Kordamisküsimused
1. Nüüdisaegse immunoloogia ja rakendusliku (sh. kliinilise) immunoloogia
arengu põhijooned. Immunoloogia areng Eestis.
Immunoloogia - teadus immuunsüsteemi funktsioonist normis ja haiguste korral, selle
mõjutamise võimalusest. Immuunsus – nakkustõvekindlus, ohustamatus, resistentsus,
infektsioonide jms suhtes. Immuunsüsteem - rakkude, kudede ja molekulide kooslus, mis
vahendab immuunreaktsioone, eeskätt infektsioonide korral.
Immunoloogia
teaduste
roll
meditsiinis ja selle erinevates
distsipliinides:
● ülesandeks on uurida neid rakulise
immuunsuse
nihkeid,
mis
määratlevad autoimmunisatsiooni
kujunemise
● immunoloogia põhieesmärgiks on
antigeensete märklaudmolekulide ja
nendega
seotud
immuunreaktsioonide uurimine rea
autoimmuunhaiguste
ja
mikroorganismide poolt indutseeritud
põletike korral.
● saada uut informatsiooni antikehade
ja rakkude poolt vahendatud
immuun-mehhanismidest
autoimmuunhäirete korral
● töötada välja uued seroloogilised ja
molekulaarsed meetodid nende
häirete
korral
esinevate
immuunreaktsioonide
iseloomustamiseks.
● olulisemaks praktiliseks ülesandeks
on
uute
immunoloogiliste
diagnostiliste ja ravi jälgimiseks
sobivate laboratoorsete meetodite
väljaarendamine
kliinikus
kasutamiseks
● immuunmehhanismide
uurimine
infektsioonide korral
● immuunmehhanismide
uurimine
mitte-infektsioossete haiguste jt.
immunoloogiliste seisundite korral
(allergia / atoopia; autoimmuunsus ja
autoimmuunhaigused;
kasvajad;
transplantatsioon)
Immunoloogia rakendamine teistes
teadusharudes:
● baasimmunoloogia (immunokeemia)
● meditsiiniline
immunoloogia
(immunogeneetika)
● kliiniline
immunoloogia
(immunoteraapia, transplantatsiooni
immunoloogia,
immunodermatoloogia, allergoloogia,
autoimmunoloogia jne)
● immunoloogilised
meetodid
-
Tänapäeva
meditsiinilise
immunoloogia põhiobjektid:
immuunsüsteem ja mikrobioom (ning
eksposoom)
immuunregulatsioon
o
kasvajad, allergia
ökoimmunoloogia,
immunotoksikoloogia jm
o
päikse poolt stim D-vit süntees
puudulik põhjamaades –
põhikomponent
immuunsüsteemi toetamiseks.
Immuunrakkude küpsemine
(eellaste külsemine) Treg-ide
just on D-vitamiinil kindel roll
o
uued ained; mikroplastik,
nanokihid-materajalid (uute
ravimite puhul kasutatakse)
organismi
immuun-homöostaasi
mõistmine
immuunsüsteemi seosed närvi- ja
endokriinsüsteemiga,
kudede immunoloogia
o
perifeerse vere kaudu
loomuliku (innate) immuunsuse
tähendus
o
loomuliku immuunsuse mälu –
letiud, et tegelt on
immunoloogilise mälu mehanismid
(sh. „treenitud immuunsus”)
o
vaktsineerimine – kui kaua
kestab mälu
reproduktiivimmunoloogia
o
viljatuse probleemid
COVID-19 immunoloogia!
haiguste vältimine immuniseerimise
abil
o
allergiad,
AI
(autoimmuun)haigused,
kasvajad
uute nn. bioloogiliste raviainete
rakendamine
jm.
immunobiotehnoloogia
geeni-, tüvirakkude jm. uusimate
ravimeetoditega
seostuvate
probleemide lahendamine
o
nt alguses efetktiivne, pärast
tundlikud ja spetsiifilised abivahendid
teistes biomeditsiini ja kliinilise
meditsiini valdkondades (põhinevad
antigeen-antikeha reaktsioonidel in
vitro; harvem in vivo)
● geneetika
● füsioloogia
● molekulaarne bioloogia
● evolutsiooniõpetus
mõju kadunud
Siin on palju detaile, mis moodustavad omavahel positiivsete ja negatiivsete
signaalidega seostatud süsteemi (võrgustiku) → IMMUNOOM
Kaasaegse immunoloogia arengusuunad: segadistsipliinid (Immunobiotehnoloogia -
immunoloogia uus suund, tegeleb kõrgefektiivsete diagnostiliste meetodite ja
ravivahendite läbitöötamisega biotehnoloogia abil)
Hiljuti arenes immunoloogia mitmete arstiteaduse valdkondade, eeskätt mikrobioloogia
rüpes. Viimastel aastatel on saadud olulisi teadmisi immuunsüsteemi erilisest
tähendusest mitteinfektsioossete põletikke tekkes, organismi üldise homöostaasi
tagamisel ja väliskeskkonna muutustega kohanemisel. Ka hiljuti kasutusele võetud
efektiivsed bioloogilised ravimid on seotud immuunsüsteemiga. Immunobiotehnoloogia -
immunoloogia uus suund, tegeleb kõrgefektiivsete diagnostiliste meetodite ja
ravivahendite läbi töötamisega biotehnoloogia abil).
2. Organismi kaitsebarjäärid (füüsikalised, keemilised, immunoloogilised).
Barjäärid, mis kaitsevad organismi on nii:
füüsilised (
nahk, kitiinkest, munakoor, lima, limaskestad, karvad, tsiiliad,
temperatuur, liikuv ripsepiteel, köha),
keemilised (mao madal pH [maohape, DNA laguneb kiiresti happelises, aluselises
lagunevad valgud], lagundavad ensüümid (
lüsosüüm, fosfolipaas, laktoferriin,
defensiinid), bioloogilised (mikrofloora).
Loomulik immuunsus (INNATE IMMUNITY)
fagotsüüdid – neutrofiilid, makrofaagid, nuumrakud
põletik –
palavik
Omandatud immuunsus (ADAPTIVE IMMUNITY)
B- ja T rakud
3. Immuunsüsteemi ehituslike komponentide iseärasused erinevates kudedes.
Primaarsete ja sekundaarsete immuunorganite tähendus ja seosed
immuunsüsteemi funktsioneerimises.
Immuunsüsteemi ehituslikud komponendid on rakud, mis on levinud üle kogu keha, olles
koondunud põhiliselt
tsentraalsetesse ja
perifeersetesse lümfoidorganitesse.
Tsentraalseteks lümfoidorganiteks on luuüdi ja tüümus. Perifeerseteks perifeersed
lümfisõlmed, põrn ja limakestade lümfoidne kude.
Primaarsed lümfoidorganid – T- ja B- rakkude küpsemise koht
Need on organid, kus toimub lümfopoees ja seal tekivad lümfotsüütide pinnale antigeeni-
retseptorid, tänu millele toimub funktsionaalne diferentseerumine.
Luuüdis toimub B-
lümfotsüütide ja
tüümuses T- lümfotsüütide valmimine ja küpsemine. Primaarsetes
lümfoidorganites omandavad lümfotsüüdid võime eristada „oma“ „võõrast“.
Sekundaarsed lümfoidorganid – immuunvastuste tekkepaik
Need on organid, kus toimub interaktsioon antigeeni ja immuunsüsteemi rakkude vahel,
aga ka immuunsüsteemi komponentide eneste vaheline interaktsioon. Peamiseks
ülesandeks on filtreerida ja kontsentreerida võõrast antigeeni. Sekundaarsed
lümfoidorganid on põrn, lümfisõlmed, MALT.
Vahel eristatakse ka tertsiaarseid lümfoidorganeid, mis on difuusselt organismis
lokaliseeruvad lümfoidkoe rakud ja nende kogumikud.
Immuunsüsteemi ehituslikest komponentidest:
APC – antigeeni haaravad ja lümfotsüütidele esitavad rakud. professionaalsed (DC,
makrofaagid, B-rakud) + mitteprofessionaalsed (epiteelirakud – pigem on passiivsed,
ei käivita immuunreaktsioone)
Lümfotsüüdid immuunsüsteem kesksete rakkudena
Efektorrakud – elimineerivad mikroorganisme (CTL), lümfotsüüdid kui ka
leukotsüüdid, fagotsüüdid ka
Organismi immuunsuses osalevad vererakkudest valgelibled e leukotsüüdid, mis
jagunevad vastavalt graanulite olemasolule. Agranulotsüüdid (u 65%) ja granulotsüüdid
(35%).
1.
Granulotsüüdid
Neutrofiilid – neid leidub veres
kõige rohkem. Neutrofiilidele
on omane hästi väljakujunenud
fagotsütoosivõime,
nad
fagotsü-teerivad
peamiselt
mikroobe, kuid põletikukolletes
ka väikesi võõrkehi ja rakkude jääke.
o Kiire haigustekitaja leviku pidurdamine
o NET – viskab graanulite võrgu välja millega hävitavad mikroobe
o „3 tuuma“
Eosinofiilid – organismi kaitsereaktsioonideks võõrvalkudele, samuti allergia
(pidurdavad histamiini eraldumist basofiilidest) ja anafülaksia korral.
o Palju graanuleid, nn kaheosaline tuum, värvub punaselt
Basofiilid – histamiini ja hepariini ainevahetuses – põhjustavad allergiliste
reaktsioonide korral turset.
o Tsütoplasmat ka, siniselt värvub, tuum on laiali, palju graanuleid siniseid
Nuumrakud – palju tüsoplasmat, ümmargune tuum – ka parasiidid ja allergeenide
vastane immuunvastus
2.
Agranulotsüüdid
Lümfotsüüdid – suure tuumaga, väiksed ümmargused
o T- ja B- rakkudel väga diverse ja spetsiifilised antigeensed retseptorid –
vastutavad immuunsüsteemi spetsiifilisuse eest ja mälu eest. Palju
pinnamarkereid
o
Tüümussõltuvad lümfotsüüdid – T-lümfotsüüdid, tekivad luuüdi
tüvirakkudest tüümuses, kust nad siirduvad perifeersete lümfoidorganite T-
tsoonidesse, kus toimub uute T-lümfotsüütide teke. T-lümfotsüüdid
kindlustavad rakulise immuunsuse, kuid reguleerivad ka humoraalse
immuunsuse kulgu. Teise liigi T-lümfotsüüte moodustavad rakud, mis
mõjutavad B-rakkude funktsioone. Siia kuuluvad T-helperid ja T
regulatoorsed rakud. Eristatakse ka T-mälurakke.
o
TH1 vastus toodab tsütokiine, mis toetab
põletiku teket ja aktiveerib
peamiselt
T-rakke ja
makrofaage.
o
TH2
vastus
aktiveerib
peamiselt
B-rakke
ja
immuunvastuseid, mis sõltuvad
antikehadest
o
TC rakud aktiveeritakse, kui nad
interakteeruvad
MHC klass I-
antigeen kompleksiga mõne
ebanormaalse oma organismi
rakuga, nt kasvajarakk, vaja on
ka
õigeid
tsütokiine.
Aktivatsioonile
järgneb
proliferatsioon,
rakkude
diferentseerumine efektoriks –
tsütolüütiline T-lümfotsüüt
CTL.
Helperitega võrreldes on CTL-de poolt sekreteeritud tsütokiinide
hulk väike. Neil on hoopistükis võime oma organismi rakke ära tunda ja
elimineerida.
o
B-lümfotsüüdid (bone marrow) kujunevad samuti vere tüvirakkudest.
Nad tagavad põhiliselt humoraalse immuunsuse, mille efektrorrakuks on
plasmarakk. B-lümfotsüütidest moodustunud plasmarakud produtseerivad
antikehasid.
o
NK rakud (Natural killer cells) – lümfotsüütide eri variandina tuntakse veel
naturaalseid hävitajaid rakke (NK-cells), mis on võimelised hävitama
muundunud rakke.
Viirusega nakatunud rakud, kasvajarakud (stressimarker peal).
NK rakud tutvuvad normaalsete MHC-dega ning eristavad
normaalseid rakke „kummalistest“
o
ILC-d – innate lymphoid cells – loomuliku immuunsuse efektorrakud
Lümfotsüütide elu kestab mõnest nädalast mõne aastani. Enamiku veres
ringlevatest lümfotsüütidest moodustavad retsirkuleeruvad pikaealised T-
lümfotsüüdid, mis funktsioneerivad kuude või aastate vältel, lühiealisi B-
lümfotsüüte on vähem. Lümfotsüüdid püsivad perifeerses veres lühikest aega.
Lümfotsüüdid
migreeruvad kudedesse, aga samuti ka õõnesorganite
limaskestade pinnale. Kudedes liiguvad nad aeglaselt. Liikumis- ja
fagotsütoosivõime on neil vähe arenenud. On kindlaks tehtud, et osa lümfotsüüte
pöördub kudedest lümfisõlmedesse ja –sõlmekestesse tagasi ning siirdub sealt
koos voolava lümfiga uuesti verre, mis kannab neid muude organite kõrval ka
tagasi luuüdisse. Seda nähtust nim lümfotsüütide retsirkulatsiooniks.
Monotsüüt (kui läheb koesse, on makrofaag) – esitavad immuunsüsteemi
rakkudele antigeene. Nad on leukotsüütidest kõige suuremad. Suur tuum, vahel
neerukujuline. Fagotsütoosi võime. Monotsüütidele on iseloomulik sõrmjate
tsütoplasmaatiliste jätkete, samuti fagotsütaarsete ja pinotsütaarsete vakuoolide
moodustumine. Monotsüüdid kuuluvad organismi makrofaagide süsteemi (MPS).
Monotsüüdid ringlevad veres ja kujutavad endast diferentseerumisvõimeliste
rakkude reservi.
Dendriitrakud – professionaalsed APC-d. Asuvad kudedes mis puutuvad paljude
väliste antigeenidega kokku. Esitavad antigeeni informatsiooni lümfotsüütidele.
Iseärarasused erinevates kudedes Rohkem kui pooled kõigist lümfoidrakkudest paiknevad gastrointestinaal- ning
respiratoorsete teede limaskestades -
MALT e limaskestadega assotseeruv
lümfoidkude.
Luuüdi. Koht, kus nii B- kui T rakud alguse saavad (T rakkud liiguvad Tüümusesse). Nad
saavad ühisest eellasest alguse. Ühtlasi toimub seal hematopoees.
Punane luuüdi on
erütrotsüütide, trombotsüütide ja granulotsüütide tekke paik. Punane luuüdi
koosneb põhiliselt retikulaarsest sidekoest, milles leidub igasugusel arenguastmel
olevaid vererakke. Kõikide vererakkude eellasteks peetakse müeloblaste –
hemotsütoblastide järglasi. Müeloblastid arenevad edasi protoerütroblastide ja
promüeloblastide liinis. Protoerütroblastidest arenevad erütrotsüüdid, promüeloblastidest
granulotsüüdid. Müeloblastidest arenevad ka suured megakarüoblastid. Luuüdi
retikulaarse sidekoe rakud võivad muutuda makrofaagideks.
Lümfisõlmed. Iga lümfisõlm koosneb koorest (core) (sisaldab B rakke, spetsiaalseid
antigeeni-esitlevaid rakke (antigen-presenting cells, APC)) ja sisemisest säsist (medulla)
(T-, B- ja plasmarakud). Lümfisõlmed on 0.1-2.5cm pikad. Lümfisõlmedes on
germinaaltsentrid, kus toimub B-rakkude küpsemine. Nendes on tume ja hele tsoon. T- ja
B rakud paiknevad lümfisõlmedes erinevates kohtades.
Lümfisoon aitab immuunsüsteemi rakkudel kiiresti liikuda (kaasasündinud immuunsuse
rakkudel eriti) infektsioonikohtadesse ja aiitab väiksel arvul lümfotsüütidel (mis on antud
antigeenile spetsiifilised) reageerida sellega – olenemata sellest kust antigeen pärit on.
Lümfisõlme ehitus: väljaspoolt on ta kaetud sidekoelise kapsliga, millest siirduvad sõlme
sisse trabekulid, vaheseinad ja põimuvad seal üksteisega võrgustikuks. Nende
vaheseinte vahekohtades asetsevad lümfifolliikulid, millest suunduvad säsisse
säsiväädid.
Säsiväätides leidub rohkesti
lümfotsüüte, hemolümfoblaste, plasmarakke.
Kõik need vaheruumid, mis jäävad lümfisõlme tema
kihnu ja folliikulite ning säsiväätide vahele ja mis on
täidetud hõredarakulise retikulaarse sidekoega,
kannavad lümfiurgete nimetust. Lümfiurgete seinteks
on väga suure fagotsütaarse aktiivsusega
retikuloendoteliaalsed rakud, mis võivad eralduda ja
muutuda makrofaagideks. Lümfiurgetes rikastub lümf
lümfotsüütide ja plasmarakkudega. Üldiselt
samasuguse ehitusega on ka Peyeri naastud ja mandlid. Lümfisõlmes toimub lümfi
filtreerimine võõrkehadest, fagotsütoos, lümfotsüütide produktsioon ja antikehade
moodustumine.
Põrn. Põrn koosneb kihnust, millest kulgevad sidekoelised põrgad elundi sisemusse ja
põrkade vahele jäävast säsist. Põrna (punane) säsi koosneb retikulaarsest sidekoest,
selles leidub leukotsüüte ning erütrotsüüte. Punase säsi retikulaarse sidekoe rakud
võivad fagotsüteerida, lammutades seega vanu erütrotsüüte ja nende osi. Põrna Malpighi
kehakesed ehk hall säsi on lümfotsüütide, monotsüütide ja plasmarakkude
tekkimispaigaks. Tavalistest lümfisõlmedest erinevad nad selle poolest, et neis puuduvad
lümfisooned ja –urked ning nende talitlus toimub ainult veresoonte vahendusel. Põrn on
vere depooks. Füüsilise pingutuse korral paisatakse põrnast vere varu vereringesse ja
suurendatakse seega hapniku transporti.
Tüümus ehk harkelund on kahest sagarast koosnev paariline elund. Ta tekib neelu
epiteelist. Oma arenemise haripunkti jõuab tüümus varajases lapseeas, suguküpsuse
saabumisel hakkab atrofeeruma ja tema kude asendub rasvkoega. Tüümus on kaetud
sidekoelise kihnuga, millest elundi sisse ulatuvad vaheseinad ning jaotavad ta
sagarikkudeks. Sagarikud koosnevad omakorda koorest ja säsist. Koores on väga palju
väikesi lümfotsüüte, säsis on neid märksa vähem. Lümfotsüüdid pärinevad luuüdist, kust
nad liiguvad veretee kaudu tüümusesse. Tüümuses lümfotsüüdid paljunevad ja “õpivad”
olema tolerantsed keha rakkude suhtes ning vaenulikud võõra vastu. Vereteedesse
lahkumisel on lümfotsüüdid muutunud immunoloogiliselt küpseteks e
immunokompetentseteks. Tüümuses tekivad looteeas rakud, mis kanduvad
lümfoidsetesse organitesse ja panevad alguse antikehasid tootvate plasmarakkude
tekkele.
Peyeri naastud. Peensoole limaskesta päriskiht koosneb põhiliselt kohevast sidekoest,
milles leidub plasmarakke, eosinofiile ja lümfisõlmi. Peensoole algusosas on lümfisõlmi
vähe, niudesooles suureneb nende arv märgatavalt. Paiguti moodustavad nad seal suuri
kogumeid – Peyeri naaste.
Limaskesta hattudes on päriskiht rikkalikult varustatud
verekapillaaride ja keskse lümfisoonega.
Jämesoole epiteeli päriskihis on rohkesti lümfoidse koe kogumeid, eriti rohkesti on
neid umbsoole ussripikus, mis on otsekui mingi lümfaatiline elund, mistõttu nimetatakse
teda mõnikord “soolemandliks”.
Makrofaagid kudedes
Alveolaarsed makrofaagid – kopsus
Histiotsüüdid – sidekoes
Kupfferi rakud – maksas
Mesangiaalrakud – neerudes
Mikrogliia rakud – ajus
Osteoklastid – luudes
Üldjuhul on makrofaag puhkavas seisundis. Fagotsütoos aktiveerub esialgselt kindlate
antigeeni toimel ja veelgi enam saab aktiivsust tõsta
tsütokiinide abil, mis on
sekreteeritud aktiveeritud TH-rakkude poolt, põletikuliste mediaatormolekulide
poolt ja bakterite rakuseinte komponentide tuvastamisel.
4. Limaskestade ja nahaga assotsieeruv lümfoidne süsteem, ehitus ja roll
immuunsüsteemis.
Lokaalsest immuunsüsteemist rääkides tuleb teda ruumiliselt lahterdada: perifeersed
lümfisõlmed ja põrn; limaskestadega assotsieerunud lümfoidkoed; serooskelmetega
kehaõõned (pleura, peritoneum); nahk. Igas lahtris algatatud immuunvastuse
efektormehhanismid toimivad vastavalt selle „lahtri” piires.
Immuunvastuse lokaalse lahterduse tagamine toimub selle kaudu, et naiivsetele T
rakkudele antigeenide esitlemisel programeeritakse T rakud igas „lahtris” erinevalt.
Seetõttu on erinevates „lahtrites” efektor-T-rakkudel erinevad adhesioonimolekulide ja
kemokiinretseptorite komplektid, mis tagavad nende rakkude koespetsiifilisemat
pesastumist. Erinevate „lahtrite” koerakud ekspresseerivad vastavaid
adhesioonimolekulide ligande ja toodavad vastavaid kemokiine. Lokaalsetes „lahtrites”
kontrollivad lümfotsüütide liikumisi koespetsiifilise adhesiooni ja kemokiinide mõjud.
IgA-l keskne osa ja Tregidel, kes immuunvastust „kasuliku“ vastu ära hoiavad
Limaskestadega assotsieeruv immuunsüsteem kuulub adaptiivse immuunsuse hulka. Ta
tagab lokaalset kaitset kogu keha ulatuses, paiknedes peamistes antigeenide
sissetungimise väravates ning moodustab 50% kogu lümfoidsest koest. Teda võib
vaadelda kahe omavahel interakteeruva osana:
a) organiseerunud koed, kus antigeene seotakse e
MALT- mucosa associated
lymphoid tissues (saab jagada veel omakorda kolmeks:
GULT- gut (sool) associated
lymphoid tissue,
nasal (nina) ehk NALT ja bronchial (bronhiaalne) ehk BALT
(viimased on seotud
respiratoorse traktiga) ja
b) difuussed lümfoidsed koed, kus vastavad rakud ja antikehad talitlevad kaitsjatena.
sIgA neutraliseerib sisse tungivad patogeenid – IgA-d sekreteerivateks plasmarakkudeks
saavad B rakud muutuda nii T rakust
sõltuval kui sõltumatul viisil. Dimeerne sIgA
transporditakse läbi epiteeli ja vabastatakse luumenisse. Kuigi osa IgA antikehadest
tehakse süsteemses immuunsüsteemis (see tähendab, et osaliselt luuüdis), valmib
enamik siiski plasmarakkude poolt toodetuna lamina proprias. See antikeha
transporditakse üle epiteliaalsete rakkude trakti luumenisse ehk valendikku
spetsialiseerunud mehhanismi abil: IgA subepiteliaalses lamina proprias seondub IgA
polü-Ig retseptoritega ja transporditakse endosoomide kaudu luumeni ehk valendiku
pinnale.
Sekretoorset IgAd leidub pisarates, süljes, rinnapiimas, kolostrumis jne. Tänu sellele, et
teatud sekretoorne osake on seotud IgA antikehaga, väheneb proteolüütiliste ensüümide
poolt degradatsiooni oht intestinaalses keskkonnas. IgA esineb nii monomeerses kui ka
dimeerses (kahte monomeeri hoitakse koos J-ahela abil) vormis. Monomeerne vorm on
tsirkulatsioonis ja ta omab kahte alaklassi. Dimeerne vorm osaleb primaarses kaitses
lokaalsete infektsioonide vastu, olles süljes, pisarates, bronhiaalsetes sekreetides,
nina limas, prostata vedelikus, vaginaalsetes sekreetides ja mukoossetes peensoole
sekreetides.
1. MALT iseloomustus: (limaskestad üldiselt)
MALTis või ära voolavates lümfisõlmedes tekitatud efektor lümfotsüüdid liiguvad verre ja
tulevad samasse organisse tagasi (homing mechanisms).
Limaskestade epiteel on õhuke, oma funktsioonide täitmiseks mingil määral läbilaskev
(gaasivahetus, toitainete absorptsioon, sensoorne või reproduktiivne funktsioon). Samas
on limaskestad kergelt haavatavad ning paljud haigusetekitajad tungivadki just
limaskestade kaudu organismi. Soole limaskest on ka suure hulga võõrantigeenide (toidu
näol) sissetungi koht. On olemas mehhanismid, mis väldivad toidu vastu jõuliste
immuunreaktsioonide teket. Lahustuvate antigeenide vastu tekib tolerants ilma
põletikuta. Samas on ka mehhanismid, mis teevad kindlaks ja reageerivad
sissetungivatele patogeenidele: jõuline ja kaitsev Th1 immuunvastus patogeenidele.
1.1. NALT ja 1.2. BALT - nasaalne ja bronhiaalne limaskestadega seonduv
lümfoidne kude: need mikroobid, mille tungimist kopsu ei suudeta ära hoida
sekreteeritud lima ja ripsmete liikumise abil, satuvad kontakti lümfoidsete agregaatidega
nagu
mandlid ja adenoidid ning
bronhiaalsed sõlmekesed, milles nende
elimineerimine toimub sarnaselt soolega. Selle puudumisel või häiretel (BALT) võib
esineda hingamishaigusi nt astma.
1.3. GULT - Soole limaskestaga seonduv lümfoidne kude:
Tuleb eristada patogeene kommensaalidest, kes on inimese soolele kasulikud.
kõige paremini uuritud MALT, mis sisaldab
Peyeri naaste (nendes olevad M- ehk
microfold rakud teostavad transtsütoosi – vahendavad partikleid ja molekule
soolevalendikust dendriitrakkudele ja lümfotsüütidele) ja teisi agregaate, mis on
spetsialiseerunud
antigeenide esitlemisele ning transpordivad mikroobe microfold
rakkude kaudu subepiteliaalsesse piirkonda (M-rakud võtavad soolevalendikust antigeeni
endotsütoosi kaudu vastu ja annavad üle APC (antigen presenting cells) rakkudele), kus
neid töödeldakse ja esitletakse
MHC II klassi dendriitrakkude ja
makrofaagide poolt
T rakkudele, mis sisenevad koesse. Selle häirel võib tekkida nt Chrons haigus –
loomuliku ja omandatud immuunsuse mehhanisme ei osata kontrollida.
Nahaga assotsieeruv lümfoidne kude (SALT)Kaitseb mikroobide läbi naha tulekut ja supresserib komensiaalsete organismide
tegevust. Nahal on mitmekihiline epiteel – kertiniseeritud – epidermis – teeb loomuliku
immuunsuse mehhanisme – füüsiline barjäär mikroobide invasiooni vastu.
Keratinotsüüdidd sekreteerivad defensiine ja põletiku tsütokiine. Naha kiht dermis
sisaldab palju immuunrakke: nt nuumrakud, makrofaagod ja dendriitrakud (DC) – nad
kõik vastavad mikroobidele ja vigastustele ja vahendavad immuunvastusi.
Naha DC-d vahendavad loomuliku immuunsuse vastuseid, transpordivad mikroobseid ja
kk antigeene mis naha kaudu sisenevad lümfisõlmedesse, kus nad alustavad T rakulist
vastust. Naha lümfisõlmedes aktiveeritud T-rakud ekspresseerivad kemokiini
retseptoreid ja adhesioonimolekuli mis soodustavad T rakkude migreerumist naha poole.
Nahas on nii CD4+ kui CD8+ T rakke. Helperitest on nii Th1, Th2 kui Th17 T helperid, kes
kaitsevad nahka invadivate patogeenide eest kuid on olulised ka põletiku protsessides
naha autoimmuun haiguste korral – psoriaas nt ja atoopiline dermatiit.
Difuusne lümfoidkude: B-rakkude kloonid lahkuvad mälurakkudena või plasmarakkude
eellasrakkudena äraviivate lümfisoonte kaudu lümfisõlmedest. Need rakud liiguvad
tsirkulatsiooni ja „home back” ehk lähevad tagasi elundite ja teiste difuusse lümfoidkoe
piirkondadesse, kus leidub peamiselt IgA plasmarakke. See pakub kaitset teiste
mikroobide sisenemiste eest. Lümfi lamina propria sisaldab T rakke ja on rikas ka
nuumrakkude poolest. CD8+ T rakke on leitud epiteliaalsete rakkude hulgas hulgas.
Samuti on nuumrakud olulised, sest nad osalevad parasiitide vastases kaitses. Kuigi
nende vastases kaitses on peamised IgE antikehad, aitab nuumrakkude degranulatsioon
tekitada ägedat põletikulist vastust, kui toimub ussnugiliste antigeenide presentatsioon.
5. Loomulikku immuunsust tagavad rakud ja humoraalsed faktorid. „Treenitud
immuunsus“.
Humoraalsed faktorid
Komplemendisüsteem
Lüsosüüm on paljude kehavedelike (pisarad,uriin) koostises olev antibakteriaalne
ensüüm. Lagundab gram-positiivsete bakterite seina.
Aktiivse faasi valgud
Tsütokiinid
Rakud
•
Neutrofiilid ja makrofaagid on fagotsüüdid, mis tapavad mikroobidega nakatunud
rakke produtseerides ROS-i, NO-d ja fagolüsosoomi ensüüme. Makrofaagid toodavad
lisaks tsütokiine, mis stimuleerivad põletiku ja nakatunud koe ümberkorraldusi.
Fagotsüüdid tunnevad mikroobseid produkte ära erinevate retseptorite kaudu (nt TLR,
NLR, scavenger retseptorid jne).
•
Dendriitrakud on olulised kaasasündinud ja omandatud immuunvastuste sidumisel.
Nad ekspresseerivad PAMPe äratundvaid retseptoreid. Aktiveerimisel toodavad
tsütokiine.
•
NK rakud ehk loomulikud tapjarakud on rakud, mis tunnevad ära nakatunud rakud
ja/või stressis rakud ning hävitavad need. NK rakud sisaldavad põletikulisi tsütokiine,
mille abil on nad võimelised sihtmärkrakke hävitama. Lisaks sisaldavad NK rakud IFN-
γ, mis on oluline makrofaagide aktiveerimiseks. NK rakkude aktivatsioon on
reguleeritud inhibeerivatelt ja aktiveerivatelt retseptoritelt tulenevate signaalide
tasakaaluga
•
Nuumrakud
Lümfoidse rea rakkudel on transkriptsioonifaktorid, mis suunavad neid arenema selleks.
6. Loomuliku immuunsuse tsütokiinide toimerajad.TNF - neutrofiilide aktivatsioon
(põletik), palavik jt
IL-1 - põletik, palavik, akuutse faasi
valkude süntees (maks), Th17
rakkude diferentseerumine jt
IL-12 - T rakkude diferentseerumine
jt, IFN sünteesi stimulatsioon
IFNα,β - MHC I ekspressiooni
stimuleerimine, NK rakkude
aktivatsioon
IL-6 - akuutse faasi valkude süntees
(maks), T ja B rakkude
stimulatsioon
IL-15 - NK rakkude paljunemine
IL-18 - NK ja T rakkude aktivatsioon,
IFN süntees
IL-23 - IL-17 produtseerivate
rakkude stimulatsioon
IL-27 – mitmesugune mõju T ja NK rakkudele
Toime sõltub rakkude kooslusest ja tsütokiini kontsentratsioonist
Lokaalsed (parakriinne jm) ja süsteemsed efektid
Makrofaagide
poolt
TNF – T rakud ka. põletik, neutrofiilide aktivatsioon, palavik
IL-1 - põletik, palavik, akuutse faasi valkude süntees (maks),
Th17 rakkude dif jt
IL-6 – T rakud. akuutse faasi valkude süntees (maks), T ja B
rakkude stimulatsioon
IL-15 - NK rakkude paljunemine
IL-18 - NK ja T rakkude aktivatsioon, IFN süntees
IL-10 – tregide poolt ka. IL-12 inhibitsioon, MHC II ekspressioon
IL-15 – NK paljunemine, CD8+ paljunemine
IL-18 – NK ja T rakkude poolt IFN γ süntees
IL-12 Th1 diferentseerumine
Dendriitrakud
ja
makrofaaagid
Tüüp I IFN (α, β) – NK aktivatsioon, MHC I ekspressioon
IL-23 – T rakud: IL-17
IL-27 – Th1 dif., Th17 inh. NK IFN γ.
+ Loomuliku immuunsuse kemokiinid
JAK-STAT signaalirada – viirusinfektsioonide korral
7. TLR keskne tähendus. NLR ja RLR. Makrofaagide ja dendriitrakkude poolt
produtseeritavad tsütokiinid. Nende tähendus immuunsüsteemi
funktsioneerimisel..
Loomuliku immuunsuse aktivatsioonis on keskne roll
rakkudel, mille pinnal on
mikroorganisme ja lagunenud keharakke äratundvad retseptorid e PRR -
pattern recognition receptors (molekulaarseid mustreid äratundvad retseptorid).
Need retseptorid reageerivad molekulidega, mida kutsutakse:
PAMP – pathogen associated molecular patterns – patogeeni molekulaarsed mustrid.
o LPS – endotoksiin
o CG rikkad (CpG) oligonukleotiidid
o dsRNA
o mannoosijääkidega molekulid jms
DAMP – damage associated molecular patterns – koekahjustuse puhul
o Ekstratsellulaarne ATP mitokondritest
o Histoonne valk rakudtuumadset
PRR retseptorid:
TLR – Toll-like receptors
CLR – C-type lectin receptors (I mannoos-R, II asialoglükoproteiin-R)
NLR – NOD-like receptors
RLR – RIG-like receptors
DNA-sensorid (tsütoplasmas)
Tolli laadsed retseptorid ehk TLR-del on oluline roll loomulikus immuunvastuses. TLR
ekspresseeritakse makrofaagide, dendriitrakkude, neutrofiilide, mukoosa epiteelrakkudel
ja endoteelrakkudel. TLR geenid on kõik glükoproteiinid ning sisaldavad
ekstratsellulaarses regioonis leutsiini kordusjärjestusi ja tsüsteiini motiive.
Tsütoplasmaatilises osas sisaldavad TLR-id TIR (Toll/ IL-1 retseptor) homoloogilist
domääni, mis on vajalik signaliseerimiseks.
Imetajate TLR on oluline erinevat tüüpi molekulide äratundmiseks, mis ekspresseeruvad
mikroobide pinnal. Kuna imetajate TLR asuvad nii rakkude pinnal kui ka
intratsellulaarsetel membraanidel on imetajate rakud võimelised mikroobe ära tundma
nii raku sees kui väljas. Imetajate rakusisesed (ER pinnal) TLR-id on 3,7,8 ja 9 ning nad
on olulised mikroobide nukleiinhapete äratundmisel.
Ekstratsellulaarseteks TLR-id: neid stimuleerivad erinevad PAMPid.
TLR1:2 – bakteriaalsed lipoproteiinid
TLR2 – gram-positiivsete bakterite peptiidoglükaan
TLR4 – LPS (lipopolüsahhariidid) – GN bakterid
TLR5 – bakteriaalne flagelliin
TLR2:6 – bakteriaalsed lipopeptiidid
Rakusisesed (endosomaalsed) TLR-id:
TLR3 – dsRNA
TLR7 – ssRNA
TLR8 – ssRNA
TLR9 – CpG DNA (metüleerimata)
TLR signaliseerimisrada: TLR seondab ligandi (milleks on PAMPi sisaldav molekul) → TLR
molekulide dimeriseerumine → adapter proteiinide aktiveerimine (TIR domään sisaldab
neid) → proteiin kinaaside aktiveerimine → transkriptsioonifaktorite aktivatsioon → geeni
transkriptsioon → ekspresseeritakse: põletikulisi tsütokiine (TNF, IL-1, IL-12), kemokiine
(IL-8), endoteliaalseid adhesiooni molekule (E-selektiin) jne. Vaja kahe retseptori poolt
signaali.
NLR –
nod like receptor (NACHT-LRR) on perekond
tsütoplasmaatilisi molekule, mis
on olulised intratsellulaarse bakteriaalse infektsiooni vastu võitlemisel. NLRid seovad
tsütoplasmas bakteri derivaate, nt peptiidoglükaani. NLRP3 inflammosoom IL-1β
tootmine
RLR (RIG-1 like receptor) on samuti perekond tsütoplasmaatilisi molekule, mis on
olulised viirusinfektsioonide vastu võitlemisel. Äratuntavaks viiruste molekuliks on
dsRNA. Osadel RLR retseptoritel on CARD (caspase activation and recruitment domain)
domään, mille kaudu aktiveeritakse signaalirajad, mille tulemusena sünteesitakse tüüp I
interferoone.
Makrofaagide ja dendriitrakkude poolt produtseeritavad tsütokiinid. Nende
tähendus immuunsüsteemi funktsioneerimisel.
Kaasasündinud immuunsüsteemi rakkude poolt toodetud tsütokiinid aktiveerivad
leukotsüüte ja põhjustavad süsteemi ümberkorraldusi; suureneb efektor rakkude süntees
ja antimikroobsete valkude süntees.
Dendriitrakud produtseerivad tsütokiine IL-12, IL-23 ja IL-27, IFN α, β,
Makrofaagid toodavad IL-1,
IL-6, IL-12, IL-15, IL-18, IL-23, IL-27, TNF, kemokiine, INF-αt.
Nendel tsütokiinidel on erinevad rollid immuunsussüsteemi reguleerimises. Nt INF-α
kutsub esile antiviraalse oleku, klass I MHC molekuli ekspressiooni aktivatsiooni ning NK
rakkude aktivatsiooni. Nt TNF aktiveerib neutrofiile, muudab hüpotalamuse temperatuuri
”set pointi” (palavik) ja põhjustab paljude rakutüüpide apoptoosi
8. Loomuliku immuunsuse lümfoidse rea rakud (ILC), NK-rakud ja nende
pinnaretseptorid –iseloomustus ja funktsioon.
ILC – innate lymphoid cell – on lümfoidse rea rakud loomulikus immuunsuses. ILC1
alatüüp on NK rakud, ILC2- on sarnased Th2-le. Id2 transkriptsioonifaktor
IL-12, IL-18
ILC1 IFN γ – viirusevastane kaitse
o IL-12, IL-15
NK rakud IFN γ
IL-25, IL-33, TSLP
ILC2 IL-4, IL-5, IL-13 – allergiline põletik
IL-23, IL-1 β
ILC3 IL-22, IL-17 – soole barjääri funktsioon,
lümfoidorganogenees
NK rakud (natural killer) ehk loomulikud tapjarakud on rakud, mis tunnevad ära
nakatunud rakud ja/või stressis rakud ning hävitavad need. NK rakud sisaldavad
põletikulisi tsütokiine, mille abil on nad võimelised sihtmärkrakke hävitama. Lisaks
sisaldavad NK rakud IFN-γ, mis on oluline makrofaagide aktiveerimiseks. NK rakud on
eriti olulised võitlemaks intratsellulaarsete mikroobide ja viirustega haiguse algstaadumis
kui CTL rakud ei ole veel täielikult aktiveeritud. NK rakud, nagu ka makrofaagid,
sisaldavad
graanuleid proteiinidega, mis algatavad sihtmärkrakkude hävitamise. Kui NK
rakud aktiveeritakse, siis eksotsüteeritakse need proteiinid sihtmärgini. Perforiin on
üheks selliseks proteiiniks ja selle proteiini tulemusena pääsevad rakku ülejäänud
gransüümid.
NK rakkude aktivatsioon on reguleeritud
inhibeerivatelt ja aktiveerivatelt
retseptoritelt tulenevate signaalide tasakaaluga. Selleks, et NK rakud ei ründaks
”omasid” peavad inhibeerivad signaalid olema ülekaalus.
Rakud, mis on stressis
ja/või nakatunud viiruse või intratsellulaarse mikroobiga ekspresseerivad
rakupinnale erinevaid ligande mida tunnevad ära NK rakkudel olevad
aktivatsiooni retseptorid. Üheks selliseks retseptoriks on NKG2D, mis seondab klass I
MHC molekuli laadseid proteiine, mida võib leida viirusega nakatunud rakkudest ja
kasvajarakkudest. Lisaks retseptoritele reguleerib NK rakkude aktiivsust tsütokiinid,
peamiselt makrofaagide poolt toodetud IL-12 ja IL-15. IL-15 on NK rakkude kasvufaktoriks
ja IL-12 on oluline interferoon γ produtseerimiseks NK rakkudes ja tsütotoksilise
aktviisuse juures.
Pinnaretseptorid:
Nendel on ITAM – mis aktiveerib need. ITIM – inhibeerib.
KIR (killer cell inhibitory receptor) – on üheks NK rakkudel olevaks inhibeerivaks
retseptoriks, mis seondub tervete rakkude klass I MHC molekuliga.
Aktiveerivad retseptorid NK rakkudel:
Fc eeta RI γ, sigma retseptor
CD16
Fc eeta RI γ, sigma, DAP12
NCR-id,
DAP12 KIR2DS
DAP23 CD94, NKG2C, E
DAP10 NKG2D
Inhibeerivad retseptorid NK
rakkudel:
NKG2A CD94, ILT2,
KIR
9. Komplementsüsteem. Tähendus, tähtsamad molekulid ja retseptorid.
Komplement (complement = C) – seerumvalkude süsteem, mille aktiveerumisel käivitub
ensüümreaktsioonide ahel, mis võib esile kutsuda rakkude lüüsi.
Komplementsüsteem
sisaldab rida seerumvalke, mis toimides koos reguleerivad põletiku teket,
aktiveerivad fagotsütoosi ja lõhuvad rakuväliste infektsioonitekitajate
rakumembraane. Süsteemi kuulub üle 20 molekuli. Moodustab 10% seerumvalkudest,
toodetakse enamuses maksas, kuid mõningaid (C2, C3, C5 jne) võivad toota ka
aktiveeritud makrofaagid põletikukoldes. Klassikalise tee komponendid on C1q, C1r, C1s,
C2, C3, C4, C5, C6, C7, C8, C9. Alternatiivse tee komponendid on properdiin, faktor B,
faktor D. Inhibiitorid on C1 inhibiitor, faktor I. Regulatoorsed proteiinid on C4-siduv
proteiin, faktor H, S proteiin (vitronektiin). Komplementsüsteemi põhifunktsioonide hulka
kuuluvad:
põletikureaktsiooni käivitamine ja kontroll (C4a, C2b, C5a),
opsonisatsiooni teel fagotsütoosi tugevdamine (C3),
rakumembraanide otsene kahjustamine (MAK-C-C9),
immuunkomplekside töötlemine,
antigeeni sidumine APC rakkudele paremini omastatavas vormis.
Komplemendi retseptorid on CR1-CR4. CR1 ja CR2 moodustuvad nn komplemendi
kontrolli proteiinist (CCP). CR3 ja CR4 on integriinid sisaldades sarnast β -ahelat (CD18).
Retseptorite funktsioonideks on IK sidumine transpordi eesmärgil (IK clearance),
opsonisatsioon, komplemendi aktivatsiooni reguleermine. Komplemendi osa põletiku
tekkes kannavad edasi anafülotoksiinid (kemoatraktandid - C3a, C5a). Nende toimel
verevoolu hulk suureneb, veresoonte läbilaskvus tõuseb, rakkude migratsioon kudedesse
suureneb ja põletiku tsütokiinide tase tõuseb, kudedes tekib nn oksüdatiivne plahvatus.
10.
Komplementsüsteemi kolm aktivatsioonirada ja nende tähendus.
Komplementsüsteemi aktivatsiooni teedeks on
klassikaline (adaptiivne immuunsus), alternatiivne
(loomulik immuunsus) tee ja lektiinne tee.
Klassikalise
tee
aktivatsioon
algab
immuunkompleksi moodustumisega. Antikeha
seondub antigeeniga. Ak võime
aktiveerida
komplementi sõltub nende klassist (ja erineb liigiti).
C1 koosneb C1q, C1r, C1s molekulidest. Iga C1 on
C1q kaudu seondunud antikehale Fc saidi kaudu.
Antikeha Fc osa seondumisel C1q alaosaga toimub
C1r-i autoaktivatsioon. C1r lõikab ja aktiveerib C1si liituvad C4 ja C2, C1s lõikab nad:
C2a + C4b tekib
C4bC2a kompleks. See kompleks toimib
C3 konvertaasina ja lõikab
C3
C3a valgud liiguvad verre ning passiivsed immuunrakud märkavad neid,
liiguvad põletikukohta, värbavad fagotsüüte, fagotsüüt seondub komplemendivalkude
abil bakteri külge.
C3b-dest osad seonduvad C4bC2a kompleksiga, Bakteri pinnal
muudab C3 kompleks jälle kuju, tekib
C5 konvertaas – mis on veel suurem, ehk
C4b2a3b, mis lõikab C5-te, C5b kinnitub C6le. C5b + C6 + C7 + C8, tekib superkompleks
(Membrane Attack Complex), suured valgud läbistavad membraani, tekitades tunneli,
mistõttu bakteri sisu voolab välja.
Alternatiivne tee algab komplemendi kokkupuutel mikroobi pinnaga.
Alternatiivne tee ei sõltu antikehadest – alt. tee on osa loomulikust immuunsusest. Selle
tee võivad aktiveerida
bakterite rakuseina osad. Aktivatsioon on kaskaadne ja toimub
inaktiivse ensüümi lõikamisel mitmeks aktiivseks komponendiks. Aktivatsioon algab
juhusliku C3 lagunemisega. Tekkinud aktiivset C3i stabiliseerivad võõrpinnad
(membraanid) ja inhibeerivad oma koed. Lahuses C3i hüdrolüüsub. Aktiveeritud C3b
seondub faktori B-ga. B sait vabaneb ja on substraadiks faktor D-le. C3b lõigatakse Bb-ks
faktor D poolt C3bBb lõhub C3 C3b-ks – seonduvad kovalentselt komplemendi
aktivatsiooni saidi ümber, tekitatakse sama reaktsiooni uuesti ja uuesti. Tulemuseks on
ka C3bBb3b kompleks, millel on C5 konvertaasne aktiivsus, nagu ka C4b2a3b-l, saab
jälle C5b, mis pinnale kinnitub.
Kontrollvalgud dissotseerivad C3bBb, mille järgselt Faktor
I fragmenteerib vaba C3b. Alates C5-st on aktivatsioon klassikalise teega sarnane viies
membraane atakeeriva kompleksi (MAK) tekkeni (nn aktivatsiooni lüütiline osa).
Lektiinne rada algab
MBL
seondumisega – mannose
binding lectin – mannoosi
jääkidele,
mis
on
mikroorganismide
pinnal
(salmonella, listeria, neisseria
nt). MBL on akuutse faasi valk,
toodetakse
põletikureaktsioonides. Ta efekt
on C1q-le sarnane, ka struktuur.
Peale seondumist tulevad talle
ligi MASP-1 ja MASP-2 – MBL
associated serine proteases.
Lektiini rada on väga
homoloogne klassikalise rajaga
kui seda aktiveeritakse antikeha
sõltuval moel. C1q kuulub
lektiinide perekonda, tuntakse
kui kollektiide (kollageensed
lektiinid). Sellesse valkude
perekonda kuuluvad mannaan-
seostuv lektiin (
MBL), mida
tuntakse ka kui mannaan-
seostuv valk (MBP). Seerumis esinev MBL on võimeline seostuma bakterite pinnal olevate
terminaalsete mannoosi rühmadega, peale seda on võimeline reageerima kahe seriini
proteinaasiga, mis on tuntud MASP ja MASP2, on homoloogiline C1q mõjuga C1r ja C1s-
ga. MBL-i koostoime MASP ja MASP2 on homoloogne C1q mõjuga C1r ja C1s-le, mis juhib
klassikalist rada antikeha sõltuvale aktivatsioonile. Esineb komplemendi puudulikkust. …
Lugu on sarnane klassikalisele, aga antikehi vaja pole, on siis kaasasündinud
immuunsuse tähtis osa.
11.
Autoinflammatoorsed protsessid.
Autoinflammatoorsete sündroomide korral = TNF retseptoriga seotud perioodiline
sündroom, podagra, Crohn’i tõbi - on monogeensed või polügeensed haigused
Autoinflammatoorsed protsessid tekivad liigse loomuliku immuunvastuse korral –
põhjuseks spontaanne põletik, mis mõjutab mitmeid organeid.
Põhjused võivad olla geneetilised – on nii monogeenseid kui polügeenseid. Haiguseid:
Chron’s disease, psoriaatiline artriit
Autoinflammatoorsete protsesside korral on DAMPide ja PAMPide tundlikkus ülene, või
üleaktiivne, mille tulemusena on kontrollimatu tsütokiinide vahendatud põletik.
12.
Loomuliku (sünnipärase) ja omandatud (adaptiivse) immuunsuse
põhilised erinevused, nende seosed.
Infektsioonis – IFN α/β, TNF α, IL-12 loomulik immuunsuse poolt NK vahendatud
nakatunud rakkude tapmine T-raku vahendatud nakatunud rakkude tapmine
Tunnus
Kaasasündinud immuunsus
Omandatud immuunsus
Spetsiifilisus
Tunneb ära PAMPe;
mannoosiretseptorid kõigil
samad.
suurem osa kaasasündinud
immuunsuse komponente on
olemas enne infektsiooni
puhkemist ja ei ole spetsiifilised
kindlale patogeenile
Tunneb ära mikroobseid molekule
(antigeene) ning samuti suudab
ära tunda mittemikroobseid
antigeene, areneb
immuunvastusega
Retseptorid
Kodeeritud juba sugurakkudes;
mitmekesisus väike (mustrit
äratundvad retseptorid)
NT: TLR, SR, mannoosi-
retseptorid ja fMet retseptorid
rakulised ja molekulaarseid
elemendid, mis tunnevad ära
molekulide klasse, mis on
omased sagedasti esinevatele
patogeenidele
Kodeeritud geenides, mis tekivad
geenisegmentide
rekombinatsioonil; mitmekesisus
suur
NT: TCR, Ig
Retseptorite
asetus
Mitteklonaalne. Sama rea
rakkudel on kõigil sama retseptor
Klonaalne. Lümfotsüütide kloonidel
on esialgsele sarnased retseptorid,
kuid erinevad omavahel
suurematse gruppides
Võõras/oma
vahettegemine
Kehaomaseid rakke ei tunta ära
või neil on ekspresseeritud
vastavad antigeenid.
Selektsioon, et kõrvaldada keha
enda antigeenidele seonduvate
retseptoritega lümfotsüüte. Samas
võimalik ka autoimmuunhaiguste
tekkeks.
Reageerimisae
g
Tunnid – esmane vastus
Päevad – võtab kauem aega
Reaktsioon
peale
esmanakatumi
st
Alati sama tugevusega
Teistkordsel nakatumisel
reageeritakse kiiremini
Humoraalsed
faktorid
Lüsosüüm, komplement, aktiivse
faasi valgud, tsütokiinid
antikehad
Rakud
Fagotsüüdid
(neutrofiilid,makrofaagid), NK
rakud
T-rakud
Mälu
Puudub
Olemas – aastaid olenevalt
haigustekitajast
13.
Valkude immunogeensust määravad tegurid. Antigeenid
immuunsüsteemi aktivaatoritena. Antigeenide epitoobid.
Antigeen
Antigeen on igasugune aine, mis spetsiifiliselt seondub talle vastava antikehaga või TCR-
ga ning indutseerib immuunvastust. Kõik antigeenid pole iseseisvalt võimelised
immuunvastust esile kutsuma, st nad pole alati immunogeenid (immunogeenid on
makromolekulid, mis stimuleerivad humoraalset immuunvastust). T-rakud on võimelised
reageerima ainult valgulise antigeeniga, antikehad või BCR-id reageerivad nii valgulise,
süsivesikulise ja lipiidse struktuuriga antigeenidega. Immunoloogilisi reaktsioone kutsub
esile mitte terve molekul, vaid selle teatud osa –
epitoop (determinantide arv erinevatel
ühenditel varieerub).
Antigeenid on: mikroorganismide antigeenid, autoantigeenid, alloantigeenid, allergeenid
+ superantigeenid
Hapteen – antigeeni mitte-immunogeenne osa, madalmolekulaarne aine, mis on
antikehaga seotud, aga ei kutsu ilma kandjata immuunvastust esile.
Kandja (”carrier”) – antigeeni immunostimulatoorne osa Eristatakse endogeenseid,
eksogeenseid ag ja allergeene
Antigeeni epitoop (=
antigeenne determinant) – koht antigeeni molekulis, millega
reageerib (“mille tunneb ära”) B raku retseptor (BCR, ka vastava spetsiifikaga
immunoglobuliini Fab fragment) või T-raku retseptor (TCR). – nt T raku puhul on see
antigeeni see osa, mis on MHC molekuli küljes (determinant)
T- ja B-raku epitoobid: – lineaarsed – järjestikku; ruumilised – pole järjestikku –
epitoobid
B-rakkude poolt äratuntavaid epitoope iseloomustab:
• Ehituselt: polümeeri primaarjärjestus, st lineaarsed või korduva järjestusega
determinandid; sekundaarne, tertsiaarne, kvaternaarne molekulstruktuur, st
konformatsioonilised.
• Suuruselt: antikeha Fab-fragment seob 4-8 aminohappejääki.
• Arvult: teoreetiliselt võiks iga aminohape 4-8st aminohappest olla eraldiseisev
antigeenne determinant, reaalselt aga on antigeensete determinantide arv antigeeni
kohta palju väiksem. Üldiselt on antigeensete determinantide arv piiratud antikehade
võimega neid ära tunda ja siduda.
T-rakkude poolt äratuntavaid antigeenseid determinante iseloomustab:
• Ehituselt: aminohappeline primaajärjestus. (NB T-rakud ei tunne ära polüsahhariide
ega nukleiinhappeid)
• Suuruselt: TCR seostab endaga 8-15 aminohappejääki
• Arvult: antigeensete determinantide arvu määrab MHC molekulide võime siduda
antigeene. (sellest tulenevad ka erinevused indiviidide tasemel)
Epitoope on:
1) Lineaarseid – denatureerimise käigus saab Ig seonduda veel ühe determinandiga,
mis konformatsioonilises vormis oleks ligipääsmatu
2) Konformatsioonilisi - determinant kaotatakse denaturatsiooni käigus
3) Neoantigeeniline determinant – proteolüüsi tulemusel on kättesaadav
Valkude immunogeensust määravad tegurid:
1) Fülogeneetiline kaugus
2) Molekulmass – suurem on immunogeensem
3) AH koostis
4) Struktuur
5) Epitoopide tihedus
6) Degradeeritavus
SUPERANTIGEEN - immuunsüsteemi mitte- spetsiifiline aktiveerija - bakteriaalne toksiin
või retroviiruse valk, mis stimuleerib T-helpereid läbi MHC II klassi molekuli ja TCR V
segmendi vahelise silla. Seeläbi võib aktiveeruda kuni 10 % organismis tsirkuleerivatest
T-helperitest -> toksiline shokk
LINEAARNE AG: MHC I saab siduda ainult ühe peptiidi korraga. MHC I peptiidi siduva vao
otsad on kinnised -> määrab, et peptiidi pikkus mitte suurem, kui 10 aminohappet (AH),
selle peptiidi AH kinnituvad a1-ja a2-domeenides olevate nn ankuraminohapete abil ->
sälitab paindlikkus
KONFORMATSIOONILINE AG: MHC II peptiidi siduv vagu on otseselt avatud -> võimalik
pikemad peptiidi seonduda (kuni 30 AH, aga enamik neist on 13-18 AH pikad). Sageli
need peptiidid konserveerunud sekundaarse struktuuriga
Antigeenid jagunevad T-rakkudest sõltuvateks ja T-rakkudest mitte-sõltuvateks. See
tähendab seda, et immuunvastus on T-rakk-vahendatud või mitte. T-sõltuvad antigeenid
ei stimuleeri otseselt antikehade produktsiooni, vajavad selleks T-rakkude abi. T-
sõltuvateks antigeenideks on valgud. Ehituselt on nendele antigeenidele iseloomulik
omada väheseid koopiaid mitmetest antigeensetest determinantidest (st neil on palju
antigeenseid determinante, kuid need on erinevad). T-rakkudest sõltumatud antigeenid
stimuleerivad otseselt B-rakke tootmaks antikehi. B-rakud aktiveeruvad polüklonaalselt,
st antigeen suudab aktiveerida B-raku kloone, mis on spetsiifilised ka teistele
antigeenidele. Selle järgi saab T-sõltumatud antigeenid jagada kaheks tüübiks. Tüüp I
suudab B-rakke polüklonaalselt aktiveerida, tüüp II mitte. T-sõltumatud antigeenid on
polüsahhariidid. Ehituselt on T-sõltumatutele antigeenidele iseloomulik polümeerne
struktuur. T-sõltumatud antigeenid on resistentsemad degradatsioonile, mistõttu nad
püsivad organismis kauem ja stimuleerivad immuunsüsteemi ka kauem.
Antigeeni immunogeensuse määrab:
• Keemiline koostis
• Molekulmass (> 10kD on head immunogeenid; mida suurem mass, seda intensiivsem
vastus)
• Struktuuri kompleksus (polümeerid!)
• Geneetiline erinevus (st eristus oma-võõras; samas on osa aineid immunogeensed
ühele liigile, kuid mitte teisele, sama võib täheldada ka indiviidide tasemel: Ühel
liigil/isendil võib olla puudus/muundunud mingid teatud geenid, mis kodeerivad B- või
T-rakkude retseptoreid või on vajalikud APC-del antigeenide esitlemiseks Th-
rakkudele),
• Organismi sattumise tee (tugevam immuunvastus on siis, kui ag sattub
subkutaanselt, vähem kui i/v või p/o (seedesüsteemi-intragastriaalselt-(kaudselt)))
• Lisafaktorid – adjuvandid – suurendavad antigeeni immunogeensust, kasutamine on
tihtipeale seotud soovimatute kõrvalekalletega nagu põletik ja palavik; vanus - väga
noored ja vanurid)
• doosi suurus,
• lahustuvus koevedelikes
• degradeeritavus
Üldiselt on immunogeensed antigeenid need molekulid, mis on organismile võõrad, suure
molekulmassiga (>10kDa), kompleksse ehitusega, mittelahustuvad koevedelikes,
denatureerunud (natiivses vormis pole nii immunogeensed), degradeeritavad (mida
kergem on antigeeni fagotsüteerida, seda immunogeensem ta on; nt T-rakkudele
antigeeni esitlemiseks on vaja see esmalt proteolüütiliselt töödelda APC-de poolt).
Parimad antigeenid on valgud, polüsahhariidid ja lipiidid on nõrgema
immunogeensusega.
Hapteen on molekul, mis pole eraldivõetuna immunogeenne, aga on võimeline
reageerima spetsiifilise immuunvastuse mehhanismidega, juhul kui eelnevalt on
immuunvastus esile kutsutud. Hapteenid on väiksed molekulid, mis ei kutsu ise esile
immuunvastust, välja arvatud juhul kui nad on seotud kandja-molekuliga. Hapteen-
kandja kompleks: hapteen ja kandja on omavahel kovalentselt seotud, immunogeense
osa moodustab kandja. Hapteen pole immunogeenne, küll aga antigeenne. Neil
kompleksidel on antigeenseteks determinantideks kandja natiivsed epitoobid, kuid ka
hapteeniga seostumisel tekkivad epitoobid. Antud kompleksis määrab kandja, kas
immuunvastus on T-vahendatud või mitte. Toodetud antikehad reageerivad ka vaba
hapteeniga (antigeenne omadus).
Adjuvandid: suurendavad mittespetsiifiliselt antigeeni immunogeensust, nt maarjas
(kaksiksulfaadi kristallhüdraat), alumiiniumhüdroksiid vaktsiinides. Kasutatakse eluta
vaktsiinides, eriti nendes, mis sisaldavad väiksemõõtmelisi molekule (teadagi on ju
suuremad immunogeensemad), et muuta neid immunogeensemateks. Mitmetel
mikroobsetel, sünteetilistel, endogeensetel preparaatidel (liposoomid) on adjuvandi
aktiivsus, hetkel kasutatakse inimese vaktsiinides siiski alumiinium- või kaaliumsoolasid.
Antigeenide kasutamine vaktsiinides.
Põhimõtteliselt võib vaktsiinides antigeenidena kasutada terveid organisme kuni väikeste
peptiidideni välja. Tegelikult sisaldavad vaktsiinid nõrgestatud, surmatud organisme,
inaktiveeritud toksiine (toksoide) või subtsellulaarseid fragmente. Enamik vaktsiine on
viirusvastased. Elusvaktsiinid sisaldavad nõrgestatud mikroorganisme (viiruseid). Nende
jaoks on kasvukeskkond muudetud vastavaks, et nende geenid muteeruks nii, et nad
kaotaksid oma virulentsuse, kuid säilitaks antigeensuse. Taolised vaktsiinid tekitavad
lokaliseerunud ja tugeva immuunvastuse ega vaja tavaliselt korduvvaktsineerimist.
Samas on elusvaktsiinide manustamine immuunkomprimeeritud isikutele raskendatud.
Antigeenide kasutamine vaktsiinides tähendab kunstlikku aktiivset immuniseerimist.
Elusvaktsiinid on tuulerõugete, lastehalvatuse, mumpsi, hepatiit A, kollapalaviku, leetrite,
punetiste vastu. Ainus bakteriaalne elusvaktsiin on tuberkuloosivastane (Mycobacterium
bovis). Surmatud viirusvastased vaktsiinid on lastehalvatuse, gripi, marutõve jne vastu.
Enamik bakterivastaseid vaktsiine sisaldavad surmatud mikroorganisme. Nt tüüfuse,
koolera, katku, läkaköha vastased vaktsiinid. Teised bakterite vastased vaktsiinid
sisaldavad bakterite rakuseina komponente. Nt läkaköha, meningokoki, pneumokoki
vastased vaktsiinid. Mõned viirusvastased vaktsiinid sisaldavad vektoreid (nt pärmiseen),
mis sisaldavad antigeenseid klonaalseid valke. Toksoide kasutatakse difteeria,
teetanuse, koolera vastastes vaktsiinides. Subühikulised vaktsiinid (subtsellulaarsed,
toksoidid) on loodud, et vähendada vaktsiini toksilisust. Pinnaantigeenid on kasutusel B-
hepatiidi vaktsiinis.
Superantigeenid. Antigeenid (valgud, mida toodavad mitmed patogeenid: bakterid,
mükoplasmad, viirused), mis suudavad aktiveerida suure hulga Th-rakke. Tähtsamad
superantigeenid on stafülokokkide endotoksiinid, mis võivad seostuda TCR Vβ-regiooniga
(T-raku retseptori varieeruva osa β-ahelaga). Th-rakkude aktivatsiooniks on vaja ka, et
superantigeen seostuks APC pinna MHC II klassi molekulidega. Superantigeene seejuures
ei töödelda ega esitleta APC-de poolt. Nad jäävad intaktseteks. Iga superantigeen võib
seostuda ühe või mitme Vβ-regiooniga, mida inimesel on 20-50. Th-rakkude
stimulatsiooni ulatus on funktsioon Th-rakkudest, mis kannavad Vβ-regiooni, mis võib
seostuda kindla superantigeeniga. Aktiveerunud Th-rakud toodavad tsütokiine, kuna
aktiveerub suur hulk T-rakke, siis toodetav tsütokiinide hulk on samuti väga suur, see
tingib toksilise šoki sündroomi. (loengus olen kirjutanud: aktiveerunud Th otsivad
organsimis märklaudu).
14.
Immunoglobuliinisubühiku ehitus. Fab ja Fc fragmendi immunoloogiline
tähendus. Immunoglobuliinide klassid ja alaklassid. Nende strukturaalsed
erinevused.
Antikehad on immunoglobuliinid. Antikehad on veres ja muudes kehavedelikes esinevad
globulaarsed glükoproteiinid, mille teke on indutseeritud antigeenide poolt ja mis on ise
võimelised vastavate antigeenidega reageerima nii in vitro kui ka in vivo. Antigeenid
võivad olla seotuna B raku pinnale või esineda sekreteeritud kujul: vereseerumis ja
kudedes, ka immuunkomplekside e Ag-Ak komplekside koostises. Igal inimesel võib
arvestuslikult olla kuni ca. 100 miljonit erinevat immunoglobuliini molekuli, mis on
määratletud Ig-de geeni-blokkide rekombineerumise kaudu. Kõrgematel imetajatel
esineb 5 klassi immunoglobuliine: IgG, IgA, IgM, IgD, IgE, mille erinevus seisneb
molekulide suuruses, laengus, aminohappelises ja valgulises koostises. Erinevused
esinevad mitte ainult klasside vahel, vaid ka iga klassi piiris. Normaalse vereseerumi
elektroforeesil esinevad immunoglobuliinid kõikides fraktsioonides α-st γ-
Ig-de strukturaalsed komponendid. (IgG näitel)
Peamine neljaahelaline Ig-de srtuktuurne ühik (monomeer) koosneb kahest kergest ja
kahest raskest polüpeptiidsest ahelast. Kerged (L-) ahelad koosnevad 212
aminohappejäägist, nende molekulaarmass on 25 kDa. Nad on identsed kõikidel Ig
klassidel. Rasked (H-) ahelad koosnevad 450 aminohappejäägist, nende molekulaarmas
on 50-77 kDa. Nad on erinevad Ig erinevatel klasidel ja alaklassidel.
Igal L-ahelal on 2 disulfiidsidet ja igal H-ahelal on 4 disulfiidsidet (-S-S-). Iga disulfiidside
suleb sõlmust, mis koosneb 60-70 aminohappejäägist. Peptiidsilmus, mis on suletud
peptiidsideme abil, moodustab domeeni. Domeenis on kokku u 110 aminohappejääki.
Mõlematel ahelatel (L ja H) on üks varieeruv regioon e domeen (VH, VL), mille
aminohappeline koostis varieerub, sellepärast on see regioon võimeline seonduma
erinevate antigeenidega. Hüpervarieeruv regioon - varieeruva regiooni alad, mis omavad
erilist variaabelsust, moodustavad immunoglobuliini antigeeniga seostuva ala ja
koosnevad CDR1-CDR3-st (complementarity determing regions).
Ülejäänud osa, mis paikneb varieeruvate regioonide all, on invariantse iseloomuga. See
on konstantne domeen. Kergel ahelal on üks (CL1) ja raskel ahelal on 3 või 4 (CH1, CH2,
CH3, CH4) konstantset domeeni.
Kerged ahelad on λ ja κ tüüpi. Nad seostuvad kõikide raskete ahelate tüüpidega, kuid
ühes Ig molekulis peavad olema 2 ühesugust tüüpi kerget ahelat.
Rasked ahelad määravad Ig klassi: γ, α, μ, δ, ε (IgG, IgA jne) ja subklassi : α1, α2 (IgA1,
IgA2 jne)
Raske ahela lõik CH1 ja CH2 domeenide vahel on hinge regioon. Selles regioonis
prevaleeriv osa on Cys ja Pro jääkidel - see annab hinge regioonile paindlikkust. Tänu
sellele võivad Fab regioonid funktsioneerida sõltumatult.
Ig-de funktsionaalsed osad.
Fab-regioon on antigeeniga seonduv osa. Seostumine - monovalentselt, bivalentselt,
polüvalentselt (IgM)-erinevad Ag-Ak seostumisviisid, mille abil on võimalik antigeeni
lõhustada ja organismist välja viia komplemendisüsteemi, fagotsüütide, eosinofiilide,
basofiilide, nuumrakkude abil. Siin on vajalik Fc-kaudne mitmesuguste rakkude
kaasamine Fc-retseptorite (FcR) abil.
Fc-regioon - nn kristalliseeruv osa. Funktsioon: seostumine rakkudega (Fc-retseptorid),
komplemendiga.
IgG prevaleerib inimese organismis . Vereplasmas on teda 75%. (12-14 g/l). IgG1- 65%,
IgG2- 25%, IgG3- 5%, IgG4- alla 5%. Esineb kaks kerget ja kaks rasket ahelat.
Homoloogsuse aste erinevate alaklasside vahel on väga suur (90-95%). Erinevus on
enamasti disulfiidsidemete arvus hingeregioonis. γ1 ja γ4 ahelates on kaks disulfiidsidet,
γ2 ahelas 4 ja γ3 ahelas 11.
IgA prevaleerib inimkeha sekreetides (süljes, maomahlas, limaskestade sekreetides,
rinnapiimas). Vereplasmas on tema sisaldus väike 10-15%. Raskel ahelal on 3 domeeni.
Eksisteerib 2 IgA alatüüpi IgA1 ja IgA2. IgA2 tüüp omab veel 2 alleelset varianti
(allotüüpi): A2m(1) ja A2m(2).
Suured erinevused on alatüüpide vahel hinge regioonis. α1 on 13 aminohappejäägi võrra
pikem kui α2-ahel. α1-ahelas järjestus 224-239 tekkis 8 aminohappe duplikatsiooni
tagajärjel. Süljes, jämesooles on ensüümid, mis lõhustavad just neid duplitseeritud
alasid. Mõned bakterid sünteesivad ka sellist ensüümi ja seega lõhustavad IgA1. IgA2-l
toimus deletsioon selles hinge regiooni fragmendis (nagu juba mainitud IgA2-l on raske
ahel 13 AH võrra lühem kui IgA1-l raske ahel) ja sellepärast ei allu IgA2 nende
proteolüütiliste ensüümide toimele.
IgA struktuurseteks iseärasusteks on see, et molekulis esineb J-ahel ja sekretoorne
komponent. J-ahela ülesandeks on molekuli polümerisatsioon. J-ahel interakteerub raske
ahela COOH lõpurühmaga (disulfiidsidemete osalusel). Selle tagajärjel tekivad IgA
polümeersed vormid (dimeerid, trimeerid jne)
IgM on kõige raskemini organiseeritud. Molekulaarmass on 950 kD. Pentameer. Koosneb
viiest monomeerist, igal on 2 rasket ahelat (μ) ahelat ja 2 kerget (κ või λ). Monomeerid
on seostunud disulfiidsildade abil. Monomeersed subühikud paiknevad radiaalselt Fc-
fragmendid sissepoole, Fab fragmendid väljapoole. Raskes ahelas on 4 domeeni (Cμ1,
Cμ2, Cμ3, Cμ4). Hinge regioon -ahel, mille ülesandeks on on monomeeride polümeri
IgD on monomeer, sisaldus veres 0.03 g/l. Raskes ahelas (δ) on 3 domeeni.
IgE on monomeer; molekulaarmassiga 190 kD. Organismis on teda väga vähe. Veres -
0.0005 g/l. Raske ahel (ε) sisaldab 5 domeeni.
Isotüübilised erinevused : need on immunoglobuliinide klassid ja alaklassid (nende
geenid esinevad normis kõikide ühe liigi esindajate genoomis).
Allotüübilised erinevused - kajastavad geneetilisi erinevusi ühe liigi esindajatel. (erinevad
alleelid).
Idiotüübilised erinevused - erinevus aminohappelises järjestuses varieeruvates ja
hüpervarieeruvates regioonides
15.
Immunoglobuliinide erinevate klasside bioloogiline tähendus. Primaarse
ja sekundaarse immuunvastuse iseloomustus. Primaarses ja sekundaarses
immuunvastuses tekkivate antikehade iseloomustus. Monokloonsed
antikehad.
IgM – sekreteeritakse enamike antigeenide korral esmase immuunvastusena, võimaldab
efektiivset esmast kaitset baktereemiate vastu. Sellest klassist on veregruppide
antikehad. Komplemendi sidumine, bakterite lüüs. Limaskestade kaitse IgA puudulikkuse
korral. IgM hulk veres tõuseb patoloogiliste seisundite korral. Naistel seerumi IgM nivoo
kõrgem. suur valk (makroglobuliin), mis koosneb viiest identsest monomeersest
alaühikust ja neid koos hoidvast väikesest proteiinist (J ahel). Alaühikutel on madal
afiinsus antigeeni suhtes aga tema pentameerne struktuur annab molekulile väga hea
antigeeni sidumise võime. Ekspresseeritakse enamuste B-lümfotsüütide pinnal.
IgG – domineeriv immunoglobuliin vereringluses ja koevedelikes, kus ta võitleb
mikroorganismide ja nende toksiinidega.
Kõrgem afiinsus antigeenide suhtes võrreldes
IgM-ga. Võime liikuda verest kudedesse. Läbib platsentat - kaitseb vastsündinut
infektsioonide vastu, mille suhtes ema oli immuunne. Komplemendi sidumise võime,
põhjustab bakterite lüüsi.
IgA – dimeerne vorm võimaldab esmase kaitse lokaalsete infektsioonide vastu, tänu selle
rohkusele süljes, pisarates, bronhiaalsekreetides, nasaalses limas, prostata vedelikes,
vaginaalsekreetides, nahas ja peensoole mukoosas. Limaskestade kaitse ja bakterite
lüüs. eksisteerib nii monomeerne kui ka dimeerne vorm (kaks monomeeri mida hoiab
koos J ahel). Kõigil neil antikehadel ühesugune struktuur: kaks identset rasket ja kerget
ahelat, kergel üks ja raskel kolm või neli konstantset domeeni. Mõlematel ahelatel on üks
varieeruv regioon e. domeen, mis seondub antigeeniga - komplementaarsust määravad
piirkonnad (CDR).
IgE – kaitseb keha välispindu, eelkõige mukoosaga kaetud pindasid antimikroobsete
rakkude `värbamise` teel. Kutsub esile akuutse põletikulise vastuse. IgE tase tõuseb
ussidega seotud infektsioonide ajal, kaitsedes selliste sissetungijate eest. Nuumrakkude
(basofiilide) aktivatsioon.
IgD – funktsioon ebaselge, arvatavasti toimib kui antigeeni retseptor, et kontrollida
lümfotsüütide aktivatsiooni ja inaktivatsiooni. Arvatakse, et IgD puudub mälurakkudes.
Esmane immuunvastus: Ag esitlevad mitmesugused APC-d (antigeeni esitlev rakk). Ag
kontsentratsioon suhteliselt kõrge. Latentne faas 5-10 päeva. Ak isotüüp – IgM. Ak
afiinsus ja spetsiifilisus madalad.
Teisene immuunvastus: Ag esitlevad peamiselt B-rakud. Ag kontsentratsioon madal.
Latentne faas 2-5 päeva. Ak isotüübid – IgG, IgA koeiseärasused. Ak afiinsus ja
spetsiifilisus kõrged. Ak kontsentratsioon kõrgem. Oluline roll märklaudrakkudel.
Monoklonaalsed antikehad (Mak). 1975 lõid Köhler ja Milstein hübridoomi meetodi, liites
hiire immuniseeritud põrnarakud hiire müeloomi rakkudega ning said hübriidsed rakud e
hübridoomid, mis sekreteerisid spetsiifilisi antikehi selle antigeeni vastu, mida kasutati
põrnadoonori immuniseerimiseks. Põrnarakud võimaldavad toota antikehi,
müeloomirakud võimaldavad kultuuris piiramatut kasvu ja eritada immunoglobuliine.
Hübriidrakud eraldatakse, tappes müeloomi ja põrnarakud, mis ei ole omavahel liitunud.
Hübridoomid kloonitakse ja seega on nende poolt toodetud antikehad identse
struktuuriga, kaasa arvatud nende antigeeni siduvad kohad. Selliseid antikehi
nimetatakse monoklonaalseteks. See võimaldab toota homogeenset antikehade
populatsiooni, mille antigeenne spetsiifilisus on teada. Mak`e kasutatakse seroloogilistel
ja diagnostilistel uuringutel ja terapeutiliste teguritena. Siiani on regulaarselt suudetud
toota vaid hiire Mak`e, katsed toota inimese Mak`e on saavutanud vähest edu.
16.
Sekretoorse IgA tekkemehhanism. Sekretoorse IgA iseloomustus ja
bioloogiline tähtsus.
Teda võib vaadelda kahe omavahel interakteeruva osana: a) organiseerunud koed, kus
antigeene seotakse e MALT- mucosa associated lymphoid tissues (saab jagada veel
omakorda kolmeks: GULT- gut (sool) associated lymphoid tissue, nasal (nina) ehk NALT ja
bronchial (bronhiaalne) ehk BALT (viimased on seotud respiratoorse traktiga) ja b)
difuussed lümfoidsed koed, kus vastavad rakud ja antikehad talitlevad kaitsjatena.
SIgA ja regulatoorsed T-rakud on olulisemad tolerantsuse ja kaitse tagajad limaskestadel.
Limaskestadel toimivad ka eraldi repertuaariga mujal ebatüüpilised T-rakud, mis ei
interakteeru tavalise MHC peptiidligandiga, vaid mitmete teiste ligandidega, näiteks MHC
klass IB molekulidega. Difuusne lümfoidkude: B-rakkude kloonid lahkuvad
mälurakkudena või plasmarakkude eellasrakkudena aferentsete lümfisoonte kaudu
lümfisõlmedest. Need rakud liiguvad tsirkulatsiooni ja „home back” ehk lähevad tagasi
elundite ja teiste difuusse lümfoidkoe piirkondade lamina propriasse (subepiteliaalne
sidekoekiht), kus leidub peamiselt IgA plasmarakke. See pakub kaitset teiste
mikrobiaalsete sisenemiste eest. Lamina propria sisaldab T rakke ja on rikas ka
nuumrakkude poolest. CD8+ T rakke ehk Tc-rakke on leitud epiteliaalsete rakkude
hulgas (intraepiteliaalsete rakkude) hulgas. Samuti on nuumrakud olulised, sest nad
osalevad ussnugiliste vastases kaitses. Kuigi nugiliste vastases kaitses on peamised IgE
antikehad, aitab nuumrakkude degranulatsioon tekitada ägedat põletikulist vastust, kui
toimub ussnugiliste antigeenide presentatsioon.
Lokaalsest immuunsüsteemist rääkides tuleb teda ruumiliselt lahterdada: perifeersed
lümfisõlmed ja põrn; limaskestadega assotsieerunud lümfoidkoed; serooskelmetega
kehaõõned (pleura, peritoneum); nahk. Igas lahtris algatatud immuunvastuse
efektormehhanismid toimivad vastavalt selle „lahtri” piires.
Immuunvastuse lokaalse lahterduse tagamine toimub selle kaudu, et naiivsetele T
rakkudele antigeenide esitlemisel programeeritakse T rakud igas „lahtris” erinevalt.
Seetõttu on erinevates „lahtrites” efektor-T-rakkudel erinevad adhesioonimolekulide ja
kemokiinretseptorite komplektid, mis tagavad nende rakkude koespetsiifilisemat
pesastumist. Erinevate „lahtrite” koerakud ekspresseerivad vastavaid
adhesioonimolekulide ligande ja toodavad vastavaid kemokiine. Lokaalsetes „lahtrites”
kontrollivad lümfotsüütide liikumisi koespetsiifilise adhesiooni ja kemokiinide mõjud.
Sekretoorne IgA: Kuigi osa IgA (teda on umbes 3 g/l s.h. IgA1 80% ja IgA2 20%)
antikehadest tehakse süsteemses immuunsüsteemis (see tähendab, et osaliselt luuüdis),
valmib enamik siiski plasmarakkude poolt toodetuna lamina proprias. See antikeha
transporditakse üle epiteliaalsete rakkude trakti luumenisse ehk valendikku
spetialiseerunud mehhanismi abil: IgA subepiteliaalses lamina proprias seondub IgA polü-
Ig retseptoritega ja transporditakse endosoomide kaudu luumeni ehk valendiku pinnale.
Mitte vähem kui 3g transporditakse päevas gastrointestinaaltrakti, millest 1/3 läheb
maksa ja sapi kaudu. See kaitseb mukoosseid pindu mikrobiaalse invasiooni eest.
Sekretoorset IgAd leidub pisarates, süljes, rinnapiimas, kolostrumis jne. Tänu sellele, et
teatud sekretoorne osake on seotud IgA antikehaga, väheneb proteolüütiliste ensüümide
poolt degradatsiooni oht intestinaalses keskkonnas.
IgA esineb nii monomeerses kui ka dimeerses (kahte monomeeri hoitakse koos J-ahela
abil) vormis. Monomeerne vorm on tsirkulatsioonis ja ta omab kahte alaklassi. Dimeerne
vorm osaleb primaarses kaitses lokaalsete infektsioonide vastu, olles süljes, pisarates,
bronhiaalsetes sekreetides, nina limas.
17.
Immunoglobuliinide geenide komponendid ja
rekombinatsioonimehhanismid. Immunoglobuliinide spetsiifilisuse
geneetiline alus. Immunoglobuliinide isotüüpide kujunemine seoses B-
rakkude küpsemisega. B-raku retseptori (BCR) iseloomustus.
Immunoglobuliinide geenide komponendid ja rekombinatsioonimehhanismid.
V-regioon või V-domeen (variaabelne) Ig raskes või kerges ahelas on kodeeritud paljude
geenisegmentide poolt. V-D-J segmendid ja V-J segmendid kerges ahelas Segmendid on
ühendatud intronitega. V(D)J rekombinatsiooni käigus intronid splaissitakse välja ja
juhuslikult ühendatakse segmendid. Kõigepealt V-J /V-D ja siis VD-J. V
regioonile/domäänile, mis on V(D)J rekombinatsiooni läbinud, liidetakse C ehk konstantne
ala. Igal V-regioonil on ka C-regioon (konstantne), mis hoolitseb efektori või signaali
funktsiooni eest.
Kerge ahelKerges ahelas on V-domeen kodeeritud 2 eraldatud DNA segmendi poolt. Esimene
kodeerib 95-101 aminohapet ja teda nimetatakse V-geeni segmendiks (inimesel 40) ja
teine kodeerib kuni 13 aminohapet ja teda nimetatakse ühendavaks (joining) või J- geeni
segmendiks (inimesel 5).
V ja J geeni segmendi ühinedes luuakse pidev ekson, mis kodeerib terve kerge ahela V
regiooni. V- segment asub kaugel C regioonist, aga J -segment on lähedal C regioonile.
Kui V-ja J-segmendid ühinevad, toob see ka V- segmendi C-regiooni lähedale. Et teha
terve Ig kerge ahela mRNAd, tuleb V-regiooni eksonil ühineda C-regiooni järjestusega
RNA splaissingu abil peale transkriptsiooni.
Immunoglobuliini ahelad on ekstratsellulaarsed proteiinid ja V-geeni segmendile eelneb
ekson, mis kodeerib juhtpeptiidi ja juhatab proteiini raku sekretoorsetesse käikudesse. C-
regiooni kerged ahelad on kodeeritud erinevate eksonite poolt ja ühendatud V-regiooni
eksoniga.
λ kerge ahela lookus paikneb 22. kromosoomis, κ kerge ahela lookus paikneb 2
kromosoomis. Raske ahela lookus paikneb 14. kromosoomis. Geenisegmentide koopiaid
on algses DNAs palju, mis juhuslikult valitakse rekombineerimiseks.
Raske ahel:
Raske ahela V-regioon on kodeeritud 3 geenisegmendi poolt: V (inimesel 51), J (inimesel
6) ja DH (diversity) (inimesel 27). DH asub V ja J vahel. Kui V-geeni segment ühineb DJ
järjestusse, siis tekib täielik VH ekson. C-regiooni geen on kodeeritud erinevate eksonite
poolt. C-regiooni ekson koos juhtjärjestusega splaissinguga töötlevad V-domeeni
järjestust, mis viiakse läbi raske ahela RNA transkriptsioonil. Juhtjärjestus eemaldatakse
pärast translatsiooni ja disulfiidsed sidemed polüpeptiidi ahelas on muutunud. Geeni-
segmendid genoomis
V-regiooni geenisegmendid ühinevad rekombinatsioonil:
DNA rekombinatsiooni mehhanism on sarnane nii kerge kui raske ahela puhul, kuigi üks
ühinemine vajab genereerimiseks kerge ahela geene, kaks vajavad raske ahela geene.
Kui kodeerimisjärjestus kahel geeni segmendil on
samasuunaline (kaks segmenti on
samasuunalised) DNAs, siis rekombinatsioon sisaldab
väljasopistust ja deletsiooni DNA-
st kahe geenisegmendi vahel.
Teine moodus rekombinatsiooniks toimub kahe
geenisegmendi vahel, mis on vastassuunalised. Mehhanism on peaaegu sama, aga DNA,
mis paikneb kahe geeni segmendi vahel – pööratakse ümber ja signaaljärjestused
(heptameerid ja nonameerid) jäävad sisse. Kui RSS-id on kokku pandud ja
rekombinatsioon toimub, siis lisandunud DNA ei lahku kromosoomist, aga säilitab
ümberpööratud orientatsiooni.
V(D)J rekombinatsioon
Kaks RSS järjestust asetsevad interaktsioonis koos proteiinidega, mis spetsiifiliselt
tunnevad ära õla pikkuse ja nii teostub 12/23 reegel. RAG-1 j RAG-2 ühilduvad
spetsiifiliselt RSS-dega. DNA molekulid on katki tehtud ja ühinevad uuesti erinevateks
vormideks. Heptameeri lõppjärjestus on ühinenud täpselt pea-pea vastu viisil ja on
tekkinud signaalühendus (
signal joint). V ja J segmendid, mis jäävad kromosoomi,
ühinevad kodeerumisühenduseks (
codering joint). Ensüümide kompleksi, mis üheselt
teostab somaatilist V(D)J rekombinatsiooni, nimetatakse V(D)J rekombinaasiks. RAG-1 ja
RAG-2 on rekombinatsiooni aktivatsiooni geenid, mis sisaldavad lümfotsüüdi-spetsiifilisi
rekombinaasi komponente. See paar geene on ekspresseeritud lümfotsüütidel, kuna nad
on ühinenud antigeeni retseptoritega. RAG proteiinid ei ole ainsad ensüümid. Veel on
tähtsad DNA ligase IV, DNA-PKA, KU, endonukleaasid jne. RAG-1 j RAG-2 ühilduvad
spetsiifiliselt RSS-dega. RAG-1 tunneb ära spetsiifilise nanomeeri RSS-s. Endonukleaas
aktiveerib RAG proteiini kompleksi, mis teeb kaks eraldi seisvat DNA katki 5` juurest,
jättes vabaks 3´-OH grupi. See 3`-OH grupp ründab fosfodiester-sidemeid teises
seisukohas (stand), tekib DNA ”juuksenõel.” Kaks RSS on ühinenud
signaalühenduseks. Kuna see ühendus ei ole puhas, siis tuleb see puhtaks teha, kas
RAG proteiinidega või Artemisega (so. mingi valk). DNA parandusensüümid (repair)
modifitseerivad avatud juuksenõela, eemaldades nukleotiidide (mingi sõna on siit puudu)
ja juhuslikke nukleotiide, kasutades ensüümi TdT (terminal deoxynucleotidyl
transferase). Lõpuks DNA ligaas IV ühineb protsessi lõpus, rekonstrueerides kromosoomi
sedasi, et see sisaldaks
rekombineeritud geene.
P ja N nukleotiidid segmentide liitekohtades.
Immunoglobuliinide proteiini ahelates on kolm hüpervariaabelset silmust (ingl.k. loops),
kaks on kodeeritud V geeni segmendi poolt. Kolmas – CDR3 sattus ühinedes V geeni
segmendi ja J-geeni segmendi vahele ja raske ahel on kodeeritud D geeni segmendi
poolt. Nii kerge kui raske ahela CDR3 erinevus on märgatavalt mitmekesistunud
nukleodiidide lisamises ja eemaldamises. Lisatud nukleotiidid on P- ja N-nukleotiidid.
Need on segmendid, mis on rekombinatsiooni käigus juhuslikult sattunud Ig-i. Kergel
ahelal lisatakse ja eemaldatakse nukleotiide V ja J segmendi liitmiskohta, mis vastab
CDR3-le. Raskel ahelal lisatakse ja eemaldatakse nukleotiide D segmendis, mis vastab
HV3-le.
P-nukleotiide kutsutakse sedasi, kuna nad loovad sarnaseid järjestusi, mis lisatakse
geenisegmentide algusesse või lõppu. N-nukleotide kutsutakse sedasi, sest nad on
nontemplate-encoded (ei oska sõna-sõnalt tõlkida.)
Geeni-segmentide rekombinatsiooni kontroll
Peab kontrollima, et V-geeni segment ühineks D või J-ga, mitte teiste V. Selliseks
kontrolliks on vaja järjestust. Seitsmest nukleotiidist koosnev
heptameer
5`CACAGTG3`, mis järgneb alati kodeerivale järjestusele. Järgneb mittekonserveerunud
järjestus, mida nimetakse õlaks, mis on 12 või 23 nukleotiidi pikk Õlale järgneb
nonameer, mis on teine konserveerunud plokk ja mis koosneb
9 nukleotiidist 5
´ACAAAAACC3`. Heptameer-õlg-nanomeer järjestust kutsutakse rekombinatsiooni
signaali järjestuseks e RSS. Et õiged segmendid oleks õiges järjekorras, toimub
rekombinatsiooni kontolliv süsteem RSS.
12/23 reegel: segment, mille RSS-s 12 bp õlg, tohib liituda ühe segmendiga,
mille kõrval asuvas RSS-s bp õlg.
Somaatilised hüpermutatsioonid.
Vastusena antigeeniga seondumisele koos T-rakkude signaalidega toimub B-lümfotsüüdis
lõplikult rekombineerunud Ig geeni VH ja VL osades somaatilisi punktmutatsioone, mille
tulemusena saadakse muteerunud BRC, mis seovad antigeeni paremini. Need rakud
valitakse antikehi sekreteerivate rakkude arengu lõppfaasi. Immunoglobuliinide
spetsiifilisuse geneetiline alus.
võimalik on 1011 erineva spetsiifilisusega antikeha. Selle aluseks on Ig geenide
segmentne olemus, millest iga segment kodeerib osa Ig polüpeptiidist. B-raku küpsedes
segmendid ühendatakse DNA rekombinatsioonil, mille tulemusel moodustub tervet Ig
varieeruvat osa kodeeriv geen.
Immunoglobuliinide isotüüpide kujunemine seoses B-rakkude küpsemisega.
B-lümfotsüüdid toodavad infektsioonist jagusaamiseks erilisi valkmolekule – antikehi. Iga
B-lümfotsüüt toodab antikehi, mis on võimelised ära tundma ainult üht kindlat epitoopi.
Kord edukalt rekombineerunud pika ahela muutlik osa (fragmendid V, J, D) enam B-raku
elueal ei muutu. Seevastu aga vahetatakse pika ahela konstantne osa (C) aktiveeritud B-
lümfotsüüdil vähemalt üks kord välja (nn. isotüübivahetus), et nakkuse iseloomu ja
asukoha põhjal anda edukamat immuunvastust. Esialgu toodavad imetaja B-
lümfotsüüdid IgM tüüpi antikehi, mille koosseisus on konstantne ala Cμ (mikro).
Isotüübivahetuse korral toimuvad DNA ümberkorraldused, mille tagajärjel eemaldatakse
mingi B-raku genoomse DNA-lõik ning antikeha muutliku osa kodeeriva regiooniga (VDJ)
liitub üks järgmistest konstantse osa geenidest. Cδ liitumise puhul hakatakse tootma IgD
tüüpi antkehi, Cγ puhul toodetakse IGg, Cη puhul IgE, Cα puhul IgA tüüpi antikehi.
Antikeha toimib esmalt B-lümfotsüüdi pinnal retseptorina. B raku
arengustaadiumid
Pro-B rakud (toimub raske ahela rekombinatsioon) – D-J rekombinatsioon.
Suured pre-B rakud (toimub raske ahela rekombinatsioon) – V-DJ rekombinatsioon.
Väiksed pre-B-rakud (toimub kerge ahela rekombinatsioon) – V-J rekombinatsioon –
rekombinatsioon κ geeni esimeses kromosoomis
o
Pre-BCR:
H ahela rekombinatsiooni takistamine + kerge ahela
rekombinatsiooni soodustamine + pre-B rakkude prolifereerumine + teise
kerge ahela (surrogaat) transkriptsiooni takistamine
Ebaküps B-rakk (rekombinatsiooni lõpp) – (kerge ahela rekombinatsioon) – Vl-Jl –
Membraanseoseline IgM. receptor editing,
negatiivne selektsioon
Küps B-rakk – membraanseoseline IgM ja IgD, perifeerias – aktivatsioon ka. rakk
ekspresserib μ:κ ja μ:κ. Esimene ekspressioon toimub siis, kui rekombinatsioon κ
geeni esimeses kromosoomis ja rekombinatsioon κ geeni teises kromosoomi ühinevad
produktiivselt
o
Alajaotused:
Marginaaltsooni B rakud (MZ)
Põrnas. Piiratud diversiteet. Vastavad polüsahhariidsetele
antigeenidele, toodavad loomulikke antikehi.
B1 B rakud
Limiteeritud diversiteediga retseptorid. Puutuvad kokku nn
„tavaliste“ antigeenidega
Follikulaarsed B rakud
IgD + IgM ekspressioon raku pinnal.
B-raku retseptori (BCR) iseloomustus.
B-raku pinnal olev monomeerne Ig (IgM, IgD) on BCR keskseks osaks. Omab
transmembraanset ja lühikest tsütoplamaatist osa. Membraanne Ig saadakse samadest
segmentidest kui antikehad, lisatakse ainult transmembraanne ja lühike
tsütoplasmaatiline osa. Koosneb kahest kergest ja kahest raskest ahelast, millel igal ühel
on V ja C regioon. BCR ise ei kanna signaali üle selleks on kaks teist molekuli,
Igα ja
Igβ , moodustavad koos antikeha molekuliga B-raku retseptori kompleksi.
Igα ja Igβ on
antigeeni mittespetsiifilised signaalimolekulid, mis ulatuvad nii raku sisse kui rakust
välja. Nendes mõlemas on järjestused, mida nimetatakse immunoreceptor tyrosine-
based activation motifs (ITAM). Tsütosoolis paiknevad sabad viivad signaali raku sisse,
kui B-retseptor on seostunud antigeeniga. Igα ja Igβ on seotud omavahel
disulfiidsidemetega ja on assotsieerunud raske ahelaga.
18.
T-raku retseptori (TCR) geenide komponendid ja rekombinatsiooni-
mehhanismid. TCR spetsiifilisuse geneetiline alus
TCR esineb T-raku pinnal. TCR-il on sarnane ehitus Ig Fab fragmentidega. TCR koosneb
kahest ahelast, mis on omavahel seotud disulfiidsidemetega –
α/β (TCR2) 90-95% T-
rakkudel ja
γ/δ (TCR1) ahelad 5-10% T-rakkudel. Mõlemad ahelad omavad kaht osa –
V (variaabel) regioon ja C (konstantne) regioon. Mõlemal ahelal on 3
komplementaarsust määravat piirkonda, hüpervariaabelsed alad - CDR1, CDR2, CDR3.
CDR3 on põhiline vastutaja antigeeni äratundmises. CDR1 α-ahelal seondub
peptiidi N-terminaalse osaga ning β-ahela CDR1 seondub C-terminusega. CDR2 on
vastutav MHC-molekuli äratundmise eest. TCR-i transmembraanne osa koosneb
positiivselt laetud aminohapetest, mis on olulised, et TCR saaks seostuda negatiivset
transmembraanset osa omavate koretseptoritega (nt. CD3, mis on oluline kompleks
signaali ülekandes). TCR-i tsütoplasmaatiline „saba“ on üsna lühike.
TCR-i CDR3-e rekombineerumine toimub tüümuses. α-ahela rekombinatsioon toimub VJ
rekombinatsiooni mehhanismi kaudu ning β-ahela rekombinatsioon VDJ rekombinatsiooni
mehhanismiga. Rekombinatsiooni järjekorda juhib RSS (recombination signal sequence).
(Toimub samamoodi kui Ig-del, vt. jooniseid loenguslaididelt). Somaatilist
hüpermutatsiooni ei esine eriti.
CD4 ja CD8 on TCR-i koretseptorid, mis on TCR-i spetsiifilisuse aluseks. Nad määravad
MHC klassi, millega T-rakk seostub. CD4 on monomeerne proteiin, mis seostub MHCII
klassi molekuliga. CD8 on disulfiidsidemetega seotud dimeer, mis seostub MHCII klassi
molekuliga
19.
T-raku retseptori (TCR) ehitus ning aktivatsioonisignaalide
transduktsioon T-rakus. TCRα/β ja TCRγ/δ retseptorid. Immunoloogiline
sünaps. Kahe signaali teooria.
TCR – T-cell receptor (T-raku retseptor)
Erinevalt antikehadest ei suuda T-lümfotsüüdid (TCR
+ koretseptorid) tunda rakku ümbritsevas
keskkonnas lahustunud vaba antigeeni. Antigeen
peab olema mõne teise keharaku pinnal ühenduses
erilise valguga, koesobivusantigeeniga (MHC, lühend
tuleb inglise keelest: major histocompatibility
complex).
T-lümfotsüüdi retseptor ei tunne antigeeni-
koesobivusantigeeni kompleksi ära üksi. Püsiva
ühenduse saamiseks T-lümfotsüüdi ja antigeeni
esitleva raku või T-lümfotsüüdi ja märklaudraku vahel
on vajalikud veel mitu T-raku pinnal olevat lisavalku.
Iga T-rakk kannab oma pinnal ~30 000 identset
antigeen-retseptorit (TCR). Iga T-raku retseptor on
heterodimeer,
mis
koosneb
kahest
transmembraansest glükoproteiinahelast (α/β või
γ/δ). Retseptoril eristatakse:
Ekstatsellulaarosa
Hinge regiooni
Transmembraanset osa
lühikest tsütoplasmaatiline osa
Retseptori ekstatsellulaarosa koosneb kahest variaabelsest regioonist/domeenist (N-
terminuse pool) ja kahest kontantsest regioonist/domeenist. Kummagi polüpeptiidahela
ekstatsellulaarosa igale domeenile (nii konstantsele kui variaabelsele) kinnitub üks
suhkrujääk. Retseptori ekstatsellulaarosa on sarnane antikeha Fab regiooniga. Hinge
regioon ühendab retseptori polüpeptiidahelate ekstratsellulaarosa T-raku membraaniga.
Samuti toimub selles piirkonnas TCR polüpeptiidahelate ühendamine ühe
disulfiidsidemega – side, mis formeerub kahe tsüsteiinijäägi abil. Retseptori
polüpeptiidahelate transmembraanne osa, millel on heeliksi kuju, sisaldab positiivse
laenguga aminohappeid. α–ahelas on 2 positiivselt laetud aminohapet, β-ahelas aga üks
positiivselt laetud aminohape.
Mõlemat tüüpi TCR erinevad membraanseotud immunoglobuliinist (BCR):
TCR-il on ainult üks antigeeni siduv regioon, samas BCR-il on neid kaks
Kui immunoglobuliini (BCR) sekreteeritakse antikehadena, siis TCR ei sekreteerita
kunagi.
TCR seostub antigeeniga vaid MHC vahendusel
BCR (seal hulgas antikeha) tunneb ära konformatsioonilisi epitoope antigeenil, samas
TCR vajab antigeeni eelnevat töötlust, kuna TCR tunneb ära lineaarseid epitoope.
TCR on kahte tüüpi:
α/β – seostub MHC I ja MHC II klassi molekulidega ning immuunvastuses selle
tavalises mõttes. Seda retseptorit kannavad 90-95 % T-rakkudest
γ/δ – sarnane kujult α/β retseptorile. Kuid seda tüüpi retseptor seostub spetsiifiliste
ligandidega - kuuma-šoki valkudega, mitte-peptiidsete ligandidega sh fosforüleeritud
ligandid või mükobakteriaalsed lipiidsed antigeenid. γ/δ tüüpi
retseptor ei vaja
seostumiseks antigeeniga MHC klassi molekule, vaid ta võib seostuda ka
vabade antigeenidega, aga ka mitte-klassikaliste MHC-sarnaste molekulidega.
Üldiselt ei teata veel γ/δ retseptori rolli immuunvastuses. γ/δ retseptorit kannavad 5-
10 % T-rakkudest. Saab valmis enne α β t
CDR-id TCR-is
CDR alad moodustavad TCR pinna (ülevalt vaadates)
CDR1 lingud katavad MHC seondunud peptiidi otsasid
CDR2 lingud katavad MHC molekulide α heelikseid (külgmised)
CDR3 ulatub üle peptiidi keskkoha - määrab seondumise — pannakse erinevatest
segmentidest kokku ja pannakse nt juurde.
TCR koostises on tegelikult mitu erinevat sorti retseptoreid – juba mainitud
heterodimeer (α-ahel ja β-ahel, mille variaabelne osa on antigeeni spetsiifiline)
CD3, mille 4 polüpeptiidahelat moodustavad 3 erinevat retseptori kompleksi – ε/δ ja ε/
γ heterodimeerid omavad nii ekstratsellulaarset, transmembraanset ja
intratsellulaarset osa
ζ/ζ (sigma) homodimeer, mis omab põhiliselt intratsellulaarosa, ent ζ/ζ homodimeeril
eksisteerib ka transmembraanne osa ning lühike ekstratsellulaarne osa. Igal
polüpeptiidahelal on vähemalt üks signaaljärjestusala – iminoretseptori
türosiinipõhine aktivatsiooni järjestus (iminoreceptor tyrosine based activation motif)
ehk lühendina
ITAM. ITAM piirkonnad on CD3 polüpeptiidahelate intratsellulaarsetes
osades. T-raku puhul on ITAM piirkondi kokku 10.
o ITAM motiiivid on olulised signaaliülekandes (ζ ahelatel 3 tk, CD3-l 1)
TCR kompleksi kuuluvad ka Src perekonda kuuluvad proteiin
türosiin kinaasid,
millest kõige tähtsamad on
Fynid.
CD45, mille tsütoplasmaatiline domeen sisaldab türosiin fosfataasi (ensüüm) -
Naiivsed T rakud ekspresseerivad CD45RA vormi, mälu T rakud ekspresseerivad
CD45RO vormi.
o Tsütoplasmaatiline osa omab türosinaasfosfataasi aktiivsust
defosforüleerib Lck kinaasi (see aktiivne!)
TCR koretseptorid –
CD4 või CD8. Igal T-rakul ekspresseeritakse neist ainult ühte
tüüpi! Koretseptori tsütoplasmaatilisele domeenile on seostunud samuti
türosiin
kinaas – sedapuhku tüüp Lck.
o Lck kinaas – fosforüleerib CD3 ja sigma ahelaid annab edasi T raku
aktivatsiooni signaali
Ka tsütosoolne ensüüm ZAP70 mängib olulist rolli T-raku aktivatsioonis
CD28 – kostimulatoorne molekul osaleb ka signaaliülekandes – seondub B7-1
(=CD80)/B7-2 (=CD86)-ga
CD2: (LFA-3)
Inhibeerijad on kas CTLA-4 (inhibeerib T raku aktivatsiooni – vastand CD28-le) või PD-
1 valgud
Adhesioonimolekul – LFA-1 R: ICAM1
TCR tuleb pinnale ainult siis, kui kõik kompleksi osad on olemas. Kõigepealt moodustub
TCR ER transporditakse läbi Golgi rakumbmbraaniile.
CD4 ja CD8 molekulide struktuur:
CD8 seondub MHC I külge kõrvale. mittekovalentne seondumine (valk-valk). põrkavad
erinevate rakkude oksa - kui sobib tekib tõmbumine. CD8-l α β (joonisel α α) ahelad
kaks 1 Ig ahelat.
CD4'l 4 Ig domääni – üksteise otsas
Kui T-helper aktiveeritakse, hakkavad igasugused vastused tööle. Nii rakulised kui
humoraalsed vastused.
Immunoloogiline sünaps:
Immunoloogilise sünapsi moodustumine tagab
molekulide kontsentreerumise –
korrastumine – tõmbavad lähestikku – sisaldab TCR, CD3, ζ ahelaid, CD4/CD8 ja
kostimulatoorseid nagu CD28 molekule. MHC ja TCR kompleksi molekulid
agregeerivad. Integriinid (LFA1 ICAM-1) hoiavad kompleksi koos.
peptiidi seostumine MHC I või II klassi molekuliga
MHC seostumine TCR-iga
T-helperi või T-tsütotoksilise raku aktiveerumine
patogeeni hävitamine
või B-lümfotsüüdi aktiveerimine.
Kahe signaali teooria:
T-raku aktivatsiooniks on vajalik kõigepealt TCR ja peptiidi (MHC küljes presenteerituna)
interaktsioon. Lisaks on vaja
sekundaarset
signaali, mida vahendavad
kostimulatoorsed molekulid. Kostimulatoorset signaali T-raku pinnal vahendab
CD28
molekul.
Naiivne T rakk vajab aktivatsiooniks kostimulatoorseid signaale. Mitteaktiveeritud APC
rakk ekspresseerib vähesel määral kostimulatoorseid molekule. APCde Aktivatsioon
tõstab kostimulatoorsete molekulide ekspressiooni (APC-l B7). Kõige tähtsam on T rakul
CD28 – ilma selleta ei toimu midagi.
CD28 interakteerub APC pinnal B7 (inimesel vastavalt CD80/86) molekuliga ning
alles nende kahe retseptori interaktsioon annab edasi signaali.
Selleks et dendriitrakk ekspresseeriks
B7 kõrgel tasemel, aktiveerub sünapsi tekke
tagajärjel
T-raku pinnal CD40L molekul, mis on CD40-retseptori ligandiks
dendriitraku pinnal. CD40-CD40L seondumise tagajärjel aktiveerib APC rakk oma
pinnale suurel hulgal B7 molekule, mis võimaldavad maksimaalset sekundaarset signaali.
Vastupidine toime on CTLA-4-l mis ei lase T rakul aktiveeruda. Kolmas signaal IL2/IL2R
interaktsioon, ilma selleta täismahulist aktivatsiooni ei ole võimalik saavutada.
T raku aktivatsioonisignaali transduktsioon:
TCR variaabel domeenid (α ja β ahel) seostuvad vastava MHC klassi molekuliga (see
sõltub sellest, millist koretseptorit T-rakk kannab, vastavalt kas CD4 või CD8) ning
sellega seostunud peptiidiga. Vaja APC-d (nt DC, MF, B rakk).
Koretseptori ekstatsellulaarosa (kas CD4 või CD8) seostub samuti MHC klassi
molekuliga. Selle seondumise tagajärjel nihkuvad üksteise lähedusse Lck ja ITAM
fosforüleeritud alaga seostunud ZAP70. Lck fosforüleerib ZAP70 ja seega aktiveerib
ZAP70.
Lck fosforüleerib ennast, seejärel fosforüleerib CD3 ja ζ ahela ITAM-ide türosiinijäägid
(Fosforüleeritud ITAMid on seostumiskohtadeks kinaas ZAP70-le)
o Fyn (kinaas) fosforüleerib CD3 molekule
ZAP70 seostub ζ ahela ITAM alade fosfotürosiinidele.
Aktiveeritud ZAP70 saab nüüd seostuda ja seejärel aktiveerida e fosforüleerida teisi
intratsellulaarseid signaaliülekandes osalevaid adaptiivseid molekule, nagu näiteks
LAT (linker of activation in T cells)
Fosforüüülitud LAT-le seondub
PLC-γ (fosfolipaas γ), ning mitmed adaptorvalgud,
millele omakorda seonduvad GEFid (GDP, GTP - Guanine-nucleotide exchange factors)
PLC-γ toimel vabanevad
PIP2 lõhustamise tulemusena DAG
(diatsüülglütserool) ja IP3 (inositooltrifosfaat).
IP3 tõstab rakusisest
Ca2+ kontsentratsiooni ja aktiveerib fosfataaside hulka
kuuluva
kaltsineuriini, mis aktiveerib trankriptsioonifaktori
NFAT (nuclear factor of
activated T-cell).
IP3 toimel vabanenud Ca2+ koos DAG-ga aktiveerivad
PKC (proteiini kinaas C), mis
omakorda aktiveerib transkriptsioonifaktori
NFκBB.
GEF-d aktiveerivad Rasi-GDP, mis omakorda aktiveerib
MAP kinaaside kaskaadi. Ras
algatatud kinaaside kaskaad toob kaasa Fosi sünteesi ja Fosi aktiveerimise, mis
kuulub ühe osana
AP-1 transkriptsioonifaktori koosseisu
Lõpuks – transkriptsioonifaktorid
NFκBB,
NFAT ja
AT-1 kutsuvad esile spetsiifilise
geenitranskriptsiooni (peamiselt täiesti uute geenide trankriptsiooni alustamine),
mille tulemusena T-rakk prolifereerub ja diferentseerub.
Põhiline – aktiveeritakse NFkB, NFAT ja AT-1 transkriptsioonifaktorid
20.
Peamise koesobivuskompleksi (MHC) iseloomustus. Tähendus
immuunsüsteemi funktsioneerimisel. HLA I ja II klassi antigeenid. HLA
alleelide pärilikkus ja assotsieerumine immunoloogiliste haigustega. MHC
restriktsioon T-helper ja T-tsütotoksiliste rakkude suhtes.
MHC iseloomustus
MHC haplotüüp on teatud MHC geeni alleelide kombinatsiooniks, mis sageli päranduvad
koos – iga inimene ekspresseerib mõlemalt alleelilt kodeeritavaid MHC geene. Nende
molekulide (ehk antigeenide) abil on organism võimeline eristama oma rakke võõrastest,
sest igal elusorganismil on need liigiti ja indiviiditi erinevad. Inimese puhul nimetatakse
seda kompleksi leukotsüütide antigeenide kompleksiks ehk HLA-ks (human leucocyte
antigen).
Peamine koesobivussüsteem e MHC molekulid on glükoproteiinid, mis on kodeeritud
suure geeniklastri poolt. Klassikalistel MHC molekulidel on oluline roll immunoloogilise
kompleksi moodustamisel, mis peab ilmnema T-raku ja
teiste keharakkude vahel immuunvastuse tekkimiseks.
Enamus MHC molekule täidavad oma ülesannet raku
pinnal, kus nad esitavad polüpeptiide T-rakule. Need
esitatavad polüpeptiidid võivad pärineda valgust, mis on
toodetud raku enda sees, või teise võimalusena
võõrvalgust bakterilt või viiruselt. "Täiskäigul"
immuunsüsteemi aktiveerimine nõuab tavaliselt B-raku
aktiveerimist BCRi kaudu ja T-raku aktivatsiooni MHC-TCR
kompleksi tekkel.
MHC molekulid on glükoproteiinid, mis jagunevad I ja II klassi.
MHC I molekul on heterodimeer, koosnedes rakumembraani läbivast α -ahelast ja seda
mitteläbivast β2 -mikroglobuliinist. α-ahelal on
kolm osa: ekstratsellulaarne, ning
transmembraanne osa. Ekstratsellulaarsel osal on omakorda kolm domeeni:
α1, α2 ja
α3, millest T-raku Ag-i äratundmise määravad kaks esimest. Need moodustavad nimelt
peptiidi siduva vao.
Kui MHC I molekulid on sünteesitud (seda tehakse rakus pidevalt), siis "ootavad" nad
peptiidi seondumiseni ER-s. Peptiidid lagundatakse tsütosoolis proteasoomide poolt
(proteolüütilise aktiivsusega makromolekulid, mis degradeerivad intratsellulaarseid
proteiine väiksemateks peptiidideks, mis seejärel
vabastatakse tsütosooli) tükkideks. Peptiidid liiguvad
tsütosoolist
endoplasmaatilisse retiikulumi (ER), et
nad kohtuks seal MHC I klassi molekulidega. MHC I klassi
molekul edastab info CD8+ T rakule. Kuna viirused
nakatavad rakku tsütoplasmasse sisenedes selle
tsütosooli, siis Kuna MHC I-ga esitatakse peptiide, mis on
pärit tsütosoolist, nimetatakse ka
tsütosooli või
endogeenseks rajaks. MHC I-st sõltuv antigeeni
presentatsiooni rada ongi primaarne viirus-nakatatud
raku ja T-raku vahelises signalisatsioonis. (tsütotoksilise
T rakud hävitavad sihtmärkraku)
MHC II molekuli mõlemal ahelal on kolm osa: ekstra- ja intratsellulaarne ning
transmembraanne osa. Rakuvälised osad moodustavad omakorda neli domeeni: α1, α2
ja β1, β2. Ag-i siduva vao moodustavad α1ja β1. MHC II molekulid genereeritakse
happelistes endotsüütilistes vesiikulites.
MHC II-sõltuvat antigeeni presentatsiooni rada nimetatakse ka endotsütootiliseks või
endogeenseks rajaks. Selleks siseneb ekstratsellulaarne patogeen endotsütoosi teel
rakku, kus ta seeditakse lüsosüümides. Invariant clipi abil hoitakse MHC II koos – muidu
laguneb ära. Seejärel MHC II-ga seotuna transporditakse proteiin raku
plasmamembraanile. MHC II klassi molekul edastab info rakule, millel on CD4 ja TCR e. T
helperile (Th-rakk). T helper aitab tagada vajaliku immuunvastuse tekitades lokaalset
põletikku (makrofaagid) ja aktiveerides B-lümfotsüüte. MHC II klassi molekulid on olulised
bakterite kui ekstratsellulaarsete patogeenide hävitamisel.
MHC funktsioon:
MHC molekulide ekspressiooni intensiivsus rakkude pinnal omab suurt tähtsust T-
rakulises immuunvastuses, sest need on võimelised Ag-ga seostuma vaid
MHC-
vahendatult. Eriti oluline on aga nende molekulide hulk, mistõttu on organismis välja
kujunenud erilised MHC II molekulide ekspressiooni mehhanismid. Näiteks INF-γ
suurendab tavaliselt
makrofaagidel ja dendriitrakkudel olevat MHC molekulide
arvu ning on samas võimeline esile kutsuma MHC II ekspressiooni selliste rakkude
pinnal, kus need tavaliselt puuduvad (nt endoteeli-, epiteeli- ja endokriinrakud).
Vajadusel muutuvad need rakud Ag esitlevateks rakkudeks, olles normaalselt MHC II
negatiivsed. B-lümfotsüütidel suureneb MHC II hulk IL-4 toimel. MHC I klassi molekulide
ekspressiooni suurendavad INF-α, INF-β , INF-γ ja TNF.
MHC seondumine peptiidiga pole nii spetsiifiline. Kuna peab mahutama igast peptiide,
aga on steerilise piiranguga – st iga peptiid, mida esitatakse, peab sobima MHC
molekuliga – MHC-ga seonduvatel peptiididel on sarnaseid
ankur AH-sid – nende
kaudu kinnituvad. Peptiidid kinnituvad ankurpositsioonide kaudu (need MHC küljes).
Alasid, kus valk MHC-ga seondub, nimetatakse epitoopidesk – klass I ja II erinevad.
Peptiid on lõdvalt - seondub aeglaselt ja eemaldub veel aeglasemalt (pole spetsiifiline).
MHC ei tee vahet oma ja võõraste peptiidide vahel!
MHC restriktsioon T-helper ja T-tsütotoksiliste rakkude suhtes
MHC I (α3) saab seonduda ainult CD8+ T rakkudega (tsütotoksilised)
MHC I klassi molekulid ekspresseeruvad
peaaegu kõigi tuumaga rakkude ja ka
trombotsüütide pinnal. Rakkudeks, kus neid molekule ei ole või on vähe on näiteks
KNS-i rakud, cornea ja enamus endokriinrakke (k.a. pankrease B-rakud).
Klass I ekspressioonitase tõuseb
tsütokiinide ja
interferoonide stimulatsiooni
tagajärjel
MHC II (β2) saab seonduda ainult CD4+ T rakkudega (helper)
MHC II molekulid ekspresseeruvad APC-de (B-rakkude, aktiveeritud T-rakkude,
monotsüütide, makrofaagide ja dendriitrakkude) pinnal.
Klass II stimuleeritakse IFN γ ja TNF α-ga. Oluline transkriptsiooniline regulaator on
CIITA
T-rakud tunnevad ära ainult peptiide. Ning nad on peptiidi järjestustele spetsiifilised –
tunnevad ainult MHC küljes olevaid peptiide! ST et kui antigeen on vabalt ringlev, ei
tunne T rakk teda ära.
MHC I ja II saavad antigeeni ka koospresenteerida. Nt viiruse antigeenid klass II-ga ja kui
viiruse antigeen tsütosoolis, klass I-ga ja viirus-spetsiifilise CD8+ T rakuga T-rakuline
vastus.
Antigeeni esitavad rakud
Klass II
Kostimulatoorne
signaal
Funktsioon
Dendriitrak
ud
Konstitutiivne,
IFN γ tõstab
Konstitutiivne, IFN γ
tõstab,
ka
CD40CD40L
Initsieerib T raku!ise vastuse,
aktiveerides naiivseid T rakke
Makrofaagi
d
Madal, IFN γ
tõstab
Indutseeritav LPSi,
IFN γ ja CD40-CD40L
Effektor T rakkude aktivatsioon
B rakud
Konstitutiivne,
11--4 tôstab
Indutseeritud
T
rakkude
poolt,
CD40-CD40L
Humoraalne immuunvastus
Endoteelira
kud
Konstitutiivne,
Indut-seeritav
IFN-γ-ga
Konstitutiivne
Võimendavad T raku kohale
kutsumist
ja
vastust
antigeenide suhtes
HLA I ja II klassi antigeenid:
Kontrollimatu MHC II ekspressioon rakkude pinnal võib põhjustada autoimmuunseid
haigusi. HLA geenid paiknevad 6. kromosoomis ja kodeerivad kolme klassi molekule (HLA
I-III). HLA III geenid kodeerivad mõningaid
komplemendi komponente, kuumašoki valke ja
TNF.
Inimesel:
Klass I geenid on HLA-A, HLA-B ja HLA-C
Klass II geenid on HLA-DR, HLA-DQ ja HLA-DP
Hiirel:
Klass I geenid on H-2K, H2-D ja H2-L
Klass II geenid on I-A ja I-E (Ir immune response)
Erinevad HLA alleelid määravad indiviidi võime immuunvastuseks, vastuvõtu või
resistentsuse nakkushaigustele, autoimmuunhaigustele ja allergiatele.
Iga inimene saab kummaltki vanemalt ühe HLA geenikompleksi => genoomis on 2 HLA
kompleksi, ning HLA geenid avalduvad kodominantselt. HLA geenilookus on polügeenne
ja polümorfne -> St igal indiviidil ekspresseeruvad mõlemad, nii emalt kui ka isalt saadud
HLA geeni haplotüübid. Iga I ja II klassi HLA geeni lookused on populatsioonis esindatud
mitmete alleelsete variatsioonidena, kuna igas geenilookuses sisalduvad veel omakorda
erinevad geenid. Seega on HLA polümorfne. S.t, et sel on liigisiseselt palju alleele. See on
oluline seepärast, et siis saab isend esitleda laia valikut Ag-e ja tõenäolisemalt tagada
efektiivset immuunvastust.
HLA I ja II klassi alleelid määravad ära peptiidid, mida esitatakse immuunsüsteemile,
kusjuures peptiidi sobivuse konkreetse HLA-alleeliga määravad ära selle peptiidi
vastavas positsioonis olevad aminohapped.
HLA roll haiguste tekkes on seotud selle teatud alleelidega: Näiteks HLA-B27 alleeliga
geeni omavatel inimestel on 90 korda suurem tõenäosus haigestuda anküloseerivasse
spondülartriiti kui neil, kellel vastavat alleeli pole (juhul kui nad muus osas on sarnased).
Nimelt esitab see alleel peptiide, mis on pärit lülisamba kudedest, kuid sarnanevad
nimetatud haigust põhjustava veel kindlakstegemata patogeeni peptiididega, muutes
seega lülisamba antigeensed struktuurid immuunreaktsiooni märklauaks.
Teisi häireid:
MHC I defititsiit seotud TAP-geeni mutatsiooniga, mille tagajärjel puuduvad
immuunsüsteemis Tc-rakud. Selle tagajärjel suureneb vastuvõtlikkus kroonilisele
kopsu- ja nahapõletikule.
MHC II defitsiit on seotud MHC II molekulide endi puudumisega, mistõttu puuduvad
Th-rakud ja organismi iseloomustab üldine tundlikkus.
MHC I-ga on seotud näiteks herpesviirus, mis infitseerib epiteeli ja levib sensoorse
neuronini, kus persisteerub latentselt. Latentne viirus toodab vähe proteiine ja neid
presenteeritakse MHC I kaudu minimaalselt. Kuna neuronite pinnal on MHC I molekule
liiga vähe, siis tunnevad Tc-rakud neid halvasti ära. Seega võib madal MHC tase olla
neuronitele kasulik vältimaks nende destruktsiooni, kuid muudab nad samas ka
vastuvõtlikuks infektsioonidele. Tsütomegaloviiruse (TCV) korral on vähenenud MHC I
ekspressioon ja viirusvalgud "nüristavad" immuunvastuse. (loeng 10, slaid 26,28)
Teised MHC-ga seotud häired, mille korral spetsiifiline viga on teadmata, on näiteks
üldine varieeruv immuundefitsiit (defektne IgA, IgG produktsioon; esineb tundlikkus
ekstratsellulaarsete bakterite suhtes) ja selektiivne IgA puudulikkus (puudub IgA
süntees; tundlikkus hingamisteede infektsioonide suhtes). (loeng 7, slaidid 29, 30)
MHC II on seotud ka autoimmuunhaigustega, kus Ag ekspresseeritakse koos MHC II
molekuliga rakkudel, kus seda tavaliselt ei esine.
MHC I ja II alleelid DR3/DQ2 ja DR4/DQ8 on seotud I tüüpi (insuliinsõltuva) diabeedi
(DR2 alleeli puhul väiksem risk) ja tsöliaakiaga (siin lisaks ka HLA alleelid DQ2 ja B8).
HLA serotüübiga seotud tundlikkus autoimmuunhaigustele: DR4 - reumatoidartiit, DR3
- süsteemne erütematoosne luupus ja Gravesi tõbi (loeng11, slaid 21)
Reproduktsiooni ülesanne on suurendada järglaste HLA polümorfsust! Ehk mida
erinevad on meie vanemad, seda paremini jääme meie ellu infektsioonide vastu.
Platsenta koosneb pool ema ja pool isa rakkudest. Ema ja loote vereringe ei ole
segatud, aga loote veresooned on ümber uhatud pidevalt ema vereringega. Nende
vahel on platsentaarbarjäär. Keksmine platsenta kiht on HLA negatiivne, et ema
immuunsüsteem ei näeks isa HLA-sid. Kõige emapoolsem kannab mitteklassikalisi
HLA I klassi molekule. Lootepoolses osas loote enda klassikalised HLA I ja II klassi
molekulid.
Lokaalne platsentaarbarjäär: aktiivselt toodetakse põletikuvastust reguleerivaid
tsütokiine, HLA ekspressiooni eripärad, trofoblasti rakk saab ise alustada põletiku,
mitteküpsed dendriitrakud, mis fagotsüteerivad pidevalt „prahti“. Perifeerne
platsentaarbarjäär: pidevalt lekib perifeeriasse antigeene, sellega koos läheb ka
isapoolne HLA molekul. Trofoblasti eksosoomid sisaldavad igasugu miRNAsid ja isa
HLA-sid. Ajutiselt tekib isapoolsetele HLA-dele tolerants ning ajutine ema rakulise
immuunsuse langus.
21.
Antigeeni esitavate rakkude ja T-rakkude vahelised signaalmolekulid.
“Kahe signaali teooria”. Immunoloogiline sünaps. eespool
22.
Tüümuse ehitus. T-rakkude küpsemise põhimõtted ja tüümuse roll selles.
Tüümus on immuunsussüsteemi primaarne organ. Tüümus koosneb sagarikest, mille
välist osa nimetatakse kooreks ning sisemist osa säsiks. Säsi osad on omavahel
ühenduses säsiväätide abil, milles kulgevad veresooned ja närvid. Tüümus muutub
väiksemaks vanusega ning kahaneb, asendudes rasvarakkudega. Koores peamiselt
tümotsüütide varajased arengufaasid. Medullas küpsevad. Tüümus on keskkond, kus
valitsevad tümotsüütide proliferatsiooniks ja küpsemiseks vajalikud stiimulid. Need
stiimulid tulevad tüümuses olevatest dendriitrakkudelt, luuüdist pärinevatelt
makrofaagidelt ja tüümuse epiteelirakkudelt
Teised olulised rakud on tüümuse epiteeli rakud (koore ja säsi ehk
kortikaalsed ja
medullaarsed), epiteliaalsed tugirakud (epithelial nurse cells) ja dendriitrakud.
T-lümfotsüütide prekursorid pärinevad maksast (lootel) või luuüdist (täiskasvanul) ning
migreeruvad sealt veresooni pidi tüümuse koore sisse. Tümotsüütide migratsioon ja
küpsemine toimub
koorest – säsi suunas. Algselt on koores tegemist pro-T rakkudega
(=
double negative), millel ei ekspresseerita TCR-i, CD4 ega CD8. Koores migreerudes
küpseb antud rakk pre-T rakuks (=
double negative), mis endiselt ei ekspresseeri CD4+
ja CD8+, kuid milles on rekombinatsiooni tulemusena raku pinnale ekspresseeritud
TCR
β- ahel ja pre-α ahel.
Edasi säsi poole migreerudes areneb rakk
double positive rakuks, mille membraani
pinnal on ekspresseeritud
dimeerne TCR ning CD4+ ja CD8+. Sellele järgneb
positiivne (liigub edasi kui on piisavalt tugev signaal „kallistus“ enda antigeenide vastu)
ja negatiivne selektsioon (DP T cells bone marrow derived antigeenidele) tüümuses ning
rakud, mis selektsiooni läbivad küpsevad
single positive tümotsüütideks, mis
ekspresseerivad
CD4+ või CD8+. Küpsed rakud liiguvad seejärel
perifeersetesse
kudedesse.
Kui T rakk tüümuses antigeeniga ei kohtu (MHC kaudu), siis ta sureb.
Positiivne selektsioon määrab, kas temast saab CD4+ või CD8+ T rakk (MHC II vs I
määrab selle).
Tüvirakk pro-T pre-T DP SP küps T rakk
CD8- CD4- - double negatiivne
CD8+CD4+ - double positiive (perifeerias ei esine, ainult tüümuses)
CD8+/CD4+ - single positive (kui liigub tüümusest välja)
naiivne küps T-rakk
(Antud T-rakkude küpsemise põhimõte on αβ T-rakkude põhjal kirja pandud. αβ
retseptoreid omavad T-rakud moodustavad u 95% kogu T-rakkudest)
23.
Tüümuse roll organismi enese kudede vastase tolerantsuse kujunemisel.
AIRE ja tema tähendus.Tüümuses toimub nn tsentraalse tolerantsuse kujunemine - T-rakkude positiivse ja
negatiivse selektsiooni kaudu. Positiivse selektsiooni käigus elimineeritakse T-rakud, mis
ei tunne ära MHC molekule. Samas ei pääse positiivsest selektsioonist läbi T-rakud, mis
MHC-peptiid kompleksiga
liiga tugevasti seonduvad ehk oluline on seondumise tugevus.
Negatiivse selektsiooni käigus elimineeritakse autoreaktiivsete retseptoritega
(retseptorid, mis tunnevad ära kehaomaseid antigeene) T-rakud, mille eemaldamine on
oluline, et tagada organismi enese kudede vastane tolerantsus. Negatiivne selektsioonil
osutub määravaks antigeeni seondumise tugevus T-rakuga, liiga tugev signaal viib raku
apotooosi. Ehk siis perifeeriasse pääsevad vaid need T-rakud, mis tunnevad ära oma
MHC-peptiid molekule kuid ei oma olulist afiinsust self peptiidi suhtes.
AIRE – autoimmuunregulaator, mis ekspresseerub tüümuses. Tänu AIRE’le
ekspresseeruvad tüümuses kehaomased antigeenid ning tänu sellele saab tüümuses
toimuda T-rakkude negatiivne selektsioon. Mutatsioonid AIRE geenid võivad esile
kutsuda
autoimmuunhaigusi, kuna tekivad autoreaktiivsed rakud.
24.
Immunoloogiline tolerantsus (tsentraalne ja perifeerne), selle kujunemise
mehhanismid.
Immuunsüsteemi funktsioon: kaitse haigustekitajate eest (neid elimineerides);
tolerantsus oma kudede suhtes (tüvirakkudest genereeritud T ja B lümfotsüütide abil;
TCR ja BCR oluline roll); st immuunsüsteemi põhiülesanne on eristada oma ja võõrast.
Immuuntolerantsus – mehhanismid, mis tagavad antigeenspetsiifilise adaptiivse
immuunvastuse (B ja T-rakud) vältimise või supresseerimise kehaomaste antigeenide
ehk potensiaalsete autoantigeenide suhtes kahjutute võõrsubstantside (näit
kommensaalsed toidubakterid, toiduantigeenid) suhtes.
Tsentraalne tolerantsus – arenevate (ebaküpsete) lümfotsüütide positiivne ja
negatiivne selektsioon primaarsetes lümfoidorganites:
o T-rakud tüümuses
Autoantigeeni ära tundmisel kas rakusurm/retseptori muutmine /või ka
T-regiks muutumine – saab tugeva signaali aga ei sure ära.
o B-rakud luuüdis
Mitte autoreaktiivne B rakk (läbi retseptori muutmise selliseks, mis ei
reageeri) või/ deletsioon (läbi apoptoosi) / anergia (reduced signaling,
reduced receptor expression)
Perifeerne tolerantsus: immunoloogiline ignorantsus, autoreaktiivsete
lümfotsüütide anergia või deletsioon sekundaarsetes lümfoidorganites, regulatoorsete
T-rakkude poolt vahendatud supressioon.
o Anergia/apoptoos/Tregide poolt supressioon
Anergia – signaal blokeeritakse, T rakk ei reageeri – või on tegu
inhibitoorsete retseptoritega
Kostimulatoorse signaali puudumine on oluline anergia tekkeks
o Tregide poolt supressioon – FOXP3 transkripsioonifaktori mõjul tekivad Tregid,
mis inhibeerivad teisi rakke – kas rakk-rakk kontaktide või tsütokiinide abil (IL-
10, TGF-β)
o AICD: Fas ja FasL ekspressioon T raku pinnal viib selle surmani
o B rakkudel: anergia (inaktivatsioon), deletsioon (apoptoos), inhibitoorsete
retseptorite läbi regulatsioon
Kui naiivne T rakk kohtab esimene kord antigeeni ilma kostimulatoorset signaali (B7-
CD28), võib T rakk jääda antigeeni suhtes anergiliseks
Tümotsüüdi positiivne ja negatiivne selektsioon on klonaalse deletsiooni
aluseks
Positiivne selektsioon: valitakse T rakud, mis tunnevad ära MHC
Negatiivne selektsioon: deleteeritakse T rakud, mis reageerivad tugevalt enda
antigeeni suhtes
Tolerantsuse säilitamine:
Klonaalne deletsioon (tsentraalne tolerantsus) (Tüümus - T ja luuüdi - B)
o - primaarsed immuunorganid o - kõige suurem tolerantsus
Klonaalne regulatsioon (enamasti perifeerne tolerantsus)
o - kõik mis jääb välja tüümusest ja luuüdist
o Kostimulatoorse signaali puudumine
o Anergia: inaktiivsus
o Regulatoorsed T rakud – Treg CD4+ (FoxP3 transkriptsioonifaktorit kasutavad)
– määrab regulatoorse võime, mõjuvad teistele T-rakkudele, suruvad aktiivsust
maha
Ignorantsus
o Priviligeeritud koed (peaaju – seal pole, CD8 lähevad Alzheimeri puhul ajju ja
tekitavad põletikku; silmamuna sisemus; testis)
o Peidetud antigeenid (midagi MHC-st)
T-rakk vajab elamiseks kallistust. Kui ei saa, sureb ära (apoptoos), kui kallistus on liiga
tugev, sureb ka ära.
Kallistus = TCR kaudu signaal – kui ei sobi MHC-ga kokku – ei saa. TEC (thymic
epithelial cells), cTEC (core), mTEC (medulla), DC.
Signaal liiga tugev = reageerivad enda valkudega, enda antigeenidega. (
negatiivne
selektsioon, anergia). agonist selektsioon: Treg, CD8 α α + IEL-id (intraepiteliaalsed
lümfotsüüdid)
, NKT rakud
Signaal paras: lahkuvad õnnelikuna tüümusest – 3% algmaterjalist. - positiivne
selektsioon cTEC rakud. (algul positiivne negatiivne medullas). Reageerib signaali
tugevusele
⇒
Antigeeni afiinsus ja signaali tugevus määrab äara tümotsüütide
selektsiooni.
Aire defect causes escape of tissues specific self-reactive T-cells from thymus – tekib
autoimmuunsus
Autoimmuunsus – immuunreaktsioonide esinemine organismi enese antigeenide
(autoantigeenide) vastu. Selle tekkes on oluliseim tolerantsus mehhanismide häirumine.
Autoimmuunne reaktsioon võib areneda autoimmuunseks haiguseks.
Autoimmuunhaigus: reaktsioon omaenda kudede/rakkude vastu.
Lisa nt tüümuses tekkiva tolerantsuse ja B-rakkude tolerantsuse teket.
T rakud
B rakud
Koht
Tüümus
BM
Tolerantsus
sensitiivsus
DP: CD4+CD8+
Immatuurne IgM+IgD-
Stimulatsioonid
Tsentraalne:
Tugev signaal
Tugev signaal, multivalentne
antigeen
Perifeerne:
Korduv
presentatsioon või
T rakulise abi puudumine
ilma
kostimulatsioonita
Peamine
mehhanism
Tsentraalne:
Klonaalne deletsioon Klonaalne deletsioon,
retseptori ümberkorraldus
Perifeerne:
Anergia, AICD, Tregs Anergia (signaaliblokk),
väljajäämine follikulitest
25.
Immuunreaktsioonides osalevate rakkude tsirkulatsiooni tagavad
mehhanismid.
Vere- ja lümfiringe tagab selle, et immuunrakud saavad organismis tsirkuleeruda. Lisaks
on olulised erinevad molekulid (selektiinid, integriinid, kemokiinid jne).
Neutrofiilide ja monotsüütide tsirkulatsioon
Infektsiooni puudumise korral tsirkuleerivad neutrofiilid ja monotsüüdid veres ja ei
migreeru kudedesse. Kui infektsioon tekib, toimub mitmeastmelise protsessi tulemusena
neutrofiilide ja monotstüütide adhesioon endoteeli seinale ning migratsioon läbi selle.
Kõigepealt produtseeritakse vastusena infektsioonile tsütokiine (IL-1 ja TNF), mille
tulemusena hakatakse endoteeli rakkudel tootma adhesiooni molekule (nt kemokiine,
selektiini). Selle tagajärjel neutrofiilid ja monotsüüdid adheeruvad endoteeli seinale ning
algab rakkude rullumine endoteeli seinal (selektiini retseptor endoteeli seinal ja selektiini
ligand leukotsüütide pinnal). Seejärel seonduvad leukotsüüdid integriini molekulide
kaudu endoteeli IgA molekulidega, sidumist suurendavad kemokiinid. Kemokiine
toodetakse infektsiooni kohas ning reguleerivad leukotsüütide transporti veresoone
seinast infektsiooni saiti.
Lümfotsüütide tsirkulatsioon
Naiivsed lümfotsüüdid liiguvad vereringest lümfisõlmedesse ja lümfisoonte kaudu tagasi
vereringesse. Nii toimub see mitmeid kordi kuni nad kohtuvad lümfisõlmes spetsiifilise
antigeeniga, mille nad ära tunnevad. Sellist naiivsete lümfotsüütide tsirkulatsiooni nim
lümfotsüütide retsirkulatsiooniks. Retsirkulatsioon võimaldab lümfotsüütidel leida üle
kogu keha endale spetsiifilist antigeeni. Lümfotsüüdid, mis on spetsiifilise antigeeniga
reageerinud ja diferentseerunud efektor- ja mälurakkudeks liiguvad tagasi vereringesse
ning migreeruvad sinna, kus on parasjagu infektsioon ja/või põletik. Mõned lümfotsüüdid
migreeruvad kindlatesse kudedesse, nt nahka. Sellist selektiivset protsessi nim
lümfotsüütide ”homing”. Erinevate homing mustrite olemasolu tagab selle, et
konkreetsed lümfotsüüdid liiguksid sinna koesse, kus on neile sobiv mikrokeskkond
adaptiivse immuunvastuse esile kutsumiseks.
Liikumine veresoontest lümfisõlmedesse: sarnaselt neutrofiilide ja monotsüütidega.
Lümfotsüütide pinnal ekspresseeritud adhesiooni molekule nimetatakse sageli homing
retseptoriteks. Retseptorid endoteeli pinnal millega homing retseptorid interakteeruvad
nim aadressiinideks (ingl k addressins).
26.
Omandatud immuunsuse iseloomujooned (spetsiifilisus,
“iTmmunoloogiline mälu” jm) ja tähendus. Omandatud immuunsuse
kujunemisel osalevad rakud ja signaalmolekulid. Omandatud immuunsuse
kujunemise põhilised etapid. Tagasiside-mehhanism.
Immuunvastuse tekkemehhanism
Omandatud immuunvastuse iseloomujooned:
"oma" ja "võõra" eristamine (mitteohtliku/ohtliku eristamine) - [tegelikult teeb seda
suures osas juba loomuliku immuunsuse alla liigitatav APC rakk]
kõrge (kitsas) spetsiifilisus - [pole absoluutne]
"mälu" - [antigeeni püsimine või pikaealiste mälurakkude teke]
Adaptiivse immuunvastuse tekkimise eelduseks on:
Naiivsete B- ja T rakkude pidev toomine primaarsete immuunorganite poolt ja
perifeeriasse saatmine
Lümfotsüütide aktivatsioon ja klonaalne selektsioon (kontakt „võõra“ antigeeniga
APC-de kaudu
Autoreaktiivsete T-lümfotsüütide deletsioon toimub põhiliselt juba tüümuses.
Klooni anergia - lümfotsüütide klooni olek inaktiivses seisundis; aktivatsiooniks vajalik
lisasignaal.
Esmaseks aktiveeruvaks rakuks on peamiselt
T-lümfotsüüt (aktiveerijaks valgulise
antigeeni epitoop APC pinnal), harva B-lümfotsüüt (siis vahetuks aktiveerijaks
polüepitoopsed antigeenid - näiteks endotoksiinid).
Lümfotsüütide pidev tsirkulatsioon versus "homing". Lümfotsüütidel "homing"
("pesastumis") retseptorid. Tagab, et lümfotsüüdid liiguksid sinna, kus on aktiivne kolle.
Endoteliaalsetes veenulites vaskulaarsed adressiinid.
Immuunvastuse etapid:
1. antigeeni töötlus ("protsessing") antigeeni esitava raku (APC) poolt
a. APC- (antigen-presenting cell):
i. dendriitrakk (epidermaalne Langerhansi rakk jt) kannab antigeeni
kudedest perifeersetesse lümfoidorganitesse (samal ajal toimub raku
"küpsemine"). NB! Dendriitrakkudel (DC1 ja DC2) on keskne roll kogu
immuunvastuse seadistamisel (st kas tekib eelistatult rakuline või
antikehade poolt vahendatud immuunvastus)
ii. makrofaag (sh. erinevates kudedes erinev nimi)
iii. B-lümfotsüüt
2. antigeeni (õigemini: antigeense peptiidi) esitlus Th-rakule toimub läbi II klassi HLA
(hiirel H-2) molekuli. Lisaks eksogeensele antigeeni "protsessingule" ja esitlusele (HLA
II molekuli kaudu Th-le) võib toimuda ka endogeenne antigeeni "protsessing" ja
esitlus (HLA I molekuli kaudu Tc-le) - antigeenide lõhustumine proteasoomides,
peptiidide transport raku pinnale TAP1 ja TAP2 (transporters associated with antigen
processing) abil. Hiljuti avastati, et Tc-le võidakse antigeen esitleda nii HLA I kui ka
HLA II kaudu
3. Th aktiveerumine: Th-raku aktivatsiooniks on vaja kahte signaali:
a. signaal MHC (HLA) ja TCR sideme kaudu
b. signaal CD80/CD86 ja CD28 (CTLA-4) sideme kaudu (või teiste
molekulpaaride kaudu)
c. Teine signaal võib toimuda ka CD40 ja CD40L (B-raku ja T-raku kontakti
korral) või ICAM-1 (CD54) ja LFA-1 kaudu. APC ja Th raku vahelist kontakti ja
stimulatsioonisignaali tugevdavad samuti muud adhesioonimolekulid.
Aktivatsiooni tulemus:
TCR+ / CD28+ Th raku aktiveerumine
TCR+ / CD28- Th anergia
TCR- / CD28+ efektita
CD40L geneetiline defekt - hüper-IgM sündroom - B-rakud stimuleeritud ilma Th
vahenduseta.
Aktivatsioonisignaalid kanduvad Th-raku sisemusse proteiini kinaaside (fyn, lck, ZAP-70)
kaudu
27.
Omandatud immuunsuse efektormehhanismid (mikroorganismi ja
antigeeni kahjutuks tegemise ja elimineerimise võimalikud mehhanismid).
Eelmine küss?
Erinevatel T rakkudel on erinevad signatuursed tsütokiinid.
Th1 rakud toodavad IFN-γ, mis aktiveerib makrofaage ja IgG produktsiooni.
Th2 rakud toodavad IL-4, IL-5 ja IL-13, mis aktiveerivad nuumrakke, eosinofiile ja IgE
produktsiooni. Need on tähtsad helmintide vastases kaitses. Praegusel ajal kui parasiite
eriti pole, on nad allergia mediaatorid.
Th17 rakud toodavad IL-17A, IL-17F ja IL-22, mis aktiveerivad neutrofiile ja monotsüüte.
Tsütotoksilise T raku toime
Vajalik TCR-MHC I peptiidi kompleks
LFA-1 ja ICAM-1 toetab (adhesioonimolekulid)
Moodustub sünaps ja CTL aktiveerub
Märklaudraku hävitamine –
graanulite eksotsütoos
o Perforiin aitab gransüümidel paremini rakku pääseda
o Saab ka tappa Fas/Fas L vahendusel – FasL CTL-il interakteerub Fasiga
märklaudrakul.
Fs aktiveerib DISC kompleksi ja kaspaaside kaskaadi ⇒ rakusurm
28.
Tsütokiinide iseloomustus ja toime põhimõtted. Tsütokiinide tähendus
loomuliku ja adaptiivse immuunsuse reaktsioonides, lümfotsüütide kasvu
ning diferentseerumise kontrollis.
Tsütokiinide iseloomustus ja toime põhimõtted.
Tsütokiin on üldnimetus kõigile immuunsüsteemi rakkudevahelistele mediaatoritele, mis
tagavad rakkudevahelise kontakti immuunreaktsiooni, põletiku ja reparatsiooni korral.
Tsütokiinid on väiksed, umbes 25 kDa glükoproteiinid, mida sekreteerivad keha mitmed
rakud reeglina vastusena aktiveerivale stiimulile. Mõjutatavates rakkudes kutsuvad
tsütokiinid spetsiifilise retseptoriga seostudes esile vastuseid. Rakupinna spetsiifiliste
kõrge afiinsusega retseptoritega seostumise tõttu stimuleerivad tsütokiinid rakkude
diferentseerumist, proliferatsiooni ja funktsionaalset aktiivsust. Tsütokiinid võivad
toimida autokriinselt või parakriinselt; mõned tsütokiinid on ka endokriinseks toimimiseks
piisavalt stabiilsed.
Nad esinevad vabal kujul, rakumembraaniga (retseptori kaudu ja ilma) või
valkkompleksidesse (näiteks autoantikehadega) seotult. Oluline on märkida, et
tsütokiinide toime on
lühiajaline ja
piiratud: tsütokiinide preformeeritud molekulidena
ei säilitata, süntees toimub lühiajalise transkriptsiooni ja kiiresti hävitatava mRNA kaudu.
Erinevalt hormoonidest on tsütokiinide toime lokaalne. Neid produtseerivad eri liiki
rakud, kuid kõige sagedamini leukotsüüdid. Tsütokiinid avaldavad toimet vastavate
rakuretseptoritega seostudes, kusjuures ühel rakuliigil on retseptorid mitme tsütokiini
jaoks.
Mis on tsütokiinide ülesanded?
1. Vereloome
2. Loomuliku ja omandatud immuunsuse aktivatsioon
3. Lümfotsüütide kasvu ja diferentseerumise kontroll immuunreaktsioonide korral
a. T-helperite polarisatsioon kalduvus teatud tüüpi immuunkaitse ja patoloogia
eelsoodumusele
4. Mõju teiste kudede rakkudele (närvikude, endokriinsüsteem, haavad (TGF-β)
Tsütokiinide toime üldpõhimõtted on:
Üks tsütokiin,
mitu märklaudrakku ja funktsiooni. Tsütokiine toodavad nii
loomuliku kui ka adaptiivse immuunsuse rakud.
Nii lokaalne (auto- ja parakriinne – auto – tagasimõju teda tootnud rakule, parakriinne
– kutsub esile teiste tsütokiinide tootmist omakorda) kui süsteemne efekt
Erinevate tsütokiinide antagonistlik ja sünergistlik kaasmõju
o Tsütokiinide sünergistlik toime – kaks või enam tsütokiini mõjuvad
koos
o Antagonistlik toime – vastupidine toime.
Tsütokiinid omavad toimet ka teiste organsüsteemide rakkudesse
Rakulised vastused tsütokiinidele on reeglina aeglased (tundides), kuna vastus eeldab
mRNA ja valgu sünteesimist.
Tsütokiinide tähendus loomuliku ja adaptiivsetes immuunreaktsioonides.
Kui patogeen suudab läbida epiteelibarjääri ning koes paljunema hakata, tuntakse ta
reeglina ära makrofaagide poolt. Teine tähtis fagotsütoosivõimeliste rakkude perekond
on neutrofiilid (või polümorfonukleaarsed neutrofiilsed leukotsüüdid), mida leidub
rohkesti veres, kuid normis kudedes mitte. Mõlemad fagotsütoosivõimeliste rakkude
grupid mängivad võtmerolli loomuliku immuunsuse reaktsioonides.
Esmalt tunneb makrofaag mikroorganismi ära oma pinnaretseptoritega, mis on
võimelised eristama oma pinnamolekule võõrast. Need retseptorid on mannoosiretseptor
(ainult makrofaagidel), scavenger-retseptor, mis seostub lipoteihhoidhappele, CD14
lipopolüsahhariidiga seostumiseks. Mikroorganismi seostumisel makrofaagidega
hakkavad MF vabastama tsütokiine, tekitamaks koes põletikuseisundit ning meelitamaks
infektsioonikoldesse neutrofiile ja plasmavalke. Arvatakse, et mikroorganism indutseerib
tsütokiinide-kemokiinide sekretsiooni retseptorite kaudu, millele ta seostub.
Lisaks on retseptoritel veel roll, indutseerimaks
kostimulatoorsete molekulide
ekspressiooni makrofaagidel ja dendriitrakkudel – antigeeni esitlevatel rakkudel on nii
võimalus initsieerida adaptiivse immuunsuse reaktsioone.
Põletik.
Põletikul on infektsioonivastases võitluses kolm üliolulist rolli:
(a) tuua kohale uusi efektormolekule ja –rakke, võimendamaks sisse tungivate
mikroorganismide tapmist;
(b) luua füüsiline barjäär mikrovaskulaarse koagulatsiooni näol, takistamaks infektsiooni
levikut vereringesse;
(c) edendada kahjustatud koe parandamist (täiesti mitteimmunoloogiline nähtus).
Põletiku korral tekib kudedes esmalt suurenenud, kuid samas aeglustunud lokaalne
verevoolutus veresoone diameetri tõusu kaudu (milles punasus ja temperatuuritõus).
Teiseks hakkavad endoteelirakud ekspresseerima adhesioonimolekule, mis soodustavad
ringlevate leukotsüütide seostumist. Need kaks efekti kokku võimaldavad leukotsüütide
ekstravasatsiooni. Loetletud muutusi põhjustavad aktiveeritud makrofaagide toodetud
tsüto- ja kemokiinid.
Tsütokiinide tähendus lümfotsüütide kasvu ning diferentseerumise kontrollis.
Varaseimad lümfotsüütide prekursorrakud ekspresseerivad pinnal
IL-7 retseptorit, mis
on hädavajalik T-rakkude kasvuks ja ellujäämiseks inimesel ja hiirel, B-rakkude
säilimiseks hiirel.
Lümfotsüütide arengut ja organiseerumist perifeerses lümfoidkoes kontrollivad TNF
perekonna molekulid. B- ja T-rakkudel on erisugused retseptorite komplektid, mis
reageerivad kemokiinidele, mille sekretsioon T- ja B-tsoonis on erinev. Lisaks sõltub
lümfisõlmede areng TNF-perekonna molekulide ekspressioonist. B- ja T-rakkude täpne
lokalisatsioon (homing) perifeerses lümfoidkoes sõltub strooma- ja luuüdist pärit rakkude
toodetud kemokiinidest. Vastselt perifeeriasse emigreerunud lümfotsüüdid peavad
läbima perifeerse tolerantsuse mehhanismid, elimineerimaks (inaktiveerimaks)
autoreaktiivseid rakke nende autoantigeenide suhtes, mida tsentraalselt ei
ekspresseeritud. Perifeersete tolerantsusmehhanismide seas on ka tsütokiinidest
sõltuvad regulatsioonimehhanismid.
Dendriitrakud võivad perifeersetes kudedes üsna kaua oma tähetundi oodata.
Infektsiooni ilmnemisel stimuleeritakse nende migratsiooni kohalikku lümfoidkoesse, kus
nad enam ei suuda antigeeni fagotsüteerida, küll aga ekspresseerivad suurt hulka
pikaealisi MHC I ja II klassi molekule, mis võimaldavad patogeenide proteiini esitleda.
Pärast antigeeni haaramist toimuvaks küpsemiseks ja migratsiooniks annavad signaale
tsütokiinid (või otsene kontakt patogeeni pinnaga).
Küpsed dendriitrakud
sekreteerivad naiivseid T-rakke meelitavat kemokiini DC-CK (
CCL18), mis esineb ainult
lümfoidkoe dendriitrakkudel.
Hematopoietiinide perekonda kuuluvad kõik järgmised
IL-märgistatud tsütokiinid.
Ülipüüdlikele päheõppimiseks, ülejäänud võiksid ehk
IL-2 ja IL-7 teada ja on rohkem
kui küll. IL-2 on tähtsaim T-rakkude proliferatsiooni mõjutav tsütokiin, nimetatakse ka T-
raku kasvufaktoriks. IL-2 indutseerib T-rakkude prolifereerumist, teda toodab aktiveeritud
T-rakk.
IL-1 – aktiveerib T-rakke
IL-2 – stimuleerib antigeenide poolt aktiveeritud T- ja B-rakkude proliferatsiooni
IL-4-6 funktsioonide hulgas on B-rakkude aktiveerimine.
IL-4: Th0 Th2
IL-6 toimib T- ja B-rakkude kasvule ja diferentseerumisele.
IL-7 funktsioon on pre-T- ja pre-B-rakkude kasvu mõjutamine.
IL-9 stimuleerib TH2.
IL-12 (MF) – Th0 Th1
IL-13 aktiveerib B-rakkude kasvu ja diferentseerumist, samas TH1 rakkude tootmist
pärssides.
IL-21 indutseerib B, T ja NK-rakkude proliferatsiooni.
INF-γ – aktiveerib makrofaage
IL-3, IL-7 ja GM-CSF – stimuleerivad hematopoeesi.
Mälurakkude säilimist ja funktsioneerimist kontrollivad mitmed tsütokiinid, peamiselt IL-7
ja/või IL-15, kombinatsioonis T-raku retseptori signaalidega.
TNF-perekonda kuuluvatest tsütokiinidest aktiveerivad CD40 ligandid B-rakke, CD27
ligand stimuleerib T-rakkude ja CD30 ligand T- ja B-rakkude proliferatsiooni, APRIL, BlyS
B-rakkude proliferatsiooni.
TH1 ja TH2 inhibeerivad teineteist vastastikku IFNγ, IL-4, ja IL-10-ga. IFNγ toodavad TH1-
(Tc-rakud, NK-rakud), IL-4 TH2-rakud (nuumrakud) ja IL-10 TH2-rakud (makrofaagid). TH1
sekreteerib makrofaagide aktiveerimiseks IFNγ ning TNFβ. Nimetatud tsütokiinid
toimivad ka B-rakke inhibeerivalt.
TH2 sekreteerib B-rakkude aktiveerimiseks IL-4 ja IL-6 ning makrofaagide aktivatsiooni
vältimiseks, TH1 aktiveerimiseks IL-10.
29.
Tsütokiinide määramise põhilised in vitro meetodid.
CD4+ T-rakkude funktsioon hõlmab reeglina antigeeni kandvate rakkude aktiveerimist,
mitte aga tapmist. CD4 T-rakkude aktiveerivat toimet B-rakkudele ja makrofaagidele
vahendavad enamasti mittespetsiifilised vahendajavalgud tsütokiinid, mida T-rakk
vabastab vastusena antigeeni äratundmisele. Seega hinnatakse CD4 rakkude
funktsioonivõimet reeglina
sekreteeritud tsütokiinide hulga ja tüübi järgi.
Tsütokiine saab määrata nende aktiivsuse järgi bioloogilistes
rakukasvu analüüsides,
kus nad toimivad kas kasvufaktorite või –inhibiitoritena.
Spetsiifilisem on
ELISA modifikatsioon sandwich ELISA, milles tsütokiini
iseloomustatakse tema võime kaudu siduda kaht erinevat monoklonaalset antikeha, mis
tsütokiini erinevate epitoopidega seostuvad. Tsütokiin-spetsiifiline antikeha on kinnitatud
plastikplaadile, sellega seostub tsütokiin, millega omakorda seostub teine antikeha.
T-raku vastuste uurimiseks on kasutusel ka
ELISA modifikatsioon – ELISPOT. T-
raku populatsioone stimuleeritakse meid huvitava antigeeniga ning asetatakse siis
tsütokiinidele vastavate antikehadega kaetud plastikplaadile. Kui aktiveeritud T-rakk
tsütokiini sekreteerib, seotakse ta plaadile kinnitatud antikeha külge. Pärast teatud aega
eemaldatakse rakud plaadilt, lisatakse teine ports tsütokiin-spetsiifilisi ensüüm-
märgistatud antikehi, paljastamaks tsütokiinide ringe plaadil iga aktiveeritud T-raku
positsiooni ümber. Seejärel loetakse täpid (ringid) kokku; teades algset T-rakkude arvu,
saab arvutada antud tsütokiini sekreteerivate T-rakkude sageduse. ELISPOT-i saab
kasutada ka spetsiifiliste antikehade sekretsiooni määramiseks B-rakkude poolt,
kasutades antigeen-kaetud pindu ja anti-immunoglobuliine.
Sandwich ELISA ja ELISPOT väldivad tsütokiinide bioanalüüsimise suurimat probleemi –
erinevate tsütokiinide võimet analüüsis sama reaktsiooni esile kutsuda.
Teatud tsütokiini tootvaid rakke on võimalik tuvastada ka
rakkude
fluorestsentsmärgistamisel anti-tsütokiinse monoklonaalse antikehaga.
Märgistatud rakud eristatakse fluorestsents-aktiveeritud
rakusorteriga (FACS,
fluorescence-activated cell sorter; rakkude jagamise võimega voolutsütomeeter).
Tsütokiinide tootmist on võimalik määrata ka
tsütokiinide mRNA olemasolu ja hulga
järgi
aktiveeritud T-rakkudes. mRNA-d saab määrata üksikute rakkude puhul in situ
hübridisatsiooniga,
rakupopulatsioonides
pöördtranskriptaasi
polümeraasi
ahelreaktsioonil (
RT-PCR). RT-PCR-il isoleeritakse rakust mRNA, mille baasil
sünteesitakse pöördtranskriptaasi abil cDNA. Soovitud DNA-lõiku paljundatakse PCR-il,
kasutades järjestus-spetsiifilisi praimereid. Reaktsiooni saadusi geelelektroforeesil
lahutades saadakse spetsiifilise kujuga bändid. Amplifitseerunud DNA-järjestuse hulk
vastab tema esindatusele mRNA-s: aktiivselt teatud tsütokiini tootvates T-rakkudes on
hulgaliselt konkreetset mRNA-d, seega ka vastavalt suur hulk DNA-d RT-PCR-il.
Tsütokiini-mRNA taseme hindamiseks algses koes võrreldakse saadud tulemust RT-PCR-il
saadud kõigis rakkudes ekspresseeritud
housekeeping-geenide mRNA hulgaga.
Tsütokiinide tootmise määramise probleem ühe raku tasandil seisneb selles, et T-rakud
eritavad tsütokiini ümbritsevasse ainesse ning igasugune kontakt teda tootnud rakuga
kaob. Üksikute rakkude toodetud tsütokiinide määramiseks on kaks meetodit:
rakusisene tsütokiinide märkimine (intracellular cytokine staining) ja
tsütokiinide vangistamine (cytokine capture).
Rakusisene tsütokiinide märkimine on meetod, mis põhineb valkude rakust
väljutamist pärssivate metaboolsete mürkide kasutamisel. Tulemusena kuhjub
tsütokiin endoplasmaatilisse retiikulumi, vesiikulivõrgustikku. Kui järgnevalt rakud
fikseerida, kindlustamaks valkude säilumine järgneval töötlemisel, ning pehmete
detergentidega antikehadele läbitavaks muuta, saavad antikehad võimaluse tungida
rakusiseste ruumideni (kompartmentideni) ning ära tuntu märgistada. Antikehad on
eelnevalt fluorokroom-märgistatud. T-rakke samaaegselt teiste märgistega märkides,
saab näiteks leida IL-10 tootvate CD25+ CD4 T-rakkude sageduse. Meetodi
puuduseks on see, et uuritavad rakud tapetakse.
Tsütokiinide vangistamine on meetod, mis kasutab hübriidantikehi, milles on seotud
kaks erinevat kergete ja raskete ahelate paari erinevatest antikehadest. Saadud
antikehal on seega kaks antigeeni siduvat saiti erinevate ligandide jaoks. Selliseid
bispetsiifilisi antikehi kasutatakse tsütokiinide tootmise määramiseks: üks antigeeni
siduv sait seostub spetsiifilise T-raku pinnamarkeriga, teine uuritava tsütokiiniga. Kui
rakk antud tsütokiini sekreteerib, haarab antikeha selle enda külge enne, kui see
jõuab raku pinnalt eemale difundeeruda. Detekteerimiseks lisatakse fluorokroom-
märgistatud teine, tsütokiinspetsiifiline antikeha; märgistatud rakke võib siis eristada
näiteks voolutsütomeetriaga. Meetodi eelis eelmisega võrreldes on see, et kasutada
saab elavaid rakke.
30.
Interferoonide (I, II, III klass) iseloomustus ja nende tähendus erinevate
immunopatoloogiliste protsesside korral.
Interferoonid on eriti tähtsad tsütokiinid, mis limiteerivad teatud viirusinfektsioonide
levikut inimorganismis.
Tüüp I: Üks interferoonide grupp (IFN-α ja IFN-β) sekreteeritakse viirus-infitseeritud
rakkude poolt. See interferoonide rühm on loomuliku immuunsuse osa. Sekretsiooni
keskne stimulaator on dsRNA (kaheahelaline RNA, mis on viiruse genoomne või tekib
infektsioonitsükli käigus).
IFN-α on kodeeritud 20-st geenist koosneva geenide perekonna poolt, mis asub 9.
kromosoomis. Molekul on monomeerne ja koosneb 166-st aminohappejäägist. Selle
molekuli retseptoriteks on CD 118 ja IFNAR-2. IFN-α produtseerivad leukotsüüdid ja
dendriitraud.
IFN-β on kodeeritud ainukese geeni poolt 9. kromosoomis. Molekul on samuti
monomeerne ja koosneb 166-st aminohappejäägist. Selle molekuli retseptoriteks on
CD118 ja IFNAR-2. IFN-β produtseerivad fibroblastid.
Tüüp II:
IFN-γ e „immuuninterferoon“ – sekreteeritakse makrofaagide, NK rakkude ja
lümfotsüütide poolt, mitte ainult viirusinfektsioonide korral. Ta on adaptiivse
immuunsuse osa.
Funktsioonid.
IFN-α/β produktsioon aktiveerib viirusvastaseid mehhanisme kõrvalolevates rakkudes.
Interferoonid aktiveerivad hulk geene, millest 2 on viirusvastase aktiivsusega:
1) Geen, mis kodeerib proteiini kinaasi, mis inhibeerib eIF-2 fosforüülimist. See eIF-2 on
valgu translatsiooni initsiatsiooni faktor, st selle mehhanismiga pärsitakse
valgutranslatsiooni;
2) Geen, mis kodeerib 2’,5’-oligoadenülaadi süntetaasi, mis aktiveerib latentset
endonukleaasi, mis omakorda degradeerib RNA-d.
IFN-γ ja IFN-α/β stimuleerivad MHC I ja II klassi molekulide suurenenud produktsiooni ning
mõlemad need interferoonid on võimelised aktiveerima makrofaage ja NK rakke.
IFN-γ’l ei ole mitte ainult viirusinfektsioonide vastase immuunsuse roll, tal on ka hulk teisi
funktsioone. Nimelt:
Indutseerib (koos TNF-α ja IL-1ga) põletikuregioonides mõningaid endoteeli
adhesioonimolekule, nimelt ICAM-1 ja VCAM-1.
Põletiku regioonis võivad lümfotsüüdid moduleerida hilisemat lümfotsüütide
migratsiooni vabastades kemokiine ja inflammatoorseid/põletiku tsütokiine, just
IFN-γ’t.
Naiivsed CD4+ T-rakud, mis on aktiveeritud IL-12 ja IFN-γ juuresolekul, on
sunnitud diferentseeruma Th1 rakkudeks.
IFN-γ manustamine koos antigeeniga suurendab antikehade afiinsust
antigeenidele.
NK rakud vabastades IFN-γ’t aktiveerivad makrofaage.
IFN-γ, mis on produtseeritud Th1 rakkude poolt, inhibeerib Th2 rakkude
proliferatsiooni ja kutsub esile makrofaagide aktivatsiooni.
Tüüp III -
IFN-λ1,
IFN-λ2,
IFN-λ3,
IFN-λ4
31.
Vereloome kasvufaktorid, hematopoeesi stimuleerivad tsütokiinid ja
nende tähendus lümfotsüütide arengus.
Tsütokiinid üldjuhul stimuleerivad hematopoeetilistesse rakuliinidesse kuuluvate rakkude
proliferatsiooni ja diferentseerumist või osalevad organismi immuun- ja põletikulise
vastuse tekkega seotud mehhanismides. Hematopoeesi mõjutavad tsütokiinid on näiteks
erütropoietiin (EPO), mis stimuleerib punaste vereliblede teket, trombopoietiin, mis 20
stimuleerib megakarüotsüütide rakuliinidesse kuuluvate rakkude teket, ja
granulotsüütide kolooniaid stimuleeriv faktor (G-CSF), mis stimuleerib neutrofiilide teket.
Need tsütokiinid on kasulikud vereliblede normaalse taseme taastamisel patsientidel, kes
põevad aneemiat, trombotsütopeeniat ja neutropeeniat või kes saavad vähktõve vastast
keemiaravi.
Kasvufaktorid:
CSF-id – colony stimulating factors
Nim kolooniaid stimuleerivateks faktoriteks (CSF). Granulotsüütide (G-CSF),
monotsüütide (M-CSF), granulotsüütide-monotsüütide (GM-CSF) kasvufaktorit
toodavad immuunvastuse käigus
aktiveeritud T-rakud, makrofaagid,
endoteelirakud ja lüüdi stroomarakud. Nende toimel
suureneb immuunrakkude
produktsioon lüüdis, lisaks kiirendab lüüdi rakkude diferentseerumist
dendriitrakkudeks ja monotsüütideks. Põletikukoldes toodetud
G-CSF stimuleerib
rakkude väljapaiskamist lüüdist.
Veel kasvufaktorid EPO (erütrotsüüt), TPO (trombotsüüt), G-CSF (neutrofiil), IL-5
(eosinofiil), tüvirakk + SCF → pluripotentne tüvirakk + IL-7 → (lümfoidne tüvirakk +
IL-15 (NK)/ IL-7+ IL-4 (B rakk)/ IL-7 + IL-2 (T rakk)
IL-7: stimuleerib lümgotsütaarse ellasraku elulemust ja paljunemist. Toodavad lüüdi
stroomarakud.
IL-3: universaalne CSF paljudele rakkudele.
32.
Tsütokiinide retseptorid ja signaalrajad.
Erinevate tsütokiinide retseptorid on struktuurilt üsna varieeruvad, kuid peaaegu kõik
kuuluvad ühte viiest retseptorite
perekonnast:
Kõik tsütokiini retseptorid
koosnevad vähemalt ühest
transmembraansest valgust, mille
rakuväline
osa
seondub
tsütokiiniga ja rakusisene osa
algatab signaaliraja.
Immunoglobuliini
superperekonna
retseptorid (IL-1 perekond)
o IL-1, IL-18
o M-CSF
o C-Kit
Tüüp I tsütokiini retseptorid ( ehk hematopoietiini retseptorid) – JAK-STAT
signaalirada
o IL-2, IL-4, IL-9, IL-15, IL-21
o IL-6, IL-7,
o IL-3, IL-5,
o IL-11, IL-12, IL-13
o GM-CSF, G-CSF
Tüüp II tsütokiini retseptorite perekond (ehk interferooni retseptorid) – JAK-STAT
signaalirada
o IFN-α, IFN-β, IFN
-γ, IL-10, IL-22
TNF retseptorite perekond – NF-κBB signaalirada
o TNF-α, TNF-β,
o CD40, FasL,
o BAFF, jne
Kemokiini retseptorid – G valgu vahendatud
o IL-8,
o CCR1, CXCR4
Enamus tsütokiini retseptoritest kuulub klassi I või II.
Paljud tsütokiin-seonduvad retseptorid, mis esinevad immuun- ja hematopoeetilistes
süsteemides, kuuluvad klass I tsütokiini retseptorite hulka. Nendel retseptoritel on
konserveerunud aminohappelised järjestused, mis koosnevad
neljast tsüsteiinist
(CCCC) ja konserveerunud järjestusest trüptofaan-seriin-(juhuslik aminohape)-
trüptofaan-seriin (
WSXWS), kus X on mittekonserveerunud aminohape.
Klass II tsütokiinide retseptoritel on
konserveerunud CCCC järjestused,
kuid WSXWS
järjestused puuduvad.
Signaalrajad
Algavad – ligand-retseptori seondumise järgse retseptori klasterdumisega, (mitu
retseptori molekuli liiguvad lähemale)
JAK-STAT
NF-κBBG-valk vahendatud (MAP kinaas)
RAS/RAC/CDC42/RHO
MAPKKK (MAP kinaaside kinaaside kinaas)
MAPKK
(MAP kinaasi kinaas)
MAP kinaas
transkriptsioonifaktorid (nt AP-1)
33.
Tähtsamate loomulikku immuunsust mõjutavate tsütokiinide toimerajad.
Cytokine
Principal Cell Source
Principal Cellular Targets and Biologic Effects
Tumor
necrosis
factor (TNF)
(α,β)
Macrophages, T cells Endothelial cells: activation (inflammation,
coagulation) Neutrophils: activation
Hypothalamus: fever
Liver: synthesis of acute-phase proteins
Muscle, fat: catabolism (cachexia)
Many cell types: apoptosis
Interleukin-
1 (IL-1)
Macrophages,
endothelial cells, some
epithelial cells
Endothelial cells: activation (inflammation,
coagulation) Hypothalamus: fever Liver:
synthesis of acute-phase proteins
Chemokine
s (see Table
3-2)
Macrophages,
endothelial cells, T
cells, fibroblasts,
platelets
Leukocytes: chemotaxis, activation; migration
into tissues
Interleukin-
12 (IL-12)
Macrophages,
dendritic cells
T cells: differentiation NK cells and T cells: IFN-
y synthesis, increased cytotoxic activity
Type I
interferons
(IFN-o., IFN-
ß)
IFN-o: macrophages
IFN-B: fibroblasts
All cells: antiviral state, increased class I MHC
expression NK cells: activation
Interleukin-
10 (IL-10)
Macrophages, T cells
(mainly regulatory T
cells)
Macrophages, dendritic cells: inhibition of IL-
12 production and expression of costimulators
and class II MHC molecules
Interleukin- Macrophages,
Liver: synthesis of acute-phase proteins B
6 (IL-6)
endothelial cells, T
cells
cells: proliferation of antibody-producing cells
Interleukin-
15 (IL-15)
Macrophages, others NK cells: proliferation T cells: proliferation
(memory CD8' cells)
Interleukin-
18 (IL-18)
Macrophages
NK cells and T cells: IFN-y synthesis
Interleukin-
23 (IL-23)
Macrophages and
dendritic cells
T cells: maintenance of IL-17-producing T cells
Interleukin-
27 (IL-27)
Macrophages and
dendritic cells
T cells: Tw1 differentiation; inhibition of Tw1
cells NK cells: IFN-y synthesis
34.
Tähtsamate omandatud immuunsust mõjutavad tsütokiinide toimerajad.
Tähtsamad tsütokiinid omandatud immuunsuses:
1. IL-2 – toodavad T rakud – kasvufaktor T-rakkudele – põhjustab T rakkude klonaalset
ekspansiooni pärast antigeeni äratundmist, samuti stimuleerib B rakkude
proliferatsiooni ning aktiveerib NK rakke ja monotsüüte.
2. IL-4 – peamiselt toodavad seda Th2 rakud – stimuleerib B rakkude proliferatsiooni ja
diferentseerumist, indutseerib B rakke tootma IgE, tõstab MHC II ekspressiooni
professionaalsetel APCdel.
a. IL-4 on vajalik IgE produktsiooniks
3. IL-5 – peamiselt toodavad seda Th2 rakud – stimuleerib B rakkude ja eosinofiilide
proliferatsiooni, samuti aktiveerib eosinofiile.
i. Vajalik L-5 eosinofiilide aktivatsiooniks – eosinofiilid tunnevad ära IgE-ga
kaetud parasiidid.
4. IL-10 – toodavad makrofaagid ja T rakud – inhibeerib aktiveeritud makrofaage ja
dendriitrakke; IL-12, MHC II ja kostimulaatorite inhibitsioon
5. TGF-β – toodavad antigeeniga stimuleeritud T rakud –
inhibeerib T rakkude
proliferatsiooni ja diferentseerumist ja makrofaagide aktiveerumist, stimuleerib IgA
tootmist
a. Tregide ja Th17 diferentseerumine
6. IFN-γ – toodavad efektor T rakud ja NK rakud – stimuleerib TNF-α ja IL-12
produktsiooni – TAGASISIDE MEHHANISM
a. Aktiveerib makrofaage, B rakkude klassivahetus, Th1 diferentseerumine, MHC I
ja II ekspressiooni tõstmine, antigeeni esitlemine T rakkudele ↗
35.
Kemokiinide ja nende retseptorite üldine iseloomustus, tähendus
rakuliste immuunreaktsioonide kujunemises.
Kemokiinid on tsütokiinide perekond, mis stimuleerivad immuunrakkude liikumist ja
reguleerivad leukotsüütide migratsiooni verest kudedesse. („Chemotactic cytokine“).
Võtmereaktsioon rakkude migratsioonis on
rakkude kinnitumine (sulandamine,
adhesioon) põletikus oleva koe veresoonte endoteelile. See toimub tänu erilistele
molekulidele leukotsüütide pinnal ja aktiveeritud endoteliaalsete rakkude vastastikulisele
mõjule. Koe sisse tungides migreeruvad rakud põletiku piirkonda, mis omakorda toimub
tänu sihipärasele keemilisele külgetõmbele –
kemotaksisele.
Kemoatraktantsus tähendab, et nad indutseerivad kemotaksist sellesse piirkonda
(mida rohkem neid molekule on, seda rohkem leukotsüüte sinna külge tõmmatakse). Nad
määravad ära, milline rakk
kinnitub endoteelile ja läbib selle ning
kuhu ta koes
liigub. Mõned kemokiinid ainult aktiveerivad rakke, teised omavad
kemotaktilist
iseloomu, kolmandad aga omavad mõlemat funktsiooni.
Kemokiinid, mis on vabastatud fagotsüütide ja dendriit-rakkude poolt ja on
ekspresseritud endoteeli pinnal, meelitavad rakke
verest põletiku piirkondadesse
(recruitment). Nad võivad olla vabastatud paljude raku- tüüpide poolt. Mõned
kemokiinid osalevad lümfotsüütide arengus ja migratsioonis ning angiogeneesis (uute
veresoonte kasv).
Erinevad kemokiini retseptorite kombinatsioonid, mis
leukotsüütide pinnal ekspresseeruvad, võimaldavad kindlaid migratsiooniteid
rakkudele
Kemokiinil on kaks sidumiskohta: spetsiaalne sidumiskoht kemokiini retseptori tarvis ning
kinnitumaks endoteelile (suhkruahelate kaudu). Kemokiinid jagunevad nelja perekonda
vastavalt konserveerunud tsüsteiini (C) jäägile:
CC
o CCL4 (T rakud)
o CCL2,3,5,7,8,12,13,15 – leukotsüüdi
o CCR7 T rakkudel (nii h kui ctl) kutsub
lümfisõlme
CXC
o Neutrofiilid, efektor T rakud
XC – XCL1 – T rakud, NK rakud
CX3C (=CXXXC) - mixed
* X on aminohape (tavaliselt tripeptiid).
Aminohappeline järjestus seab kõik kemokiinid sugulusse. Erinevad perekonnad
interakteeruvad erinevate retseptoritega, seega on ka nende toime spesiifiline. Nende
perekondadele vastavad retseptorid: CCR 1-9, CXCR 1-6 jne.
Kemokiini retseptor
paikneb leukotsüütide pinnal. Kõikide nende retseptorid on integraalsed
(trans)membraansed proteiinid, mis koosnevad seitsmest membraani läbivast heeliksist
(segmentidest).
Kemokiinid võivad olla produtseeritud vastuseks bakteriaalsetele produktidele, viirustele
ja agentidele, mis põhjustavad füüsilist kahjustust.
* Neutrofiile meelitavad: kemokiinid, komplemendi fragmendid, bakteriaalsed peptiidid.
Need on esimesed rakud, mis jõuavad põletiku piirkonda ning suurtes kogustes.
36.
Põletikureaktsioone stimuleerivad ja pidurdavad tsütokiinid.
Stimuleerivad:
TNF-α – aktiveerib endoteeli, neutrofiile ja põhjustab palavikku ning akuutse faasi
valkude tootmist.
IL-1 – aktiveerib endoteeli ja põhjustab palavikku ning akuutse faasi valkude tootmist
IL-12 – aktiveerib T rakke diferentseeruma Th1 suunas ning NK rakke, sellega toimub
tagasiside mehhanism
IFN-α ja IFN-β – paneb lähedal olevaid rakke tootma antiviraalseid valke ja MHC I,
aktiveerib NK rakke
IL-6 – soodustab B rakkude prolifereerumist
IL-15 – soodustab NK rakkude proliferatsiooni
IL-18 – T rakke tootma IFN γt
IL-23 – aitab IL-17 tootvaid T rakke
IL-8 – kemokiin, mis kutsub kohale neutrofiilid
IL-2 – kasvufaktor T-rakkudele – põhjustab T rakkude klonaalset ekspansiooni pärast
antigeeni äratundmist, samuti stimuleerib B rakkude proliferatsiooni ning aktiveerib
NK rakke ja monotsüüte.
IL-4 – peamiselt toodavad seda Th2 rakud – stimuleerib B rakkude proliferatsiooni ja
diferentseerumist, indutseerib B rakke tootma IgE, tõstab MHC II ekspressiooni
professionaalsetel APCdel.
IL-5 – peamiselt toodavad seda Th2 rakud – stimuleerib B rakkude ja eosinofiilide
proliferatsiooni, samuti aktiveerib eosinofiile.
o IL-4 on vajalik IgE produktsiooniks ja IL-5 eosinofiilide aktivatsiooniks –
eosinofiilid tunnevad ära IgE-ga kaetud parasiidid.
IFN-γ – toodavad efektor T rakud ja NK rakud – stimuleerib TNF-α ja IL-12
produktsiooni – TAGASISIDE MEHHANISM
Pidurdavad:
IL-10 – inhibeerib IL-12 produtseerimist
TGF-β – toodavad antigeeniga stimuleeritud T rakud – inhibeerib T rakkude
proliferatsiooni ja diferentseerumist ja makrofaagide aktiveerumist
IL-1RA – IL-1 tsütokiini retseptori antagonist
37.
Makrofaagide iseloomustus.
Makrofaagid on valged vererakud, mis tekivad monotsüütide diferentseerumisel. Nad on
kõige efektiivsemad ja suuremad fagotsüüdid. Kõrvaldavad näiteks paranevast koest
surnud rakkude jäänused, mikroobid, kasvajarakud jt. Samuti on neil oluline roll teiste
immuunsüsteemi rakkude stimulatsioonil patogeenide vastu. Makrofaag esitab peptiide
(APC). Tekib monotsüütidest. Kudedes alles muutuvad makrofaagideks.
Makrofaagid jagunevad M1 ja M2 tüüpideks.
M1 on esialgses koekahjustuses väga olulised. M1 makrofaagidele on iseloomulikud
põletiku näitajad: olulised Th1 tekkeks. M1 on kasvajavastase toimega, inhibeerivad
kasvajate edasist kasvamist, koekahjustus, proinflammatoorne, patogeeni tappev.
Toodab ROS-e, NO, lüsosüüme. Põletiku rajas toodab IL-1, IL-12, IL-23, kemokiine.
M2 tüüpi (alternatiivselt aktiveeritud makrofaag) soodustavad kasvajate arenemist, nad
on anti inflammatoorsed, haava paranemises, kõrge fagotsütoos. Nad tüüpiliselt
osalevad koe kahjustuse parandamises. Olulisel määral toetavad uute veresoonte teket.
Töötavad immuunvastusele vastu tasakesi. Olulised on paranemisfaasis. Kasvajad
kavalal moel muudavad makrofaagi eellased M2 tüüpi makrofaagideks. Kasvajad on
üleparanejad. Toodab IL-10, TGF-β.
Th1 rakud vastutavad makrofaagide poolt fagotsüteeritavate rakusiseste bakterite
hävitamise eest. Toodetud IFN γ stimuleerib ka komplemendi seondumist, mis aitab
kaasa fagotsütoosile. Lisaks stimuleerivad LT ja TNF α neutrofiilide aktivatsiooni.
Makrofaagi aktivatsioon: Kohtumine koes T rakuga (nii Th1 CD4+ kui ka CD8+) muudab
verest sinna liikunud
monotsüüdi makrofaagiks. Toimub oluline muutus raku
fenotüübis, mille tagajärjel on makrofaagil võime hävitada mikroobe. Lisaks aktiveerivad
TLR-id makrofaagi. Makrofaagi aktivatsioon toimub
CD40-CD40L ning IFNγ
vahendusel. Toimub
TNF-R kaudu vahendatav NF-kappaB ja SAPK signaalraja
aktivatsioon. Makrofaagi aktiveerib ka LPS. Edasises aktivatsioonis osalevad STAT1 ja
IRF1. Vabade hapniku radikaalide abil ja lämmastikoksiidide abil hävitavad baktereid ja
esitavad seda siis T-rakkudele.
38.
Dendriitrakkude iseloomustus. Dendriitrakkude roll adaptiivsetes
immuunreaktsioonides. Antigeeni töötlus antigeeni esitavas rakus.
Dendriitrakud on kõige tähtsamad antigeeni esitajad – ka professionaalsed ja
kandjad perifeersetesse lümfoid-organitesse (samal ajal toimub raku
“küpsemine”). Ühendavad adaptiivse ja loomuliku immuunsuse. Neil on pikad
dendriidid – nagu kombitsad ja sensorid, millega korjavad antigeene üles. Fagotsüütilised
omadused. Elavad lümfoidkoes, limaskesta epiteelis, organite parenhüümis. Müeloidset
päritolu. Sekreteerivad tsütokiine. Nad patrullivad, ja viivad lümfisõlmedesse antigeene
esitlemiseks. CCR7 T rakkudel kutsub neid sinna. DC-del vastavalt CCL21 kemokiin.
Konventsionaalne DC – vastavad mikroobidele minnes lümfisõlmedesse esitavad T
rakkudele nende antigeene.
Plasmatsütoidsed DC-d vastavad vara viiruslikule infektsioonile.
T-rakkude aktivatsiooniks on vaja APC-sid (peamised on kudedest lümfisõlmidesse
liikunud DC). DC esitavad antigeeni naiivsetele T-rakkudele
MHC I/II kompleksis – kuna
on professionaalsed, on neil mõlemad. DC epiteliaalsetes ja teistes kudedes (+põrnas ja
veres) ebaküpsed, selles staadiumis nad fagotsüteerivad ja töötlevad antigeeni. Samal
ajal patogeene äratundvate retseptorite (nt TLR) seondumine signaal küpsemiseks,
lümfisõlmidesse liikumiseks ja suureneb võime antigeeni esitada, kostimulatoorse B7
ekspressioon. DC on võimelised ka presenteerima CD8+ väliseid antigeene koos MHC I
(cross-presentation).
Dendriitrakkudel on ka ohuretseptorid pinnal – TLR nt, saab sealt signaali ja otsustab kas
aktiveerub või mitte. Kui aktiveerub, migreerub lümfisõlme, ekspresseerib CD80
molekule, et aidata T rakku aktiveerida (t rakul CD28).
39.
T-, B- ja NK-rakkude teke ning võrdlev iseloomustus. T-helperid (Th) ja
Ttsütotoksilised (CTL, Tc) rakud. Pinnamarkerid. NKT-rakud. Mälurakud.
Teke:
T-rakud: Arenevad luuüdis tüvirakkudest ning küpsevad ja diferentseeruvad tüümuses.
Ekspresseeritakse erinevaid antigeeni retseptoreid raku pinnal. Rakud, mis pole
võimelised neid ekspresseerima surevad ja eemaldatakse makrofaagide poolt.
Positiivselt selekteeritud rakud jäävad ellu ja hakkavad ekspresseerima madalal tasemel
nii CD4 kui CD8. Toimub veel kord selektsioon ja selle järel ellu jäänud rakud hakkavad
produtseerima kas: CD4 (Th-rakk) või CD8 (Tc-rakk), nad ja lahkuvad tüümusest
perifeersetesse kudedesse. Selektsiooni kaudu jääb järgi suhteliselt vähe T-rakke – need,
mis on võimelised ära tundma MHC molekuli koos antigeeniga ja ei ole liigreaktiivsed
keha enda rakkude suhtes.
Kui Th rakk esitletakse nii antigeenile kui ka õigele tsütokiinile, saab temast aktiveeritud
Th-rakk.
CD8+ T rakkude aktiveerimine toimub kas Th rakkude abil (mis aktiveerivad APC) või
lümfisõlmedes paiknevate dendriitrakkude abil, mis esitlevad MHC I CD8 rakule.
B-rakud: tekivad samuti luuüdi tüvirakkudest, küpsevad lümfikoes ja kehavedelikku
ilmunud antigeen aktiveerib nad immuunglobuliine produtseerivateks plasmatsüütideks.
Väike osa B-rakke tekib ka eneseuuenduslikult. Iga individuaalne B-rakk omab täiesti
kindlat, unikaalse spetsiifikaga antikeha. See on saavutatud sellega, et B-lümfotsüütide
diferentseerumise ajal kombineeruvad omavahel erinevad geenide segmendid. See
toimub aga igas konkreetses rakus isemoodi. Kui B-rakk on puhkeolekus (pole kohanud
Ag), siis tema poolt sünteesitud antikehamolekulid on plasmamembraanis. Kui aga B-
lümfotsüüt aktiveeritakse antigeeni poolt, siis ta hakkab neid samu molekule
sekreteerima.
NK-rakud pärinevad suurtest granulaarsetest lümfotsüütidest. Neil puuduvad antigeeni
retseptorid.
T- ja B- rakkudele ühine
Tsirkuleerivad kehas verest kudedesse ja sealt tagasi verre. Lümfisõlmedesse
lahkuvad lümfotsüüdid tsirkulatsioonist homing retseptorite abil, nad võivad tagasi
vereringesse tulla kohtades, kust leiavad antigeeni, millega seostuda.
Lümfotsüütidel on individuaalsed retseptorid, mis muudavad ta spetsiifiliseks teatud
antigeeni suhtes
Kui retseptor aktiveeritakse spetsiifilise antigeeniga, algab klonaalne proliferatsioon.
Neil on mälu, mis tagab suurenenud immuunvastuse sama antigeeniga kohtudes.
B-rakud
B-rakkudel on immuunoglobuliinid (Ak) pinnal, mis talitlevad Ag retseptoritena.
Kui B-lümfotsüüte stimuleerida nende antigeen-retseptorite kaudu, arenevad neist
antikehasid sekreteerivad plasmarakud. Plasmarakud on normaalselt leitavad
lümfoidkoes ja väga vähesel hulgal vereringes.
Stimuleeritud B-rakk võib areneda B-mälurakuks.
B-rakud käituvad ka kui antigeeni esitavad rakud (APC) T-lümfotsüütidele.
Subpopulatsioonid B1, B2.
T-rakud
Kui tunnevad ära oma antigeeni, vastavad nad klonaalse proliferatsiooniga ja
kannavad üht kahest funktsioonist:
Th rakud (CD4), T-helper-rakud amplifitseerivad/aitavad immuunvastusele kaasa
peamiselt tsütokiinide produktsiooniga. Nad interakteeruvad makrofaagidega ja
produtseerivad ka tsütokiine, seeläbi nad aktiveerivad immuunsüsteemi rakke
(stimuleerivad B-, Tc-rakkude, makrofaagide ja enda proliferatsiooni). Th rakud
jagunevad Th1 ja Th2
CTL T-rakud (CD8); T-tsütotoksilised rakud – tapavad rakke antigeen-vahendatud
mehhanismide kaudu. Seostuvad pinnaretseptorite antigeensete sidumiskohtade abil
anormaalsete pinnaomadustega rakuga (organismi muundunud raku või võõrrakuga)
ja hävitavad selle
CD4 ja CD8 molekulid juhatavad T-raku õige märklauani, mis vastavalt
interakteeruvad MHC II ja I klassi molekulidega
CD8 prolifereerudes võib 50-100 000 korda tõusta arv. CD4 puhul 100-1000 korda.
Loomulikud killerrakud (NK-rakud)
Põhifunktsiooniks on viirusega infitseeritud rakkude ja kasvajarakkude äratundmine ja
hävitamine
Oma (Self) MHC molekule kandvad rakud on NK ründe eest kaitstud
Toime erivormiks Ak kaudne rakuline tsütotoksilisus
Aktiveeritud NK toodavad tsütokiine reguleerivad immuunvastust
Efektormehhanismid sarnased T-lümfotsüütidele, aga märklaudraku äratundmine
toimub teisiti: nad liiguvad kehas ringi ja otsivad nakatunud rakke.
Rakkude pinnamarkerid:
T-rakud: TCR(/ või /); CD3, CD2, CD5, lisaks võivad olla CD28 jne
T-helper-rakkudel CD4
CTL T-rakkudel CD8
B-rakud: BCR (pinnaimmuunoglobuliinid), CD19, CD20, lisaks võivad olla CD40, CD5,
FcgRII, CR1 jne
NK-rakud: CD2, CD16 (FcgRIII) CD56 , CD94 (killeri Ig-sarnane retseptor-KIR), puudub
CD3
40.
I ja II tüüpi T-helperite (Th1 ja Th2) ja Th17 rakkude iseloomustus ja
tähendus immuunvastuses. Antigeenide, dendriitrakkude ja tsütokiinide roll
T-rakkude polariseerumisel.
CD4+ markereid kandvad T-helperid on regulatoorse funktsiooniga. Nende ülesandeks
on tunda ära antigeen koos II tüüpi koesobivusantigeenidega (MHC II), signaliseerides
seejärel B ja Tc lümfotsüüte. Th rakud sekreteerivad antikehade produktsiooni
suurendavaid ning Tc rakke ja makrofaage stimuleerivaid immunomodulaatoreid.
Viimaste produktsioonispektri alusel jagunevadki Th lümfotsüüdid kaheks grupiks:
Th1 – toodavad IL-2, TNFβ,
INFγ. Th1 aktiveerib
rakulisi immuunreaktsioone.
Th2 – toodavad IL-4, IL-5, IL-6, IL-10. Th2 aktiveerib
antikehade tootmist (k.a. IgE).
Allergilised reaktsioonid
Th17 – toodavad IL-17, IL-22, IL-21. Th17 aktiveerivad neutrofiile, tõstab põletikku
ning IL-17 kaudu aitab antimikroobseid aineid toota. Limaskestad, nahal (seen,
bakter)
Funktsioon täpsemalt:
Th1 ja Th2 rakkude polarisatsioon immuunsüsteemi erinevate aktivatsiooniseisundite
puhul (ei ole absoluutne, võib sõltuda haigusstaadiumist jne.
Th1:
rakusisene bakteriaalne infektsioon, viirusinfektsioon, organ-spetsiifilised
autoimmuunhaigused;
o
IL-2: soodustab teiste lümfokiinide produktsiooni, suurendab Tc rakkude
diferentseerumist ja proliferatsiooni, aktiveerib NK rakke, B-rakkude
proliferatsiooni, on immunoglobuliinide sekretsiooni kofaktor.
o Toodab: INFγ: aktiveerib makrofaage, NK-rakke, suurendab IgG tüüpi
antikehade tootmist, suurendab MHC II ekspressiooni raku pinnal, pärsib Th2
lümfotsüüte.
Th2:
rakuväline bakteriaalne infektsioon, atoopiline allergia, tolerants loote kudede
suhtes. Helmindid algselt
o IL-4: soodustab B-rakkude proliferatsiooni ja
IgE tüüpi antikehade sünteesi,
aktiveerib Th2-rakke ja pärsib Th1-rakke, tema toimel tekivad B-rakkude
pinnale IgE ja MHC II retseptorid, soodustab nuumrakkude paljunemist.
Alternatiivne (M2) makrofaagide areng, surub tavalisi makrofaagide
aktivatsiooni alla
o IL-5: indutseerib IgA ja IgM tüüpi antikehade sünteesi, suurendab eosinofiilide
produktsiooni ja aktiveerib neid.
o IL-6: indutseerib aktiveeritud B-rakkudes immunoglobuliinide sünteesi,
indutseerib palavikku, ägeda põletiku faasi proteiinide sünteesi ja ACTH
vabanemist.
o IL-10: makrofaagide aktiivsuse pärssimine, B-rakkude proliferatsiooni ja
immunoglobuliinide tootmise stimulatsioon.
Th17:
o Arenevad vastusena bakterite ja seentele. Haigustest võivad tekitaad
autoimmuunseid põletikuhaiguseid
Th0 sekreteerivad nii IL-4 (B-rakkudele) kui ka INF-γ. IL-4 inhibeerib Th1 produktsiooni ja
INF-γ inhibeerib Th2 produktsiooni.
Immuunreaktsioon võib aeg-ajalt muutuda tugevalt polariseerituks Th1 või Th2
produktsiooni suhtes, nii et üks või teine hakkab domineerima. Eriti võib see juhtuda
kroonilise immuunvastuse puhul, näiteks parasiitsel infektsioonil.
41.
Immuunvastuse regulatsioonimehhanismid. Regulatoorsed T-rakud.
Foxp3. CTLA-4 ja teised regulatoorse funktsiooniga molekulid.
Immuunvastuse regulatsioonis osalevad:
Tsütokiinid (autonoomne-, sünergistlik- ja antagonistlik toime). – eespool
Tolerants reguleerib- tsentraalne ja perifeerne
Mitut signaali vaja et aktiveerida – kindlat konteksti vaja antigeeni esitamiseks
T rakkude poolt antigeeni ära tundmine – tasakaal aktiveerivate ja inhibeerivate
retseptorite vahel.
Cytotoxic T-lymphocyte associated protein 4 (CTLA4)
Molekul on CD4+ ja CD8+ T-rakkude ja Treg-ide pinnal. Ta on CD28 homoloog, seondub
B7-ga – takistab CD28-ga seondumast tal. CTLA-4 ekspressioon on aktiveerimata
rakkudel madal. Ekspresseeritakse ka Tregide pinnal. CTLA-4 endotsütoosi teel sööb ära
B7 molekulid, vähendab seega APC-del nende ekspressiooni ⇒
pärsib T-rakkude
aktivatsiooni. Kui CTLA-4 on defektne, toimub kontrollimatu lümfotsüütide aktivatsioon,
lümfisõlmed suurenevad süsteemne autoimmuunsus perifeerse tolerantsuse läbi
kukkumine ⇒ T cell mediated disease
PD-1 ja PD-1L – programmed death 1
PD-1 tunneb ära PD-L1 (ligand) ning PD-L2. Neid ligande ekspresseeritakse nii APC-de
peal (L2) kui ka muude peal (L1 ja L2). Kui toimub PD-1 seondumine PD-L (mõlemad) T
rakk inaktiveeritakse. PD-1 defektsuse korral viib see autoimmuunhaigusteni – nt
luupusesarnane neeruhaigus, artriit. PD-1 inhibeerib T rakulist vastust, indutseerides T
rakus inhibitoorseid signaale.
Regulatoorsed T-rakud (Treg, CD4+CD25+FOXP3+)
Tekivad tüümuses tugeva signaali mõjul. Liiguvad perifeeriasse ja on olulised
perifeerse tolerantsi tekitamisel TGF β mõjul. Omavad TCR ja aktiveeruvad ainult siis,
kui seostuvad MHC II molekuliga ja saavad lisasignaali APC B7 molekulidelt.
Ekspresseerivad palju IL-2 retseptorit
CD25. Tarbides IL-2, inhibeerivad teisi T-rakke,
(IL-2 on vaja T rakkude arenguks) et nad ei algataks immuunvastust organismi enda
komponentide vastu, st kaitsevad autoimmuunsuse vastu.
Ekspresseerivad transkiptsioonifaktorit FOXP3 – Tregide diferentseerumiseks väga
oluline!
Kasutavad IL-7 asemel
IL-2, et ellu jääda!
Kõrge CTLA-4 ekspressioon
Aktiveerituna sekreteerivad
immuunsupressiivseid tsütokiine
IL-10 ja TGF-β.
Need mõlemad on tugevad immuunsupressandid – nad inhibeerivad Th1 ja Th2.
Regulatoorsed T-rakud mängivad olulist rolli organismi immuuntolerantsuse säilitamisel
ning autoimmuunhaiguste ära hoidmisel. Regulatoorsed T-rakud on vajalikud säilitamaks
immuunsüsteemi homöostaasi. T-regulatoorsete rakkude arvu vähenemine või
funktsiooni kadumine võib viia autoimmuunhaiguste tekkele. Teisalt, nende liigsel
kuhjumisel on leitud kahjulikku mõju kasvajate ja krooniliste põletikkude korral.
Kõige tähtsam Treg-ide tunnusjoon on nende immuunsüsteemi pärssiv toime, kuid lisaks
eelnevale iseloomustab neid ka sõltuvus
IL-2-st ning abi-T-rakkudega seotud
efektor
tsütokiinide ekspressiooni puudumine (IFN-γ, IL-4, IL-17). Treg-ide supresseeriv
toime seisneb läheduses olevate efektor-T-rakkude funktsiooni maha surumises.
Regulatoorsete funktsioonidega T-rakke on kirjeldatud mitmeid, kuid kõige olulisemateks
peetakse ning kõige paremini on kirjeldatud
CD4+CD25+ Treg-e, mille
diferentseerumiseks on vajalik transkriptsioonifaktor
FOXP3. Treg- supresseerivad
tsütokiinidega. Tõmbavad keskkonnast IL-2 ära, kui seda pole siis T-rakk ei aktiveeru.
42.
Loommudelid immuunsüsteemi uuringus. Võimalused ja piirangud
immuunsüsteemi funktsiooni hindamisel.
Hiir on kõige tavalisem katseloom, vähem rotti ja küülikut kasutatakse ka. Looma valik
sõltub uuritavast teemast.
Hiir: eelised:
• Kosmopoliit – laia levikuga
• Sarnane immuunsüsteem (kuigi ka mitmed olulised erinevused)
• Lihtne kasvatada, standardiseeritud toit-jook, vähe ruumi, odav
• Lihtne paljundada, väike, sobib laboritingimustesse
• Genoom, anatoomia ja füsioloogia väga hästi teada
• Reaktiivide baas
• Väljatöötatud meetodid
Hiir ja rott on väga erinevad – erinev käitumine ja ülalpidamine.
Rott:
• Laborirott on rändrott Rattus norvegicus
• Populaarsuselt hiire järel teine mudel immunoloogias
• Traditsiooniline mudel füsioloogias, eriti aju ja endokriinsüsteemi uurimisel
• Füsioloogiliselt rohkem inimese moodi.
• Immuunsüsteemis ka rohkem sarnasusi
– T rakud ekspresseerivad MHC class II
– Foxp3 (Treg marker) on ka aktiveeritud T rakkudel
– Makrofaagidel on CD4 ja CD8 markereid
• Rottidest kasutatakse liine/tüvesid (geneetiliselt identsed) kui ka geneetilise
variatsiooniga tõuge.
• Rott on sotsiaalsem ja intelligentsem kui hiir ja väga uudishimulik
• Ei vasta alati inimesele
Küülik
Antikehade tegemine
Pürogeenid
Väga tundlikud pupillid
Väga hea kuulmine ja lõhnaaisting
Suurloomad
-: hind, aeg, reaktiivide vähesus
On kasutatud hobuseid, lambaid, vasikaid erinevate haiguste uurimisel
43.
Tähtsamad loommudelid immuunsüsteemi funktsiooni hindamisel.
Immuunpuudulikud hiired – SCID
• RAG hiirel on deleteeritud
Rag1 või koos
Rag2 geeniga.
• Puuduvad T ja B lümfotsüüdid.
• SCID/Rag eriti sobiv “humaniseerimiseks”
• Võimaldavad lisada uusi lümfotsüüte, näiteks inimese omasid
• Nude ehk FoxN1-puudulik hiir. Puudub tüümus ja seega ka T rakud
Transgeenne hiir
Genoomis on juhuslikku kohta integreerunud geen või DNA fragment
Knockout tehnoloogia hiirel (Cre-lox) süsteem
Spetsiifilise geeni deletsioon
• Sama geeni deletsioon erineval viisil võib anda erinevaid tulemusi
• Hiirte geneetiline taust mõjutab sageli
Mutatsioon ühes geenis võib põhjustada muutusi teistes geenides, ka naabergeenides
• Hiirte konkreetsed elutingimused võivad mõjutada fenotüübi teket
• Mutatsioonid mis mõjutavad organi (LN; tüümus, põrn) arengut võivad mõjutada
teiste geenide funktsiooni
Knock-IN mudelidSpetsiiflise mutatsiooni tekitamiseks või mingi geeni asendamiseks teise geeniga. Hiir on
tavaliselt normaalse fenotüübiga.
CRISPR-CAS mudelid.
T-rakkude selektsiooni uurimise mudelid
1) OT-I (CD8) ja OT-II (CD4). Ovalbumin on koespetsiifiline geen.
2) H-Y – transgeenne hiir. H-Y peptiid kloneeritakse TCR geenidesse. MHC on H-2Db
tüüpi.
B-rakkude selektsiooni uurimise mudel: OVA ja BCR (ovalbumiin)
44.
Autoimmuunhaiguste loommudelid (sh AIRE-/-, NOD, BB, d3T jt).
EAE (experimental autoimmune encephalomyelitis) hiir
Hulgikoldelise skleroosi (sclerosis multiplexi mudel, MS)
Immuniseerides müeliini spetsiifilist valku FCA-ga – (myelin basic protein, MBP;
proteolipid protein, PLP; myelin oligodendrocyte glycoprotein, MOG)
Tekib entsefalomüeliit (pea- ja seljaajupõletik) koos lümfotsüütide ja makrofaagide
infiltratsiooniga – võetakse MBP-d ja nende spetsiifilise T raku kloonid, pannakse
normaalsesse rohtti EAE rott.
Demüelinatsioon ca 1-2 nädalat peale immuniseerimist
NOD (non obese diabetes) hiir
T1D mudel, üsna sarnane inimese T1D-le
NOD hiirel on spetsiifiline I-Ag7, mis on struktuurilt sarnane DQB1*0302 (Asp57 b-
ahelal)
Ilmselt on osa haigusgeene samad mis inimese T1D aga mitte tingimata kõik
Tekib insuliit (8 nädal), mis edasi areneb diabeediks
Esineb rohkem emastel (80%) isastel vähem (20%)
Haigus on üle kantav vanematest hiirtest noorematele NOD hiirtele
Haiguse teke sõltub palju pidamistingimustest
SLE (systemic lupus erythematosus)
Spontaansed hiiremudeliks on (NZBxNZW)F1
Tekivad antinukleaarsed ja anti-DNA antikehad
Sõltub T rakkudest, aga B rakud olulised kuna antikehad moodustavad
immuunkomplekse (tuumavalgud, DNA-valgu kompleksid, mis tekivad apoptootilistest
rakkudest)
Oluline on geneetiline taust, MHC seotud haigus
Hiired hüperreageerivad võõrastele antigeenidele
AIRE -/- mutatasioonid AIRE geenis viivad tsirkuleerivate autoantikehadeni erinevate
organite vastu
BB – biobreeding rott – spontaalselt saab tüüp I diabeedi – kasutatakse loommudelina
diabeedi puhul – peamiselt uuritakse T1D geene
45.
Viirusinfektsioonide immunoloogia.
Viirustel on peamised sissetungi kohad – limaskestad, nahk, hingamisteed,
urogenitaaltrakt. Nad kinnituvad raku pinnale kindlate retseptorite kaudu.
Viirusinfektsioonid jäävad tihti asümptomaatiliseks, mis aga ei tähenda, et ei tekiks
immuunsust. Viiruste puhul on kaitses suurem osa
interferoonidel (mittespetsiifilised
mehhanismid). Kuid kuna viirusinfektsiooni korral on tegemist
intratsellulaarselt
asetseva haigustekitajaga, on haigusprotsessi generaliseerumise vältimisel ja
likvideerimisel põhiosa
rakulisel immuunsusel. Immuunreaktsioonid saab jaotada
mittespetsiifilisteks ja spetsiifilisteks.
Viiruse antigeene korjatakse üles makrofaagide ja DC-de poolt. Kõigepealt aktiveerub
loomulik immuunsus – NK rakud (IFN α, β, TNF-afla, IL-12) – kontrollitakse viiruse
replikatsiooni, aga ei elimineeri seda. Viiruse eemaldab CD8+ T rakk.
Interferoonide ülesanded:
1. Olulised sammud on inflammosoomi ja põletiku pidurdamine (koekahjustuse
vältimine)
2. Naaberrakkude teavitamine barjääride püstitamiseks
3. Nakatunud rakkude surm
4. Immuunrakkude värbamine pikaajalise immuunsuse tagamiseks
Loomulik immuunsus – IFN α ja IFN β viirusvastane toime
TLR-ide kaudu tuntakse ära viiruse materjal IRF geeni transkriptsioon
infterferoonide geenitranskriptsioon
IFN-de toimel aktiveerub JAK-STAT signaalirada, aktiveeritakse ribonukleaas L süntees
(mRNA lagundamine), APK (proteiinkinaas)ide inhibeerimine – viirusvalgu translatsiooni
inhibeerimine. Üldiselt on eesmärk viirusinfektsioonist jagu saada. –
piirata viiruse
levikut varases staadiumis.
• Algatatakse p53 kaudu apoptoos
• MHC klass I ekspressiooni suurendamine (kaitseb rakke NK eest: väiksema MHC I
molekulide tihedusega rakkude hävitamine aktiveerunud NK-rakkude poolt on
tõenäosem kui kõrge MHC I tihedusega rakkude hävitamine; kõrgem MHC I tihedus
suurendab tõenäosust saada “ära tuntud” CD8+ T rakkude poolt. Nakatunud rakud ei
suuda MHC I ekspressiooni piisavalt suurendada.)
• Stimuleeritakse NK-rakke teisi rakke hävitama
• Stimuleerib Th1 teket
• Inhibeerib rakkude proliferatsiooni
Inflammasoomid
Olulised IL-1 ja IL-18 küpsemiseks läbi selle, et rakk viiakse
püruptoosi (rakusurm). Kui
detetkeeritakse ssRNA (TLR7/8), aktiveeritakse kompleksid (MYD88, IKK α/β), mis
tulemusena aktiveeritakse NF kappa B signaaalirada. ⇒ IL-1 β tootmine. Tuntakse ära ka
viiruse kattevalke ja polümeraase. Rakust väljuvad küpsed IL-1 β ja IL-18.
NK rakud
Vahendavad immuunvastust – läbi tsütokiinide, antikehade, ligandide (stressimolekul),
viiruse ligandide. Tapavad nakatunud rakke
Antikehad
Ak-d ei ole võimelised tungima infitseeritud rakku ning on seetõttu ebafektiivsed
latentsete ja otseselt rakust rakku levivate viirusinfektsioonide suhtes. Küll seostuvad
nad aga ekstratsellulaarsete viiruslike antigeenidega. Sellisteks Ag-deks võivad olla
organismi rakul paiknevad viirusvalgud ja vabalt vereringes tsirkuleerivad intaktsed
viirused.
Immuunvastusena tekivad inimesel antikehad – IgA olulised limaskestadel, eriti
hingamisteedes ja soolestikus. IgG1 – kaitse kopsudes.
Võivad olla nii neutraliseerivad kui mitteneutraliseerivad (nnAb). nnAb ainult märgistavad
nakatunud rakku, sesostudes tema antigeenidega. Seejärel tunnevad NK rakkude
retseptorid AK ära ja saadavad rakusurma signaali apoptoos
Viirusevastases kaitses on pikaajalise kaitse tekkeks olulised
mälurakud. Lühiealised
plasmarakud mälu-B rakk ⇒ pikaealine plasmarakk.
Neutraliseerivad antikehad limaskestadel hoiavad ära nakatumise –
viiruspartiklite külge seondumine ei lase neil kinnituda rakule.
Ak seostumine viirusosakesega pärsib
viiruse adhesiooni peremeesorganismi
rakulemembraanile ja edasist penetratsiooni – neutralisatsioon. Veres ja kehavedelikes
täidavad neid ülesandeid IgG ja IgM, limaskestadel aga IgA. Isegi kui Ak-dega kaetud
viirus suudab tungida rakku, on sageli takistatud tema desintegratsioon ja nukleiinhappe
vabanemine kapsiidist. Ak-d moodustavad tsirkuleerivate viirustega komplekse, piirates
nii viiruste levikut. Kompleksid on paremini fagotsüteeritavad ning äratuntav
komplemendile. Komplement võib viiruse otseselt lüüsida, purustades viiruspartikli, aga
ka opsoneerida.
T-rakuline immuunsus
Kaitseb raske haiguse eest. CTL ründab nakatunud raku vastu – toimib kõikide „tüvede
vastu“, isegi kui viirus muutub. CD8+ T rakkudel on perforiinid ja gransüümid, mis siis,
kui MHC I-ga seondumine on toimunud, tekitavad poore, mistõttu nakatunud rakk sureb.
See vastus püsib hästi.
AGA krooniliste viirusinfektsioonide korral võivad T rakud olla kurnatud – PD-1 seondub
PD-L1-ga ja ei lase funktsioneerida T rakul.
Viirusinfektsioonide immuunopatoloogia.
Immuunsüsteemi rakkude põhjustatud patoloogia: autoimmuunsus; hüperreaktsioonid,
koekahjustuse teke; tsütokiinide torm.
Tuumorite arenemine – HCV, HBV, HIV, papilloomiviirus; HIV,
ADE – antibody dependent enhancement – nt COVID puhul viirus saab antikeha Fc kaudu
siseneda. Viiruse ja viirust
mitte neutraliseeriva antikeha - IgG - kompleks seondub
antikehal oleva Fc domääni kaudu Fc Gamma retseptorile, mis on immuunrakkude, nt
makrofaagide. Retseptori kaudu saab rakku sisse ning saab seal sees paljuneda.
46.
Seeninfektsioonide immunoloogia.
Seened on heterogeensed ja eukarüootsed. Suhteliselt vähe neist on inimpatogeensed
ning vähe on neist teada. Kasvades produtseerivad ensüüme, mis lagundavad
ümbritsevat keskkonda. Rakuseinas on kolesterooli asemel ergosterool. Seene
infektsiooni nimetatakse mükoosiks. Mõned infektsioonid on endeemsed, mõned on
oportunistlikud (tervetes inimestes kerge haigus, nõrgematel (ID inimestel) äge
haigus). Eriti sagenevad infektsioonid (tihti oportunustlikud) pidurdunud reaktiivsusega
immuunsuse korral, T-lümfotsüütide defektiga, HIV haigetel. Neutrofiilide puudus on üks
kaasuv faktor mis võib viia seenhaigusteni.
Seened on kaetud CLR-ga C-type lectin receptor – nende kaudu saab seonduda MF, DC.
Aga seentel on palju erinevaid molekule, mille abil seonduda. Erinevate kinaaside abil
aktiveeritakse NFAT, NF-kappaB, ERK, IRF-3 rajad sünteesitakse
IL-1β, IL-18.
Patoloogiliste muutuste põhjuseks makroorganismis on kas tekitaja ise, tema poolt
produtseeritud toksiinid või antigeen-antikeha reaktsioonid, mis lõpevad
immuunkompleksi moodustamisega ja see kutsub esile allergilisi reaktsioone.
Seenhaiguste tekitajad põhjustavad ülitundlikkuse, mükotoksikoosi, mütsetismi ja
seeneinfektsiooni.
Esimesena märkavad seent neutrofiilid ja makrofaagid, kes vabastavad TLR-idega
fungitsiidseid ensüüme (ROS-id, lüsosüüm) ning fagotsüteerivad seene. Mõned
seeneliigid inhibeerivad tsütokiinide nagu TNF ja IL-12 sünteesi makrofaagides ja
stimuleerivad hoopis IL-10, mis inhibeerib makrofaagide tegevust
Peamine kaitsemehhanism on siiski rakulised immuunreaktsioonid, osalevad ka
humoraalsed reaktsioonid. Intratsellulaarsete bakterite tõrjega sarnased mehhanismid –
CD8+ ja CD4+ T rakud ühinevad et nt pärmseene kolooniaid eemaldada. Lisaks
tekitatakse tugev Th17 vastus, mis tekib tugeva DC aktivatsiooni tõttu (seene glükaan
dektiin-1 (seene PS retseptor). DC-d toodavad Th17 indutseerivaid tsütokiine (IL-6, IL-23)
⇒
põletik, värbavad neutrofiile ja monotsüüte, mis seent hävitavad
Seega seenehaiguste korral aktiveeruvad makrofaagid, neutrofiilid, CD4 Th1 ja antikehad
(aga enamikul puudub protektiivne toime). Koostoimivad loomulik ja omandatud
immuunvastused.
Subkutaansete ja süsteemsete mükooside korral ilmuvad erinevad antikehad, aga neil
puudub oluline roll, nad ainult annavad teada, et infektsioon on olemas. Kutaansete ja
pindmiste mükooside korral immuunreaktsioonid ei ole nii tugevad.
47.
Parasiitinfektsioonide immunoloogia.
Loomsed parasiidid – protozoa, helmindid, ektoparasiidid (puugid, lestad). Arengumaades
on nad 30% suremuse põhjustajateks. Malaaria nt käib siia alla. Parasiitidel on keerulised
elutsüklid, osa toimub peremehe sees. Inimesi mõjutab enim hammustustest esile
kutsutud – puugid, kärbsed, teod. Paljud parasiit infektsioonid on peremehe nõrga
immuunsüsteemi tõttu kroonilised.
Parasiite on palju raskem hävitada kuna suured + tekkinud kaitse: tugev välispind.
Ainuraksetel parsiitidel on muutlik elutsükkel, kus nad muudavad oma vormi, peremeest
ja antigeenset väljanägemist.
Loomulik immuunsuus: kuigi kutsuvad selle välja, aga parasiidid oskavad seda hästi
vältida ja peremehes ikkagi paljuneda. Fagotsüüdid neelavad ainurakseid, kuid võivad
hätta jääda nende rakusisese hävitamisega. Parasiidi on võimelised fagolüsosoomi teket
inhibeerima, mille tõttu nad jäävad eluvõimeliseks fagotsüüdi sees ning seal paljuneda.
Mõned parasiidid võivad isegi ekspresseerida pinnamolekule, mis tuntakse TLR-ide poolt
ära Suuri hulkrakseid parasiite fagotsüüt ei ole võimeline neelama, seda püütakse
kompenseerida makrofaagide sekreteeritud bioaktiivsete ainetega, aga paljudel
parasiitidel on liiga tugev kutiikula ning on keemilisele rünnakule nad on vastupidavad.
Lisaks need parasiidid, kes on nakatunud peremeesrakust ellu jäänud, võivad muutuda
resistentseks.
Omandatud immuunsus: kuna parasiite on erinevaid, on neil ka erinevad omandatud
immuunsuse mehhanismid (CTL-id, antikehad, antikehad + fagotsütoos, jne).
Fagotsüütide sees elavate parasiitide vastu on pearõhk
rakuliste immuunmehhanismide
aktiveerumisel, kuna paljud patogeensed protistid on arenenud peremeesrakkudes
elama, on vaja mehhanisme, mis on sarnased intratsellulaarsete bakterite ning
viirustega.
Fagotsüüdid tuleb hüperaktiveerida CD4+ Th1 toodetud tsütokiinide (põhiliselt IFN-γ)
abil, et nad suudaksid alla neelatud parasiidi hävitada. Kui aktiveeritakse hoopis Th2
vastus, tõstab see parasiidi elulemust sest Th2 surub makrofaagide tegevuse alla.
Antikehadest on kasu, kui parasiit satub rakust välja (kuigi eluvormi vahetades parasiit
võib kergesti põgeneda). Tugev humoraalne immuunvastus tekib rakuväliste ja
hulkraksete parasiitide vastu. Aktiveeritud Th2 stimuleerivad antikehade sekretsiooni ja
Ig klassi lülitamist IgE tootmisele. IgE antikehad kinnituvad parasiitidele ning aktiveerivad
eosonofiile ja nuumrakke. Tekib tugev põletik ja eosonifiilide graanulites olevad valgud
(MBL. ECP) kahjustavad parasiiti välismembraani. Olulised on veel tsütokiinid IL-4 ja IL-5.
Adaptiivne IV võib viia ka koekahjustuseni, kuna CD4+, kes aktiveerivad makrofaage,
indutseerivad DTH (delayed type hypersensitivity) reaktsiooni.
48.
Rakuväliste bakteriaalsete infektsioonide immunoloogia.
Rakuvälised e ekstratsellulaarsed bakterid on võimelised väljaspool peremeest
paljunema, nt veres või siedekoes ning hingamisteede ja soolestiku luumenites. Haigus
tekib
endotoksiinide (rakukesta komp.) või
eksotoksiinide (sekreteeritakse bakterite
poolt) toimega ja
põletikust põhjustatud koekahjustustega. Endotoksiiniks bakteritel
(GN) on LPS – lipopolüsahhariid – see on ka potentsiaalne makrofaagide, DC ja
endoteelirakkude aktiveerija. Eksotoksiinid on
väga patogeensed, lülitades krutsiaalseid
osi raku elutegevusest välja.
– Teetanus – tetanospasmiin
– Koolera – kolerogeen
– Botulismitekitaja neurotoksiin
Loomuliku immuunsuse kaitsemehhanismid:
Fagotsüüdid kasutavad pinnaretseptoreid, et rakuväliseid baktereid ära tunda,
kasutavad oma Fc retseptoreid, ja komplemendi retseptoreid, et antikehadega
opsoniseeritud baktereid ja komplemendi valke ära tunda
1)
komplemendi aktivatsioon, põletiku teke ja fagotsütoos. Peptidoglükaanid ja LPS
aktiveerivad komplimendi alternatiivsed teed. Komplemendi aktiivsed komponendid
kinnituvad bakteritele ja märgistavad (opsoneerivad) neid efektiivsemaks
fagotsütoosiks. Lisaks sellele on komplimendi lüütilise tee komponentidel on
membraani kahjustav toime.
2)
Põletiku teke. Aktiveeritakse TLR-id fagotsüütidel, vastavalt sellele, milline
pinnamolekul bakteril on. Komplement aktiveerib ka põletiku. C5a ja C3b
komponendid meelitavad põletikus osalevaid rakke. Ohuretseptorite (C3b-ga kaetud
bakter) kaudu aktiveerivad fagotsüüdid neelavad ja hävitavad bakterid ja toodavad
põletikutsütokiine.
3) DC ja fagotsüüdid sekreteerivad tsütokiine, mis meelitavad põletikus leukotsüüte
kohta.
TNF – väikestes kogustes – lokaalne põletik < süsteemsed mõjud < septiline šokk.
Omandatud immuunsus:
Tähtis on ka
humoraalne immuunsus. Tekkivad antikehad bakterite toksiinide või LPS
või patogeeni välismembraani vastu. Antikeha kinnitumisel blokeeritakse infektsioon,
häiritakse bakteri elutegevus või neutraliseeritakse toksiini (eriti head neutraliseerijad
IgG ja IgA). Antikehad võimendavad fagotsütoosi, sest antikehadega kaetud bakteri osad
fagotsüteeritakse kiiremini ja efektiivsemalt. Komplemendi aktiveerivad IgM ja IgG.
Polüsahhariididega kaetud bakteritega võitlemisel on väga oluline roll B rakkudel, kuna T
rakkudel pole sellist võimet neid ära tunda. Antikehad neutraliseeritakse,
opsoniseeritakse (fagotsütoos) ja aktiveeritakse komplemendi rada. Neutralisatsioon –
IgG, IgM, IGA – opsonisatsioon – IgG. (ja IgM).
T-rakud aktiveeritakse bakterite valguliste antigeenide poolt. CD4+ T rakud eritavad
tsütokiine, mis indutseerivad lokaalset põletikku, tõstavad makrofaagide ja neutrofiilide
fagotsütoosi võimet; ning stimuleerivad antikehade tootmist. Th efektormehhanismid
sõltuvad IL-4, IL-5 ja IL-10 stimulatsioonist. Th17 vastus värbab neutrofiile ja monotsüüte
et lokaalset põletikku teha. Lisaks indutseeritakse Th1 vastus, mis mõjul toodetakse IFN-
γ aktiveeritakse makrofaagid ja neutrofiilid.
T-rakust sõltumatud antigeenid on G+ bakterite peptidoglükaanid, neile immuunsust
ei võimenda Th rakud. Nendevastase immuunsuse tekkeks on vaja palju B rakke. T-
rakust sõltumatud antigeenide suhtes tekkiv immunoloogiline mälu on nõrgem ja
lühiajalisem, võimalikud reinfektsioonid.
T rakust sõltuvad on G- bakterite antigeenid –
valgulised, aktiveerivad hästi Th
süsteemi, mis aktiveerivad tsütokiine ja need omakorda antikehi. T rakust sõltuvate
antigeenide suhtes tekib pikaajaline mälu.
Ekstratsellularsete bakterite vastane IV võib kahjustada org-mi. Tekkib palju
tsütokiine tugeva reaktsoioni tulemusena (tsütokiinide torm) võim tekkida tugev põletik
ja
septiline šokk. Võivad tekkida ka hiliskomplikatsiooni nagu autoantikehade teke. Nt
betta hemolüütilise streptokokki M valgu vastased antikehad mis risti reageerivad
südamelihase sarkolemm ja müosiinvalkudega, võib tekkida südamepõletik.
MALT – Peyeri naastud. Hattudes on limakuht mis kaitseb. DC; T rakud patrullivad, IgA
antikehad on igal pool.
49.
Rakusiseste – intratsellulaarsete – bakteriaalsete infektsioonide
immunoloogia.
Loomuliku immuunsuse mehhanismid ei aita. 😊 Sest nad saavad elada ja
paljuneda ka fagotsüütides. Need mikroobid on suutnud leida niši, kus nad on
antikehadele kättesaamatud, seega on vaja: rakulisi mehhanisme!
Loomuliku immuunsuse rakulised mehhanismid: esimene kaitseliin fagotsüüdid ja
NK rakud. Sellised bakterid on fagotsüüdi sisesele hävitamisele resistentsed. Bakterite
produkte tuntakse ära TLR-ide ja NLR-idega nis aktiveerib fagotsüüdid. Hävitamine
muutub edukakas vaid
hüperaktiveeritud neutrofiilis ja makrofaagis. Nakatunud
rakkudel indutseeritakse NK rakke aktiveerivate ligandide ekspressioone, stimuleerides
makrofaagide ja DC-de
IL-12 ja IL-15 tootmist (mõlemad aktiveerivad NK rakke). NK
toodavad tsütokiine, tähtis on IFN-γ, mis omakorda saadab stiimuli MF ja fagotsütoositud
bakterite tapmise. Varane kaitse enne omandatud immuunsüsteemi sisse lülitust!
Omandatud immuunsuse rakulistest mehhanismidest on olulised: CD4+ T
rakkude poolne makrofaagide hüperaktivatsioon (läbi IFN γ ja CD40L – CD40 on APC-l) ja
infitseeritud rakkude lüüs Tc abil. Hüperaktiveerudes hävitab fagotsüüt rakusise bakteri
koos rakuga. Hüperaktiveerimiseks vaja 2 signaali: fagotsüütid esitavad T rakkudele
antigeeni. Makrofaagid või DC toodavad IL-12, mis suunab naiivseid Th diferentseeruma
Th1suunas, Th1 tiidab IFNγ ja oma pinnal CD40L fagotsüteeritud rakkude tapmine VÕI
CTL rakkude poolt nende tapmine, kes fagotsüüdis ellu jäid. Läbi IL-2 Th1 rakud
sekreteerivad IFN-γ aktiveeritakse makrofaage tootma antimikroobseid aineid, - ROS,
NO, lüsosümaalseid ensüüme. IFN-γ stimuleerib komplementi aktiveerivate AK isotüüpide
tootmist, mis ka opsoniseerivad bakteri fagotsütoosiks (aitab MF)
CD8+ T rakulinerada aktiveeritakse siis kui bakter põgeneb fagosoomist või kui bakter
transpordil liigub tsütosooli. Tsütosoolis peab CTL nakatunud raku tapma.
Põhiline erinevus fagotsüüte aktiveeritakse erinevate radade kaudu.
50.
Bakterite ja viiruste mehhanismid, mis aitavad vältida
immuunreaktsioone.
Viiruste peamised põgenemise mehhanismid
Peitumine
(varjumine)
Rakusisene eluvorm, varjatud levik, nn. privilegeeritud organisse
peitumine, peremehe antigeenidega katmine (ümbris), antigeenne
mimikri,
MHC kaudu viirusepitoopide esitamise piiramine
Antigeenne
variatsioon
Geenide mutatsioon ja rekombinatsioon, viiruse seroloogiline
plastilisus, geenide sisse-välja lülitumine → antigeenne nihe
(antigenic drift). (Punktmutatsioonid)
Drift – mutatsioonid nt pinnavalkudes, modifitseeritud antigeenid
Shift – suuremad muutused antigeenis, uued viirused nt à la
seagripp, linnugripp. Gripi uued tüved
Immuunvast
use
häirimine
Mittespetsiifiline immuunosupressioon
Spetsiifiline immuunosupressioon (makrofaagide, Treg. kaudu,
mikroorganismi produktidega)
Viirusinfektsioonid võivad provotseerida autoimmuunsust :
Viirusindutseeritud kohekahjustus, „krüptilised“ peidetud autoantigeenid saavad
immuunsüsteemile kättesaadavaks
Molekulaarne mimikri – viirusvalgu mingi epitoop on identne või homoloogne
mingisuguse kehaomase AH järjestusega, avaneb tee immuunvastuse tekkeks ja
edasiseks epitoobi levimiseks.
Mõned viirused persisteerivad in vivo latentselt – st on sümptomiteta ning ei replitseeru.
Nt herpes simplex, mis pesitseb priviligeeritud koes – sensoorse neuronina, kus ta
persisteerub latentselt. Ta aktiveerub vaid UV; bakteriaalsete inf., hormonaalsete
muutuste jms korral. Seal on hea pesitseda, kuna neuronite pinnal on väga vähe MHC I
molekule ja CTL-id ei tunne neid hästi ära. Madal MHC tase võib olla neuronitele kasulik
vältimaks nende destruktsiooni kuid muudab nad haavatavamaks infektsioonidele.
Latentne viirus toodab vähe proteiine ja neid presenteeritakse MHC I kaudu minimaalselt.
„Virokiinid” ja „viroretseptorid”
Viirused (suured DNA viirused) kodeerivad ja toodavad tsütokiinide, kemokiinide ja nende
retseptorite homolooge – nt on nad ära varastanud peremehe geenid ja kasutavad neid,
et paremini paituda.
Epstein-Barr viirus kodeerib IL-10-sarnast valku (stimuleerib B-rakke paljunema;
supresseerib rakulist immuunsust).
Herpes saimiri (T-raku troopiline viirus) kodeerib IL-17 sarnast valku, mis soodustab T-
rakkude proliferatsiooni (T3).
Poxviirused kodeerivad valke, mis meenutavad tsütokiini retseptoreid (IL-1, TNFα,
IFNγ või kemokiine), mis seovad ja inhibeerivad neid tsütokiine, seeläbi
supresseerides immuunreaktsioone.
Inimese herpesviirus 8 (HHV-8, põhjustab Kaposi sarkoomi) kodeerib kemokiini-
sarnaseid valke, mis seovad CC- and CXC-retseptoreid.
Bakterite immuunvastuse vältimise mehhanismid:
Rakuväliste bakterite puhul:
o Antigeenne variatsioon
o Komplemendi aktivatsiooni inhibitsioon
o Resistentsus fagotsütoosile
o ROS-ide eemaldamine
Rakusiseste bakterite puhul:
o Fagolüsosoomi inhibitsioon
o ROS-ide ja NO inaktiveerimine
o Fagosoomi membraani lõhkumine ja tsütoplasmasse põgenemine
Fagotsütoosi vältimise viisid:
Kemotaksise repellendid.
Kapsel, mis vähendab kleepumist
fagotsüüdile.
Bakter toodab ainet, mis inhibeerib
Komplemendi vältimise viisid
Bakteri kapsel väldib komplemendi
aktivatsiooni.
Bakteri välispinna molekulid takistavad
fagotsüüdi ligipääsu komplemendile.
lüsosoomi fusiooni või prootonpumpa.
Bakteri katalaas lõhub vesinikperoksiidi.
Äärmiselt tugev välis-membraan
(mükobakter).
Lipoarabinomannaanid blokeerivad IFNde
toime.
Antigeeni esitamise häirimine.
Põgenemine tsütoplasmasse (listeria).
Bakteri molekulid juhivad MAK pinnast
eemale.
Bakteri ensüümid lagundavad kinnitunud
komplemendi.
Bakteri pind ei lase MAK kinnituda.
Bakteri poolt sekreteeritavad “peibutis-
molekulid” seovad komplemendi lahuses.
Antikehade vältimise viisid
Tekivad antikehad, mis blokeerivad efektiivseid antikehi
Bakteri proteaas lõhub sIgA
Bakterimembraani tükid adsorbeerivad antikehi
Pinna-antigeenide variatsioon: LPS sialüülimine; faasivariatsioon, ripsme valkude
variandid geneetilise rekombinatsiooni tulemusel.
51.
Infektsioonhaiguste korral esinevate immuunreaktsioonide diagnostika.
52.
Kasvajate immunoloogia üldised põhimõtted.
1. Kasvajat infiltreerivate T rakkude suurem hulk korreleerub munasarja vähki põdevate
patsientide parema elulemusega
2. Immuunsupresseeritud isikutel on teatud tüüpi kasvajaid
rohkem kui terve
immuunsüsteemiga isikutel
3. Kasvaja ülekanne organi siirdamisel doonorilt retsipiendile.
Immuunseires osalevad loomuliku ja omandatud immuunsuse mehhanismid
Molekulid: Tsütokiinid (IL-12, interferoonid γ, α/β); Retseptorid (NKG2D, MHCI);
Tsütotoksilised molekulid (perforiin)
Leukotsüüdid: NK rakud, M1 makrofaagid, dendriitrakud, CD4+ and
CD8+ T rakud
NK rakkude roll kasvajate vastases kaitses
NK rakud võitlevad hematopoeetiliste kasvajatega, võimalik et ka metastaseeruvate
rakkudega tsirkulatsioonis. Kui tuumorirakk on MHC molekulide ekspressiooni kaotanud,
aktiveeritakse NK rakud. Kasvaja mikrokeskkonnas on hulgaliselt inhibeerivaid faktoreid.
Kui aktiveerivad signaalid on ülekaalus, aktiveeritakse ka NK. (stress)
CD8+ T rakud kasvajavastases kaitses: kasvaja antigeeni esitletakse T
aktivatsioon CTL infiltreerub kasvajasse Tunneb kasvajarakud hävitab
kasvajarakud. CD4+ T rakud abistavad CD8+: IFN-γ (NK aktiveerimine, M1 makrofaagide
toetamine).
Makrofaagid ja kasvajad
Makrofaagid suudavad olenevalt tüübist kasvajaid nii
inhibeerida kui ka nende kasvu
soodustada. Osalevad koekahjustuse erinevates faasides. M1 Äge faas – et eemaldada
koekahjustuse põhjus. M2 Paranemisfaas – koe regeneratsioon.
(IFN γ, LPS, TLR-L) ⇒
M1: MHC II , CD68, CD80, CD86, IL-6, IL-12, IL-23, TNF-Α ⇒
pro-
inflammatoorsed, tsütotoksilised,
antikasvaja!
IL-4-, IL-13 ⇒
M2: CD163, CD200R, MGL-1, MGL-2, IL-10, TGF-Β ⇒
anti-
inflammatoorsed,
prokasvaja!
IL-4, IL-10, IL-13 ⇒
TAM: (tuumor induced macrophage) – CD163, IgG Fc fragment, CTL
domään, kuumašokk
prokasvaja
Põletik ja kasvajad:
Kasvaja indutseerimine. Põletikurakkudest vabanevad hapniku vabad radikaalid (ROS)
põhjustavad naabruses asuvates epiteelirakkudes mutatsioone. Tsütokiinid omakorda
soodustavad vabade radikaalide vabanemist ja epigeneetilisi muutusi.
Kasvaja soodustamine. Põletikutsütokiinid aktiveerivad transkriptsioonifaktoreid
NF-
κBB (TNF-α) ja
STAT3 (IL-6) kasvajaliseks muutuvates rakkudes. Need soodustavad
proliferatsiooni, angioneogeneesi, inhibeerivad apoptoosi ning indutseerivad üha uute
põletikurakkude invasiooni. Lisaks indutseerivad need TF kemokiine, mis tõmbaks ligi
veel rohkem T rakke, MF ja DC.
Immuunseire:
53.
Immuunreaktsioonide vältimine kasvajarakkude poolt. Kasvajate
hoidumine immuunsüsteemi rünnaku eest
1. Ebapiisav immuunvastuse induktsioon
a. Negatiivne selektsioon tüümuses
Paljud
kasvaja-antigeenid, mida T rakud võiksid ära tunda, on väga sarnased
normaalsetele
auto-antigeenidele: sellised T rakud on enamasti juba tüümuses
elimineeritud/madala afiinsusega.
b. Kasvajatel puudub eriti kasvu algfaasis “ohu signaal”, dendriitrakud ei küpse,
puudub kostimulatsioon ning T-rakud toleriseeritkse. (T raku aktiveerimine
nõuab 3+ signaali)
c. Kasvajale spetsiifiline epitoope puudub või pole nähtav
i. Punktmutatsioonid onkogeenides, mis
tekitavad tugevaid muutusi
valgu funktsioons ei pruugi tekitada ühtegi uut epitoopi
ii. MHC molekulid kaovad kasvajarakkudelt
d. Immuunsüsteemi selektiivne mõju: soosides mutatsioone, mis on vähem
immunogeensed.
Immuunsüsteem ise viib sellise selektsioonini, et rakud, mis on vähem
immunogeensed jäävad ellu „tugevas“ organismis – paradoks, et kasvajad
tekivad ka tugeva immuunsüsteemiga organimsis
Cancer immunoediting: 3E
Eliminatsioon: Immuunsüsteem leiab üles ja hävitab arenevad
kasvajarakud enne, kui tekivad kliinilised nähud
Tasakaal (equilibrum): Immunogeensed kasvajarakud endiselt
hävitatakse, kuid tasapisi selekteerub välja teatud hulk rakke, mis on
juhuslike mutatsioonide tagajärjel kaotanud oma immunogeensuse.
Pääsemine (escape): selliselt selekteeritud rakud jäävad hävitamata,
kasvavad ja tekitavad enda ümber immuunosupresseeriva keskkonna.
Kliiniliselt avaldunud kasvaja kujunemine
2. Aktiivne immuunvastuse inhibeerimine
a. Immuunsupressiivsete tsütokiinide tootmine (IL-10, TGFβ, prostaglandiinid)
b. Metaboolne konkurents – kasvajakoes on toitainete nappus pärsib
immuunrakkude funktsiooni. Kasvajarakud vabastavad immuunsupresseerivaid
metaboliite ⇒ T rakkude aktivatsiooni pidurdavad signaalid
c. M2 makrofaagide indutseerimine immuunsupressioon (IL-10, TGF-β),
invasioon ja metastaasid (MMP), angiogenes, lümfangiogenees (VEGF)
d. Regulatoorsete T rakkude genereerimine
i. CTLA-4 pinnal. FOXP3. Inhibeerib CD8+, CD4+, B, APC, NK rakke. ☹
ii. Veel rohkem eelmisi samme selle tõttu.
e. Immuunsüsteemi kontrollpunktide kaaperdamine kasvaja poolt – T rakkude
aktivatsiooni pidurdamine!
i. Vajalikud autoimmuunsuse vältimiseks
ii. Inhibeerivad aktivatsioonisignaali
iii. Soodustavad „kurnatud“ T rakkude arenemist
iv. Kasvajad kaaperdavad selle loomuliku tolerantsuse mehhanismi
v. Tuntuimad
kontrollpunktid: CTLA4 ja PD1
54.
Kasvaja-antigeenid.
Kasvaja-antigeenid: nende klassifikatsioon baseerub
ekspresioonimustril.
Kasvaja-spetsiifilised antigeenid: antigeenid, mida leidub
kasvajarakkkudes, aga
mitte normaalsetes rakkudes; nad võivad olla unikaalsed ühele kasvajale või esineda
mitmes erinevas kasvajas. Kõrge spetsiifilisus kasvajale
Kasvaja-seoselised antigeenid: kasvaja-antigeenid, mida leidub ka normaalsetes
rakkudes. Neil on madal spetsiifilisus kasvajatele
Onkogeensetest viirustest pärit epitoobid – tekivad mutatsioonide järel. DNA
viirustest – HPV, EBV, HPV, RNA viirustest – HLTV1, HCV.
Muteerunud p53 (tuumori supressorgeen)
Ras mutatsioonid (onkogeen)
55.
Kasvajate immunoloogilise ravi võimalused. – immunoteraapia
Kostimulatoorsete signaalide CTLA-4 ja PD-1 blokeerimist kasutatakse kasvajate
immuunoteraapias.
1. Mittespetsiifiline immunoteraapia (tsütokiinid, BCG vaktsiin)
o Interleukiinid: IL-2, IL-12 – T- ja NK rakkude aktivatsioon
o Interferoonid: antiproliferatiivne toime, NK rakkude ja makrofaagide aktivatsioon,
MHC ekspressiooni tõus
o Kasvufaktorid: GM-CSF et indutseerida kasvajarakkude diferentseerumist ja
müeloidsete rakkude toodangut luuüdis
2. Passiivne immuunteraapia monokloonsete antikehadega
o (Rituximab: anti- CD20 non-Hodgkini B rakulise lümfoomi raviks)
o Humaniseeritud antikehadega
o Antikehadest sõltuv tsütotoksilisus – ravim on antikeha küljes
o Radioimmunokonjugaadid ja antikeha-toksiini konjugaadid
o Angiogeneesi inhibeerimine: antikehad VEGF või VEGF-R2 vastu
3. Terapeutilised vaktsiinid – on olnud palju lootusi...
o Eesmärk on immuunrakke stimuleerida. Sipuleucel-T on ainus registreeritud
vähivaktsiin (FDA 2010). Väljaspool keha toodetakse APCsid ja pannakse kehasse
tagasi.
Vaktsiinid pole edukad olnud, sest vähi mikrokeskkond on tugevasti
immuunsupresseeriv: seal toodetakse immuunsupressiivseid tsütokiine, seal on T
rakkude aktivatsiooni pidurdavad signaalid ning indutseeritakse protuumorigenseid M2
makrofaage+ T rege mis suruvad immuunvastust alla.
Onkolüütilised viirused
Sisenevad retseptorite kaudu, mis vähirakkudel on üleekspresseeritud. Kasutavad ära
asjaolu, et viirusevastased mehhanismid on kasvajates immuunsüsteemi eest
peitumise käigus inhibeeritud. Viirusi modifitseeritakse paljunema eelkõige
kasvajarakkudes ning ekspresseerima lisastiimuled immuunsüsteemi aktiveerimiseks.
Manustatakse otse kasvajakoesse. Esimene onkolüütiline viirusvaktsiin registreeriti
2015.a, seni ka ainus.
4. T rakuline teraapia
Kasvajale spetsiifiliste T rakkude paljundamine ex vivo. Kimäärne antigeeni retseptor.
Personaalne ning suhteliselt eksperimentaalne.
CD19 spetsiifiline CAR. CD19 on B
rakkudele iseloomulik pinnamarker, mis ekspresseerub pidevalt B raku arengu käigus.
Seetõttu on ta olemas ka enamusel
B-rakulistel kasvajatel. Esimesed CAR-põhised
terapeutikumid on suunatud just CD19 vastu. Ravi tagajärjel kaovad ka normaalsed B
rakud. Asendusravi immuunoglobuliinidega. 300-400 000 $ maksab ravi.
5. Kontrollpunkti inhibiitorid (Immune checkpoint blockade)
o Koos TCR signaaliga saab T rakk pidurdavaid signaale (kontrollpunktid) ja
aktiveerivaid signaale (kostimulatoorsed molekulid). Ühed pidurdavad
immuunvastust, teised tugevdavad.
o Checkpoint valgud (CTLA-4, PD-1) kontrollivad immuunvastuse teket, vajalikud
autoimmuunreaktsioonide vältimiseks – vähirakud kasutavad neid
immuunvastuse pärssimiseks
o Checkpoint inhibiitorid takistavad T-rakkude aktivatsiooni allasurumist ⇒
aktiveerivad kasvajavastase immuunvastuse
o Vajalikud autoimmuunsuse vältimiseks. Ka krooniliste infektsioonide korral
koekahjustuse limiteerimiseks. Inhibeerivad aktivatsioonisignaali. Soodustavad
„kurnatud“ T rakkude arenemist. Kasvajad kaaperdavad selle
loomuliku
tolerantsuse mehhanismi
o CTLA-4 blokeeriv antikeha oli esimene kontrollpunkti inhibiitor, mis kasvajate
ravis kasutusele võeti, Ipilimumab (2011)
Peale esmast aktivatsiooni tuleb T raku pinnale inhibitoorne CTLA-4.
Kuigi CTLA-4 ja aktiveeriva CD28 ligand on sama – CD80/CD86 tõrjub
CTLA-4 suurema afiinsuse tõttu CTLA-4 välja.
Priming kontrollpunktis – aktivatsiooni inhibitsioon
o PD-1 blokeerivad antikehad Nivolumab ja Pembrolizumab said loa 2014 ja on
veelgi parema efektiga
Efektor faasi kontrollpunktis – efektor funktsiooni takistamine
56.
Immuunsüsteemi kontroll-punktide kaudne vähiravi. CAR-T eespool
57.
Transplantatsiooniimmunoloogia üldised põhimõtted.
Transplantatsiooni abil asendatakse mittefunktsioneerivad koed või organid
funktsioneerivatega. Enamasti ühelt isikult teisele:
doonor – retsipient. Transplantaadi
sobivus sõltub indiviidide geneetilisest sarnasusest.
Siirata saab verd, luuüdi, südameklappe, nahka, luud, sarvkesta, neer, maks, süda, kops,
pankreas, sool.
Transplataatide tüübid.
Autograft – ühelt kehaosalt teisele (sama inimene)
Isograft – loomad geneetiliselt identsed – võib olla ka monosügootsed kaksikud
Allograft – ühelt isikult teisele
Ksenograft – ühelt liigilt teisele – from monkey to man
Äratõukereaktsioonil olulised:
Peamise koesobivuskompleksi alloantigeenid: MHC molekulid
o I klass: A B C
o II klass: -DP -DQ -DR – identsed HLA geenid õdedel/vendadel sagedusega 1:4,
mittesuguluses olevatel isikutel 1:100 000
o III klass: komplemendi komponendid C2, C4, jm.
Minor histocompatibility alloantigeenid
o Mitmesugused polümorfsed geenid.
o Ka identsete HLA geenide korral tekib äratõuge (aeglasemalt)
Veregruppide antigeenid
HLA alleelid – mõlemalt vanemalt pärit, ekspresseeruvad kodominantselt.
Alloantigeenidele reageerimine otse/kaudne
1. T rakk tunneb ära töötlemata allogeense MHC molekuli siiriku APC-l – kiire reaktsioon
2. Protsessitud allogeense MHC molekuli esitlemine enda MHC molekulile T rakule –
vajab töötlemist
Otsene ära tundmine
Enda MHC esitab võõrast peptiidi T rakule.
Isegi infektsiooni korral on ühe raku pinnal piiratud hulk (tublisti alla 1%) MHC molekule,
mis esitavad sama peptiidi. Tugev seondumine, kuid väike arv kontakte.
Alloreaktsiooni korral on raku pinnal suur hulk MHC molekule, mida tunneb ära sama
TCR. Tugeva reaktsiooni tagab kontaktide rohkus, mis kompenseerib madala afiinsusega
seondumise. Korraga reageerib suur hulk T-rakke: umbes 2% T-rakkudest on
alloreaktiivsed (infektsiooni korral reageerib 100 -1000 korda vähem rakke)
Kõigil T-raku retseptoritel on teatud
nõrk afiinsus oma MHC molekulide suhtes.
(Positiivne selektsioon tüümuses). Iga rakk omab tuhandeid koopiaid MHC molekule ja
kuna seondunud peptiid ei oma alloreaktsiooni korral enamasti tähtsust, siis
loovad
alloreaktiivsed T rakud tunduvalt rohkem kontakte kui seda näiteks
infektsiooni korral (tekitajast pärit peptiidid satuvad korraga vaid vähestesse MHC
molekulidesse)
Paljud alloreaktiivsed rakud on mälurakud ja ei sõltu enam kostimulatoorsetest
signaalidest
Otsene alloreaktiivsus genereerib nii CD4+ kui CD8+ alloreaktiivseid T-rakke
CD8+ rakud ründavad transplantaati otse, CD4+ rakud toodavad
põletikutsütokiine aktiveerides makrofaage ja indutseerides alloreaktiivsete
antikehade teket.
Alloantigeenide kaudne äratundmine Retsipiendi antigeeni esitavad rakud töötlevad ja esitavad doonori MHC molekulide
materjali samal moel, nagu see toimub näiteks infektsioonide korral. Allo-MHC-dest pärit
järjestused satuvad oma MHC molekulide süvenditesse.
Kaudne äratundmine viib eelkõige CD4+ T rakkudest põhjustatud alloreaktiivsusele, sest
alloantigeeni fagotsüteeritakse ning see satub endosomaalsesse süsteemi, mille kaudu
toimub materjali esitamine MHC II molekulide kaudu.
Krooniline äratõukereaktsioon sõltub
kaudsest äratundmisest rohkem kui
akuutne
äratõuge. CD4+ rakud ei ole otseselt tsütotoksilised, vaid mõjuvad põletikotsütokiinide
sekretsiooni ja B rakkude abistamise kaudu.
58.
Hüperakuutsed ja akuutsed äratõukereaktsioonid. Krooniline
äratõukereaktsioon
Üliäge äratõuge (hüperakuutne)
o Minutite jooksul
o Tsirkuleerivad olemasolevad antikehad
o Antikeha seostub transplantaadi veresoone
endoteeliga
o Tromboos,
komplementsüsteemi
aktivatsioon, põletik
o Üliägeda äratõuke ravi puudub
o Saab vältida määrates enne transplatatsiooni
transplantaadi
vastaste
antikehade
olemasolu: Komplement
lüüs
sekundaarne antikeha
o Antikehad
Äge äratõuge
o Päevade jooksul
o CTL ja TH
Krooniline äratõuge
o Kuid ja aastaid
peale
transplantatsiooni.
o Aeglaselt
progresseeruv.
o Arerioskleroos ja
sulgus.
o Tsütokiinide toime.
o Puudub ravi
59.
Kroonilised äratõukereaktsioonid
60.
Äratõukereaktsioonide ennetamise võimalused.
Äratõuke vältimiseks:
Perekonnasisene siirdamine
o identsete kaksikute vahel
äratõuget ei esine;
o hea sobivus on tõenäolisem
lähisugulaste puhul;
ABO veregruppide määramine
HLA I/II tüpiseerimine
Cross-match – olemasolevate HLA-
vastaste antikehade määramine)
Tolerantsuse saavutamine – „Püha
Graal“
Üliägeda ennetamine: määrates enne
transplatatsiooni
transplantaadi
vastaste antikehade olemasolu
Äge: immunosupressiivne ravi
Kroonilisele puudub ravi.
Ravimid äratõukereaktsiooni maha
surumiseks:
T-rakkude signaaliradade inhibiitorid – kaltsineuriini inh, rapamütsiin
Tsütostaatikumid – asatiopriin, mükofenoolhape
Lümfotsüütide vastased monoklonaalsed antikehad – anti CD3, -CD25, -CD52
Kostimulastiooni blokeerimine – CTLA4-Ig
Põletikuvastased ravimid
o Nende kombosid kasutatakse, et iga üksiku ravimi doosi ja kõrvalnähtusid
vähendada.
Immunosupressiooni tagajärjel tõuseb vastuvõtlikkus infektsioonidele ja
viirus-seoseliste kasvajate tekkimise tõenäosus
Otsene toksilisus, mis pole seotud immunosupressiooniga. Mõnikord on
ravim toksiline sama koe suhtes, mida transplanteeriti: nt. Tsüklosporiin
neerutuubulite epiteelile.
61.
Äratõukereaktsioonide immunoloogiline diagnosika.
62.
Hematopoeetiliste tüvirakkude siirdamine ja äratõukereaktsioonide
iseärasused. =luuüdi siirdamine
Leukeemiate puhul tehakse. Leukeemia puhul on toimunud geenimutatsioonid, mis
mõjutavad hematopoeetilisi tüvirakke – adenosiini deaminaasi puudulikkus, SCID,
hemoglobiini defektid.
Autoloogne - peale kiiritamist ja/või kemoteraapiat transplanteeritakse oma vereloome
tüvirakke. Äratõuke ohtu pole.
Allogeenne - väga oluline on MHC võimalikult täpne sobitamine, muidu tõugatakse
transplanteeritud rakud ära
Probleemid: GVHD ja immuunpuudulikkus peale transplantatsiooni – graft vs host disease
GVHD:
Luuüdi sisaldab ka küpseid lümfotsüüte – siirdatud koe reaktsioonid retsipiendi kudede
vastu
Akuutne: naha, maksa ja seedetrakti epiteeli kahjustus Kliiniliselt palavik, lööve,
kõhulahtisus. Võib olla fataalne.
Krooniline: tekib > 3 kuud peale luuüdi siirdamist ja meenutab naha, kopsude, maksa ja
soolestiku autoimmuunkahjustust; esinevad autoantikehad, immuunkompleksid. Kujuneb
fibroos ja koe atroofia.
Kui transplantaadist T-rakud elimineerida, siis GVHD oht väheneb, kuid jääb ära ka efekt
transplantaat leukeemia vastu. Samuti implanteeruvad tüvirakud ilma T-rakkudeta
kehvemini.
63.
Vereülekande immunoloogia.64.
Immunotehnoloogia võimalused
Humaniseeritud Monoklonaasled antikehad – hiire CDR3 – liidetakse inimese Ig-le –
oluliselt väiksem immunogeensus
Afiinsuskromatograafia
ELISA (immuunsadestamine)
-
Rakulüsaat/valgusegu
-
Inkubatsioon koos antikehadega
-
Pesu
-
Analüüs
-
Sandwich elisa: sekundaarne antikeha, detektsiooni antikeha, capture antibody.
Fluo märgis.
Antigeeni epitoopide identifitseerimine
• ELISA rekombinantsete või sünteetiliste peptiididega
• Biosensorite abil (immobiliseeritud antikeha, sünteetiline/rekombinantne peptiid)
• ProteinChip® tehnoloogia
Immunoblott
Fluorestsentsmärgistega antikehade kasutamine
Immnohistokeemias, voolutsütomeetrias (lümfotsüütide klasside ja arengustaadiumi
määramiseks; sorteerimiseks CD-de järgi).
Rakuline immuunteraapia: autoloogne, allogeenne, keemiaravi, geeniteraapia
-
DC ülekanne
-
T rakkude ülekanne
-
NK rakkude ülekanne
Vähi immuunteraapia
Alloreaktiivsete NK rakkude ülekanne
Geeniülekanne – transfektsioon. Plasmiid, kus on geen, integratsioon kromosoomi.
Tekib ka episoom (plasmiid + kromosoom – geeniülekannet pole toimunud). Geeni
suurus ei ole limiteeritud. Töötab k6igis rakutüüpides mingil määral. Põhiliselt
episomaalne – in vivo ebaefektiivne
Transduktsioon – geeniülekanne viirusvektori abil – integreeruv või episomaalne –
geenisuurus piiratud + rakutüübi spetsiifiline + vektori spetsiifiline
Viirusvektorid: AAV, AdV, HSV-1, SV40, Alphavirus, Retrovirus, Lentivirus
perekonnad.
Geeniülekande ohud: palju – integratsioon tüvirakku.
65.
Immunotehnoloogilised võtted kliinikumis
66.
Immuunpuudulikkuse sündroomide klassifikatsioon. Tekkemehhanismid.
Immuunpuudulikkus või immuundefitsiit (ID) – organismi kaitsemehhanismide häire, mis
on tingitud immuunsüsteemi mingi lüli defektist – elund, rakk või nende produkt. Häire
võib olla nii ühes kui mitmes immuunsüsteemi osas. Kliiniline pilt: ID väljendub korduvate
või püsivate infektsioonidena. Samas võib kaitsemehhanismide häire aga olla tingitud ka
patogeeni võimest vältida või alla suruda immuunvastust (vt infektsioon).
Immuunpuudulikkushaiguseid jaotatakse:
Primaarsed e kaasasündinud – kaasasündinud immuunsüsteemi rakkude defekt(id),
enamuses geneetiliselt determineeritud
Sekundaarsed e omandatud – sünnijärgsed välisfaktorites (ravimid, kiiritus,
nälgimine, infektsioonid) tingitud immuunsüsteemi rakkude defektid või funktsiooni
häired. Infektsioonid, AIDS, kroonilised põletikud, vaegtoitumus, pahaloomulised
kasvajad, ravim-indutseeritud ja radiatsioonist indutseeritud.
Esinevad ka vanuselistest iseärasustest tingitud immuunpuudulikkuse seisundid
(lapseeas, raugaes).
Primaarsed immuundefitsiidid:
Neid ei esine eriti sageli – keskmine esinemissagedus 1:500 sünni kohta. 300+
geenitdefekti kirjeldatud, rohkem kui 400 PID vormi põhjustajateks. Arstid on puudulikult
informeeritud ID kliinilistest tunnustest ja diagnoosimise võimalustest → diagnoosimine ja
ravi venib.
1.
Geneetilised
Autosomaalsed kromosoomi defektid (retsessiivsed/dominantsed)
Sookromosoomi defektid (nt X-liitelised)
Üksikute geenide mutatsioonid
2. Biokeemilised ja metaboolsed
Adenosiindeaminaasi defitsiit
Puriini nukleosiidfosforülaasi
defitsiit
Biotiin-sõltuva karboksülaasi
defitsiit
Membraani glükoproteiidide
defitsiit
3. Embrüogeneesi häired – loote
arenguhäire tõttu on lisaks muule
häiritud ka IS mingi osa areng
Sekundaarsed
immuundefitsiidid:• Toitumisdefitsiidid ja ainevahetushäired
o alatoitlus, valgudefitsiit (nt enteropaatia)
o vitamiinide ja mineraalainete defitsiidid (biotiin, B12, tsink jt)
o suhkrutõbi
o ureemia jt kroonilised neeruhaigused
o nefrootiline sündroom
• Eksogeensed tegurid (toksilised defitsiidid)
o narkomaania ja alkoholismi foonil tekkiv ID
o ioniseeriv kiirgus
o ravimite tagajärjel tekkivad ID-d (immunosupressandid, tsütotokslised ravimid,
antilümfotsütaarne globuliin)
• Nakkused, infektsioonid
o bakteriaalsed (tuberkuloos, leepra jt)
o viiruslikud ( HIV, punetised, leetrid, CMV, EBV jt)
o algloomad (toksoplasmoos jt)
o seennakkused ( Candida, Aspergillus jt)
• Kasvajad –
peamiselt lümfisüsteemi tuumorid – plasmotsüstoom,
makroglobulineemiad, krooniline lümfoleukeemia, äge leukeemia, mitte-Hodgkini
lümfoom, Hodgkini lümfoom)
• Immuunrakkude kaotus – splenektoomia, verekaotus jne
• Traumad, põletused
• Transfusioonide järgsed defitsiidid (koesobivuskonfliktiga seotud)
• Autoimmuunhaigustega seotud (passiivne, aktiivne, autontikehadest sõltuv (IFN-γ
vastased), rakuline)
Enamusel juhtudest on sekundaarne ID ajutise iseloomuga ja põhihaiguse paranedes või
kahjustava teguri kadumisel immuunsus normaliseerub.
Mis kaasneb ID-ga?
Vererakkude arengu- aktivatsiooni või efektormehhanismide häied ⇒ suureneb
vastuvõtlikkus infektsioonidele, kasvajate sagedus (onkgeensed viirused, vererakkude
kasvajad jne), sagenevad autoimmuunhaigused
Immuunpuudulikkuse kliinilised tunnused:
Sagedased ja väga kahtlust äratavad
sümptomid:
Krooniline infektsioon
Sageli korduvad infektsioonid
Ebatavaline infektsiooni tekitaja
Mittetäielik paranemine infektsioonide
vaheajal v. puudulik vastus ravile
Sagedased kuid keskmiselt kahtlased
sümptomid:
Nahalööve (ekseem, kandidoos)
Krooniline kõhulahtisus
Kaalulangus
Maksa-põrna suurenemine
Korduvad abstsessid
Korduv osteomüeliit
Autoimmuunsed nähud
Ravi:
Infektsioonivastane,
immuunasendus
(IgG,
komplement,
ensüüm,
tsütokiinid/kasvufaktorid, luuüdi transplantatsioon, geeniteraapia).
Keda uurida ID suhtes?
Imikud peredes, kus on esinenud primaarsed ID
Väikelapsed, kelle õel või vennal on kindlaks tehtud primaarne ID või kahtlustatakse
seda
Väikelapsed sündroomide või haigustega, mis võivad kombineeruda ID-ga
Väikelapsed, kelle areng on pidurdunud või kellel esinevad ebatavalised nakkused,
nahalööbed, püsiv kõhulahtisus
Haiged retsidiveeruvate või püsivate nakkustega, mis ei allu antibiootikumravile
(kõige sagedamini haiged retsidiveeruvate sinopulmonaarsete infektsioonidega või
krooniliste obstruktiivsete kopsuhaigustega)
Haiged retsidiveeruvate nahainfektsioonide, abstsesside, periodontiidi või ebatavalise
haavade paranemisega (fagotsütoosihäire kahtlus)
Haiged retsidiveeruvate Neisseria infektsioonidega või süsteemse
erütematoosluupusega (komplemendi defitsiidi kahtlus)
Elusvaktsiiniga vaktsineerimisjärgsete reaktsioonidega imikud ja väikelapsed
Immuunpuudulikkuse klassifikatsioon
:
Kombineeritud immuunpuudulikkus
Antikehade e B-rakuline (nt Brutoni tõbi) – peamine
T-rakuline (nt DiGeorge sündroom)
Teiste arengudefektidega kombineeritud ID vormid
Komplemendi puudulikkus (nt C1 inhibiitori puudulikkus)
Fagotsütoosi puudulikkus (nt krooniline granulomatoostõbi)
Immuunpuudulikkuse eri vormid võivad tekkida rakkude
diferentseerumishäiretest (B
ja T), komplemendi defektidest ja fagotsütoosi defektidest.
ID komplikatsioonid:
Autoimmuunnähud
Kasvajad
Kudede remodelleerumine
67.
Omandatud immuunpuudulikkuse sündroom - AIDS. AIDSi korral tekkivad
immunoloogilised muutused, nende dünaamika. AIDSi immunodiagnostika.
Omandatud immuunpuudulikkuse sündroom ( AIDS )
Üldiselt: põhjustab HIV (inimese immuunpuudulikkuse viirus) infektsioon. HIV on
retroviirus, mis replitseerub immuunsüsteemi rakkudes. Viiruse tsükkel on sõltuv
aktiveeritud T-rakkude transkriptioonifaktoritest. HIV infitseerib CD4+ rakke,
makrofaage, dendriitrakke. Viirus siseneb rakku teatud kemokiinide retseptorite abil.
HIV nakatunud inimestel esineb 4 perioodi haiguse kulus:
Esimesel 4-8 nädalal pärast nakatumist ilmneb tavaliselt gripisarnane haigus,
viiruse tase võib järsult tõusta, tõuseb ka HIV-spetsiifilise CTL-i tase , HIV p24
antikehade teke, HIV Env antikehade tase.
Asümptomaatiline faas. Tavaliselt 2-12 aastat (võiks olla ka 24-25 aastat, enne
1982.a haigestunud inimeste kohta pole andmeid). HIV Env ja HIV p24 antikehade
tase püsivalt kõrge , enam-vähem püsival tasemel ka HIV spetsiifilise CTL. Viiruse
tase plasma on headel juhtudel (ravimid) kergelt kõikuv ja madal, kuid võib olla ka
kõrgemal tasemel.
Pre-AIDS-i periood, sümptomaatiline faas, 2-3 aastat. Viiruse tase hakkab tõusma,
antikehade ja CTL tase langema.
AIDS, 0-1 aastat, viiruse tase tõuseb, muu langeb, surm.
I faasis langeb CD4+ tase järsult, seejärel tõuseb veidi, kuni langeb aeglaselt
nullini – surm.
Mõningaid HIV infektsiooni iseloomujooni:
HIV paljuneb kiiresti kõigis infektsioonifaasides
infitseerib ja kahjustab eelkõige limaskesta T-helpereid (aktiivsed, kõrge CCR5), aga
mitte ainult
infitseerumine ja kahjustus sõltub T raku aktivatsioonist ja kemokiini retseptorite
olemasolust
HIV on tsütotoksiline T rakkude suhtes, mitte aga makrofaagide ja dendriitrakkude
suhtes (püsiv depoo)
HIV „kirjutab“ end peremehe genoomi DNA-sse (proviirus)
veres on viirusest väike osa (enamus on rakkude pinnal immuunkompleksina või
limaskestas)
HIV infektsioon püsib ka tugeva rakulise ja humoraalse immuunsuse foonil
HIV- on mitmeid IS rünnaku eest kõrvale „hiilimise“ mehhanisme
enne AIDS-i teket muudab viirus oma variandi lümfotsüüditroopseks ja hakkab
hävitama ka naiivseid T rakke
T-helperite languse põhjus
T rakkude lagunemine
o viiruse otsene tsütopatogeenne toime (infitseeritud, aga puhkavad rakud ei
tooda viirust)
o infitseeritud raku apoptoos
o T-tsütotoksilise raku rünnak
o infitseerimata raku apoptoos viiruse antigeeni toimel
o enne otsest T rakkude arvu langust tekib tõsine funktsiooni häire (eriti
mälurakkudes)
T rakkude taastootmise häired
HIV kui limaskesta haigus
Infitseeritakse eeskätt soole ja reproduktiivsüsteemi limaskest. Tekib limaskesta
kahjustus.
Limaskesta hakkab läbima enam antigeene ( s.h toidu Ag) ja aktiveeritakse enam T
rakke
Limaskesta aktiveeritud T rakud infitseeruvad ja hävivad
Immuunkaitse langeb ja tekivad limaskestade oportunistlikud infektsioonid
Infektsioonid süvendavad limaskesta kahjustust
HIV progresseeumist soodustavad:
o põletikulised soolehaigused
o teised seksuaalselt levivad haigused
o toidu allergeenid?
Diagnostika
Serokonversioon on pöördepunkt, kus antigeen seerumist kaob ja asemele ilmub vastav
antikeha. Infektsiooni alguses esineb antigeneemia (vireemia e viirusveresus),
antikehade tootmine ei ületa viiruspartiklite tootmist ja tekkivad antikehad tarvitatakse
viiruspartikliga immuunkompleksi moodustamiseks. Serokonversiooni ajal on oht, et
haigust ei diagnoosita, sest diagnostilised antikehad on negatiivsed (alla määramispiiri).
Diagnostilised markerid
HIV antikehad (HIV 1 / 2 antigeenidel põhinevad testid)
primaartest: nt ELISA rekombinatsete viiruskapsiidi valkude ( nt p24, gp120, gp41)
alusel
positiivse tulemuse kontroll: nt Western blot meetod
HIV antigeenid verest
p24 antigeneemia enne serokonversiooni ja AIDSi staadiumis
Prognostilised markerid:
CD4+ rakkude hulk langeb ja CD4+ / CD8+ suhe langeb p24 antikehade tiitri tase langeb
– p24 antigeneemia - AIDS
68.
Teistest viirusinfektsioonidest tingitud immuunpuudulikkused (COVID-19
jt.).
69.
Komplementsüsteemi ja fagotsütaarse funktsiooni puudulikkuse
sündroomide iseloomustus ja diagnostika.
Komplemendi puudulikkused
Jaotatakse kolme suurde klassi:
1. Komplemendi komponentiede puudulikkus
a. Pärilik angioneuriootiline ödeem (Quincke ödeem)– C1 esteraasi inhibiitori
puudulikkus, kõige tavalisem komplemendi defitsiitsus – veresoonte
dilatatsioon ja läbilaskvuse tõus, tursed. Kaasasündinud vorm ja omandatud
vorm. Ravi värske plasmaga, C1 inhibiitori preparaadiga (asendusravi) või
androgeensete hormoonidega (stimuleerib inhibiitori sünteesi)
2. Kontrollvalkude deftitsiit
3. Komplemendi retseptorite puudulikkus
Primaarsed puudulikkused on harvad. Esineb kliiniliste alavormide paljusus olenevalt
puuduvast või mittefunktsionaalsest komplemendi komponendist
Iseloomulik on bakteriaalsed nakkused ja immuunkomplekside elimineerimisest tingitud
häired. Suureneb vastuvõtlikkus Neisseria meningitidis’e, Streptococcus pneumoniae ja
stafülokokkide suhtes
Korduvad naha mädapõletikud, sinusiidid ja meningiidid
Immuunkomplekshaigustest esinevad süsteemne luupus, glomerulonefriit, vaskuliidid
Komplemendi puudulikkuse ravi on tavaliselt (mõõduka kliinilise pildi korral)
sümptomaatiline
Fagotsütoosi puhul:
Neutrofiilid/mononukleaarsed fagotsüüdid arv langeb/düsfunktsioon
Jaotus:
o Primaarsed neutropeeniad
o Neutrofiilide liikuvuse häired
o Fagotsütoosi defektid mikroobide rakusisesel hävitamisel (Fagosoomid, mis
sisaldavad fagotsüteeritud mikroobe, ei „sulandu“ lüsosoomidega, mis
sisaldavad tapja-agente (tegureid) (killing agents). Chediak-Higashi sündroom
Kliiniliselt iseloomulik retsidiveeruvad pikaajalised bakteriaalsed infektsioonid
(sagedamini stafülokokkidest) nahal ja limaskestadel. Komplitseerub sageli mädase
lümfadenopaatiaga. Reageerivad antibiootikumide ravile halvasti
Neutrofiilid ei ole võimelised vastama
kemotaktilisele stiimulile nt komplemendi
poolt vahendatud opsonisatsioonist (laiskade leukotsüütide sündroom) – fagotsüüdi
ekstravasatsiooni ei toimu
Neutrofiilid ei ole võimelised piiristuma spetsiifiliste
adhesioonimolekulide
puuduse tõttu, mille abil seostuda veresoone endoteeliga (leukotsüütide adhesiooni
defitsiit, LAD)
Bakterid ei ole
opsoniseeritud (vt klassikalise komplemendi teed läbi C1-e, C
puudulikkus)
Fagolüsosooomi ei moodustu
Patogeeni lagundamist ei toimu
Krooniline granulomatoostõbi – fagotsüütidel on bakteritsiidse tegevuse häire,
radikaalide tootmine häiritud.
70.
Antikehade puudulikkuse sündroomide iseloomustus ja diagnostika.
Antikehade puudulikkus moodustab enamiku primaarse immuunpuudulikkuse juhtudest.
Antikehade puudulikkuse eri vormidel varieeruvad immunoloogilise defekti suurus ja
vastavad kliinilised sümptomid. Sagedaseim on Iga defitsiit – harvim B rakkude täielik
puudumine.
Kõige raskema antikehade puudulikkuse vormi – aγglobulineemia – korral puuduvad nii
immuunglobuliinid kui ka B-lümfotsüüdid ja esinevad rasked, peamiselt bakteriaalsed
infektsioonid. Samas võivad aga kergemad antikehade puudulikkuse vormid, nagu IgA
või IgG alaklasside puudulikkus, kulgeda ka asümptomaatiliselt.
Immunoloogilistest uuringutest on põhiuuringud:
immuunglobuliinide põhiklassid (IgG, IgA, IgM)
IgG alaklassid
B-lümfotsüütide arv
isohemaglutiniinid
antikehad enne ja pärast vaktsineerimist
spetsiifilised antikehad (nt polüsahhariidide vastu)
Lisauuringud:
o T-lümfotsüütide üldarv ja T-lümfotsüütide subpopulatsioonide arv
IgE, IgD
Kõige suurema informatiivsusega on
immuunglobuliinide põhiklasside määramine.
Nende olulise languse korral aitab immuunpuudulikkuse vormi
diferentsiaaldiagnostiliselt selgitada B-lümfotsüütide arvu määramine. Kergemate
antikehadefektide avastamiseks on oluline IgG alaklasside ja spetsiifiliste antikehade
määramine. Harvem kasutatakse isohemaglutiniinide määramist ja antikehade analüüsi
enne ja pärast vaktsineerimist. T-lümfotsüütide hindamine on oluline nii
diferentsiaaldiagnostiliselt kui antikehade puudulikkuse täpsemaks iseloomustamiseks
seoses immuunsüsteemi teiste osade funktsiooniga. IgD tase on oluline antikehade
puudulikkuse mõningate vormide diferentsiaaldiagnostikas. Geneetilised uuringud on
olulised aγglobulineemia täpsel diagnoosimisel, samuti hüper-IgM-sündroomi
diagnostikas.
X-liiteline agammaglobulineemia – Brutoni tõbi (XLA)
• X-kromosoomiga seotud ID, mis avaldub poistel. Heterosügootsed naised kannavad
geeni. Btk geeni (B cell tyrosine kinase, Brutoni türosiinkinaas) defekt. Geen kodeerib
ensüümi, mis osaleb pre-B raku retseptori signaali ülekandes (vajalik raku
küpsemisprotsessis). Levimus ~1:10000. – B rakude ei küpse
• Perifeerses veres ja lümfisõlmedes B rakud puuduvad. Trakud tavaliselt normis.
• IgG <200 mg/dL, puuduvad ka IgM, IgA ja IgE.
• Sümptomid: rekurrentsed püogeensed infektsioonid (kummalised pneumooniad,
otiidid, dermatiidid, maladsorbtsiooni sdr. (lamblioosist), entsefaliidid (ECHO viirused)
v. autoimmuunhaigus (reumatoidartriidi pilt) 5.-6. elukuul (20%).
• Reageerib hästi immuunoglobuliinide asendusravile.
• Kaugprognoos halb (kroonilised kopsuhaigused ja KNS viirusinfektsioonid).
Selektiivne IgA puudulikkus
Esinemissagedus 1:200-1:800. Kõige sagedamini esinev primaarse immuunpuudulikkuse
vorm, mida iseloomustab IgA madal tase (< 0,05 g/l) või selle puudumine vereseerumis
ja sekreetides. Võib kaasneda IgG alaklasside puudulikkus, teised immunoloogilised
näitajad ei erine oluliselt normi väärtustest.
Multigeenne ja lõplikult selgitamata etioloogia, mitmete immuunsüsteemi tööga seotud
geenide mutatsioonide koosmõju võib viia IgA plasmarakkude
lõppdiferentseerumise
häireni. Limaskestadel ja veres IgA puudub või on väga madal (<0,005 g/l). Võib olla
kombineeritud IgG 2/(4) defitsiidiga.
• Sageli asümptomaatiline. Esinevad korduvad seedetrakti ja sinopulmonaarsed
infektsioonid (mis võivad viia krooniliste kopsuhaigusteni).
• Sagedased allergiad ja autoimmuunhaigused. Kliiniliselt väljendunum kui
kombineerub IgG defitsiit (IgG2 (IgG4)).
• Ravi sümptomaatiline, ka IgG asendusravi. IgA-d ei saa asendada, sest esinevad IgA
vastased antikehad
Üldine variaabel immuunpuudulikkus (CVID)
Esinemissagedus 1:10 000-1:50 000. ilmneb kõige sagedamini 20.-30. eluaastatel.
Iseloomustab immuunglobuliinide kõigi põhiklasside langus, kusjuures geneetiline defekt
ei ole päris selge. Haige sugulastel esineb tavalisest sagedamini üldist variaablit
immuunpuudulikkust. Seos erinevate HLA antigeenidega.
Tekkemehhanismid: – B-raku arengupeetus. – T-raku abistava funktsiooni puudulikkus. –
Autoantikehad T- ja B-rakkude vastu.
Põhitunnused: hüpogammaglobulineemia; immuunglobuliini põhiklasside IgG, IgA, IgM-i
langus, antikehade sünteesivõime langus; sagedased (hingamisteede)nakkused, eriti
bakter- ja seennakkused; varieeruv haigustunnuste ilmnemine varases lapseeas või
täiskasvanul. On kolm alavormi, millest enamikul on B-rakkude defekt. B-rakkude arv
veres on enamasti normaalne, need rakud tunnevad ära antigeenid ja prolifereeruvad,
kui ei diferentseeru plasmarakkudeks. Suureneb pahaloomuliste kasvajate tekke risk.
Immunoloogilised näitajad: -globuliini langus (< 7 g/l), IgG, IgA ja IgM-i madal tase.
• Kliiniliselt sarnane Brutoni tõvele. Sagedased mädased bakteriaalsed nakkused
hingamiselundites ja koljuurgetes, aga ka viirus- ja ja seeninfektsioonid. Võib esineda
ka lamblioosist põhjustatud kõhulahtisus ja malabsorbtsiooni sündroom.
• Iseloomulik kõikide immuunglobuliini põhiklasside langus (IgG <250 mg/dL). B
rakkude arv enamasti normis. Võib lisanduda CD8 T-rakkude arvu tõus. Nõrgenenud
on võime reageerida polüsahhariidseid antigeene sisaldavatele vaktsiinidele.
• Komplitseeruvad sageli autoimmuunsete (reumatoidartriit, hemolüütiline ja
pernitsioosne aneemia, trombotsütopeenia, autoimmuunne hepatiit, alopeetsia jt.) ja
lümfo-proliferatiivsete haigustega (lümforetikulaarsed ja mao-sooletrakti kasvajad).
• Ravi: IgG asendusravi, IL-2 preparaadid konjugeerituna polüetüleenglükooliga,
tüsistuste ravi (infektsioonid).
Veel
• Wiskott-Aldrichi sündroom (WAS)
• Ataksia-telangiektaasia
• DiGeorge’i sündroom – kaasasündinud tüümuse aplaasia (CATCH 22 sündroomi osa)
71.
Kombineeritud immuunpuudulikkus. SCID.
Geenidefektid immuunpuudulikkus (SCID)
seriini/treoniini - spetsiifiliste proteiini fosfaatide kadu (kaltsineuriini toimel aktiveerub
IL-2 geen). türosiini fosfataasi (CD45) kaudu. CD45 isovormid peegeldavad T-raku
seisundit ja nende sünteesiga on seotud lümfotsüütide migratsioon.
o CD45RA+ puhkeolekus (naiivne) Th : L-selektiin "homing" lümfisõlmedes
o CD45RO+ aktiveeritud Th : integriin VLA-4 vaskulaarne endoteel, samuti mälu
mehhanismid
B-raku ja tsütotoksilise T-raku (Tc) aktiveerumine ja mälurakkude teke
immuunvastuse lõppfaas = antigeeni neutraliseerimine (enamasti lisandub
põletikureaktsioon) ja tulem antigeeni neutraliseerimine või eliminatsioon
o Tc rakkude kaudu
o antikehade kaudu (neutralisatsioon, opsionisatsioon, komplement-süsteemi
aktivatsioon, NK-rakkude poolt vahendatud tsütotoksilisus)
o põletikureaktsioon
Immuunvastusega kaasneb põletik, mille esilekutsujaks on põletikumediaatorid
(eeskätt põletiku tsütokiinid, inflammatory cytokines) -
IL-1, IL-6, IFN, IFN, TNF.
NB! Tsütokiin TGF omab põletikku moduleerivat toimet.
Sümptomaatika: alates kuuendast elukuust korduvad ja väga ägedad infektsioonid
väga erinevate ja ebatavaliste tekitajatega (viirused, seened, bakterid, algloomad).
Labor: T- ja NK-rakkude küpsemise häired (arv vähenenud või puuduvad), B-rakkude
tase normis või suurenenud.
Funktsionaalsed häired: immuunsus praktiliselt puudub nii T- kui B-süsteemis:
häiritud T-rakuline immuunvastus, antikehade madal tase kuni täieliku puudumiseni
Th funktsiooni puudumise tõttu Brakule, lümfoidse koe alaareng.
Haiged vajavad kiiret diagnoosi ja agressiivset ravi (kuni luuüdi
transplantatsioonini).
72.
T-rakkude aktivatsiooni ja funktsiooni puudulikkus.
MHC ekspressiooni defektid MHC II ekspressiooni
defektid CD4+ T rakkudes;
defektne rakuline
immuunsus ja T rakust
sõltuvad humoraalsed IV
MHC II geeniekspressiooni
reguleerivad TF
TCR signaali defektid
rakulise IV ja T rakulise ak
vastuse def.
WAS
DiGeorge sündroom
APS1 – APECED
Pärilik haigus. Endokriinnäärmete düsfunktsioon autoimmuususe tõttu. AR pärandumine.
Defekt AIRE geenis.
Sümptomid: krooniline mükotaanne
kandidoos, hüpoparatüreoidism,
Addisoni tõbi, ektodermaalne
düstroofia jms.
Immuuntolerants on häiritud – IFN-α ja
IFN omega reaktsioon + IL-22 vastu
reaktsioon.
MHC I ja II klassi defitsiidid
MHCII puudus on harv ja suremisega lõppev CID.
Autoimmune polyendocrine syndrome
type 1
Other
names
Autoimmune polyendocrinopathy-
candidiasis–ectodermal
dystrophy/dysplasia (
APECED),
SCID
73.
Immuunsüsteemi funktsiooni iseärasused lapseeas. Füsioloogiline hüpo-
γ-globulineemia.
4ndal lootenädalal – granulotsüütide ja monotsüütide eellased maksas – 5.-6. nädalal NK
rakkude teke maksakoes, lootel võib lümfoidset kude tüümuses täheldada esmakordselt
8.elunädalal (B rakud). Seega tekivad loote maksakoes juba 7.-8. nädalal T- ja B-rakkude
eellased. 12.-14. nädalal neutrofiilid, hiljem T-rakud. Peyer’i naastud ilmuvad 5.
raseduskuul, antikehi produtseerivad rakud on lümfisõlmedes ja põrnas kindlaks
tehtavad 20. rasedusnädalal. 6. raseduskuul tekib luuüdi. Immunoglobuliinide
produktsioon embrüonaaleas on minimaalne. Alates sellest algab loote organismis IgM ja
IgD tüüpi antikehade süntees.
Vastsündinul – nagu looteeas – on
loomuliku immuunsüsteemi mehhanismid väga
olulised – komplemendisüsteem mitte, sest erinevatel komponentidel erinev küpsus.
Lapseeas (eriti esimesel kahel eluaastal) toimuvad IS-i funktsioonis väga olulised
muutused.
Sünnimomendiks on vastsündinu vereseerumis IgG antikehade kontsentratsioon
võrreldav täiskasvanu omaga. Kuna IgG läbib platsentat. Enneaegsetel lastel on IgG hulk
siiski oluliselt madalam ema IgG hulgast veres. Kõrge kontsentratsioon on saavutatud
tänu passiivsele transplatsentaarsele ema antikehade ülekandumisele.
Samaaegselt sünniga algab ka IgM antikehade süntees, kui umbes aasta vältel jääb
sünteesikiirus siiski täiskasvanud omale alla. Tunduvalt aeglustunud on ka IgG ja IgA
antikehade süntees ning IgG antikehad saavutavad täiskasvanu taseme alles
kolmandaks eluaastaks, IgA aga isegi hiljem.
Tänu immunoglobuliinide sünteesi aeglusele ja passiivselt omandatud (ka auto-)
antikehade elimineerumisele organismist tekib väikelapse varases eluperioodis
ajavahemik, mille kestel humoraalne immuunsus on nõrgendatud. Eriti ohtlik periood on
alates kolmandast elukuust kuni esimese eluaastani, millal puuduvad emalt saadud
antikehad, organism pole aga suuteline neid ise veel täies mahus tootma. See vastab
lapse eluperioodile, mille vältel ta on eriti vastuvõtlik infektsioonhaigustele <
füsioloogiline (arenguline) hüpoγglobulineemia.
Ka rakuline immuunvastus on vastsündinul pärsitud, hakates täisvõimsusel tööle
alles mõne aasta jooksul pärast sündi. Selle tõttu on täheldatav lümfotsüütide,
sealhulgas T-lümfotsüütide, NK-rakkude suhtelise hulga
langus perifeerses veres. Ka
iseloomustab laste rakulist immuunsust suur immuunrakkude hulga varieeruvus.
Kuna ema komplement ei läbi platsentat on vastsündinul kuni 75% täiskasvanu nivoost.
Eriti puudulik on C8 ja C9 süntees, mistõttu vastsündinud on väga vastuvõtlikud E.colist
põhjustatud infektsioonidele. Alates 11-13 eluaastast sarnaneb lapse immuunsüsteem
täiskasvanu omale, sellel on tihe seos lapse üldise füüsilise ja hormonaalse arenguga.
Füsioloogilise hüpoγglobulineemia kompensatsiooniks on ema rinnapiim. Rinnapiima
immunoaktiivsed komponendid:
Põhjus – CD40L omavate T rakkude vähesus, B rakkude puudulikkus (B1 CD5+ enamus).
Kui testab pikemalt – väikelaste transitoorne hüpogammaglobulineemia.
Sekretoorne IgA (sIgA) – kaitseb eeskätt seedetrakti; valmistab ette võõrvalkusid
sisaldavatele toitudele üleminekuks
T-rakud – sensibiliseeritud; osa aitab immuunsüsteemi stimuleerida, osad on
antikehade suhtes neutraliseerivad
Neutrofiilid ja makrofaagid
Tsütokiinid (põletiku tsütokiinid + IL-10, G-CSF jt)
Lisaks:
Hormoonid
Laktoferriin, lüsosüüm, proteaaside inhibiitorid jne.
74.
Rinnapiima immunoaktiivsed komponendid
Füsioloogilise puudulikkuse kompensatsiooniks ema rinnapiim.
Rinnapiima
kaitsefunktsioon (loomulik, spetsiifiline immuunsus) ja regulatoorne toime:
Sekretoorne IgA (sIgA) ja sIgA antikehad
T-rakud
Neutrofiilid, makrofaagid
Tsütokiinid (põletiku tsütokiinid + IL-10, G-CSF jt.)
sCD14
Hormoonid (insuliin, kortisool, adipokiniinid jt.)
Laktoferriin, lüsosüüm, proteaaside inhibiitorid jt.
Epiteliaalsed kasvufaktorid, kasvufaktorid kommensaalse mikrofloora tarbeks
75.
Immunoloogilised muutused seoses vananemisega. Vanemaealiste
immuunsüsteemi iseärasused.
Sagenevad infektsioonid, nende kulg on raskem
Sagenevad kasvajad
Sagenevad põletikulised protsessid (sh autoimmuunhaigused)
Vaktsineerimine on sageli ebaefektiivne (Nt. HBV, teetanus, gripp)
Samal ajal on kliiniliste analüüside üldnäitajad tervetel vanemaealistel eriliste
iseärasusteta
Tüümuse funktsiooni puudulikkuse tulemusena – asendub rasvkoega
Väheneb „naiivsete” (CD45RA+) T-lümfotsüütide teke ja osakaal perifeerses veres
Väheneb immuunsüsteemi „paindlikkus” (T-rakkude variaabelsus, TCR
retseptoritega rakkude hulk on vähenenud)
Väheneb CD4+ rakkude produktsioon
Puudulik CD8+ efektorrakkude funktsioon
Väheneb regulatoorsete T-lümfotsüütide teke
I ja II tüüpi T-helper-lümfotsüütide omavahelise hulga muutus kutsub esile:
Immunoregulatsiooni häired
Th2 tsütokiinide suurema produktsiooni
suureneb vastuvõtlikkus rakusisestele infektsioonidele
Luuüdi:
Luuüdis täheldatakse CD5+ B-lümfotsüütide (B1 alaklass) hulga suurenemist, mis tingib:
IgM tüüpi antikehade ning madala afiinsuse ja reaktsioonivõimega antikehade tekke
Mitmesuguste autoantikehade tekke
Põhjuseks luuüdi hemopoeetilise võimekuse langus – punane asendub kollasega. Vähem
naiivseid B rakke, vähem mBC-de kloone.
Põhjuseks B-rakkude küpsemise häired (ka klonaalne B-rakkude ekspansioon
monoklonaalsete immunoglobuliinide teke)
Samuti vananemisega:
NK-rakkude hulk ja funktsionaalne aktiivsus suureneb, et kompenseerida T-rakkude
puudulikkust.
o Kemokiinide ja tsütokiinide produktsioon vähenenud
o Reaktiiivsus IL-2 suhtes vähenenud
põletikutsütokiinide TNFa, IL-6 ja IL-1 produktsiooni suurenemine:
o madaldab organismis põletikuliste protsesside
tekke läve
o muudab
põletikulised
protsessid
raskekujulisemaks
AAK esinemine sageneb
Mälurakkude osakaal tõuseb
Suitsetamine, madal füüsiline aktiivsus võimendab seda
kõike!!!
Lisaks: antikehade produktsiooni erinevused vanuritel
Kõrgesse vanusesse jõudnutel väga efektiivne immuunsüsteem.
Milline on pikaealisuse „immunoloogiline allikas”?
Vere CD4+/CD8+ lümfotsüütide suhtarv üle 1.0
Vere NK-rakkude kõrge aktiivsus
Patogeenseid autoantikehi vähe, neid neutraliseerivaid AK palju
Antikehade olemasolu enamlevinud mikroorganismide vastu (sh autoantikehade
puudumine).
76.
Immunoloogilised erinevused meeste ja naiste vahel.
♀ seerumi IgM nivoo kõrgem kui meestel, samuti leitakse neil sagedamini ja kõrgemas
tiitris haigustekitajate vastaseid antikehi, ka autoantikehi ja autoimmuunhaigusi esineb
sagedamini. T reg tase langeb
♂ immuunsüsteemi funktsionaalne aktiivsus on tagasihoidlikum – vähem IL-2 tootmist,
aga IL-1 β ja TNF α produktsioon > kui naistel. Tuleb meessuguhormoonide
immunosupressiivsest mõju. Immuunpuudulikkuse sündroomid esinevad rohkem. X
kromosoomis mutatsioonid tõenäolisemad avalduma. X liiteline a γglobulineemia – B
rakkude defekt. IPEX sündroom – FOXP3 tranksriptsioonifaktori defekt – CD4 T rakud T
reg. See katki.
Reproduktsioon ja immuunsüsteem tihedalt seotud – suguteede haigused ja mitte
infektsioossed haigused (PCOS) mõjutavad, AI haigused mõjutavad – viljakust nt.
Rasedus ja selle kulg mõjutab immuunsüsteemi. reprodktiivimmunoloogia.
Hüpotaalamuse – hüpofüüsi – ovariaal telg
Immuunsüsteemi rakkudel hormooniretseptorid – ja ka neuroendokriin rakkudel tsütokiini
retseptorid. Nihked teljes mõjutavad kogu süsteemi. Suguhormoonid võivad inhibeerida –
progesteroon või aktiveerida autoimmuunseid protsesse.
77.
Toitumise ja toidu iseärasuste mõju immuunsüsteemi funktsioonile.
Rasvumine ja immuunsüsteem.
Alatoitlus on sagedasemaid (globaalsest aspektist vaadatuna) sekundaarse
immuunpuudulikkuse põhjusi. Alatoitluse korral häirub immunokompetentsete rakkude
proliferatsioon ja väheneb valgu süntees, mis viib immuunpuudulikkuse tekkimiseni.
Üldise proteiinide ja kaloraaži
puudulikkuse (primaarne või sekundaarne) tagajärjel:
tüümuse degenereerumine („toitumuslik tümektoomia“)
perifeerse vere T-lümfotsüütide osatähtsuse langus lümfotsüütide seas 15-25%
CD4+ ja CD8+ rakkude suhtenäitaja langus (nagu AIDSi korral)
fagotsütaarse aktiivsuse langus
komplemendi hulga langus
immunoglobuliinide nivoo perifeerses veres on normi piires, vaid sIgA on langenud
Tulemuseks on suur rakusiseste infektsioonide ja kasvajate oht.
Lastel võib puuduliku või mittetäisväärtusliku toitumise korral täheldada kurgumandlite
atroofiat, lümfopeeniat; väheneb T-lümfotsüütide arv ja mitogeenide toimel nende
proliferatiivne aktiivsus; neutrofiilide kemotaksis on normaalne, aga bakteritsiidne
aktiivsus langenud; immunoglobuliinide tase vereseerumis tavaliselt ei muutu, kuid
häirub oluliselt nende süntees (kõige sagedamini täheldatakse sekretoorse IgA taseme
langust).
Toidu üksikkomponentide puuduse tagajärjel:
Fe – madal fagotsütoos ja vähenenud T-rakkude hulk
Zn – lümfoidse koe involutsioon kuni atroofia, tsütotoksiliste reaktsioonide häire
vit. A – rakulise immuunsuse supressioon
vit. E – rakulise ja humoraalse immuunsuse puudus (eriti kui ka seleeni puudus)
Teatud komponentide liigsus viib ka väga mitmesuguste muutusteni immuunsüsteemis.
Muude komponentide mõju toidus.
„caramel food colorant III“ – sfingosiin-1-fosfaadi hulga tõus kudedes → immuun-
supressioon
polüküllastamata rasvhapped (PUFA) – ω-6 rasvhapped (taimeõlid) reeglina
soodustavad põletikku; ω-3 rasvhapped (kalaõlid) reeglina pidurdavad põletikku
glutamiin, arginiin, nukleotiidid – vajalikud ISi kesksete komponentide sünteesiks
probiootikumid – reeglina stimuleerivad eeskätt loomulikku immuunsust
(erinevatel tüvedel erinev toime)
Rasvumine aga – suurendab immuunsüsteemi rakkude hulka. Lipiidide metabolism
kiireneb, DAMP signaalirajad. M1 e
proinflammatoorsed makrofaagid võtavad õle
(kõhnadel on M2 MF ja IL-10).
78.
Immunotoksilised ühendid ja nende mõju iseärasused. Radiatsiooni mõju
immuunsüsteemile.
Immunotoksilised ühendid väliskeskkonnast: pestitsiidid, halogeensed aromaatsed
süsivesinikud (PCB jt.), lahustid (benseen), metallioonid (Ni, Be, Cd, Hg jt.). Neil on kõigil
väga mitmekülgne mõju immuunsüsteemile.
Radiatsiooni mõju iseärasused
Esmalt häired T-rakkude süsteemis (CD4+/CD8+ langus jne.), hiljem ulatuslikumad
muutused.
79.
Närvisüsteemi ja immuunsüsteemi vahelised seosed. Stressi mõju
immuunsüsteemi funktsioonile.
Immuun-, närvi ja endokriinsüsteemi seoste kujunemisel ja toimumisel on väga oluline, et
ühe süsteemi kudede raku oleks võimelised reageerima teiste kudede rakkude
poolt produtseeritud mediaatorile e virgatsainele.
Nt tsütokiinidel on toime vastavate retseptorite kaudu mitte ainult IS rakkudele vaid
närvi ja endokriin rakkudele. Nt Ajukoes on rohkesti tsütokiineretseptoreid. Sama
puudutab ka neurotransmittereid ja hormoone, mis omakorda toimivad IS rakkudele.
Ravi mõeldud tsütokinide või nende retseptoritele mõjutamiseks annab kõrvalnähud ka
närvisüsteemi talitluses. Nt c hepatiiti IFNα ravi võib põhjustada depressivsuse ilmingud.
Immuun, närvi ja endokriin rakkudel on tihe kontakt perefeerses kudes, arvatakse, et
selline kontakt aitab paremaini mobilisatsieerida kaitsemehh välisfaktorite kahjuliku
toime puhul.
Lühiagsel stressil on immuunreaktsioone pidurdav toime. Selle põhjuseks võivad olla
nii psühhogeensed, füüsikalised jm faktorid, aktiveerub hüpotaalamus-hüpofüüsi telg
(HPA-telg), mille tulemusel suureneb ajus adrenokortokotroopse hormooni (AKTH) ja
sümpaatilist närvisüsteemi stimuleerivate mediaatorite produktsioon. Organismis kasvab
neerupealise hormoonide (kortisoolide) ja katteholamiinide (adrenaliini) hulk. Tulemusel
on põletikuliste protsesside pidurdamine. Kroonilise stressi korral on muutused IS
ülatuslikumad ja enamasti seotud üldise immunfunktsiooni pärsimisega.
Psühhogenne stress: pikaajaline magamatus põhj Th1/Th2 rakkude tasakaalu Th2
aktivatsiooni suunas, suureneb
põletikutsütokiinide (IL-6) spontanne teke. Valu
mehhanismid on näideks kuidas närvi ja immuunsüsteem töötavad koos, akuutse valu
tekkes pärast koekahjustust osalevad põletikutsütokiinid, kemokiinid ja komplemendi
komponendid.
80.
Füüsilise koormuse mõju immuunsüsteemile. Endokriinsüsteemi
interaktsioonid immuunsüsteemiga.
IL-6 tootmine kontrakteeruvate müotsüütide poolt.
Mõõdukas kehaline koormus stimuleerib immuunsüsteemi eeskätt sünnipärase
immuunsuse osas (nt. NK-rakkude hulga ja aktiivsuse tõus, T-rakkude aktiveerumise
lävend väheneb).
Ülemäärase kehalise koormuse (tippvormis sportlased) tulemuseks on immuunsüsteemi
stress.
Kehaline koormus: reageerib sõltuvalt tugevusest ja kestvusest, märkimisväärne osa
ka emotsionaalsel komponendil. Vahetult pärast koormust on enamik loomuliku ja
omandatud immuunsuse funktsiooni näitajaid on supresseritud, mis 24h hiljem taastub
koormuse-eelse tasemeni.
Akuutne kehaline koormus: iseloomulik perifeerse vere leukotsüütide hulga
suurenemine, T rakud reageerivad kergemini nii mitogeenide kui antigeenide
stimulatsioonile. Akuutse koormuse järgset IS depressiooni seostatakse IL-6 hulga
suurenemisega organismis, mis võib olla üheks põhjuseks kortisooli hulga suurenemise ja
sellest tuleneva immuunsupressioni põhjuseks.
Krooniline kehaline koormus: tekib loomuliku ja adaptiivse immuunsuse
reaktsioonides rida muutusi, muutub perifeerses vere NK rakkude funktsioonid ja hulk,
suuresti muutub perifeerse T-rakkude ja vähemal määral B rakkude hulk, häirib
tavapärane tasakaal tsütokiinide produktsioonis. Vähenenud rea TLR-ide ekspressiooni
monotsüütidel, nii tsirkuleerivate Th1 rakkude hulk, kui ka T ja B rakkude funktsionaalne
aktivsus. Ig produktsioonis olulisi muutusi pole. Haigustumised hingamisteede
põletikulistesse haigustesse. Positiivsetest nihketest: suureneb Nk rakkude arv aktiivsus,
põletikku soodustavate tsütokiinide produktsioon piirdub (vähem kroonilisi haigusi).
Paraneb kudede verevarustus mis soodustab immuunrakkude tsirkalatsiooni, füüsilise
aktiivsuse tõttu väheneb ka rasvkude (jällegi seos põletikutsütokiinidega).
81.
Naise organismis raseduse ajal toimuvad immunoloogilised muutused.
Spetsiifilised immunoloogilised muutused, mis kaitsevat loodet ema
immuunsüsteemi kahjuliku toime eest.Naiste immuunsüsteem on seotud menstruaal tsükliga. Kuna sõltub nt östrogeeni
tasemest. Östr raseduse ajal kasvab minimaalseltmaksimaalse tasemeli kuni
platsentani. Östrogeenid indutseerivad B rakkude diferentseerumist plasmarakkudeks.
Stimuleerivad ka T regide teket ja Th2 polarisatsiooni (IL-4) B rakud. Kui Östrogeenide
hulk madal – th1, Th17 reaktsioonid
Prolaktiin – on korraga nii hormoon kui tsütokiin – suhtlus neuroendokriin ja
immuunsüsteemide vahel. Toodetakse ajuripatsis e hüpofüüsis – ka neuronite, emaka ja
rinnanäärme epiteeli rakkude ja naha ja immuunsüsteemi rakkude poolt – Stimuleerib T-
rakkude proliferatsiooni. Aitab toota T-rakkudel tsütokiine IL-1,4,5,6, IFN α
stimuleerivad aktiveeritud B-rakkude jagunemist ja diferentseerumist autoantikehade
tootmine tõuseb. Tolerants kaob prolaktiini tõttu – kui prolaktiin tõuseb raseduse ajal –
rasedus võib katkeda..
Raseduse ajal tekkiv tolerants
Loote suhtes on vaja tolerantsi tekitada, sest loode on emale võõras – tolerants on vaja
enne rasedust tekitada. See algab gameedi tasemel. HLA antigeeni ekspressioon,
tsütokiinid, integriinid, spermatosoidide vastased antikehad, emaka NK rakud. Ema
immuunsüsteemi komponendid mõjutavad munaraku küpsemist ja ovulatsioonid.
Naise-mehe tolerantsus enne rasedust
Oluline kui sarnased on HLA antigeenid. Reproduktsioon püüdleb geneetilise
varieeruvuse poole. Ei tohi liiga erinev ka olla. Sõltub suguteede limaskesta seisundist,
immuunsüsteem regulatsioonist (kui see on vale, ei saa loote suhtes tolerantsus
tekkida), mehe seemnevedeliku seisundist (kui on terve, toodetakse TGF β ja IL 10
põletiku alla suruvaid markereid.
Munasarja corpus lütseumi progesterooni toimed emakale – mõjutab detsidualisatsiooni,
makrofaagide tsütotoksilisus alla, toodetakse IL-10. Mehel – seemnevedeliku roll –
lühiajaliselt immunosupressiivsete tsütokiinide juurdevool, põletiku aktivaatorite
juurdevool (meelitavad makrofaage ja DC, et esitleksid partneri antigeenide esitlus T
rakkudele signaal neile mitte aktiveeruda ja mitte spermatosoide hävitada (kuna on
„rahu aeg“.) HLA I ja II lahustunud isa omad – tolereerivad.
Limaskesta põletik algab tolerantsuse asemel immuunreaktsiooni – kui tavapärane
sugutee – IL-10, TGF-β üles reguleeritud – DC esitlevad T rakkudele antigeeni – muutuvad
deregulatoorseteks. Kui on põletik – aktiveeritud DC suunavad CD4 T rakud Th17 suunas
– soodustavad põletikku ja tolerantsuse kadumist.
Kuidas põhjustab autoimmuunsus viljatust?
Aktiveeritakse autoimmuunsus, reproduktiivorganite vastu suunatud autoimmuunsus
häiritakse partnerspetsiifilise tolerantsi teket ning kahjustatakse reproduktiivorganite
funktsiooni. Reproductive autoimmune failure – endometrioos, autoantikehad, viljatus.
Antiovariaalsed aak – raseduse katkemised, POF; endometrioos, PCOS.
Munasarjade vastu – FSH b alaühik – anti FSH β alaühiku vastu. (folliikuleid stimuleeriv
hormoon) – suurenenud produktsioon viljatuse korral
Endometrioos – emaka kude väljaspool emakat. Infertiilsus, düsmenorröa. – autoimmuun
sündroom kuna on alla reguleeritud T regide ja NK rakkude akt. Suurenenud B
lümfotsüüdid ja AAK.
AEA – anti endometriaalsed antikehad.
ASA – spermatosoidide vastased AAK – munandid on immuunpriviligeeritud –
immuunsüsteemi eest kaitstud. Tüümuses ei presenteerita spermatosoididega seotud
antigeene – seda soodustavad traumad, põletikud. Antikehad spermatosoidide vastu. IgA
seemnevedelikus – takistavad selle liikumist ja munaraku viljastumist.
Raseduse aegne tolerantsi säilitamine
Raseduse ajal programmeeritakse emaka immuunsüsteem rakud ümber kaotades nende
tsütotoksilised omadused, luugakse tolerogeenne, immunosupressiivne keskkond.
Platsenta võtab üle likaalse emaka leukotsüütide käitumise.
Immunoloogilised kaitsemehhanismid naise genitaaltraktis, tolerants spermatotsüütide ja
loote suhtes. Limaskesta tüüpilised kaitse- ja regulatoorsed mehhanismid: sIgA,
intraepiteliaalsed T-lümfotsüüdid (CD8+), subepiteliaalsed T-lümfotsüüdid (CD4+), TGFβ.
Spetsiifilised immunoloogilised muutused, mis kaitsevat loodet ema
immuunsüsteemi kahjuliku toime eest.
Loode on ema organismi jaoks võõras (sest MHC geenid on pärit isast) ja et ema T-
helperid ei saaks loodet ära tunda ja rünnata, ei oma loote rakud MHC I ja II. Koed, mis
omavad vähe MHCI võivad olla rünnatud NK rakkude poolt. Et seda vältida, on
produtseeritud MHC Ib klassi HLA-G (seondub kahe peamiste NK inhib. retseptoritega
KIR1 ja KIR2).
Rasedusse ajal haigused:
Infektsioonid sagenevad ning kulgevad raskemini, autoimmuun taanduvad,
AAK, ja autoimmuun Haigused tekivad ka.
82.
Loote immunoloogilised mehhanismid. Platsenta immunoloogiline
tähendus.
Platsenta on aktiivne immuunorgan!
Platsenta emapoolses osas on
emaka makrofaagid ja mitteküpsed DC –
fagotsüteerivad implatatsioonil tekkinud platsenta prahti, esitavad neid antigeene T-
rakkudele (õpetab tolerantsust loote rakkude suhtes), toodavad aktiveerivaid
tsütokiine (IL-1 β, TNF), ja nende inhibiitoreid – raseduse ajal on rohkem tsütokiinide
inhibiitoreid.
Emaka NK rakud (uNK) – CD56+CD16- (kui 16+ on loote äratõuge). Koloniseerivad
endomeetriumi ovulatsioonijärgselt kuni varase raseduse staadiumini. Vahendavad
platsenta arengut, spiraalarterite moodustumist – minimaalne toksilisus –
supresseeritud lokaalne rakuline immuunvastus (IL-4, IL-5, IL-13, KIR) – ema Th2
immuunreaktsioonid.
Γdelta T – rakuddelta T – rakud – lokaalne immunomodulatsioon.
CD4+CD25+ T regid – toodavad Il-10 ja TGF β – migreeruvad emakasse lümfiteede
kaudu – tagavad tolerantsust.
Trofoblasti rakud – loote rakud
o Toimivad sarnaselt loomuliku immuunsüsteemi rakkudele – TLR 1-10 (lokaalne
infektsiooni vastane kaitse). Steroidsete ja proteiinhormoonide tootmine –
immuunosupressioon – süntsüütiotrofoblast toodab tsütotrofoblastiga peptiide.
o Lootepoolses osas ekspresseerivad
enda pinnale isapoolseid HLA molekule
tolerantsi säilitamiseks.
Emakas olevaid T rakke õpetama mitte hävitama loote rakke.
IL10, IL4, PGE2 produktsioon stimuleerib ema Th2 rakkude hulga suurenemist.
Platsentabarjäär:
1. Lootepoolne tsütotrofoblast – loote klassikaline HLA I ja II
2. Süntsüütiotrofoblast – HLA negatiivne (toodavad HLA G molekule)
3. Hatuväline tsütotrofoblast – loote mitteklassikaline HLA I
Platsentale ainuomase
MHC I klassi HLA-G ekspressioon (alloimmuunne reaktsioon
ema IS-I poolt immuunsupressioon). Isovormid 5 ja 6. Mitteklassikaline HLA.
Vähendavad CD8 ekspressiooni T raku pinnal – CD8 oluline MHC I seondumisega.
Vähendavad põletikumediaatorite produktsiooni (IL-1, TNF). Suurendavad IL-10 taset,
tühistab põletikutsütokiinide tootmise aktiveeritud makrofaagide poolt. Täiesti nulli ei
saa ema immuunsüsteemi viia, kuna ema peab ka terveks jääma.
Pre-eklampsia
Histoloogiliselt ja funktsionaalselt ebapiisav platsenta hattude invasioon
Prekliiniline lokaalne staadium – 6 -18 rasedusnädalal, platsenta invasiooni
puudulikkuse tulemusena suurenenud lagumenine ja nekrootilise platsentaarkoe leke
ema vereringesse:
o Platsenta invasiooni puudulikkus geneetilistest põhjustest ja/või
immunoloogilise partner-spetsiifilise tolerantsi häiretest
o Antifosfolipiidsündroom, -antikehad
o Proinflammatoorsed tsütokiinid tõusevad
o Treg aktiivsus langeb
Kliiniline staadium – antiangiogeensete ja proinflammatoorsete faktorite vabanemise
tõttu stressis platsenta
o Põhjustatud süsteemsest üldisest põletikulisest vastusest, mis erineb
normaalse rasedusega kaasnevast põletikulisest aktivatsioonist vaid
kvantitatiivselt, mitte kvalitatiivselt (III trimester normaalne põletiku
tsütokiinide tõus)
o Ei ole tegu
antigeen -spetsiifilise vastusega loote või platsenta antigeenidele
! peamiselt loomuliku immuunsüsteemi poolt vahendatud (komplemendi
süsteem, verehüübimise süsteem)
o Samaaegselt võimalik autoimmuunne aktivatsioon nekrootilise platsenta lekke
tõttu – hilisemad AI haigused ema
Barjäärfunktsioon immunoloogilistele efektoorsetele olekulidele (v.a. IgG – läbib
platsentat)
Steroidsete (östrogeen, progesteroon) ja proteiinhormoonide (korioon-gonadotropiin:
HCG) produktsioon (→immunosupressioon jms.)
IL-10 produktsioon (stimuleerib ema immuunsüsteemi Th2 poolt vahendatud
reaktsioonide prevaleerimise suunas)
Loote antigeenide mõju ema immuunsüsteemile.
Rh faktor – põhjustab punaliblede opsonisatsiooni ja lõhustumist. D antigeen on Rh
positiivsetel olemas ja Rh neg puudub. Konflikt on siis kui Rh+ loode on Rh- emas.
Ohustab loodet. Immuunsüsteemi supressioon, et ta ei hävitaks loodet. Üks patoloogiline
mõju on reesuskonflikt. Rh- ema ja Rh+ laps loote reesuspositiivse vere D-antigeenid
sattuvad ema vereringesse. Nende toimel on reesusnegatiivse ema organismis tekkinud
anti-D antikehad, mis lootesse jõudes põhjustavad seal antigeen-antikeha reaktsiooni.
Tulemuseks on loote hemolüüs. raviks on loote vereülekannet, konflikti ohu korral
süstitakse emasse anti-D-immunoglobuliine.
Ema antikehade mõju lootele.
Ema ja loote vaheline mõlemasuunaline rakkude ja DNA transport – sünnituse järgselt
püsivad 1: 1 000 000 aastakümneid. Vajalik, et koed saaksid uueneda ning kaitsevad
kasvajate eest. Kui loote ja HLA on liiga kattuvad, või on platsenta moodustamise häired
võivad viia allo-autoimmuunhaiguste tekkimiseni.
Ema IgG läbib platsentat ja satub loote vereringesse. II ja III trimestril – passiivne
immuniseerimine nende haigustekitajate vastu, kellega ema on kokku puutunud.
Imikute plasma IgG tase pärast sündi on sama, mis emal ja sama antigeen spetsiifilisus
diaposooniga. Seega, selle spetsiaalse transportsüsteemi abil saavad lapsed sünni
alguses Ak kõigi tavaliste patogeenide vastu. Ka IgG4 (antikehad allergeenide vastu)
transporditakse lootesse.
Erinevad haigused aak üle kandumisel – neonataalne luupus, Graves’i tõbi, Hashimoto
türoidiit, neonataalne müasteenia
Raseduse mõju emale
Infektsioonhaigused sagenevad, autoimmuunhaigused taanduvad, osad halvenevad –
SLE, müasteenia. Th alapopulatsioonide nihkest – prevaleeruvad Th2 vs Th1 tüüpi rakud (
rasedusel Th1 immuunreaktsioonid alla surutud.)
Viljatusravi võib autoimmuunhaiguse ägenemise, aga immuunsupressiivne ravi
vähendab nt aak haigustel.
83.
Loomulik (kommensaalne) mikrofloora ja selle tähendus immuunsüsteemi
arengus.
Mikroorganismid, mis on keha pindadel ja on „väliskeskkonnaga” kontaktis (seedetrakt,
vagiina, nahk, hingamisteed). Nad on arenenud koos peremehega, aga nad on nö
ületanud peremehe kaitsemehhanismid ning pole patoloogse mõjuga. Kõige suurem osa
kommensaale on sooles. Kommensaalidel on oluline roll immuunsüsteemi töötamises ja
selle arengus. Limaskesta immuunsüsteemil on regulatoorsed ja antiinflammatoorsed
mehhanismid, et tolereerida ohutuid, toidust antigeene ja kommensaaalseid
mikroorganisme.
Limaskesta immuunsusel on tugevad loomuliku immuunsuse mehhanismid: tagavad
limaskesta barjääri töötamise, lümfotsüütide ja nende produktide olemasolu limaskestas,
Ig-de sekreteerimine (sIgA), ja MALTist pärit rakkude migreerumine ja
homing.
84.
Koekahjustuse mehhanismide (ülitundlikkuse) neli tüüpi (Coombs-Gelli j).
Antikehade ja T-rakkude koekahjustuse efektormehhanismid.
Ülitundlikkusreaktsioonid võivad ilmneda immuunsüsteemi aktivatsiooni käigus ja on
kahjulikud. Lisaks immuunsüsteem aktiveerumisele siis kui on vaja kaitsta patogeensete
mikroobide vm välisfaktorite eest. Ülitundlikkusreaktsioonides osalevad
antikehad
ja/või immuunrakud. Ülitundlikkusreaktsioonid on: oma kudede vastu (autoimmuunsus), mikroorganismide
vastu, või väliskeskkonnast pärit mitte-mikroobsete antigeenide vastu (allergia).
4 tüüpi:
I kiiret tüüpi reaktsioonid
II antikehade poolt tekitatud reaktsioonid
III immuunkomplekside e. antigeen-antikeha komplekside poolt tekitatud reaktsioonid
IV rakkude poolt tekitatud reaktsioonid, tuntud ka nn aeglast tüüpi
immuunreaktsioonidena.
Esimesed kolm tüüpi (I-III) reaktsioone on esile kutsutud antikehade, IV tüüp - rakkude
poolt. Kõigis on oluline roll põletikuga seotud rakkudel.
Gell-Coombs järgi:
Tüüp
Kliiniline
väljendus
Reaktsiooni
kiirus
Mehhanism
I (kiiret
tüüpi)
Anafülaksia,
atoopia
minutid
IgE-vahendatud (Th2, nuumrakud
ka olulised)
II
(tsüto-
toksilin
e)
Hemolüütiline
aneemia
Varieeruv
IgG, IgM (opsonisatsioon –
fagotsütoos]
III
Arthuse reaktsioon,
vaskuliit
Varieeruv
(tundides)
IgG ja IgM immuunkomplekside
vahendatud (leukotsüütide
aktivatsioon)
IV
(aeglan
e)
Naha ülitundlikkus-
reaktsioon
12-48 h
Sensibiliseeerunud lümfotsüütide
poolt vahendatud
85.
Allergilise e anafülaktilise (I tüüpi) koekahjustuse (ülitundlikkuse)
tekkemehhanism. Osalevad rakud, rakumembraanil paiknevad retseptorid,
rakkudevahelised mediaatorid ja antikehad. Tsütokiinide tähtsus.
Nuumrakkude aktiveerumisel vabanevad mediaatorid ja nende bioloogiline
toime. Nuumrakkude aktiveerumisel sünteesitavad mediaatorid ja nende
bioloogiline toime.
Allergia tekkepõhjuseks on ülemäärane IgE vastus välisantigeenidele.
Antigeen (allergeen) siseneb organismi. Ag esitletakse antigeeni esitlevate rakkude poolt
Th0- le, mis diferentseeruvad allergilise reaktsiooni korral
Th2-ks , need omakorda
vabastavad erinevaid tsütokiine (IL-4 ja IL-13), mis
inhibeerivad Th1 rakkude
aktivatsiooni ja intensiivistavad IgE produktsiooni. Käsklus B- lümfotsüütidele. IgE
antikehad kinnituvad
nuumrakkudel/basofiilidel/lagerhansi rakkudel olevale FcRI-
le (veel kliinilisi sümptomeid pole). Et kliinilised sümptomid tekiks, on vaja vastava Ag
teistkordne sisenemine organismi kontakteerumine kahe IgE- ga aktiveeritakse
nuumrakk ja mediaatorid vabanevad.
Mediaatorid: histamiin - bronhide ahenemine, veresoonte dilatatsioon, lima eritus,
veresoonte permeaabluse suurenemine (tänu plasmaproteiinide sattumisel
interstitsiaalvedelikku tekib ka paistetus).
Trüptaas - aktiveerib retseptorid endoteelirakul, mis valikuliselt “meelitavad” eosinofiile
ja basofiile, C3 aktivatsioon
Nuumraku aktiveerumisel vabanevad ka
leukotrieenid (LTD4, LTC4 jt) (silelihaskoe
kontraktsioon, veresoonte permeaabluse tõus)
PGD2, PGE2, PGI2 – bronhilihaste kontraktsioon ja lõtvumine, vasodilatatsioon,
trombotsüütide agregatsioon
PAF(trombotsüüte aktiveeriv faktor)- trombotsüütide agregatsioon , vasoamiinide
produktsioon
Tsütokiinide tähtsus: nuumrakud ja basofiilid toodavad allergia hilises faasis, mis
algab 6-24h peale Ag –ga kokkupuudet TNF, IL-1, IL-4 ja IL-5 ning kemokiine nagu MIP-1
ja MIP-1 . Need “meelitavad” omakorda CD4+, monotsüüte ja eosinofiile, mis omakorda
vabastavad mitmeid Th2- tüüpi tsütokiine, eriti IL-4 ja IL-5, mis võimendavad põletikku
allergeenile.
Allergiahaigused (allergilised reaktsioonid): bronhiaalastma (kopsus olevatest
nuumrakkudest vabanevad mediaatorid, mis põhjustavad bronhilihaste kontraktsiooni ja
lima produktsiooni. Tekivad hingamisraskused). Allergiline rinokonjunktiviit
(heinapalavik), ekseem(atoopiline dermatiit), urtikaaria, toiduallergia, anafülaksia.
86.
Antikehade poolt vahendatud (II tüüpi) koekahjustuse mehhanism.
Koekahjustusmehhanism:
Fagotsütoosi kaudu
o Komplement aktiveeritakse opsoniseeritud rakul C3b: fagotsüüdil C3bR +
antigeen-antikeha seondumine opsoniseeritud rakk fagotsüteeritakse
Komplementsüsteemi aktivatsiooni kaudu
o Komplemendi kõrvalproduktid – C5a, C3a kaudu seonduvad Fc
retseptoritega fagotsüüdid ja viib põletiku ja koekahjustuseni
Anikehade poolt vahendatud NK rakkude tsütotoksilisuse kaudu (ADCC – antibody
dependent cellular cytotoxicity)
o Abnormaalne füsioloogiline vastus:
o TSH (thyroid stimulating hormone) + selle retseptori vastane antikeha
seonduvad kilpnäärme epiteelirakule ak stimuleerib ilma hormoonita
retseptorit
o Kui Ak seondub nt ACh (atsetüülkoliiin närvirakus) retseptorile lihasrakus,
inhibeerib ta neurotransmitteri seondumist) pidurdab raku funktsiooni.
o Nt autoantikehad ioonkanalite molekulide vastu
87.
Immuunkomplekside poolt vahendatud (III tüüpi) koekahjustuse
tekkemehhanism. Immuunkomplekside tekkemehhanismid ja iseloom
sõltuvalt antigeeni ja antikeha hulgast. Meditsiinilisi näiteid
immuunkompleksidest vahendatud reaktsioonidest. Immuunkompleksidest
tekkinud neerukahjustuse mehhanism.
Immuunkompleksid on antigeenide ja antikehade kompleksid. Tavaliselt eemaldavad
makrofaagid põrnas ja Kupfferi rakud maksas immuunkompleksid vereringest kiiresti.
Mõningatel juhtudel jäävad aga nad veres ringlema. Lõpuks jäävad nad lõksu neerudes,
kopsudes, nahas, liigestes või veresoontes. Koht, kus nad lõksu jäävad sõltub antigeeni
loomusest. Seal kohtades põhjustavad nad reaktsioone, mille tulemused tekivad
põletikud ja koekahjustused.
Immuunkomplekside poolt vahendatud ülitundlikkusreaktsiooni kutsuvad esile
bakteriaalsed, viiruste ja võõrseerumiste
antigeenid. Nende abil tekivad
immuunkopleksid, mis fikseeruvad (ladestuvad) kudedesse ( lokaalne või süsteemne
kahjustus)
Etapid:
Ag-Ak kompleksi teke
Ak Fc fragmendi seostumine komplemendi C1-ga
Komplemendi aktivatsioon
C5a (anafülotoksiin) veresoonte läbilaskvuse tõus, neutrofiilide aktivatsioon
Komplekside fagotsütoos (hapnikuradikaalide ja lüsosomaalsete ensüümide
vabanemine)
Koekahjustus (fibrinoosne nekroos)
Ülitundlikkusreaktsioonid tekivad lahustuvate antigeenidega. Antigeen-antikeha
kompleksid ehk immuunkompleksid ladestuvad kudedesse ning see põhjustab
patoloogia. Immuunkompleksid tekivad iga immuunvastuse korral, kuid patoloogia teke
sõltub nende suurusest, hulgast, afiinsusest ning reageeriva antikeha isotüübist.
Suuremad agregaadid fikseerivad komplemendi ning eemaldatakse kiirelt vereringest
mononukleaarse fagotsüüdisüsteemi poolt. Väiksemad kompleksid aga ladestuvad
veresoonte seintes. Seal nad seostuvad Fc retseptoritega leukotsüütidel ning
põhjustavad leukotsüütide aktivatsiooni ja koekahjustuse.
III tüüpi koekahjustusmehhanism võib vallanduda nahas, kui on olemas IgG antikehad
sensibiliseeritud antigeeni vastu. Kui antigeeni süstida nahka, siis veres ringlev ning
kudedesse difundeerunund IgG antikeha moodustab lokaalselt immuunkompleksi.
Immuunkompleks seostub Fc retseptoriga leukotsüütidel, mis põhjustavad lokaalse
põletikureaktsiooni. Sellega kaasneb suurenenud veresoonte permeaablus. Suurenenud
vaskulaarne permeaablus võimaldab vedelikel ja rakkudel, eriti polümorfonukleaarsetel
leukotsüütidel, siseneda veresoonde. Seda reaktsiooni nimetatakse Arthuse
reaktsiooniks. Immuunkompleksid aktiveerivad ka komplemendi, vabastades C5a, mis
aitab kaasa põletikulistele reaktsioonidele ligeerudes C5a retseptoritega leukotsüütidel.
III tüüpi koekahjustusmehhanismi tuntakse ka seerumtõve all ning see resulteerub kui
süstida suurtes kogustes halvasti kataboliseeritud võõrast antigeeni. Seerumtõbi ilmneb
7 – 10 päeva jooksul pärast hobuse seerumi süstimist, ajavahemik, mis vastab ajale, mis
on vajalik paigaldamaks primaarset immuunvastust. See lülitub IgM antikehalt IgG-le
hobuse seerumi võõraste antigeenide vastu. Seerumtõve kliinilised nähud on
külmavärinad, palavik, lööve, artriit ning mõnikord ka glomerulonefriit. Haiguse
puhkemine langeb kokku antikehade kujunemisega võõrseerumis olevate lahustuvate
proteiinide vastu; need antikehad moodustavad immuunkomplekse üle kogu keha. Need
immuunkompleksid fikseerivad komplemendi ning võivad seostuda ja aktiveerida Fc
retseptoreid kandvaid leukotsüüte ja komplemendi retseptoreid; need põhjustavad
ulatuslikke koekahjustusi.
Arthuse reaktsioon
Arthuse reaktsioon esineb inimestel väga harva. Hemorraagiline nekroos ravimi
süstekohal, putuka hammustus või nõelamine võib soodustada Arthuse reaktsiooni.
Arthuse reaktsiooni limiteeritud vorm võib esineda allergia desensitiseeritud injektsiooni
kohal, kui süstitud allergeenid on hakanud genereerima IgG blokeerivaid antikehi. Kuna
IgG tase on väga madal, siis kutaanne ning subkutaanne koekahjustus põhjustab kerge
nahapunetuse või kõvastumise.
Immunoloogiline patogenees sõltub antigeeni ja antikeha kontsentratsioonist, mis on
vajalik moodustamaks immuunkompleksi, mis oleks võimeline aktiveerima komplementi.
Vahepealse suurusega kompleksid aktiveerivad komplemendi ning seetõttu on nad ka
kudedele kõige ohtlikumad. Suured lahustuvad kompleksid eemaldatakse
mononukleaarse fagotsüütide süsteemi poolt ning väiksed kompleksid ei suuda
aktiveerida komplemendi retseptoreid. Immuunkompleksid aktiveerivad komplemendi
antikeha Fc seostumisel komplemendi C1q-ga. C3a ja C5a anafülotoksiinid on
vabastatud. Need molekulid aktiveerivad nuumrakud. Aktiveeritakse ka neutrofiilid, mis
on Arthuse reaktsiooni puhul väga tähtsad, sest nad vabastavad toksilisi kemikaale ning
toimub immuunkompleksi fagotsütoos.
Glomerulonefriit
Äge glomerulonefriit on päsmakeste immuunpatoloogilise põletikuga kulgev
neeruhaigus. Selle riskigruppi kuuluvad peamiselt nooremas täiskasvanueas inimesed,
mehed haigestuvad sagedamini kui naised.
Etioloogia - ägeda glomerulonefriidi tekkepõhjused on sageli teadmata. Suuremal osal
juhtudest kutsub haiguse esile angiin või tonsilliit. Glomerulonefriidile iseloomulikud
sümptomid ilmnevd üldjuhul 1 – 2 nädala pärast. Lisaks sellele kuuluvad etioloogiliste
faktorite hulka ka B-hepatiit, mumps, malaria jne.
Patogenees - ägeda glomerulonefriidi põhjuseks on immuunpatoloogiline puudulikkus.
Haigust iseloomustavad T-lümfotsüütide defitsiit ning organismi võimetus antigeeni
elimineerimiseks. Glomerulonefriidil võib immuunpatoloogilise mehhanismi alusel
eristada kahte vormi:
Antibasaalmembraani – glomerulonefriidi korral moodustuvad basaalmembraani-
vastased antikehad. IgM ja IgG tungivad läbi päsmakeste kapillaari endoteeli ja
reageerivad basaalmembraani retseptoritega. Selle tagajärjel kinnituvad leukotsüüdid
integriinide ja spetsiaalsete valkude (ICAM-1 ja VCAM-2) vahendusel endoteelile,
võimendades HVR ja proteolüütilise ensüümide produktsiooni, mille tulemusena
kahjustuvad neerupäsmakesed. Päsmakeste kahjustusele aitavad kaasa ka
trombotsüüdid, mis nende kapillaarides agregeerudes kutsuvad esile mikrotrombide
teke. Lisaks soodustab trombotsüütidest vabanev fosfolipiid PAF veresoonte läbilaskvust
ja avaldab tsütotoksilist toimet. Tsütokiinide tootmine aktiveerib omakorda leukotsüüte
ning kahjustus süveneb.
Immuunkompleksidest põhjustatud neerupõletik kujuneb välja juhul, kui antigeen-
antikeha komplekside hulk ületab retikuloendoteliaalsüsteemi rakkude
fagotsütoosivõime. Seda vormi iseloomustavad immuunkomplekside ladestused
basaalmembraani välisküljel. Neutrofiilsed leukotsüüdid kutsuvad esile kapillaari
permeaabluse suurenemine ja proliferatiivsed protsessid neerukoes.
Morfoloogiline pilt ja sümptomatoloogia - Morfoloogiliste muutuste põhjal võib ägedal
glomerulonefriidil eristada kuut alavormi, millest alljärgnevalt tutvustatakse lühidalt
nelja:
Membranoosne glomerulonefriit – glomerulaarkapillaari sein on tugevalt
paksenenud, kapillari seina epiteliaalsel küljel granulaarsed IgG ja C3 ladestused ning
kliinilistest sümptomitest esinevad nefrootiline sündroom ning proteinuuria.
Membranoproliferatiivne glomerulonefriit – päsmakestes on näha proliferatiivsed
mesangiumirakud, seina paksenemine ja IgG, IgM ning C3 ladestused.
Endokapillaarne proliferatiivne glomerulonefriit – iseloomustavad proliferatsioon,
polünukleaarsed leukotsüüdid, IgG ja C3 ladestused.
Mesangiaalne IgA ladestusega glomerulonefriit – esinevad mesangiaalsed IgA
ladestused, proliferatsioon ja hüalinoos. Lisaks ilmneb sageli mikrohematuuria.
Kliinilistest sümptomitest figureerivad ägeda glomerulonefriidi korral:
tursed
glomerulaarse filtratsiooni vähenemine
naatriumi reabsorptsiooni suurenemine
arteriaalse vererõhu kõrgenemine
vasaku vatsakese puudulikkus
oliguuria
hematuuria
proteinuuria
88.
T-rakkude poolt vahendatud (IV tüüpi) koekahjustuse mehhanism.
Th1 ja Th17 – kaudsed reaktsioonid.
o Th1 aktiveerib makrofaage
o Th17 aktiveerib neutrofiile – IL-17; GM-CSF
Tsütotoksiliste CD8+ T rakkude (CTL) poolt vahendatud reaktsioonid
o Infektsioonid, kemikaalid/looduslikud mürgid, metalliioonid, ravimid,
autoimmunisatsioon.
Mükobakteri antigeen. Primaarne infektsioon, 1-2 ndl pärast süstitakse mikroobset
antigeeni, tekib punane ring.
Aeglast tüüpi ülitundlikusreaktsiooni puhul infiltreeruvad lümfotsüüdid ja makrofaagid
perivaskulaarselt. Infiltreerunud rakud on CD4+ positiivsed.
Näited:
- tuberkuloos
- Candida infektsioon
- kontaktdermatiit
- reumatoidartriit
- polüskleroos (sclerosis multiplex)
- 1. tüüpi diabeet
- psoriaas
89.
Autoimmuunsuse tekkemehhanismid, antigeenide ja
immuunregulatsiooni tähendus. Välisfaktorite ja pärilikkuse roll
autoimmuunsuse tekkes. Oluliseimad autoimmuunhaiguste tekkega
assotsieeruvad HLA alleelid. Infektsioonid ja autoimmuunsus.
Autoantigeen on organismi endogeenne antigeen, mis stimuleerib autoantikehade
produktsiooni
autoimmuunreaktsioonis.
Autoantigeen
võib
põhjustada
autoimmuunhaiguseid.
Autoantigeenid jaotatakse:
Organspetsiifilised autoantigeenid:
Rakumembraaniretseptorid
o Atsetüülkoliinretseptor
o
hormoonide retseptorid
(TSH,INS)
Hormoonid
o T3, T4
Ensüümid
o kilpnäärme peroksüdaas
o mao HK-ATPaas
o tsütokroom P450
o glutaamhappe dekarboksülaas
Autoantigeenide organspetsiifilisus
määrab sageli ära haiguse
Organspetsiifikata autoantigeenid:
Rakutuuma komponendid
o DNA
o RNP
o histoonid, tsükliin, Sp-100
Tsütoskeleti komponendid
o aktiin, tubuliin, keratin
Ensüümid
o 2-oksohapete dehüdrogenaasid
o aminoatsüül tRNA süntetaasid
o müeloperoksidaas
jm (IgG Fc, adhesioonimolekulid,
kollageenid, HSP valgud)
organspetsiifilisuse.
Autoimmuunsuse tekkes osalevatel välisfaktoritel on kas:
otsene mõju (ristreaktsioonid, immuunsüsteemi aktiveerumine jm): mikroorganismid,
toiduained
kaudne mõju:UV kiirgus, radiatsioon, kemikaalid jms
Välisfaktorid on olulised, kuna samast kohast pärit inimestel on suur erinevus haiguse
avaldumisel, kuigi geneetika on sama. Põhja-lõuna gradient on esinev, põhja pool
rohkem esinevad.
Infektsioonid:
Autoimmuunhaiguste (AIH) loommudelid on näidanud, et infektsioonid võivad AIH esile
kutsuda. Adjuvantidega südamevalkude manustamisel hiirtele võib indutseerida
müokardiiti, st AIH välja kujunemiseks ei ole tingimata vajalik aktiivne infektsioon.
Samuti on näidatud, et erinevad mikroobid võivad põhjustada ühte ja sama AIH sarnaste
mehhanismide abil.
AIH ja infektsiooni vahelist seost kutsutakse sageli
molekulaarseks mimikriks. St mikroorganismi antigeenid sarnanevad
autoantigeenidega. Seega toimub ristreaktsioon immuunvastuse tulemusena
genereeritud Ak-de või T rakkude ning autoantigeenide vahel ja AIH esile kutsumise.
Teine võimalik seos AIH ja infektsiooni vahel: mikroorganismid kahjustavad otseselt
kudesid ja sel viisil eksponeerivad autoantigeene immuunsüsteemile (“bystander
effect”).
Mudel
AIH tekke üldmudel: mingil määral pärilik eelsoodumus + päästikud
väliskeskkonnas
autosensibiliseerunud T rakud, autoantikehad
nihked
biokeemias
sümptomid (koekajustus)
Tsentraalse ja/või perifeerse tolerantsuse häirumine. Tregide teke T rakkudel
tüümuses või Retseptori parandamine B rakkudel.
Lisasignaali (aktivatsiooniks) puudumise tõttu ei aktiveeru T B rakul reaktiivsuseks
vajalikud rakupinna retseptorid, adhesioonimolekulid jne ⇒ anergia/deletsioon
(aktiveeritud rakkude apoptoos)
Alla suruvad signaalid –
o CTLA-4, PD-1, TIGIT aktivatsioon T rakkude pinnal CD4+CD25+FOXP3+ Treg
rakke
o CD22 kaudu B rakkude aktivatsioon; aktiveerivad IL-10 produtseerivaid
regulatoorseid B rakke e B10 rakke
⇒ SUPRESSIOON --- IL-10, TGF-Β, IL-35
T rege on kahte tüüpi:
nTreg – loomulikud, tunnevad enda autoantigeeni ära
tüümuses (IL-2
FOXP3)
iTreg – indutseeritud – tunnevad self antigeeni ära perifeersetes kudedes, LN nt;
ka IL-2 mõjul Tregiks
⇒
mõlemad inibeerivad T-raku aktivatsiooni VÕI aktiveeritud efetor T raku
efektorfunktsioone
HLA alleelid:
HLA-DR3-DQ2 haplotüüp, HLA-DRB1. Erinevatel AIH erinevad alleelid, mis teket
soodustavad.
HLA-B27, C06, HLA-DR1, DR3, DR4, DR8, DR15
Märklaudkoe kahjustus:
autosensibilseeritud T rakkude vahendatud mehhanismid
o CD4+ - põletikutsütokiinid
o CD8+ - tsütotoksiline toime
Autoantikehade poolt vahendatud
o Otsene mõju märklaud antigeeni molekulile
Konformatsiooni muutus
Funktsionaalselt oluliste osade
blokeerimine/inhibeerimine/stimuleerimine
o Immuunkomplekside moodustamine komplemendi aktivatsioon
o NK rakkude poolt vahendatud ADCC reaktsioonid
Loomulik immuunsus: NK rakkude otsene mõju/IFN α, IL-1 mõju
90.
Organ-spetsiifiliste autoimmuunhaiguste iseloomustus. Nende
immunopatogeneesi mehhanismid ja immunodiagnostika.
Immunogeneetilised aspektid.
Autoimmuunhaigused moodustavad nn. spektrumi, mille ühes äärmuses on
organspetsiifilised ja teises organspetsiifikata haigused. Esineb ka haigusi, mis haaravad
primaarselt ühte organit, kuigi immuunreaktsioonid on ilma organspetsiifikata
(primaarne biliaarne tsirroos). Organ-spetsiifilised AIH-d hõlmavad spetsiifilisi organeid,
kus leidub autoantigeene. Endokriinsüsteemi haigused
Autoimmuunhaiguste diagnostikas põhinetakse sümptomitele ja antikehade leiule
mis reageerivad haaratud kudede ja rakkudega. Antikehad raku/koe vastu, mis
reageerivad antigeenidega, määratakse immunofluorestsentsiga. AK-si lahustuvate AG-
de vastu leitakse ELISAga või radioimmunoloogilistel meetoditel, vahel ka biokeemiliselt
või bioloogiliselt.
IIF; ELISA; FEIA; Immunoblot
Organspetsiifilised autoantigeenid:
rakumembraani retseptorid (atsetüülkoliinretseptor)
hormoonid (T3, T4)
ensüümid (kilpnäärme perokidaas, tsütokroom P4, glutaamhappe dekarboksülaas,
maohappe APTaas)
Tänu AAKdele saame diagnoosida haigust, ennustada kliinilise haiguse teket,
prognoosida haiguse kulgu, hinnata haiguse aktiivsust ja ravi efektiivsust.
Kasutatakse:
kaudne immunofluorestsents
ensüümkaudne immunosorbent-test (ELISA)
radioimmunopretsipitatsioonitest (RIP)
Western blot e. immunoblotanalüüs
automaatanalüsaator
Raviks on praegusel ajal põhiliselt kortikosteroidid ja immunosupressiivne (tsüklosporiin)
ravimteraapia. Senised ravimeetodid ei ole suunatud tekkepõhjuste vastu vaid on
immunomodulaatorid, retseptorite, tsütokiinide jm. toimet pidurdavad ravimid.
Olulised uute spetsiifiliste ravivõimaluste otsingud:
autoantigeenide või nende osade vastase immunoloogilise tolerantsuse
esilekutsumine:
limaskestade kaudu manustatud autoantigeenid (suukaudsed,
ninnapihustatavad preparaadid üksi või koos immunomodulaatoritega)
autoantigeenide cDNA manustamine (koos immunomodulatsiooni esile
kutsuvate mehhanismide aktivatsiooniga)
tolerantsi kujundamine autoreaktiivsete regulatoorsete T rakkude abil jne.
Autoimmuune türeoidiit (Hashimoto türeoidiit) e Hashimoto türeoidiit –
kilpnäärme lümfotsütaarne infiltratsioon (B, CD4+)
immuunreaktsioonid kilpnäärme perosidaasi vastu
immuunreaktsioonid türeoglobuliini vastu
Diagnostikas olulised anti-TPO, anti-TG, anti-TSHR
-
näärmekoe destruktsioon
-
lõpptulemusena hüpotüreoos
-
N:M 6:1
-
autoantigeenid : türeoglobuliin, kilpnäärme peroksüdaas, TSH retseptor
Mõiste autoimmuunne türeoidiit hõlmab enda alla Hashimoto türeoidiidi, Graves´i tõve ja
teisi kilpnäärme haigusi. Krooniline autoimmuune türeoidiit (Hashimoto türeoidiit 85% on
naised) on autoimmuune haigus, mis on tingitud sellest, et immuunsüsteem käsitleb
kilpnääret kui kehavõõrast kude ning ründab teda
autoantikehade kaudu. Haiguse
tekkes on oluline osa geneetilisel eelsoodumusel. Võib avalduda igas vanuses, kuid
sagedamini esineb keskealiste naiste hulgas. Kahjustavad kilpnäärmevastased antikehad
kilpnäärme rakke nii, et hormooni produktsioon väheneb.
Hashimoto türeoidiiti esineb ligikaudu 5% populatsioonist. Graves’i tõve puhul
produtseerib immuunsüsteem kilpnäärme vastaseid antikehi ja selle tulemusena hakkab
kilpnääre produtseerima liiga palju kilpnäärme hormooni. Naistel esineb
autoimmuunhaigusi 5 korda rohkem kui meestel.
Sümptomid: Ägeda türeoidiidi puhul esineb tugev valu kaela eesosas, mis kiirgub
kõrvadesse. Palavik võib tõusta 38-39ºC, esineda võib ka neelamise valulikkus ning naha
punetus kaelal kilpnäärme kohal. Enamasti häireid kilpnäärme talitluses (hormoonide
tootmises) ei ole.
Alaägedale türeoidiile on iseloomulik järsk algus kõrge palaviku, kõrvadesse kiirguva valu
ning valuliku sõlmega kilpnäärmes. Esinevad ka üldised viirushaiguse sümptoomid -
nõrkus, lihasvalud. Umbes pooltel juhtudel võivad haiguse esimesel nädalal esineda
kilpnäärme ületalitluse nähud, mis on tingitud hormoonide vabanemisest viirusest
kahjustatud kilpnäärmest. Haigus kestab 1-6 kuud ning lõpeb kilpnäärme normaalse
talitluse taastumisega.
Krooniline autoimmuune türeoidiit - olenevalt kilpnäärme suurusest eristatakse kahte
haigusvormi: esimesel juhul kilpnääret kaelal katsudes tunda ei ole, autoimmuunse
protsessi tagajärjel on kilpnääre muutunud väiksemaks; teisel juhul on tegemist
kilpnäärme kompenseeritud suurenemisega. Haiguse kulg võib olla stabiilne,
progresseeruv (viib kilpnäärme alatalitluse tekkele) või ägenemistega.
Lisaks on olemas veel sünnitusjärgne türeoidiit.
Loomkatsed:
spontaanselt tekkinud N: NOD (mittepaks diabeetiline hiir), BB rotid jt
neonataalse tümektoomia teel indutseeritud
antigeenide või nende osistega immuniseerimisel saadud
antigeenide cDNAga immuniseerimisel saadud
transgeensed ja Knock-out mudelid
Immunoloogilised eksperimendid:
BB rotid – spontaanne autoimmuunne diabeet ja autoimmuunne türeoidiit
Buffalo (pühvel) rotid – autoimmuunne türeoidiit ja pankrease β -rakkude hävimine
Obese (turske) kana – hashimoto türeoidiit.
T1d – I tüüpi diabeet
Pankrease β-rakkude kahjustumine ja kolm haiguse staadiumi.
1. toodetakse CTL T rakke suureneb β-rakkude hulk väheneb aastatega. kliinilised
nähud.
Geenid, mis soodustavad: HLA/Dq/DR haplogrupp, INS, CTLA4, PTPN22, CD226 jm
mutatsioonid. Olulised on ka välisfaktorid – viirused, stress jm. Toidukompoenndid,
toksiinid, mikroorganismid (bakterid, viirused) JM.
GADA + IA-2A Autoantikehad on prognostilised!
Tsöliaakia
Tekitajad – nisu, rukis, oder, kaer. Geenid: HLA-DR3, DQ2, DR4, DQ8. Peensoole
limaskesta kahjustus. Raviks on ainult range gluteenivaba dieet.
Mehhanism: glyteeni peptiid seondub HLA molekulile CTL-id aktiveeritakse, hakatakse
tootma gluteeni vastaseid autoantikehasid. Ifn γ IL-21 mõjul vastus suureneb.
91.
Organ-spetsiifilikata autoimmuunhaiguste iseloomustus. Nende
immunopatogeneesi mehhanismid ja immunodiagnostika.
Organspetsiifikata haiguste puhul on autoantigeenid jaotunud üle kogu keha laiali. Ja
mõjutataud on mitmed organid. Erinevate haiguste korral võivad koekahjustuse
immuunmehhanismid olla erinevad
Organspetsiifikata e generaliseerunud autoimmuunhaigused e süsteemsed
sidekoehaigused: nt süsteemne erütematoosne luupus (SLE), reumatoidartriit,
polümüosiit ja dermatomüosiit, süsteemne skleroos, Sjörgeni sündroom.
Neile on iseloomulik patoloogilise protsessi süsteemsus, mis on tingitud
sellest, et autoantikehad reageerivad mitmesuguse lokalisatsiooniga
rakkudega.
Organspetsiifikata autoimmuunhaiguste diagnoosimiseks kasutatakse:
Kui on tegemist süsteemsete nähtudega, on alati vajalikud
verepilt, uriinianalüüs,
neeru ja maksa funktsiooni näitajad. Selle kõrval on enamlevinud testiks (just
reumatoloogias)
ANA (anti nuclear antibodies) määramine. ANA testi sensitiivsus
ning spetsiifilisus on kõige kõrgem just SLE korral, oluliselt madalam on see aga teiste
süsteemsete sidekoehaiguste korral ja samuti vanemaealistel patsientidel.
Spetsiifiliste autoantikehade uuringud (nt anti-dsDNA, anti-Ro, anti- La jne) ei ole
rutiinuuringud. Koesobivusantigeenidest on enam levinud HLA-B27 määramine, mis
on oluline marker spondüloartropaaatiate korral.
otsest immunofluorestsentsmeetodit, milles uuritavaks materjaliks on koelõik.
Määratakse antigeeni olemasolu ja lokalisatsiooni koelõigus, nt glomerulonefriit –
neerubiopsia külmutuslõigul määratakse neerupäsmakeses immuunkomplekside
olemasolu, lokalisatsioon ja koostis (Ak isotüüp);
kaudset immunoflorestsentsmeetodit: määratakse järgnevaid autoantikehi – ANA
e tuumavastased antikehad; SMA e anti-smooth muscle cell antibodies; nii ANA kui ka
SMA võib leida ka tervetel inimestel, seetõttu nende leid ei viita veel väga kindlale
haigusele. ELISA: antigeenidena kasutatakse nt ssDNA-d - SLE puhul on spetsiifilisus
>90%, reumatoidartriidi puhul 40%; kardiolipiini – SLE puhul spets. 40%
Skriining poolkvantitatiivse latekstestiga - määratakse reumatoidfaktorit (fS-RF) – see
on oma olemuselt heterogeenne autoantikehade grupp, mis tekib IgG-molekuli Fc-
fragmendi vastu. Need autoantikehad võivad kuuluda nii IgG, IgM, kui ka IgA klassi,
kuid peamine immuunglobuliinide klass, millest moodustub veres tsirkuleeriv
reumatoidfaktor, on IgM klass. Reumatoidfaktor toodetakse aktiveeritud B-
lümfotsüütide poolt, mis on infiltreerinud haige liigese. Moodustuvad
immuunkompleksid, mis aktiveerivad komplemendi ja põhjustavad koekahjustust.
Uurimiseks kasutatakse seerumit. Latekstest muutub positiivseks kontsentratsioonil
üle 20 IU/mL. Positiivse tulemuse korral teostatakse kvantitatiivne
immuunturbidimeetriline määramine. Normaalselt RF seerumis ei leidu. Kasutatakse
artriitide ja autoimmuunhaiguste diferentsiaaldiagnostikas (koos tuumavastaste
antikehade määramisega). Positiivse testitulemuse sagedaseim põhjus on
reumatoidartriit, mille puhul on test positiivne 75-80% haigetel - seropositiivne artriit.
Haiguse alguses on reumatoidartriit umbes kolm kuud seronegatiivne. Juveniilse
reumatoidartriidi korral leidub RF seerumis vaid ~20%-l juhtudest. RF võib esineda
haigete veres ka teiste reumaatiliste haiguste ning samuti mõningate
mittereumaatiliste haiguste puhul. RF leidub seerumis ~35% süsteemse
erütematoosse luupusega patsientidest, ~95%-l Sjögreni sündroomiga patsientidest,
võib esineda sklerodermia, dermatomüosiidi, maksatsirroosi, sarkoidoosi ja
Waldenströmi makroglobulineemia puhul. On leitud haigete veres bakteriaalsete
infektsioonide ja parasitaarhaiguste, samuti mõningate viirusinfektsioonide
(infektsioosne mononukleoos, hepatiit) korral ja vaktsinatsioonide järgselt.
Infektsioonide puhul kaob RF seerumist pärast infektsiooni likvideerumist. Vähene RF
leid võib esineda ka ~4% haigussümptomiteta isikutel, eriti üle 60-aastastel
Koagulomeetriline määramine -määratakse luupus-antikoagulante (P-LA) - Luupus-
antikoagulandid (lupus(-type) anticoagulants, LA või ka LTA) on fosfolipiididevastased,
tavaliselt polüklonaalsed IgG ja/või IgM tüüpi antikehad, mis in vitro inhibeerivad
kämbu teket (pikendavad tekkimise aega) fosfolipiid-sõltuvates
koagulatsioonitestides (nt APTT). Luupus-antikoagulandid võivad plasmas esineda ka
isikutel, kes ei põe SLE-d. LA leiuga (eriti SLE) haigetel on kõrgenenud risk
trombooside tekkeks. Määramisel on sisuliselt tegemist APTT-ga, kuid
reaktsioonisegule on lisatud hepariini inhibiitorit (muudab reaktsiooni keskkonna
tundetuks hepariinile kuni 1 IU/mL) ja normaalset inimplasmat (korrigeerib kämbu
tekkimise aja pikenemise, mis võiks esineda hüübimisfaktorite defitsiidi korral
uuritavas plasmas). Määratakse APTT kahes katsutis, millest ühte on lisatud ka
heksagonaalse faasi lipiidi (fosfatidüületanoolamiini, HPE), mis seob spetsiifiliselt LA ja
neutraliseerib selle efekti ning võrreldakse hüübimisaegade erinevust neis kahes
katsutis. Referentsväärtused: Normaalleid on negatiivne. Luupus-antikoagulandid
esinevad sagedamini SLE ja ka teiste autoimmuunhaiguste, infektsioonide (nt HIV) ja
pahaloomuliste kasvajate puhul, samuti mõnede ravimite (nt fenotiasiinid) tarvitamise
korral.
Röntgenoloogiline diagnostika – nt reumatoidartriidi puhul
Spektrofotomeetriline määramine – määratakse adenosiini desaminaasi (S-ADA, jt)
Adenosiini desaminaas on kõigis kudedes esinev ensüüm, mis osaleb puriinide
ainevahetuses. Eriti palju ADA on lümfaatilises koes, kus tal on keskne roll rakkude
diferentseerumisel. Rakulise immuunsuse aktiveerumise puhul on ADA aktiivsus
kõrgem, ADA vähene aktiivsus viitab rakulise immuunsuse puudulikkusele. Uuritavaks
materjaliks kasutatakse Hemolüüsivaba seerum/plasma, pleuravedelik, liikvor. Kõrge
S-ADA (normväärtus S – ADA < 21U/L) viitab rakulise immuunsuse aktiveerumisele
(mõned infektsioonhaigused, autoimmuunhaigused, pahaloomulised kasvajad)
Turbidimeetriline meetod – määratakse immuunglobuliine A, G ja M. Uuritakse
seerumit või plasmat. IgG tõus: autoimmuunhaigused (krooniline hepatiit,
maksatsirroos. Väga väljendunud kontsentratsiooni tõusu võivad põhjustada
mõningad parasitaarsed haigused ja kroonilised ning retsidiveeruvad infektsioonid.)
Süsteemsete sidekoehaiguste ravis
kasutatakse nt. mittesteroidseid
põletikuvastaseid aineid (MSPVA-d). Nende üldine toimemehhanism – kapillaaride
permeaabluse vähendamine, põletikumediaatorite inaktiveerimine ja sünteesi piiramine,
lüsosoomide membraani stabiliseerimine, mis piirab põletikku soodustavate ainete
(tsütokiinide) rakust väljumist, põletiku proliferatiivse faasi pidurdamine,
prostaglandiinide sünteesi inhibeerimine. Veel kasutatakse haigust modifitseerivaid
reumaravimeid e II rea preparaate, mis põhiliselt on immunoaktiivsed ained. Nende
eesmärk on immunosupressiivne toime autoimmuunsele protsessile. Ravikuurid haigust
modifitseerivate ravimitega kestavad poolest aastast mitme aastani.
Nüüdisajal pööratakse suurt tähelepanu kombineeritud tsütostaatilisele ravile. Sel puhul
kasutatakse üheaegselt 2-5 haigust modifitseerivat ravimit. Immuunteraapias on leidnud
kasutamist monoklonaalsed antikehad, nt anti-CD4, anti-CD5, anti-CD7, anti-CD52, anti-
TNFα ja interferooni preparaadid. Sellisel ravil on aga olnud palju kõrvalnähte.
Perspektiivseks on osutunud luuüdi ja vere tüvirakkude autoloogne transplantatsion.
Näidustusteks oleksid SLE, raskete tüsistustega reumatiodartriit, dermatomüosiit jt. R
Süsteemne erütematoosne luupus (SLE)
SLE kuulub mitteelundispetsiifiliste autoimmuunhaiguste hulka, mille puhul esineb lai
tsirkuleerivate antikehade spekter. Häiritud on immunoloogiline homöostaas, põhjuseks
defekt rakulise immuniteedi lülis (T-lümfotsüüdid) ja humoraalse immuniteedi
hüperaktiivsus (B-lümfotsüüdid).
Peamised immunoloogilised tunnused:
Esinevad – tuumavastased antikehad (ANA); anti-ds-DNA; anti-Sm antikehad
Komplemendi tase seerumis on langenud
Immunoglobuliinid ja komplment on ladestunud glomeeruli basaalmembraanile ja
dermaal-epidermaal joonele
Võib esineda palju teisi autoantikehasid
Tuumavastaste antikehade positiivne näit esineb veel ka tervetel isikutel (kuid
madalas tiitris), erinevate krooniliste haigustega isikutel (kasvajad) ning vanematel
inimestel. Kui tuumavastased antikehad on kõrges tiitris, seostuvad need SLE ning teiste
süsteemsete sidekoehaigustega ja ANA alatüübid on nende haiguste diagnostiliseks
markeriks. Süsteemse erütematoosse luupuse diagnoosi vastu on negatiivne ANA.
SLE patsientide seerumis esineb kolme tüüpi anti-DNA antikehasid:
anti-single stranded e denature-DNA (=anti-ssDNA)
anti-double stranded e native DNA (= anti-dsDNA)
reageerivad nii ss-DNA-ga kui ds-DNA-ga
dsDNA väga spetsiifiline SLE-le. dsDNA autoantikehade määramiseks kasutatakse testi
Crithidiae luciliae. Antikehad võivad olla nii IgG kui ka IgM klassist. Üheks oluliseks
tunnuseks SLE haigetel on anti-dsDNA antikehade esinemine kõrges tiitris. anti-dsDNA
antikehad esinevad 70% aktiivse luupusega haigetest. Positiivse testi spetsiifilisus on
95%. Anti-RNP – 30-50% patsientidel eeskätt “overlap” sündroomi markerina
Anti-Ro (30%) ja anti-La (15%) võivad esineda ka ANA-negatiivse SLE korral. Sagedamini
leitakse neid primaarse Sjörgeni sündroomi korral
Reumatoidfaktor – 30-60%-l tavaliselt madalamas tiitris kui RA (reumatoid artriidi) puhul
SLE etioloogia on ebaselge. On leitud HLA-DR2 ja HLA-DR3 suuremat esinemissagedust.
Vallandavateks teguriteks on peetud infektsioone, stressi, dieeti, keskkonna tegureid,
UV-kiirgust. Kliiniline pilt haiguse alguses võib olla suhteliselt ebamäärane ja
mittespetsiifiline. Klassikaline triaad: dermatiit, artriit, polüserosiit esineb 90% haigetest.
Haiguse korral võib tekkida:
Nahakahjustus – tekib päikese käest katmata kehaosadel ning intensiivistub
ultraviolettkiirguse toimel; sagedamini esineb liblikakujuline erüteem ninaselja ja
põskede piirkonnas
tugiaparaadikahjustus – artralgia ja artriit võib esineda 63-90% patsientidel
serosiidid – kuni 30% SLE patsientidel areneb haiguse käigus (sagedamini
perikardiit ja/või pleuriit)
neerukahjustus – aluseks immuunkompleksne põletik: glomerulonefriit;
nefrootiline sündroom kujuneb ~10% patsientidel
närvisüsteemikahjustus – esineb kuni 40 % haigestunutest, põhjuseks erinevad
patogeneetilised mehhanismid k.a antineuraalsed antikehad, tromboos, vaskuliit
varia – autoimmuunne hemolüütiline aneemia, mille korral esinevad autoantikehad
ja positiivne Coombsi test; tsütopeenia võimalik igas rakuliinis, sagedamini
leukopeenia; autoimmuunne trombotsütopeenia põhjustab harva veritsust ja
reageerib GK-ravile; hüperkoagulatsiooni seostatakse antikardiolipiinantikehade
olemasoluga
SLE-I medikamentoosses ravis on kasutusel 4 ravimite põhigruppi ja nende
kombinatsiooni: mittesteroidsed põletikuvastased ained, hydroxychloroquine,
kortikosteroidid ja immunosupressandid.
SLE on seotud mitme sõltumatu geeniga, mis mõjutavad immunoregulatsiooni,
immuunvastust, komplemendi taset, komplemendiga seotud faktoreid,
retikuloendoteliaalsüsteemi funktsiooni, immunoglobuliinide teket ja hormoonide
metabolismi. Haigus on geneetiliselt determineeritud immunoregulatoorsete
mehhanismide mittetäisväärtuslikkusega: esineb C2 ja C4 defitsiit, kindlad Gm allotüübid
ja madal CR1 tase. SLE-I haigetel esineb sagedamini kui populatsioonis antigeenne HLA-
A1, B8, DR2, DR3, DQ1 ja DQ2.
92.
Autoantikehade testide kliiniline tähendus diagnostikas, haiguse
prognoosis, ravi hindamisel jm. Tähtsamad kliinilises praksises kasutatavad
autoantikehad.
Autoantikehad (AAK) võivad põhjustada haigust, aga ka kaasa aidata selle patogeneesile.
AAK-d on pigem haiguse kui selle põhjuse tulemus. Mõnikord leitakse neid ka haiguse
puudumisel (eriti vanematel inimestel). Testides kasutatakse
immuunfluorestsentsi,
hinnatakse kvantitatiivselt immuunkomplekside moodustumise tagajärjel tekkivat
helendust või positiivset-negatiivset hindamist. Testidel on subjektiivne tõlgendus,
seetõttu peab omama kogemusi ja oskusi nende läbiviimiseks. Kui antigeenid (AG) on
iseloomustatud ja isoleeritud, asendatakse immunofluorestsents mõnikord teiste
meetoditega (Western blotting, aglutinatsioon jne).
Kliiniline tähendus: AAK-d on diagnostilised markerid, analüüsitava aine sidujad
kliinilise keemia uuringutes, immunoloogilised reaktandid uute biomolekulide
avastamiseks, lihtsustavad diagnoosimist (näiteks: mitokondriaalsed antikehad
primaarse biliaarse tsiroosi puhul; enne nende kasutuselevõttu oli haiguse
diagnoosimiseks vaja laparotoomiat, mis oli sageli kahjustav patsiendi ea või seisu tõttu).
Samuti on heaks diagnostiliseks markeriteks üht tüüpi AAK-de kogus seerumis.
Võimaldavad haiguse kulu jälgimist ja ravi mõjususe hindamist. Selleks kasutatakse
vähemspetsiifilisi teste (reaktsioonid kompleksse koeekstraktiga). Ennetav väärtus:
näiteks inimene, kellel on insuliini ja glutamiinhappe karboksülaasi vastased antikehad,
omab suuremat riski haigestuda insuliinsõltuvasse diabeeti.
AAK-de grupid: membraani liporoteiinivastased, silelihaste vastased,
mitokondrite vastased, mikrosoomide vastased, tuumavastased,
tsütoplasmavastased, DNA vastased, endoteelivastased,
türeoidnäärmevastased, granulotsüütide vastased, erütrotsüütide vastased
jne.
93.
Geneetiliste ja välisfaktorite osa allergia tekkes. Tähtsamate allergiaga
seotud geenide iseloomustus. Välisfaktorid, mis kutsuvad esile Th2 rakkude
aktivatsiooni. Hügieenihüpotees.
Allergia teke: APC presenteerib Th0-le antigeeni (allergeeni) IL-4 mõjul areneb Th2 (IL-
12 mõjul Th1), aktiveeritakse eosinofiilid, B rakud (IFN γ mõjul), Nuumrakud (B rakkude
mõjul), antikejad (IgE) seonduvad nuumrakkude FcR-le. Allergeen spetsiifilised Tregid
toodavad IL-10, TGF β, mis inhibeerivad nuum, baso, eosinofiile. Toodavad Brege, mis
toodab IgG mis seondub IgEle ja blokib ära selle.
Allergilised reaktsioonid on spetsiifiliste IgE antikehade tootmise tulemus tavalistele
(kahjututele) antigeenidele. Muutused IgE produktsioonis on mõjutatud nii geneetiliste
kui ka keskkonna faktorite poolt.
Keskkonna faktorid, mis on seotud allergia esinemissageduse suurenemisega:
Laste immuunsüsteem polariseeritud Th2 reaktsioonide suunas (lastel esineb vähe
infektsioone, mis stimuleerivad Th1 rakke). Nt lapsed, kellel on olnud bronhioliit koos
respiratoorse süntsütsiaalviirusega (RSV), on hiljem rohkem vastuvõtlikud astma
suhtes. (Selle haiguse tõttu muutub tsütokiinide tootmine: ei produtseerita enam nii
palju IFN-γ ja suureneb IL-4 tootmine, mis omakorda põhjustab Th2 immuunvastust.)
On võimalik, et infektsioonid, mis kutsuvad esile Th1 immuunvastuse, võivad
vähendada Th2 immuunvastuse tõenäosust hilisemas elus.
Ülekaalulisus
RSV viirusinfektsiooni ja võõrantigeenide (toiduallergeenid) koosmõju lastel (Th0
rakud polariseeruvad seetõttu Th2 rakkude tekkele, mis omakorda soodustab allergia
kujunemist)
Keskkonnafaktorid (diiselkütus, bensopüreenid); SAMAS: Janeway’st => keskkonna
saastatus ei kutsu esile ei atoopia ega astma esinemise suurenemist, pigem esineb
rohkem atoopiat puhtama õhuga piirkondades (kuigi ei välista saastatud õhu
kahulikku mõju kopsudele, kuid see ei olevat enamasti allergeense päritoluga)
Antibiootikumide kergekäeline kasutamine (mikrofloora muutus)
Uued toiduainete tootmistehnoloogiad
Tubakasuits, tolmud (taimed, kodutolm), putukamürgid, loomsed valgud, ravimid
(penitsilliin jt).
Muutusi organismi mikroobikoosluses peetakse kõige
usutavamaks allergia leviku suurenemise põhjuseks, sealhulgas muutused
antibiootikumide kasutamise tõttu ja lastel Th2 reaktsioonide esinemine Th1
stimuleerivate infektsioonide puudumise tõttu. Allergia teket mikroobikoosluse
muutuste tõttu toetab ka allergia tekke hügieeni hüpotees: liigne hügieen põhjustab
allergiate leviku suurenemist (hävitatakse organismi normaalne mikroobikooslus, mis
stimuleerib immuunsüsteemi).
Geneetilised faktorid:
Allergia tekkes on oluline osa pärilikkusel. Allergiliste vanemate laps on allergiale
ohustatud 4x rohkem kui ilma selle foonita. (Lastel, kellel on üks vanem allergik, on 30%
ja lastel, kelle mõlemad vanemad on allergikud, on 50% suurem tõenäosus allergia
kujunemiseks, võrreldes lastega, kelle vanemad pole allergikud.). Allergiliste haiguste (nt
astma) kujunemisega on seotud mitmed geenid, esineb keskkonna-geenide teatud
koosmõju- mõned geenid soodustavad allergia kujunemist ainult konkreetsete keskkonna
tingimuste ja elustiili juures.
Geneetiline vastuvõtlikkus erinevatele allergilistele haigustele on määratud erinevates
populatsioonides erinevate geenide poolt. Tõenäoliselt mõjutavad ühe haiguse erinevaid
külgi ka erinevad geenid. Nt. astma puhul on erinevate geenide poolt määratud IgE
tootmine, põletikuline vastus ja vastus ravile (ei ole ühte konkreetset geeni, mis
kujundaks välja astma kui terviku). Pärilikkuse tulemusena tekkinud IgE teatud
variatsioonid võivad koos MHC II klassi molekulidega soodustada tugeva Th2 tüüpi
immuunvastuse kujunemist. Paljud inimesed on seetõttu eelsoodumusega Th2 tüüpi
immuunvastusele ning reageerivad ka mõnedele allergeenidele rohkem kui teistele.
Ka IL-4 geenide variatsioonide tõttu võib esineda eelsoodumus allergiale.
Teatud pärilikku geneetilist variatsiooni IL-4 promootori piirkonnas on seostatud IgE
kõrgemate tasemetega atoopia puhul.
Tähtsamate allergiaga seotud geenide iseloomustus
Kromosoom 5 (5q31…33) – sisaldab omavahel tihedasti seostunud geenide klastrit,
mis sisaldab IL-3, IL-4, IL-5, IL-9, IL-12, IL-13 ja granulotsüütide-makrofaagide
kolooniad stimuleeriva faktori (GM-CSF) geene. Need tsütokiinid mõjutavad IgE
isotüübi muutumist, eosinofiilide ellujäämist, nuumrakkude proliferatsiooni. 5.
kromosoomis on ka β2 adrenergilise retseptori ja glükokortikoidretseptori (GCR) geen,
mille muutumisel võib tekkida glükokortikoidresistentne astma.
Kromosoom 6 (6p) - sisaldab HLA geenide erinevaid alleele, mille ekspresseerumisel
suureneb eelsoodumus nt. heinapalavikule (alleelid HLA-DR5, HLA-DR3), aspiriin-
astmale (HLA-DQ2); sisaldab ka TNF-α geene.
Kromosoom 11 (11q13) - kodeerib FcεRI β- subühiku (FcεRI- IgE esimest tüüpi
retspetor)
Kromosoom 12 - sisaldab IFN-γ, STAT6 (STAT- Signal Transducers and Activators of
Transription- tsütoplasmas olevad signaalmolekulid)
Kromosoom 14 - sisaldab NFκB ja TCR α- ja δ- ahela geene.
Välisfaktorid, mis kutsuvad esile Th2 rakkude aktivatsiooni.
Th2 stimuleerivad harvaesinevate parasiitide (nt. Toxoara, Anisakis simplex) poolt
põhjustatud infektsioonid. Penitsilliini ja keha valkude konjugaadid võivad kutsuda esile
Th2 tüüpi immuunvastuse mõnedel inimestel
Hügieenihüpotees
- “hygiene hypothesis” (ei ole enam kokkupuudet tavaliste haigustekitajatega)
Th2 poole suunamine: IL-3, IL-5, GM-CSF EOS↗
Tüümuse stromaalne lümfopetiin (TSLP): - mõjutab teisi rakke eritama tsütokiine, mis
suunavad Th2 poole diferentseeruma.
Allergia ravi:
Mittespetsiifiline
o Immuunsupressioon – glükokortikoidid
o Mediaatoritele suunatud – antihistamiin
o IgE-ga reageeriv antikeha
Spetsiifiline
desensibiliseeriv ravi
o
Omalizumab – seondub vabale ige-le
o
94.
IgE iseloomustus ja bioloogiline tähendus. IgE tekkemehhanism atoopia
korral. IgE retseptorite iseloomustus. IgE-retseptorite aktivatsioonist
tulenevad rakusisesed muutused. IgG4 ja allergia.
IgE iseloomustus ja bioloogiline tähendus. ~0,004% seerumi antikehadest; monomeerne
(immunoglobuliinidest suurima molekulmassiga); immunoglobuliinidest lühim
poolväärtusaeg; allotüüpne variatsioon. Oluline kaitse mitmerakuliste parasiitide
infektsioonide korral (IgE antikehade poolt vahendatud tsütotoksilisus). IgE tüüpi
immuunvastus on välja kujunenud mitmerakuliste parasiitide (N: soolenugiliste) eest
kaitsmiseks. Oluline roll on siin ka IgG4-l.
Mürkide suhtes (mesilase, taimede). Evolutsiooniliselt oluline meh.
IgE tekkemehhanism atoopia korral.
Atoopia – IgE vahendatud allergia.
Allergeen satub nahale ja limaskestale ja on fagotsüteeritud makrofaagide poolt.
Makrofaagid töötlevad allergeeni ja presenteerivad selle T-helperitele. T-helperid
produtseerivad tsütokiine, mis stimuleerivad B-lümfotsüütide proliferatsiooni ja
indutseerivad IgE sekretsiooni.
Th1 poolt inhibiitorid (IFN-γ, IL-12), lisainhibiitorid: TGF-β, IL-10, IFN-α, PGE2
Th2 poolt indutseerijad (IL-4, IL-13), lisaindutseerijad: IL-6, IL-1, CD23
IgE-retseptorite (FceRI ja FceRII) iseloomustus.FceRI – kõrge afiinsusega, esineb nuumrakkudel, basofiilidel, Langerhansi rakkudel
FceRII – madala afiinsusega retseptor, 2 tüübi: FceRIIa – B rakkudel (stimuleerivad
kasvu), FceRIIb – T, NK rakkudel, makrofaagidel.
IgE-retseptorite aktivatsioonist tulenevad rakusisesed muutused.
IgE seostub nuumraku retseptoriga (FceRI, FceRII), kui allergeen kontakteerub kahe IgE-
ga toimub mediaatorite vabanemibe nuumrakust läbi: Ca++ influksi, seriini proteaasi
aktivatsiooni, cAMP, graanulite transpordi raku pinnale.
Käivitatud protsesside tulemused on:
eksotsütoos (olemasolevate mediaatorite vabastamine: NCF, ECFA –
kemotaktilised faktorid; histamiin, serotoniin; trüptaas (→C3), kininogennas
(→kiniin))
uute ainete süntees: leukotrieenid (LTD4, LTC4), prostaglandiinid (PGD2, PGE2,
PGI2), PAF (trombots. aktiveeriv faktor)
95.
Allergia in vivo ja in vitro testide üldine iseloomustus. Üld-IgE ja
spetsiifilise IgE määramise kliiniline tähendus. Rekombinantsed allergeenid
- uued võimalused täpsemaks diagnostikaks ja raviks.
Allergia in vivo ja in vitro testide üldine iseloomustus.
Testide eesmärk on põhjusliku allergeeni väljaselgitamine ja etioloogilise diagnoosi
kinnitamine. Samuti on testid vajalikud ravitaktika valikuks nt. konkreetse allegreeni
vältimiseks ja spetsiifilise immuunravi määramiseks. Allergiat identifitseerivaid teste on
vähe ja nende kasutamine on piiratud kliinistes laboratooriumides.
In vitro testid:
üld-IgE määramine, ECP-eosinofiilne katioonne proteiin, nuumraku trüptaas
spetsiifiline IgE (RAST jt)
rafiefekti hindamiseks IgG tüüpi antikehad
IgG4 antikehad
basofiilse histamiini vabanemine, IL-4 produktsiooni hindamne. Senini laialt
kasutusel allergeen ekstraktid.
In vivo testid:
nahatestid
provokatsioonitestid jm kliinilised mitte-immunoloogilised testid
Üld-IgE ja spetsiifilise IgE määramise kliiniline tähendus
Üld IgE 1967
Radioimmuunmeetod (RIA)
Ensüümiimmuunmeetod (EIA) –
ELISA (enzyme linked immunosorbent assay)
FEIA (fluoroscence enzyme immuno assay)
CLIA (chemiluminescence immuno assay)
Allergeenspetsiifiline IgE-
Phadebas-RAST, Pharmacia CAP SystemTegemist on uute, automatiseeritud, hästi standardiseeritud laborisüsteemidega.
Tagavad kõrge tundlikkuse ja spetsiifilisuse. Skriiningtestide efektiivsus on >85%.
Paneelid on mitme allergeeniga. S-IgE tiiter ei ole seotud sümptomite raskusega. Kui on
madal spetsiifiline IgE tiiter, siis see ei ole kliiniliselt oluline.
Konjugaat sisaldab radioaktiivset märgistust. EUROIMMUN mikroplate ELISA – üld-IGE
Allergeenspetsiifilise IgE määramist mõjutavad faktorid:
Reaktiivid (kvaliteetsed, standardiseeritud, allergeeniekstraktid, anti-IgE ak)
Optimaalne määramissüsteem (kalibratsioonikõverad, kontrollseerumid)
Uued ja tehniliselt väga tundlikud meetodid-määravad kliiniliselt mitteolulisi rist-
reaktsioone (IgE antikehad mõne epitoobi suhtes)
Ag muutunud vormide ak (töödeldud toit)
Üld IgE ↑ (mittespetsiifiline seostumine)
SIR patsientidel IgG ↑ (vale negatiivsed testid)
Antigeeni eliminatsioonil ak ↓
IgE sessoonsetele allergeenidele väiksemas kontsentratsioonis või määramatu
asümptomaatilises perioodis
Allergeenspetsiifilise IgE eelistus:
Väljendunud dermografism
Ihtüoos, generaliseerunud ekseem
Pikaaegsel antihistamiinsel, antidepressiivsel ravil patsiendid
Halb koostöö patsiendiga
Täiendav laboratoorne test –parasitaarhaiguste korral ja allergilise
bronhopulmonaarse aspergilloosi aktiivsuse hindamisel
Allergeeni pole NTT allergeenekstraktide nimekirjas
Anamnees viitab kõrgenenud anafülaktilise reaktsiooni riskile.
Eosinofiilne katioonne proteiin (ECP) - ECP mõõtmine seerumist on üks allergia
reaktsiooni markereid. ECP on allergilise reaktsiooni mediaator ja põletikurakkude
marker
Trüptaaas – nuumrakkudelt vabanev allergia mediaator – selle märamist kasutatakse
anafülaksia diferentsiaaldiagnostikas, mastotsütoosi või selle kahtluse korral.
Keemiliselt puhtad allergeenid - uued võimalused täpsemaks diagnostikaks ja
raviks.
Seerumiproteiinid
Metallid nt nikkel, kroom
Ravimpreparaadid nt antibiootikumid, sulfaniilamiidid
Lisaained
Jood
bensiin
Loomsed valgud
Ensüümid nt trüpsiin
Ravimid tulevikus: allergeenide fragmendid, tran
sgeensed taimed, IgE blokeerivad
antikehad. Allergia profülaktikaks saaks kasutada taimseid toiduaineid, kus puuduvad
allergeenid nt. taimed kus looduslikult ei esine allergeeni geeni või taimed, mis on
radiatsiooni vms abil muudetud või siis geneetliselt muundatud taimed.
96.
Üldpõhimõtted immunoloogia laboratooriumi töö korralduses.
Laboratoorsete immunoloogiliste uuringute läbiviimise ja nende tulemuste kasutamise
üldised põhimõtted.
Laboratoorse uuringu faasid:
Preanalüütiline (kliinilise probleemi analüüs materjali kogumine);
analüütiline (lab. uuring tulemuste raport);
postanalüütiline (kliiniline raport kliiniline otsustus).
Selles protsessis on väga oluline koht laborispetsialistija raviarstikontaktidel–olulisem kui
teiste laboratoorsete uuringute määramisel ja kliinilisel hindamisel.
Preanalüütilinefaas
Standarduuringud
Mittestandardsed uuringud
Lisaks haiguse olemuse ja diagnostikaga seotud küsimustele (seda otsustab raviarst!)
tuleb koos raviarstiga lahendada sellised olulised küsimused, nagu...
OLULINE! Üldised põhimõtted materjali kogumiseks.
Täpsed andmed uuritava isiku kohta (sugu, vanus, rass ja päritolu jm),
immuunsüsteemi moduleerivatehaiguste, ravitoimingutekohta.
Materjali võtt ja säilitamistingimused –ülitähtsad. Labor peab andma eeskirjad
materjali kogumiseks, transpordiks
Näiteks:
1. Erisused (eeskätt: missugune lisand vereplasma/rakkude saamiseks, mis temperatuuril
materjali säilitada, uuritava materjali kogus, kas on vaja vere võtmine
endotoksiinivabadesse katsutitesse jm);
2. Kui suures hulgas on uuritat materjali vaja (sjvarumiseks kordusanalüüsiks)
Uuringud arhiveeritud materjalist (seerumpank, DNA pank, kudede pank)
Kas materjal sobib uuringuks?
-
Uurimismaterjali peab võtma nii palju kui on hädavajalik, et teha uuring täpses
vastavuses eeskirjadele ja NII VÄHE KUI VÕIMALIK.
Analüütiline faas
Iga testsüsteemi puhul on oluline:
-
analüütiline tundlikkus
-
analüütiline spetsiifilisus (peegeldavad testi tehnilist külge, mitte segamini ajada
kliinilise (diagn) tundlikkusega jm)
Antigeen-antikeha reaktsioonides on oluline reaktantide hulkade omavaheline sobivus
(vältida antikeha ülehulka).
„Standardi EVS-NE ISO 15189 kohaselt peavad meditsiinilised laborid oma
analüüsi tulemuiste usaldatavuse kinnitamiseks osalema ajakohastes
laborivahelistes võrdlustes“ (EAK)
Kõige paremini on see saavutatav vastavate rahvusvaheliste organisatsioonide abil,
kes hoiavad vastavaid kontroll-süsteeme töös (laborid osalevad neis makstes
vastavat tasu).
Ka diagnostilisi komplekte tootvad firma võivad kontrollsüsteeme pakkuda (N:
Immudex –multimer proficiency panel)
Mitmeid immunodiagnostilisi testsüsteeme evalveerivad organisatsioonid -UKNEQAS,
Labqualityjt
Spetsiifilisi testsüsteeme evalveerivad organisatsioonid -IASP jt. (diabeediga
seostuvate autoantikehade testid)
Diagnostilisi meetodeid komplekselt evalveerivadorganisatsioonid
-EASI jt.
(autoimmuunhaigused ja nende diagnostika)
Uuringud jaotatakse CLIA alusel keerulisuse järgi alljärgnevalt:
lihtsad(„waived“) uuringud st. kus on väike võimalus eksimiseks
o Need analüüsid tehakse testide valmistajate eeskirju ja
kontrollimisepõhimõtteid jälgides (HIV, EBV jm antikehad, ravimite monitooring
in vivo jt)
mõõdukalt keerulised uuringud (siia kuulub rida mikroskoopilisi uuringuid)
väga keeruliseduuringud (siin on vajalik v hea tehniline ja teaduslik ettevalmistus)
o N: ANA testid IF-mikroskoopia abil, vererakkude eristamine voolutsütomeetria
abil jt.
Postanalüütiline faas
Labori referentsväärtused(sõltuvad uuritavate vanuselisest, soolisest koosseisust).
Immunoloogiliste (jm. labori-) testide referents-intervall kätkeb tavaliselt 95%
(harvem 99%) tervetel leitavatest uuringutulemustest.
Soovitatav on alati töötada välja testsüsteemi referentsväärtused selle elanikkonna
kohta, kus uuringuid läbi viiakse. Piiripealsetel juhtudel (testitulemuste “hall tsoon”)
määrata sama analüüti teise testsüsteemiga (või saata uurimismaterjal teise
laborisse).
97.
Kvaliteedikontrolli üldised põhimõtted. Referentsväärtused ja nende
saamise tingimused. Uurimistöö ja rahvusvaheline koostöö olulisus.
• Seadme kvaliteedi tagamine
– regulaarne ennetav hooldus kvalifitseeritud tehniku poolt
– igapäevane seadistamine ja kontroll kasutaja poolt
• Tööprotsessi kvaliteedi tagamine
– bioloogiline kontrollmaterjal
• referentsmaterjal, kontrollmaterjal
– antikehade stabiilsus
• ajas ja partiide lõikes
• bioloogiline kontrollmaterjal, sünteetilised kerakesed (beads)
• Median Fluorescence Intensity (MFI)
98.
Faktorid, mis mõjutavad immunoloogiliste uuringute tulemusi laboris.
Analüüsidel tuleb arvestada mitmeid immunoloogilisi faktoreid, mis võivad analüüside
lõpptulemusi mõjutada (kahtlemata on mõju ka tehnilistel mõjudel, nagu aparatuuri
kalibratsioon jms). Lisaks ISO nõuetele:
kasutatavate reaktiivide mittesobivus (kvaliteet! kõrvalekalded toote standardis –eriti
vähetuntud firmade puhul, kus tootmisel puudub piisav kontroll)
NB! uuringut mõjutavad lisakomponendid analüüsitavas materjalis
-reumatoidfaktor – vanus
-autoantikehad analüütide vastu
-heterofiilsed antikehad: looma IgG vastased ak (ravi –OKT3 jm)
-immuunkompleksid
-immunoglobuliinide agregaadid (säilitustingimused!)
ERINEVATE ANALÜÜTIDE MÄÄRAMISEL TULEB ARVESTADA
-Testide valikuga:
N: tsütokiinide puhul
o keemilise struktuuri järgi
o biol. aktiivsuse järgi
N: autoantikehade puhul reageeriva epitoobi struktuuri:
o lineaarne (sobib ELISA, immunoblot)
o ruumiline(sobib RIP,
LIPS)
NB! GADA -SPS puhul ELISA, diabeedi puhul RIP.
-Kas uuringutulemusi võivad mõjutada lisakomponendid uuritavas proovis
agregaadid (mitmekordne seerumi sulatus)jms
muud komponendid (ensüümid)
-Vajalik labori „sisemine kontroll“ (testsüsteemi kontroll võib olla „kunstlik“)
99.
Teadusuuringute tähendus igapäevases laboritöös.
???
100. Immunopretsipitatsioonimeetod antikehade määramiseks.
LIPS – lutsiferaas immunopretsipitatsioon
RIP – RNA immunoprecipitation
101. In vitro testid lümfotsüütide funktsiooni hindamiseks.
In vitro rakulise immuunsuse määramine.
Võimalik määrata rakkude hulka. Seda tehakse voolutsütomeetria (FACS) abil.
Kasutatakse monoklonaalseid antikehi, mida märgistatakse fluorestsentsmikroskoopias.
E rosetid.
(PE), CD3-le (FITC). CD8 retseptoreid võib leida NK rakkude pinnalt. Lümfotsüüdid on
juhendaja sõnul kõige väiksemad ja kõige väiksemate graanulite sisaldusega. Seda on
võimeline voolutsütomeeter eristama teistest klassidest. Lümfotsüütide alaklasse aga
see voolutsütomeeter üksteisest eraldada ei oska. Nendeks klassideks on T, B ja NK. CD3
retseptor on T raku pinnal. B rakke iseloomustab CD19, NK rakke CD16 ja CD56. Need on
pinnamolekulid, mille abil on võimalik klasse eristada. Selleks kasutatakse
monoklonaalseid antikehi. Meie lisasime kahte antikeha – CD-le saab eristada teistest
rakkudest CD14 retseptoritega. Kõikide leukotsüütide pinnal on CD45.
Definitsioon (FACS). Osakesed viiakse vedeliku laminaarse voolu abil tsütomeetri sisse
detekteerimse punkti. Rakud peavad olema vedelikus. FACS – flourescence-activated cell
sorting. Koosneb – voolutusosast, optilisest osast (genereerib laseritega valgussignaale),
elektroonika osa – võimendab signaale, konverteerib neid ja salvestab. Nõel, mis ulatub
katseklaasi. Selle abil detekteerimispunkti. Sinna sattunud osake murrab, hajutab
valgust. Osa aga peegeldab organellidelt ja graanulitelt tagasi – seda osa nimetatakse
side scatter e SSC. Mida rohkem on osakese sees graanuleid, seda suurem on side
scatter. Nende valguse hajutamise parameetrite järgi identifitseritakse lümfotsüütide,
granulotsüütide ja leukotsüütide gruppe.
Samuti florestseeruvate gruppide eristamine – emiteeritava valguse intensiivsus on
võrdeline osakeste poolt seotud florokroomi hulgaga. Need antikehad, mis on seotud
värvainega, seostuvad vastavate retseptoritega (CD-d), ja laserid aktiveerivad seda
värvainet, ja selle püüab optiline osa kinni. Sellest tulebki tulemus. Florestsents signaalid
filtreeritakse valgusfiltritega. Iga signaal jõuab seega oma detektori juurde. Mõned
tuntumad florokloorid – sinine laser – 488 nm lainepikkus.
Molekulid erinevad üksteisest nii ergastus kui emissiooni spektri poolest. Sellega peavad
kasutatavad florokroomid olema ergastatavad vastava laseri valguse lainepikkuse poolt.
Ühel osakesel saab määrata – suurus, granulaarsus, nelja erinevat florestsentsi. Detektor
genereerib elektrilise impulsi, mis on proportsionaalne valgussignaali tugevusega.
Analoogdigitaal konverter ja salvestaja. Üheks võimaluseks on näidata tulemusi
sageduste jaotuse e. histogrammiga. Ühel teljel rakkude arv, teisel signaali tugevus.
Teine võimalus on näha tulemust dot plotina. Võrreldakse kahte parameetrit omavahel.
Igale rakule vastab tabelis üks punkt. Edasi analüüsitakse järgmistes katsutites ainult
lümfotsüüte – sinna antikehad – isotüübi kontroll. Vastavalt IgG1 või IgG2.
Mittespetsiifilise seostumise näitamiseks. Järgmisse antikehad CD3 ja CD19 vastu.
Sellega saab eristada T ja B lümfotsüüte. Järgmine CD3 ja CD4 antikehad. CD3/CD8. Siis
CD3, CD16/CD56. Arstile trükitakse välja vastus koos normiväärtustega.
102. In vitro testid rakkude tsütotoksilise funktsiooni hindamiseks.
CDC – komplement sõltuv tsütotoksilisus
HLA klass I vastaste tsütotoksiliste antikehade olemasolu ja hulga hindamine ning
antikehade spetsiifilisuse määramine komplemendist sõltuval lümfotsütotoksilisel
meetodil:
mikrotiiterplaat standardiseeritud lümfotsüütidega
uuritav seerum
komplement
värvaine
reaktsiooni hindamine (mikroskoop)
Vastuse vorm: PRA% (panel reactive antibodies)
Kui on võimalik tuvastada, millise konkreetse antigeeni vastu antikehad esinevad,
märgitakse vastav antigeen
103. Lümfotsüütide uuringute kliiniline kasutamine.
Diagnoos.
104. Lümfotsüütide intratsellulaarsete ja pinnamarkerite määramine.
Tähendus.
FACS – allpool
antikehadega inkubeerimine, lüüsimine, pesemine, seadmes loendamine
105. Voolutsütomeetria (FACS) põhimõte ja kasutamine.
Voolutsütomeeter saab määrata koerakkude tüüpi. Suspensioonis rakke mõõdab
ukshaava. Rakkud peavad olema ühekaupa, kudede puhul töödeldatakse neid
mehhaniliselt või ensümaatiliselt. FACS fluorescense activated cell sorting.
– Laserkiirgust kasutav meetod, mille abil on võimalik iseloomustada rakkude füüsilisi ja
keemilisi tunnuseid
– Mitmed tunnused samaaegselt
• Struktuursed tunnused (suurus, tsütoplasmaatiline koostis, fotosünteetilised
pigmendid)
• Funktsionaalsed tunnused (pinna- ja tsütoplasmaatilised retseptorid, DNA-RNA sisaldu
– kvalitiivsed ja kvantitatiivsed
Põhiprintsiip
Rakkude märgistamine fluorokroomidega
Märgistatud rakkude suunamine laservalgusesse
Fluorokroomi ergastamine ja fluorestsentsi detekteerimine
Fluorestsentssignaali muutmine digitaalseks
Voolutsütomeetria kliinilises diagnostikas
• Immunoloogia
– primaarsete immuunpuudulikkuste diagnoosimine (T-, B-, NK-rakud)
– CD4+ T-rakkude hulga määramine
• Hematopatoloogia
– leukeemiate/lümfoomide immuunfenotüpeerimine
– vereloome tüvirakkude loendamine ja hindamine
• Transfusioonimeditsiin
– fitreeritud verepoduktidest jääkleukotsüütide loendamine
106. In vitro testide põhimõtted antikehade ja antigeenide määramiseks.
Avidiin-biotiinkompleksi tekkel põhinevate meetodite rakendus.
Antikehade omadused, mis on olulised analüütilistes testides:
Antikeha võime tunda ära väga laia spektrit erinevaid antigeene, nii looduses
esinevaid kui sünteetilisi molekule. Selle tagab antikeha hüpervariaabelset
regiooni moodustavate lingude aminohapete võimalike järjestuste
kombinatsioonivariantide hiigelsuur arv.
Erakordne spetsiifilisus substantsi suhtes, millega antikehad reageerivad. See
võimaldab detekteerida väga sarnaseid aineid.
Antigeen ja antikeha seondumise tugevus e afiinsus. Antikeha ja antigeen
seonduvad mittekovalentselt. Siiski piisavalt tugevalt, et side säiluks inkubatsiooni
ja signaali tekkimise etappides.
Immuunmeetodites kasutatakse antikehi, et detekteerida analüüdi molekule.
Kasutatakse polüklonaalseid või monoklonaalseid antikehi. Polüklonaalsed antikehad
toodetakse katselooma korduval immuniseerimisega puhastatud antigeenidega.
Katseloomal areneb tugev immuunvastus, mis annab kõrge afiinsusega ja ja
spetsiifilisusega antikehad. Need antikehad on erinevate parameetritega, tunnevad ära
erinevaid epitoope samal antigeenil. Katseloomalt kogutakse seejärel korduvalt seerumit,
millest eraldatakse afiinsuskromatograafia abil välja meid huvitavad antikehad. Nii saab
toota ka antikehade vastaseid antikehi, nt kui küülikut immuniseerida inimese IgG
fraktsiooniga. Nii saab toota inimese IgG vastaseid küüliku polüklonaalseid antikehi.
Monoklonaalseid antikehi saab ühe raku kloonimisega. Katselooma immuniseeritakse
korduvalt puhaste antigeenidega. Põrnast eraldatakse B-lümfotsüüte, mida
stimuleeritakse paljunema ja antikehi tootma vastava antigeeni vastu. Kuna B-lümfotsüüt
on diferentseerunud rakk, siis selle eluiga on piiratud, samuti paljunemiste arv.
Lümfotsüüdi rakud segatakse müeloomi rakkudega, millele on omane piiramatu
jagunemiste arv. Juhuslikult kõrvuti sattunud müeloomi ja lümfotsüüdi rakud ühendavad
oma membraanid, tekib hübriidrakk. Segusse jäävad müeloomi rakud, lümfotsüüdid ja
hübriidid. Segu kultiveeritakse. Teatud ainetega (HAT) blokeeritakse müeloomirakkude
metaboolsed rajad, samuti hukkuvad diferentseerunud lümfotsüüdid. Allesjäänud rakud
jagatakse mikrotiiterplaadile nii, et ühte auku satub 1 rakk. Edasi selekteeritakse
antigeenile vastavad antikehi toottvad hübriidrakud, neid paljundatakse rakukultuuri
keskkonnas. Nii saab selekteerida enim kloone tootvaid hübriidrakke, spetsiaalse
epitoobiga rakke, kõrge afiinsusega antikehi tootvaid rakke, saab välistada
ristreaktsioonid. NB sel juhul pärinevad saadud antikehad kõik ühest eellasrakust, st
ühes mikrotiiterplaadi augus olevast hübriidrakust.
Immuunmeetodeid saab klassifitseerida kahe kategooria alusel. Esiteks
ülesehitusprintsiibi alusel, teiseks testid, kus testi täpsuse, inkubatsiooniaja lühendamise,
lihtsustamise või parema automatiseerimise saavutamiseks kasutatakse esmaste
ülesehtusprintsiipide variante. Tänapäeval on vajalik, et primaarne reaktsioon oleks
nähtav, mõõdetav. Primaarseks reaktsiooniks on antikeha ja antigeeni seondumine, mis
põhjustab mingeid füüsikaliste parameetrite muutusi nagu pretsipitatsioon,
aglutinatsioon, mis oleksid ühtlasi sekundaarseks reaktsiooniks. Immuunmeetodid, mis
määravad otseselt primaarreaktsiooni on tundlikumad ja täpsemad, kui
sekundaarreaktsioonid määravad reaktsioonid. Kaks peamist ülesehitusprintsiipi on
sändvitš- ja konkurentsiprintsiip. Eristatakse veel homogeenseid ja heterogeenseid
immuunmeetodeid. Heterogeensetes immuunmeetodites on kasutusel mitu
inkubatsioonietappi, mille vahel on pesuetapid mitteseondunud ja seondunud
fraktsioonide eraldamiseks. Homogeensed immuunmeetodid ei vaja fraktsioonide
eraldamist, on üheetapilised. Immuunteste saab kasutada kas antigeeni või antikeha
määramiseks. Üks reaktsioonipool on testi standardiseeritud komponent, teine aga
analüüt. Analüüt tuleb uuritavast materjalist. Lisaks osaleb testides tavaliselt veel mõni
komponent (antigeen ja antikeha).
Sändvitšprintsiip: tahkele faasile on kantud antigeenid (standardiseeritud komponent),
millele valatakse peale uuritavat materjali e analüüsitavaid antikehi. Nende antikehade
vastaseid antikehi kasutatakse esimeste detekteerimiseks. Antikehade vastased
antikehad on märgistatud nt fluorestseeruvate osakestega. Sändvitšmeetodiga ei saa
määrata madalmolekulaarseid antikehi. Mida rohkem märgistatud antikehad ühinevad
vastavate antikehadega, seda tugevama signaali saab detekteerida. Enne signaali
detekteerimist toimub seondumata antikehade eemaldamine/pesemine. Kuna antigeene
tahkel kandjal on kindel hulk, siis tegemist on küllastuva protsessiga, analüüdi
konstentratsiooni ja antigeen-antikeha-antikeha seostumine on võrdelises seoses.
Konkurentsiprintsiip: tahkele kandjale on seostatud antikeha, millega seondumise pärast
konkureerivad omavahel analüüt ja analüüdiga identne, kuid märgistusega molekul
(tracer). Antud printsiibi puhul on teada tahkele faasile seotud antikeha kontsentratsioon
ja traceri konstentratsioon. Kuna traceri ja analüüdi afiinsus antikehale on sama, siis
sõltub seondumine konstentratsioonide suhtest. Signaali tugevuse (sõltub traceri
seostumisest antikehaga) ja analüüdi kontsentratsiooni vahel valitseb pöördvõrdeline
seos.
Meditsiinis kasutatakse immuunmeetodeid nakkushaiguste, autoimmuunhaiguste,
atoopilise allergia diagnostikas. Väga mitmeid biomolekule määratakse antigeen-
antikeha reaktsioonil põhinevate meetoditega. Samuti kasutatakse immuunmeetodeid
mitmete patoloogiate kindlakstegemisel ja diagnostikas.
Immunoblot-analüüs koosneb kolmest etapist. Esiteks toimub valkude elektroforees,
mille tulemusena lahutatakse antigeenid, kasutatakse SDS-PAGE. Toimub ca 1h. Teiseks
etapiks on blotting e lahutatud antigeenide ülekanne geelist (SDS-PAGE)
nitrotselluloosfiltrile. Kolmandaks viiakse läbi antigeen-antikeha reaktsioon. Selleks
blokeeritakse nitrotselluloos „neutraalsete“ valkudega (nt rasvatu piimapulber).
Primaarne antikeha lisatakse süsteemi, st seerumlahjendus. Seejärel lastakse sellel
seista, st inkubatsioon seerumlahjendusega. Pärast inkubatsiooniaega detekteeritakse
tekkinud antigeen-antikeha kompleksid. Sekundaarne antikeha on märgistatud kas
ensüümmärgistusega (peroksüdaas, alkaalne fosfataas), mille korral toimub
detekteerimine kromogeensuse või kemoluminestsentsi alusel, või radioaktiivse
isotoobiga, detekteeritakse autoradiograafia abil. Immunoblott-analüüsil on kõrge
spetsiifilisus, sest toimub antigeenide lahutamine elektroforeesiga. Kõrge tundlikkuse
annab antigeen-antikeha vaheline immunoloogiline reaktsioon. SDS denatureerib valgud,
st säilivad lineaarsed epitoobid, konformatsioonilised kaovad. Kliinilises diagnostikas on
immunobloti kasutamine piiratud, sest ta on aeganõudev ja raskelt automatiseeritav.
100% puhaste antigeenide kasutamine iseloomustab ELISA meetodit, immunoblott
võimaldab aga analüüsida nn musti antigeene. Immunoblotti kasutatakse
referentstestina mitmete nakkushaiguste serodiagnostikas (HIV, H. pylori, Borrelia
burgdorferii). Skriinimiseks immunoblott ei sobi. Veel kasutatakse seda meetodit
autoantikehade määramiseks (tuumaantikehade leiu diferentseerimiseks, ENA-
Extractable Nuclear Antigens- antikehad), teaduslikes eksperimentides haiguste
patogeneesimehhanismide selgitamisel, autoantigeenide kindlakstegemisel,
molekulaarbioloogias.
Serodiagnostika annab tõendeid infektsiooni olemasolust või läbipõdemisest. Põhineb
seerumis määratavatel antikehadel. Antikehade olemasolu aga ei võimalda eristada, kas
leitud antikehad pärinevad antud hetkel olevast või varasemast infektsioonist. Haiguse
ajal kogutakse kaks seerumit kahe või kolme nädalase vahega. Samaaegselt määratakse
seerumites antikehade taseme e tiitri tõus. Tiitri tõus infektsiooni käigus peab ületama
teatud kriitilise väärtuse. Antikehade isotüüpide dünaamika uurimisel saab teha järledusi
infektsiooni kulu kohta. IgM tekib ägeda infektsiooni korral esimesena, paranemisel see
kaob seerumist kiiresti. IgG tekib aeglasemalt, kuid jääb seerumisse kauemaks.
Serokonversioon on hetk, mil antigeen seerumist kaob ja selle asemele tulevad
antikehad. Infektsiooni alguses esineb antigeneemia, sel ajal antikehade tootmine ei
ületa viiruspartiklite tootmist ja tekkivad antikehad kasutatakse viiruspartikliga
immuunkompleksi tekkeks. Serokonversiooni ajal on oht, et haigust ei diagnoosita, sest
diagnostilised antigeen ja antikeha on negatiivsed, st alla määramispiiri.
Täisautomaatsetele immunoanalüsaatoritele on omane käsitöö minimaalsus. Neil on iga
proovi jaoks individuaalsed testühikud. Samaaegselt saab teha mitut analüüsi ja vastus
saadakse samal päeval. Ei vaja igakordset kalibreerimist. Võimaldavad kvantitatiivselt
mõõta erinevaid antikehi ja antigeene (hormoone, tuumormarkereid, allergeenspetsiifilist
IgE-d). Kõrge analüütilise tundlikkuse tagavad märgise ja selle signaali
detekteerimisviisid. Valdavalt on kasutusel ECL e võimendatud kemoluminestsents.
Automaatsete immunoanalüsaatorite analüütiline tundlikkus ja täpsus on sarnased RIA-
le, mistõttu on ta RIA-l baseeruvaid teste asendanud. Automatiseeritud
immunoanalüsaatorites kasutatakse kõrge tundlikkusega mitteradioaktiivseid
detekteerimissüsteeme nagu aeg-lahutatud fluorestsents (time-resolved fluorescence),
võimendatud kemoluminestsents (ECL), FEIA- FluoroEnzymeImmunoAssay. Aeg-
lahutatud fluorestsentsmeetodil kasutatakse märgiseks lantanoidi aatomi.
Fluorestsentssignaali mõõtmine toimub aeg-lahutatult, st ergastamise ja signaali
mõõtmise vahel on paus. See on vajalik selleks, et vaibuksid seerumjääkide (NADH,
proteiinid, bilirubiin) autofluorestsents, väheneb taustfluorestsents. Võimalik on mõõta ka
väga nõrka signaali. Signaali võimendamine toimub võimendava lahusega, mille toimel
lantanoidmärgis dissotsieerub antikeha küljest ning tema ümber tekib mitsell. Sel viisil
võimendub fluorestsents ca 105 korda. Võimaldab detekteerida üksikuid antigeen-
antikeha reaktsioone. Võimendatud kemoluminestsentsmeetodil on kõrge analüütiline
tundlikkus tänu enhanceri kasutamisele.
Immuundifusioon-meetod. Immuundifusioon viiakse läbi geelis (agargeel Petri tassis).
Antigeenid ja antikehad liiguvad geelil soojusliikumise ja kontsentratsiooni gradiendi
tõttu, moodustades kohtudes immuunkomplekse. Immuunkomplekside suurused
sõltuvad anitgeenide ja antikehade kontsentratsioonide suhtest reaktsioonikeskkonnas.
Suuremad immuunkompleksid murravad valgust oluliselt rohkem. On
topeltimmunodifusioon (Ouchterlony meetod), mis võimaldab kvalitatiivselt määrata
antigeene või antikehi (st vastab küsimusele jah/ei, üks või teine). Radiaalse
immunodifusiooniga (Mancini meetodiga) saab kvantitaviiselt määrata antigeene või
antikehi, st nende kontsentratsioonide määramist. Radiaalse immuunodifusiooni puhul
on üks komponent valatud geeli sisse, puudub kontsentratsioonigradient ning difusiooni
ei esine. Uuritava kontsentratsiooniga komponent asetatakse geeli süvenditesse, uuritav
komponent hakkab difundeeruma kontsentratsiooni vähenemise suunas. Seos
kontsentratsiooni languse ja difundeerumisraadiuse vahel on logaritmiline, kusjuures ka
mõõtmisviga võimendub logaritmiliselt. Kolmandaks immuundifusiooni meetodi
võimaluseks on selle kombineerimine elektroforeesiga. Selle juures eristatakse immuno-,
rakett- ja vastuvoolu elektroforeesi. Immuundifusioonimeetodite analüütiline tundlikkus
on tagasihoidlik, vajab mõlemat reaktsioonipoolt küllaltki suures koguses.
Antigeenide määramine veres ja kudedes:
Turbido- ja nefelomeetrias mõõdetakse immuunreaktsioon valguse hajumise fenomeni
kaudu. Nefelomeetrias lahuse hägususe suurenedes valgusvoog detektorisse suureneb.
Turbidomeetrias lahuse hägususe suurenedes valgusvoog detektorisse väheneb. Neid
meetodeid kasutatakse vereplasma valkude kvantitatiivseks määramiseks, ka uriinist,
tserebrospinaalvedelikust.
Komplemendi fiksatsioonitest: antigeen-antikeha kompleksi tekte detekteerimiseks
kasutatakse komplemendi funktsiooni, st aktiveerumine immuunkompleksi toimel, lüüsiv
toime.
Aglutinatsioonil põhinevad meetodid:
Hemaglutinatsioonitestid: antigeen-antikeha kompleksi teke toimub erütrotsüüdi pinnal,
mistõttu tekib erütrotsüütide sadestumine- hemaglutinatsioon. Selle visualiseerimiseks
on mitmeid võimalusi. Hemaglutinatsioonteste kasutatakse veregruppide määramisel.
Coombs`i testid: Otsene Coombs`i test määrab immuunkompleksidega kaetud
erütrotsüütide olemasolu. (Nt reesuskonflikti korral võetakse lootelt verd, kus
eraldatakse erütrotsüüdid, mis on kaetud ema antikehadega. Neile lisatakse katselooma
anti-antikeha. Tekib aglutinatsioon.) Kaudne aga määrab erütrotsüütide membraani
antigeeni vastase antikeha olemasolu uuritavas seerumis. (Nt reesuskonflikti korral
võetakse ema seerum, kuhu lisatakse Rh+ erütrotsüüte. Pestakse ära seondumata
antikehad. Segule lisatakse katselooma anti-inimese antikehi. Toimub aglutinatsioon.)
Lateksaglutinatsioon: pole kõige usaldusväärsem. Immuunfluorestsentsmeetod: põhineb
sändvitš- või konkurentsiprintsiibil. Märgistusena kasutatakse fluorestseeruvaid osakesi.
Märgistatud antikehad seostuvad antigeeniga, tekib immuunkompleks, mida saab
märgistuse tõttu detekteerida. Meetodit saab kasutada erinevate antigeenide või
antikehade uurimiseks fluorestsentsmikroskoobis. (Wikipeedia ütles, et enamasti
kasutatakse sändvitšprintsiipi immunofluorestsentsmeetodis, et siis esmalt seostuvad
uuritava komponendi antikehad tahke faasiga seondunud antigeenidega. Seejärel
pestakse mitte-seostunud antikehad maha ja lisatakse märgistatud antikehi, mis peaks
olema anti-antikehad. Seostunud jäävad, mitte-seondunud pestakse maha. Edasi
vaadeldakse tulemust mikroskoobis.)
ELISA (Enzyme Linked Immunosorbent Assay): ensüümseotud immuunsorbent-
meetod.
Kliinilises praktikas baseerub ELISAl paljude antikehade ja haigustekitajate antigeenide
määramine. ELISA koosneb mitmest etapist:
ELISA plaadi (96-auguga mikrotiiterplaat) augud kaetakse antigeenilahusega,
toimub antigeeni valgu seondumine plastikpinna külge. Edasi välditakse
mittespetsiifiline seostumine, nt valgu kaseiini abil.
Kantakse peale analüüsitav seerumlahjendus (primaarne antikeha) ja
inkubeeritakse.
Inkubatsioon ensüümiga (AP- alkaalne fosfataas või HRP- peroksüdaas), mis on
konjugeerunud sekundaarse antikehaga.
Märgise hulga mõõtmine- ensüümaktiivsus määratakse kromogeense substraadi
abil. Substraadilahus muutub ensüümaktiivsuse toimel värviliseks, spetsiaalse
kolorimeetri (ELISA plaadi lugeja) abil mõõdetakse igas augus värvuse intensiivsus
(optiline tihedus).
ELISA sobib
suurte analüüside hulga skriinimiseks, tema analüütiline tundlikkus on
väiksem kui radioimmuunmeetodil, kasutatavad reaktiivid on aga stabiilsemad.
Kasutatakse ära ensüümi katalüüsivaid omadusi, neutraalse substraadi ensümaatilisel
töötlemisel tekib värviline produkt, mida saab detekteerida. Kasutatavad ensüümid pole
substraadi suhtes valivad.Tegemist on põhimõtteliselt sändvitšmeetodiga. Plaat võib
kaetud olla ühe antigeeni või mitme erineva antigeeniga. Antigeeni valgu seondumine
plaadiga on mittespetsiifiline, kuid piisavalt tugev, et komponendid ei lahkneks, neid
hoiavad koos van der Waalsi jõud.
RIA (Radioimmunoassay): radioimmuunmeetod. RIA baseerub konkurentsiprintsiibil. Kaks
antigeeni (märgistamata analüüt ja märgistatud antigeen e tracer), mille antigeensed
omadused on identsed või väga sarnased, konkureerivad antikeha sidumiskohtade
pärast. Pärast konkurentsi seostumisele on antigeenide suhe võrdeline nende suhtega
testsüsteemis. Kuna traceri hulk on teada, siis mõõtes radioaktiivsust, saab
kalibratsioonigraafiku abil määrata analüüdi konstentratsiooni uuritavas materjalis. RIA
on täpsem kui ELISA, kõrgema analüütilise tundlikkusega. RIAt kasutatakse ravimite ja
metaboliitide, hormoonide, tuumormarkerite kvantitatiivseks määramiseks. RIA puhul on
minimaalselt probleeme mittespetsiifiliste taustreaktsioonide või valepositiivsusega.
Samas on RIA kasutamisel puudusteks isotoopide poolestusajad, pikk analüüside
kogumisaeg, radiatsioonirisk, mitteautomatiseeritavus. RIA ei sobi üksikseerumi
analüüsiks.
Kiirtestid: Immunokromatograafial põhinevates kiirtestides kasutatakse suunatud
vedelikuvoolu immuunkromatograafia ribal, seda inkubatsioonietapi asemel. Testid
põhinevad kas sändvitš- või konkurentsiprintsiibil. Tulemus visualiseeritakse värviliste
mikropartikkel-märgiste abil. Immuunkromatograafial põhinev uriini hCG testis toimub
immuunkromatograafia filtermaterjali ribal, kus tekib suunatud vedelikuvool filtri
kapillaarse märgumise tõttu. Voolav vedelik tirib kaasa lüofiliseeritud anti-αhCG, mis on
märgistatud sinise latekspartikliga. Filtermaterjalil on triibuna kantud anti-βhCG ja hiire
IgG-vastane antikeha. Antigeeni (hCG) olemasolul uuritavas materjalis (uriinis) tekib teise
antikeha triibu (anti-βhCG) kohale silmaga nähtav sinine joon. Antigeen (hCG) seob
esmalt anti-αhCG ja seejärel anti-βhCG, mistõttu tekibki viimase antikeha
lokalisatsioonikohta silmaga nähtav sinine joon (anti-αhCG on ju märgistatud
107. Rekombinantsed antikehad, üheahelalised antikehad (scFv).
Rekombinantsed antikehad on antikeha fragmendid, mis on saadud, kasutades
rekombinantse antikeha kodeerivaid geene. Üldjuhul koosnevad Ig varieeruvate
regioonide kergest ja raskest ahelast. Kõige tavalisem on üheahelaline varieeruv
fragment (scFv), sellel on kõige lubavamad tulemused nii meditsiinis kui ka teaduses.
Rekombinantsed antikehad, mis on toodetud faagi displei kaudu säilitavad kõrge
spetsiifilisuse ja madala immunogeensuse. See on erinev monoklonaalsetest
antikehadest, mis võivad kaotada oma võime toota antikehi pika aja jooksul, samuti
võivad nad läbida muutusi.
On mitmeid erinevaid rekombinantseid antikehi:
-
Fab rekombinantsed antikehad
-
scFv
-
diakehad
-
anti-idiopaatilised
-
jne
Igaühel neist on erinev rakenduslik potentsiaal ja neid võib kasutada nii inimeste kui ka
loomade meditsiinis. Anti-idiopaatilised seonduvad teise spetsiifilise antikeha
paratoobile. Seetõttu saab neid kasutada antikehade hulga määramiseks patsiendi
seerumis.
scFv on kõige väiksemad rekombinantsed antikehad, mis suudavad antigeeni siduda.
Kaks ahelat, kerge ja raske, on ühendatud paindliku linkeriga, mis stabiliseerib antikeha.
Seondumise aktiivsuse hindamiseks kasutatakse ELISA testi.
Rekombinantsete antikehad tootmine
Kõigil enam-vähem samamoodi:
-
järjestuse kindlakstegemine
-
geeni süntees
-
kokkupanek
-
ekspresseerivate konstruktide kokkupanek
-
transfektsiooniga rakkudesse
-
kui ekspresseerivad vastavat antikeha, siis puhatsamine ja analüüs
Peab olema tagatud õige voltumine ja scFv puhul ka õige kokkupanek raskest ja kergest
ahelast.
Hübridoome kasutatakse monoklonaalsete antikehade tootmiseks. Monoklonaalsetest
saab panna kokku Fab fragmente, scFv või bispetsiifilisi antikehi.
Kõige tavalisemalt kasutatakse rekombinantsete antikehade tootmiseks faagi displeid.
See on meetod, mille puhul huvipakkuv rekombinantne antikeha toodetakse
bakteriofaagi peal. See võimaldab kiiret rekombinantse valgu tootmist ja kerget
manipuleerimist labori tingimustes. Nii scFv kui ka Fab toodetakse nii igapäevaselt. Kõige
tavalisem on E.coli süsteem, kuna paljuneb kiiresti, kasvab hästi, on odav.
scFv saamiseks kasutatakse kahte strateegiat:
-
mittekolineaarne lähenemine: kahe ahela heterodimeriseerimine. Selle käigus
toodetakse diakehasid ja rekombinantseid antikehi, millel on kaks spetsiifilisust
-
kolineaarne lähenemine: kahe erineva scFv ühendamine bioloogiliselt aktiivs valgu
abil
Rekombinantsed antikehad teevad haiguste diagnoosimise mitmekesisemaks.
108. Antikehade rakendusi kromatograafias.
Immobiliseeritud antikehad kandjal (geeli-graanulid, magnetiseeruvad
osakesed jt.)
Kiirtestid: Immunokromatograafial põhinevates kiirtestides kasutatakse suunatud
vedelikuvoolu immuunkromatograafia ribal, seda inkubatsioonietapi asemel. Testid
põhinevad kas sändvitš- või konkurentsiprintsiibil. Tulemus visualiseeritakse värviliste
mikropartikkel-märgiste abil. Immuunkromatograafial põhinev uriini hCG testis toimub
immuunkromatograafia filtermaterjali ribal, kus tekib suunatud vedelikuvool filtri
kapillaarse märgumise tõttu. Voolav vedelik tirib kaasa lüofiliseeritud anti-αhCG, mis on
märgistatud sinise latekspartikliga. Filtermaterjalil on triibuna kantud anti-βhCG ja hiire
IgG-vastane antikeha. Antigeeni (hCG) olemasolul uuritavas materjalis (uriinis) tekib teise
antikeha triibu (anti-βhCG) kohale silmaga nähtav sinine joon. Antigeen (hCG) seob
esmalt anti-αhCG ja seejärel anti-βhCG, mistõttu tekibki viimase antikeha
lokalisatsioonikohta silmaga nähtav sinine joon (anti-αhCG on ju märgistatud).
109. Antikehade iseloomustamine biosensorite abil.
Immuunosensorid
Immobiliseeritakse antikehad (polü- või monokloonsed)
Immobiliseeritakse antikeha fragmendid (Fab , scFv)
Immobilisseritakse antikehadega sarnased molekulid
Antikehadel kasutatakse erinevaid märgiseid (AP, HRP, fluorokroomid jpt)
Detekteeritakse
Elektrokeemiliste meetoditega (potentsiomeetria, amperomeetria, konduktomeetria)
Optiliste meetoditega (peegeldus-spektroskoopia, ellipsomeetria, pinna plasmon
resonants, fluoresents)
110. Antikeha-kiibid.
Pinnale immobiliseeritakse antikeha
Märgistatud proov kantakse kiibile (Cy3, Cy5)
Seondumine detekteeritakse fluorestsentsi mõõtmisega
Tulemuste alusel saab võrrelda valkude ekspressiooni- taseme muutusi erinevates
rakkudes
111. Antikehade rakendusi proteoomikas. HUPO.
Human Proteome Organisation
Antikehade kasutamine inimese proteoomi kaardistamiseks.
Valguatlas – inimese valgud normaalsetes/haiguse kudedes. Valideeritud antikehad.
Eesmärgiks identifitseerida inimese valgud ja nende lokalisatsioon erinevates kudedes ja
rakkudes
– Vereplasma
– Aju
– Maks
– Neerud ja uriin
– Tüvirakkude bioloogia
– Kardiovaskulaarproteoomika
– Haiguste hiiremudelid
– Haiguste biomarkerid
Antikehade initsiatiivi eesmärk- luua kõigi valkude vastased antikehad (rekombinantsed
valgud mono- v polükloonsed ak-d
112. Autoantikehad töövahenditena (cDNA raamatukogude skriining jms.).
Vähispetsiifiliste autoantigeenide vastased autoantikehad
SEREX analüüs – vähikoest valmistatud cDNA raamatukogu skriining haige
vereseerumiga
113. Immunofluorestsents- ja immunoensüüm meetodid immunohistokeemias.
Immunohistokeemiliste meetodite üldiseloomustus. Immunohistokeemiliste meetodite
kasutusvõimalusi. IHC hõlmab endas koelõigus selektiivselt antigeeni esitlemist tavalist
antigeen-antikeha printsiipi kasutades. IHC kasutatakse laialdaselt ebanormaalsete
rakkude diagnoosimiseks. Seda kasuatakse ka biomarkerite ja nende asukoha
kindlakstegemisel. Selle kasutamiseks on mitmeid meetodeid, kõige tavalisem
peroksidaasidega, mis katalüüsivad värvi tootvat reaktsiooni. Samas saab antikeha
panna ka fluorokroomi külge.
Kasutatakse polü- või monoklonaalseid antikehi. Kasutatkse primaarseid ehk antigeeni
vastu, märgiseta ja sekundaarseid ehk primaarse Ig vastu, märgistatud antikehi.
Sekundaarsel antikehal võivad olla küljes linkerid, nt biotiin.
Reporterid võivad olla kromogeensed (ensüümid) või fluorestseeruvad (fluoropoor).
Kromogeensete puhul moodustab ensüüm substraadiga reageerides intensiivse värvi,
mida saab vaadelda valgusmikroskoobis. Kasutatakse aluselist fosfotaasi (AP) või
mädarõika peroksüdaasi (HRP). Moodustuvad pruun (huvipakkuv marker, rakk) ja lillakas
värv (ümbritsevad rakud).
Fluorestseeruvad reporterid on väikesed, orgaanilised molekulid, nt FITC,
TRITC, AMCA, on ka kommersiaalseid, nt Alexa Fluorid.
Mõlema meetodi puhul saadakse kvantitatiivsed andmed, võrreldakse reporteri signaali
ning valgu ekspressiooni ja/või asukohta.
Otsene meetod: hõlmab üheastmelist värvimist, kasutatakse märgistatud antikeha
(FITC), mis reageerib otse antigeeniga koelõikudel. Tundlikkus madal.
Kaudne meetod: hõlmab märgiseta primaarset antikeha, mis seondub antigeeniga koes
ning märgisega sekundaarset antikeha, mis reageerib primaarse antikehaga. On
spetsiifilisem kui otsene.
Võidakse kasutada lisaks IHC lisavärvimist, et esmane paistaks paremini välja.
Kasutatakse selleks hematoksüliini, DAPI.
IHC on suurepärane detekteerimise meetod, sest näitab väga täpselt ära, kus uuritav
valk koes asub. Kasutatakse palju neuroteaduses. Miinuseks on see, et ei ole millegi
vastu võrrelda, kas värvus tuleb ikka õigelt valgult. Seetõttu tuleb primaarset antikeha
hoolikalt valida.
IHC-d kasutatakse kasvajate immuunofenotüpiseerimises. Ka süljenäärmete, pea ja kaela
kartsinoomide diagnoosimiseks.
IHC saab määrata, milline ravi võiks olla konkreetsele kasvajale parim.
IHC saab kasutada ka valkude paremaks kirjeldamiseks.
114. Immunofluorestsents- ja immunoensüüm meetodid immunohistokeemias.
Eelmises?
115. Uued võimalused immunotehnoloogilisteks arendusteks.
Immumotehnoloogia on tehnoloogia, mis baseerub imuunsüsteemi molekulide ja rakkude
rakendamisel ravis, diagnostikas ja teaduses. Võimaldab erinevaid immuunmeetodeid
kasutades teha kindlaks vastavate antigeen-antikeha komplekside moodustumine ja
selle alusel näiteks hinnata, millise haigustekitajaga (patogeeniga) on tegemist. Välja on
töötatud mitmeid erinevaid viise haiguste diagnoosimiseks ehk siis laboratoorseid teiste
antigeenide ja antikehade määramiseks, nt nahatestid, ülitundlikkustestid,
imminotoksikoloogilised uuringud; immunosupressiivsed ravimid, mis võimaldavad
imuunvastust alla suruda... ühesõnaga kõik meetodid, vahendid, sünteesitud ravimid
vms. mis võimaldavad organismi immuunsust hinnata, selle efektiivsust parandada,
vajadusel immuunvastust supresseerida, tugevdada antigeenivastased mehanisme jne.
Immunotehnoloogia võimaldab nö teha erinevaid antikehasid, milla kasutamine
meditsiinis on ilmselgelt olulisel kohal. Rekombinantsete, polüklonaalsete,
monoklonaalsete antikehade valmistamine ja kasutamine ravieesmärkidel on laialt
levinud, kuid siiski on tegemist valdkonnaga, mis vajab veel uurimist, kasvõi juba
seetõttu, et in vitro läbiviidud katsed antikehade võimest nõrgestada antigeene ei
tähenda seda, et need samad antikehad oleksid kompetentsed in vivo. Senised
laboratoorsed testid ja nende teadvustatud rakendamine ravis, profülaktikas on aga väga
efektiivsed.
Mõned näited monoklonaalsete antikehade kasutamise kohta kaasaegses meditsiinis:
Immunosupressiivsed monoklonaalsed antikehad
sihtmärk-valkude funktsiooni blokeerimine- kasvufaktorid, tsütokiinid (TNF-, IFN-,
IL-12, IL-2) ja nende retseptorid
pinnamarkerite, retseptorite vastu (CD4, CD52, CD11)
lümfotsüütide huku indutseerimine in vivo
autoimmuunhaiguste allasurumine (anti TNF-, anti-CD18)
tolerantsi taastamine autoantigeenidele (spetsiifilise vastuse tekke blokeerimine
autoantigeenideks)
Tuumorite immunoteraapia
tuumorspetsiifiliste antigeenide (CD20, CD52, HER-2, EGRF) vastased
monoklonaalsed antikehad, millele võib olla lisatud toksiini molekul
Transplantaadi äratõukereaktsiooni allasurumine
monoklonaalne anti-CD3 ja anti-CD4 antikehad
Kasutatakse transportmolekulina- antigeeni transport antigeeni esitlevatele rakkudele
(APC), T-rakkude aktivatsioon.
116. Voolutsütomeetrilise uuringu rakendamise põhimõtted kliinilises laboris.
Kvaliteedikontrolli põhimõtted.
Kliiniline voolutsütomeetria
Standardiseerimine
Kvaliteedikontroll 1.– sisemine 2.– välimine
Nõuded töövahenditele
Standardiseerimine
• Pre-analüütilised tingimused – nõutekohane proovimaterjali võtmine, transport,
säilitamine
• Analüütilised tingimused – materjali värvimine
• sobivad antikehad, kloonid, fluorokroomid – materjali lüüsimine
• kas enne või pärast – andmete kogumine ja analüüs
• Post-analüüstilised tingimused – vastuse väljastamine arstidele
Kvaliteedikontroll
• Sisemine kvaliteedikontroll – seadme töö tagamine
• regulaarne ennetav hooldus kvalifitseeritud tehniku poolt
• igapäevane seadistamine ja kontroll kasutaja poolt – tööprotsessi kvaliteedi tagamine
• bioloogiline kontrollmaterjal
• antikehade stabiilsus nii ajas kui partiide lõikes
• Väline kvaliteedikontroll – võrdlus teiste laboritega, aitab avastada “vigu”
Nõuded töövahenditele
• Eelistatult IVD-märgisega seaded (in vitro diagnostics)
• Eelistatult IVD-märgisega reagendid
• Antikehade “kokteilid”
• Lüofiliseeritud antikehad/“kokteilid“
• Eelistatult IVD-märgisega tarkvara
• lihtne ja automatiseeritud analüüs
• väheneb analüüsija subjektiivne faktor
117. Transplantoloogias kasutatavate laboratoorsete uuringute valik,
kvaliteedikontroll ja rakendamise põhimõtted.
Kasutatakse veregrupi AB0 määramist - doonoril ja retsipiendil peab olema sama või see,
mis sobib. Rh-faktor ei mängi rolli.
HLA test - kudede sarnasus peresiseselt võib olla väga suur, transplantaadil peavad
olema samad HLA antigeenid doonoriga.
Antikehade test - ega pole organismis antikehi, mis võiksid siirikut tappa
Veretestid - ülekantavate haiguste uurimiseks, nt HIV, CMV, hepatiit
Retsipiendi tausta uuring, haiguste ajalugu!
Võidakse teha testid, et uurida organi tööd, nt uriiniproov neerude töö kontrolliks.
Röntgen, et uurida kopse, südant, EKG ka.
Arteriogramm - süstitakse kontrastainet transplaneeritava organi veresoontesse, et
veenduda transplanteerimiseks sobilikkuses.
Naistele tehakse alati günekoloogiline läbivaatus.
Viimane veretest 48h enne, et veenduda antikehades, et pole selliseid, mis ründaksid
siirikut.
118. Vereülekandeeelsed seroloogilised testid, nende interpreteerimine ja
kvaliteedikontroll.
-
veretüübi kindlakstegemine koos antikehade testiga (crossmatch)
-
antikehade skriining, kas miski võib põhjustada soovimatut efekti pärast ülekannet
-
infektsiooniliste ainete test
Kohustuslikud testid enne vereülekannet:
-
AB0 ja Rh tüüp
-
Veregrupi antikehade skriining
-
Seroloogilised testid retroviiruste osas: HIV, HTLV (human T-lymphototicvirus)
-
Seroloogilised testid hepatiidi osas: B, C
-
Muud infektsioonilised ained: süüfilis, West Nile Virus
Ainult siis, kui kõik need markerid on negatiivsed, võib verd üle kanda, võib selle panka
saata. Kui miski positiivne teavitatakse doonorit. Võimalik, et ei tohi enam kunagi verd
annetada.
Näitavad in vitro erütrotsüüdi antigeeni-antikeha reaktsioone. reaktsiooni tulemus on
aglutineerumine
Lisaks uuritakse ka doonori meditsiinilist tausta, et olla kindlad, et veri on ohutu.
119. Autoantikehade määramiseks kasutatavad testid ja nende võrdlus (IIF,
ELISA, FEIA, immunoblot).
IIF - indirect immunofluorescence. Erinevad organid
Krüostaatlõigud (4-5 um) koeplokist
Inkubeerimine uuritava seerumiga (patsient)
pesu
Inkubeerimine
sekundaarse antikehaga, millel fluorestseeruv märgis (näiteks
inimese IgG vastased küüliku antikehad, konjugeeritud FITC-iga (fluorescein
isotiocyonate)
Pesu
Katteklaas
Reaktsiooni hindamine UV-mikroskoobis
Tundlikkus piisavalt suur kliiniliselt oluliste ak leidmiseks.
ELISA
ELISA (Enzyme Linked Immunosorbent Assay): ensüümseotud immuunsorbent-
meetod.
1. Antigeen (kaetakse ELISA plaadile)
2. Blokeerimine (veiseseerum, seaseerum, Tween)
3. Seerumilahjendus e. primaarne ak (Ak+ või Ak-)
4. Sekundaarne antikeha e. konjugaat ensüümmärgisega
5. Substraat (mida sekundaarse antikeha küljes olev ensüüm lõhustab nii, et tekib
värvuse muutus)
6. Optilise tiheduse mõõtmine (värvuse tugevuse erinevus!);
7. Kalibratsiooni kõvera tegemine ja vastuse tööühikute kalkuleerimine
Kliinilises praktikas baseerub ELISAl paljude antikehade ja haigustekitajate antigeenide
määramine. ELISA koosneb mitmest etapist:
ELISA plaadi (96-auguga mikrotiiterplaat) augud kaetakse antigeenilahusega,
toimub antigeeni valgu seondumine plastikpinna külge. Edasi välditakse
mittespetsiifiline seostumine, nt valgu kaseiini abil.
Kantakse peale analüüsitav seerumlahjendus (primaarne antikeha) ja
inkubeeritakse.
Inkubatsioon ensüümiga (AP- alkaalne fosfataas või HRP- peroksüdaas), mis on
konjugeerunud sekundaarse antikehaga.
Märgise hulga mõõtmine- ensüümaktiivsus määratakse kromogeense substraadi
abil. Substraadilahus muutub ensüümaktiivsuse toimel värviliseks, spetsiaalse
kolorimeetri (ELISA plaadi lugeja) abil mõõdetakse igas augus värvuse intensiivsus
(optiline tihedus).
ELISA sobib
suurte analüüside hulga skriinimiseks, tema analüütiline tundlikkus on
väiksem kui radioimmuunmeetodil, kasutatavad reaktiivid on aga stabiilsemad.
Kasutatakse ära ensüümi katalüüsivaid omadusi, neutraalse substraadi ensümaatilisel
töötlemisel tekib värviline produkt, mida saab detekteerida. Kasutatavad ensüümid pole
substraadi suhtes valivad.Tegemist on põhimõtteliselt sändvitšmeetodiga. Plaat võib
kaetud olla ühe antigeeni või mitme erineva antigeeniga. Antigeeni valgu seondumine
plaadiga on mittespetsiifiline, kuid piisavalt tugev, et komponendid ei lahkneks, neid
hoiavad koos van der Waalsi jõud.
FEIA
Immunoblot
Immunoblot-analüüs koosneb kolmest etapist. Esiteks toimub valkude elektroforees,
mille tulemusena lahutatakse antigeenid, kasutatakse SDS-PAGE. Toimub ca 1h. Teiseks
etapiks on blotting e lahutatud antigeenide ülekanne geelist (SDS-PAGE)
nitrotselluloosfiltrile. Kolmandaks viiakse läbi antigeen-antikeha reaktsioon. Selleks
blokeeritakse nitrotselluloos „neutraalsete“ valkudega (nt rasvatu piimapulber).
Primaarne antikeha lisatakse süsteemi, st seerumlahjendus. Seejärel lastakse sellel
seista, st inkubatsioon seerumlahjendusega. Pärast inkubatsiooniaega detekteeritakse
tekkinud antigeen-antikeha kompleksid. Sekundaarne antikeha on märgistatud kas
ensüümmärgistusega (peroksüdaas, alkaalne fosfataas), mille korral toimub
detekteerimine kromogeensuse või kemoluminestsentsi alusel, või radioaktiivse
isotoobiga, detekteeritakse autoradiograafia abil. Immunoblott-analüüsil on kõrge
spetsiifilisus, sest toimub antigeenide lahutamine elektroforeesiga. Kõrge tundlikkuse
annab antigeen-antikeha vaheline immunoloogiline reaktsioon. SDS denatureerib valgud,
st säilivad lineaarsed epitoobid, konformatsioonilised kaovad. Kliinilises diagnostikas on
immunobloti kasutamine piiratud, sest ta on aeganõudev ja raskelt automatiseeritav.
100% puhaste antigeenide kasutamine iseloomustab ELISA meetodit, immunoblott
võimaldab aga analüüsida nn musti antigeene. Immunoblotti kasutatakse
referentstestina mitmete nakkushaiguste serodiagnostikas (HIV, H. pylori, Borrelia
burgdorferii). Skriinimiseks immunoblott ei sobi. Veel kasutatakse seda meetodit
autoantikehade määramiseks (tuumaantikehade leiu diferentseerimiseks, ENA-
Extractable Nuclear Antigens- antikehad), teaduslikes eksperimentides haiguste
patogeneesimehhanismide selgitamisel, autoantigeenide kindlakstegemisel,
molekulaarbioloogias.
Rakuliinid
HEp-2 – kõri epiteeli kartsinoomi rakuliin
HEp-20-10 – nagu HEp2 aga rohkem mitootilisi rakke-
120. Autoantikehade kasutamine esmaseks sõeltestimiseks ja täpsustavaks
testimiseks autoimmuunhaiguste diagnostikas.
ANA IF on esimene test, mida tehakse, kui kahtlustatakse AIH. See on väga hea, kuna on
väga tundlik. Positiivne ANA IF võimaldab hakata uurima täpsemalt. Kasutatakse Hep-2
rakuliini ANA IF-s. Selle testiga saab uurida paljusid autoantikehi. Pärast ANA testi
tehakse veel ka tiitri uuring, kui kõrge tiiter on.
ANA IF pos, tiiter pos, hakatakse uurima põhjust. Alustatakse kõige tõenäolisemast ja
liigutakse kuni saadakse positiivne tulemus või kui kõik testid annavad negatiivse
tulemuse.
Arstidel on tabelid, mille kaudu saavad hakata teste määrama ja tulemusi hindama.
Tehakse veel ka reumatoidfaktori ja erütrotsüütide sadestamise teste.
Siiski tuleb teada, mida tehakse, mida uuritakse, muidu raisatakse kõigi raha ja aega,
samuti võib patsient saada vale diagnoosi või jääda diagnoosita.
121. Autoantikehade testide rahvusvaheline standardiseerimine. ICAP
soovitused ANA määramiseks.
ICAP - International Consensus on ANA patterns
ICAP kodulehel olemas tabel, kust saab vaadata, milline ANA muster vastab millisele
haigusele, mida mis järjekorras testida.
122. Viis olulisemat allergoloogias kasutatavat diagnostilist meetodit. Kuidas
neid kasutada?
Anamnees! Ülduuringud
Nahatestid, üld-IgE
Spetsiifline IgE/komponentanalüüs
Eliminatsioonitestid
Provokatsioonitestid
Astma korral kopsufunktsiooni uuringud
Ülitundlikkusreaktsioonid
IgE vahendatud (tüüp I) Kiire: minutid-tunnid
IgE nuumrakkude ja basofiilide pinnal
Astmahoog, anafülaksia, urtikaaria, äge riniit, konjunktiviit jt
•Tsütotoksiliste lümfotsüütide poolt (tüüp IV) vahendatud –Aeglane: tunnid-ööpäevad
–Krooniline astma ja riniit, kontaktdermatiit
•IgG (IgM) vahendatud (tüübid II-III) –eReaktsioon antigeenile rakkude pinnal või sees
(transfusioonireaktsioon, siiriku äraõtukereaktsioon jt)
–Autoimmuunhaigused
Nahatorketest
Kiire (15 min)
Suhteliselt ohutu
Odav
Vajab spetsiaalselt treenitud personali
Antigeenide lahused
Saab kasutada ka oletatavat allergeeni
Tervel nahal, antihistamiinikum segab!
Positiivne (histamiin) ja negatiivne (füs. Lahus) kontroll
Epikutaantest
Mõõdab rakulist vastust! (Allergiline kontaktdermatiit)
Standardseeria: 31 olme ja tööstuskemikaali
o Kokku 96h: 0h: paigaldatakse kannud kemikaalidega
o 48h: eemaldatakse, esmane hindamine
o 96h: lõplik hindamine
Üld IgE
Tõus viitab allergilistele haigustele
AGA KA:
Parasiidid (nahal, soolestikus jm)
Geneetilised immuunpuudulikkusega kulgevad haigused – Hüper-IgE sündroomid
(tuhanded kU/L)
Spetsiifilised IgE-d
Kliiniliselt samaväärne nahatorketestidega
Veeniverest (lastel raskem võtta)
Aeganõudvam
Kallim
Täpsem, mõõdetav tulemus (kvantitatiivne)
Suurem IgE repertuaar
Komponentanalüüs
Toiduainetes on erineva allergeensed valgud – “molekulaarsed komponendid”
Kogu allergeeni ekstrakti asemel uuritakse kindlatele valkudele spetsiifiliste IgE
olemasolu
Erinevad komponendid kutsuvad esile erineva tugevusega või kliinilise pildiga
reaktsioone
Analüüsitakse kas ImmunoCAP või immunoblot meetodil.
Immunoblot analüüs
Võimaldab analüüsida mitut allergeeni korraga
Automatiseeritav (ettevalmistusest kuni hindamiseni)
Masinaga lugemisel kvantitatiivne
Eliminatsioonitest
Juhul kui spetsiifiline IgE positiivne
Spetsiaalne dieet, kus potentsiaalne allergeen on välja lülitatud 1-6 nädalat
Histamiini sisaldavate ning histamiini vabastavate toiduainete vaene dieet.
Kakaotooted, Tsitruselised, Paprika, Maasikad, Kunstlikud värv- ja aroomiained, Mesi,
Viinamarjad, rosinad, vein
Provokatsioonitest
Juhul kui spetsiifiline IgE positiivne
Manustatakse allergeeni suu limaskestale või nahale väikeses koguses
Arstliku järelevalve all! Anafülaksia oht!
Nii avalik kui pimetest, platseebo kontrolliga
Võib järgneda eliminatsioonitestile.
Astma
NO väljahingatavas õhus - allergilise astma korral, eosinofiilide aktivatsiooni ja
allergilise põletiku indikaator
ECP (eosinofiilide tsütotoksiline valk, mõõdetakse vereseerumist, ) – vabaneb
eosinofiilide aktivatsioonil, hingamisteede põletiku aktiivsuse indikaator
Pulmonoloogilised (ekspiiriumi e väljahingamise takistus) FEV1 langenud
FVC langenud
Lisauuringud
Kliiniline veri Eosinofiilia (kroonilised) allergilised haigused
AGA KA:
Parasiidid
Addisoni tõbi
Ravimitest indutseeritud
Kasvajalised haigused
Teatud immuunpuudulikkussündroomid
Teatud autoimmuunhaigused
Trüptaas – anafülaksia indikaator, vabaneb nuumrakkude aktivatsioonil
123. Tavalised ja molekulaarsete komponentide põhised allergiatestid. Nende
võrdlus ja eelistused (põhjendused).
Tavalised: Nahatorketest(kiire), Epikutaantest, Üld IgE(näitab ainult antikehade hulga ja
midagi muud mitte, kõrgel IgE on ka teisi põhjusi peale allergiat), Spetsiifilised IgE-d
(Kliiniliselt samaväärne nahatorketestidega),
Komponendid: Komponentanalüüs otsitskse spets valkude vastu IgE nt valkud toidus,
Immunoblot analüüs: väga spetsiifiline uuritakse mitu antigeeni, eliminatsioonitest ja
provokatsioonitestid põhimõte, oletatav allergeen alguses ei tarvita ja kontroll tarvitakse
ja vaadatakse mis juhtub, ei sobi väga just komponentideuurimise hulka aga nt saab
teada saada et on sellise toudus on korrelatsioon et äki neil on ühine komponent mille
vastu allergis.
Õpikud: A. Abbas & A. Lichtman: Cellular and Molecular Immunology (soovitatav 2012 –
2017, 2021); Rakubioloogia. Toim. T. Maimets, S. Kuuse. Tartu 2018 (immunoloogia osa);
R.
Uibo, K. Kisand, P. Peterson, K. Reimand Immunoloogia, Tartu 2015.
Loengud ja seminarid (sh Journal Club materjal juhendajate poolt).
Muud
41.
Adenosiin signaalmolekulina.
Tema tase on tavaliselt väga madal, kuid kudede lagunemine ja hüpoksia põhjustavad
rakuvälise adenosiini kõrge taseme.
Kasvaja kudedes on adenosiini tase 10-20
korda kõrgem, kui ümbritsevates kudedes. Rakuväline adenosiin võib signaali läbi viia
nelja
G-valguga seotud retseptorite komplekti: A1, A2a, A2b ja A3.
A2a retseptor võib negatiivselt reguleerida ülereageerivaid immuunrakke, kaitstes
seeläbi kudesid põletikuliste kahjustuste eest.
A2a retseptorid on T-regulatoorsete rakkude, CTL-ide, dendriitrakkude, makrofaagide ja
NK-rakkude pinnal. A2aR vahendab intratsellulaarse cAMP kogunemist immuunrakkudes.
cAMP on tsükliline adenosiinmonofosfaat, mis on sekundaarne virgatsaine ning osaleb
rakusiseses närvisignaali ülekandes. cAMP-l on palju immunosupressiivseid toimeid:
suurenenud immuunosupressiivsete tsütokiinide tootmine (TGF-β , IL-10); alternatiivsete
immuunkontrollpunktide radade retseptorite ülesreguleerimine (PD-1 ja LAG-3);
suurendab FOXP3-e ekspressiooni CD4 T-rakkudes, et toota T-regulatoorseid rakke.
A2aR, the ligand of which is adenosine, inhibits T cell responses, in part by driving CD4+
T cells to express FOXP3 and hence to develop into TReg cells115
B7-H3 – kasvajaendoteelirakkudes
B7-H4 – tumor-associated makrofaagides
KIR- T-rakkude ja APC-de pinnal
LAG3 – suurendab Treg funktsiooni ja inhibeerib CD8+ effektor T-raku funktsiooni
TIM3 – galektiin (rinnavähk), inhibeerib Th1 funktsiooni
4?. Viruse kohta küsimus
Mittespetsiifilised reaktsioonid – interferooni tekkevõime on viirustega infitseeritud
rakkudel. Infitseeritud rakust vabanev INF toimib tema naabruses asuvatele rakkudele
(piisab üliväikesest kogusest). INF on suuteline mõjutama erinevaid viiruste elutsükli
etappe, nagu nt viiruste adhesiooni organismi rakkudele (viiruse adhesioon toimub
rakkude pinnal olevate retseptorite kaudu), viiruspartiklite desintegratsiooni,
transkriptsiooni, translatsiooni, valgusünteesi ja viiruspartiklite väljumist rakust. Lisaks
viiruste replikatsiooni otseselt pärssivale toimele omavad INF-id ka immunomoduleerivat
toimet. Nimelt INF-ide võime reguleerida MHC esinemist rakkude pinnal. Kõik
interferoonid on suutelised suurendama MHC I hulka, ka IFN-γ ja MHC II hulka. Lisaks
MHC molekulide mõjutamisele indutseerivad INF-id ka Fc ja tsütokiinide retseptoreid.
Kõik need protsessid tõstavad Ag äratundmise efektiivsust. Interferoonil on võime
suurendada efektoorsete makrofaagide, neutrofiilide, T-lümfotsüütide ja NK-rakkude
tsütotoksilisi omadusi. Lisaks tsütokiinidele pakuvad viirusinfektsiooni eest kaitset ka
rakulised reaktsioonid. Eeskätt seostatakse mittespetsiifilisi rakulisi reaktsioone NK-
rakkudega. Viimased Ühinevad infitseeritud rakuga, järgneb tsütotoksiliste molekulide
vabanemine ja sihtraku häving. NK-rakkude viirus hävitavat tomet suurendab INF-ide
vabanemine.
Spetsiifilised reaktsioonid – immuunvastus viiruslikule Ag-le on peaaegu 100% T-
rakkudest sõltuv, kuid see ei tähenda, et immuunvastus on ainult rakulist tüüpi.
Viirusinfektsiooni likvideerimiseks kasutab organism nii rakulisi kui ka humoraalseid
mehhanisme. B-lümfotsüütide BCR-retseptorid ja Ak-d ühinevad viiruspartiklil asetsevate
Ag-dega, kuna aga „töödeldud” lineaarsed epitoobid, esitletuna koos MHC molekulidega,
on äratuntavad T-lümfotsüütidele. B-lümfotsüütidel on viirusinfektsiooni korral kahene
roll. Lisaks otsesele viirust äratundvale funktsioonile ja Ak-de produktsioonile on nas
lisaks võimelised viirust internaliseerima, töötlema ja esitlema oma pinnal koos MHC II
molekuliga T4-lümfotsüütidele. Samaaegselt sekreteerivad antigeense stimulatsiooni
saanud T4-lümfotsüüdid B-lümfotsüüte aktiveerivaid tsütokiine.
Ak-d ei ole võimelised tungima infitseeritud rakku ning on seetõttu ebafektiivsed
latentsete ja otseselt rakust rakku levivate viirusinfektsioonide suhtes. Küll seostuvad
nad aga ekstratsellulaarsete viiruslike antigeenidega. Sellisteks Ag-deks võivad olla
organismi rakul paiknevad viirusvalgud ja vabalt vereringes tsirkuleerivad intaktsed
viirused. Ak seostumine viirusosakesega pärsib viiruse adhesiooni peremeesorganismi
rakulemembraanile ja edasist penetratsiooni. Veres ja kehavedelikes täidavad neid
ülesandeid IgG ja IgM, limaskestadel aga IgA. Isegi kui Ak-dega kaetud viirus suudab
tungida rakku, on sageli takistatud tema desintegratsioon ja nukleiinhappe vabanemine
kapsiidist. Ak-d moodustavad tsirkuleerivate viirustega komplekse, piirates nii viiruste
levikut. Kompleksid on paremini fagotsüteeritavad ning äratuntav komplemendile.
Komplement võib viiruse otseselt lüüsida, purustades viiruspartikli, aga ka opsoneerida.
Teatud viirused, nagu herpesviirus, ei allu Ak-de neutraliseerivale toimele, sest nad
levivad intratsellulaarselt närvikiude pidi (paikneb ganglion trigeminales`es). Ak-d on
võimetud ka viirupartikli antigeenide muutuste tõttu.
Humoraalsel immuunsusel on oluline osa reinfektsioonide vältimiseks nende haiguste
korral, mis nõuavad haiguse tekkkeks viiruste tsirkulatsiooni vereringe : poliomüeliit,
hepatiit A ja B, tuulerõuged, leetrid.
Viirustel esineb mitmeid mehhanisme, mille abil nad neutraliseerivad immuunsüsteemi
mõju: Ajutise immuundefitsiidi teke infektsiooniga mitteseotud antigeenide suhtes,
immuunvastust esilekutsuva viiruse suhtes reaktsioon tavaliselt ei nõrgene. Nii on teada,
et gripp, punetised, leetrid ja tsütomehaloviirusinfektsioonid loovad soodsa pinnase
bakteriaalsete ja teiste infektsioonide tekkeks.
Püsiva immuundefitsiidi teke infitseerivate agensite suhtes. Tuntuimaks näiteks on HIV-
viirus, mis muudab organismi vastuvõtlikuks suhteliselt ohutute mikroorganismide
suhtes (oportunistlikud haigusetekitajad).
Viiruste persisteerumine. See on omapärane dünaamilinse tasakaaluseisund ühelt poolt
viiruste proliferatsioonivõime ja teiselt poolt organismi immuunsüsteemi vahel. Teatud
viirushaiguste puhul jäävad viirused peale esmast ägedat perioodi organismi püsima,
persisteerides rakkudes. Sellistena käituvad herpesviiruste perekonda kuuluvad viirused,
tsütomegaloviirus, Epstein-Barri viirus. Peristeerivad viirused on enamuse inimese
eluajast immuunsüsteemi kontrolli all. Kuid kui toimub immuunsüsteemi pärssumine,
aktiveerub seni latentsena püsinud viirusinfektsioon.
B-rakkude puhul:
Eellas-B-rakud (pre-B cells), mis sisaldavad tüstoplasmaatilist IgM-i, ei arene
täiskasvanud B-rakkudeks, mis ekspresseeriksid rakupinna antikeha retseptoreid
(Brutoni tõbi). Selle tulemusena ole enam ringluses B-rakke ja antikehasid
(aγglobulineemia).
Täiskasvanud B-rakkude võimetus areneda plasmarakkudeks
T-rakkude puhul:
Tüümuse puudumine arenguhäirete tõttu
Adenosiini või guanosiini akumulatsioon defektse adenosiini või puriini nukleosiidi
fosforülaasi tõttu.
IgG alaklasside puudulikkus
Esinemissagedus kõigub suurtes piirides. Ühe või mitme IgG alaklassi taseme oluline
langus või puudumine, kusjuures samal ajal on IgG üldine tase normaalne või isegi
tõusnud. IgG alaklasside bioloogilised omadused ja immuunvastus antigeenidele on
samuti erinevad. Nende puudulikkus on tingitud raske ahela konstantse segmendi
geenide deletsioonist või isotüüpide ümberlülitumise defektist. IgG alaklasside
puudulikkus võib olla ka sekundaarse immuunpuudulikkuse ilming. Võib olla
asümptomaatiline, kuid enamasti sagedased infektsioonid. IgG alaklasside puudulikkus
võib olla ajutise iseloomuga, eriti väikelastel.
Selektiivne antikehade puudulikkus
Vaktsineerimisel (infektsioonijärgselt) ei teki antikehi polüsahhariidsete antigeenide
suhtes. Miks nii on, see on teadmata. Põhjustab peamiselt hingamiselundite
retsidiveeruvaid infektsioone ja sinusiite. Immunoloogilistel uuringutel leitakse IgG, IgA,
IgM-i normaalne tase, mõnel võib IgG2 tase olla langenud. Võib esineda ka primaarse
immuunpuudulikkuse teiste vormide korral (Wiskott-Aldrichi, DiGeorge’i sündroom) ja
sekundaarse immuunpuudulikkusena aspleenia esinemisel.
T-rakkude puudulikkusele lisandub enamasti ka antikehade puudulikkus, mistõttu on
vähe vorme, mille puhul esineb prevaleeruvalt T-rakkude defekt. Siia rühma kuuluvad
mitmesugused sündroomid, mille patogenees ja geneetilise defekti iseloom ei ole veel
selgitatud.
PS! Kombineeritud (B- ja T-rakuline) immuunpuudulikkus
Immuunsüsteemi elundid on puudulikult arenenud, tüümus puudub või on hüpoplaasia.
Tüümuses on peamiselt retikulaarrakud, esineb vaid üksikuid lümfotsüüte. Antigeenide
mõjul lümfoidne kude ei arene, lümfisõlmed ja kurgumandlid puuduvad. Haigustunnused
ilmnevad tavaliselt 3.-6. elukuul, imikul tekib püsiv lööve, mis väljendab
immuunreaktsiooni emalt ülekandunud lümfotsüütidele. Sageli esinevad soor suu
limaskestal, naha mäda- ja seennakkused, respiratoorsed ja seedetrakti infektsioonid.
Iseloomulik on diarröa.
Vereanalüüsid näitavad tavaliselt lümfopeeniat (< 1000 lümfotsüüti/mm3). T-
lümfotsüütide arv on väga väike, võime rakulisteks immuunreaktsioonideks puudub.
Reaktsioon mitogeenidele kas puudub täielikult või on tunduvalt langenud. B-
lümfotsüütide arv ja immuunglobuliinide tase on kombineeritud immuunpuudulikkuse
vormist olenevalt erinev. Oluline on varajane diagnostika ja ravi. Raske kombineeritud
immuunpuudulikkus
Jaotub mitmeks alavormiks, enamlevinud on X-liiteline immuunpuudulikkus (tsütokiinide
retseptorite ühise -ahela defekt). Üldse on põhjuseks vähemalt 10 erinevat geenidefekti.
Rakuline immuunsus on häiritud, mis põhjustab kandidoosi, püsivat diarröad,
arengupeetust. B-lümfotsüütide arv on normaalne või isegi suurenenud, kuid B-rakud ei
küpse ega funktsioneeri normaalselt. Seetõttu on immuunglobuliinide tase enamasti
madal ja spetsiifili antikehi ei moodustu. NK rakkude arv on normaalne või suurenenud.
HIV variandid – tropism kemokiini retseptori järgi
Makrofaagi-troopne viirus – iseloomulik varajasele nakatumisele limaskestade kaudu
o infitseerib aktiivseid T rakke (sh mälurakke), dendriidirakke, makrofaage
o koretseptoriks on CCR5
Lümfotsüüdi-troopne viirus
o vajab kinnitumiseks kõrget CD4 tihedust
o infitseerib naiivseid T rakke
o koretseptoriks on CXCR4
Geneetiline defekt CCR5 kemokiini retseptoris võib tuua kaasa resistentsuse HIV
infektsioonile
On näidatud, et:
makrofaagi-troopne HIV ei suuda defektse CCR5 puhul rakku tungida
kemokiinide (RANTES, MIP-1) kõrgem kontsentratsioon geendefektiga isikutel
konkureerib viirusega CCR5 pärast
mutantse CCR5 homosügoodid ei infitseeru makrofaagitroopse viirusega,
heterosügootidel on infektsiooni progresseerumine aeglasem
Euroopas CCR5 geeni defekt ~10 % rahvastikust, ~1 % neist homosügoodid
Veidi täpsemalt HIV:
HIV kinnitumine ja rakku tungimine toimub gp120 ja gp41 abil ning märklaudraku
pinnal asuvate CD4 ja kemokiini retseptorite kaudu. HIV infitseerib T-rakke,
makrofaage ja dendriitrakke.
Viiruse partikkel seostub CD4 ja koretseptoriga T-rakul
Viiruse ümbris sulandub rakumembraaniga, lastes viiruse genoomil rakku siseneda
Pöördtranskriptsioon kopeerib viiruse RNA genoomi topeltahelaliseks cDNA-ks
Viiruse cDNA siseneb raku tuuma ja integreerub peremeesraku DNA-ga
T-raku aktivatsioon indutseerib madalatasemelist proviiruse transkriptsiooni
RNA transkriptid paljundatakse ja seotakse, saab toimuda varajaste geenide tat ja rev
translatsioon
Tat amplifitseerib viiruse RNA sünteesi, rev indutseerib üksikult seotud või
mitteseotud viiruse RNA transporti tsütoplasmasse
Hilised valgud Gag, Pol, ja Env transleeritakse ja viiakse viiruse partiklitesse, mis
rakust välja punduvad
Immuunsüsteem kontrollib HIV hulka, kuid ei suuda viirust lõplikult elimineerida.
Document Outline
- Kordamisküsimused
- 1. Nüüdisaegse immunoloogia ja rakendusliku (sh. kliinilise) immunoloogia arengu põhijooned. Immunoloogia areng Eestis.
- 2. Organismi kaitsebarjäärid (füüsikalised, keemilised, immunoloogilised).
- 3. Immuunsüsteemi ehituslike komponentide iseärasused erinevates kudedes. Primaarsete ja sekundaarsete immuunorganite tähendus ja seosed immuunsüsteemi funktsioneerimises.
- 4. Limaskestade ja nahaga assotsieeruv lümfoidne süsteem, ehitus ja roll immuunsüsteemis.
- 5. Loomulikku immuunsust tagavad rakud ja humoraalsed faktorid. „Treenitud immuunsus“.
- 6. Loomuliku immuunsuse tsütokiinide toimerajad.
- 7. TLR keskne tähendus. NLR ja RLR. Makrofaagide ja dendriitrakkude poolt produtseeritavad tsütokiinid. Nende tähendus immuunsüsteemi funktsioneerimisel..
- 8. Loomuliku immuunsuse lümfoidse rea rakud (ILC), NK-rakud ja nende pinnaretseptorid –iseloomustus ja funktsioon.
- 9. Komplementsüsteem. Tähendus, tähtsamad molekulid ja retseptorid.
- 10. Komplementsüsteemi kolm aktivatsioonirada ja nende tähendus.
- 11. Autoinflammatoorsed protsessid.
- 12. Loomuliku (sünnipärase) ja omandatud (adaptiivse) immuunsuse põhilised erinevused, nende seosed.
- 13. Valkude immunogeensust määravad tegurid. Antigeenid immuunsüsteemi aktivaatoritena. Antigeenide epitoobid.
- 14. Immunoglobuliinisubühiku ehitus. Fab ja Fc fragmendi immunoloogiline tähendus. Immunoglobuliinide klassid ja alaklassid. Nende strukturaalsed erinevused.
- 15. Immunoglobuliinide erinevate klasside bioloogiline tähendus. Primaarse ja sekundaarse immuunvastuse iseloomustus. Primaarses ja sekundaarses immuunvastuses tekkivate antikehade iseloomustus. Monokloonsed antikehad.
- 16. Sekretoorse IgA tekkemehhanism. Sekretoorse IgA iseloomustus ja bioloogiline tähtsus.
- 17. Immunoglobuliinide geenide komponendid ja rekombinatsioonimehhanismid. Immunoglobuliinide spetsiifilisuse geneetiline alus. Immunoglobuliinide isotüüpide kujunemine seoses B-rakkude küpsemisega. B-raku retseptori (BCR) iseloomustus.
- 18. T-raku retseptori (TCR) geenide komponendid ja rekombinatsiooni-mehhanismid. TCR spetsiifilisuse geneetiline alus
- 19. T-raku retseptori (TCR) ehitus ning aktivatsioonisignaalide transduktsioon T-rakus. TCRα/β ja TCRγ/δ retseptorid. Immunoloogiline sünaps. Kahe signaali teooria.
- 20. Peamise koesobivuskompleksi (MHC) iseloomustus. Tähendus immuunsüsteemi funktsioneerimisel. HLA I ja II klassi antigeenid. HLA alleelide pärilikkus ja assotsieerumine immunoloogiliste haigustega. MHC restriktsioon T-helper ja T-tsütotoksiliste rakkude suhtes.
- 21. Antigeeni esitavate rakkude ja T-rakkude vahelised signaalmolekulid. “Kahe signaali teooria”. Immunoloogiline sünaps. eespool
- 22. Tüümuse ehitus. T-rakkude küpsemise põhimõtted ja tüümuse roll selles.
- 23. Tüümuse roll organismi enese kudede vastase tolerantsuse kujunemisel. AIRE ja tema tähendus.
- 24. Immunoloogiline tolerantsus (tsentraalne ja perifeerne), selle kujunemise mehhanismid.
- 25. Immuunreaktsioonides osalevate rakkude tsirkulatsiooni tagavad mehhanismid.
- 26. Omandatud immuunsuse iseloomujooned (spetsiifilisus, “i
- 27. Omandatud immuunsuse efektormehhanismid (mikroorganismi ja antigeeni kahjutuks tegemise ja elimineerimise võimalikud mehhanismid).
- 28. Tsütokiinide iseloomustus ja toime põhimõtted. Tsütokiinide tähendus loomuliku ja adaptiivse immuunsuse reaktsioonides, lümfotsüütide kasvu ning diferentseerumise kontrollis.
- 29. Tsütokiinide määramise põhilised in vitro meetodid.
- 30. Interferoonide (I, II, III klass) iseloomustus ja nende tähendus erinevate immunopatoloogiliste protsesside korral.
- 31. Vereloome kasvufaktorid, hematopoeesi stimuleerivad tsütokiinid ja nende tähendus lümfotsüütide arengus.
- 32. Tsütokiinide retseptorid ja signaalrajad.
- 33. Tähtsamate loomulikku immuunsust mõjutavate tsütokiinide toimerajad.
- 34. Tähtsamate omandatud immuunsust mõjutavad tsütokiinide toimerajad.
- 35. Kemokiinide ja nende retseptorite üldine iseloomustus, tähendus rakuliste immuunreaktsioonide kujunemises.
- 36. Põletikureaktsioone stimuleerivad ja pidurdavad tsütokiinid.
- 37. Makrofaagide iseloomustus.
- 38. Dendriitrakkude iseloomustus. Dendriitrakkude roll adaptiivsetes immuunreaktsioonides. Antigeeni töötlus antigeeni esitavas rakus.
- 39. T-, B- ja NK-rakkude teke ning võrdlev iseloomustus. T-helperid (Th) ja Ttsütotoksilised (CTL, Tc) rakud. Pinnamarkerid. NKT-rakud. Mälurakud.
- 40. I ja II tüüpi T-helperite (Th1 ja Th2) ja Th17 rakkude iseloomustus ja tähendus immuunvastuses. Antigeenide, dendriitrakkude ja tsütokiinide roll T-rakkude polariseerumisel.
- 41. Immuunvastuse regulatsioonimehhanismid. Regulatoorsed T-rakud. Foxp3. CTLA-4 ja teised regulatoorse funktsiooniga molekulid.
- 42. Loommudelid immuunsüsteemi uuringus. Võimalused ja piirangud immuunsüsteemi funktsiooni hindamisel.
- 43. Tähtsamad loommudelid immuunsüsteemi funktsiooni hindamisel.
- 44. Autoimmuunhaiguste loommudelid (sh AIRE-/-, NOD, BB, d3T jt).
- 45. Viirusinfektsioonide immunoloogia.
- 46. Seeninfektsioonide immunoloogia.
- 47. Parasiitinfektsioonide immunoloogia.
- 48. Rakuväliste bakteriaalsete infektsioonide immunoloogia.
- 49. Rakusiseste – intratsellulaarsete – bakteriaalsete infektsioonide immunoloogia.
- 50. Bakterite ja viiruste mehhanismid, mis aitavad vältida immuunreaktsioone.
- 51. Infektsioonhaiguste korral esinevate immuunreaktsioonide diagnostika.
- 52. Kasvajate immunoloogia üldised põhimõtted.
- 53. Immuunreaktsioonide vältimine kasvajarakkude poolt. Kasvajate hoidumine immuunsüsteemi rünnaku eest
- 54. Kasvaja-antigeenid.
- 55. Kasvajate immunoloogilise ravi võimalused. – immunoteraapia
- 56. Immuunsüsteemi kontroll-punktide kaudne vähiravi. CAR-T eespool
- 57. Transplantatsiooniimmunoloogia üldised põhimõtted.
- 58. Hüperakuutsed ja akuutsed äratõukereaktsioonid. Krooniline äratõukereaktsioon
- 59. Kroonilised äratõukereaktsioonid
- 60. Äratõukereaktsioonide ennetamise võimalused.
- 61. Äratõukereaktsioonide immunoloogiline diagnosika.
- 62. Hematopoeetiliste tüvirakkude siirdamine ja äratõukereaktsioonide iseärasused. =luuüdi siirdamine
- 63. Vereülekande immunoloogia.
- 64. Immunotehnoloogia võimalused
- 65. Immunotehnoloogilised võtted kliinikumis
- 66. Immuunpuudulikkuse sündroomide klassifikatsioon. Tekkemehhanismid.
- 67. Omandatud immuunpuudulikkuse sündroom - AIDS. AIDSi korral tekkivad immunoloogilised muutused, nende dünaamika. AIDSi immunodiagnostika.
- HIV nakatunud inimestel esineb 4 perioodi haiguse kulus:
- Mõningaid HIV infektsiooni iseloomujooni:
- 68. Teistest viirusinfektsioonidest tingitud immuunpuudulikkused (COVID-19 jt.).
- 69. Komplementsüsteemi ja fagotsütaarse funktsiooni puudulikkuse sündroomide iseloomustus ja diagnostika.
- 70. Antikehade puudulikkuse sündroomide iseloomustus ja diagnostika.
- X-liiteline agammaglobulineemia – Brutoni tõbi (XLA)
- Selektiivne IgA puudulikkus
- Üldine variaabel immuunpuudulikkus (CVID)
- Veel
- 71. Kombineeritud immuunpuudulikkus. SCID.
- 72. T-rakkude aktivatsiooni ja funktsiooni puudulikkus.
- DiGeorge sündroom
- APS1 – APECED
- MHC I ja II klassi defitsiidid
- SCID
- 73. Immuunsüsteemi funktsiooni iseärasused lapseeas. Füsioloogiline hüpo-γ-globulineemia.
- 74. Rinnapiima immunoaktiivsed komponendid
- 75. Immunoloogilised muutused seoses vananemisega. Vanemaealiste immuunsüsteemi iseärasused.
- 76. Immunoloogilised erinevused meeste ja naiste vahel.
- 77. Toitumise ja toidu iseärasuste mõju immuunsüsteemi funktsioonile. Rasvumine ja immuunsüsteem.
- 78. Immunotoksilised ühendid ja nende mõju iseärasused. Radiatsiooni mõju immuunsüsteemile.
- 79. Närvisüsteemi ja immuunsüsteemi vahelised seosed. Stressi mõju immuunsüsteemi funktsioonile.
- 80. Füüsilise koormuse mõju immuunsüsteemile. Endokriinsüsteemi interaktsioonid immuunsüsteemiga.
- 81. Naise organismis raseduse ajal toimuvad immunoloogilised muutused. Spetsiifilised immunoloogilised muutused, mis kaitsevat loodet ema immuunsüsteemi kahjuliku toime eest.
- 82. Loote immunoloogilised mehhanismid. Platsenta immunoloogiline tähendus.
- 83. Loomulik (kommensaalne) mikrofloora ja selle tähendus immuunsüsteemi arengus.
- 84. Koekahjustuse mehhanismide (ülitundlikkuse) neli tüüpi (Coombs-Gelli j). Antikehade ja T-rakkude koekahjustuse efektormehhanismid.
- 85. Allergilise e anafülaktilise (I tüüpi) koekahjustuse (ülitundlikkuse) tekkemehhanism. Osalevad rakud, rakumembraanil paiknevad retseptorid, rakkudevahelised mediaatorid ja antikehad. Tsütokiinide tähtsus. Nuumrakkude aktiveerumisel vabanevad mediaatorid ja nende bioloogiline toime. Nuumrakkude aktiveerumisel sünteesitavad mediaatorid ja nende bioloogiline toime.
- 86. Antikehade poolt vahendatud (II tüüpi) koekahjustuse mehhanism.
- 87. Immuunkomplekside poolt vahendatud (III tüüpi) koekahjustuse tekkemehhanism. Immuunkomplekside tekkemehhanismid ja iseloom sõltuvalt antigeeni ja antikeha hulgast. Meditsiinilisi näiteid immuunkompleksidest vahendatud reaktsioonidest. Immuunkompleksidest tekkinud neerukahjustuse mehhanism.
- 88. T-rakkude poolt vahendatud (IV tüüpi) koekahjustuse mehhanism.
- 89. Autoimmuunsuse tekkemehhanismid, antigeenide ja immuunregulatsiooni tähendus. Välisfaktorite ja pärilikkuse roll autoimmuunsuse tekkes. Oluliseimad autoimmuunhaiguste tekkega assotsieeruvad HLA alleelid. Infektsioonid ja autoimmuunsus.
- 90. Organ-spetsiifiliste autoimmuunhaiguste iseloomustus. Nende immunopatogeneesi mehhanismid ja immunodiagnostika. Immunogeneetilised aspektid.
- Autoimmuune türeoidiit (Hashimoto türeoidiit) e Hashimoto türeoidiit – kilpnäärme lümfotsütaarne infiltratsioon (B, CD4+)
- T1d – I tüüpi diabeet
- Tsöliaakia
- 91. Organ-spetsiifilikata autoimmuunhaiguste iseloomustus. Nende immunopatogeneesi mehhanismid ja immunodiagnostika.
- Süsteemne erütematoosne luupus (SLE)
- 92. Autoantikehade testide kliiniline tähendus diagnostikas, haiguse prognoosis, ravi hindamisel jm. Tähtsamad kliinilises praksises kasutatavad autoantikehad.
- 93. Geneetiliste ja välisfaktorite osa allergia tekkes. Tähtsamate allergiaga seotud geenide iseloomustus. Välisfaktorid, mis kutsuvad esile Th2 rakkude aktivatsiooni. Hügieenihüpotees.
- 94. IgE iseloomustus ja bioloogiline tähendus. IgE tekkemehhanism atoopia korral. IgE retseptorite iseloomustus. IgE-retseptorite aktivatsioonist tulenevad rakusisesed muutused. IgG4 ja allergia.
- 95. Allergia in vivo ja in vitro testide üldine iseloomustus. Üld-IgE ja spetsiifilise IgE määramise kliiniline tähendus. Rekombinantsed allergeenid - uued võimalused täpsemaks diagnostikaks ja raviks.
- 96. Üldpõhimõtted immunoloogia laboratooriumi töö korralduses.
- 97. Kvaliteedikontrolli üldised põhimõtted. Referentsväärtused ja nende saamise tingimused. Uurimistöö ja rahvusvaheline koostöö olulisus.
- 98. Faktorid, mis mõjutavad immunoloogiliste uuringute tulemusi laboris.
- 99. Teadusuuringute tähendus igapäevases laboritöös.
- 100. Immunopretsipitatsioonimeetod antikehade määramiseks.
- 101. In vitro testid lümfotsüütide funktsiooni hindamiseks.
- 102. In vitro testid rakkude tsütotoksilise funktsiooni hindamiseks.
- 103. Lümfotsüütide uuringute kliiniline kasutamine.
- 104. Lümfotsüütide intratsellulaarsete ja pinnamarkerite määramine. Tähendus.
- 105. Voolutsütomeetria (FACS) põhimõte ja kasutamine.
- 106. In vitro testide põhimõtted antikehade ja antigeenide määramiseks. Avidiin-biotiinkompleksi tekkel põhinevate meetodite rakendus.
- 107. Rekombinantsed antikehad, üheahelalised antikehad (scFv).
- 108. Antikehade rakendusi kromatograafias.
- 109. Antikehade iseloomustamine biosensorite abil.
- 110. Antikeha-kiibid.
- 111. Antikehade rakendusi proteoomikas. HUPO.
- 112. Autoantikehad töövahenditena (cDNA raamatukogude skriining jms.).
- 113. Immunofluorestsents- ja immunoensüüm meetodid immunohistokeemias.
- 114. Immunofluorestsents- ja immunoensüüm meetodid immunohistokeemias.
- 115. Uued võimalused immunotehnoloogilisteks arendusteks.
- 116. Voolutsütomeetrilise uuringu rakendamise põhimõtted kliinilises laboris. Kvaliteedikontrolli põhimõtted.
- 117. Transplantoloogias kasutatavate laboratoorsete uuringute valik, kvaliteedikontroll ja rakendamise põhimõtted.
- 118. Vereülekandeeelsed seroloogilised testid, nende interpreteerimine ja kvaliteedikontroll.
- 119. Autoantikehade määramiseks kasutatavad testid ja nende võrdlus (IIF, ELISA, FEIA, immunoblot).
- 120. Autoantikehade kasutamine esmaseks sõeltestimiseks ja täpsustavaks testimiseks autoimmuunhaiguste diagnostikas.
- 121. Autoantikehade testide rahvusvaheline standardiseerimine. ICAP soovitused ANA määramiseks.
- 122. Viis olulisemat allergoloogias kasutatavat diagnostilist meetodit. Kuidas neid kasutada?
- 123. Tavalised ja molekulaarsete komponentide põhised allergiatestid. Nende võrdlus ja eelistused (põhjendused).
- Muud
- IgG alaklasside puudulikkus
- Selektiivne antikehade puudulikkus
- PS! Kombineeritud (B- ja T-rakuline) immuunpuudulikkus
- Veidi täpsemalt HIV:












































































































































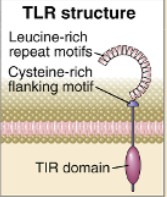
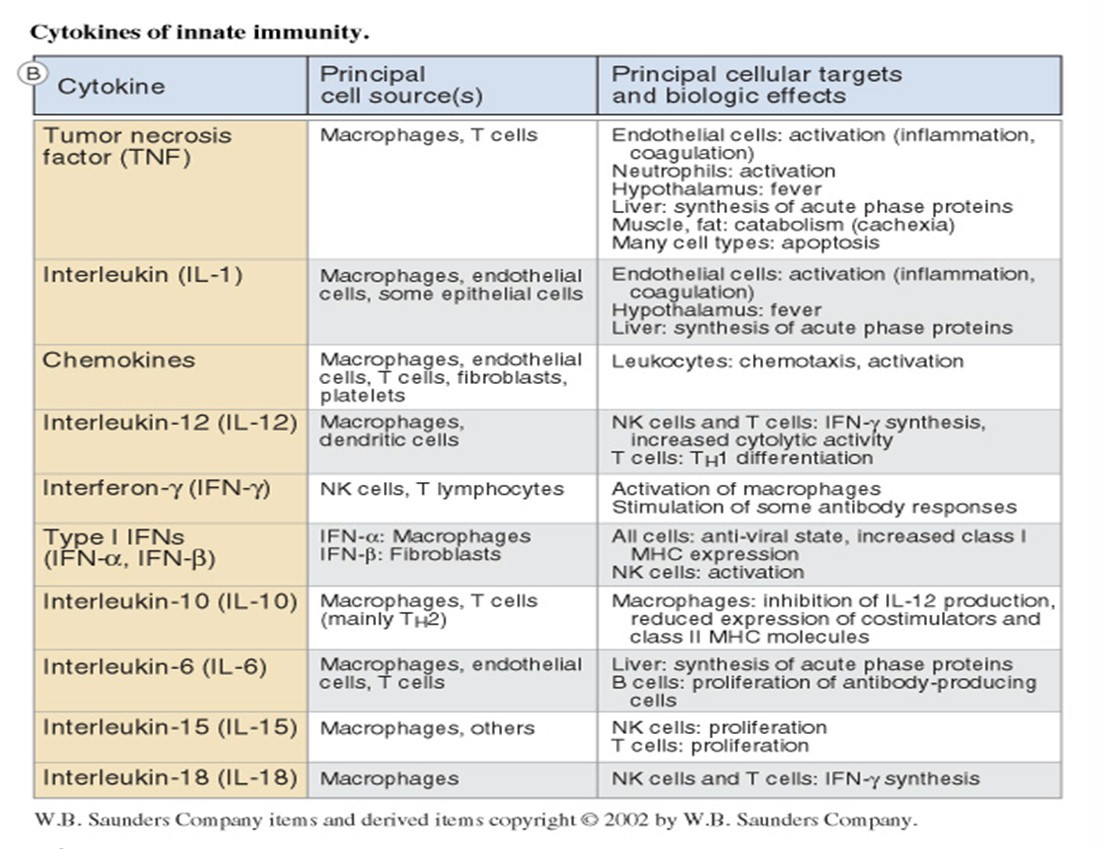
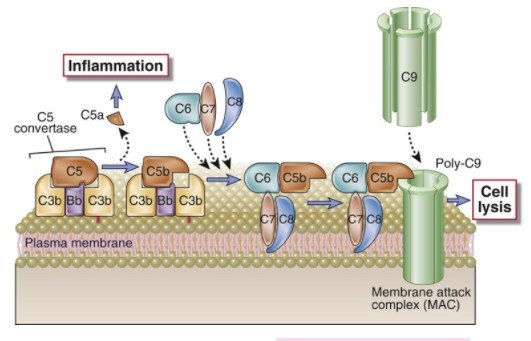
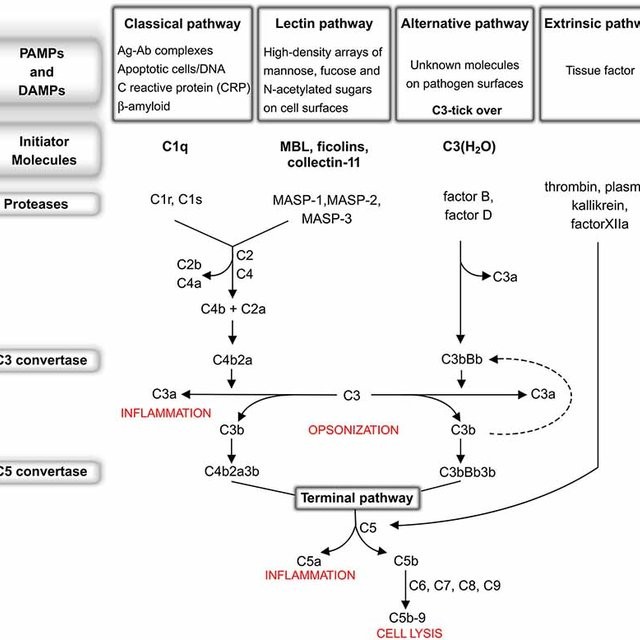
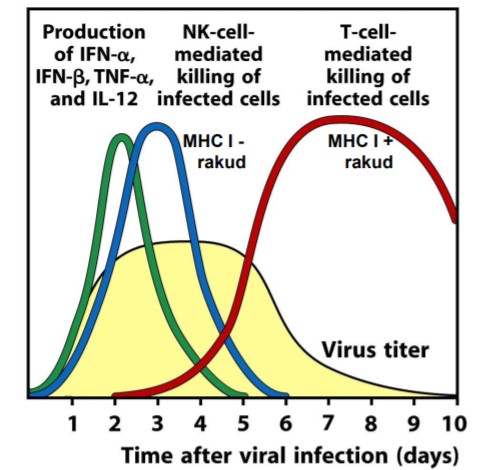
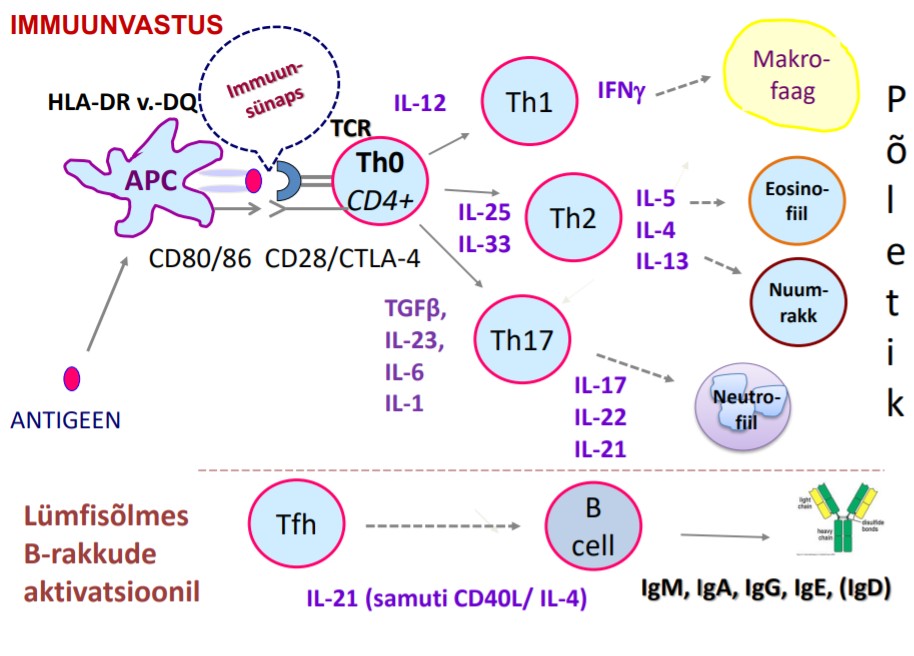
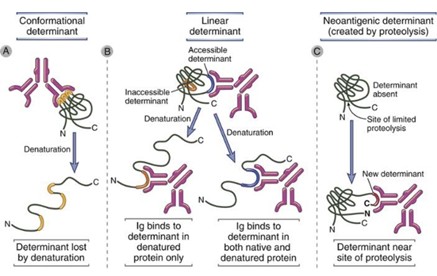
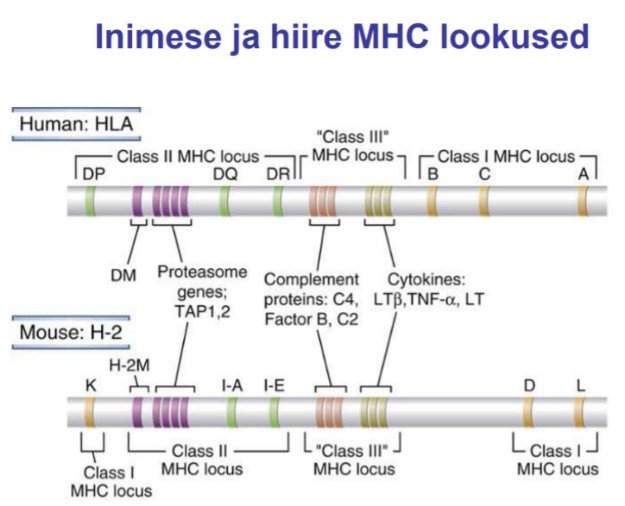
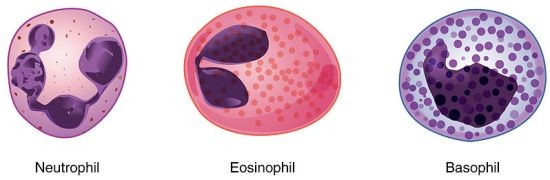
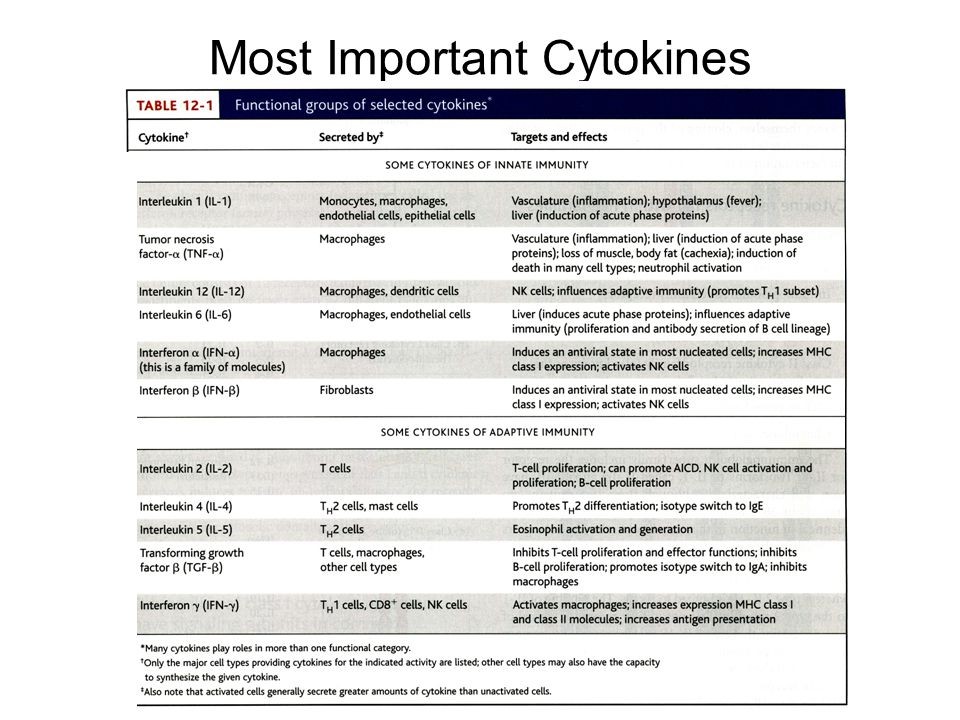

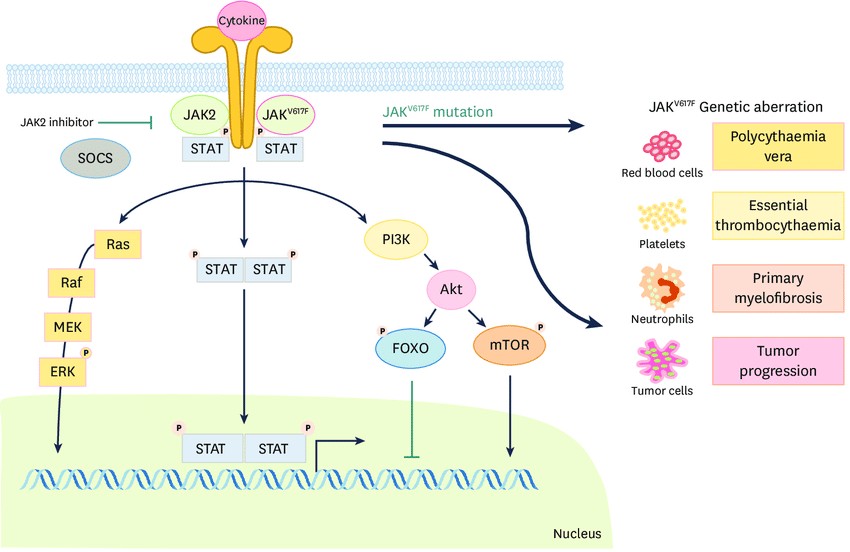
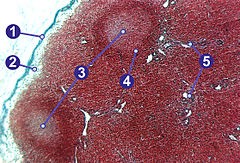
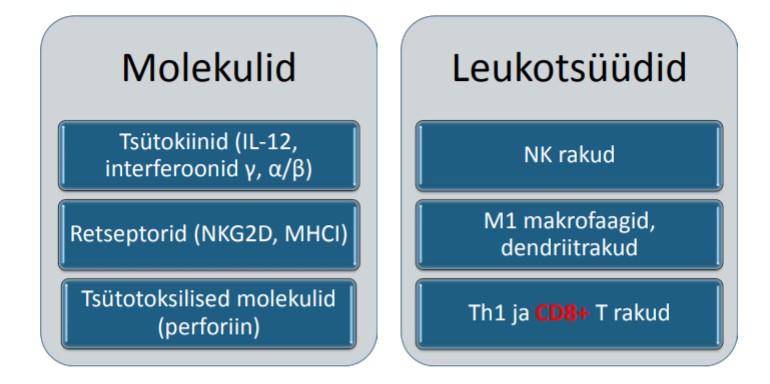



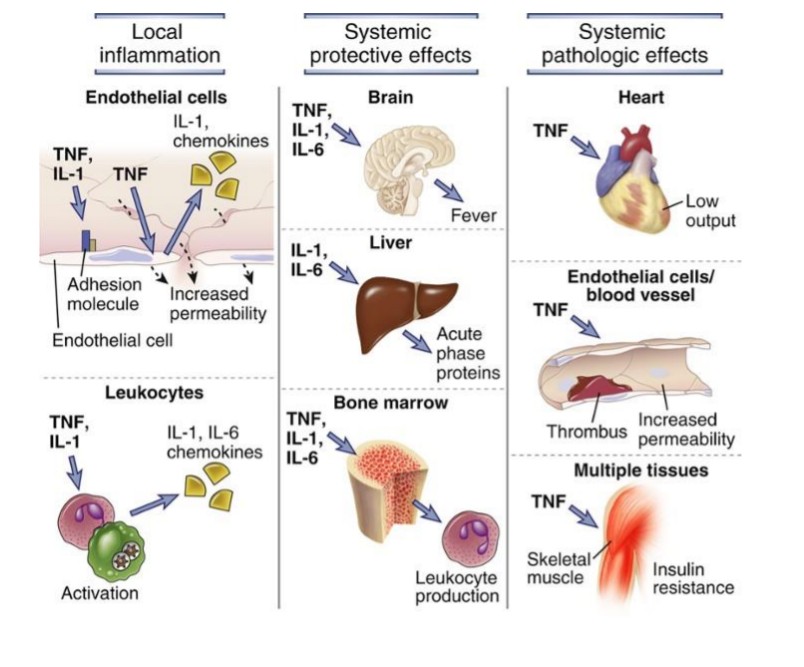
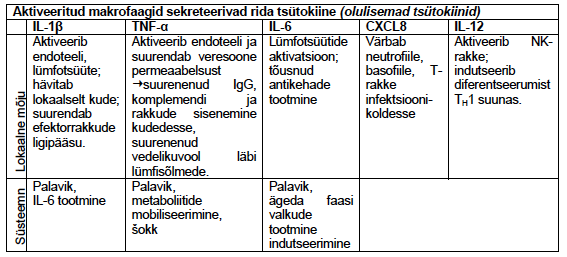
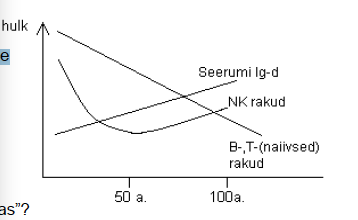









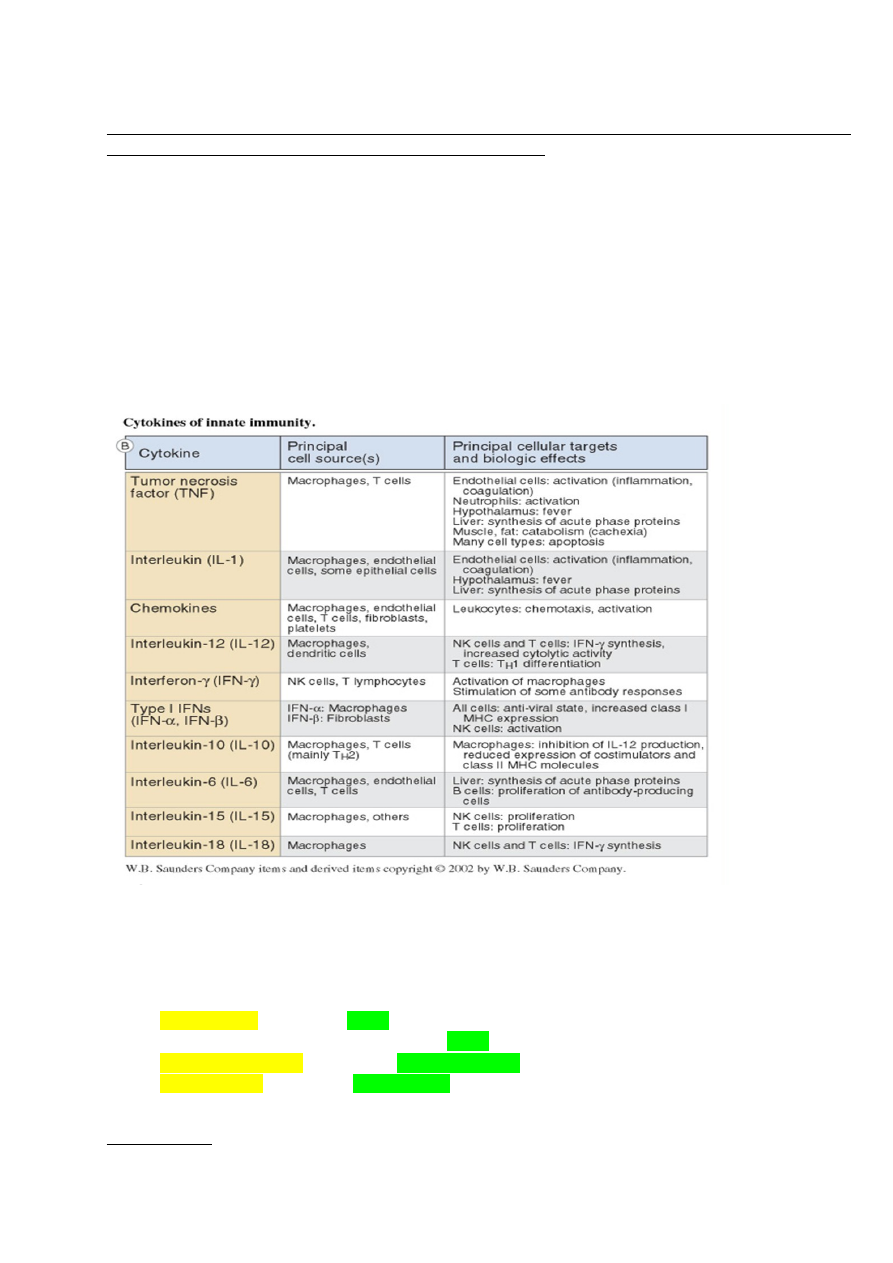


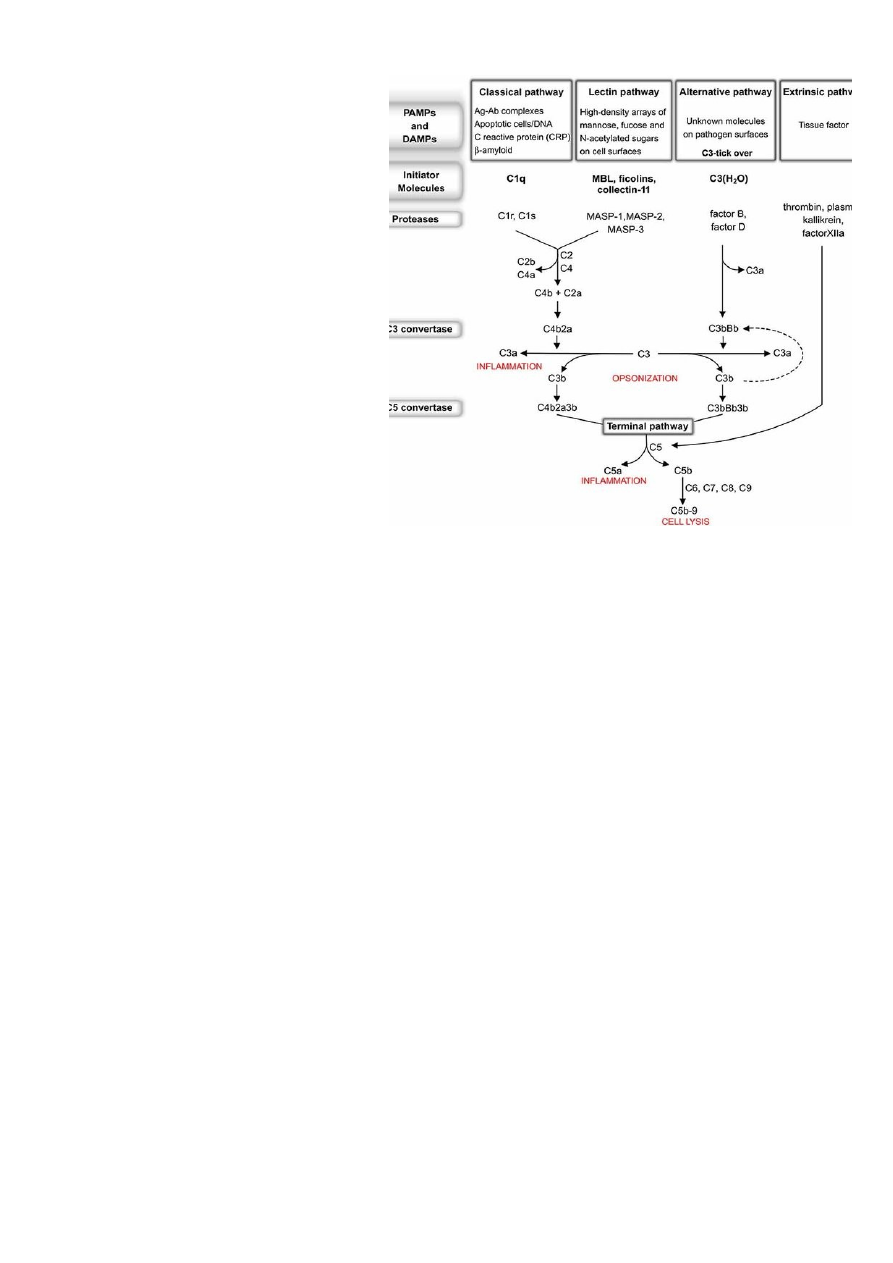
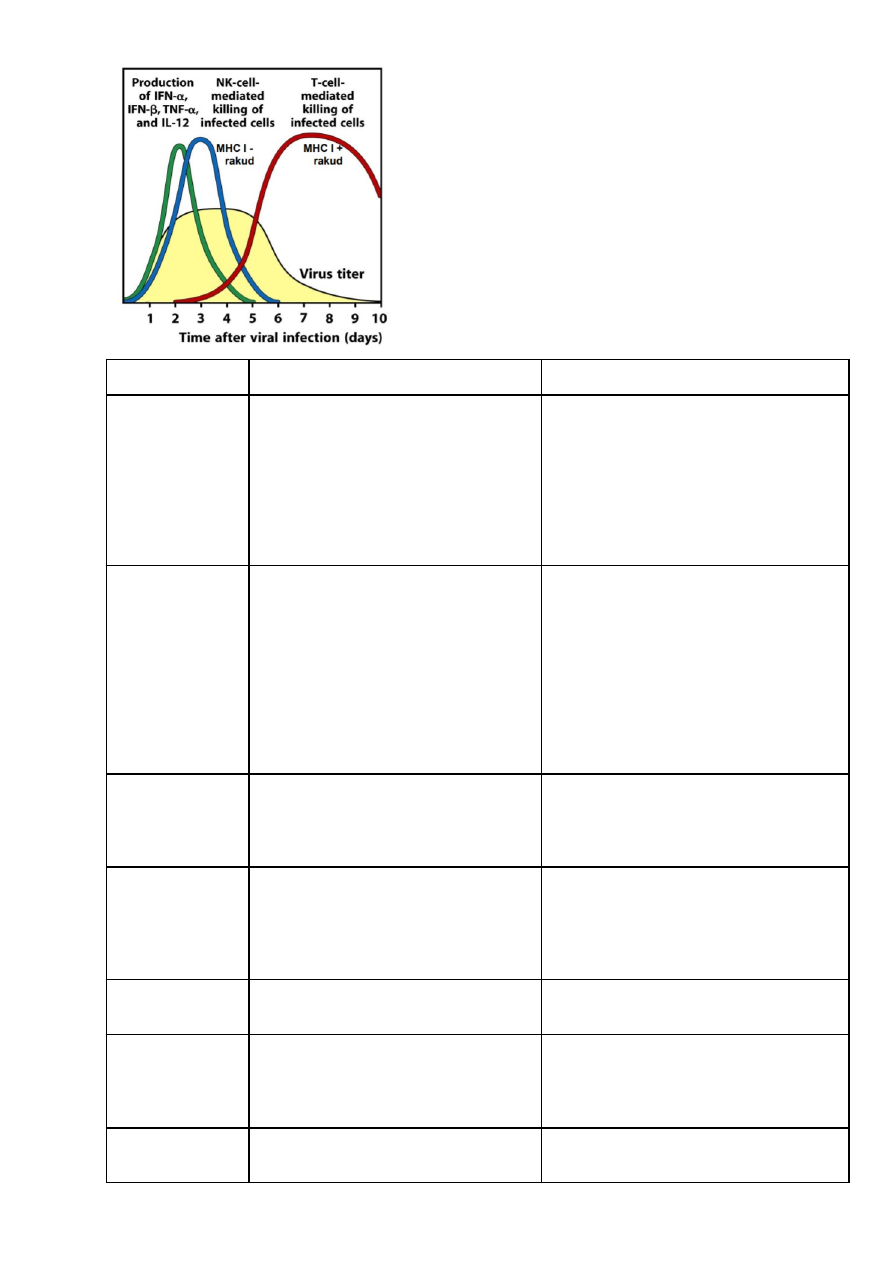
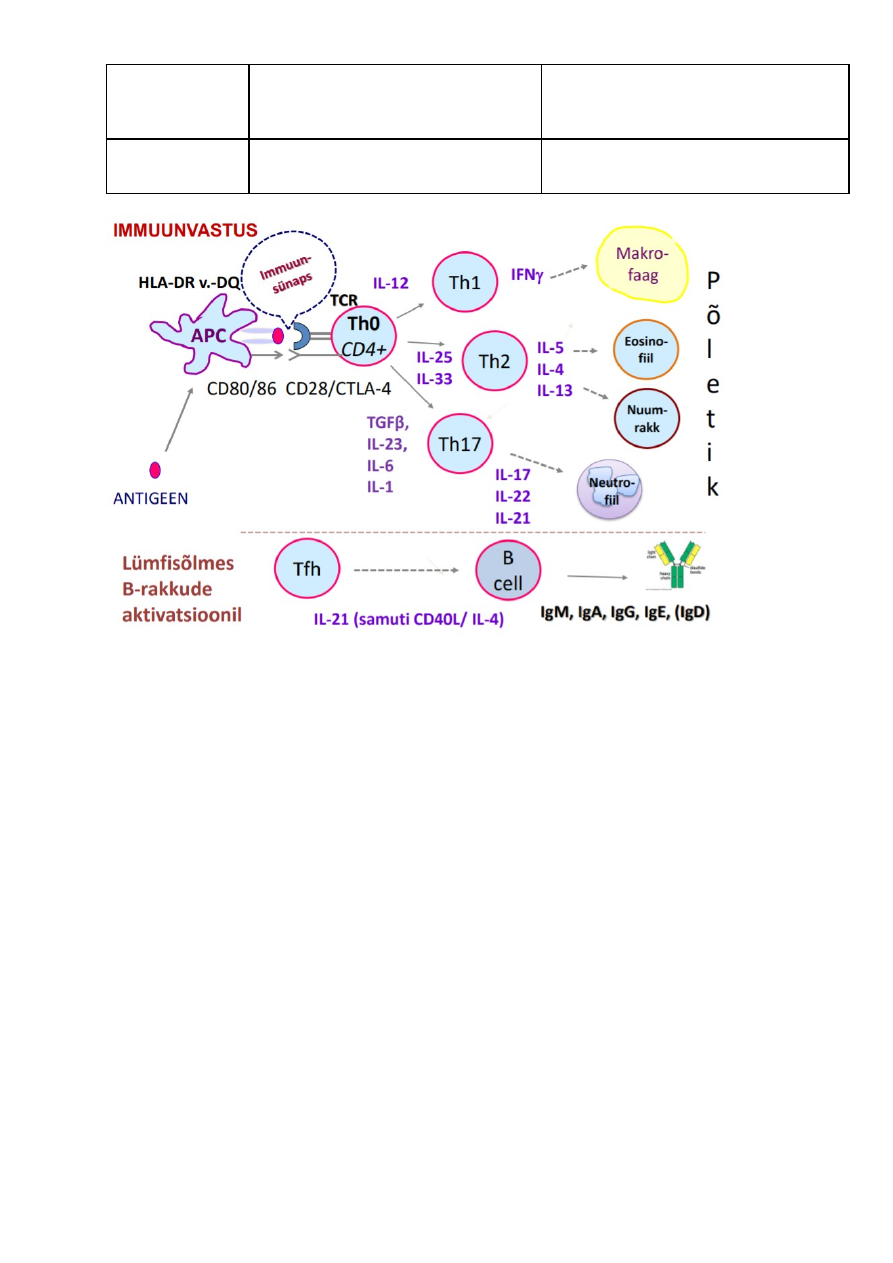
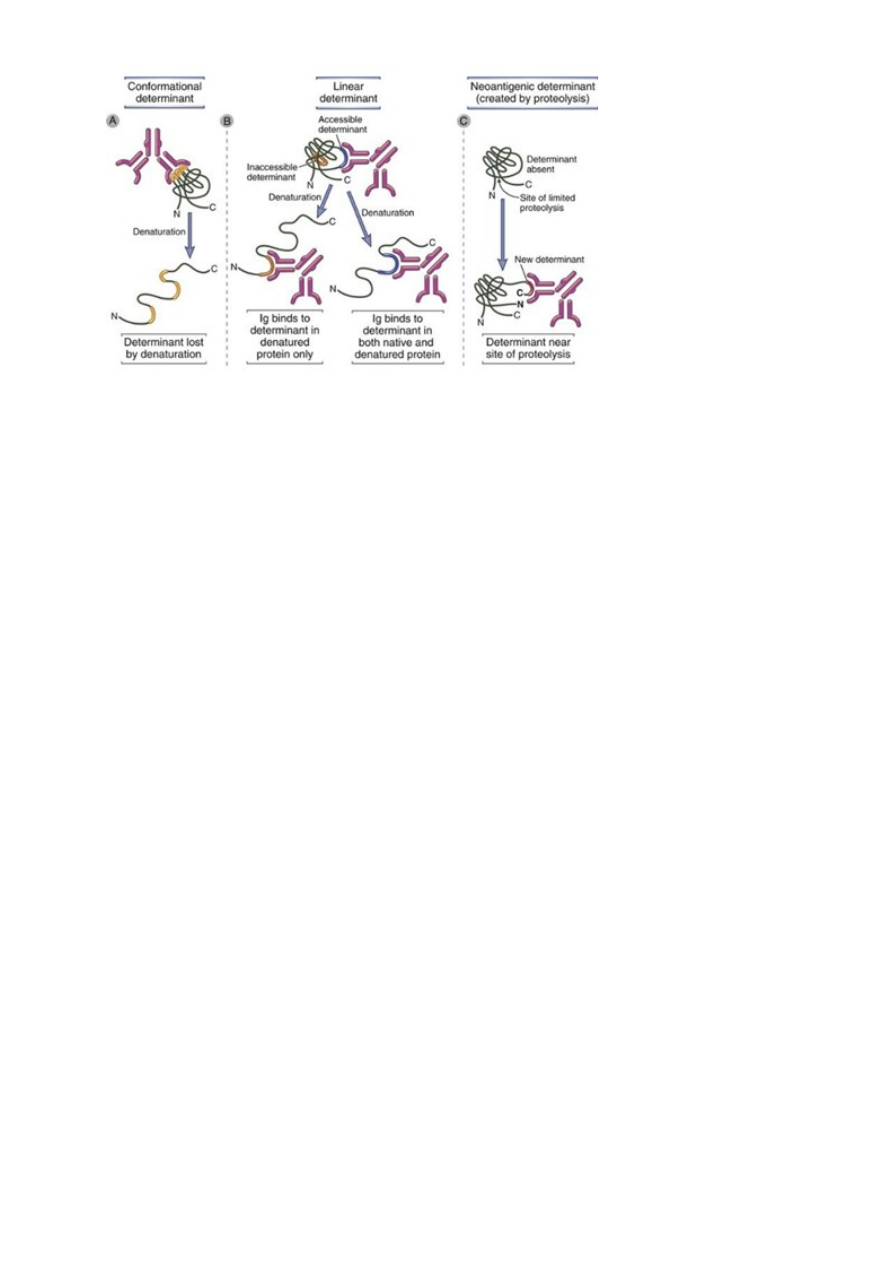

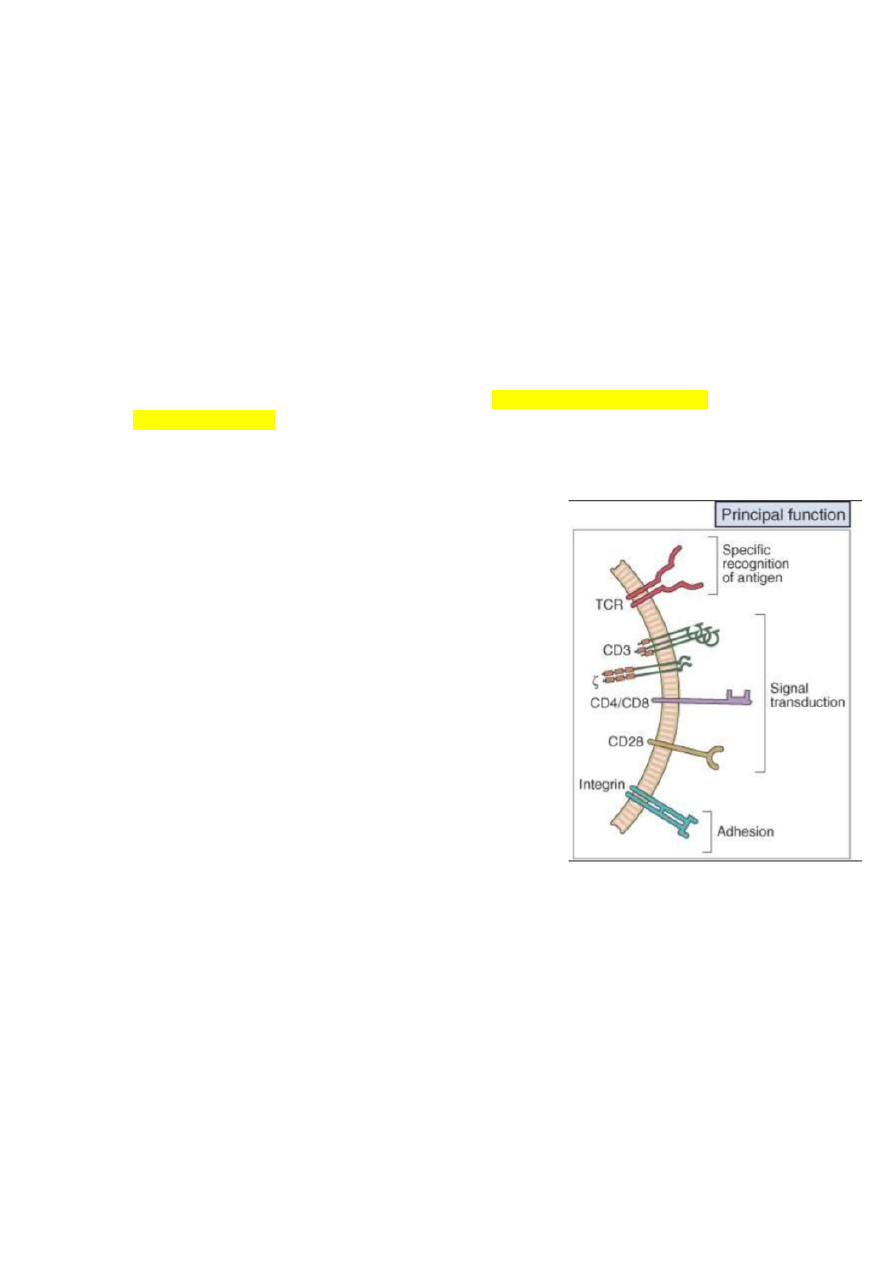














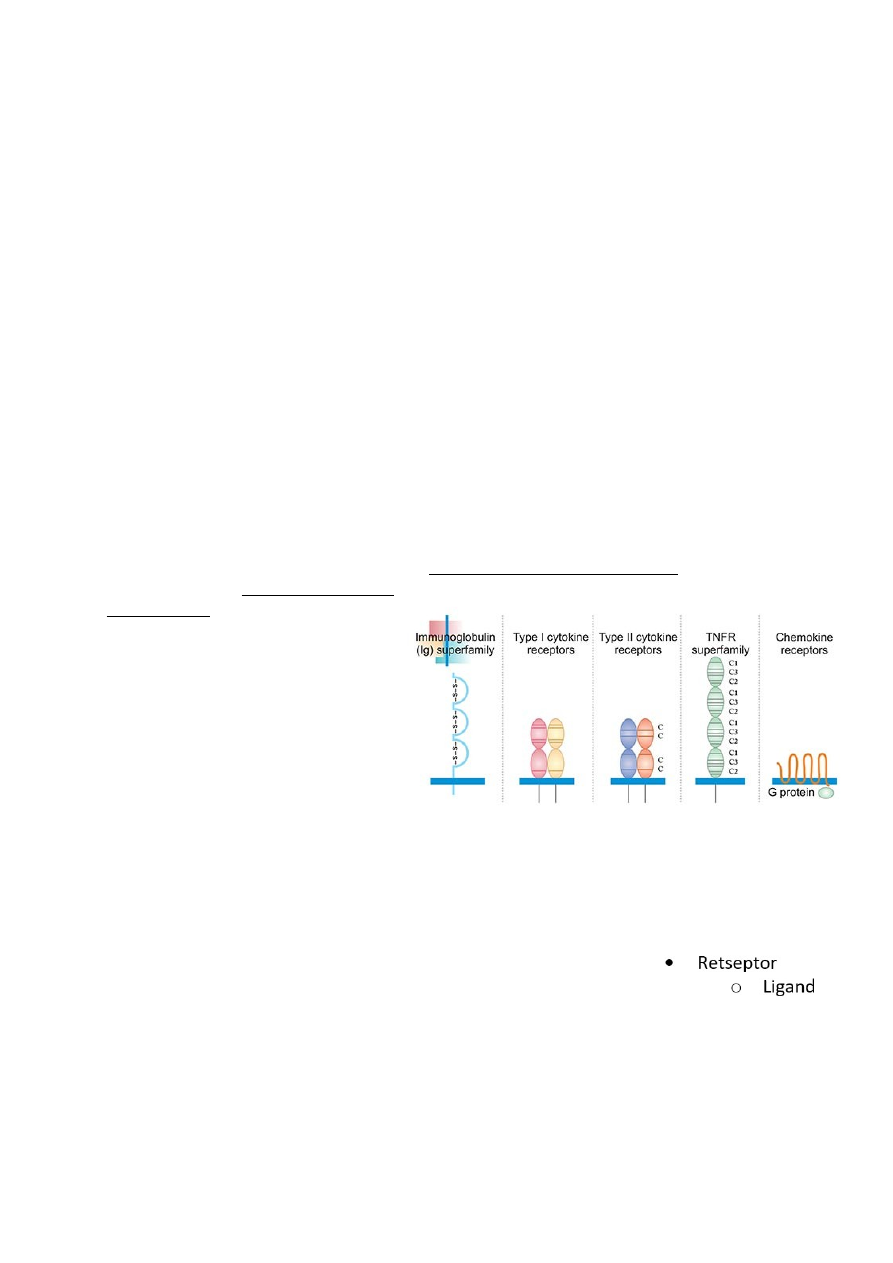
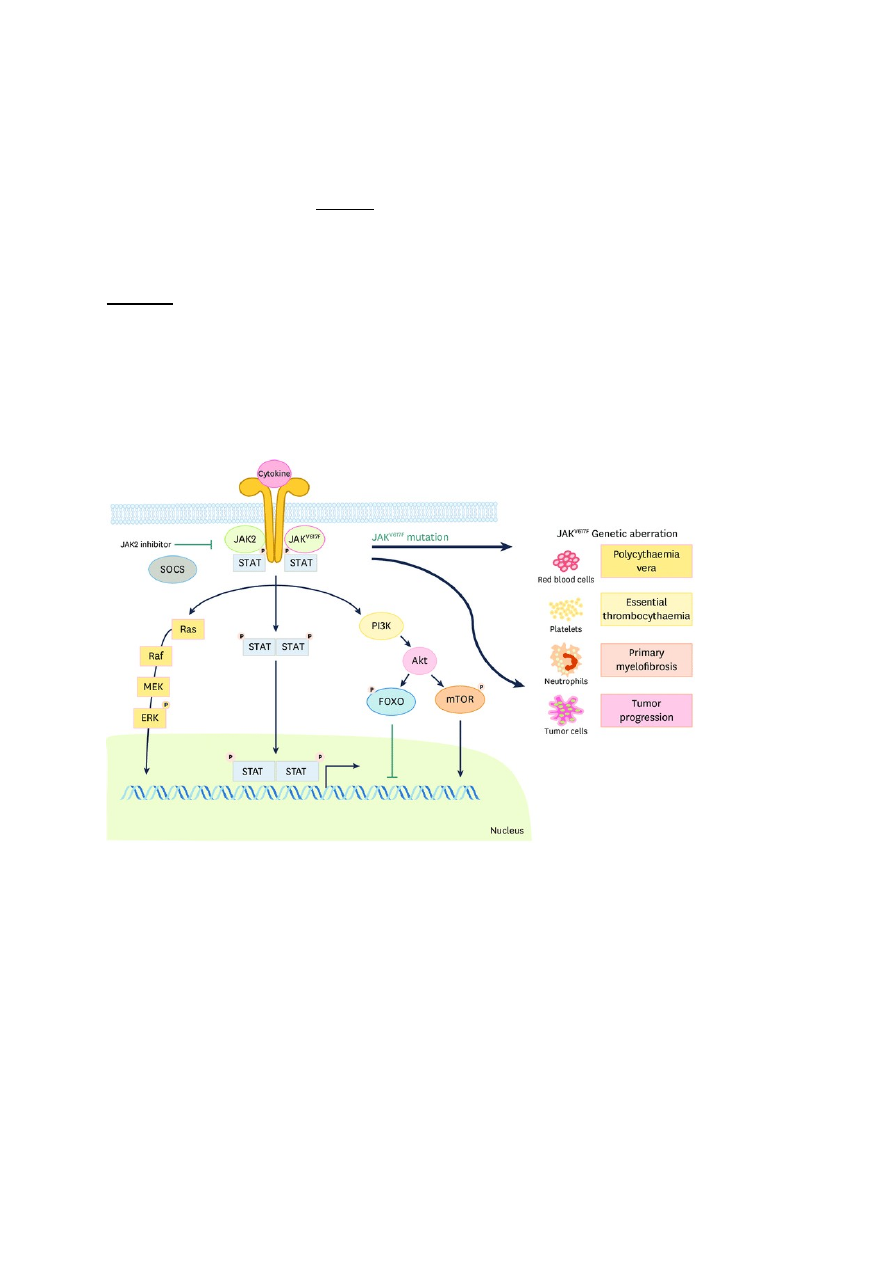
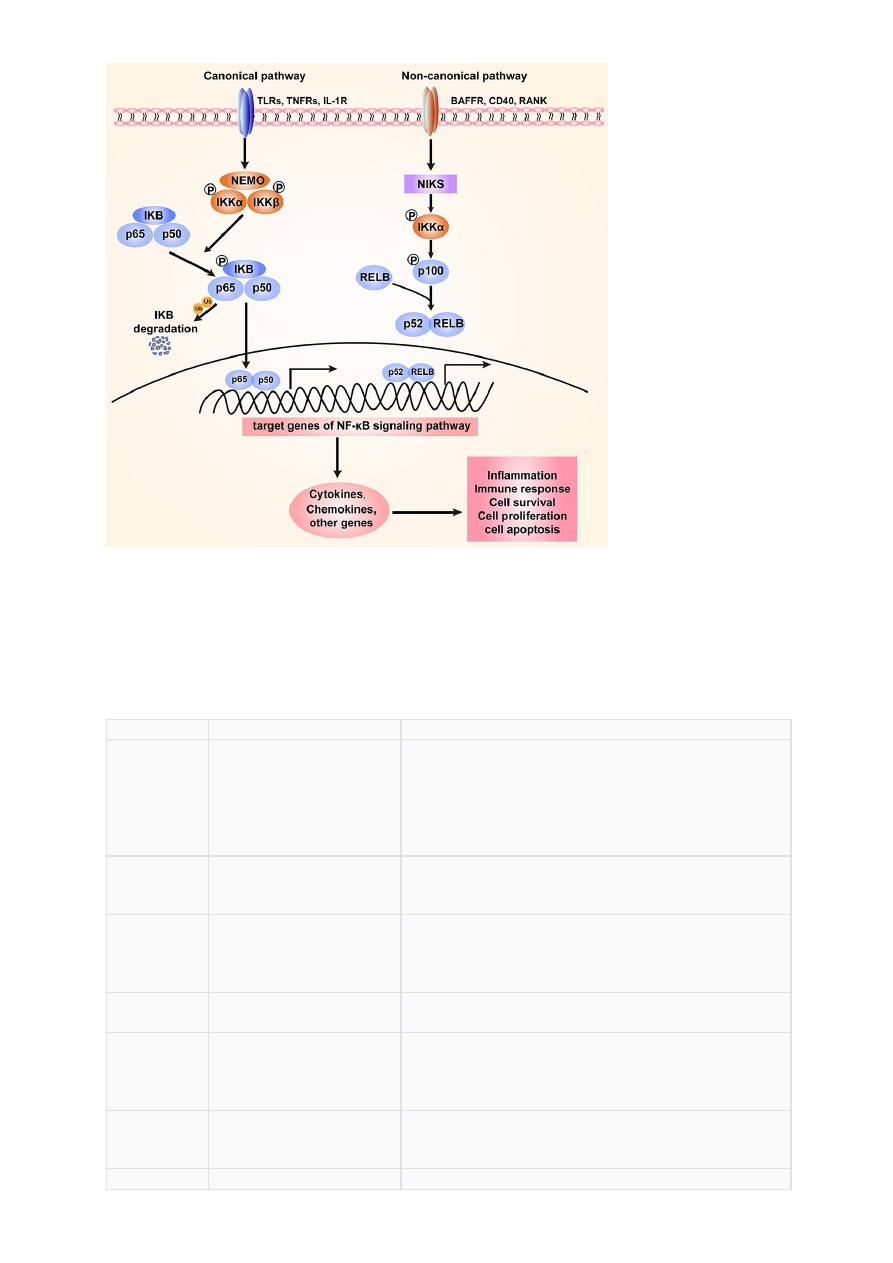
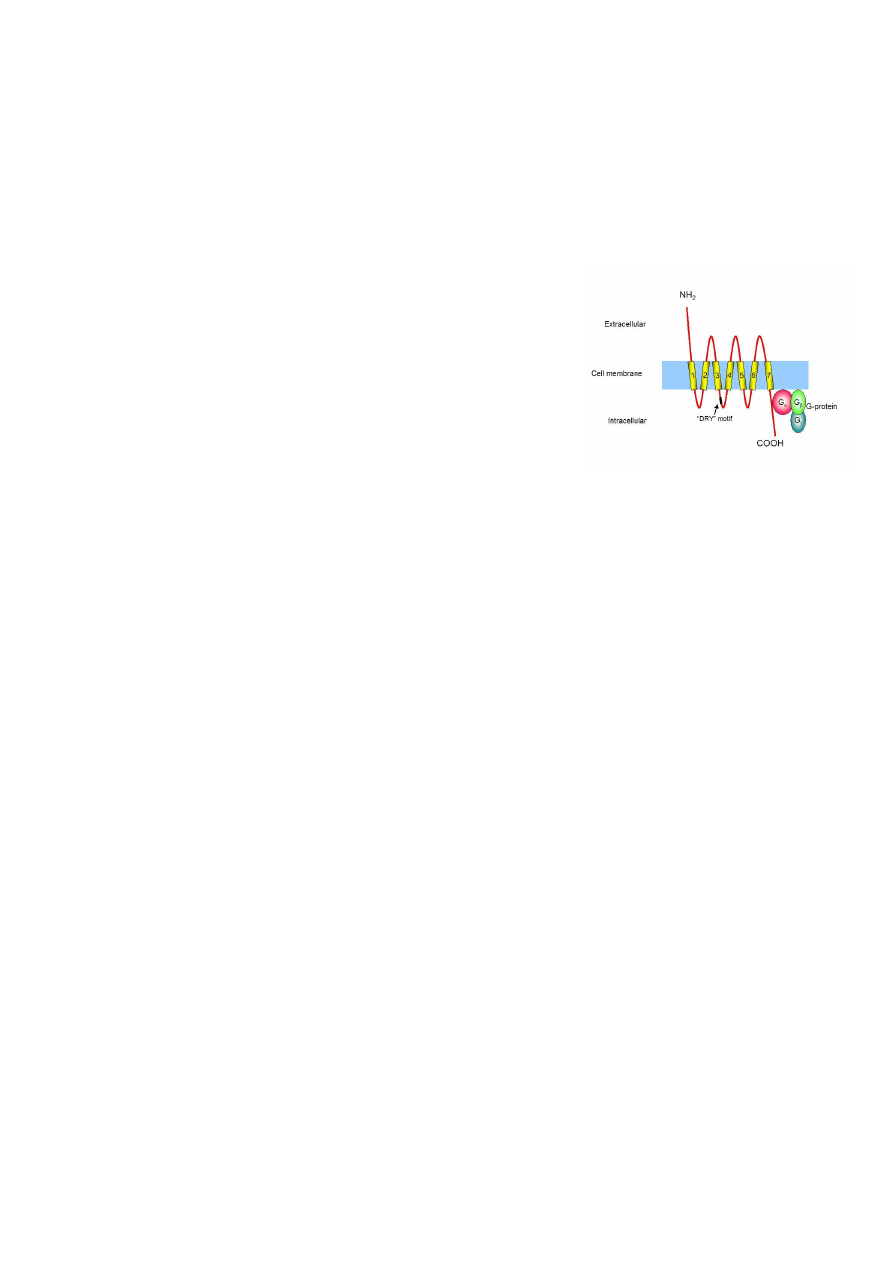


















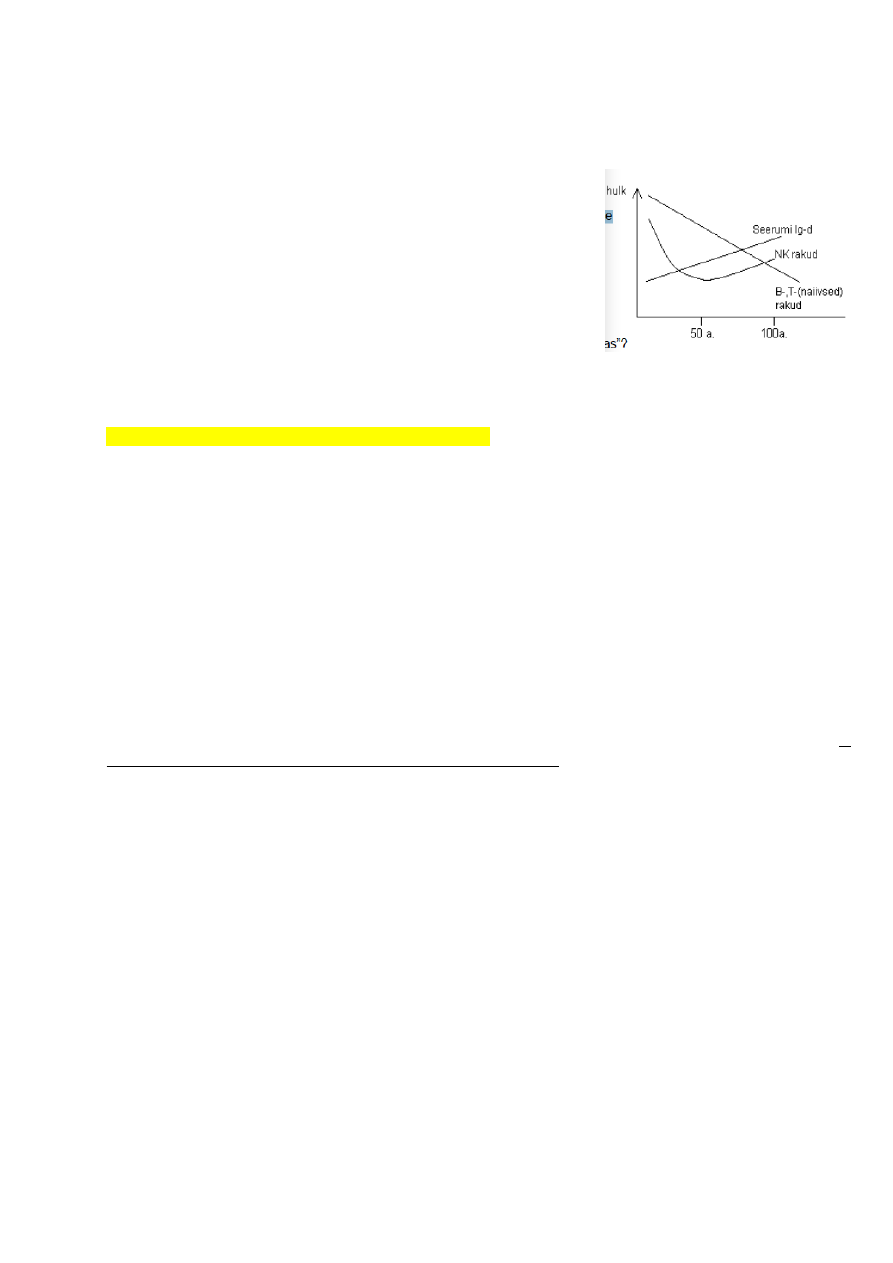

















Kõik kommentaarid