Kordamisküsimused
Immunoloogia I
1.
Mis on
immuunsus ? Kirjelda organismi kaitsemehhanisme inimese näitel!
Immuunsus
on organismi võime muuta kahjutuks mitmesuguseid haigustekitajaid,
nende mürke ja kõrvaldada surnud rakke enne, kui need haigust
põhjustavad. Samuti reageerida siirdatud kudedele ja muundunud
rakkudele (kasvajatele).
Immuunsüsteem
ei ole meie arenguks hädavajalik, kuid ilma selleta jääksime ellu
vaid steriilsetes tingimustes.Kaitsemehhanismid:
Barjäärid-
nahk ja limaskestad , ensüümid, antibakteriaalsed peptiidid , konkurents Nahk:
mehhaaniline tõke ja happeline keskkond (pH 3-5), RNAsid, sebum
(sebum- triglütseriidide, vaha ja õli segu mis teeb nii naha kui
juuksed veekindlamaks)
Limaskestad:
Sealne normaalne
mikrofloora on patogeenidele kinnitumiskoha ja
toitainete konkurent. Enamus patogeene pole vôimelised
tervet limaskesta läbima. Ripsepiteel väljutab
mikroobe . Lima koosneb
põhiliselt mutsiinidest, mis on klass tugevalt glükosüleeritud
valkusid (glükoproteiine).
Patogeenid kleepuvad sinna kinni, ei saa
edasi. Lisaks on lima koostises ka antimikrobiaalsed peptiidid
(näiteks defensiinid- nahal, limaskestadel, lüsosüüm-pisarates,
süljes), võivad lõhkuda bakteri membraani, aktiveerida ensüümid,
peatada nukleiinhapete sünteesi
Madal
pH (nahal ~5.5, mao pH 2). Lima ja ripsmed. Antibakteriaalsed
peptiidid. Mikrofloora. Immuunrakud epiteelkoes
(alveolaarmakrofaagid,
nuumrakud ). Sülg ja pisarad, ensüümid.
Kehatemperatuur .
Retsptorid,
mis võimaldavad infektsioone ära tunda- Mitte
spetsiifiline (innate) TLR, LRR
- Spetsiifiline
(adaptive) TCR, BCR
Signaali
ülekande rajad Immuunsüsteemi
aktivatsiooni teedEfektormehhanismid infektsioonide hävitamiseksOtsene
destruktsioon (antibakteriaalsed peptiidid (lüsosüüm jne),
komplemendi tekitatud lüüs, fagotsütoos, hapniku ja lämmastiku
radikaalide
produktsioon , NK rakkude tekitatud lüüs),
patogeeni leviku
blokeerimine (
opsoniseerimine C3b, neutraliseerimine Ab),
patogeeni paljunemise blokeerimine (piiratud ligipääs toitainetele,
viirusega nakatunud rakkude
apoptoos )
Efektormehhanismide
regulatsiooni süsteemidNormaalseisundi
taastamine (puhkeolek): kudede parandamine, homoöstaasi taastaminePõletikKui
patogeen tungib koesse, siis algab põletik: kude tursub, läheb
soojaks ja punaseks. Seda põhjustavad veresoonte muutused, mis
tekivad immuunsüsteemi aktivatsioonil.
Veresooned muutuvad
läbilaskvateks ja niiviisi saavad immuunsüsteemi komponendid
paremini infektsioonikohta jõuda. Rollid: tuua infektsioonikoldesse
uusi immuunrakke ja efektormolekule, luua füüsiline barjäär
mikrovaskulaarse koagulatsiooni näol takistamaks infektsiooni
levikut vereringesse, soodustada kahjustatud kudedes regeneratsiooni.
Kui
põletikulised reaktsioonid toimuvad vales kohas, valel ajal või on
liiga tugevad, siis on tagajärjeks näiteks allergia ja astma .2.
Selgita,
mis on loomulik (=innate; = sünnipärane) immuunsus ja mis on
omandatud (=adaptiivne) immuunsus.
Innate
immuunsus: mitte spetsiifiline, kiire (
minutid , tunnid), mälu ei
teki. Ta on alati olemas ja kohe valmis atakeerima. Näiteks
fagotsütoos,
komplement , interferooni süntees.
Adaptiivne
immunsus: väga spetsiifiline, aeglane (päevad) ja ei ole valmis
kohe atakeerima. Adaptiivne immuunsus stimuleeritakse innate
immuunsuse poolt patogeeni sisenemisel, oma magnituudilt on ta
võimsam kui innate, kuid vajab aega aktivatsiooniks. Näiteks T- ja
B-
rakud .
Innate
immuunsuse puhul tekib ühetaoline
kaitsereaktsioon samatüübiliste
mikroobide vastu, eristamata eri liike kuid eristades gruppe
(bakterite vastu tekivad üht liiki kaitsereaktsioonid ja viiruste
vastu teist tüüpi). Adaptiivse immuunsuse puhul tekib mikroobi tüve
spetsiifiline
immuunvastus ca 5-7 päevaga. Adaptiivne immuunvastus
tekib kiiremini, kui täpselt sama
mikroob teist korda siseneb.
Omandatud
immuunsusel on kahte tüüpi vastust: Ühe reaktsioonina (
humoraalne vastus) sekreteerivad B-lümfotsüüdid antikehasid ja teise
reaktsioonina (
rakuline vastus) tapavad tsütotoksilised
T-lümfotsüüdid viirusinfekteeritud rakke.
Kuidas
kaasasündinud immuunsus
nakkuse ära tunneb?
Nii
selgrootutel kui ka selgroogsetel on Pattern Recognition Receptors
(PRR), mis
tunnevad ära patogeenidele
omaseid molekulaarseid
struktuure (Pathogen Associated Molecular
Patterns - PAMP). PAMP –
need on kindlad molekulid ja
struktuurid , mida leidub vaid
prokarüootidel. PAMP-e toodavad vaid
mikroobid ja seega
kaasasündinud immuunsüsteem on võimeline tegema vahet oma ja võõra
vahel.
3.
Joonista adaptiivses immunsuses osalevad tähtsaimad
molekulid:
Ig, TCR,
MHCI ja
MHCII ,
millistel rakkudel need ekspresseeruvad,
millistele struktuuridele seonduvad/milliseid peptiide seovad ja mis
neid iseloomustab?
alfa beeta
Ig
(BCR) ekspresseerub B- raku pinnal. B-
rakk tunneb ära kas
lineaarseid või konformatsioonilisi epitoope (valgulisi,
karbohüdraatseid ja lipiidseid).
TCR
ekspresseerub T- raku pinnal. T
rakk tunneb ära ainult lineaarseid
peptiidseid epitoope mis on seondunud MHCII või MHCI molekulidele.
TCR spetsiifilisus MHC suhtes: Kui autoloogne MHC presenteerib
viirust mille suhtes T rakk on spetsiifiline, siis toimub raku lüüs.
Kui allogeenne MHC presenteerib sama viirust või kui autoloogne MHC
presenteerib teist viirust, siis lüüsi ei toimu.
T
rakkude aktivatsiooniks on vajalik kolmikkompleks:
peptiid (9 aa) +
MHC I+TCR-Tc või peptiid (12-25 aa) + MHC II+TCR-Th.
MHCI
esineb kõikidel keha tuumsetel rakkudel, selleks et immuunrakkudele
esitleda raku sees toodetud (= endogeenseid) valke.
MHCII
esineb nn “antigeeni esitlevate rakkude” (APC- DC, MQ, B) pinnal,
selleks et immuunrakkudele esitleda rakuvälisest keskkonnast sisse
võetud (= eksogeenseid) valke.
4.
Kirjelda
primaarset ja sekundaarset lümfoidset kudet ja
mis protsessid seal toimuvad.
Nimeta lümfoidsed
organid ja nende funktsioonid. Mis sarnasused ja
erinevused on vere ja lümfiringel, mis on lümfi funktsioon?
Primaarsed
lümfoidorganid: luuüdi ja tüümus, IMMUUNRAKKUDE GENEREERIMINE.
Nendes toimub algselt lümfotsüütide teke
vereloome tüvirakkudest
ja nende funktsionaalne küpsemine,
diferentseerumine T- ja B-
lümfotsüütideks.
Sekundaarsed
lümfoidsed organid ja koed: põrn, lümfisõlmed ja lümfoidse koe
kogumikud (MALT, GALT), IMMUUNVASTUSE GENEREERIMINE.
Spetsiifilised ,
peamiselt lümfoidkoest koosnevad elundid, mis on paigutunud teiste
kudede suhtes
selliselt , et oleks tagatud maksimaalne tõenäosus
patogeenide ja lümfotsüütide kontaktiks. Sekundaarsed
lümfoidorganid on seega sageli
spetsiifiliste immuunreaktsioonide
toimumispaigaks.
Tertsiaarne lümfoidne kude: ag poolt sekundaarsetes organites aktiveeritud
immuunrakud võivad lümfisõlmedesse jääda või pöörduda
lähteorganisse (kops, maks, aju) tagasi efektor- ja mälurakkudena
ja moodustada seal organiseeritud lümfoidse koe kogumikke.
Luuüdi:
Luuüdis asuvatest vereloome tüvirakkudest (HSC) arenevad välja
kõik punased ja valged
vererakud ning trombotsüüdid. Luuüdis
valmivad ka B- rakud. Luuüdi
strooma rakud toodavad vereloomeks ja
immuunrakkude arenguks vajalikke tsütokiine: IL3, IL7, CSF.
Tüümus:
Tüümus asub südame kohal otse
rinnakuluu all. Ainus lümfoidorgan,
kus immuunvastust ei genereerita. Tüümuse epiteelirakud toodavad
hormoone: tümosiini, mis on lümfoidsete rakkude paljunemist
reguleeriv
hormoon . Tüümuses toimub T- rakkude valmimine.
Põrn:
Veres leiduvate antigeenide vastu immuunvastuse initsieerimine,
vanade ja defektsete RBC eraldamine, uute RBC ja Mo
reservuaar , Fe
varu. Põrna kude on pehme ja vererikas- punane üdi (
pulp ):
erütrotsüüdid (RBC), vere filtratsioon, valge üdi (pulp):
lümfotsüüdid, spetsiifilised immuunreaktsioonid, lümfotsüütide
aktiveerumine, proliferatsioon ja diferentsioon ning antikehade
süntees.
Lümfisõlmed:
Püüavad antigeene kudedest, vastavalt
koele tuleneb ka nende
nimetus. Aferentsoone kaudu saabub ag ja eferentsoone kaudu
lahkuvad lümfivedelik,
antikehad ja rakud.
Lümfi
funktsioon: Lümfisõlmede
filtreerimine (bakterite, viiruste ja
vähirakkude transport). Liigse vedeliku transport kudedest verre.
Toitainete, eriti lipiidide transport.
Lümfiringe
ja
vereringe võrdlus: Lümfiringes voolab lümf, vereringes
veri .
Lümfiringe on ühesuunaline (kudedest südame poole), vereringe
kahesuunaline. Lümfikapillaarid on läbilaskvamad kui
verekapillaarid .
5.
Kirjelda vereloome arengut ning joonista vastav skeem. Nimeta
tüvirakust algavad arenguteed ja kõik terminaalselt
diferentseerunud vererakud, nende kuuluvus adaptiivsesse või innate
süsteemi, ning funktsioon organismis.
Granülotsüüdid:
Neutrofiilid
(N)- 50-70% vere leukotsüütidest, N arvu tõus ja noorvormid veres
näitavad põletikku, põletikukoldes lõhkevad, vähesel määral ka
fagotsüteerivad,
hukkunud neutrofiilidest moodustub mäda.
Basofiilid
(B)- Veres alla 1%, olulised
parasiitide vastu ja allergilistes
reaktsioonides, graanulites
histamiin .
Nuumrakud
(MC)- Esinevad nahas, respiratoor- ja seedetraktis.
Graanulid sisaldavad
histamiini jt aktiivseid substantse. Nuumrakud ja vere
basofiilid indutseervad allergiaid. Nuumrakud tekivad luuüdis
vereloome käigus ja kudedes differentseeruvad limaskoe või sidekoe
nuumrakkudeks.
Eosinofiilid (E)- Veres alla 1%, esinevad luuüdis, soole limaskestades,
eskpresseerivad tsütokiine, olulised
parasiit -infektsioonide vastu.
Monotsüüdid
–tsirkuleerivad veres ca 8 tundi, suurenevad ja migreeruvad
kudedesse, kus diferentseeruvad koespetsiifilisteks makrofaagideks -
paikne lokalisatsioon, kindel funktsioon. Luuüdist verre saabunud
osad monotsüüdid migreeruvad kudedesse ja arenevad seal
makrofaagideks. M1- inflammatoorsed- sisenevad kudedesse infektsiooni
korral. M2-patrullivad- liiguvad aeglaselt mööda veresooni,
moodustavad koe-
resident monotsüütide reservuaari.
Makrofaagid
(MQ)-koristajad. M1- inflammatoorsed. M2- koe parandajad, kasvajaid
toetavad . Aktiveeritud makrofaagid on parimad fagotsüüdid. Toodavad tsütokiine.
Dendriitrakud
(DC) on parimad APC. DC esinevad erinevates kudedes: nahas,
seedeelundite sisekestas. Funktsioon: ag esitlus Th rakkudele. DC
rakud ei vaja
eelnevat aktivatsiooni ag-ni esitlemiseks, nad
ekspresseerivad MHCII pidevalt.
Follikulaarsed
dendriitrakud (FDC): FDC ei ekspresseeri oma pinnal MHC II-te, seega
pole APC –rakud ega esitle ag-ni Th-le. Lokaliseeruvad lümfisõlme
folliikulites, mis on B-rakkude poolest rikkad. FDC ekspresseerivad
kõrgel tasemel antikeha ja komplementi siduvaid retseptoreid ja
seovad vabalt tsirkuleerivaid ag-Ab komplekse. FDC aktiveerivad
B-rakke lümfisõlmedes.Follikulaarsed
dendriitrakud (FDC) ja dendriitakud (DC) on vaatamata välisele
sarnasusele täiesti erineva funktsiooniga rakud: FDC “hoiavad
antigeeni (Ab-ag kompleksidena)” oma pinnal B- rakkudele, DC
esitlevad (MHC-TCR interaktsioonis) antigeeni T rakkudele.NK
rakud-5-10% lümfotsüütidest, suured, granuleeritud. Puudub TCR ja
MHC klass I/II
restriktsioon . Reageerivad MHC I puudumisele teistel
rakkudel. Ei vaja eelnevat aktiveerimist. Sihtmärkraku lüüsimine
toimub perforiini abil. Toodavad tsütokiine: IFNgamma.
Intratsellulaarsete infektsioonide ja kasvajate vastane kaitse.
Mõningane immunoloogiline mälu.
Esmaseks ülesandeks on kiiresti
ära tunda ja hävitada kasvajarakke ja viirusega
infitseeritud rakke. Teiseks tähtsaks funktsiooniks on immuunvastuse kulu
regulatsioon tsütokiinide tootmise kaudu (IFNgamma).
6.
Mis on lümfisõlme funktsioon? Kirjelda lümfisõlmes toimuvaid
protsesse, rakkude tüüpe ja nende terminaalset differentseerumist,
GC kujunemist, klonaalset selektsiooni ja FDC osa selles.
Lümfisõlmede
funktsiooniks on lümfi filtreerimine. Lümfisõlmedes toimub
lümfogeensete ehk kudedest lümfiga saabunud patogeenide
kahjutustamine. Seal toimub immuunvastuse algatamine.
Immuunaktivatsioon toimub lümfisõlmedes, peale antigeeni
kontakteerumist.Lümfisõlme
koores ehk korteksis asuvad B- rakud, makrofaagid ja follikulaarsed
dendriitrakud, mis moodustavad rakkude kogumikke- primaarseid
folliikuleid . Immuunvastuse tekkimisel muutuvad need sekundaarseteks
folliikuliteks, mille keskel asub follikulaarsete dendriitrakkude,
aktiveeritud T- helperite ja paljunevate B- rakkude moodustatud
germinaaltsenter (GC) ehk idukeskus.
Korteksi
all
asuvas parakorteksis on B- rakke vähe, kuid leidub hulgaliselt
T- rakke (toimub T- rakkude
aktivatsioon ). Samuti leidub seal
hulgaliselt dendriitrakke, mis ekspresseerivad MHCII molekule,
esitamaks antigeeni Th- rakkudele. Aktiveeritud T- helperite ja B-
rakkude esmased
interaktsioonid toimuvad samuti parakorteksis.
Sellele järgnev B- rakkude paljunemine toimub juba korteksi ja
parakorteksi
piiril , samuti idukeskuste ümber.
Afiinsusküpsemise
ja isotüüp- ümberlülituse järel tekkinud plasmarakud liiguvad
medullasse, samuti vereringe kaudu luuüdisse.
Germinaalsetes
keskustes toimub:
1.
Afiinsuse küpsemine (SM)- Tugevam seondumine ag-le FDC pinnal tagab
B- rakule pääsu GC
keskele , rohkem toitu ja
edukama paljunemise.
2.
Isotüübi vahetus (IgM-ist erinevaks)
3.
B- plasmarakkude ja mälurakkude teke
FDC-d
seovad ja säilitavad immuunkomplekse. FDC-d on tähtsad follikulaar-
ja germinaaltsentrite struktuuri püsimises ja antigeeni esitlemisel
diferentseeruvale B-rakule.
Klonaalne
selektsioon : Antigeeni kokkupuutel seda äratundva retseptoriga
vastav rakk stimuliseeritakse, millele järgneb raku paljunemine
klonaalsel teel, st kõigil raku järgnevatel generatsioonidel on
samasugune antigeeni
retseptor , et luua antigeeni reaktiivsete
rakkude
populatsioon .
T-rakud
lümfisõlmes
T-rakud,
mis sirvivad lümfisõlme, kuid ei seondu MHC-peptiididele, väljuvad
eferentsete lümfisoonte kaudu. T-rakud, mille TCR
seondub APC raku
MHC-
peptiidi kompleksile lümfisõlmes, peatavad migreerumise ja
jäävad mõneks päevaks lümfisõlme. Siin see paljuneb ja APC-lt
saadud vihjete alusel diferentseeruvad järglasrakud erinevate
funktsioonidega efektorrakkudeks. CD8- T-rakud omandavad võime tappa
sihtmärkrakke. CD4- T-rakud võivad diferentseeruda mitut erinevat
tüüpi efektorrakkudeks, sh nendeks, mis võivad aktiveerida
makrofaage, CD8- T-rakke ja B-rakke.
B-rakud
lümfisõlmes
Lümfisõlmes
B-rakud aktiveeritakse ja diferentseeritakse kõrge afiinsusega
antikehasid sekreteerivateks plasmarakkudeks. B-raku aktiveerumine
vajab nii antigeeni seondumist B-raku BCR-retseptorile kui ka otsest
kontakti aktiveeritud Th- rakuga. Mõlemad sündmused on tagatud
lümfisõlme anatoomiaga. B-rakud erinevad T-rakkudest selle poolest,
et nende
retseptorid saavad ära tunda vaba antigeeni. B-rakk kohtub
antigeeniga tavaliselt folliiklis. Kui tema BCR seondub antigeenile,
aktiveerub B-rakk osaliselt ning „neelab alla“ ja protsessib
selle antigeeni. B-rakud on spetsialiseerunud antigeeni esitlevad
rakud, mis esitlevad protsessitud peptiid-MHC komplekse oma pinnal
Th- rakkudele. B-rakud, mis on antigeeni edukalt protsessinud,
muudavad migreerumiskäitumist ning liiguvad T-raku-rikkase
parakorteksisse, kus suureneb nende võimalus kohata aktiveeritud Th-
rakku, mis tunneb ära B-raku esitletava MHC-
antigeen kompleksi. Kui
B-rakk interakteerub edukalt Th- rakuga, jäävad nad kontakti
mitmeks tunniks, muutudes täielikult aktiveerituks ja
saades signaale, mis indutseerivad B-raku jagunemist. Mõned aktiveeritud
B-rakud diferentseeruvad otse antikeha tootvaks rakuks
(plasmarakuks), kuid osad taassisenevad folliiklisse, et moodustada
GC.
7.
Immunogeensus
v
ersus
antigeensus. Mis teeb antigeenist immunogeeni?
Adjuvandid
ja miks neid kasutatakse.
Antigeensus
– on võime spetsiifiliselt seonduda BCR-i või TCR-iga (hoolimata
kas immuunvastus järgneb).
Immunogeensus-
võime esile kutsuda spetsiifilise immuunvastuse (
humoraalse või
rakulisel tasandil).
Antigeenid -
ained, mis seonduvad antikehadega. Immunogeenid – need antigeenid,
mis tekitavad immuunvastuse.
Head
immunogeenid on reeglina makromolekulid (
valgud ja
suhkrud ).
Puudulikud immunogeenid on väikesed molekulid – hapteenid, selleks
et hapteeni immunogeensust tõsta, seotakse ta suuremale kandjale.
Immunogeensust
mõjutavad faktorid :•
Immunogeeni
poolt vaadelduna1. võõraks
tunnistamine 2. suurus 3. keemiline koostis 4. füüsikaline seis
( natiivne , denatureeritud) 5. sobivus antigeeni protsessinguks
(fagotsütoosile allumine ja MHC-le seondumine)
•
Bioloogilise
süsteemi poolt vaadatuna: geneetilised faktorid ja organismi vanus•
Antigeeni
sisestamise meetodid (kogus, viis, adjuvandid)Mis
teeb antigeenist immunogeeni?
- Antigeen peab olema organismile võõras. Selle määrab antigeeni ja organismi analoogi geneetiline erinevus. Mida kaugemal fülogeneetiliselt asetsevad 2 liiki teineteisest, seda suurem on nende vahel struktuuriline ja järelikult ka antigeenne erinevus.
- Suur molekulmass
1000-6000
Da – immunogeensus on
varieeruv >
6000 Da – tavaliselt immunogeenid
50
-100 000 Da tavaliselt parimad immunogeenid
- Antigeen peab olema keemiliselt keeruline.
peaaegu kõik valgud on immunogeensed b) karbohüdraadid (polüsahhariidid) on potentsiaalselt immunogeensed (näiteks: ABO veregrupid ) c) lipiidid pole tavaliselt immunogeenid d) nukleiinhapped on nõrgad immunogeenid
Adjuvandid
on vaktsineerimisel kasutatavad lisaained (immuunvastuse
tõhustamiseks). Adjuvante kasutatakse immuunvastuse tõstmiseks, kui
antigeen on nõrk immunogeen või antigeeni on vähe. Adjuvant : a)
pikendab antigeeni püsimist b) võimendab ko-stimulatsiooni signaale
c) indutseerib granuloomi teket d) stimuleerib põletikku ja
migratsioooni.
Adjuvant
-klassikaliselt sisaldab mineraalõli ja surmatud gr- baktereid.
Inimese puhul kasutatakse alumiiniumi ja Al(OH)3 geeli (CD4+
kostimulaator).
8.
IgG. Joonista IgG molekuli skeem ja märgista H ja L ahelad , ahelate vahelised ja sisesed disulfiidsed sidemed, “hinge” piirkond, Fab
ja Fc piirkonnad ja milline osa seondub antigeenile.
“Hinge”
regioon on antikeha molekuli kõige altim koht proteolüütiliste
ensüümide toimele.
Fab- fragment tagab algse antikeha võime seonduda spetsiifiliselt
antigeenile.
Fc-
fragment, seostudes komplemendiga ja Fc- retseptoritega
immuunrakkudel määrab antikeha bioloogilise sekundaarse efekti
tüübi, efektorfunktsiooni.
9.
Inimese Ig isotüübid, struktuur, esinemine vere seerumis . Antikeha
funktsioonid organismis.
Immuunoglobuliinid
on valgud, mida toodavad B- rakud vastuseks immunogeenile.
Variaabel regioon erineb erinevate B- rakkude poolt produtseeritud antikehade
vahel, aga sama raku poolt produtseeritud antikehades on samasugune. Konstantne regioon on identne kõikides sama isotüübi antikehades,
aga erineb erinevate isotüüpide antikehades. Konstantne regioon
determineerib mehhanismi, mis on vajalik antigeeni hävitamiseks.
Ig
H ahela konstantne osa on isotüüpidel IgM ja IgE ühe C domääni
võrra pikem.
IgM
–5-15% seerumi Ig-dest. Monomeerne IgM- membraanseotud vorm,
ekspreseeritakse esimese antikehana B-raku pinnale. Sekreteeritav IgM
–pentameer, J ( joint chain ) osaleb pentameeri polümerisatsioonis.
Erütrotsüüdid+IgM agregatsioon -agglutinatsioon. Parim
komplemendi aktivaator !
Viirus +IgM – neutraliseerib võimsalt. IgM suur molekul , vähe
liikuv, vähene kontsentratsioon veres, võib läbida epiteelrakud.
Iseloomulik
primaarsele immuunvastusele.
IgG-80%
seerumi Ig-dest. Inimesel 4 IgG alamklassi (IgG1, IgG2, IgG3, IgG4),
aa erinevused γ –ahelas. IgG1, IgG3 ja IgG4 –läbivad platsenta ja neil on tähtis roll loote arengul. IgG3- efektiivne komplemendi
aktivaator. IgG1 ja IgG2 vähem. IgG4 ei aktiveeri komplementi. IgG1
ja IgG3 seonduvad makrofaagide ja neutrofiilide Fc- retseptoritega ja
seega vahendavad opsoniseerimist. IgG4 madalam afiinsus Fc retseptori
suhtes, IgG2-l see puudub. Iseloomulik
sekundaarsele immuunvastusele. Bakteriaalsete toksiinide
neutraliseerimine ja bakterite opsoniseerimine.
IgD:
0,2%,
IgD-d leidub ainult B- lümfotsüütide pinnal, osaleb mälurakkude
väljakujunemisel.
IgE:
0,001%,
IgE on seerumis, seostub Fc retseptori abil eosinofiilidele,
nuumrakkudele ja basofiilidele. Põhjustab allergia reaktsioone ning
aktiveerib otse need rakud, mille membraanile IgE on kinnitunud kui
antigeen IgE-le seondub. Esineb monomeerina. IgE
– seotud allergiliste reaktsioonidega.
IgE tunneb ära ka helminte ja eosinofiilid omakorda selle kompleksi
(IgE FcR abil) ja tapavad helmindi. IgE ei seo komplementi.
IgA:
10-15%
seerumi Ig-dest. Põhiliselt esineb sekretsioonides
(sülg, pisarad, bronhide limas), seedetraktis.
Inimesel on kaks IgA alamklassi: (IgA1 ja IgA2). IgA võib esineda
monomeeri ja dimeerina. Sekretoorset IgA-d on palju ternespiimas.
Koos IgG-dega kaitsevad vastsündinut infektsioonide eest.
Sekretoorne IgA kaitseb efektiivselt näiteks salmonella,
lastehalvatuse, gripi ja retroviiruste eest. sIgA
väldib bakterite adhesiooni epiteelirakkudele ja hoiab neid limas,
blokeerib bakterite adhesiine ja viiruste retseptoreid.
Antikehadel
on 2 põhilist funktsiooni:
1)
seondub võõrantigeeniga
2) vahendab efektor-funktsioone, et elimineerida või neutraliseerida
antigeeni
- Neutraliseerimine – antikeha seondumisel viiruste, bakterite ja toksiinide külge Fab osa kaudu takistatakse nende seondumist oma retseptoritega ja kahjustava toime avaldamist. (IgG, IgM, IgA)
- Opsoniseerimine ja selle kaudu fagotsütoosi võimendamine makrofaagide ja neutrofiilide poolt. Antikehad nn märgistavad antigeene, rakke, kudesid seondudes neile. Fagotsüütidel on antikeha (IgG) spetsiigilised Fcγ retseptorid, millega nad antikehale seonduvad. Järgnevalt initsieeritakse signaalülekanne, ja märklaud antigeeni, raku, koe fagotsütoos. (IgM, IgG, IgA)
- Komplemendi aktivatsioon: Peamiselt IgM ja kordi kehvemini IgG aktiveerivad komplementi. Antikeha, mis on seondunud oma antigeenile aktiveerib komplementi klassikalisel teel: komplemendi esimene komponent C1q aktiveeritakse, kui läheduses on piisavalt palju Ab Fc piirkondi ja seda on IgM puhul, mis on pentameer. (IgM, IgG)
- Antikehadest sõltuv rakkude tsütotoksilisus (ADCC)- antikeha märgistab antigeeni, raku või koe: tekib Ab-ag kompleks . Fagotsüüdid ja NK rakud seonduvad antikehale oma Fcγ retseptoriga. Järgneb märklaud raku tsütotoksiline lõhustamine. Makrofaagid, monotsüüdid, neutrofiilid ja eosinofiilid omavad Fcγ retseptorit (FcγR). Peale FcγR-ga seondumist toimub nende rakkude metabolismi aktivatsioon (nende lüütiliste ensüümide kontsentratsioon lüsosoomides ja graanulites tõuseb ja ensüümid suunatakse Fc ühenduspiirkonda ning märklaudrakk hävitatakse. (ADCC puhul ei rakendata komplemendi poolt vahendatud lüüsi). NK ja eosinofiilid – graanulites perforiin, mis tapab . NK, monotsüüdid ja makrofaagid sekreteerivad TNF-i, millel ka tsütotoksiline efekt. (IgG)
10.
Immunoglobuliini
raske ahela geeni ümberkorraldumine. Millised mehhanismid genereerivad Ig variabiilust. N nukleotiidide lisamise eelis ja
puudus.
Ig
lookuse ümberkorraldamine toimub alati kindla korra järgi:
VH:
D+J
DJ+V
VL:
V+J
form
a signal joint.
Variabiilsuse
genereerimine:
- Kombinatsioonide arv- Iga funktsionaalne VH segment võib rekombineeruda iga DH ja iga JH segemendiga. Iga funktsionaalne VL segment võib rekombineeruda iga JL segmendiga. 106 on minimaalne vôimalik B- raku variantide arv. Siia lisanduvad veel: junktsionaalne variabiilsus (CDR3), trimming (nukleotiidide deletsioon ), P ja N nukleotiidide lisamine, D kolm raami. Totaalne Ig kombinatsioonide arv on 1013 .
- Junctional diversity - Kodeeriva ala ühenduskohtadesse lisatakse nukleotiide .
- H ja L ahelate kombineerumine
- Somaatilised hüpermutatsioonid- Toimuvad põrnas ja LN-des (eelnevad kolm protsessi toimuvad luuüdis). B- raku rekombineerunud DNA ahela V piirkonda tekitatakse punktmutatsioonid ( single nt mutations). Suureneb Ab ja ag vaheline afiinsus (seondumise tugevus).
Ensüüm
AID (activation induced cytidine deaminase) tekitab mutatsioone.
Eelis:
tekitab lisa mitmekesisuse CDR3 hüpervariaablis regioonis
Puudus:
võivad tekkida paardumata nukleotiidid , lugemisraam võib minna
katki.
11.
Monoklonaalsete
antikehade valmistamise põhimõte ja kasutamine teraapias. Milline
peaks olema edukas ravim ?
Monoklonaalne
antikeha
on antikeha, mis on produtseeritud ühe raku või rakuliini poolt,
mis on ühest rakust arenenud. On identsed ja omavad unikaalset
aminohappe järjestust, kasvavad identselt. Reageerivad vaid ühe
epitoobiga antigeeni molekulis.
Hiirt
immuniseeritakse antigeeniga, eraldatakse põrnarakud, fuseeritakse
müeloomi rakkudega, kasvatatakse HAT söötmel (Aminopteriin
blokeerib nukleotiidide sünteesi põhiraja ning HGPRT defekti tõttu
ei saa müeloomirakud kasutada alternatiivseid substraate ja surevad,
vanuse tõttu hukkuvad ka põrnarakud. Alles jäävad vaid
hübridoomid.), isoleeritakse üksikutest hübridoomidest pärinevad
rakukloonid, mis produtseerivad ühesuguste omadustega monokloonseid
antikehi immuniseerimisel kasutatud antigeeni vastu, paljundatakse,
säilitatakse ja kasutatakse antikehade tootmiseks koekultuuris.
Kasutatakse
astma, autoimmuunhaiguste ja kasvajate ravis.
Edukas
ravim
peab olema spetsiifiline- toimima ainult teatud rakkude grupile. Ei
tohi olla immuunogeenne (ei tohi põhjustada enda vastast
immuunvastust). Peab olema ilma kõrvaltoimeteta. Peab jõudma mõju
avaldada enne kui ta organismis lagundatakse. Ei tohi ladestuda ja
peab olema süsteemist täielikult kõrvaldatav.
12.
Tsütokiinid.
Kirjelda tütokiinide üldiseid omadusi. Pleiotroopia, redantsus.
Nimeta pro- ja anti-inflammatoorseid tsütokiine ning kirjelda nende
funktsioone organismis.
Tsütokiinid
on väikeste sekreteeritavate signaalvalkude üldnimetus, mis
vahendavad immuunvastuse käigus kommunikatsiooni rakkude vahel.
Omadused:
1.
Tsütokiinid on mitte-spetsiifilised
Spetsiifilisus
on tagatud retseptori kaudu- retseptorid on ekspresseeritud ainult
aktiveeritud rakkudel, rakkude lähikontakti kaudu- tsütokiinide
kontsentratsioon on väga lokaalne , ainult interakteerunud rakud
aktiveeruvad, tsütokiinide ebastabiilsuse kaudu.
2.
Võivad mõjuda parakriinselt, autokriinselt, juktakriinselt,
endokriiselt ja intrakriinselt
Parakriinne-
tsütokiin mõjub sekreteeriva raku vahetusläheduses. Autokriinne-
tsütokiin mõjub sekreteerivale rakule. Endokriinne- suur mõju kogu
organismile, levib igale poole. Juktakriinne- mõjub lähikontaktis.
Intrakriinne- mõjub raku sees.
3.
Pleiotroopia ja redantsus
Pleiotroopia-
Üks tsütokiin mõjutab mitut erinevat bioloogilist funktsiooni
(IL-4
B-raku aktivatsioon ja proliferatsioon, nuumraku proliferatsioon).
Redantsus-
tsütokiinide funktsioon kattub (IL-2,
IL-4 ja IL-5 indutseerivad B- rakkude proliferatsiooni.)
4.
« In vivo » toimivad tsütokiinid alati koostöös teiste
tsütokiinide, hormoonide ja neurotrofiinidega
5.
Mõjuvad madalates kontsentratsioonis 10-8
kuni 10-10
M-1.
Juba üksikute molekulide sektretsioonil võib olla suur efekt.
Funktsioonid:
1.
Reguleerivad immuunvastuse algatamist, intensiivsust ja kestvust
2.
Vahendavad märklaud-raku aktivatsiooni, proliferatsiooni ja
diferentseerumist
3.
Vahendavad antikehade või tsütokiinide sekretsiooni ja tsütokiinide
retseptorite ekspressiooni
Pro-inflammatorsed:
IL1, IL6, TNFα, IFNγ
IL1
(Mo, MQ) põhjustab palaviku ja valu teket, stimuleerib CRP sünteesi
ja Th2 proliferatsiooni, mobiliseerib luuüdist leukotsüüte,
stimuleerib T- ja B-rakkude proliferatsiooni, indutseerib IL-6
sünteesi.
IL-6
(T, Mo) aktiveerib B- rakke. IL6 on tuntud kui lihasrakkude poolt
toodetud tsütokiin, kuna ta tase tõuseb lihaste treeninguga.
TNFα
(MQ, Mc) aktiveerib endoteeli rakke, indutseerib ka palavikku, valu.
TNFα suur kogus põhjustab šokki. Aktiveerib neutrofiile,
makrofaage ja dendriitrakke. Stimuleerib endoteelirakke ja makrofaage tootma kemokiine.
IFNγ
(T) keskne makrofaage aktiveeriv tsütokiin, osaleb nii loomuliku kui
omandatud immuunvastuse reguleerimises, stimuleerib MHCI ja II klassi
molekulide ekspressiooni APC-del, soodustab fagotsütoosi
võimendavate IgG- alaklasside tootmist, aktiveerib neutrofiile.
Anti-inflammatoorsed: IL10 , transforming growth factor (TGFβ), IL1 retseptori antagonist
(IL1Ra)
IL10-
enamasti immunosupressiivne (pidurdab teiste rakkude funktsioone),
inhibeerib pro- inflammatoorseid tsütokiine (TNFa, IL1, IL6, IL12 ,
IFNγ), inhibeerib MHCII ekspressiooni.
TGFβ-
fibroblastide kasvufaktor, armkoe tekitaja, inhibeerib B- rakkude
proliferatsiooni, inhibeerib aktiveeritud makrofaage, blokeerib
rakutsükli G1 faasi.
(IL-1Ra)
ei lase intensiivsel põletikulisel vastusel lisakahju tekitada,
inhibeerides pro-inflammatoorsete tsütokiinide produktsiooni ning
põletikku soodustavaid signaale.
Perekonnad:
interleukiin 1, hematopoietiinid, interferoonid , TNF-id, interleukiin
17, kemokiinid.
13.
Kemokiinid,
retseptorid, üldine iseloomustus, funktsioon ja nende
klassifikatsiooni põhimõte.
Neid
toodavad peamiselt põletiku käigus aktiveeritud leukotsüüdid ja
kudedes olevad rakud (endoteelirakud, fibroblastid , epiteelirakud).
Peamine
roll on käituda kui kemoatraktant, et juhtida rakkude migratsiooni .
Liigutavad rakke ühest kohast teise, kemokiini kontsentratsiooni gradiendi suunas. Reguleerivad rakkude liikumist, ka pikkuse kasvu, haavade paranemist, kasvajate metastaseerumist.
- Immuunrakkude põletikukoldesse meelitamine - see protsess algab leukotsüütide kinnitumisega endoteelile, mille järel nad liiguvad kemokiini gradiendi kasvu suunas põletikukoldesse.
- Immuunrakkude liikumise regulatsioon läbi immuunorganite ja kudede- kemokiinide kaudu suunatakse diferentseeruvaid rakke ühest organist teise või ühest organipiirkonnast teise.
IL8
(Mo, MQ) mobiliseerib N, on põletikku indutseeriv faktor ja osaleb
veresoonte tekkes .
Kemokiine
klassifitseeritakse struktuuri järgi- selle aluseks on
tsüsteiinijääkide paigutus molekuli teatud konserveerunud alas .
C-
kemokiinid
CC-
kemokiinid
CXC-
kemokiinid (eraldatud ühe aminohappe poolt)
CX3C-
kemokiinid (eraldatud kolme aminohappe poolt)
Kemokiini
retseptorid on seitse korda membraani läbivad G- valk seotud
retseptorid, klassifitseeritakse vastavalt sellele, milliseid
kemokiine seovad (CCR, CXCR).
14.
Fagotsütoos ja selle tähtsus. Patogeeni lõhustamine fagotsüüdi
sees. Erinevates kudedes leiduvad fagotsüüdid.
Fagotsütoos-
partikulaarsete substantside ja organismide õgimine rakkude poolt.
Fagotsütoosil on kaitsefunktsioon, kui õgitakse mikroorganisme või
muid organismile kahjulikke partikleid, ja puhastusfunktsioon, kui
eemaldatakse surnuid või surevaid keharakke ja nende osiseid.
Fagotsüteeritud materjal enamasti lagundatakse fagotsüüdi sees.
Fagotsüüdid
ekspresseerivad oma pinnal erinevaid retsetoreid, mis tunnevad ära
spetsiiflisi molekulaarseid komponente mikroobide pinnal. Nii
selgrootutel kui ka selgroogsetel on Pattern Recognition Receptors
(PRR), mis tunnevad ära patogeenidele omaseid molekulaarseid
struktuure (Pathogen Associated Molecular Patterns - PAMP).
Fagotsüüdid:
neutrofiilid, monotsüüdid
makrofaagid. Makrofaagid on parimad fagotsüüdid: mikrogliiarakud
ajus, Kuppferi rakud maksas , osteoklastid luus , histiotsüüdid sidekoes , Langerhansi rakud nahas, alveolaarsed ja pleura makrofaagid kopsus , intraglomerulaarsed mesangiaalsed rakud neerus, makrofaagid
põrnas, luuüdis ja lümfisõlmedes.
Etapid:
Fagotsüüdi liikumine põletikukoldesse ehk kemotaksis - kemoaktraktantideks on komplemendi komponendid (C5a ja C3a), kemokiinid
Neelatava objekti ära tundmine ja seostamine rakumembraaniga, mis viib fagotsüüdi aktivatsioonile
Vahetu neelamine ja fagosoomi moodustamine tsütoplasmas- fagotsüüt sopistab oma rakumembraani neelatava objekti ümber
Lüsosoomi liitumine fagosoomiga
Bakteri hävitamine- lüütilised ensüümid: lüsosüüm, proteaasid , NADPH oksüdaasid, mille toimel tekivad reaktiivsed hapnikuosakesed (ROS). Bakterit aitab veel hävitada ka madal pH.
Laguproduktide väljutamine
Opsoniinidega
(antikehad, komplemendi aktiivsed komponendid- C3b) märgistatud
patogeen on tunduvalt paremini allaneelatav kui märgistamata
infektsioonitekitaja.
15.
Komplemendi
aktivatsiooni teed ja peamised funktsioonid organismis
Komplement
on grupp seerumivalke (30+), mis on tavaolekus inaktiivsed ja
moodustavad 5% vereseerumi globuliini fraktsiooni massist.
Komplemendi
aktiveerimisel käivitub kaskaad , mille lõpptulemuseks on patogeeni
märgistamine ja/või patogeeni membraani lõhkumine.
1. Antikehade
poolt märgistatud rakkude lüüs
2. Opsonisatsioon
–> fagotsütoos
3. Kemotaksis,
põletikuvastuse induktsioon, patogeenide hävitamine
4. Immuunkompleksidele
seondumine muudab need lahustuvaks ja kergesti lagundatavaks-
immuunkomplekside koristamine
Klassikaline
tee- Aktiveerivad immuunkompleksid, mis tekivad antikehade
seondumisel märklaudantigeenidega. Antigeenile seondunud pentameerne
IgM on parim komplemendi klassikalise tee aktivaator. Esimeses etapis seonduvad erinevad C1 alakomponendid antikeha Fc- osale- viimases
toimub antigeeni sidumise järel konformatsiooniline muutus, mis
paljastab vastava sidumiskoha C1-le. Järgnevalt liituvad erinevad C1
alakomponendid ja tekkiv C1qr2s2- kompleks lõhustab nii C4 kui ka C2
kaheks aktiivseks alaosaks a ja b. C4b ja C2a moodustavad C3-
konvestaasi, mis asub lõikama C3 molekule.
Lektiini
tee- Komplemendi aktivatsioon lektiinide kaudu sarnaneb klassikalise teega . Erinevuseks on see, et immuunkompleksi asemel seondub
märklauaga vereplasmas leiduv lektiin - mannoosi siduv valk MBL- MBL
tunneb ära bakteri pinnal olevaid suhkrujääke. MBL on võimeline
sarnaselt C1-ga aktiveerima C4 ja käivitama komplemendi kaskaadi.
Alternatiivne tee- algatajaks mikroobide membraanid , kobra mürk. Seerumis toimub
ka spontaanne C3 hüdrolüüs C3(H2O)-ks. C3(H2O) seob faktor B-d,
mis omakorda võimaldab faktor D-l selle lõigata Ba-ks ja Bb-ks. Bb
ja C3(H2O) moodustavad C3 konvertaasi C3(H2O)Bb, mis lõikab C3
C3a-ks (põletik) ja C3b-ks (seda tekib vähe, kuid on efektiivne).
C3b (opsonisatsioon) ja Bb moodustavad uue C3 konvertaasi C3bBb, mida
stabiliseerib seerumi valk properdiin.
Kõik
komplemendi kolm aktivatsiooniteed lõpevad ühesuguse lüütilise
aktivatsiooniteena. C5- konvertaasid lõikavad inaktiivse C5 kaheks
aktiivseks alaosaks C5a (põletik) ja C5b (lüüs). C5b seob endaga
C6 ja C7 ja kinnitub mikroorganismi membraanile, kinnitumise
stabiliseerib C8. Tekkinud kompleks seob endaga 10-16 C9 molekuli,
mis polümeriseerudes moodustab membraani läbiva poori . Tekkinud
kompleksi nimetatakse membraani atakeerivaks kompleksiks ehk MAC.
16.
Kirjelda
MHC I ja II molekulide strukturaalseid ja funktsionaalseid erinevusi.
Inimesel
paikneb kromosoomis 6 “HLA kompleks”. Hiirel paikneb kromosoomis
17 “H-2 kompleks”.
MHCI
esineb kõikidel keha tuumsetel rakkudel, selleks et immuunrakkudele
esitleda raku sees toodetud (= endogeenseid) valke. MHCI esitleb
peptiide Tc (CD8+) rakkudele.
MHCII
esineb nn “antigeeni esitlevate rakkude” (APC- dendriitrakud,
makrofaagid, B-rakud) pinnal selleks et immuunrakkudele esitleda
rakuvälisest keskkonnast sisse võetud (= eksogeenseid) valke. MHCII
esitleb protsessitud antigeenseid peptiide Th (CD4+) rakkudele.
MHCI:
α ahel ja β2 mikroglobuliin. MHCII: α ja β ahelad. Iga α ja β ahelal on TM regioon ja C- terminaalne tsütoplasmaatiline osa. β2m
seondub ainult α3-ga. α2, α3 ja β2 on disulfiidsed sidemed
konformatsiooni hoidmiseks. Peptiidi siduv osa on MHCI α1 ja α2,
MHCII α1 ja β1.
MHCI
peptiidi siduva vao otsad on kinnised, mis määrab ära selle, et seonduv peptiid ei saa olla pikem kui 10aa, antigeeni esitamiseks lõigatakse valkude polüpeptiidid väiksemateks peptiidideks.
Peptiidi aminohapped kinnituvad α1 ja α2 domeenides olevate
ankuraminohapete abil. Reeglina MHCI, mis on seondunud organismi enda
peptiidiga, ei aktiveeri T- rakke, sest sellised T- rakud on kas juba
tüümuses eemaldatud või muudetud tolerantsusmehhanismide kaudu
anergiliseks.
MHCII
peptiidi siduv vagu on otstest avatud, võimaldades pikemate
peptiidide seondumist, 12-25aa. T- rakud aktiveeruvad, kui neile
esitatakse organismile võõrast peptiidi MHCII pinnal.
MHCI
ja MHCII funktsioonid:
Oma
ja võõra eristamine- T- rakkude põhiline roll organismis
Ag
esitlus (ja esitlemata jätmine)
Millised
rollid on MHC molekulil just raku pinnal? Esitleda oma MHC I-te, et
näidata, et rakk on terve. Esitleda võõrast peptiidi MHC I-ga, et
näidata, et rakk on nakatunud ja Tc- rakud peavad selle lüüsima.
Esitleda oma peptiidi klass I ja II MHC-ga T- rakkude arengu käigus
tüümuses. Kontrollida tolerantsi organismi oma valkude suhtes, st
esitleda oma peptiide MHC I ja II-ga (toimub perifeerias). Esitleda
võõrast peptiidi MHC II-ga, et näidata, et organism on nakatunud
ja vajab Th –rakkude aktiveerimist ja abi.
Kuidas
saab väike kogus MHC molekule esitada suurt arvu peptiide TCR-le ära
tundmiseks?
1.
Polügeenne- igal indiviidil on palju MHC geene
2.
Polümorfne- populatsiooni lookustes on palju erinevaid alleelle
3.
Ko- ekspressioon - üks MHC molekul seob erinevaid peptiide
17.
Antigeeni
töötlemine (protsessimine) ja esitlus. Ekso - ja endogeensed
antigeenid. Professionaalsed ja mitte professionaalsed antigeeni
esitlevad rakud.
MHC
I antigeense peptiidi töötlemine ja esitlemine:
Valkude lagundamine proteosoomides. TAP on ER-i läbiv kompleks, mis
suunab peptiidid tsütsoolist ER-i luumenisse. Peptiidid on
transpordi ajal kaitstud HSP (kuumashoki) valkudega. MHCI α ahel
sünteesitakse ER-is asuva ribosoomi poolt, mis ER-i membaanil
seondub kaitsevalgu kalneksiiniga, mis aitab α ahelal leida
partneriks β2 mikroglobuliini. Kalneksiin vahetatakse kalretikuliini
vastu. Seejärel toimub MHC seondumine peptiidiga, mille käigus
tekib ka interaktsioon TAP kompleksiga, mille stabiliseerib tapasiin.
Viiakse Golgi kompleksi, kust viiakse edasi plasmamembraani. MHCI
esitab enamasti oma keha valkude peptiide va juhul kui raku sees
sünteesitakse viiruse või kasvajate valke.
Tulemus:
Võõra endogeense antigeeni esitamine, raku märgistamine
lagundamiseks.
MHC
II antigeense peptiidi töötlemine ja esitlemine:
MHCII esitab rakuväliseid peptiide, millest enamus on samuti enda
peptiidid. Infektsioonide korral esitatakse peptiide, mis on raku
välistest bakteritest, toksiinidest või parasiitidest. Olnevalt
APC-st võetakse valgud rakku sisse kas endotsütoosi või
fagotsütoosi teel. Membraanilt sisse sopistunud vesiikul koos
fagotsüteeritud materjaliga liitub endosoomidega, milles on madal
pH. Endosoomid sisaldavad proteaase ja algab valkude lagundamine
10-30 aa fragmentideks, mis kantakse transportvesiikuli abil nn
hilisesse endosoomi, kus toimub peptiidide seondumine MHCIIga. Nagu
MHCI nii ka MHCII ahelad viiakse kohe sünteesi järel ER-i. MHCII
katab kolmas valk “ invariant chain” (IC), mis kaitseb peptiidi
siduvat osa. MHCII + IC liigub Golgi kompleksi ja sealt
endolüsosoomi, kus IC laguneb. Alles jääb CLIP osa, mis
vahetatakse peptiidi vastu välja. Klass II MHC- peptiid viiakse
plasmamembraani.
Tulemus:
Võõra eksogeense antigeeni esitamine, indutseerib antikehade
tootmise, meelitab immuunrakke infektsiooni piirkonda.
Enamikel
juhtudel esitlevad klass I molekulid protsessitud endogeenset
antigeeni CD8+ Tc rakkudele ja klass II molekulid protsessitud
eksogeenset antigeeni CD4+ Th rakkudele.
Endogeenne
rada
Endogeensed
antigeenid degradeeritakse tsütosoolis proteasoomide poolt
väiksemateks peptiidideks, pannakse MHC klass I molekulidega kokku
karedapinnalises ER-s ja esitletakse membraanil CD8+ Tc rakkudele.
Eksogeenne
rada
Eksogeensed
antigeenid võetakse sisse ja degradeeritakse endotsüütilises
kompartmendis happeliste, pH-sõltuvate endosomaalsete ja
lüsosomaalsete ensüümide poolt ja need kombineeruvad MHC klass II
molekulidega CD4+ Th rakkudele esitlemiseks.
Professional
APCs specialize in presenting antigen to T cells. They are very
efficient at internalizing antigens, either by phagocytosis
(macrophages and dendritic cells) or by receptor-mediated endocytosis
(B cells), processing the antigen into peptide fragments and then
displaying those peptides, bound to a class II MHC molecule, on their
membrane.
Non-professional
antigen presenting cells include all nucleated cell types in the body . They use an MHC class I molecule coupled to beta-2
microglobulin to display endogenous peptides on the cell membrane.
18.
T rakkude areng. Positiivne ja negatiivne selektsioon tüümuses. AIRE . MHC restriktsioon. Rakkude
populatsioonid tüümuses (FACS analüüs hiire tüümusest).
T
rakk (α/β) tunneb ära ainult lineaarseid peptiidseid epitoope, mis
on seondunud MHCII või MHCI molekulidele.
TCR-ist
olenevalt esineb kahesuguseid T rakke: αβ vôi γδ. Inimese ja
hiire veres on enamus (90%) αβ Τ rakud, ülejäänud (10%) on γδ.
Môlemat tüüpi T rakud (αβ ja γδ) vôivad olla nii “helper”
kui “ killer ” funktsiooniga. Kôik TCR αβ ja γδ rakud on CD3+,
2/3 αβ T rakkudest on CD4+, 1/3 αβ T rakkudest on CD8+.
Enamus
γδ T rakke on CD4-/CD8-. 1-10% γδ on CD4+, 1/3 γδ on CD8+.
Toodavad palju tsütokiine. Esinevad
peamiselt nahas ja limaskestades. Ekspresseeruvad veres rohkem varajases ontogeneesis (kuni paar kuud peale sündi).
Tunnevad
ära mitte lipiidseid ligande (bakteriaalseid fosfolipiide) ja HSP.
Erinevalt
Ab-st TCR (va γδ) ei tunne vaba antigeeni.
TCR
variabiilsust ei suurendata somaatiliste hüpermutatsioonide abil.
Tüümuses
toimub: T rakkude areng. T rakkude astmeline küpsemine, mis on
fenotüübi ja funktsionaalsuse poolest määratletavad. TCR
ümberkorraldus. T(α/β) rakkude positiivne ja negatiivne
selektsioon.
T
raku eellane rändab luuüdist tüümusesse ja siseneb Notch /Notch
ligand interaktsiooni abil.
Positiivne
selektsioon (koores): seondumine
oma MHC-le ja oma peptiidile päästab raku (kui ei seondu, või
seondub liiga nõrgalt
apoptoos).
Negatiivne
selektsioon (säsis): liiga
tugev seondumine oma MHC-le ja peptiidile viib apoptoosi .
Positiivse
selektsiooni eesmärk on elimineerida ebaefektiivsed ja negatiivse
selektsiooni eesmärk on elimineerida autoimmuunsust põhjustavad
T-rakud.
AIRE-
transkriptsiooni faktor tüümuses, mis vastutab koespetsiifiliste
antigeenide ekspressiooni eest tüümuses. Pakib kromatiini lahti ja
vastutab keha valkude ekspressiooni eest.
Expression
of non- local proteins by AIRE in the thymus reduces the threat of
autoimmunity by promoting the elimination of auto-reactive T cells
that bind antigens not normally found in the thymus.
MHC
restriktsioon- T rakud tunnevad antigeeni ära, kui see on seondunud
MHC molekulile, Tc (CD8+) - rakud MHCI, Th (CD4+)- rakud MHCII.
Rakupopulatsioonid
tüümuses:
CD8+ ehk T- killer rakud, CD4+ ehk T- helper rakud, topeltnegatiivsed
rakud (alles tüümusesse sisenenud).
19.
T raku aktivatsioon. T
raku ko-retseptor molekulid.
T
raku aktivatsioon toimub sekundaarses lümfoidses koes naiivse T-
raku ja APC (eriti DC) interaktsiooni tagajärjel. Positiivse
tagasiside saamise korral partner -rakult järgneb T- rakkude
diferentseerumine efektor- ja mälurakkudeks. Efektorrakud
migreeruvad põletikukoldesse (kemokiini gradiendi suunas), mälurakud
migreeruvad peamiselt nahka ja limaskestadesse (paika, kust sama
antigeen võib uuesti saabuda) ja resideeruvad seal aeglaselt
paljunedes kuni paarkümmend aastat.
Naiivse
T raku aktivatsiooniks on vaja kolme signaali.
APC
rakk annab T- rakule
1.
(MHC-TCR) ja 2. (B7-CD28, CD40-CD40L) signaali,
3.
signaali (IL2- IL2R ) annab T- rakk endale ise.
Kui
blokeerida 2. signaal (CD28-B7 seondumine) siis T- raku aktivatsiooni
ei järgne. T- rakk muutub “anergseks” - mitte reageerivaks,
immuunsüsteemile kasutuks.
T-
rakud sisenevad lümfisõlmedesse spetsiaalsete endoteelirakkude
(HEV) kaudu ja moodustavad seal APC-dega kontakte. Edasine TCR ja MHC
interaktsioon viib TCR kompleksiga seotud CD3 rakusiseste domeenide
(ITAM) fosforüülimisele. CD3 on vajalik TCR membraani
ekspressiooniks ja signaaliülekandeks (peale TCR-ag-MHC seondumist).
Lck poolt fosforüülitud türosiinid on võimelised enda külge
siduma teisi kinaase, mis omakorda fosforüülivad lähedal olevaid
signaaliülekannet vahendavaid valke. Aktiveerub biokeemiline
kaskaad, mis võimaldab signaali liikumist membraani pinnal ja sealt
edasi tsütosooli ning sealt omakorda rakutuuma (NFkB). TCR-i
interaktsioon MHC-ga ei ole võimeline aktiveerima T- rakku, kui kahe
retseptori omavaheline seondumisvõime on selleks liiga nõrk ja
seondumine lühiajaline. Kui TCR ja MHC on moodustanud kontakti, siis
kokkupuute piirkonda tulevad teised sama raku TCR kompleksid ja
moodustavad kontakti MHC kompleksidega, moodustub immunoloogiline
sünaps. Aktivatsiooniks vajavad T- rakud veel teist signaali, mida
vahendavad kostimulatoorsed molekulid. Kostimulatoorset signaali T-
raku pinnal vahendab CD28 molekul. CD28 interakteerub dendriitraku
pinnal B7 molekuliga ning alles nende kahe retseptori interaktsioon
annab edasi signaali. Kui T- raku ja APC vaheline sünaps on alles
algusjärgus, siis dendriitraku pinnal olev B7 molekulide vähene arv
ei suuda täiemahulist signaali edasi anda. Selleks, et dendriitrakk
ekspresseeriks B7 kõrgel tasemel, aktiveerub sünapsi tekke
tagajärjel T- raku pinnal CD40L molekul, mis on CD40- retseptori
ligandiks dendriitraku pinnal. CD28 vahendataval signaalil on mitu
hästi läbi uuritud tagajärge: transkriptsioonifaktorite
aktivatsioon, IL-2 süntees, IL-2R retseptori aktivatsioon.
Ko-retseptorid:
CD4 ja CD8, seonduvad MHC molekulidele ja osalevad samuti signaali
ülekandes, võimendavad TCR-i poolt saadud signaali, CD4 seondub
MHCII-ga ja CD8 MHCI-ga.
20.
Nimeta
ja kirjelda T helper rakkude alatüüpe, nimeta nende poolt toodetud
tsütokiine ja kirjelda nende funktsioone organismis.
Th1
ja Th17- rakuline immuunsus. Th2 ja TFH- humoraalne immuunsus,
antikehade tootmise toetamine . Treg- inhibeerib kõigi T- rakkude
vastuse.
Th1:
IL-2 võimendab T- ja B- rakkude proliferatsiooni, aktiveerib
lümfisõlmedes ja põletikukoldes CTL-e. IFNγ võimendab
makrofaagide aktivatsiooni (suureneb fagotsütoos), stimuleerib B-
rakke sünteesima IgG1 ja IgG3 (opsoniseerimine, fagotsütoos,
komplemendi aktivatsioon).
Th2:
IL-4 IgG toodang ja IgE toodang (nuumraku aktivatsioon), IL-5
eosinofiili aktivatsioon, IL-10 pärsib proinflammatoorsete
tsütokiinide tootmist, inhibeerib MHCII ekspressiooni.
Tc-
rakud: tsütotoksilised ensüümid.
21.
B raku aktivatsiooniks vajalikud signaalid. Mis järgneb B raku
aktivatsioonile? Mille poolest erinevad B1 ja B2 rakud?
B
rakk tunneb ära kas lineaarseid või konformatsioonilisi epitoope
(valgulisi, karbohüdraatseid ja lipiidseid).
B-
raku aktivatsiooniks on vaja kolme signaali. Esiteks antigeeni
seondumine BCR-iga, teiseks CD40 ja CD40L kostimulatoorne signaal,
mis tuleneb B- raku kontaktist CD4+ Th- rakuga ning kolmandaks Th-
rakkudes toimuv tsütokiinide süntees.
Antigeeni
seondumine ja signaali edastamine eeldab B- raku pinnal mitme BCR-i
kaasamist. Ig-α
ja Ig-β edastavad B- raku aktivatsioonisignaali ITAM kaudu.
Ig-α
ja Ig-β ITAM valmistab B- rakku ette edasist T- raku abi vastu
võtma. Umbes 12 tundi peale antigeeni ära tundmist
B-
rakud suurenevad, kasvab nende transkriptsiooniline aktiivsus, nad
sisenevad rakutsüklisse. B- rakud hakkavad ekspresseerima rohkem
MHCII ja kostimulatoorseid molekule. Peamine kostimulatoorne signaal
B- raku pinnal on CD40, mis interakteerub T- raku pinnal oleva CD40L
molekuliga. Kolmas B- raku aktivatsiooni signaal tuleneb T- raku
poolt sünteesitavate tsütokiinide olemasolust, ilma Th2 poolsete
tsütokiinide olemasoluta jääb B- rakkude proliferatsioonivõime
nõrgaks (IL-4, IL-5, IL10).
Osad
aktiveerunud B- rakud diferentseeruvad plasmarakkudeks ja
mälurakkudeks, mis toodavad antikehi. Osades B- rakkudes toimuvad
somaatilised hüpermutatsioonid ja isotüübi vahetus.
Madala
afiinsusega ja autoimmuunse retseptoriga B-rakud surevad.
Somaatilise
hüpermutatsiooni (viib läbi ensüüm AID) käigus toimuvad muutused
antikehade V- regioonides, peamiselt tekivad mutatsioonid neis
geenisegmentides, mis kodeerivad CDR3 alasid ja tõstavad antikeha
afiinsust antigeeni suhtes. Selle kaudu paraneb oluliselt antikehade
afiinsus antigeeniga seondumisel.
Isotüübi vahetuse käigus vahetatakse antikeha konstantset regiooni kodeeriv
segment. Algul on kõik antikehad IgM konstantse regiooniga, isotüübi
vahetamise käigus toimub B- rakus genoomne reorganiseerumine
konstantse regiooni kodeerivate geenide piirkonnas.
Isotüübi
vahetus muudab Ab effektor funktsiooni muutmata Ab spetsiifilisust ag
suhtes.
22.
Kirjelda I ja II tüüpi ülitundlikkuse reaktsioone. Tüüp I IgE
vahendatud ülitundlikkus. Tüüp II IgG vahendatud tsütotoksiline
ülitundlikkus (näiteks reesus konflikt).
Allergia,
ülitundlikkus: Immuunsüsteemi sobimatult suur reaktsioon väikesele
ja ohutule tekitajale.
Tüüp
I: tekib väliskeskkonnast organismi sattunud antigeenide toimel juba
varem nende antigeenide suhtes sensibiliseerunud isikutel, kellel on
vastavad IgE- klassi antikehad.
1.
Süsteemne: anafülaktiline šokk- raske, generaliseerunud (mitut
elundkonda haarav ) ülitundlikkusreaktsioon. Kujuneb mõne minutiga,
vajab kohest esmaabi (adrenaliini süst- kitsendab veresooni, avab
hingamisteed, kõrgendab vererõhku, alandab sügelust).
2.
Lokaalne: allergiline nohu , astma, toiduallergia
A
type I hypersensitive reaction is mediated by IgE antibodies, whose
Fc region binds to receptors on mast cells or blood basophils.
Crosslinkage of the fixed IgE by allergen leads to mast cell or
basophil degranulation with release of pharmacologically active mediators. The principal effects of these mediators are smooth -muscle
contraction and vasodilation.
Tüüp
II: tekivad enamasti autoantikehade mõjust kudedele, harvem on need
tingitud mikroobidevastastest antikehadest. Koekahjustus tekib
seeläbi, et antikehad (IgG1, IgG3) aktiveerivad kudedes leukotsüüte,
seostudes nende pinnal olevate vastavate Fc- retseptoritega, võivad
aktiveerida ka komplemendisüsteemi klassikalise tee kaudu.
Antikehade mõju tulemusena sekreteeritakse aktiveeritud
leukotsüütidest mitmesuguseid koerakke lõhustavaid substantse
(ensüümid, ROS). Antikehade kinnitumisel rakkude membraanidele
toimub aga märklaudrakkude opsonisatsioon ja fagotsüteerimine,
otsene komplemendisüsteemi aktivatsioonist tingitud hemolüüs,
samuti ka NK-rakkude poolt FcR-i vahendusel esilekutsutud tsütolüüs.
Kõiki neid reaktsioone võib leida: alloimmuunsete hemolüütiliste
aneemiate, autoimmuunsete (vereülekandest tingitud) aneemiate,
kudede siirdamisel tekkinud kiiret tüüpi äratõukereaktsioonide ja
mõningate autoimmuunhaiguste korral (Goodpasture sündroom).
Reesus
konflikt: Ema ja loote reesus- või veregrupi erinevusest tingitud
loote haigusseisund . Reesuskonflikt tekib ema ja loote vahel, juhul
kui ema veri on reesusnegatiivne ja loote veri reesuspositiivne .
Loote verd võib sattuda raseduse ajal ema verre, kuid suurema
tõenäosusega satub see ema verre sünnituse, kirurgiliste
manipulatsioonide, abordi käigus. Võõra antigeeni (võõrvalgu) sattudes ema organismi hakatakse selle vastu tootma antikehi. Toimub
niinimetatud sensibiliseerumine. Värsked antikehad on suure
molekulmassiga ja ei suuda seetõttu läbida platsentaarbarjääri
ning seetõttu jääb esimesel korral loode puutumatuks. Järgneva
reesuspositiivse loote puhul on aga antikehad sellise molekulmassiga,
et läbivad platsenta ja satuvad loote organismi. Antikehad hakkavad
loote punaliblesid kui võõrvalke lammutama. Punalibled , mis
normaalselt transpordivad hapnikku, lammutatakse ja loote koed
kannatavad hapnikupuuduse all, samuti vabaneb lagunevatest
punalibledest bioaktiivne aine bilirubiin , mille kõrged
kontsentratsioonid kahjustavad loote kesknärvisüsteemi.
23.
Kirjelda
III ja IV tüüpi ülitundlikkuse reaktsioone. Tüüp III:
immunkomplekside poolt tekitatud ülitundlikkus (ja koe kahjustused).
Tüüp IV: rakkude (T & MQ) vahendatud ülitundlikkus (TBC, leepra , Ni -allergia)
Tüüp
III: antigeen- antikeha- kompleksidest vahendatud koekahjustus.
Tekivad suured mittelahustuvad immuunkompleksid, mis peetuvad kudede
piirkondades, kus nende tsirkulatsioon on takistatud. Seeläbi
aktiveerub komplemendisüsteem ja protsessi lisanduvad Fc-
retseptorite kaudu mitut liiki rakud. Koekahjustus võib tekkida
autoantigeenidest, bakteriaalsetest antigeenidest ja
viirusantigeenidest.
SLE-
immuunkompleksid ladestuvad naha, neeru, liigeste kapillaaridesse
ning põhjustavad põletikulise reaktsiooni.
Tüüp
IV: esinevad autoimmuunhaiguste kui mikrobiaalsete haiguste puhul,
kuid on olulised ka transplantaadi hilistes äratõukereaktsioonides.
Autoantigeenide või mikrobiaalsete antigeenide suhtes
sensibiliseerunud CD4+ T-rakud produtseerivad kudedes tsütokiine,
mille tulemusena aktiveeruvad makrofaagid, kujunevad erinevad
T-rakkude populatsioonid (Th1, Th17), kahjustuse piirkonda migreerub
erinevate omadustega leukotsüüte. Kahjustuse otseseks põhjustajaks
on neutrofiilid ja makrofaagid.
TBC
( tuberkuloos )- kudesid lagundav põletik kopsus.
Leepra-
nahka ja närvisüsteemi kahjustav nakkushaigus , lööve ja
haavandid, sõrmede kahjustused ja amputatsioonid.
Ni
allergia- põhjustab dermatiiti.
24.
Autoimmuunhaiguste klassifikatsioon , tekke põhjused ja ravi.
„Oma“
vastased (autoreaktiivsed) B- ja T-rakud tekivad ka täiesti tervetel
inimestel. Nende eest kaitseb meid immunoloogiline tolerants .
Autoimmuunsuse põhjustavad need autoreaktiivsed rakud, mida
tolerants ei ole elimineerinud. Immunoloogiline
tolerants-lümfotsüütide poolne antigeeni ignoreerimine.
Aegunud klassifikatsiooni järgi jagati autoimmuunhaigused süsteemseteks ja
organspetsiifilisteks või paikseteks (sõltuvalt iga haiguse
peamistest kliinilis -patoloogilistest tunnustest). Lokaalsed võivad
olla näiteks endokrinoloogilised (I tüüpi suhkurtõbi, Hashimoto
türeoidiit), dermatoloogilised (harilik vill-lööve) või
hematoloogilised (autoimmuunne hemolüütiline aneemia ). Süsteemsed
autoimmuunhaigused hõlmavad SLE-d, reumatoid artriiti. Need haigused
on seotud autoantikehadega antigeenide vastu, mis ei ole
koespetsiifilised.
Autoantikehad
oma koe struktuuride vastu
Autoantikehad
põhjustavad ebanormaalse füsioloogilise vastuse: retseptori aktiveerimine (agonism). Graves ’i tõbi.
Autoantikehad
põhjustavad ebanormaalse füsioloogilise vastuse: retseptori
blokeerimine (antagonism). Müasteenia.
Tsöliaakia
Autoimmuunne
hemolüütiline aneemia
Hashimoto
türeoidiit
Immunkomplekside
poolt pôhjustatud kahjustused
koekahjustus
rakkude surm
uus ag vabaneb
Immuunkompleksid
ladestuvad veresoontesse, kopsudesse, neerudesse. Neutrofiilide
kemotaksis, hapniku radikaalide vabanemine .
Luupus:
võimalikud on liigeste, naha, neerude, südame, kopsude, veresoonte
ja aju turse ning kahjustus, “libliklööve” ninal ja põskedel,
lööved kehal ka teistes kohtades, liigesed valulikud ja turses,
tundlikkus päikese suhtes. Immuunsüsteemile esitletakse surnud
rakkude tuuma komponentide (DNA, histoonid jne) vastaseid
antikehasid. Tekkinud immuunkompleksid ladestuvad kudedesse ja
põhjustavad põletikku ning rakusurma. Sealt vabaneb juba uut
rakutuuma materjali. SLE puhul võivad olla puudulikud apoptoos,
fagotsütoos, DNAaas, komplement. Raviks kasutatakse mittesteroidseid
põletikuvastaseid ja malaaria vastaseid ravimeid.
Rakuline patoloogia
(valdavalt T rakkude poolt põhjustatud autoimmuunhaigused)
Reumatoidartriit
(RA): põletik algab liigesekoest ja seejärel haarab kogu liigest .
Tüüp
I diabeet: rakuline patoloogia-praktiliselt ainuke AI, mis on meestel
veidi sagedasem . Insuliinist sõltuv diabeet ( juveniilne )- krooniline
inflammatoorne insuliini tootvate ß-rakkude hävitamine pankreases
(CD8+T). Ravi: Igapäevased süstid/ dieet .
Tekkepõhjused:
- Kaitstud (segregeeritud) antigeenide vabanemine (kõhre, südame ja närvikoe antigeenid). Näiteks autoimmuunne müokardiit tekib peale südamekahjustusi.
- Molekulaarne mimikri (reumapalavik peale streptokoki infektsiooni).
- B- ja T- rakkude polüklonaalne aktivatsioon (superantigeenid).
- Vigane MHCII esitlus mitte APC poolt. Ravimid ( penitsilliin – seondub erütrotsüütidele- põhjustab hemolüütilist aneemiat).
AI
soodustavad: Soolised iseärasused (AI haigused valdavalt naistel
kuna östrogeen on imuunrakkudele stimuleeriv), vanuselised
iseärasused (puudulik tolerants, Treg), keskkonnast tingitud
faktorid (neurogeenne stress , toit, mikrofloora), geneetilised
(AIRE).
Ravi:
Eesmärk-
taastatakse immuuntolerants.
Manustatakse
puuduvat hormooni (türoksiini, insuliini)
Kortikosteroidid
(Tänapäeva
meditsiinis kasutatakse sünteetilisi kortikosteroide, kuna nad
seonduvad retseptoritele tugevamini. Mõju: Peatab allergilised
reaktsioonid. Tugevalt anti-inflammatoorne kuna: Inhibeerib rakkude
jagunemise vôi DNA sünteesi, pikendab haava paranemise aega, peatab
lastel kasvu.)
Immuunostaatikumid
Anti
TNFα- RA
Spetsiaalsed Treg
TCR,
BCR blokeerimine (ravi efekt on eri haigete puhul varieeruv)
Plasmaferees
Aspiriin ja analoogid- RA: põletiku pidurdamine
Glükokortikosteroidid
– infektsioonide ja kasvajate tekke oht
Multiple
sclerosis (MS)
Patoloogia:
neuroneid kaitsva müeliini kihi kahjustus. Müeliini kihti
lagundavad organismi enda immuunrakud. Neurodegeneratiivne protsess
on kroonilise põletiku tagajärg.
25.
Nimeta infektsioonidega ( viirused , bakterid , parasiidid , seenhaigused ) võitlemise peamised innate ja adaptiivsed meetodid
inimese näitel ja kirjelda neid.
Mikroobide
vastase immuunsuse üldine printsiip 1: Infektsioonide vastane kaitse
on vahendatud varajase kaasasündinud immuunvastuse ja hilise
omandatud immuunvastuse kombinatsioonist. Varajane innate immuunsus
teeb kahjutuks enamuse sissetungijatest ja kontrollib infektsiooni
seni kuni adaptiivne vastus aktiveeritakse. Osa mikroobe on innate
immuunsusele resistentsed (või paljunevad liiga kiiresti)- selliste
puhul aktiveeritakse adaptiivne immuunsus. Mikroobi vastane
spetsiifiline adaptiivne immuunvastus on alati efektiivsem, kuna B-
ja T- rakud paljunevad klonaalselt.
Printsiip
2
Erinevat
tüüpi mikroobide vastu genereeritakse erinevat tüüpi
immuunvastused.
– Viirused:
IFNI, CTL, NK, Ab
– Ekstratsellulaarsed
mikroobid: Ab, fagotsüüdid, TH17, (TH1)
– Intratsellullarsed
mikroobid: fagotsüüdid + TH1, CTL
– Parasiit- infektsioonid (helmindid): IgE, eosinofiilid, TH2
Printsiip
3
Infektsioonide
ja immuunvastuse evolutsioon - nii mikroobid kui peremehed võitlevad
oma ellujäämise eest. Infektsioonide tulemus on tasakaal peremehe
immuunvastuse ja mikroobide võime vahel seda taluda. Peremehe
immuunvastus (eriti viiruste vastane) põhjustab peremehele endale
koekahjustusi.
26.
Immuunvastus
viirushaigustele. Millised faktorid reguleerivad immuunvastust
viirushaigustele. Kirjelda
gripiviiruse näitel, mis toimub, kui esimesed partiklid sisenevad
kurgu limaskesta rakkudesse. Millised
tsütokiinid siin osalevad ja millised rakud neid toodavad?
IFN-id
takistavad viiruse paljunemist.
NK-
rakud ründavad ja hävitavad nakatunud rakke, kui nende pinnal on
vähenenud MHCI hulk.
Antikeha
tõkestab viiruse sisenemise rakku, soodustab fagotsütoosi ja
viiruse lüüsi komplemendi vahendusel.
Tc-
rakud kahjustavad märklaudraku membraane perforiinide ja seriini proteaasi - gransüümi abil, samuti aktiveerivad apoptoosiprotsesse
surmaretseptorite kaudu. Spetsiifiline immuunvastus algab
viirusantigeenide esitamisega rakkudele professionaalsete antigeeni
esitavate rakkude kaudu. Th- rakud aktiveeruvad ja sekreteerivad
tsütokiine, mis on vajalikud Tc- rakkude kasvuks ja
diferentseerumiseks. Küpsedes efektorrakkudeks, hakkavad Tc- rakud
ära tundma viirusega infitseeritud rakke.
Interferoon
alpha ja beeta (Tüüp I) IFNα, IFNß: Toodetakse viirusega
infekteeritud rakkude poolt, difundeerub naaberrakkudele ja soodustab
nendel antiviraalsete valkude tootmist (antiviral proteins -AVPs).
Peamised tüüp I IFN tootjad on pDC, epiteeli rakud ja fibroblastid.
IFNα/β
süntees peatab viiruse replikatsiooni (inhibeerivad translatsiooni,
degradeerivad RNA-d, indutseerivad apoptoosi) ja muudab naaberrakud
viirusresistentseks.
Interferoon gamma (Tüüp II) IFNγ: Toodetakse lümfotsüütide (Th1), NK ja MQ
poolt. Akiveerib neutrofiile ja makrofaage.
Interferooni
poolt indutseeritud viiruse vastased mehhanismid- restriktsiooni
faktorid (RF): inhibeerivad viiruse replikatsiooni. RF on näiteks
raku sisesed membraan -seoselised PRRs, TLRs ( toll like receptors) või
raku sisesed tsütoplasmaatilised NLRs (NOD like receptors).
Sünteesitakse
ka TNFα (MQ), soodustab apoptoosi, inhibeerib viiruse
replikatsiooni.
IL12
toodetakse MQ ja DC poolt, aktiveerib NK rakke ja Th1 rakke.
27.
Ekstra ja intratsellulaarsete bakterite vastane kaitse
Peamisteks
loomuliku immuunsuse kaitsemehhanismideks rakuväliste tekitajate
vastu on komplemendi aktivatsioon, fagotsütoos ja põletiku teke.
Gramposiitvsete bakterite seina peptidoglükaanid aktiveerivad
komplemendi alternatiivset teed. Sama toimega on gramnegatiivsete
bakterite endotoksiin- LPS. Komplemendi aktiivsed komponendid
kinnituvad bakteritele ja märgistavad (opsoniseerivad) neid
efektiivsemaks fagotsütoosiks. Komplement aktiveerib ka põletikku,
sest komplemendi aktiivsed komponendid, anafülatoksiinid (C5a, C3a)
meelitavad kohale põletikus osalevaid rakke. Ohuretseptorite kaudu
aktiveeritud fagotsüüdid neelavad ja hävitavad baktereid ja
toodavad põletikukoldes märkimisväärses koguses tsütokiine.
Antikehade kinnitumisel neutraliseeritakse bakteri toksiinide toime
või häiritakse bakterite elutegevust (nt kinnitumist). Antikehad
võimendavad ka fagotsütoosi (opsoniseeriv efekt) ja nende kaudu
aktiveerub komplement.
Esimese
kaitseliini intratsellulaarsete bakterite invasioonile moodustavad
fagotsüüdid ja NK rakud. Kuigi fagotsüüdid õgivad edukalt
paljusid haigusetekitajaid, on viimastel mitmeid võimalusi
fagotsüüdi sees ellu jääda. Rakusisene bakteri hävitamine muutub
edukaks vaid hüperaktiveeritud neutrofiilis või makrofaagis.
Sellise aktivatsiooni saavutavad fagotsüüdid tsütokiin IFNgamma
toimel. Seda toodavad infektsioonikoldes olevad NK rakud, mis
omakorda saavad stiimuli baktereid neelanud fagotsüütidelt IL-12
kaudu.
Omandatud
immuunsuse rakulistest mehhanismidest on põhilised kaks järgmist:
Th- rakkude poolne makrofaagide hüperaktivatsioon ja infitseeritud
rakkude lüüs tsütotoksiliste T-rakkude abil. Hüperaktiveerudes
hävitab fagotsüüt tavaliselt rakusisese bakteri. Kui see siiski
ebaõnnestub või bakter põgeneb raku tsütoplasmasse, siis
hävitavad nakatunud fagotsüüdi Tc- rakud ja rakuvälisesse ruumi
sattunud bakterit rünnatakse antikehade ja komplemendi abil.
28.
Kuidas kujuneb välja bakteriaalne septiline šokk? Mis on septilise
šoki kliinilised tunnused?
1.
Bacterial toxins or LPS bind to TLRs on monocytes, systemically.
2.
This results in the production of TNF, IL-1, and other pro-inflammatory cytokines. Fever .
3.
Neutrophils are recruited to the blood stream
4.
Endothelial cells respond to the TNF, IL-1, lowing blood pressure and
resulting in the expression of chemokines by the endothelial cells.
5.
Activation and egress of neutrophils from the vessels leads to an
oxidative response and more vascular leakage, lowering blood pressure even more.
6.
The kinin proteolytic cascade is induced, resulting in production of
bradykinin and increased vascular permeability.
7.
The coagulation cascade is initiated. Disseminated intravascular
clotting occludes small vessels, reducing perfusion of all major organs . Eventually, this contributes to organ failure .
8.
Other organs (especially the liver ) are acted upon by the
pro-inflammatory cytokines, and begin to produce inflammatory
molecules themselves.
9.
Myocardial function is reduced by TNF and IL-1, exacerbating the
problems with vascular leakage.
10. Death in 50-90% of cases.
Sümptomid:
• Palavik /hüpotermia
(üle 38° vôi alla 36°)
• Tahhükardia
(90 x min)
• Tahhüpnoe
• Leukotsütoos,
leukopeenia või esineb üle 10% leukotsüütide noorvorme veres
• Kahvatus
• Valu
• Paistetus
• Vasodilatsioon
(vererõhu langus)
• Organi
talitluse häire
29.
Ainuraksete ja hulkraksete parasiitide vastane kaitse. Malaaria ja
ehinokokkoosi näited.
Eukarüoodi
vastu on raske võidelda, kuna haigustekitajal on organismile sarnane
biokeemia.
Samuti
on hulkrakne parasiit tavaliselt hiiglasuur objekt, sageli tugeva
välispinnaga.
Ainuraksetel
parasiitidel on sageli keerukas elutsükkel, mille käigus muudetakse
korduvalt eluvormi, peremeest ja oma antigeenset väljanägemist.
Loomulik
immuunsus: Fagotsüüdid neelavad ainurakseid parasiite, kuid sageli
ei suuda neid hävitada. Parasiidid inhibeerivad fagolüsosoomi
tekke. Parasiidid jäävad fagotsüüdi sees ellu ja isegi
paljunevad. Hulkraksete parasiitide puhul makrofaagid aktiveeruvad ja
sekreteerivad bioaktiivseid aineid. Parasiitide kutiikula on sageli
neile resistentne.
Ainuraksete
parasiitide vastane kaitse on nii humoraalne kui rakuline. Humoraalne
- on aktuaalne kui parasiit on vabalt veres. Rakuline - siis kui
parasiit on rakus sees.
Ainuraksed
parasiidid väldivad immuunsüsteemi: muutes pinna valke (genetic switch ,
Trypanosoma),
kesta vahetades ( Plasmodium ).
Malaaria
vastane immuunvastus sarnaneb rakusiseste bakterite vastase
immuunvastusega:
Fagotsüüdid
aktiveeruvad Th1 tsütokiinide (IFNgamma) abil. (Th2 eelistusega indiviidid on malaariale vastuvõtlikumad). Antikehad toimivad parasiidi rakuvälises faasis, kuid malaaria vahetab eluvormi ja
põgeneb sellega antikehade eest.
Hulkraksete
parasiitide vastane kaitse põhineb
peamiselt IgE tüüpi antikehadel ja eosinofiilidel. Aktiveeritud
Th2- rakud stimuleerivad antikehade sekretsiooni ja immunoglobuliini
klassi lülitumist IgE tootmisele. IgE antikehad kinnituvad
parasiidile ning aktiveerivad eosinofiile ja nuumrakke. Tekib tugev
põletik, eosinofiilide graanulites olevad valgud kahjustavad
parasiidi välismembraani. Sellise Th2- tüüpi immuunreaktsiooni
tekkel on olulised tsütokiinid IL-4 ja IL-5.
Ehhinokokoosi
vastane immuunvastus: kõrgem tase -IL3, IL4, IL5, IL6, IL9, IL10,
IL13, IgE süntees – allergilised reaktsioonid.
30.
Primaarne ja sekundaarne immuunpuudulikkus
Primaarne
immuunpuudulikkus: geneetilistest
või arengulistest defektidest tulenev.
Klassifikatsioon:
Puudub/
puuduvad
• T-
või rakuline immuunsus
• B
– või antikeha vahendatud immuunsus
• T-
ja B- rakuline immuunsus
• Fagotsüütide
ja NK vahendatud mittespetsiifiline immuunsus
• Komplemendi
aktivatsioon
Esimese
4 punkti juurde lisanduvad ka tsütokiinide, kemokiinide või nende
retseptorite puudulik funktsioon.
Mida
varasemas diferentseerumise järgus on rakkude areng peetunud, seda
enam on puudu immuunrakkude alapopulatsioone ja seda raskem on
immuunsüsteemi häire.
Lümfoidne arengutee - mõjutatud on T- ja B- rakud
SCID-
IL2, IL4, IL7, IL9, IL15 retseptori γ ahela defekt
WAS-
CD43 defekt
IFN-γ
puudulikkus
X-liiteline
agammaglobulineemia- B rakkude signaaliülekande defekt
X-liiteline
hüper-IgM sündroom- defekt CD40L kodeerivas geenis
Tavaline
variabiilne IP (CVI)- vähenenud plasmarakkude hulk
Hüper-IgE
sündroom (Job sündroom)- kõrgenenud IgE tase
Selektiivne
Ig puudulikkus
Ataksia
telangiestaasia- vaskulaarne dilatatsioon
Lümfoidne
immuunpuudulikkus võib kahjustada B- rakke, T- rakke või neid
mõlemaid korraga. B- raku immuunpuudulikkuse korral on inimene nö
märklauaks bakteritele. Samas aga viiruste ja seente vastu on
immuunkaitse normaalne. Kuna T- rakkudel on immuunsüsteemis keskne
osa, siis nende puudulikkus võib kahjustada nii humoraalset kui
rakuvahendatud immuunvastust. Kõige tõsisem on kombineeritud puudulikkus ehk korraga nii T- kui B- rakkude kahjustus.
Infektsioonid puhkevad juba varases imikueas ning võivad
põhjustada
imiku surma, kui ei hakata kohe varakult aktiivselt ravima.
Müeloidne
arengutee
Neutrofiilide
hulga vähenemine- müeloid prekursori defekt
Krooniline
granulomatoosne haigus- defekt oksüdatiivses rajas
Chediak-Higashi
sündroom- mutatsioon LYST valgus
Leukotsüütide
adhesiooni puudulikkus-defekt adhesioonimolekulide β ahelas
Immuunpuudulikkus
on meestel sagedasem, sest paljud IP mutatsioonid on X-kromosoomis.
Sekundaarne
immuunpuudulikkus: mingi
välise teguri poolt põhjustatud. Toidudefitsiit, nakkushaigused, kasvajad , rasked traumad ja põletused, kiirgused, mürgid,
immuunrakkude kaotus.
AIDS:
põhjustajaks HIV. AIDSi haigetel on rakulise immuunvastuse
puudulikkus ja Th- rakkude vähenemine.
Immuunpuudulikkust
võib ravida puuduoleva valgu, geeni või raku asendamisega. Tihti
kasutatakse immunoglobuliine.
IP-le
on iseloomulik kolme haigusgrupi sagenemine: infektsioonid, kasvajad
ja autoimmuunhaigused.
31.
Selgita, mida tähendab kasvajate arengus “EEE- Elimination,
Equilibrium, Escape : eliminatsioon , tasakaal ja põgenemine”,
mis rakud ja tsütokiinid selles protsessis osalevad?
Kasvajarakud
on võimelised indutseerima peamiselt just rakulist immuunvastust.
– Tc
–tunnevad ära muteerunud valgu + MHC I
– kui
MHC I ekspressioon on maha reguleeritud – siis NK atakeerivad
kasvajat (lüüs, Ab-ADCC)
– Kasvaja vastases immuunvastuses osalevad ka MQ (Ab-ADCC, TNFalfa, reaktiivsed
O, N)
Immunoediting on
protsess, mille käigus on organism kaitstud kasvaja eest nende
immuunsüsteemi poolt.
Eliminatsioon
(kasvaja kõrvaldamine):
– faas
I – põletik, mis põhjustab NK rakkude, makrofaagide ja
dendriitrakkude liikumist kasvaja juurde, toimub IFNgamma
ekspressioon
– faas
II – IFNgamma aktiveerib NK ja T rakke ja CXCL9-11 produktsiooni,
kasvaja-antigeen viiakse DC abil lümfisõlmedesse
– faas
III –B ja T rakkude differentseerumine
– faas
IV –CD4 ja CD8 positiivsed T rakud liiguvad tagasi kasvaja juurde,
B rakud toodavad antikehi, tsütotoksilised T rakud hävitavad
järelejäänud antigeene kandvaid kasvajarakke
Equilibrium
(tasakaal):
Eliminatsiooni
faasis ellujäänud kasvajarakud on selektiivse surve all
lümfotsüütide ja IFNgamma poolt. Need kasvajarakud on tavaliselt
geneetiliselt ebastabiilsed ja muteeruvad.
Escape
(põgenemine):
Rakud,
mis muutuvad eliminatsioonile resistentseks lähevad peale tasakaalu
faasi põgenemisfaasi, kus nad hakkavad piiramatult paljunema ja
põhjustavad kasvaja tekke.
32.
Kasvaja kõrvaldamise organismi poolsed viisid ja kasvajate vastane
immuunteraapia.
a) Direct tumor cell killing can be elicited by receptor agonist activity , such as an antibody binding to a tumor cell surface
receptor and activating it, leading to apoptosis (represented by the
mitochondrion). It can also be mediated by receptor antagonist
activity, such as an antibody binding to a cell surface receptor and
blocking dimerization, kinase activation and downstream signaling,
leading to reduced proliferation and apoptosis. An antibody binding
to an enzyme can lead to neutralization, signaling abrogation and
cell death, and conjugated antibodies can be used to deliver a
payload (such as a drug, toxin, small interfering RNA or
radioisotope) to a tumor cell.
b)
Immune-mediated tumor cell killing can be carried out by the
induction of phagocytosis, complement activation, antibody-dependent
cellular cytotoxicity (ADCC), genetically modified T cells being
targeted to the tumor by single-chain variable fragment (scFv), T
cells being activated by antibody-mediated cross -presentation of
antigen to dendritic cells, and inhibition of T cell inhibitory
receptors, such as cytotoxic T lymphocyte-associated antigen 4
(CTLA4).
c)
Vascular and stromal cell ablation can be induced by vasculature
receptor antagonism or ligand trapping (not shown), stromal cell
inhibition, delivery of a toxin to stromal cells, and delivery of a
toxin to the vasculature.
Immunoteraapia
1.
Madalate kostimulatoorsete signaalide võimendamine
Antigeeni
presenteerivate rakkude aktiivsuse suurendamine
– viies
kasvajarakkudesse GM-CSF-i geeni- kasvaja tootes seda aktiveerib DC
– Kasvajarakkudesse
B7 viimine ja sellega kostimulatoorse signaali võimendamine
– kasvatades
dendriitrakke koos GM-CSF-i, TNF-alfa, IL-4 ja kasvaja antigeenide
juuresolekul
2.
Immuunrünnak kasvaja stroomaraku ja vaskulaarsüsteemi vastu
3.
Tsütokiinide kasutamine
– interferoonid
suurendavad MHC molekulide arvu kasvajarakkude pinnal
– TNF-id
põhjustavad kasvaja rakkude surma, takistavad kasvaja verevarustust
4.
Monoklonaalsete antikehade kasutamine
– kasutusel
juba mitmeid, võivad olla paljalt või konjugeeritud mingi ainega,
mis tõstab effektiivsust.
– väga
sageli vähi rakud muteeruvad ja Mab muutub tarbetuks
33.
Millised ühised immuunsüsteemi komponendid on imetajatel, lindudel, roomajatel , kaladel ? Millised ühised ja millised erinevad
immuunsüsteemi komponendid on selgroogsetel ja selgrootutel?
Millised erandeid immuunsüsteemides esineb? Mille poolest erinevad
inimese ja hiire immuunsüsteem?
Ühised
immuunsüsteemi komponendid imetajatel, lindudel, roomajatel ja
kaladel:
Imetajatel:
IgM, IgG, IgD, IgA, IgE
Lindudel: IgM, IgY, IgA
Roomajatel:
IgM,
IgD, IgY, IgA
Kaladel:
luukalad- IgM, IgD, (IgW), IgT/Z, kõhrkalad- IgM1, IgM2, IgW, IgNar ,
IgX
- TCR
- MHC I ja MHC II
- Komplement
- NK- rakud
- Antimikroobsed peptiidid
- Fagotsütoos
- GALT, tüümus, põrn, luuüdi (puudub kõhrkaladel)
Lümfisõlmed
on lindudel ja imetajatel.
Lindudel
arenevad B- rakud primaarses lümfoidorganis Bursa Fabricius, Ig
geeni konversioon B- rakkudes.
Ühised
immuunsüsteemi komponendid selgroogsetel ja selgrootutel:
- Innate immuunsus
- Fagotsütoos
- Antibakteriaalsed peptiidid
- Komplement
- Kaitsefunktsiooniga retseptorid
Erinevused:
selgrootutel puudub adaptiivne immuunsus, selgrootutel puuduvad
lümfotsüüdid, selgrootutel puuduvad antikehad.
Inimese
ja hiire immuunsüsteemi erinevused:
Mouse
Human
Hemotopoiesis in spleen
Active into adulthood
Ends before birth
Presence of BALT
Significant
Largely absent in healthy tissue
Neutrophils in periph. blood
10–25%
50–70%
Lymphocytes in periph. blood
75–90%
30–50%
Serum IgA
Mostly polymeric
Mostly monomeric
Ig classes
IgA, IgD, IgE, IgG1, IgG2a*, IgG2b, IgG3, IgM * absent in C57BL/6, /10, SJL and NOD mice , which have IgG2c
IgA1, IgA2, IgD, IgE, IgG1, IgG2, IgG3, IgG4, IgM
Ig CDR-H3 region
Shorter, less diverse
Longer, more diverse
IL-13 effect on B cells
None
Induces switch to IgE
MHC II expression on T cells
Absent
Present
Mouse
lymph nodes are relatively few and organized into low numbers of simple chains. Lymph nodes of larger species are generally more
numerous and organized into more complex chains that individually
drain proportionately smaller areas of tissue.
Significant
differences in NK cell biology between mice and man have been
identified. Mouse NK cell activity in spleen and blood tends to peak early in life (4/10 weeks of age) while in man, NK cell activity is
relatively stable throughout life. Mice have high, whereas man has
low, NK cell activity in the lung . Fc receptors are easily detected
on human, but markedly less so on mouse NK cells.




















































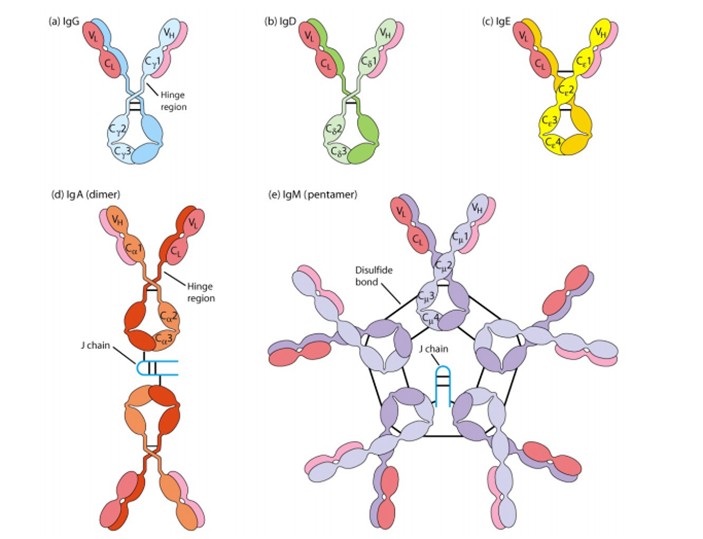
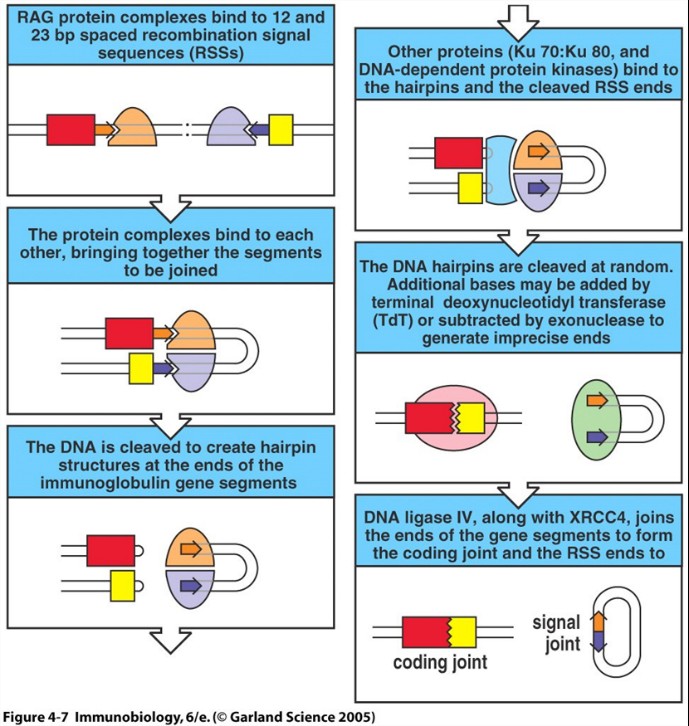
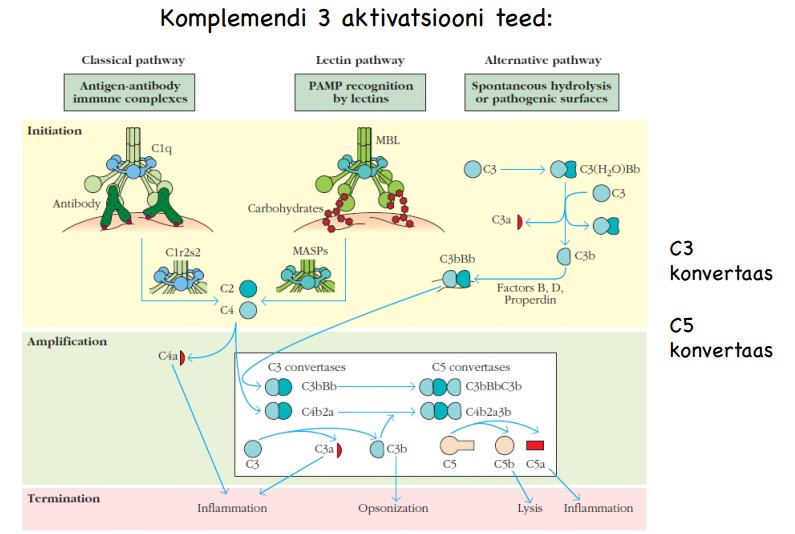

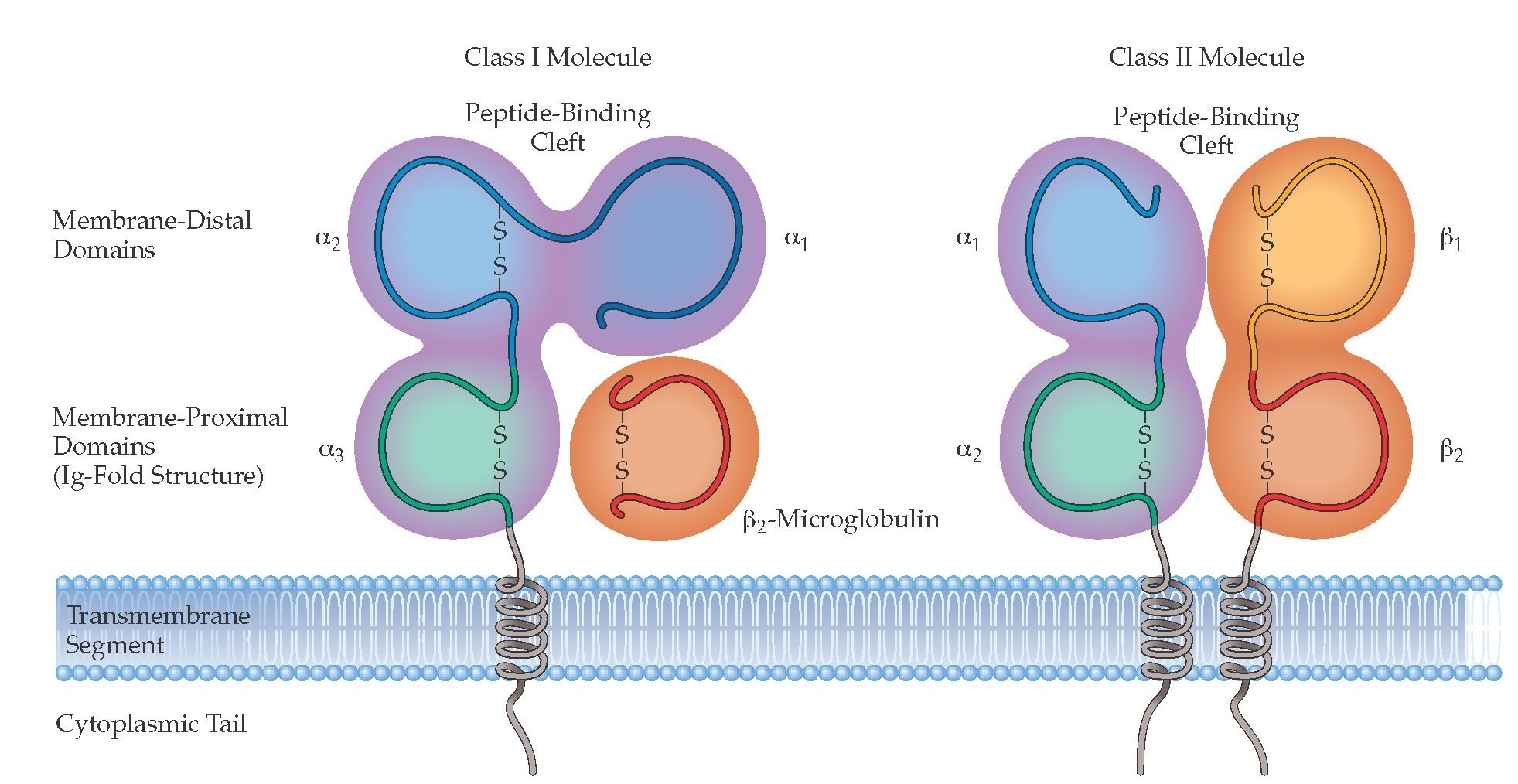
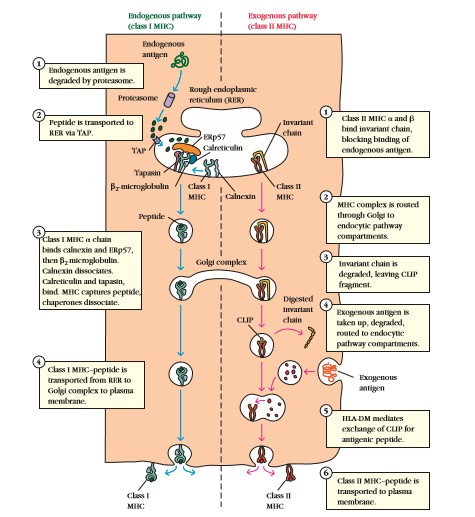
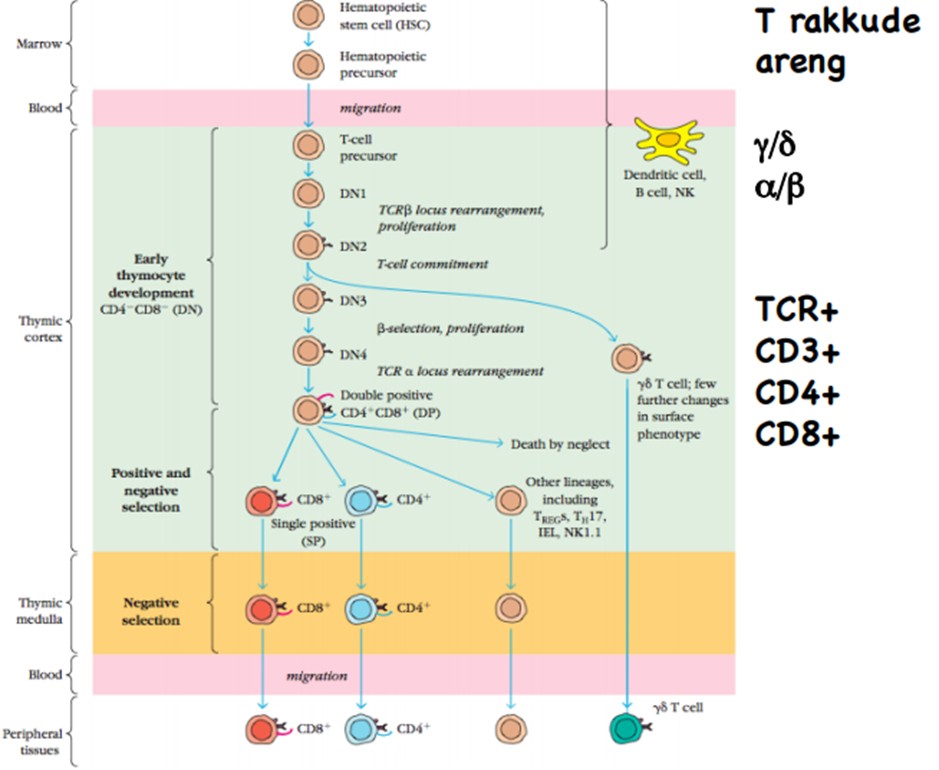
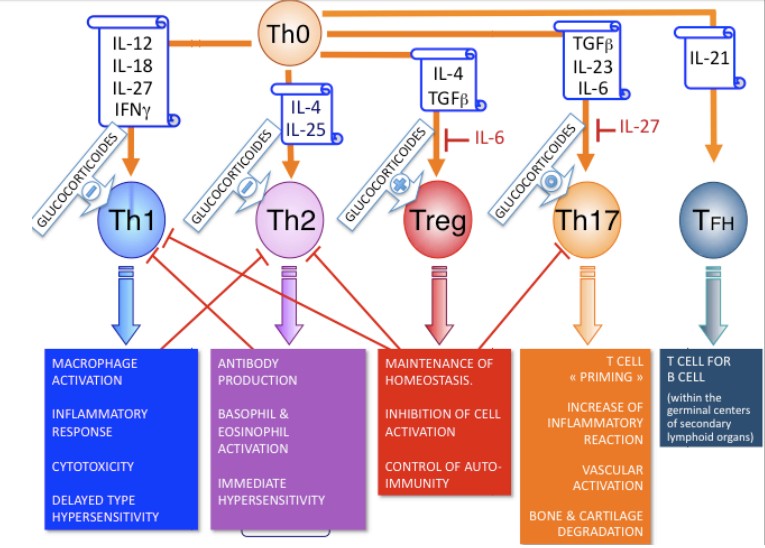
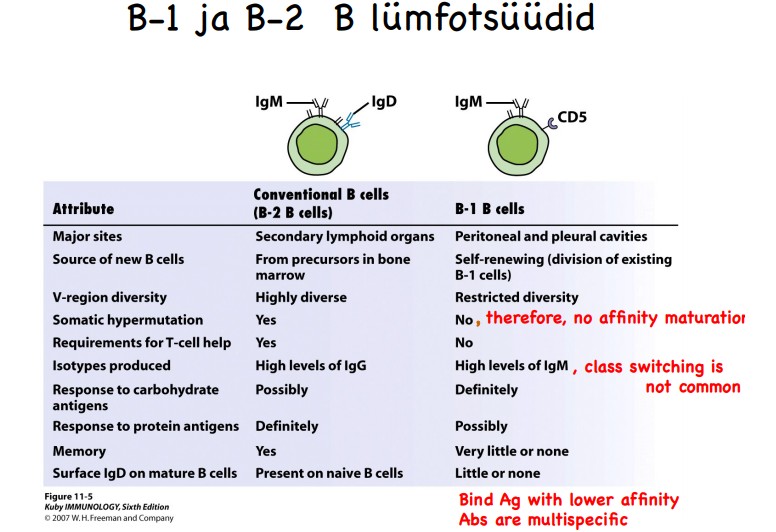
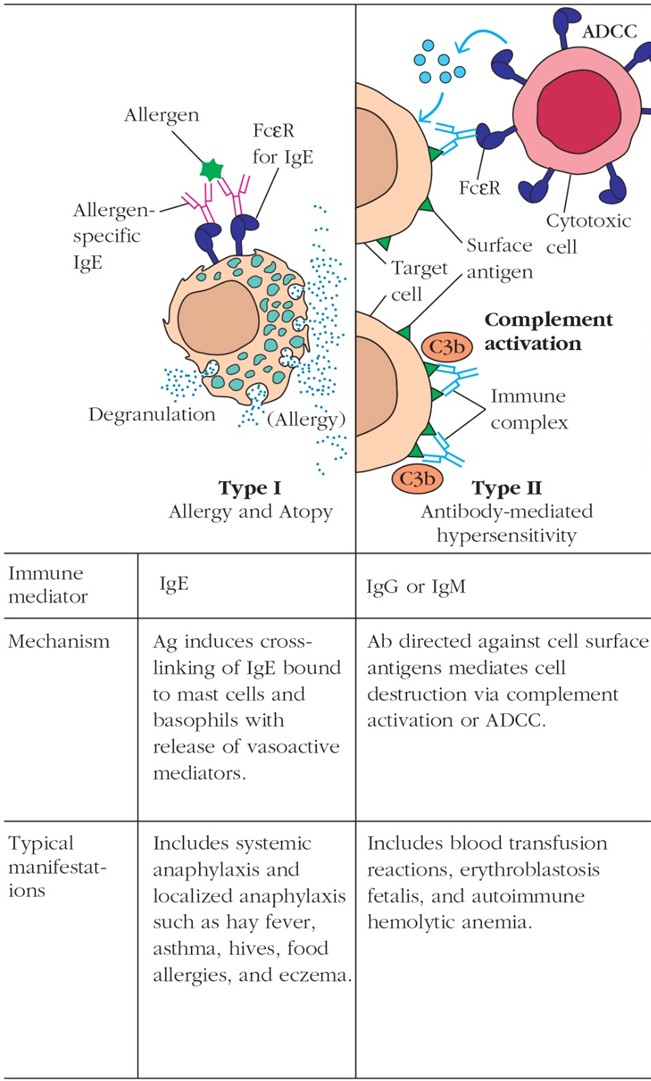
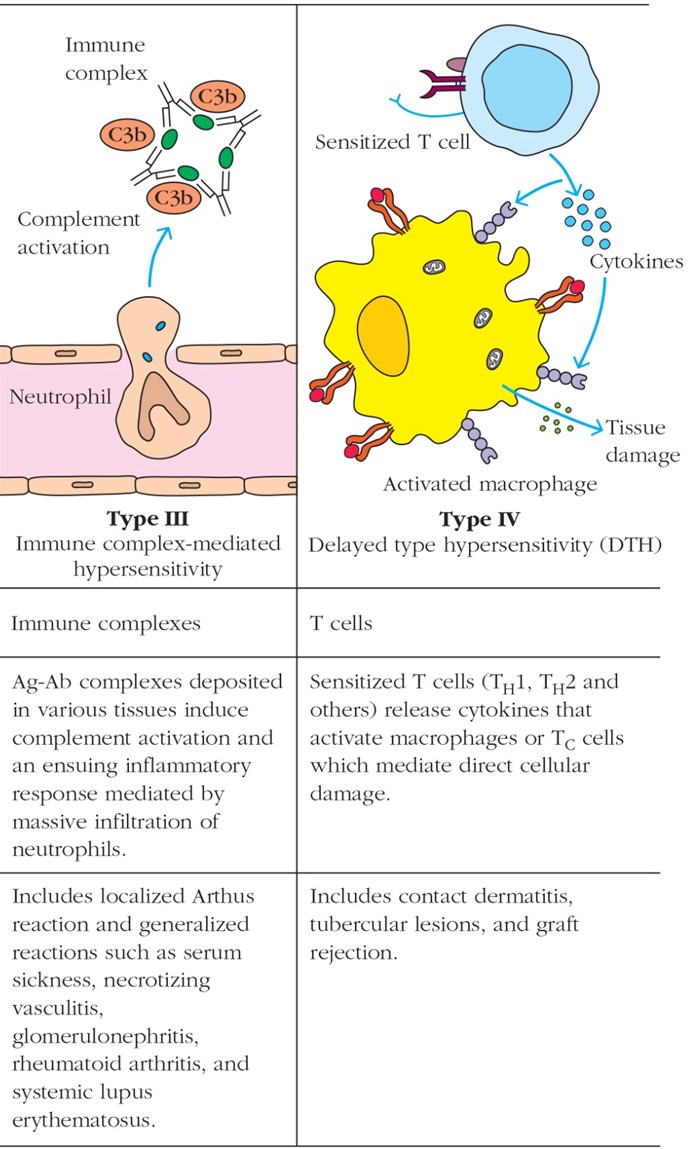
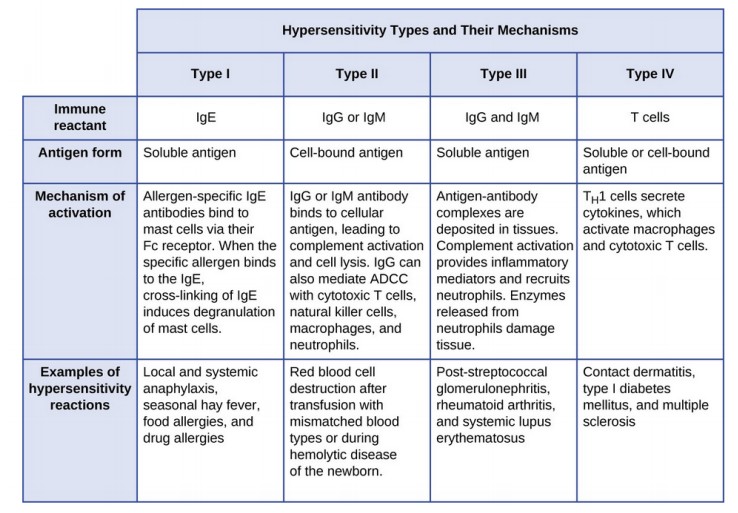


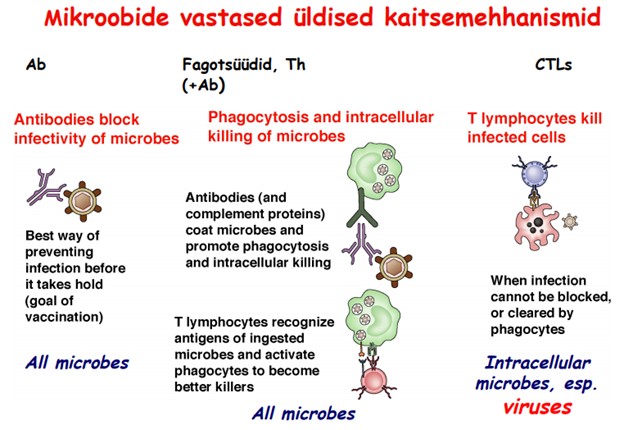
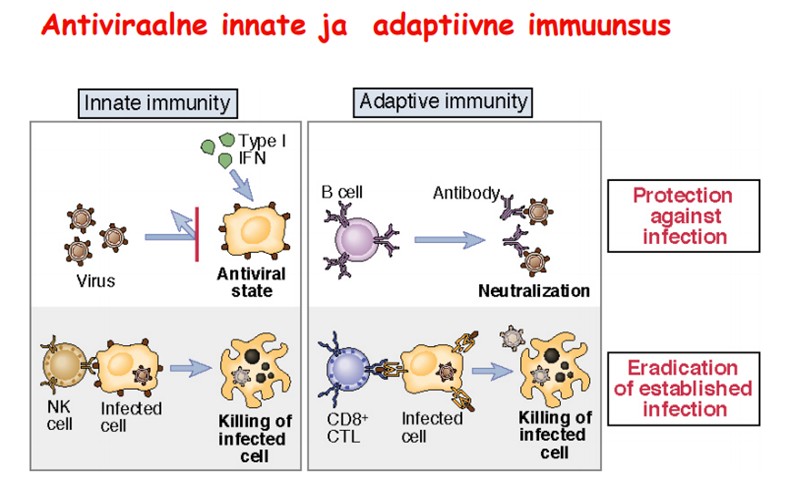
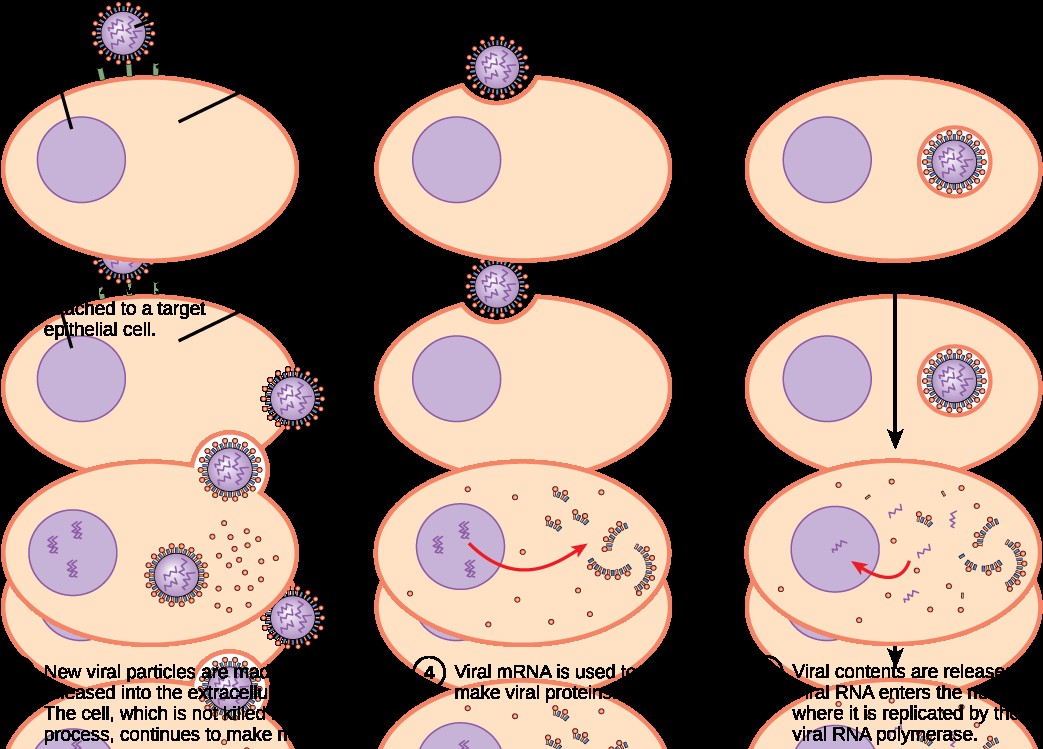
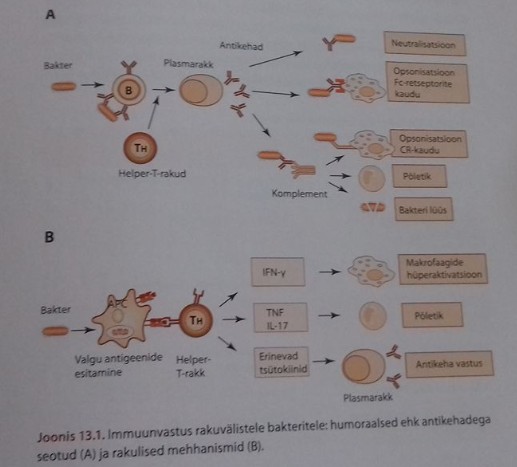
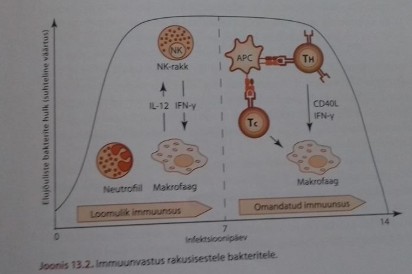

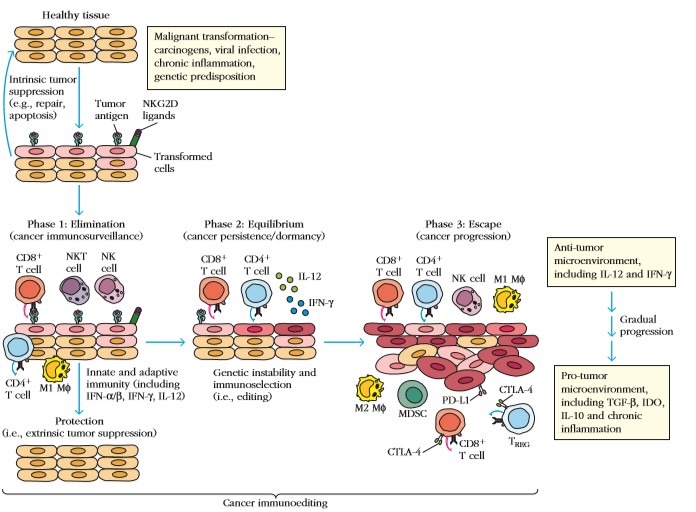
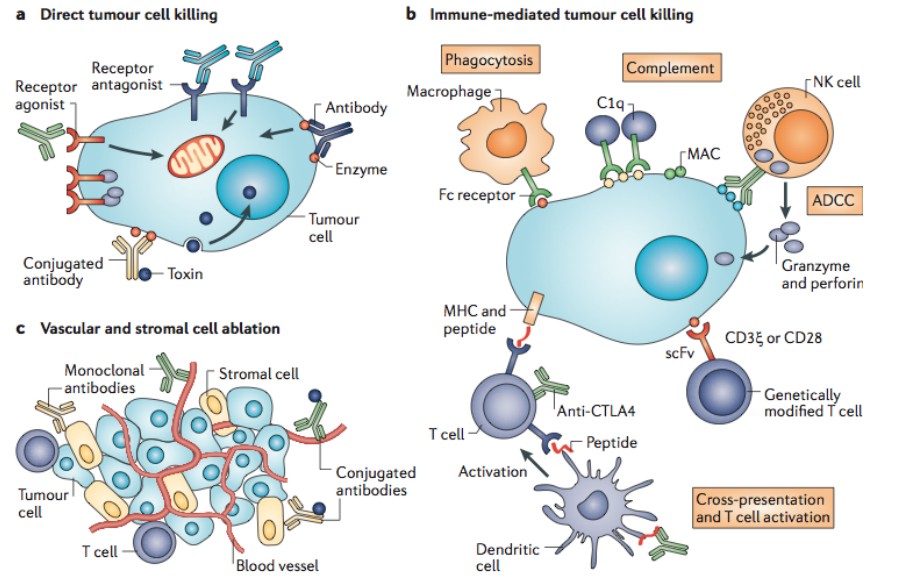
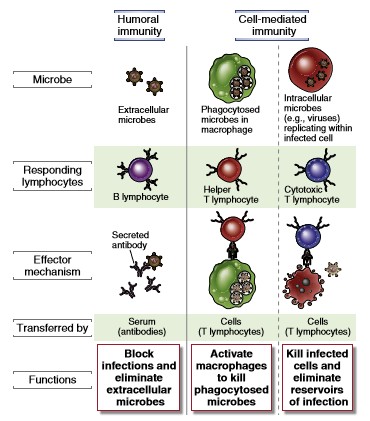
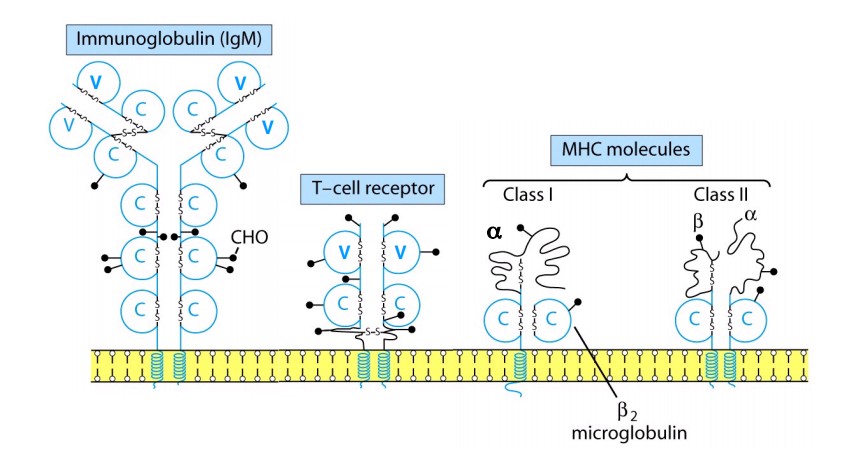
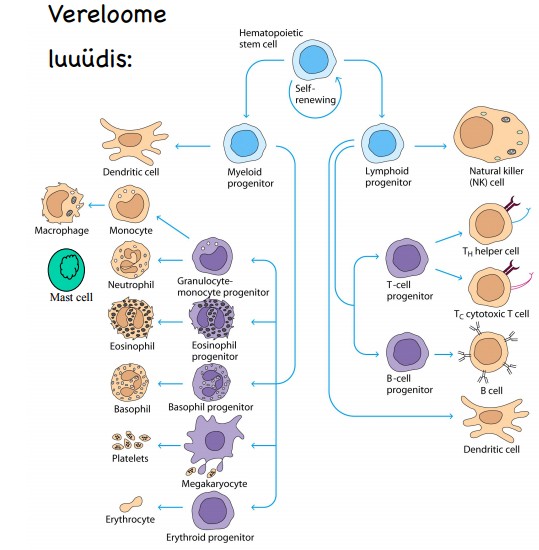
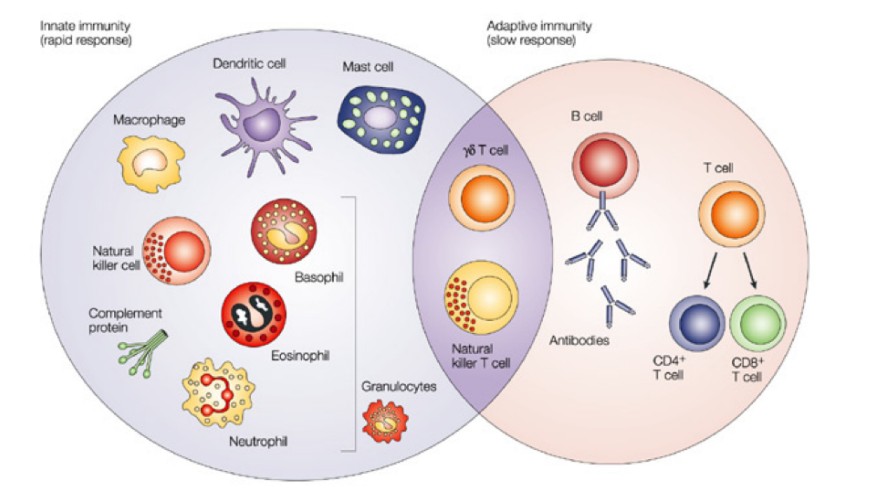
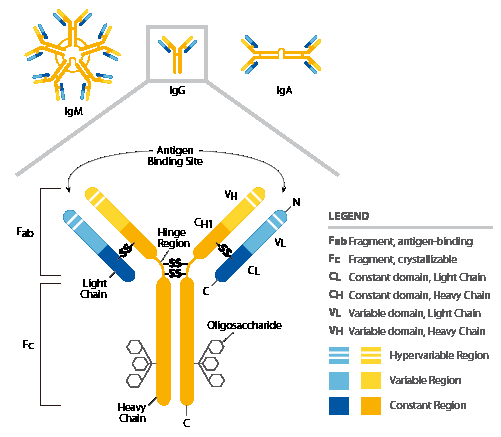

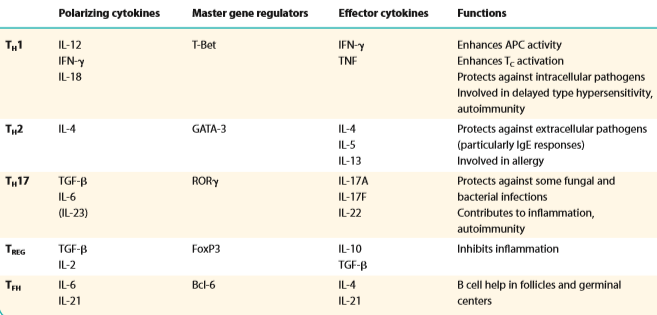
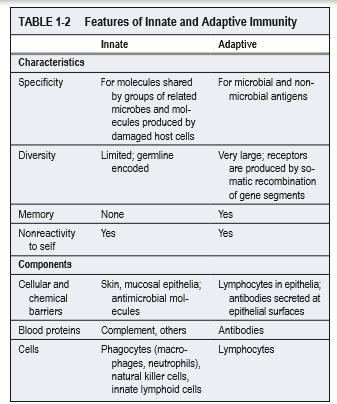




Kõik kommentaarid