Kordamine biokeemiaks. 1. Biokeemia areng ja seos teiste teadusharudega Biokeemia – teadus elava mateeria keemilisest koostisest ja biomolekulidega toimuvatest reaktsioonidest Biokeemia on väga tihedalt seotud meditsiiniga, toitumisega ja toiduainetega, metabolismiga. Meditsiinilise biokeemia baasteadmised on aluseks füsioloogiale, immunoloogiale, farmakoloogiale, farmaatsiale, endokrinoloogiale, molekulaargeneetikale, geenitehnoloogiale jt uutele spetsiifilistele arengutrendidele. 2. Keemilised ühendid ja elemendid loomorganismis Põhibioelemendid – C, H, N, O, P, S, mikroelemendid – raud, tsink, vask, mangaan, koobalt, jood jne, ja makroelemendid – kaltsium, naatrium, kaalium, magneesium, kloor. 3. Inimkeha aminohapped
1. Bioeemia areng ja seos teiste teadusharudega Esimesed sammud biokeemias tegi Scheele aastatel 1770.....1786 eraldades orgaanilisi happeid ja glütserooli. Aastatel 1770...1774 avastas Priestley hapniku- keemilise ühendi, mida loomad neelavad aga taimed toodavad. Olenevalt uurimisobjektist eristatakse biokeemias kolme erinevat suunda: staatiline, dünaamiline ja funktsionaalne biokeemia. Varasem biokeemia areng oli seotud 19. sajandi keskpaiku, kui hakkas tunnustust võitma seisukoht, et elusorganismide keemia ei ole põhimõtteliselt erinev eluta aine keemiast 20. sajandi esimesel poolel algas biokeemia kiirem areng. Võeti kasutusele kaasaegsed analüüsimeetodid, tehti kindlaks peamised ainevahetusrajad (O. Warburg, O. F. Meyerhof, H. A. Krebs, M. Calvin jpt). 1944 tõestasid Oswald Avery ja Colin MacLeod lõplikult nukleiinhapete seose geenidega. Järgnev biokeemia areng on
Biokeemia 1.Biokeemia areng ja seos teiste teadusharudega. Varasem biokeemia areng oli seotud orgaanilise keemia arenguga. Omaette uurimisvaldkonnaks hakkas ta kujunema 19. sajandi keskpaiku, kui hakkas tunnustust võitma seisukoht, et elusorganismide keemia ei ole põhimõtteliselt erinev eluta aine keemiast Meditsiinilise biokeemia baasteadmised on aluseks füsioloogiale, immunoloogiale, farmakoloogiale, farmaatsiale, endokrinoloogiale, molekulaargeneetikale, geenitehnoloogiale jt uutele spetsiifilistele arengutrendidele. 2. Keemilised elemendid ja ühendid looduses ja loomorganismis Elementaarkoostis on elava ehituse/talitluse alus. Elavast leitud üle 70 keemilise elemendi hulgas on talitlusteks vajalik miinimum 27 bioelementi, mis jaotuvad inimkehas:
31. Aine- ja energiavahetus: üldiseloomustus, põhietapid, assimilatsiooni- ja dissimilatsiooniprotsessid on katabolismi ja anabolismi integratsioon. Metabolism hõlmab seedimist, imendumist, rakus toimuvaid metaboolseid radu ja lõpp-produktide eritumist. Rakusisene metabolism toimub metaboolsete radadena, milles ensüümide toimel muunduvad/tekivad metaboliidid (biomolekulid). Metabolismi põhifunktsioonid on: · energia omastamine väliskeskkonnast toitainete vormis · toitainete omastamine ja kasutamine organismispetsiifiliste biomolekulide sünteesiks · senestsentsete biomolekulide lammutamine · lõpp-produktide väljutamine · organismi sattuvate ksenobiootikumide detoksikatsioon ja väljutamine Katabolismi staadiumid: 1. Makrotoitainete ja senestsentsete biomolekulide lõhustumine monomeerideks, ehitusüksusteks 2. Monomeeride, ehitusüksuste muundamine metabolismi võtmeühenditeks 3. Atsetüül-CoA ja Krebsi tsükli komponentide oksüdatiivne lõhustamine
1 Kordamisküsimused Biokeemia eksamiks. 1. Sissejuhatus. Bioelemendid. mis on nende olulisus ja enam-vähem funktsioonid Bioelemendid - mõiste ja jaotus: Mõiste: Bioelemendid on keemilised elemendid, mis on vajalikud elusorganismi talituseks. Jaotus: Põhibioelemendid (96-98% organismide elementaarkoostisest), Essentsiaalsed(peamised) Makroelemendid (vajatakse üle 100mg päevas nt Ca, Na, K, Mg) Essentsiaalsed Mikroelemendid Kindlapiiriliste funktsioonideta elemendid Inimkeha atomaarne koostis.
Termoregulaator (suur soojusmahtuvus ja hea soojusjuht) Organismi tasandil: Termoregulaator (higi) Transportija (veri, lümf) Hüdrostaatilise skeleti moodustaja Kaitsefunktsiooniga (pisarad, liigesvõie) Keskkond (loote areng, limakeskkond viljastumisel; laiemalt ainevahetusreakts. toimumise keskkond ja osaleja) Liiga palju vett võib olla kahjulik, kuid enamasti tekib siiski vedelikupuudus. 2. Süsivesikute/Sahhariidide biokeemia. Monosahhariidid - looduslikud süsivesikud on värvitud, veeslahustuvad, reeglina magusamaitselised kristallilised ühendid, nt glükoos, fruktoos. Glükoos (viinamrajasuhkur) on taimede ja loomade põhiline süsivesik. Ta ei ole kõige magusam suhkur. Kuulub disahhariidide koostisesse. Inimese organismis on glükoos põhiliseks energiaallikaks ja paljude teiste süsivesikute aluseks (laktoos, sahharoos, tärklis, glükogeen). Vabas olekus reguleerib ta vere osmootset rõhku.
BIOKEEMIA, II osa ORGA ANILISED AINED ORGAANILISED AINED (BIOMOLEKULID) Biomolekulid on inimkeha orgaanilised ained, millel on vähemalt üks biofunktsioon. Nad jaotuvad: ◦ lihtbiomolekulid (väikesed orgaanilised molekulid) ◦ oligomeersed biomolekulid (koosnevad väikestest ehitusüksustest nagu näiteks oligosahhariidid jt) ◦ biomakromolekulid (ehitusüksuste arv on suur nagu näiteks valgud, nukleiinhapped jt) ◦ Katabolism – ainete lammutamisprotsess, osa ainevahetuses ◦ Anabolism - ainete sünteesiprotsess VALGUD VALGUD Valgud ehk proteiinid on inimese elutegevuseks vajalikud polüpeptiidid (makromolekulaarsed orgaanilised ühendid), mis koosnevad aminohappejääkidest. Elusaine tähtsamad koostisosad, rakkude põhilised struktuursed osad, nende peamised ehitusmaterjalid. Valkude süntees toimub ribosoomides. Ööpäevas lammutub organismis u. 400 g kehavalke.
I. BIOKEEMIA AINE. RAKU EHITUS. VESI JA VESILAHUSED. (Õpik lk 3-32) 1. Bioelemendid. Bioloogilised makromolekulid. Bioelemendid: O, H, C, N, P, S. Moodustavad 99% kõikidest aatomitest inimkehas. Elemendid on molekulide tekitamiseks sobivad, sest moodustavad kovalentseid sidemeid elektronpaaride jagamisega. Biomolekulid: Valgud (ehk proteiinid, hargnemata biopolümeerid, koosnevad 20 aminohappest, moodustavad ensüümid (lipaas),retseptorid(insuliini retseptor); Nukleiinhapped (hargnemata biopolümeerid, monomeerideks nukleotiidid (dna, rna)); Süsivesikud (ehk karbohüdraadid, monomeerideks monosahhariidid, nendest tekivad polüsahhariidid mis on seotud glükosiidsidemetega; olulised energiaallikad, osalevad ka rakk-rakk äratundmisprotsessides); Lipiidid (ei moodusta polümeere!; võimelised moodustama suuri struktuure, kuid monomeerid on ühendatud nõrkade jõududega; oluline roll energiaallikana, signaalmolekulidena). Biopolümeer valgud, nukleiinhapped, süsivesikud. 2.





























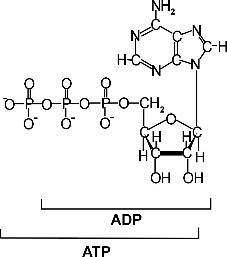




Kõik kommentaarid