-
valkude
spetsiifilisusele/mitmekesisusele- kõrgemate
struktuuritasemete kujunemisele-
molekulaarhaiguste
patogeneesileSpetsiifilisus – on valkude olulisim
tunnus. See on aluseks valkude põhifunktsioonidele (katalüütiline,
regulatoorne).
Nt. Valguline antikeha seostub spetsiifiliselt
vastava antigeeniga. Valkude spetsiifilisuses eristatakse:
- Liigispetsiifilisus – liigile iseloomulike valkude spekter
- Organismispetsiifilisus – liigi erinevate organismide valkude spekter
- Koespetsiifilisus – erinevate kudede valgud erinevad immunoloogiliselt, st ühe koe valgud kutsuvad teise koesse sattumisel esile antikehade tekke
Juba ühe aminohappejäägi asendus muudab valgu
spetsiifilisust.
Primaarstruktuur on „baasinformatsiooniks“
kõrgemate struktuuritasemete kujunemiselSee tähendab, et
aminohappejääkide kindel järjestus antud valgus on
baasiks antud
valgu kõrgemate struktuuritasemete kujunemisele.
Primaarstruktuur
ja molekulaarhaigusedGeneetilise päritoluga haiguste
põhjuseks on enamasti mõne aminohappejäägi
asendumine primaarstruktuuris. Sel puhul räägitakse molekulaarhaigustest ehk
geneetilistest haigustest.
Primaarstruktuuri määramine annab
ka kliiniliselt olulist informatsiooniPrimaarstruktuuri
selgitamine annab informatsiooni valgu võimalike kõrgemate
struktuuritasemete kohta, annab informatsiooni valgu võimalike
aktiivalade ehituse kohta, võimalike molekulaarhaiguste kohta,
näitab liitvalkude puhul mittevalgulise komponendi seostumiskohti.
6.
Valgumolekulide ruumiline ehitus, kõrgemat järku struktuurid .Valgu
sekundaarstruktuur on peamiselt vesiniksidemete abil fikseeritud
ruumikujund. Vesiniksidemete
rohkus ühes valgumolekulis tagab
struktuuri atabiilsuse.
Sekundaarstruktuuri põhivormid on
alfa-heeliks ja beeta –struktuur.
Alfa-heeliks-
polüpeptiidahelaparemale pöörduv helitseerunud
konformatsioon .
Vesiniksidemete rohkus tagab heeliksi stabiilsuse.
Beeta-struktuur-
peamiselt vesiniksidemete abil kujunenud kihilis-voldiline
konformatsioon.
Tertsiaarsruktuur –
kerajas -ellipsoidne
(gloobul) või
niitjas (fibrill) kolmemõõtmeline konformatsioon.
Formeerub polüpeptiidahela spetsiifilisel väga tihedal
kokkupakkimisel.Valgumolekul püüab võtta stabiilsemat kuju.
Tertsiaarstruktuuri hoiavad põhiliselt nõrgad sidemed. Nende väga
suur arv tagab tertsiaarstruktuuri stabiilsuse.
Kvarternaarstruktuur
– vähemalt kaks tertsiaarstruktuuriga polüpeptiidahelat.
(Subühik) Subühikuid seovad nõrgad sidemed.
7.
Valgumolekulide aluselised ja happelised omadused. Valkude
isoelektriline täpp. Katioon –
positiivne laeng
Anioon – negatiivne laeng - Amino- ja karboksüülrühma tõttu on aminohapped vesilahustes bipolaarse ioonina (tsvitter-ioonina.
- Sõltuvalt keskkonna pH-st käituvad aminohapped prootoni doonoritena (lahus on nõrk hape) või prootoni aksteptoritena (lahus on nõrk alus).
Füsioloogilise
pH (7-7,4) juures on aminorühm protoneeritud ja karboksüülrühm
karboksülaataniooni vormis. (need on laenguga, st molekulid on
bipolaarsed ioonid)
Lahused on nõrgad puhvrid. Happelises
keskkonnas on aminohapped katioonid, aluselises keskkonnas on
anioonid .
Isoelektriline täpp pI on keskkonna pH, mille korral on
valgu molekuli
summaarne laeng null. Selle juures valgud sadenevad
kõige kergemini, valgulahus on kõige ebastabiilsem. Aminohapped
elektriväljas ei liigu, on elektriliselt neutraalne, anioonsed ja
katioonsed laengud on võrdsed.
pH pH > pI – negatiivselt laetud
pI –
isoelektriline täpp
pH –
vesinikeksponent pK –
dissotsiatsioonikonstant
8. Valkude
füüsikalis-keemilised omadusedLaeng:
COOˉ NH3+ (see ei anna laengut!)
C
terminaalne ots N terminaalne
ots
Laeng on põhjustatud koostises olevate AH radikaalide
laengutest. Laeng kindlustab vaba liikumise elektriväljas ja tagab
stabiliseeruva H2O molekulkihi ümber valgu, seega ka valkude
lahustuvuse . Kui summaarne laeng on 0, sadeneb kergesti välja.
Lahustuvus veres ja veresoola lahustes. See on määratud
valkude AH koostise eripäraga: mida rohkem hüdrofoobseid AH-jääke,
seda väiksem lahustuvus. On määratud pH-st, ioontugevusest ja
tº-st. (Pm.. kolloid-
osmootne – enamik valke hüdrofiilsed ja
vesilahustuvad)
Madal difusioonikiirus. See on
seletatav nende suurte mõõtmetega.
Amfoteersed ühendid:
aminorühmad annavad aluselisi ja COOH-rühm happelisi omadusi.
Puhverdusvõime teatud piirides. See realiseerub peamiselt
H
sidumise või loovutamise tasandil.
Võime denatureeruda.
See on valgu bioloogilise aktiivsuse kadumine seoses kõrgemat järku
struktuuride lagunemisega (st alles jääb primaarstruktuur).
Denaturatsioonifaktoriteks on: tº, kiirgused,
happed , alused.
Denaturatsiooni bioloogiline tähtsus:
- Teatud määral kaitse võõrvalkude eest (nt palavik ).
- Denatureeruvad valgud muutuvad hõlpsamini hüdrolüüsivateks (nt maos HCl-line denaturatsioon).
Renaturatsioon
e denaturatsiooni pöördprotsess. Avaldub suhteliselt pehme
denaturatsiooni korral ja denaturatsioonifaktorid peavad olema
kõrvaldatud. Selle tulemusena
taastub primaarstruktuurist kõrgemat
järku struktuur ja taas ilmneb valgu bioloogiline aktiivsus.
Renaturatsiooni bioloogiline tähtsus:
- Väga lihtsate mõjutuste korral (nt pH muutused) ei toimu valkude pöördumatut inaktivatsiooni.
- Hüdrolüüs – peptiidsideme lagunemine ja vabade AH teke. See toimub kas tugevalt keemiliste mõjutustega või ensümaatiliselt (spetsiaalsete proteolüütiliste ensüümide toimel).
Püsivus –
valgulahus ei koaguleeru seismisel (ei sadene täielikult), kuna
kolloidolekut stabiliseerivad valguosakese laengud ja
hüdraatkiht.
Optiline aktiivsus ja adsorptsioonivõime –
valkude lahuste optiline aktiivsus tuleneb neis olevate AH-jääkide
optilisest aktiivsusest ja valgu konformatsioonist. Valgud võivad
adsorbeerida mitmesuguseid aineid ja ioone. See muudab need ained
lahustuvateks või blokeerib nende
toimet/toksilisust.
Makromolekulaarsus9. Valkude
klassifikatsiooni printsiibid , tähtsamad esindajadPäritolu
järgi: loomsed , taimsed,
bakteriaalsed , viiruste
valgud
Paiknemise järgi: membraan -, tsütoplasma-,
mitokondri-, ribosoomi-, lüsosoomi jne valkudest.
Füsiko-keemiline
klassifikatsioon: Polaarsed, Apolaarsed,
Amfifiilised
Struktuurne klassifikatsioon: 1. Lihtvalgud – a)fibrillaarsed –
kollageenid ,
elastiinid ,
keratiinid, fibroiinid(fibrinoogeen), müosiinid
b)globulaarsed
– albumiinid, globuliinid,
histoonid , protamiinid, prolamiinid,
gluteliinid
2. Liitvalgud – kromoproteiinid,
fosfoproteiinid, glükoproteiinid, proteolipiidid,
lipoproteiinid ,
nukleoproteiinid, metalloproteiinid ja liitensüümid
Lihtvalgud
koosnevad ainult aminohapete jääkidest
Liitvalgud sisaldavad
peale aminohapete jääkide orgaaniliste ja anorgaaniliste ainete
molekule/molekulide osi.
Funktsionaalne
klassifikatsioonid:
Ensüümid (
pepsiin , trüpsiin,
amülaas)
Transportvalgud (hemoglobiin, vereseerumi
albumiin ,
ioonpumbad)
Struktuurivalgud (kollageenid, elastiinid,
histoonid)
Regulatoorvalgud (
insuliin , histoonid)
Aktiivkaitse
valgud (immuunoglobuliinid, fibrinogeen, trombiin)
Toite- ja
varuvalgud (piima kaseiin, muna ovoalbumiin)
10. Kromatograafia . Elektroforees Kromatograafia
- üldmõiste mitmesuguste laboratoorsete füüsikalis-keemiliste
meetodite kohta, mida kasutatakse uuritavate ainete segu
komponentidel lahutamiseks paljukordse sorptsiooni ja
desorptsioon tingimustes. Mõiste kromatograafia hõlmab
vastavaid
meetodeid , protsesse ja
teadusharu .
Lihtsustatult:
praktikas kantakse ainete segu läbi sorbendi (liikumatu
faas) sobiva vedeliku või gaasi vooluga (liikuv
faas). Segu komponentide spetsiifilise sorptsiooni ja desorptsiooni
tulemusena toimub nende jaotumine liikumatu ja liikuva faasi vahel
vastavalt jaotuskoefitsientidele ning nende aktide paljude korduste
tagajärjel komponendid liiguvad edasi erinevate kiirustega. See viib
ainete lahutumisele ning moodustuvad kiiremini ja aeglasemalt
liikunud komponentide tsoonid. Protsess teostatakse kas
kolonnis ,
kapillaaris,
paberil või plaadil. Lahutunud komponendid
detakteeritakse füüsikaliste või keemiliste
meetoditega.
Eesmärgi põhjal jagunevad kromatograafilised
meetodid preparatiivseteks ja analüütilisteks. Preparatiivse
kromatograafia korral on eesmärk saada teatud hulk lahutatud
komponenti(te) edasiseks kasutamiseks. Seega on tegu
kromatograafilise puhastamisega. Analüütilise kromatograafia
korral kasutatakse oluliselt väiksemaid ainete koguseid (enamasti
mikrogrammides) ja eesmärk on määrata komponentide suhteline
sisaldus segus.
Elektroforees (
elektro +
kr
phoros kandev ) on elektriliselt laetud osakeste liikumine vedelikus elektrivälja mõjul:
positiivsed osakesed katoodile ja negatiivsed osakesed
anoodile.
Seda omadust, et erisuguse suuruse ja laenguga osakesed
liiguvad elektroodide vahel erineva kiiruse ja suunaga, rakendatakse
aineosakeste üksteisest eraldamiseks vastavalt nende laengule või
suurusele, ka sõltuvalt molekulide pikkusest.
Elektroforeesi kasutatakse nt valkude ja DNA
-analüüsis, elektroonilise paberi tehnikas, samuti
metalli pinnale kolloidosakestest kattekihi tekitamiseks
11. Nukleotiidid Nukleotiidid on
nukleiinhappe
monomeerid . Nad on nukleosiidide
mono -, di- või
trifosfaatestrid. Riboosi/desoksüriboosi esterifitseerumine
fosforhappejäägiga annab ribonukleotiidi/desoksüribonukleotiidi.
Mononukleotiidid võivad olla mono-, di- või trifosforüülitud. DNA-s võivad esineda nukleotiidid: adenosiinfosfaat A,
guanosiinfosfaat G, tsütidiinfosfaat C ja tümidiinfosfaat T. RNA-s
esineb tümidiinfosfaadi asemel uridiinfosfaat U. Nukleotiidid on
komplementaarsed (A-T/U ja C-G). Rakus esinevad nukleotiidid
anioonidena, st nad on happed, seda märgib happeanioonile
viitav nimetus (adenülaat ehk AMP, guanülaat ehk GMP jne.) Nukleotiidi
kolmetäheline sümbol (ATP, GDP jne)
viitab N-alusele ja
fosfaadijäägi arvule. Desoksüribonukleotiidid sümbol on vastavalt
dATP, gGDP jne. Nukleotiidide summaarhulk rakus jääb alati üsna
konstantseks ning nad omavad neeldumismaksimumi vahemikus
259...271nm, mille alusel saab nende hulka ahuses määrata.
12. RNA:
Ehitus, funktsioonRNA ehitus:
- Üheahelaline
- Koosneb ribonukleotiidijääkidest
- N- alusteks on A, G, C, U
RNA
primaarstrukruur ... on 3’,5’- fosfodiestersidemega seotud
ribonukleotiidjääkide lineaarahel.
DNA –A-G-C-T-A-G-
RNA
–U-C-G-A-U-C-
Eukrüootses rakus sünteesitakse RNA eritüüpide
molekulid DNAühel
ahelal komplmentaarsuse printsiibi alusel,
Seetõttu peegeldab RNA nukleotiidne järjestus komplementaarsusest
tulenevalt DNA teatud lõigu nukleotiidjärjestust.
RNA
sekundaarstruktuur - Üheahelalise RNA aluste komplementaarne
paardumine vesiniksidemete abil tekitab sekundaarstruktuuri ehk
kujuneb kaksikhelikaalsed alad. Tasapinnalisel esitamisel meenutab
tRNA sekundaarstruktuur ristikheinalehte. Teiste RNA-de
sekundaarstruktuur on sarnane tRNA järgmistes
aspektides :
- Omavad aluste paardumise tõttu kaksikhelikaalseid regioone
- Esineb vesiniksidemetevabu linge
- Nende molekulis esineb konservatiivseid nukleotiidijääkide järjestusi.
RNA
tersiaarstuktuur - Tänu vesiniksidemetele ahela erinevate osade
koplementaarsete N-aluste vahel ja hüdrofoobsetele vastastoimetele
kujuneb RNA
tertsiaarstruktuur . Ehituselt kompaktne, tihedamate ja
vähemtihedate regioonidega. rRNA tertsiaarstukruur on kerajas.tRNA
tertsiaarstuktuur on spetsiifiline ruumikujund, mis moodustub eeskätt
molekulisiseste
arvukate vesiniksidemete tõttu. mRNA helitseerunud
ja leitseerumata lõikudest kooneva ahela tertsiaarstuktuur meenutab
lihtsustatult võttes valgulisele niidirullile keritud niitjat
ruumikujundit.
Funktsioonide alusel eristatakse 3 RNA
põhitüüpi: - mRNA- asub tsütoplasmas, valgusünteesil kannab üle vajaliku geneetilise ingo DNA-lt ribosoomidele.
- rRNA- asub ribosoomides, ribosoomide nukleiinhappeline koostisosa , mis osaleb aminohapete lülitumises sünteesitavasse polüpeptiidahelasse.
- tRNA – asub tsütoplasmas, transpordib aminohappeid ribosoomidele ja võtab osa nende lülitumisest sünteesivatesse polüpeptiidahelasse.
13. DNA:
Ehitus, funktsioonDNA – e.
desoksüribonukleiinhape. On enamikus elusorganismides pärilikku
informatsiooni säilitav aine. Puhas DNA on happeline,
toatemperatuuril tahke, suhteliselt pehme ja vees hästi lahustuv
aine.
DNA on polümeer, mis koosneb
desoksüribonukleiinhappejääkidest. N-alusteks on adeniin(A),
guaniin (G), tsütosiin(C) ja tümiin(T).
Primaarstruktuur
: koosneb pentoos-fosfaatsest tüvest, millest ulatuvad välja
N-alused.
Sekundaarstruktuur : kaksikspiraalne, tema
molekul koosneb kahest lineaarsest komplementaarsest antiparalleelsest
keerdunud ahelast . Ahelaid hoiavad koos N-aluste vahelised
vesiniksidemed.
DNA asub:
1.
Tuumas – põhiülesandeks on geneetilise info säilitamine
ja edasiandmine „tütar DNA-le“ (rakujagunemisel) ja mRNA-le
(valgusünteesil).
2.
Mitokondrites - geneetiline info
mitokondrite osade valkude sünteesiks. Geneetiline informatsioon
kandub põlvkonnast põlvkonda salvestuna DNA nukleotiidses
järjestuses. Geenide avaldumine realiseerub informatsiooni
edastamise teel DNA nukleotiidselt järjestuselt valkude
aminohappelisse järjestusse. DNA asub inimorganismis
rakutuumas .
DNA funktsioon :
geneetilise informatsiooni kandja. Geneetiline informatsioon kandub
põlvkonnast põlvkonda salvestatuna DNA nukleotiidses järjestuses.
Geenide avaldumine realiseerub informatsiooni edastamise teel DNA
nukleoiidselt järjestuselt valkude aminohappelisse järjestusse.
14. Ensüümid:
üldiseloomustus, funktsioonid, klassifikatsioon.Ensüümid on
biokatalüsaatorid, kõrgelt spetsiifilise toimega liht- või
liitvalgud, endogeensed biokaktiivsed ühendid.
Funktsioon:Ensüümid on
erilise funktsiooniga valgud, sest seondavad ja muudavad keemiliselt
teisi molekule – katalüüsivad reaktsioone. Toimemehhanism seisneb
reaktsiooni kiirust limiteeriva energeetilise barjääri alandamises,
st.
reaktsioon , mis ilma ensüümita oleks toimunud aeglaselt, toimud
ensüümi osavõtul väga kiiresti. Ensüümid ei muuda reaktsiooni
suunda, ei muuda tasakaaluseisundit vaid kiirendavad selle saabumist,
ei lõhustu reaktsiooni käigus.
Klassifiatsioon:Oksüreduktaasid katalüüsivad redoksreaktsioone
Transferaasid Funktsionaalsete rühmade ülekanne
Hüdrolaasid katalüüsivad hüdrolüüsi
Lüaasid Kaksiksidemete (nt. C-C, C-O,C-N, C-S) lõhustamine
Isomeraasid Isomerisatsioonireaktsioonid
Ligaasid sünteesireaktsioonid
15.Ensüümide
toimemehhanism, substraat , ensüümiaktiivsus, aktiivtsenter,
koensüümid
Toimemehhanism –
ensüümide poolt katalüüsitud reaktsioonide aktivatsiooni alandamine saadakse ensüümi ja substraadi (ES) kompleksi
moodustamise abil:
E + S ES -> E
+P ( produkt )
ES kompleksi tekkes osalevad vesiniksidemed, hüdrofoobsed ja elektrostaatilised
vastaktoimed. Reeglina pöörduv.
Substraat – reageerivad ühendid ensüümkatalüüsis, millega ensüüm seob ja
mida muundatakse.
Aktiivtsenter on
ensüümi pinnaala, millega seostub substraat. Aktiivtsentris
paiknevad aminohappejääkide katalüütilised rühmad, mis seovad
endaga substraadi. Aktiivtsentril on kaks põhilist rolli:
o Siduv roll - seob
endaga substraadi
o Katalüütiline
roll - muudab substraadi produktiks, tänu millele toimub aktivatsioonienergia alandamine
Akriivtsenter ja
substraat peavad olema komplementaarsed, et saaks tekkida ES kompleks . Ensüüm võib muuta ka oma konformatsiooni, et siduda
substraati efektiivsemalt. Kui substraat on muundatud produkitks,
eemaldub see ensüümi aktiivtsentrist, sest kaob ensüümi ja
tekkinud produkti komplementaarsus .
Koensüümid –
madalmolekulaarsed orgaanilised ühendid, mis on tavaliselt
liitensüümis valkosaga mittekovalentselt seotud. Paljud koensüümid
on vitamiinid .
16.
Ensüümreaktsiooni kiiruse sõltuvus substraadi kontsentratsioonist,
keskkonna pH-st ja temperatuurist.
Konstantse ensüümi
kontsentratsiooni juures sõltub ensüümreaktsiooni kiirus
substraadi kontsentratsioonist hüperboolselt: Madala substraadi
kontsentratsiooni juures suureneb kiirus lineaarselt, korgemate
substraadi kontsentratsioonide juures muutub kiirus jarjest
vaiksemaks
V0 = Vmax [S] / Km +
[S]
Temperatuur, mille
juures ensüümreaktsiooni kiirus on maksimaalne, on reaktsiooni
temperatuurioptimum(imetajate ensüümidel vahemikus 37..43C)
• Optimumist
kõrgemad temperatuuris denatureerivad ensüümvalgu, madalamatel aga
langeb reaktsioonikiirus.
• Temperatuuri
toimet ensüümreaktsioonide kiirusele tuleb arvestada
- Palavik tõstab
ensüümreaktsiooni kiirust 20..30%,st suureneb biomolekulide ja
energia kulu organismis.
- Hüpotermiat. Jahutamine alandab ensüümreaktsioonide kiirust. Vähendab aine- ja energiakulu ning pikeneb rakkude eluvõime ekstreemtingimustes.
Ühekordne külmumine ja aeglane ülessulamine ei kahjusta ensüüme.’
Inimkeha ensüümide
pH optimum on vahemikus 6..8 – ensüümide kiirus maksimaalne.
Konkreetse ensüümi pH optimum sõltub ensüümi töökeskkonnast
organismis.
- ensüümi
aktiivtsentris domineerivad aminohappejäägid, mis omavad
töökeskkonna pH vahemikus ioniseeruvat funksionaalset rühma.
- Ekstreemsetel pH
väärtustel ensüümvalk denatureerub.
17. I järku
ensümaatilise reaktsiooni kineetika , Michaelise konstant.
Reaktsioonikiiruse
sõltuvust ensüümi & substraadi konsentratsioonist, keskkonna
temperatuurist ja pHst uurib ensüümreaktsioonide kineetika. Ensüümi
toimemehhanismi kindlaks tegemine ja organismis toimuva
ensüümreaktsiooni kiiruse regulatsiooni säilitamine.
Esimest järku
reaktsiooni kiirus sõltub aine konstentratsioonist, pH-st ja
katalüsaatorist
Michaelise constant kirjeldab ensüümreaktsiooni kiiruse sõltuvust substraadi
kontsentratsioonist.
Michaelise konstant
on arvuliselt võrdne substraadi kontsentratsiooniga , mille juures
kiirus on pool maksimaalsest. Mida väiksem on Km (Michaelise
konstant) , seda madalama kontsentratsiooni juures ensüüm
efektiivselt töötab.
18.Ensüümide inhibiitorid ja aktivaatorid . Ensüümide lokalisatsioon rakus.
Ensüümid on
reguleeritava aktiivsusega katalüsaatorid:
- Aktivaatorid tõstavad ensüümi reaktsiooni kiirust
- Inhibiitorid pidurdavad osaliselt või täielikult
Ensüümide inhibitsioon bioloogiliste süsteemide metabolismi oluline
regulatsiooni mehhanism . Paljude ravimite toime seisneb vastava
ensüümi inhibeerimiseks.
Inhibitsioon –
pöördumatu või pöörduv.
Pöördumatu puhul
seostub inhibiitor ensüümi aktiivtsentrisse või mujale väga
tugevalt (kovalentselt või mittekovalentselt) ning inhibiitori
dissassotsiatsioon ensüümilt on väga aeglane. Side moodustub
aminohappega, mis osaleb normaalselt ensümaatilises reaktsioonides.
Pöörduvad
inhibiitorid on nõrga mittekovalentse sidemega ja on võimelised
kompleksist dissotseeruma. Ensüüm on inaktiivne ainult siis, kui me
lahusest inhibiitorit ei eemalda.
Pöörduva
inhibitsiooni variandid:
- Ensüüm-substraadi kompleks
- Konkurentne inhibitsioon
- Inhibiitor konkureerib substraadiga ensüümi aktiivtsentrisse seostumisel
- Substraadi kontsentratsiooni tõus tõrjub aktiivtsentris seostunud inhibiitori välja
- Mittekonkurentne inhibitsioon
- Inhibiitor seostub ensüümmolekuliga väljaspool aktiivtsentrit
- Substraadi kontsentratsiooni tõus ei mõjuta
Konkurentne
inhibiitor ei muuda reaktsiooni maksimaalset kiirust, mõjutab Km
väärtust – poole maksimaalse kiiruseks on vaja tõsta substraadi
kontsentratsioon.
Mittekonkurentne
inhibiitor vähendab maksimaalset kiirust, kuid ei muuda Km väärtust
– toimib aktiivtsentri väliselt.
Allosteeriline –
regulatoorne tsenter (peale aktiivtsentrit)
- Ensüümi pinnaosa, millega seostub regulaator (ioonid ja madalmolekulaarsed ühendid)
- Paljud ravimid – allosteerilised efektorid
Allosteeriline
inhibitsioon on pöörduv.
Allosteerilised
efektorid:
- Muudab ensüümi konformatsiooni
- Muutub aktiivtsentri ruumiline ehitus
- Aktivatsiooni puhul – muutuvad substraadi sidumine ja katalüüs efektiivsemaks
- Inhibitsiooni puhul – väheneb aktiivtsentri affinus substraadile.
Ei allu
Michaelis- Menteni kineetikale. Reatsiooni kiiruse ja substraadi
kontsentratsiooni sõltuvus on S-kujuline.
19. Valkude
klassifikatsiooni printsiibid, tähtsamad esindajad.
Lihtvalgud –
koosnevad ainult aminohapete jääkidest. Jagunevad veel eraldi
fibrillaarseteks (kollageenid, elastiinid, keratiinid, fibroiinid,
müosiinid) ja globulaarseteks (albumiinid, globuliinid, histoonid,
protamiinid, prolamiinid, gluteiinid) valkudeks.
Liitvalgud - peale
valgulise osa sisaldavad veel mittevalgulist osa ehk prosteetilist
rühma. (kromoproteiiid, fosfoproteiinid, glükoproteiinid,
proteolipiidid, lipoproteiinid, nukleoproteiinid, metalloproteiinid
ja liitensüümid)
20. Sahhariidid :
üldiseloomustus, loomorganismi mono- ja disahhariidid .
Sahhariidid ehk
glütsiidid (traditsioonilise, ent ebatäpse nimega süsivesikud ehk
karbohüdraadid) on keemilised ained, mille molekulid on
biomolekulid, mis koosnevad süsiniku, vesiniku ja hapniku
aatomitest.
Süsivesikud
jagunevad kolme põhirühma:
• Monosahhariidid e monoosid – kahte või enamat hüdroksüülrühma sisaldavad
aldehüüdid või ketoonid :glükoos ( viinamarjasuhkur ); fruktoos
(puuviljasuhkur), mida leidub ohtralt mees, puuviljades ja
mahlades.
Funktsioonid inimorganismis :
*glükoosi metabolismi
vaheühend
*nukleotiidide ehitusüksused
*süsivesikute
metabolismi vaheühend
• Oligosahhariidid – liitsüsivesikud. Sisaldavad 2-10 monoosijääki, mis on seotud
omavahel glükosiidsidemetega: tuntumad esindajad on disahhariidid: sahharoos (tavaline lauasuhkur , koosneb glükoosi- ja
fruktoosijääkidest), mida on rohkelt suhkruroos ja suhkrupeedis; laktoos ( piimasuhkur , koosneb galaktoosi ja glükoosijääkidest),
mis moodustub peamiselt piimanäärmetes (lehmapiimas on ligikaudu
5%); maltoos (linnasesuhkur, koosneb kahest glükoosijäägist), mis
moodustub seemnete idanemisprotsessis.
• Polüsahhariidid
e polüoosid on liitsüsivesikud, millel on varuaine ja ehituslikud funktsioonid – monoosijäägid on seostunud, polüooside molekulid
võivad olla lineaarsed , spiraalsed või sfäärilised. Taimedes leiduv tärklis , mis koosneb glükoosi jääkidest ja laguneb
inimese seedekulglas ensüümide toimel glükoosiks,loomades ja seentes olev glükogeen. Ülekaalukalt on meie toidu peamine
süsivesik tärklis, mida me saame kartulit ja teraviljade teriseid
süües. Juhul kui me sööme maksa, liha ja seeni satub meie seedetrakti teatud kogus glükogeeni.
Monosahhariidid
loomaorganismis:
Absoluutne enamus
loomakeha moonosidest on D- isomeerid ja vaid üksikud esindajad on L
– isomeerid.
Monoosid organismis eristatakse:
Glükoos – on
loomakeha on keskne süsivesik.Veresuhkur ongi glükoos. Veri kannab
glükoosi kudedesse.kus ta kasutub peamiselt energia tootmiseks.
Glükoosi ilmnemine uriinis suhkurvõte puhul on tingitud veresuhkru
kõrgest tasemest.
Fruktoos –
(puuviljasuhkur,levuloos) on sahharoosi komponent . Pärilik fruktoosi
intolerantsus resulteerub Fru kuhjumisena maksas(tõsine
hüpoglükeemia,oksendamine,kollatõbi,hemorraagia)
Disahhariidid :
Laktoos. Laktoos
koosneb galaktoosi ja glükoosijäägist.Piimasuhkruna on ta piima
põhisüsivesinik(lehma piimas 3,8-5%, rinnapiimas 6-8%), mida
sünteesitakse piimanäärmetes laktatsiooniperioodil.inemesele on ta
oluline GaL allikas.Mõned inimesed on laktoosili intolerantsed
(seedensüüm laktaasi aktiivsus on väga madal või puudub). Laktoos
ei hüdrolüüsu, kuhjub seedekulgas ja lõhustub seedekulga
mikroobide toimel.
Sahharoos. Sahharoos
kossneb glükoosi- ja fruktoosijäägist. Ta on
toiduaine(toidusuhkur) ja magustaja. Teda lõhustab seedekulga
sahharaas ehk invertaas ja vabained Glc ning Fru imenduvad.
Maltoos. Maltoos
sisaldab kahte glükoosijääki. Ta on tärklise ja glükogeeni
struktuuriühik. Maltoos(linnasesuhkur) on tärklise
hüdrolüüsiprodukt taimedes. Sülje amülaas suudab toidu törklist
osaliselt lõhustada tootes maltoosi jt produkte. Pankrease amülaas
hüdrolüüsib toidutärklise maltoosiks. Seedeensüüm maltaas lõhustab maltoosi glükoosiks.
Biofunktsioonid:
*energeetiline funktsioon – glükoosi täielik oksüdatsioon
süsihappegaasiks ja veeks katab 55-60% inimorganismi
energiavajadustest, energeetiliseks varuaineks on maksa ja lihaste
glükogeen.
*varuaine funktsioon – glükogeen
*Biosünteetiline
funktsioon – kasutatakse rasvhapete, aminohapete jt ühendite
süsinikskeleti loomiste aluseks
*struktuurne funktsioon –
monoosid ja nende derivaadid on polüooside
monomeerid
*bioregulatoorne funktsioon – kasutatakse mõningate
hormoonide ja koensüümide komponentidena.
21.Homo- ja
heteropolüsahhariidid.
Homopolüsahhariidid
ehk homopolüoosid
Homopolüooside
ehitusüksusteks (monomeerideks) on üht-tüüpi monoosijäägid
(enamasti glükoos).
Kesksed homopolüoosid on:
- Varupolüoosid (glükoosi varud) nagu glükogeen loomsetes rakkudes (ka seentes) või tärklis taime rakkudes.
- Struktuursed polüoosid nagu tselluloos (taimede rakukestad ) või kitiin (putukate, karpide, homaaride eksoskelett).
Heteropolüsahhariidid
ehk heteropolüoosid
Heteropolüoosid
(heteroglükaanid) koosnevad reeglina korduvatest disahhariidsetest
plokkidest, need plokid koosnevad omakorda erinevate monooside
derivaatidest . Nüüdisajal nimetatakse heteropolüoose
proteoglükaanideks. Kesksed esindajad on kondroitiinsulfaadid,
dermataansulfaadid, heparaansulfaat, kerataansulfaat, hüaluroonhape.
Need biomolekulid funktsioneerivad inimkehas vaid komplekseerunult
teiste biomolekulidega.
22. Lipiidid :
omadused, klassifikatsioon.
Lipiidid – vees mittelahustuvad või raskesti lahustuvad orgaanilistes lahustes
(kloroform, eeter , kuum alkohol ) lahustuvad biomolekulid. Ei ole
polümeersed, ent moodustavad agregaate. On varieeruva struktuuriga
mittehomogeenne klass molekule. Reeglina alkoholi ja rasvhapete estrid . Koosnevad akoholist ja rasvhappest. Süsiniku ahelas on 4-36
süsinikku, lipiidide ehituskomponent. Süsiniku ahelas on paarisarv
süsiniku aatomeid. Liipiidide rasvhapped on lineaarse või hargneva
ahelaga ning küllastunud või küllastumata. Mida rohkem on lipiidis küllastumata rasvhappeid, seda madalamal temperatuuril see sulab.
Inimorganismis on lipiidide alkoholideks: glütserool (kolmevalentne alkohol , baasalkoholiks triatsüülglütseriidides ja
glütserofosfolipiidides); sfingosiin (aminoalkohol, liitlipiidide
baasalkoholiks); kolestrool (küllastumata tsükiline alkohol).
Klassifikatsioon:
• Lihtlipiidid -lihtlipiidid
on neutraalrasvad ja vahad
• Liitlipiidid -liitlipiidide
rühma kuuluvad fosfo - ja glükolipiidid
• Tsükilised
lipiidid-tsükliliste lipiidide hulka kuuluvad tsükliliste
alkoholide baasil moodustuvad lipiidid, näiteks kolesteriidid
Lipiidide
biofunktsioonid:
*energeetiline funktsioon – metaboolse energia
varu
*termoregulatoorne funktsioon – rasvkude tagab
ermoisolatsiooni
*mehhaaniline funktsioon – rasvkude kaitseb
siseorganeid
*struktuurne funktsioon – osalemine
biomembraanides
*transpordi funktsioon – rasvlahustuvate
vitamiinide ja kolesterooli transport
*glükoproteiinide ankurdamine plasmamembraani
23.
Lihtlipiidid
Need lipiidid
(triglütseriidid, vahad) koosnevad baasalkoholist ja rasvhapete
jääkidest.
Triglütseriidid - Nad on glütserooli ja
rasvhapete estrid. Rasvhapped esinevad lipiidides atsüüljääkidena,
millele viitab üldnimetuse eesliide triatsüül-.
Triatsüülglütseriidide sünonüümid on triglütseriidid ja rasvad . ( Rasv =triglütseriid ehk „rasv“ hõlmab ainult seda osa lipiididest , mida nim triglütseriidideks (TG). Looduslikud TG-d on
peamiselt segatriglütseriidid, sisaldades kahte või kolme erinevat
rasvhappejääki. Loomsed ja taimsed rasvad on segatriglütseriidide segud . Mida rohkem on lipiidis küllastamata rasvhappeid, seda
madalamal temperatuuril ta sulab. Kaksiksideme cis-kofinguratsiooni
tõttu käändub küllastumata rasvhappe süsinikahel.Ta on ka jäigem
kui küllastatud rasvhappe süsinikahel. Seetõttu on küllastamata
rasvhappeid sisaldavate TG-de kokkupakkimine raskem, st sellise rasva
sulamispunkt on madalam.(Taimsed õlid küllastamata on toatemp.
vedelevad, loomsetes lipiidides domineerivad küllastatud on
tahkemad). Rasvad on kehas tsütoplasmaatilise rasvana ja
varurasvana. (Diglütseriidid töötavad signaaliülekande
vahendajana).
Vahad -Baasalkoholiks on pikaahelaga
(16...34C) ühealuseline alkohol. Loodusvahad on estrite segud(üks ester domineerib ). Vahad on hüdrofoobsed ja vesilahustumatud. Neid
lahustavad alkohol, atsetoon või eeter. Sõltuvalt temperatuurist ja
keemilisest koostisest on vahad enamasti tahked , tunduvalt harvemini
vedelad.
Loomsed vahad:
- spermatseet(vaalavõidis).
Evolutsiooniliselt/biokeemiliselt on spermatseedi lipiidikoostis
kujunenud selliseks , et tema sulamine /tahkestumine toimub just
mereveekihtide temperatuuride vahemikus.
- lanoliin ehk villavaha
on villas kuni 10 %. Ta laguneb hästi orgaanilistes lahustites, seob
ta vett, kuid vees ei lahustu. Esineb ka linnusulgedes.
Taimsed
vahad:
- karnaubavaha on segu tserot-, montaan- ja
karnaubahapetest ning nende seostunud 26...34 C-aatomilistest
alkoholidest.
Vahasid kasutatakse: kreemides, salvides, värvide
ja lakkide koostises, korrosioonivastastes segudes, paberi
immutamiseks, poleerimiseks jne.
24.
Liitlipiidid
Lihtlipiidide
ühinemisel teiste keemiliste ühenditega moodustuvadliitlipiidid,
mis omakorda jaotuvad – fosfo- ja glükolipiidid.
Liitlipiidid
- üks rasvhappejääk on asendunud fosfaatrühmaga. Näiteks - fosfolipiid - letsitiin või biomembraanide koostelipiidid.
Fosfolipiididel on kahelaadsed omadused:
1) hüdrofoobsed –
sabad (2 RH jääki)
2) hüdrofiilsed – pea (PH jääk, madalmolekulaarne ühend, glütserooli molekul käänukoht –
küllastumatus e kaksikside
Biofunktsioonid on erinevat tüüpi
lipiididel samad:
- ehituslik, membraanides
- bioregulatoorne, fosfolipiidide olek (vedelam või tahkem) määrab ära biomembraanide omadused.
- kaitse veekaotuse eest, peegeldab kiirgust, kaitse mikroobide eest.
- energeetiline
- varuaineline, taimedel on õlid koondunud seemnetesse ( raps , kanep ) või viljadesse( oliivid , pähklid)
- lahusti, rasvlahustuvate vitamiinide jaoks (A, D, E, F + asendamatud rasvhapped K ja Q).
25. Rasvas
lahustuvad vitamiinid
• A – retinoidid – nägemisprotsess, somaatiline funktsioon (kasv ja
diferentseerumine – limaskestade epiteeli areng, naharakkude areng,
kõhre ja luukoe areng), reproduktsioon (spermatogenees, ovogenees ,
platsenta areng, embrüonaalne kasv ja areng), antioksüdantne
regulatsioon
• D – kaltsiferoolid – luukudede areng koostöös
kaltsiumi ja fosforiga.
• E – tokoferoolid – antioksüdant,
soodustab seleeni püsimist organismis, stimuleerib immuunvastust,
vähendab premenstruaalsündroomi, vereringe eba regulaarsust.
•
K – naftokinoonid – labiilsed UV-kiirguses. Vere hüübimine.
•
Q – ubikinoonid – antioksüdantide kaitse, hingamisahela keskne
komponent.
Vitameer –ühendite grupi üksikesindaja, kus üks ja
seesama täht on väga sarnase ehituse ja sama toimega. Nt A1 ja A2.
26. Vees
lahustuvad vitamiinid
Tiamiin -Taimsetest
ja loomsetest produktidest imendub ta peensoolest kandja—vahendatud
aktiivportsessina. Inimekehas 30...35mg.
Riboflaviin-Termostabiilne
(ei lammuta piima pastöriseerimise ega toidu töötlus
mikrolainetega) vesilahustav ühend. Valguse toimeb annab ta vabu radikaale. Tagavarad (maks, neer ) on minimaalsed, mistõttu teda peab
toiduga pidevalt saama.
Niatsiin -Niatsiin on üldnimetus
vesilahustuvate vitamiinsete nikotiinhappe ja nikotiinamiidi kohta.
Suhteliselt püsiv kuumutamise ,valguse,õhu, hapete ja aluste
suhtes.Niatsiiin on vaja närvikoe ja naha normaalseks talitluseks .
Pantoteenhape-Termostabiilne üõhend. Annab naatriumsooli
ja kaltsiumsooli (Ca-pantotenaat).Vajalik süsivesikute,
aminohapete,lipiidide, hukleiinhapete metabolismi esüümide
tööks.
Püridoksiin-laguneb valguse käes.
Kobalamiinid-Vähesel määral sinteesib vitamiin B12
jämesoole mikrofloora, kuid selle imendumine on
tühine.
Foolhape -Laguneb valguse toimel ja kuumutamisel
ning on vesilahustuv
Kobalamiinid-sünteesib vitamiin
jämesoole mokrofloora,kui selle imendumine on tühine. Sisaldavad
vaid loomsed produktid .
C(askorbiinhape)-on antiskorbuutne
vesilahusuv ühend. Redutseerivate omadustega üsna tugev hape, mille
bioaktiivsus kaob kuumutamisel, hapniku ja valguse toimel.Inimkehas
on 2...5g vitamiin C.Rohkesti on neerupealistes,maksas, kopsudes, aga
ka rakkudevahelises vedelikus. On vaja
naha,igemete,kapillaaride,hammaste,sidemete,luude normaalseks
funktsioneerimiseks.
27.
Vitamiinide seos ensüümidega
Vitamiinid on
paljud koensüümid – nende toime realiseerub enamasti ja olulisel
määral koensüümide kaudu. Vitamiinid on liitensüümide
ehituslik-funktsionaalsete koostisosadena (koensüümidena)
hädavajalikud ensüümkatalüüsis. Seepärast ongi need
hädavajalikud inimkeha elutegevuseks. Enamik koensüümidest on
vitamiinide derivaadid. Järelikult vitamiinide bioroll realiseerub
enamasti ja olulisel määral liitensüümi ehituse ja
funktsionaalsuse kaudu, st ensüümkatalüüsi tagamise kaudu. Samuti
mitmed ravimpreparaadid on vitamiinide koensüümvormid.
Koensüümid on
madalmolekulaarsed orgaanilised ühendid, mis on liitensüümis
valkosaga tihti mittekovalentselt seotud. Liitensüüm koosneb
tavaliselt valkosast + koensüümist
28. Hormoonid
: üldiseloomustus ja toimemehhanism
Hormoonid on
bioaktiivsed endogeensed ained, mida KNS kontrolli all sünteesitakse
spetsialiseerunud näärmetes, sekreteritakse otse verre või lümfi
ja transporditakse märklaudrakuni, millele toimides avaldubki nende
regulatoorne toime metaboolsetele protsessidele.
Signaali ülekanne
rakul rakule toimub signaalmolekulide abil.
Signaalmolekul -˃ Retseptor -˃ Sekundaarsed signaalid -˃ Spetsiifiliste ensüümide
või valkude modifitseerimine -˃ Metaboolne vastus
( Hormoon ehk
sisenõre ehk inkreet on sisenõrenäärmete poolt komplekteeritud
bioloogiliselt aktiivne ühend, mis vereringes liikudes avaldab
toimet hormooni sihtrakkudele. Hormoonid osalevad aktiivselt
imetajate (taimedel fütohormoonid) rakkude taastootmises ning
nendega seotud toimingutes. Hormoonide ringlus on katkematult seotud
väliskeskkonna ning sissesöödavate ainete ja/või sissejoodavate
vedelike ja organismi enamike elundite, kudede,rakkude ning
retseptorite ja ka geenide töös ja töötuses. Hormoonid, sarnaselt
vitamiinide ja kofaktoritega on kasvufaktorid.)
29. Hormonoidid
Koehormoonid -
Ained, mis on sarnased hormoonidele (hormonoidid)
Prostaglandiinid
(arahidoonhappe derivaadid, võivad vallandadasilelihasrakkude
kontraktsiooni, panna agregateeruma vereliistakuid või avaldada
pärssivat mõju munasarja kollakehale.
Atsetüülkoliin
(neuromediaator, substants, mis töötab närviimpulsside ülekandes
keemilise signaalkmolekulina).
Histamiin (neuromediaator,
lõõgastab veresoonte silelihasrakke, muudab veresoonte endoteeli
läbilaskvamaks ja ärritab kihelusetunnet vahendavaid sensoorseid
närvilõpmeid)
30. Antibiootikumid
Antibiootikumid
on elusorganismide (bakterite, seente) produtseeritud või
tööstuslikult sünteesitud ained, mis surmavad mikroorganisme või
pärsivad tugevalt nende kasvu ning terapeutilistes annustes ei
kahjusta makroorganismi.
Antimikroobsed preparaadid on kas
antibiootikumid või sünteetilised keemilised ühendid, mis toimivad mikroobe hävitavalt, kuid selguse mõttes kasutatakse mõlemal juhul
üldnimetust antibiootikumid.
Antibiootikumid
on tähtsaim mikroobi- ehk bakterivastase toimega ravimite rühm,
mida kasutatakse bakterite poolt põhjustatud nakkushaiguste
ravimiseks. Esimese antibiootikumi – penitsilliini – avastas Alexander Fleming 1928. aastal. Praktilisse kasutusse jõudsid
antibiootikumid 1940. aastatel ning lühikese ajaga muutsid nad
põhjalikult nakkushaiguste ja haavanakkuste ravimeetodeid, mille
tulemusena vähenes oluliselt suremus bakteriaalse päritoluga
nakkushaigustesse. Seega on oluline teada, et antibiootikumid
hävitavad ainult mikroobe ehk baktereid ning nendel puudub viiruste
vastane toime.
Algselt eraldati antibiootikumid hallitusseentest
ja mõnedest mikroorganismidest, kuid üsna varsti õpiti neid ka
kunstlikult sünteesima, mis võimaldas hakata neid massiliselt tootma . Sellega kaasnes ka antibiootikumide massiline kasutamine
ravimeditsiinis, mis tõi peagi kaasa uue ja väga tõsise probleemi
– ravimresistentsuse tekkimine.
Põllumajandusloomadel
ja -lindudel kasutatakse antibiootikume kolmel põhjusel:
nakkushaiguste ravimiseks, nakatumise vältimiseks teatud
kasvatamisperioodil ja kasvufaktoritena. Esimesel kahel juhul
manustatakse loomadele või lindudele lühikese ravikuuri jooksul
kõrges annuses antibiootikume ja kolmandal juhul – pikema aja
jooksul madalas annuses antibiootikume. Kuna kõigil nendel juhtudel
manustatakse antibiootikume suurele hulgale loomadele või lindudele,
siis on ka antibiootikumresistentsete bakterite tekkimise võimalused
suured. Loomade ja lindude organismis tekkinud antibiootikum -resistentsed bakterid võivad levida looma- ja
linnupidajate ning -hooldajate vahendusel elanikkonna sekka.
31. Aine- ja
energiavahetus: üldiseloomustus, põhietapid, assimilatsiooni (anabolismi )- ja dissimilatsiooni (katabolismi) etapid
Metabolism –
organismi elu aluseks olev biokeemiliste muutuste võrgustik.
Metabolism hõlmab seedimist, imendumist, rakus toimuvaid
metaboolseid radu ja lõpp-produktide eritumist. Põhifunktsiooniks:
energia omastamine väliskeskonnast toitainete vormis; toitainete
omastamine, lõhustamine ja kasutamine; senestsentsete biomolekulide lammutamine ; lõpp-produktide väljutamine; organismi sattuvate
ksenobiootikumide detoksikatsioon ja väljutamine.
Katabolism :
Lagundav ainevahetus – keerulisematest ainetest tekivad lihtsamad
ja vabaneb energia
Aeroobse katabolismi staadiumid:
a)
makrotoitainete (Süsiveikud, valgud, lipiidid) ja senestsentsete
biomolekulide lõhustumine monomeerideks, ehitusüksusteks.
b)
Monomeeride, ehitusüksuste muundamine vähesteks ja lihtsamateks
metabolismi võtmeühendiks.
Anabolism : Lihtsamatest
keemilistest ühenditest sünteeesitakse keerulisemaid ühendeid.
Selleks kulub energiat.
Anabolismi
staadiumid:
a)Lihtsamatest eelühenditest sünteesitakse
ehitusüksused/monomeerid.
b)Suuremate biomolekulide ja
biomakromolekulide(valgud, nukleiinhapped) süntees.
32. Seedimine,
põllumajandusloomade seede iseärasusi
Seedimise
peamiseks eesmärgiks on lagundada suure ja keerulised orgaanilise
aine molekulid selliselt , et nad oleksid aborbeeritavad ja
kasutatavad. See lagundamine toimub seedesüsteemis ensüümide
abil.
Sööda koostisosade imenduvad üksnes vesi ja mõned
veeslahustuvad ained vahetult läbi mao ja soolestiku seina looma
organism. Põhiliselt käsitletakse toitumises keemiliste
struktuuride väiksemaid ühendeid, mis lagundatakse lihtsateks
absorbeeruvateks ühenditeks või elementidek ( st juhitakse
soolekulglast looma organism). Neid protsesse nim. Seedimiseks –
eristatakse mehaanilist seedimist ja keemilist
lõhustumist.
Mehaaniline seedimine – toimub eelkõige
mäludes, aga ka lihaskontraktsiooni abil , nt lindude lihasmaos –
selle ül.on sööda peenestamine. Mälumisprotsessis segavad imetajad sööda süljega, et seda pehmendada ja osaliselt lahustada
ning muuta tükid allaneelamiseks piisavalt libedaks Mäletsejalistel
reguleerib sülg ka eesmao pH-d.
Teised
mehaanilised protsessid tagavad söödatranspordi läbi seedetrakti.
Söögitoru peristaltilised liigutused suunavad sööda makku.
Söögitorus ja sooltes toimub sööda põhjalik segamine . Sooleperistaltikal on kaks funktsiooni: toidu läbisegamine ja
transport. Transpordimehhanismid rakenduvad kogu seedetrakti ulatuses
kuni roojamiseni.
Keemiline seedimine – toimub ensüümide
abil. Seejuures eristatakse seedimist nii imetajate kehaomaste
ensüümide kui ka mikroobide poolt produtseeritud ensüümide
toimel. Viimastel on tähtis roll rohusööjate toitumises.
Ensüümid on
biokatalüsaatoritena toimivad ühendid (valgud), mis käivituvad või
kiirendavad söödaosakeste keemilist lõhustumist, Ensüümide
gruppe tähistatakse ainerühmade järgi, mida nad lõhustavad. Nii
nim. Proteiini lõhustuvaid ensüüme proteaasideks või
proteinaasideks, tärklist lõhustavaid karbohüdraasideks (nt
amülaas), rasvu lõhustavaid lipaasideks.
Seedekulgla sekreedid sisaldavad erinevaid ensüüme. Ensüümide funktsioonidele
vastavalt tekivad lõpp-produktina erinevad ühendid. Tähtsaimad
absorbeeruvad ühendid on tärklist lõhustumisel tekkiv glükoos,
rasvade lõhustumisel tekkivad rasvhapped ja glütseriin ning
proteiini lõhustumisel tekkivad aminohapped.
Suur osa
süsivesikuid esineb söödas, eriti koresöödas mitte tärklise ega
suhkruna, vaid komplekssete ühenditena, nt tselluloosi või
hemitselluloosi kujul. Selliste, ka tugiaineteks nim.ühendite
lagundamiseks pole imetajatel kehaomaseid ensüüme. Neid võivad
sünteesida ainult mikroorganismid , mida rohusööjate seedekulglas
on piisaval arvul, et söödaosakesi lagundada. Mikroorganismid
asuvad jämesooles (hobune, närilised) või mäletsejalistel vatsas ,
mis kujutab endast eelkäärimiskambrit.
Selline asmeline
seedimine annab mäletsejalistele kaks eripära: suurema osa
söödaenergiast saavad nad seedimisprotsessi lõppjärgus mikroobse
ainevahetuse produktidest, seega erinevalt lihtmaoliste
energiaoamastamisest, mis toimub seedunud toitainete, nt glükoosi
kaudu. Teiseks tagab mikroorganismide pidev kasv ja hukkumine
väärtusliku proteiini juurdevoolu makku( libedikku) ja soolestikku.
See võimaldab mäletsejalistel ka vähese proteiinisisaldusega sööda
korral elada ja toodangut anda juhul, kui vatsas on tagatud piisav
varustatus lämmastiku ja muude toitainetega .
33.
Energeetiliste protsesside spetsiifika loomorganismis, makroergilised ühendid
Katabolism ja
anabolism on energeetiliselt seostunud. Katabolismis salvestab
organism anabolismiks vajatavat energiat.
Inimkeha
biokeemia ei süüvi bioenergeetika detailidesse. Piirdutakse
üldarusaamadega lõhustatava substraadi(ka toitaine) energia
konverteerumisest inimkehas kasutatavaks metaboolseks energiaks ja
selle kasutamisest anabolismis, füsioloogilisteks funktsioonideks,
talitlustes. Vajalikke põhimõisteid interpreteerib meditsiiniline keemia lihtsustatult järgmiselt:
Vaba energia –
orgaanilise ühendi koguenergia see osa, mille arvel organismis saab
teha tööd (antud temperatuuril, konstantsel rõhul).
Vaba energia muut
– biokeemilise protsessi suuna ja võimalikkuse määrab vaba
energia muut. Negatiivse vaba energia muuduga biokeemiline protsess
kulgeb spontaanselt (kataboolsed reaktsioonid). Vaba energia muut
sõltub reageerivate ühendite ja produktide kontsentratsioonist,
temperatuurist, pH-st ja rõhust.
Vaba energia
standardmuut – vaba energia muut, kui reageerivate ühendite
kontsentratsioon on 1M, pH on 7,0 ja temperatuur 25oC.
Vajalik reaktsiooni võimalikkuse ja suuna hindamiseks.
Väike arv
orgaanilisi ühendeid, mille sideme hüdrolüüsi vaba energia
standardmuut on üle -25kJ/mol (suur vaba energia muut ei tulene
sidemest, vaid ühendi elektronkatte ja konformatsioonilisest
eripärast nagu laengute jaotuvus , resonants jne). Makroergilised
ühendid töötavad energia ülekande/doonorvormidena.
Makroergiliste
ühendite põhiesindaja on ATP. ATP pole kõige makroergilisem
fosfaat, kuid ta hüdrolüüsi vaba energia muut on tunduvalt väiksem
teiste fosfaatide omast. ATP lõhustumisega ADP-ks kaasuv terminaalse
fosforüülgrupi ülekanne biomolekulile tagab suure energiamuudu
tõttu biokmolekuli aktiveerumise. Nii rakendub ATP-s salvestatud
metaboolne energia füsioloogilisteks funktsioonideks.
Põhilised
makroergilised ühendid on makroergilised fosfaadid(ATP, GTP, UTP,
CTP, ADP, keratiinfosfaat, 1,3-bisfosfoglütseraat,
fosfoenoolpüruvaat), makroergilised tioolestrid(atsetüül-CoA,
suktsinüül-CoA jt), tsüklilised nukleotiidid( cAMP , cGMP)
UDP-glükoos, CDP- koliin , S-adenosüülmetioniin jt.
34.
Sahhariidide ainevahetuse üldiseloomustus. Sahhariidide seedimine ja
imendumine. Sahhariidide tähtsus toitumisel.
Ühe grammi
süsivesikute täielikul lõhustumisel vabaneb 17 kJ ( 4 kcal )
energiat. Kõige kiiremini kasutatav energiavaru. Aju energeetilised vajadused rahuldatakse peaaegu täies mahus glükoosi arvel.
Tasakaalustatud toidu puhul moodustub põhilisest osast verre
sattunud glükoosist energia, mida rakud kasutavad oma elutegevuses.
Ligikaudu 30% glükoosist muudetakse neutraalrasvaks ja rasvhapeteks,
ligikaudu 3% moodustub glükogeen, mis ladestub maksas ja lihastes.
Süsivesikud peaksid andma ligi 60 % päevasest energiast.
Süsivesikute defitsiidi korral muudetakse organismis talletunud rasv
energiaks, mille käigus eralduvad ketokehad ning see võib
põhjustada ketoosi.
- Süsivesikute katabolism peab andma umbes poole (50-60%) organismi toiduenergia vajadusest. Tavatingimustes on glükoos mõnede kudede, organite jaoks ainsaks sisuliseks energiasubstraadiks.
- Süsivesikute metabolism tagab veresuhkru taseme hoidmise normi piirides
- Süsivesikute metabolism tekitab monosahhariidseid eelühendeid
- Häired süsivesikute metabolismis avalduvad haiguste kujul. Nii on glükoosi metabolismi defektid põhialuseks kahele üldisele metaboolsele haigusele: suhkurtõbi ja rasvumine .
Sahhariidide
metabolism on sisuliselt glükoosi metabolism. Teiste monooside
metabolism sulandub glükoosi metabolismi. Glükoosi universaalsuse
peapõhjused:
- Ta lahustub väga hästi vees ja tema tsükliline struktuur on optimaalse stabiilsusega
- Vaba glükoos on organismis keemiliselt suhteliselt inertne (tema muundumine toimub vaid ensümaatiliselt ja seega täpse kontrolli all)
- Ta on metaboolne põhikütus enamike organismide jaoks
- Ta läbib piisava kiirusega hemato-entsefaalset barjääri tagamaks ajukoe en.vajadused
- Ta on tavatingimustes praktiliselt ainus arvestatav metaboolne „kütus“ ajukoe, testiste jaoks.
Suus
peenestatakse toit ning amülaasi toimel algab süsivesikute
seedimine. Tärklis lõhustub lihtsama ehitusega suhkruteks. Edasi
liigub toit neelust mööda söögitoru makku, kus jätkub
süsivesikute lagunemine. Toidukört liigub kaksteistsõrmiksoolde
ning peensoolde , kus lõpeb sahhariidide lagunemine ning imendumine
verre.
35. Glükolüüs
ja glükogenolüüs
Glükolüüs on
rada, mille käigus toimub heksooside, eelkõige glükoosi
oksüdatiivne lõhustamine püruvaadini. Selles protsessis
moodustub 2 ATP-d.
Lõhustumiseks peab glükolüüs sisenema rakkudesse. Inimkeha rakkudes muundatakse glükoosis olev energia
oksüdatiivse lõhustumisega metaboolseks energiaks (ATP, NADPH ) ja
toodetakse glükoosist vajalikke metaboliite. Glükolüüs on
glükoosi oksüdatiivne lõhustumine. See on glükoosi
metaboliseerumise keskne protsess. Sõltuvalt tingimusest on see
protsess osaline või täielik.
Osaline lõhustumine piimhappeks
toimub hapniku defitsiidis ja see on anaeroobne glükolüüs. Täielik
lõhustumine toimub aeroobsetes tingumustes – aeroobne
glükolüüs.
Anaeroobne glükolüüs – rakku
toodud glükoos fosforüülitakse koheselt ATP-ga Glc-6-P-ks.
Fosforüülimine on glükoosi suunamine metabolismi.
Aeroobne
glükolüüs – aeroobsus võimaldab süsivesikute täielikku
oksüdatiivset lõhustumist süsihappegaasiks ja veeks.
Glükogenolüüs toimub tsütoplasmas. Vabastab vajadusel
kiiresti/rohkesti glükolüüsi, söömisvaheaegadel, töötavas lihases . Võtmeensüüm –glükogeeni fosforülaas
Glükogenolüüs
– maksa ja lihaste glükogeeni lõhustumine pole glükogeneesi
pöördrada. Rakenduvad hoopis teised ensüümid. Glükogenolüüsi
kesksed momendid on:
1. Ahelate lühendamine (1,4 sidemete
lõhustamine)
2. Hargnemispunktide elimineerimine (1,6 sidemete
lõhustamine
3. Glükoos-1-P konversioon glükoos-6-P-ks
36.
Käärimised. Sahhariidide ainevahetuse eripära mäletsejatel.
Käärimine –
anaeroobne glükolüüs. Toimub anaeroobses keskkonnas ühenditeni,
mille edasine oksüdatsioon saab toimuda ainult hapniku osalusel.
Laktaadi teke glükoosist - piimhappe-käärimine.
Anaeroobsetes
tingimustes glükoosist etanooli teke - alkoholkäärimine.
Sahhariidide
ainevahetuse eripära mäletsejatel:
Mäletsejatel
loomadel on tselluloosi (taime rakukesta) lagundamine tüüpiline
protsess. Inimorganism tselluloosi ei omasta , vaid väga vähesel
määral toimub meie soolestikus selle bakteriaalne lagundamine. Maos
olevad bakterid muudavad tselluloosi veisele kasulikuks olluseks.
37.Püruvaadi
oksüdatiivne dekarboksüülumine. Trikarboksüülhapete tsükkel.
Glükolüüsi
käigus toimub glükoosi oksüdatiivne lõhustamine püruvaadini.
Dekarboksüülumine – keemiline reaktsioon, milles
karboksüülrühma COOH lõhustamisel eraldub CO2.
Mitokondrites
toimuv püruvaadi pöördumatu oksüdatiivne dekarboksüülimine
atsetüül-CoA(koensüüm A)-ks (kogu metabolismi keskne vaheühend)
on üks suure energiamuuduga multiastmelistest võtmeprotsessidest
inimkeha kogu metabolismis. Seda viib läbi püruvaadi
dehüdrogenaasne kompleks ( PyrDH ). Kompleksi 3 ensüümi ja 5
koensüümi kontakteeruvad füüsiliselt ja seetõttu vaheühendid vahepeal ei vabane.
Atsetüül CoA peamine ülesanne on süsiniku
aatomite toomine atsetüülgrupina tsitraaditsüklisse, kus seda
energia saamiseks oksüdeeritakse.
Trikarboksüülhappe
tsükkel (TKT, tsitraaditsükkel, Krebsi tsükkel) on mitokondrites
toimuv tegelikult organismi keskne metaboolne rada, mis seostab
süsivesikute, lipiidide, aminohapete metabolismi. Kataboolses
mõistes on TKT põhirolliks atsetüül-CoA täielik lõhustumine
süsihappegaasiks ja veeks ning lõhustumine energiamuudu arvelt
rohke ATP tootmine. Glükoosi täielik oksüdatsioon toimub TKT ja
hingamisahela koostöös.
Tsitraaditsükli käigus oksüdeeritakse
enamik sahhariide , rasvu ja valke CO2ks ja veeks. Selle
käigus vabaneb suur osa organismi elutegevuseks vajalikust
energiast, mitmete anabolismi reaktsioonide eelduseks .
38.
Pentoosfosfaaditsükkel ja selle bioloogiline tähtsus.
Glükoosi
aeroobse oksüdatsiooni rada, toimub tsütoplasmas
• I etapp:
pöördumatu, oksüdatiivne, tekib NADPH, võtmeensüümiks G6PDH
• II etapp:
pöörduv, tekib riboos -5-P
Toodab
pentoosfasfaate ja NADPH( redutseeriv metaboolne energia)
NADPH on
redutseeriv energia, mis kasutub paljudes sünteesides, on vajalik
ensüümidele ja GSH taseme hoidmiseks (GSH kaitseb rakke
oksüdatiivse stressi eest)
Hõlmab 15-30% kogu glükoosi
katabolismist (toimub aktiivselt maksas, neerupealistes,lakteerivas
piimanäärmes, rasvkoes , erütrotsüütides)
Toodab peaaegu poole
redutseerivast energiast
NADPH kasutub erütrotsüütides GSH
taseme hoidmiseks(kaitseb neid kahjuliku oksüdatiivse stressi
eest/stabiliseerib hemoglobiini)
Toodab riboos-5-P
Riboos-5-P
on vajalik nukleotiidsete koensüümide ja nukleotiidide
sünteesiks
Võib anda panuse ATP sünteesiks
Võimaldab
utiliseerida toiduga organismi sattuvaid pentoose
G6PDH defitsiiti
põhjustab X-liiteline geenmutatsioon
PFT ja glükolüüs
on väga tihedalt integreeritud.
PTF on tihedalt seotud
nukleotiidide sünteesiga.
39. Sahhariidide
biosüntees
Pentoosid on
nukleiinhapete ehituskompleksiks. Süsivesikute ainevahetuse
vaheproduktidest algavad osade aminohapete ja lipiidide
sünteesirajad. Kergesti omastavate süsivesikute ülekülluse korral
suunatakse ülejääk lipiidide biosünteesile (rasvumine), seda
kontrollib insulin, samuti säilitab ta rasvkudet. Süsivesik
(ribuloos-1,5- difosfaat ) on fotosünteesis CO2 esmane siduja.
Süsivesikute
metaboliidid laktaat . Püruvaat jt on rasvhapete, asendatavate
aminohapete jt ühendite süsinikskeleti loomise aluseks
Loomaorganismid pole
võimelised süsivesikuid anorgaaniilistest komponentidest sünteesima
ja neid suuremates kogustes deüoneerima. Loomses ainevahetuses
iseloomustab süsivesikute käivet laegunemisreaktsioon : C6H12O6+602
-> 6CO2+ 6H2O , rohelisetes taimedest toimub aga vastupidine protsess. Süsivesikud moodustavad taimorganismides klorofülli
katalüütilisel toime päikeseenergia arvel ning ladestatakse mono-,
di- ja polüsahhariididena mitmesugustes taimeosades varuainetena’
40.Bioloogiline
oksüdatsioon. Hingamisahela ensüümid.
Bioloogiline
oksüdatsioon ehk bio-oksüdatsioon on organismides toimuv
paljuetapiline redoksreaktsioonide ahel, mille tagajärjel moodustub
lõpp-saadustena peamiselt süsihappegaas, vesi ja ammoniaak , mis
organismidest väljutatakse.
Bio-oksüdatsiooni
eripära võrreldes paljude keemiliste oksüdatsiooniprotsessidega
(näiteks põlemisega) on see, et need toimuvad tunduvalt madalamal
temperatuuril – selle tagab vastavate ensüümide katalüütiline
toime.
Hingamisahela
ensüümid. Mitokondrite hingamisahel on ETA-põhise bioloogilise
oksüdatsiooni keskne variant inimkehas. Tema põhiroll on metaboolse
energia (ATP) tootmine. Häired tema töös on organismi jaoks
enamasti kriitilised ja tihti fataaslsed
Hingamisahela
ensüümid :
Oksüdaasid – katalüüsivad otsereaktsiooni
õhuhapnikuga.
Oksügenaasid – lülitavad hapnikumolekulid
substraadimolekuli
Hüdroksüperoksüdaasid – katalüüsivad
vesinikperoksiidi või lipiidide hüdroperoksiide
Dehüdrogenaasid
– Bioloogilise oksüdatsiooni kesksed ensüümid. Hingamisahela
komponendid on NAD-dehüdrogenaasid, FMN-, FAD- ja
CoQ-dehüdrogenaasid – võtvad substraadilt vesinikuaatomeid.
Hingamisahela keskne komponent on koensüüm Q, sest kõik
ahela variandid läbivad oma teel koensüüm Q süsteemi.
41.
Oksüdatiivne fosforüülumine.
Elektronide
transpordil hingamisahelas toimuvad redoksreaktsioonid (oksüdatsioon)
annavad energia prootonite elektrokeemilise gradiendi tekkeks läbi
mitokondrite seisemembraani. Prootonite liikumine gradienti pidi
maatriksisse annab ATP süntaasi tööks (ADP fosforüülimine
ATP-ks) vajaliku energia.
42.Lipiidide
tähtsus toitumisel. Lipiidide muundumine seedetraktis, sapi tähtsus
seedimisel. Imendumine.
Lipiidide
põhiülesanded:
Rasvhaoete katabolism (oksüdantsioon) annab
inimkehas põhiosa vajatavast metaboolsest energiast.Lipiidid annavad
25-30% toitelisest energiast ja rasvkoe triglütseriidid moodustuvad
83-87% inimkeha energiavarudest.
Rasvhapete ja
regulaatormolekulide(eikosandoidid,lipiidide hüdroperoksiidid jt)
süntees.
Inimkeha-spetsiifiliste triglütseriidide,liitlipiidide
ja tsükliliste lipiidide süntees.
Ketokehade süntees ja
lõhustamine.
Lipiidi-sarnaste
biomolekulide( kolesterool , steroidid ,vitamiin , sapphapped jt) süntees.
Vere lipoproteiinide süntees lipiidide,
lipiidi-sarnaste ühendite,vitamiinide jne
taransport.
Triglütseriididel on kõrge energeetiline (oksüdatiivne) väärtus ja nad on inimkeha energia põhivaru.
Täidavad energeetilisi, mehaanilisi kaitsefunktsioone ja
spetsiifilisi biofunktsioone. Biomembraani moodustavad fosfolipiidid ,
lipiidid on rasvlahustuvate ühendite (vitamiinid, paljud ravimid)
lahustiks ja transportijaks. Normaalne metabolism sõltub lipiidide
hulgast, vahekorrast, tüübist. Mitmed rasvhapped on otsesed geenide
regulaatorid. Häired lipiidide metabolismis tekitab haigusi
(ateroskleroos, rasvumine).
Lipiidide
lõhustamisel on vaja emulgeerimist, põhiemulgaatoriteks on
sapphapped ja nende soolad . Sapphapete kaudu viiakse suur osa
kolesterooli inimkehast välja.
43.Glütserooli
ja rasvhapete oksüdatsioon kudedes. Rasvhapete β-oksüdatsioon.
Oksüdatsiooniks
kasutatavad rasvhapped pärinevad põhikoguses varurasvade
mobilisatsioonist (rasvkoe TG’de lõhustumine rasvhapeteks ja
glütserooliks). Adipotsüütides toimub pidev süntees ja
lõhustumine. Mobilisatsioonil (TG’de lipolüüsil) toimub rasvkoe
TG’de hüdrolüüs rasvhapeteks ja glütserooliks lipaasidega
hormoonide kontrolli all. Protsessi algatab hormoon-sõltuv TG lipaas , mis vabastab asendist 1 või 3 rasvhappejäägi. Tekkinud
diglütseriidi (DG) lõhustab DG lipaas. Saadud monoglütseriidi
lõhustab MG’i lipaas rasvhappeks ja glütserooliks. Ehk TG
lõhustumine vabadeks rasvhapeteks ja glütserooliks toimub
adipotsüütides, mil lipolüüsi käivitab TG lipaas, mis aktiveerub lipolüütiliste hormoonide toimel.
Rasvhapete B-OX
on rasvhappe aktiivvormi (atsüül-CoA) oksüdatsiooni beeta-süsiniku
osalusel mitokondrite maatriksis. 1 B-OX ring eraldab rasvhappe
ahelast 2-süsinikulise aktiivse atsetüüljäägi (atsetüül-CoA
vormis), st 1 ringiga lüheneb rasvhappe ahel 2 C võrra. Iga B-OX
ring lubab toota kuni 15 ATP molekuli koostöös TKT
(trikarboksüülhapete tsükkel) ja hingamisahela.
Rasvhapete
β-oksüdatsioon on organismi energiatootmise üks peamistest
ainevahetuse radadest ning skeletilihaste ja südamelihase peamiseks energiallikaks . Rasvhapete oksüdatsioon hakkab mängima olulist
rolli organismi energiaga varustamisel just metaboolse stressi
tingimustes (kaasnev infektsioonhaigus, paastumine , suur füüsiline
koormus), kui glükogeenivarud on ära tarvitatud.
44. Rasvhapete
ja triglütseriidide biosüntees. Ketoosid .
Rasvhapete
vajaduse katab organism kahel viisil: suurem määral tasakaalustatud
toiduga või sünteesib ise. Üle vajaduse saadud toidusüsivesikud
konserveeritakse rasvhapeteks ja salvestatakse rasvkoe
triglütseriididena.
Rasvkoe rasv pole
staatiline varu. Toimub pidev süntees ja lõhustamine. Rasvhappe de
novo süntees (lihtsamatest eelühenditest toimuv rasvhappe uuesti
süntees) toimub maksas ja lakteerivas piimanäärmes. Ei ole B-OX
pöördprotsess. Toimub tsütoplasmas ja aktiivne
ekstramitokondoriaalne rasvhappe süntaas viib läbi kogu sünteesi.
Lähteühend on atsüül-CoA. Vajab redutseerivat energiat (NADPH),
ATP. Prevaleerib süsivesikuterikka toidu tarbimisjärgselt ja teda
soodustab kõrge insuliin/glükagoon.
Ketoosid on monosahhariidid,
mis sisaldavad ketorühma (RC(=O)R'). Ketooside hulka kuulub fruktoos
ehk puuviljasuhkur.
45. Steroidide
ja fosfolipiidide ainevahetus
Steroidid on
lipiidide üks alarühm, mille hulka kuuluvad ka steroolid , milledest
tuntuim on kolesterool.
Fosfolipiidide baasalkohol on
glütserool. Jaotus: fosfatidüülkoliinid ehk letsitiinid,
fosfatidüületanoolamiinid, fosfatidüülseriinid,
fosfatidüülinositoolid, fosfatodüülglütseroolid, kardiolipiidid
ja plasmalogeenid.
Süntees: toimub kõikide rakkude (peale
erütrotsüütide) siledapinnalisel endoplasmaatilisel retiikulumil.
Sünteesitud fosfolipiide viivad transportvalgud Golgi kompleksi
kaudu plasmamembraani, rakuorganellide membraani või nad
eksotsüteeritakse. Sünteesiks vajalik fosfatidaat tekib samal
viisil nagu triglütseriidide sünteesil: ta on TG-de ja
fosfolipiidide sünteesi eelühend. Kuna mõnede fosfolipiidide
uuenemine peab toimuma ülikiiresti, töötab abimehhanism:
fosfolipiidide kiirsüntees membraanis peagrupi vahetuse või
modifitseerimisega.
Fosfolipiidid on head detergendid hoidmaks
sapis olevat kolesterooli lahustuvana. Häired fosfolipiidide
sünteesis soodustavad kolesteroolist ja sapipigmentidest koosnevate sapikivide teket.
Kasutades sünteesi põhiradu, peagruppide
vahetust ja modifikatsiooni ning remodelleerimist, on inimkeha
võimeline sünteesima vajalikes kogustes ja vahekordades nii
tüüpilisi fosfolipiide kui ka spetsiifilisi fosfolipiide
biofunktsioonide täitmiseks.
Katabolism: Fosfolipiide
lammutavad kudede fosfolipaasid ja iga ensüüm lõhustab kindlat
sidet. PLA1 eraldab estersideme lõhkumisega asendist 1
rasvhappejäägi. Seda leidub paljudes kudedes, aga ka mesilase- ja
maomürgis.
Kaltsium-aktiveeritud PLA2 eraldab estersideme
lõhkumisega rasvhappejäägi asendist 2 (reeglina küllastamata
rasvhappejääk). Teda leidub paljudes kudedes. PLA2 toimet pärsivad
glükokortikoidid, ühtlasi on oluline ka fosfolipiidide
remodelleerimises.
46. Rasvhapete
peroksiidne oksüdeerumine. Arahhidoonhape . Prostaglandiinid.
Rasvhapete
oksüdeerimise põhikoht on mitokondrid . Kui toidus on liigselt
triglütseriide, intensiivistub maksarakkude peroksüsoomides
rasvhapete oksüdatsiooni ensüümide süntees. Peroksümaalse
B-OX(rasvhappe oksüdeerumise) mõte on: 1) tekitada väga
pikkade ahelatega rasvhapetest lühemaid hõlpsasti edasi lõhustuvaid
rasvhappeid; 2) osaleda rasvhapete liigsuse likvideerimises tootes
samal ajal ATPd; 3) lühendada sapphapete formeerumiseks kolesteriidi
rasvhappeahelat.
Arahhidoonhape on omega 6 rasvhape,
loomse päritoluga. Arahhidoonhapet talletatakse rakumembraanides
ning tema ülesanne on saata signaale muutusteks juhul, kui tegemist
on lihaskahjustustega. Arahhidoonhape on väga oluliselt seotud
valgusünteesimise protsessis ning lihashüpertoofias peale
treeninguid.
Arahhidoonhape samuti võimendab IGF-1 (Insulin-Like Growth Factor ) signaalide saatmist, parandab keha lihasmassi
taastumist. Samuti suurendab Arahhidoonhape lämmastik oksiidi (NO)
tootlikkust lihastes.
Lisaks parandab arahhidoonhape lihastes
veresoonkonda, kuna stimuleerib ka veresoonte
taastumist.
Prostaglandiine toodetakse paljude rakkude
poolt; põhjustab silelihaste kontraktsiooni, trombotsüütide
agregatsiooni, põletikku. Erinevalt teistest signaalmolekulidest
eksotsüteeritakse prostaglandiine konstitutiivselt; kui aga rakke
aktiveerida koe vigastamise või mingi keemilise signaali poolt, siis
nende süntees tõuseb. Teatud prostaglandiine toodetakse suures
hulgas emakas , kui algab sünnitustegevus. Need on olulised emaka
silelihaste kontraktsiooni stimuleerimisel. Neid kasutatakse ka
meditsiinis kunstliku abordi esilekutsumiseks. Prostaglandiinid
indutseerivad ka näiteks lima eritumist mao limaskestast.
Prostaglandiinide sünteesi aga omakorda indutseerib näiteks
alkohol. Siit on arusaadav, miks alkoholi joomisel eritub maos
rohkesti lima.
47. Valkude
ainevahetus: üldiseloomustus, valkude tähtsus toitumisel. Organismi
lämmastikubilanss.
Valkude pidevat
lammutamist ja sünteesi inimkehas nimetatakse kehavalkude käibeks.
Sisuliselt on tegemist aminohapete metabolismiga. Aminohapete kaudu
tuleb toiduga kehasse lämmastik. Lämmastik väljub karbamiidi,
kusihappe, ammooniumsoolade kujul, mis on aminohapete metabolismi
lõpp-produktideks.
Valgud täidavad kehas unikaalseid
biofunktsioone. Plasmavalkude põhifunktsioonid: vere kolloid-
osmootse rõhu säilitamine; transport; vere pH säilitamine;
kaitsefunktsioon; ensümaatiline roll.
Lämmastiku balanss -
Lämmastik on keha üks põhibioelementidest. Tasakaalustatud
segatoidu sööval on lämmastiku ringlus tasakaalus. Tasakaalustatud
toitumine tagab, et praktiliselt kogu lämmastik pärineb
toiduvalkudest. Väljutav lämmastik lahkub inimkehast põhiliselt
aminohapete metabolismi lõpp- produktidena.
48.
Aminohapete üldine ainevahetus
On kudedes
toimuva valguainevahetuse olulisimad substraadid. Neist sünteestakse:
valku, ensüüme, nukeiinhappeid, hormoone jne. Võimalik kasutada
ka enegeria saamiseks süsinikuahela oksüdeerimisel. Vere
aminohapete (AH) sisaldus on võrdelmisi püsiv.Tasakaalu võimalik
lühiaegselt höirida AH viimisega verre või proteiinirikka
ratsiooniga.
Ainevahetuslike aminohapete peamised allikad:
-
vatsas mikroobide poolt lõhustamata jäänud
valgud
-mikroorganismide poolt sünteesitud valgud
-vatsavedeliku
vabad aminohapped
seedenõredega peensoolde jõudnud valgud ja
aminohapped.
Inimese
lämmastiku ainevahetuse jääkprodukt on uurea . Uurea sünteesitakse
maksas siis, kui organismi on liigselt aminohappeid. Aminohapetest
tekib algul ammoniaak, sellest sünteesitakse uurea.
49. Valkude
muundumine seedetraktis. Valkude ainevahetuse eripära mäletsejatel.
Roiskumine
Seedetraktis
lõhustatakse valgud üle mitmete hüdrolüüsi vaheproduktide kuni
aminohapeteni ja sellistena kasutatakse organismis kas plastiliseks
otstarbeks või energeetilise lähteainena. Suus ei toimu erilisi
muutusi, sest süljes puuduvad proteolüütilised ensüümid. Valkude
seedimine algab maos, kus toiduvalkudele toimib maonõre selles
leiduvate ensüümidega. Maonõre kuivaine orgaaniline osa sisaldab
mitmeid valke (mütsiini, seerumialbumiine ja –globuliine),
ensüüme, aminohappeid ja vitamiine. Pepsiin on tähtsaim ensüüm,
mis katalüüsib valkude lõhustumist. Pepsiini toimel lõhustuvad
valgu molekulid üle peptoonide ja teiste hüdrolüüsi
vaheproduktide osaliselt kuni aminohapeteni. Peensooles toimub
toitainete edasine lõhustumine ja laguproduktide imendumine.
Seedetrakti ülemistes osades osaliselt hüdrolüüsunud toidu
koostisosadele avaldavad peensooles mõju pankrease (olulisim
osa,toimides kõikidesse toidu koostisosadesse) ja peensoole nõre
ensüümid.
Bioloogiliselt madala väärtusega taimsed valgud
muudetakse kõrgeväärtuslikuks bakteriaalseks valguks, mis
seedetrakti alumistes osades kergesti lõhusub ja pärast
resorptsiooni lülitub looma ainevahetusse. Mäletsejaliste
eesmagudes toimivad valkudesse bakterite, infusiooride ja pärmide
ensüümid. Toime iseloomu poolest sarnanevad bakteriallsed ensüümid
trüpsiiniga, lõhustades valgu molekulides pepsiidsidemeid pH 6-7
juures. Osade ensüümide toimel lõhustuvad peptiidahelad
aminohapeteni. Osa aminohappeid allutatakse ka desamiinimisele.
Roiskumine algab valkude lõhustumisega mikroobide ensüümide
toimel aminohapeteni ja edasi nende mitmesuguste laguproduktideni,
mis on tihti toksilise iseloomuga . Roiskumisel täheldatakse
hüdrolüüsi, desamiinumist, dekarboksüülumist, oksüdatsiooni,
reduktsiooni, metüülumist ja demetüülumist. Nende protsesside
tulemusena tekib ammoniaaki, orgaanilisi happeid, aldehüüde,
alkohole, ketoone, amiine jt ühendeid. Türosiin võib roiskumisel
muutuda 2 viisil – desamiinumise voi dekarboksüülumise kaudu,
lõpp-produktiks on mõlemal juhul fenool ja kresool. Trüptofaani
lõpp-produktiks on indolüüläädikhape, skatool ja indool.
Need
ühendid on toksilised ja võivad pärast resorbeerumist esile
kutsuda mürgitust.
50. Ammoniaagi
eemaldamine organismist
Põhiline osa
lämmastikust eemaldatakse organismist imetajatel kusiainena ning
lindudel ja reptiilidel kusihappena ning teise võimalusema seotakse
ammoniaak valkude karboksüülrühmadega, nii kaitstakse organismi
ammoniaagi toksilise mõju eest kui primaarsed mehhanismid . Kusiaine süntees maksas on ammoniaagi mürgistustumise ja organismist
eemaldumise protsessi viimane etapp. Kusiaine moodustamiselt
kasutatakse lisaks vabale ammoniaagile ka mitmesuguseid ogaaniliste
ühendite aminorühmi. Kusiaine eemaldatakse organismist neerude
kaudu uriiniga.
51. Valkude
biosüntees
Organismi rakud
ei suuda sünteesida mitmeid aminohappeid, neid nimetatakse
asendamatuteks aminohapeteks (Val, Ile, His, Met, Leu, Lys, Phe, Thr,
Trp). Biosünteesivõimetuse põhjuseks on nendele aminohapetele
vastavate α-ketohapete puudumine või vähesus organismis.
Ülalnimetatud aminohapete saamine toiduga on hädavajalik.
Asendavate
aminohapete sünteesi üldrada on nende süntees α-ketohapetest.
Transamiinimisega sünteesitakse Pyr-st alaniini, OAA-st aspartaati
ja AKG-st glutamaati. Argiini sünteesitakse neerudes.
52.
Nukleiinhapete ainevahetus
Nukleiinhapped on
biopolümeerid, milles nukleotiidijäägid on seostunud
fosfodiestersidemetega.
Nukleiinhapete
moodustamiseks vajalikud lähteained pärinevad toidust
(nukleiinhapete hüdrolüüsiproduktid) ning süsivesikute ja
lipiidide ainevahetusest. Puriin- ja pürimidiinalused ehitatakse
vajaduse korral üles lihtsamatest ainevahetuse vaheproduktidest.
Nukleiinhapete hüdrolüüs algab maos, kus pepsiini toimel
lõhustuvad sidemed valguliste komponentide ja nukleiinhapete vahel.
Protsess jätkud peensooles, kus algab ka lihtvalkude järkjärguline
hüdrolüüs aminohapete vabanemiseni. Need resorbeeruvad ja nad
lülitatakse edasistesse aminohapete ainevahetuse reaktsioonidesse.
Nukleiinhapped alluvad pankrease nõre nukleaaside toimele. Rakkude
nukleiinhapped lagunevad vastavate nukleaaside (DNA-aaside,
RNA-aaside) katalüütilisel toimel mononukleotiidideni. Osa neist
kasutatakse kudedes moodustuvate uute rakkude nukleiinhapete sünteesi
lähteainetena, teine osa lõhustatakse nukleosiidideni ja
fosfaatideni või pentoosfosfaatideni ja lämmastikalusteni.
Nukleotiidide hüdrolüüsil saadakse puriin- ja pürimidiinalused,
pentoosid ja fosforhape .
53. Vee- ja
mineraalainevahetus
Vee jaotus kudede
ja organite vahel pole ühtlane. Vesi on koe koostisosade solvendiks
ja kõikide kudede ning rakkude ehituselemendiks. Enamik ainevahetuse
reaktsioonidest toimub vesilahuses või vees dispergeerunud
aineosakeste piirpindadel. Vesi on toitainete ja ainevahetuse jääkide
põhiliseks transportijaks. Vee otsesel osavõtul toimuvad
organismisarvukad keemilised reaktsioonid – hüdrolüüs,
oksüdatsioon jt.
Termoregulatsioonimehhanismis võimaldab
nahakaudne vee auramine keha kõrge välistemp korral jahutada. Vesi
eemaldatakse organismist neerude, kopsude, naha ja pärasoole kaudu.
Veevahetus seostub organismis tihedalt toitainete vahetusega.
Veevahetust reguleeritakse organismis närvisüsteemi ja kilpnäärme
suprarenaalnäärmete koore, hüpofüüsi, pankrease ja sugunäärmete
hormoonide kooskõlastatud talitluse kaudu.
Kuigi mineraalainete
sisaldus on kudedes kvantitatiivselt orgaaniliste aine hulgaga võrreldes märgatavalt väiksem, osutavad nad absoluutselt
eluvajalikeks. Organismis esinevad mineraalained lahustumatute
sooladena peamiselt luudes, lahustuvate ühenditena kudedes ja
koevedelikes ning seotult orgaanilise ainega. Organism saab
mineraalaineid toidu ja joogiveega. Enamik neist resorbeerub
peensoolest verre ja antakse edasi kudedele ning koevedelikele.
Mineraalainete vahetus vere, kudede ja koevedelike vahel toimub osmoosi seaduspärasuste alusel – ioonid liiguvad üldjuhul
madalama kontsentratsiooni suunas, Mineraalained erituvad põhiliselt
uriini, higi ja roojaga.
54.
Ainevahetuse põhiradede vahelised seosed
Ainevahetus e.
metabolism. Metabolismi moodustab metaboolsete radade võrgustik.
Rakusisene
metabolism toimub metaboolsete radadena, kus ensüümide toimel muunduvad/tekivad metaboliidid. Metaboolne rada on reaktsioonide jada
(nt glükoneogeenees on üksikreaktsioonide jada, mille tulemusena
piimhappest sünteesitakse glükoos). Raja iga reaktsiooni katalüüsib
vastav ensüüm. Metaboolse raja ensüümid on tihti organiseerunud
multiensüümsüsteemideks nii, et ühe reaktsiooni produkt on
järgmise reaktsiooni substraat. Metaboolse raja astmelisus
võimaldab:
- Kataboliseerutava biomolekuli energia järk-järgulist ja kontrollitud konversiooni maksimaalseks arvuks ATP molekulideks
- Sobiva energiakoguse ajastatud kasutamist (suurt energiakogust pole võimalik korraga inkorporeerida!) biomolekulide sünteesiks anabolismi käigus
- Lammutamise ja biosünteesi peenregulatsioon (reguleerida saab igat astet!)
Põhirajad
(krebsi tsükkel jt) on erinevates organismides praktiliselt
ühesugused. Spetsrajad täidavad spetsfunktsioone (nt karbamiidi
süntees maksas kahjutustab ammoniaagi).
Metabolismi
integeeritus
Metabilism on
biomolekulide peenreguleeritud lammutamine ja süntees tagamaks
inimkeha elutegevuseks vajalikke sisetingimusi (homeostaasi).
Metabolismi integratsiooni iseloomustavad: Radadevahelised
sõlmpunktid, üleminekud (metabolism on võrgustik).
Võrksus
lubab alternatiivseid kulgemisvõimalusi ( metaboliit võib anda sama
produkti läbides erinevaid radu)
Ülikõrge koordineeritus:
energiat andvad kataboolsed protsessid ja energiat vajavad anaboolsed
protsessid eksisteerivad vaid üheskoos, sest:
- Rakufunktsioonideks vajalikke biomolekule saadakse nii lõhustumise kui ka sünteesi abil:
- Substraatide oksüdatiivne lammutamine annab energia valkude, peptiidede jt sünteesiks.
Anabolism ja
katabolism pole teineteise lihtsad pöördprotsessid. Nimelt,
kataboolse raja suure neg. Energiamuuduga võtmereaktsioonid pole
lihtpööratavad (anabolism kasutab neis kohtades kaudseud
pöördreaktdiooni). Teisisõnu: radade pöörduvus on
võtmereaktsioonide puhul kaudune ja põõrdprotsee toimub raku
teises komparmendis (nt glükoosi täielik lammutamine toimub
mitokondrites, glükoos biosüntees aga tsütoplasmas). Katabolsete
ja anaboolsete radade peenregulatsioon on ka erinevalt
organiseeritud. Nii garanteeritakse nende reaktsioonide vajalik suunitlus ja üheaegse toimumise võimalus.
Metabolismi
regulatsioon on kiire, paindlik, efektiivne. Regulatsiooni
tervilikkus tagastajse rakkudevahelise (neurohormonaalne
regulatsioon) ja rakusisese kommunikatsiooni vahendusel (signaali
ülekande rajad , sekundaarsed ülekandjad, reaktiivsed osakesed jne)
Rakusisese
regulatsiooni üks printsiipe on teatud valkude/ensüümide
aktivatsioon ja inhibitsioon nende fosforüülimise/defosforüülimisega. Tervikregulatsiooni põhimehhanismid ja varjandid on :
- Biomolekulide ja bioelementide (ioonide) rakku tuleku kiirus ja raku kompartmenide vahelise liikumise täpne regulatsioon ioonpumpate, kanalite , gradientide
- Ensüümide lokalisatsioon koes, rakus
- Ensüümide sünteesi induktsioon ja repressioon ja ensüümide de novo sünteesi ja degradatsioon kiiruste vahel; see kõik kindlustab enüümide vajaliku kontsentratsiooni rakkudes
- Ensüümide kineetiline regulatsioon
- Metaboolsete radade võtmeensüümide allosteeriline regulatsioon: pos. Ja neg tagasisidestus, rist -regulatsioon (vt.ensüümid)
- Igal metaboolsel rajal on „päästikreaktsioonid“, mis kulgeb nii suure vaba energia muutusega, et tema toimumine viib raja käivitumisele/kulgemisele (see reaktsioon on otseselt pöördumatu ja seda katalüüsivat ensüümi reguleeritakse allosteetiliselt)
55. Lihaskoe
biokeemia: biokeemiline koostis, kontraktsiooni mehhanism
Lihaskude
moodustab







































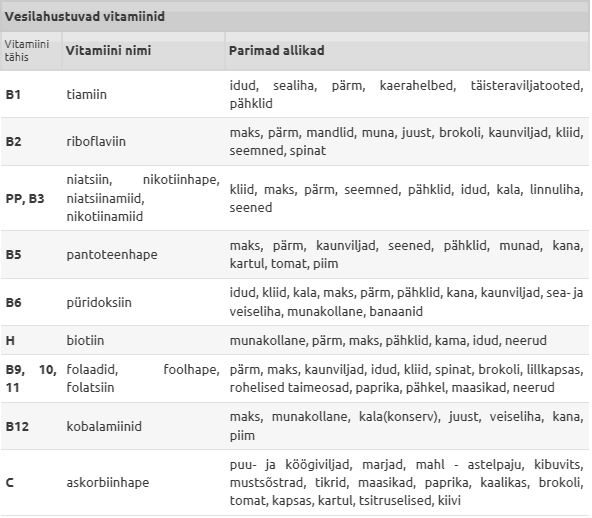




Kõik kommentaarid