1.
loengAkronüümid:
TH1 – t
lümfotsüüdid helper,
IL
interleukiin
CD –
diferentseerumise klaster, raku pinnal olev
marker , valguline v
lipo.. marker. Seos diferentseerumisega – erinevatel rakkudel eri
omad.
DSCAM-
Down Syndrome Cell Adhesion Molecule, neuronid ei tunne üksteist
õigesti ära, liiga palju DSCAM valku Down sündroomiga imetajatel.
Drosophilal erinevad valgu variandid –
tunnevad ära eri patogeene.
PPR-
molekulaarset mustrit ära tundvad
TLRs- Tol
like retseptors
PAMPs-
patogeeniga seotud molekulaarsed mustrid
NLRs-
bakterite äratundmiseks mitte rakkudde pinnal vaid sees
EvolutsioonVarasemad
– ei kaasne mälu, iga kord samaga kokku puutudes sama vastus sama
tugev. Kuni kõhrkaladeni.
Omandatud
ehk adaptiivne alates kõhrakaladest hakkab tekkima,
antikehad , T ja
B lümfotsüüdid.
Komplement – Ehhinodermidest, seotud ka kaasasündinud immunoloogiaga,
kaasasündinud
retseptorid võivad midagi väga spetsiifiliselt ära
tunda, enne kui
patogeeni ei hävita ei juhtu midagi, et märklaudua
hävitada võib kasutada ka komplemendi süsteemi, ilma T
lümfotsüütide ja antikehadeta.
Immuungeneetika
osas ennetavast immuunsussüsteemist.
Immunoloogilise
kaitse põhimehhanismid:Geneetiline
polümorfism ,
erinevad rakupinna retseptorid, valgulised retseptorid, raku pinnal glükolipiidsed retseptorid, immuunreaktsioonides osalevad valgud . Vaja – ükski viirus ei tungi rakku ilma retseptorita, kui on mitu varianti, viirus ei saa nakatada tervikuna liiku , näide herpesiviiruste kohta, mõningaid suudab mõningaid mitte.
Valgud
Glükosiidsed, glükolipiidsed jne ABO ja teised veregrupid
Immuunreaktsioonides osalevad valgud.
Organism
võib hävitada ennast kui patogeeniga kokku satub, bakter ei saa
edasi levida, organism sureb aga poluplatsioon võida, inimese korral
on seda väha – hemorraagilised viirused Aafrikas, kaitse
forseeritakse nii tugevalt et see hävitab enda. Kaitsemehhanismid on
tuigevad ja need hävitavad.
Omandatud immuunsus omav väikset osa, tema käivitamine võtab kaua aega, ta
on efektiivne aga aeglane nö.
Enamiku
koormuse saab kaasasündinud immuunsus, see ei tee vigu. Omadatud
teeb vigu – autoimmuuhaigused.
Kaasasündinud
immuunsus
- barjäärid – Füüsilised: nahk, kitiinkest, munakoor,lima, temperatuur bakteritel kitsas temp vahemik kus saavad paljuneda, kehatemp kõrgemaks ja viirused ja bakterid ei saa paljuneda, liikuv ripsepiteel, köha .
Keemilised:
pH lüüsib edukalt paljusid patogeene (maos 2-3), lagundavad
ensüümid (lüsotsüüm – bakterite kestade lõhkumine), defensiinid .
Millise pH
juures saab lahti nukleiinhapetest – aluselises keskkonnast lasevad ahelad lahti, pH normaliseerudes tõmbuvad kokku tagasi, Alpides
kõrge pH ja neandertaallased näevad välja nagu uued ja DNA ka
säilid, Taani rabades jälle inimesed super hästi säilinud aga DNA
on juppideks. Valkudest saab vabaneda pH 13. Ja nukleiinhapetest pH
happeline juures. Mõlemal korral juppideks.
- Humoraalsed ( signaalid mida antakse edasi vere või lümfiga) ja tsütokiinide (signaalmolekulid, kiire süntees, peavad kiirelt retseptorile seonduma muidu lagunevad) abil vahendatud reaktsioonid.
- Immuunsüsteemi rakkude poolt vahendatud reaktsioonid ( fagotsütoos , NK rakud , põletik )
Kus on immunoloogia piirid? Kus saame hakata rääkima äratundmisest ja
kaitsest.
- Bakterid-restriktaasid ja metülaasid (arhedel ka), oma DNAd ei restrikteerita, äratundmispiirkond palindroom (alati?).
- Seened-antibiootikumid,
- Aktinomütseedid ja linnud -biotiini kõrvaldamine. Avidiin ja streptavidiin . avidiin (linnumuna munavalged, vaba biotiin sealt ära) ja streptavidiin (streptomütses, kasutab kui antibiootikumi seda valku) – tetrameersed valfud, ülikõrge afiinsusega vitamiin biotiini suhtes. Ilma biotiinina ei saa keegi elada, võtada ära siis suurepärne kaitse. Mida madalam dissotsiatsioonikompleks seda madalamatel kontsidel moodustavad ühendid ja ei lahku üksteisest, kiiremini toimuvad reakstioonid, siin ääretult kõrge afiinsus Kd= 10-15 , teised valgud transpordivad vitamiini B12 – ei ole kaitsesüsteemi valgud, suur afiinsus samuti Kd= 10-15. Teistel bioloogilistel molekulidel on suuremad diss konstandid.
- Ainuraksed – tsüklopeptiidid jne. Kasutavad neid samu mehhanisme oma elupaiga leidmiseks, peremeesorganismi rünnakute eest.
Imetajate
kujunemine
Imetajate
kujunemisel oli vaja ületad mitmeid väga keerulisi immuunsüsteemiga
seotud ülesandeid.
- Loode-oma/võõras, pinnal teistsugused markerid kui endal?
- NK rakkude vaigistamine – imetajad linnud kontrollivad oma rakke läbi T raku retseptorite ka son oma või võõras, üks võimalus siis et ei näita retseptoreid, aga siis tapavad NK rakud ära kui ei näita mis sees on. Näidatatkse ainult monomorfseid??
- Emakas - viirused, kasvajad sagedamini. Kaitsemehhanismid on siiski allasurutud, sp kasvajad. Kaitsesüsteemid alla surutud.
- IgA ja piim – lindudel antakse antikehad kaasa IgY tüüpi kaudu, munakollase kaudu, imetajatel selliseid antikehasid ei ole, IgG tüüpi suudavad läbida platsentat. Esmaste imetajatega on keerulised. Kaitsepomm esimeste päevade piim, mitte eelkõigetoit vaid IgA antikehad.
Viirused.
Liiga steriiline elu?
Oo
ei. Me leiame viirusi kõikjal mida puudutame, hingame või sööme.
Nad on ka osaks meie genoomist. Igas ml merevees on >1 miljon
viiruse osakest. Maailma joogivee reservuaarides on 1030
bakteriofaagi , igaühe kaal umbes femtogramm.Bakteriviiruste mass
ületab elevandi kaalu rohkem kui 1000 korda. Maailmas võib leida
umbes 1016
HIV viiruse partiklit. Vaal eritab merevette igal päeval 1013
Calciviridae (vaala diarröa) partiklit! Võib nakatada inimest ja on
soolases merevees üsna püsiv. Hepatiit E , gastroenteriit,
kõhulahtisus.
Defensiinid
Imetajatel,
Putukatel, Taimedel. Kõik kes vahepeale jäävad.
Väiksed
valgud, kaitsevad bakterite ja viiruste eest edukalt.
- Opsoniseerivad – märklaua märgistamine, mis on vaja kõrvladada, beeta defesniiniga, anitkeha seostumisegaa, komplemendiga, kui tulevad fagotsüüdid siis opsoniseeritud materjal kiirelt fagotsüteeritakse.
- Kemotaktilised, gradiendis võivad dentriitarkud liikuda nt
- Histamiinid midagi??
- Raku kasvu muutmine
- Märklaudu otseselt hävitada.
Mõningad
defensiinid on ajas muutunud veidral moel. Mürgised imetajad.
Kuubalt ja Haitilit – Solendon
cubanus –
putuktoiduline, süljenäärmetesse suudavad sünteesida mürgist
sülge, hammustavad loomi ja need kangestuvad. Ka isased nokkloomad
omavad mürginääret, tagakäpa eraldi küünis. Olemuselt on mürgid beeta-defensiinid.
Imetajatel
on tavaliselt mürgiseid asju vähe, tavaliselt on mürki reptiilidel
näiteks. Solenodon perekonna esindajatel on alalõua süljenäärmetes
mürginäärmed. Teist looma hammustades ohver kangestub.
Teeta(
θ ) defensiinid
Kui oleks
funktsionaalne, siis inhibeeriks HIV viiruse sisenemist. Ta on olemas
reesusahvil. Seda ei kasutata HIV teraapias. Teeta-defensiini
inimesel ei sünteesita, geen on muutunud pseudogeeniks, retrocyclin,
mis on saadud selle pseudogeeni aktiveerimisel, inhibeerib viiruse
HIV-1 rakku sisenemise. Kui poleks pseudogeenistunud siis HIV ei
saaks inimest kahjustada. On olemas reesus makaagil ja toimib.
Retrotsükliin
Brilacidin
– antibiootik, defensiin, ravim sarnane defensiinile mimetik. Ei
ole laias kasutused, viimase astme kliinilised katsetused. Abi MRSA
S. aureus ravi. Ka
Enterococcus
ja Klebsiella
raviks. Vähihaigete seennakkuste raviks.
Sfenistsiin
– pingviinide kõhus et see roiskuma ei läheks. Beeta defensiin.
Immunoloogia
ajalugu
Edward Jenner – immunoloogia isa. Loodusuurija, uuris kägude bioloogiat
et and teiste pesadesse lähevad, avastas selle, et linnud ei lähegi
mutta talveks vaid rändavad, ja tegi ka siilidest huvitavaid töid.
Vaktsiinidega seoses midagi.
Variolatsioon
– rõuge kärnade pealt materjali lasti sisse hingata, saadi
immuunsus sageli, vahel mitte. Hiinas Song dünastia, siiditeelt
inglismaa esimese feministi, oli konstantinoopolis ja lasi seda
variolatsiooni teha oma lastele, tõi Inglismaale selle. Euroopas
tehti nahapeale haavand ja panid kuivatatud kärnamaterjali sinna.
Orjakaubandusega Ameerikasse.
Leedi
suri ära kui Jenner oli 13. Nii et Jenner ei olnudki päris esimene.
Toll – retseptorid, leiti Drosophila arengut uurides alguses. Toll –
saksakeeles kärbse vastse kuju järg .
Polly
Matzinger – Ohuteooria,
Omandatud
ehk adaptiivne immunsus
- Lümfotsüüdid ja nende spetsiifilised retseptorid
- B lümfotsüüdid-immuunoglobuliinid (sekreteerituna antikehad)
- T lümfotsüüdid -TCR retseptorid
- Antigeeni esitlevad rakud ja MHC I ja MHC II
- Immunoloogiline mälu-see on B jaT lümfotsüütide klonaalne proliferatsioon
2. loeng
Kaasasündinud
(Innate) immuunsuse elemente
TOLL retseptorid ( PRR retsetorid)
Avastati Drosophila m. arengut uurides 1985a. (toll saksa keeles kummaline,võigas, imelik
1997 tuvastati imetajatel, Bruce Beutler 1998 TLR 4 ja LPS seos
–Nobeli preemia 2011 a.
Käivitavad NF-κB ja MAP kinaaside aktivatsiooni raja
Osa raku välismembraanil-osa endosomaalselt
N-Formylmethionin
MiIline on tähtsus eukarüootidele?
Scavenger (prügivedaja, raisasööja, püüdur) retseptorid inimesel
Tsütoplasmaatilised PPR (pattern recognition receptors)
NOD –(Nucleotide oligomerization domain) retseptorid . Üle 20
imetajatel (NOD2-muramüül dipeptiid)
RLR retseptorid
RNA helikaasid
Taimede PPR retseptorid(XA21, FLS2)
MBL (mannan-binding lectin)
C-reaktiivne valk
Komplemendi retseptorid
RNA helikaasid RIG-I ja MDA5
RIG-I ja akronüümid immunoloogilistes tekstides
RIG-I(retinoic-acid-inducible protein ) retseptorite (tsütosoolsed
RNA helikaasid) käivitatud signaalirajad
Taimede immuunsüsteemi elemente
PAMPS- Pathogen-associated molecular pattern
LRR- leucine-rich repeat (leutsiini rikaste kordustega retseptorid)
R genes (resistentsuse geenid )
RNA interference
Lihtsamad keemilised mediaatorid ja signaalnolekulid (salitsüül hape ,jasmon hape, etüleen.....)
Immuunreaktsioonid taimedel
miRNA
ssRNA molekule mõnest tuhandest kuni 40,000 molekulini raku kohta
Leitud kõigis metazoa liikides, 0.5-1% geenidest
siRNA märklauaks on geenid, millest on ise pärit miRNA reguleerib
erinevaid geene
100 erinevat mRNA molekuli võib olla reguleeritud ühe miRNA poolt
3. loeng
Immuunsüsteemi
rakud ja koed
Primaarsed lümfoidsed organid – spetsiifilised immuunsussüsteemi
organid – alates kõhrkaladest.
Sekundaarsed lümfoidsed organid – seotud lümfotsüütide
küpsemisega.
Kõik saab alguse pluripotentsetest tüvirakkudest, millest arenevad
erinevad rakud (pankrease närvi jne).
Vereloome tüvirakk annab aluse lümfoidsele ja müeloidsele rajale.
Müelodsest reast arenevad erütrotsüüdid ja teised vererakud va
need mis kuuluvad lümfoidsesse ritta.
Lümfoidses koes on b lümfotsüüdid, need suudavad sekreteerid ja
oma pinnal presenteerida antikehi. T- lümfotsüüdid, mis
diferentseeruvad eri suundadeks olulisemad t helper ja TC ehk
tsütolüütilised lümfotsüüdid. Samuti on siin NK rakud, siin
tuleb ka üks osa dentriitrakke.
Müeloidne ride – erütrotsüüdid, basofiilid , eosinofiilid,
vereliisatkud, neutrofiilid ja monotsüüdid.
Leukotsüüdid – tuumaga rakud (imetajatel). Trombotsüütidel ja
erütrotsüütidel pole tuuma. Nii lümfoidse kui müeloidse koe
rakud millel on tuum on siis leukotsüüdid. Kolm suurusjärku vähem
kui erütrotsüüte. Lümfotsüüdid – spetsiifilise immuunsüsteemi
rakud on tuumaga rakkudest 20-40%.
Rakkude nimed on seotud värvimisega. Basofiil – värvid mida
kasutati annavad aluselises keskkonnas reaktsiooni, neutrofiiilid
jne. Väiksed lümfotsüüdid - naiivsed, kui opn juba aktiveerunud antigeeniga kokku puutunud siis võib olla juba suurem.
Neutrofiilid (granulotsüüdid)
- 5x10¹º tsirkuleerib, eluiga mõned päevad põletiku korral, muidu 5-6 päeva
- 60-70% vere tuumaga rakkudest
- Tsirkulatsioonis eluiga 4-6 päeva, põletiku korral kudedes 1-2 päeva
- Graanulites laktoferrin, müeloperoksüdaas, defensiinid, elastaasid jne. Graanulites on valmis sünteesitud hulgaliselt tugevamtoimelisi ühendeid, mida kaitsereakstioonides kasutatakse, bioloogiliselt kõrge aktiivsusega ühendid.
Eosinofiilid
- 2-4% vere tuumaga rakkudest
- Parasiitide ja allergia korral oluliselt rohkem, seenhaiguste jms
- Ringes 8-12 tundi, kudedes 8-12 päeva
- Graanulites histamiin ,lipaasid, ribonukleaasid. Graanulites neutrofiilidega võrreldes veelgi tugevama toimega ühendid.
Basolfiilid
- 0,01-0,3% vere tuumaga rakkudest. Kudedes rohkem.
- Kudedes mast rakkud, nimetatkse nii
- Graanulites histamiin , hepariin. Sarnane eosinofiilife graanulite sisaldisele, aga võivad sekreteerida signaalmolekule vt alla.
- IgE retseptorid – allergilised reakstioonid
- Sekreteerivad prostaglandiine, leukotrieene
MONONUKLEAARSED FAGOTSÜÜDID
- MONOTSÜÜDID VERES
- MAKROFAAGID KUDEDES:
- ALVEOLAARSED MAKROFAAGID - KOPS
- HISTIOTSÜÜDID - SIDEKOES
- KUPFFERI RAKUD – MAKSAS
- MESANGIAALRAKUD - NEERUDES
- MIKROGLIIA RAKUD - AJUS
- OSTEOKLASTID - LUUDES
- OPSONISEERUMINE-(4000X ERINEVUS)
- väljanägemine võib olla erinev kudedes paiknevates makrofaagides, aga kõigile omane hea fagotsüteerimisvõime ja opsoniseeritud juhtudel fagotsüteerivad paremini.
- Hulgaliselt fago ja lüsosoome. Kokku fagolüsosoom – fagotsüteeritud materjali surmamine lõhustamine.
Erinevad tapmismehhanismid neil:
MÜELOPEROKSIDAAS- sünteesitakse sellest HÜPOKLORITit,
VESINIKPEROKSIIDIST JA KLORIIDI IOONIST
NITRIK OKSIIDI SÜNTETAAS-ARGINIINIST
NO(gaas) JA TSITRULLIIN (kõrvalprodukt)
- HAPNIKUST SÕLTUMATU TAPMINE
Erinevad ensüümid: LÜSOTSÜÜM, HÜDROLÜÜTILISI ENSÜÜME
DEFENSIINID - CYS RIKKAD KATIOONSED PEPTIIDID , suhtelised lühikesed
TNF – tugev tsütokiin
Osa lagundatud tükkidest pannakse MHCga raku pinnale, et
lümfotsüütidele näidata mida võõrast on kätte saadud.
Lümfoidne rida:
B – lüfotsüüdid
- BURSA OF FABRICIUS AINULT LINDUDEL, nime saanud siit, B-lümfotsüütide küpsemine lindudel ühes kindlas organis, inimestel sellist kindalt paika ei ole.
- ÜHEL B RAKUL 1.5x105 Ig MOLEKULI – spetsiifiliste retseptorite hulk
- B 220 (CD45 VARIANT – kas naiivne või küpsenud lümfotsüüt selle järgi) – Claster of differentiation. Praegu teada üle 300 eri markeri . Mõnu valguline mõnu sahharidne. Saab hinnata diferentseerumise astet jne.
- CD35 (CR1 complemendi retseptor )
- CD40 SEOSTUB CD40 LIGANDIGA T HELPERITEL
T- lümfotsüüdid
- KÜPSEVAD TÜÜMUSES nii lidnudel kui imetajatel, plaju läheb sisse vähe tuleb välja, kõik mis tunneb ära organimi oma antiugeene hävitatakse.
- HELPER JA TSÜTOLÜÜTILISED – omakorda veel mälu helper, helper 1, 2 jne.
- SEOSTUB ANTIGEENSE PEPTIIDIGA, MIS ESITLETUD MHC KAUDU, suudavad tuvatsada antigeene mis neile esitletakse makrofaagide poolt MHC kaudu. Tükeldatud bakter läheb MHCI ja viiruse poolt nakatatud rakk mis sünteesib viiruse valke MHCII . Mis ise see MHCII ja mis võõras MHCI.
- CD4
- CD8
- CD28 RETSEPTOR KOSTIMULAATOR B7-1(CD80)
NK rakud.
- AVASTATI 1976, hilja võrreldes B ja Tdega.
- MÄRKLAUAKS ON MUUTUNUD MHC ESITLEMISEGA RAKUD ( KASVAJA VÕI VIIRUSEGA NAKATUNUD)
- CD16( ANTIKEHAGA MÄRGISTATUD RAKUD), antikehade poolt opsoniseeritud sellised rakud.
- NK1-T -TCR,CD1 INTERAKTEERUMINE, CD16.
Dendriit rakud
Avastas Paul Langerhans 1865 a.kasutades kuldkloriidi naha rakkude
värvimiseks. Jätkelised rakud, seotud immuunsüsteemiga. Tema arvas
olevat närvisüsteemi osa.
1973 Ralph M. Steinman näitas, et nad on antigeene esitlevad rakud
(APC).
Mehhanismid mille kaudu kasvaja rakku NK rakud hävitavad. Perforiin
lüüsib (vaatem hiljem). Kasutatakde antikehi rakkude ära tunmiseks
ja apoptoodi indutseerimiseks Fas ligande. Tappa üritatakse mitmel
viisil, mitte kunagi 1 moodus.
Nk rakud tegelevad viirusega nakatuanud rakkude hävitaisega.
Omandatud immuunsuse tase – T rakkude tase kasvab 7-8 päeval ja
antikehade tiiter löövad käiku hiljem.
Esimese ja suure töö teevad ära innate immuunsussüsteem .
Omandatud immuunsus käivitud aeglaselt.
Pärast
aktiveerumist muuuvad T ja B lümfotsüüdid suuremaks, hakkab
moodustuma klöonaalne plajunemine. OsaB lümfokaid sekreteerib
akehasid verre, on palju suuremad kui naiivne B lümfotsüüt. Plasmarakk on valgusünteesi masin. Sureb 4-5 päeva pärast
apoptootiliselt. Samasugused nagu väiksed
lümfotsüüdid on
ka mälurakud, ei muutu suureks.
B lümfotsüüt puutub kokku antigeeniga, see käivitab rakutsüklisse
mineku. Tetud hjulk rakke väljub ja muutub mälurakkudeks, osa
efektorrakkudeks – mõned päevad a.kehade süntees.
Organid,
mis on imuunsüsteemi seisukohalt olulised:
Lümfisõlmed , lümfisüsteem .
Tüümus imetajatel, põrn ja luuüdi .
Lümfisüsteem
vereringest kudedesse, rakkude vahele siseneb 2-4 liitrit vedelikku.
Kuidagi peab see vedelik vereringesse tagais jõudma, see toimub
lümfiteede kaudu. Lümfisüsteem töötab erinevalt vereringest ühes
suunas. Suundub enne südant veresoonkonda. Lümfiringe ie ole päris sümmeetriline .
Mandlid
Suuremad lümfisõlmed, paiknevad sellel teel kus hingatakse õhku ja
neelatakse toitu. Kurgumandlite pind on väga sur, sest on ahtuline.
Lihtsatel organismidel, näiteks silmul on seotud immuunkoed seotud
seedesüsteemiga (GALT-gut associated lymphoid tissue). Kaladel
mängivad olulist rolli neerud .
Kõige lihtsam ehitus on kõhrkaladel, silmul
Organid on seotud seedesüsteemiga. Eraldi tüümus ja organite
jaotus on imetajatel ja ka lindudel.
Kahepaisketel on luuüdi ja tüümus.
Kaladel
ei ole luuüdi, sest luid
ei ole. Vereloomes
mängivad olulist rolli neerud.
Akronüüme erinevate kudede koosluste nimetamisel, mis osalevad
rakkude diferentsuuruvad
MALT – mukoosaga seotud lümfoidne kude, ei küsi neid.
Hematopoees
Esimestel nädalatel on lootel olemas yolc sac. Luuüdis on rakke
1/50 000, mis on tüvirakk. Kui luuüdi on hävitatud(nt vähiravi),
siis piisab umbes 0,1%, et kude taastada. Küpsemiseks on vaja
tsütokiinide signaali ja strooma rakkude osalust.
- LOOTEL - YOLK SAC- TÜVIRAKUD ESIMESTEL NÄDALATEL
- KOLMAS KUU - MAKS, PÕRN
- SEITSMES LOOTE KUU- LUUÜDI
- TÜVIRAKUD MULTIPOTENTSED E. PLURIPOTENTSED – palju on vaja multipotenseid luuüdirakke? VÄHE---ÜKS 50 000 LUUÜDI RAKUST ON TÜVIRAKK.
- FUNKTSIOONI TAASTAMISEKS PIISAB 0,1-0,01% . Nt vaja vereloome kasvaja tõttu vaja hävitada ja siis taastada.
- KÜPSEMISEKS VAJA TSÜTOKIINIDE SIGNAALI JA , et tüvirakkudest diferentseeruksid rakud
- STROOMA RAKKUDE OSALUST lisaks vajalik diferentseerumiseks
- (KOLOONIAID STIMULEERIV FAKTOR-CSF JA ERÜTROPOIETIIN-EPO JT.)
- REGULATSIOON GEENIDE (TRANSKRIPTSIOONIFAKTORITE) TASANDIL, kui plaju milliseid rakke teha.
- 3,7x10¹¹ LEUKOTSÜÜTI PÄEVAS
Luuüdi
Strooma rakud, adipotsüüdid. Erinevate lümfoidse ja müelodse rea
rakkude valmimine.
Proovi võtmine, analüüsi või siirdamiseks. Vaagnaluu, seal sees
on käsnjas struktur, mis on luuüdi, seal toimub vereloome.
VERELOOME RAKKUDE KULTIVEERIMINE PEHMES AGARIS
Tüüpiline on pehme agari kultuurid. Vajalikud strooma rakud, sest
vereloome rakke ei saa kasvatada ilma abirakkudeta.
Transkriptsioonilisi faktoreid on teada väga palju, vaatame
olulisemaid.
Gata -1 erütroidse rea puhul
Gata-2 erütrodne, müoloidse ja lümfoidse rea omad.
Ikaros
Oct2 – paneb B lümfotsüüdid dif, antikehi sekret plasma rakkudeks.
Oluline on nende kindel kombinatsioon pigem.
Apoptoosi puhul toimub kogu DNA lagundamine, ei ole võimalus et
viirus saaks edasi minna. Nekroosi puhul seda ei juhtu ja vabaneb
selliseid markereid, mi käivitavad põletiku reaktsiooni.
Apoptootiline rada on normaalse elutegevuse osa.
Faktorid , mis apoptoosi reguleerivad.
Caspaasid – olemuselt proteaasid
Bcl faktorid – nii inhibeerivad kui toetavad
lõpptulemus oleneb erinevtae faktorite taskaalust
Oluline ka Fas faktor ja fas ligand
Ühe konkreetse antikeha saamiseks hübriseeritakse kasvajarakk ja B
lümfotsüüt ja saaba mahukalt antikehasid.
Makrofaagides olevad toksilised ühendid. Slaid.
Lümfiringe ei ole sümmeetriline, imetajatel paremas poolest pool
pead ja üks käsi, osa rindkerest suundub ühte kohta ja ülejäänud
kolmveernad organsimi teise kohta. Lümf liigub lümfisõlmedess tänu
keha liikumisele, lihaste liikumisele, saab tänu klapikestele
liikuda ainult südame suunas. Kudede vahel olev vedelik saab imbuda
lümfisoontesse.
Tüümus
- Digeorge sündroom – inimesel kui tüümust pole, raske haigus.
- Nude mice – kui tüümust pole hiirel, kui steriilsetes tingimustes kasvatades sudab elada vanaks, kui aasta vana leitud mõni t lümfotsüüte kust kurat keegi ei tea.
- 70g lapsel - hiljem 3g, vastsündinul on suur organ, täiskasvanul väiksem
- Sisestruktuuris oluline: erinevad makrofaagid, spetsiifilised epiteliaal ja dentriitrakdu, suurel hulgal apoptootilis rakke, (naiivsed T rakud kes ei sobi),
- Hassall´scorpuscles—TSLP tüümuse strommalne lümfopoietiin-reg.T lümfotsüüdid
Bursa
Lindudel B lümfotsüütide küpsemine. Imetajatel suvalistes
lümfisõlmedes.
Lümfisõlm
- Lümfi kapilaaridest tulevad lümfiteed lümfisõlme
- Lümfisõlmes : vesiikuleid, antigeeni esitlevad rakud, lümfotsüüidi, seal toimub neile antigeen esitlemine ja transformatsioon.
- Lümfisõlmede suurus võib muutuda oluliselt seoses põletikuga
- Sinisega on terminaal tsentrid, lümfotsüütide aktivatsioon plasttransforatsioon
- välja viib lümfijuha.
- Kahes lümfoidses organis, meenutavad lümfisõlme, aga erinevad selle poolest et pole sissetulevaid lümfiteid, põrn ja tüümus
- Põrn on filter vereringe süsteemile, nagu lümfisõlm, samad fn, aga ka lisafn – vererakkude depoo, kõrvaldab ringest vanaks jäänud erütrotüüdid, raud pannakse tallale, hemoglobiin läheb maksa sapi komponentideks bilirubiiniks. Põrnas on valge ja punane pulpo . Punane on erütrotüütide oma – võetakse välja, valge pultp – sarnane tavalistelelümfisõlmedele, põrn on lümfisõlm vereringe jaoks.
- keelel olevas lüfoidses koes ja kurgumandlites
- Kõik mis on seotud massaasiga on seotud lümfiringe aktiveerimisega. Lümf peab liikuma õiges suunas, ei tohi pressida vastupidi.
4. loeng
ANTIKEHADE
STRUKTUUR JA FUNKTSIOON
ANTIGEENID
1. SUBSTANTSID,MIDA TUNNEVAD ÄRA, KAS
B LÜMFOTSÜÜTIDE IMMUUNOGLOBULIIN RETSEPTORID või
2. PEPTIIDID, MIDA TUNNEVAD ÄRA T LÜMFO- TSÜÜTIDE RETSEPTORID
(PEPTIIDID ESITLETAKSE MHC MOLEKULIDEGA KOMPLEKSIS)
IMMUNOGEENSUS—OMADUS INDUTSEERIDA HUMORAALSET JA / VÕI RAKULIST
IMMUUNVASTUST
ANTIGEENSUS –VÕIME REAGEERIDA KAS ANTIKEHADE VÕI T LÜMFOTSÜÜTIDE
RETSEPTORITEGA
HAPTEENID-VÄIKESED MOLEKULID,MIS VÕIVAD OLLA IMMUNOGEENID KUI ON
SEOTUD SUUREMALE “ KANDJALE”. Nt penitsilliin võib seonduda
meie oma vere valgule(albuliinile) ja muutuda antigeeniks siis.
Immuunoglobuliin domään
70-110 amiinohapet, võileiva taoline kahest kihist
antiparallelsetest beeta ahelatest (7-9) struktuur, kus on väga
konserveerunud disulfiid sild (2 tsüsteiini).
Antikehades võime ristada konstantseid domääne aga aminoterminuses
on piirkonnad mis vastutavad äratundmise eest, tegemist on
variaablite domäänidega. On ka teisi valke kus võib leida
immunoglobuliinide domääne (CD48helperitel) ja
CD8(tsütotoksilistel)). Adhesioonimolekulid – ICAM.
Siirdamisreaktsioonid
on keerulised, sest MHCII
klassi molekule kontrollitakse,
kui esitlatakse võõra isendi omasid (võõras rakk), neid on väga
palju variante inimestel. Klass I MHC – oma materjal ja klass II
võõras.
Koaalade polümorfsim on väike ilmselt pudelikaela läbi elanud.
IgG tüüpi antikeha koosneb kahest raskest ahelast mis on
disuldiidsildadega seotid, kerged ahelad on omakorda seotud
disuldiidsildadega raskete ahelate külge. Saame eristada antikehaga
seonduvad piirkonnad ja bioloogiliseaktiivsiuse piirkonnad sabaosas,
mida tutakse ära komplemendi valkude poolt vms, peale antigeeni
äratundmist et midagi toimuks siis sabaosas on see piirkond.
Tähistamine:
C-konstantne, V-variaable, H- raske, L-light kerged ahelad.
Konstantseid osasid võib raksetel ahelatel olla rohkem IgG del kolm
ja IgM neli näiteks. Raksete ahelate klasse tähistatakse kreeka
tähtedega. Imetajatel on viis. Gamma (IgG), delta (IgD), alfa (IgA),
müü (IgM), epsilon (IgE). Kahte tüüpi kergeid ahelaid kappa või
lambda.
Antikehade funktsioon ei sõltu sellest kas nad on glükosüleeritud
või mitte. Sellest sõltub nende eluiga. Et püsiks orgaismis (nt
kui toodame bakterites) peab olema glükosüleeritud.
Variaable domään – halliga eriti varaablid domäänid. Mustaga on
tähist kahe tsüsteiini vahel ole disulfiidsild.
EPITOOBID – piirkonnad millele antikehad seonduvad
1.STRUKTUURSED – see võib olla struktuur, mingi jupp valgust mida
ära tuntakse, pole oluline kas on natiivne või denatureeritud,
enamus antikehasid
2.KONFORMATSIOONILISED – tunnevad ära anult natiivseid antigeene,
tunnevad ära struktuuris olevaid aminohappeid mis ona struktuuris
üksteise kõrval.
3.NEO – avanevad kui oleme antigeeni proteaasidega töödelnud,
midagi avanud ära lõiganud.
Müoglobiin ja tema antigeensed piirkonnad.
Piirkonnad mis jäävad antigeeni pinnale, pöördekohad – need on
need alad mida antikeha meelsamini ära tunneb.
Antigeen antikeha omavaheline seostumine. L1 L2 L3 – kerge ahela
variaablid domäänid, H1 H2 H3 – raskete ahelate variaablid.
Loopid. Seondumine ei ole kovalentne aga on väga tugev.
Vereseerumis
on kõige rohkem albumiini valku, kuidas lahutada valgud? Lahutamine
sõötub nende isoelektrilisest täpst. Albumiini isoelektriline täpp
on happelises piirkonnas. Isoelektrriline täpp – pH väärtus kui
tema laeng on null.
Albumiiin liigub kiirelt siis.
Grupp valke mis liigub vähe, isoelektriline punkt pH 8. Gamma
globuliini mõiste on seotud elektroforeesi tehnikaga.
Inimese
viis erinevat klassi antikehasid, eri tüüpi raske ahel. IgM tüüpi
antikegadel (ja IgA tüüpi) on peale raskete ja kergete ahelte veel
J-segment
valk (ahel)??
esimene antikeha mida sekreteeritakse on IgM, kui patogeen ründab.
Saa kindlaks teha nt kui millal borrelioos ründas, IgM tekib algul
ja mõne nädala jooksul langeb ja toimub ümberlülitumine IgG tüüp
antikehadele.
IgE
tüüpi seotud ülitundlikkusreakstioonidega. IgA sekreteeritud siis
on dimeerid, neljavalentsed, veres?
on niisama kahevalentne. Antikeha mida sekreteeritakse kopsudes,
limaskestades, pisarates süljes. Suures hulgas ka soolde sekreteeritakse seda.
IgGd on veres levinuimad. Klassi sees on alamklassid. Erinevus on
hinge piirkonnas (disulfiidsildade arv). Subklasside hulk on liigiti
erinev, inimestel on neli.
IgA -l on ka alamklassid, neid saab eristada ühe lektiini abil.
IgG
alamklassid
Erinevus
on hinge piirkonnas olevate disulfiidsildade arvus, ehk erinevus on
bioloogilises piirkonnas, mis põhjustab erinevatele retseptoritele
seondumist(näiteks komplement) Antigeeni äratundev piirkond on suht
sama.
IgA
sekreteerimine
Plasma rakk toodab IgA molekuli, mis seondub polü-ig retseptorile
mis asub epiteelraku pinnal. Siis võetakse ta sisse vesiikuliga.
Toimub ensümaatiline lõikamine retseptoris ning IgA vabaneb
sekretoorselt, kusjuures retseptor on temaga seotud, aga lõigatud
kujul. Pilt ka.
Millised olid esimesed lektiinid mis Tartus puhastati?
Sekretoorne komponent – et saaks vereringets mujale transportida.
Kuidas see toimub? Nt soole raku pinnal on veresoonte seespool PolyIg
retseptor 8koosneb struktuurselt imuunoglobuliini somäänidest -
sugulasvalk), võimaldavad transporti ensüümid mis lõikavad osa
PolyIgst ära ja see jääb IgA dimeeri külge. Vastsündnutel
esimestel päevadel see protsess toimub vastupidi.
Kolostrum – imetajatel peale sünnitaist esimestel päevadel
sekreteeritav piim – toitaineid suht vähe, ülikõrges kontsis
IgA-sid, defensiine ja valgud mis aitavad vabaneda materjalist mis
looteeas on kujunenud soolde.
Antikehaline vastus väiksetel ühenditel – suurem kandevalk,
organismi enda valkudet levinud albumiin. Kui tahame immuniseerida hiiri vms, siis kasutatakse kandevalguna suurt valku – ürgvähiliste
hemolümfist puhastuat valk, sinna külge seondatakse või seondub.
Võib ära tunda seda väikset molekuli, seondumiskohta vms. Palju
katseid on thetud dinitrofenüüliga. Antikehad mis teevad vahet eri
asendites bensoehapete karboksüülrühmadel. Antikehde
spetsiifilisus võib olla väga kõrge, eritavad peenstruktuure.
Ravimite muutumine immuunogeenideks.
Erinevate antikehade toime uurimine. Kuidas kolme erinevat antikeha
saavad antigeenidga seonduda. Dinitrofenüülid seotud mingi
spetssidemega.
Kolostrum
Pärast
sündi esimestel päeval sekreteeritav piim, see on täiesti teise
koostisega ja ka teist värvi. Ülikõrges kontsis on seal IgA
antikehasid, defensiine ja muid kaitsevalke. Lisaks veel selliseid
valke, mis aitavad vastsündinul vabaneda soolde kogunenud
materjalist. Ravim Travelan on tehtud puhastatud IgA molekulidest.
See ravim aitab reisijatel vältida seedehäireid.
Väikesed
antigeenid on seotud kandjaga. Antikehad tekivad väikesele
molekulile, kandjale kui ka mõlema segu konjugaadile.
Dinitrofenüülfosfaat
on aine, mille kaudu saab mõõta hinge piirkonna pikkust.
IgE
IgE tüüpi
antikehasid on veres vähe, aga nad kutsuvad kohale kiire
reaktsiooni. IgE on seostunud mast rakkude või basofiilidega ja
ootab seal antigeeni. Nende tihedus rakupinnal on suur ja kui allergeen seob kahte antikeha raku pinnal ristseoseliselt, siis
vabanevad graanulid, mis sisaldavad histamiine ja muid allergilise
reaktsiooni vahendavaid ühendeid.
Nuumrakud vabastavad histamiini ja mõningate minutite jooksel võivad
tekkida kopsuturse jne. IgE tüüpi antikehad on spetsiifilstel
rakkudel ja ootavad kokkusaamist antigeenidega.
Paar slaidi lasime üle.
Biotehnoloogias on antikehade kasutamisel probleem. Neid kasutades,
võib juhtuda probleem, et mingi kude seondab antikeha, mitte
antikeha ei seonda antigeeni. Selle vältimiseks saame rasked ahelad
ära lõigata teatud ensüümidega. Alles jääb fab osa.
Kui tahame antikehasid kasutada mingite asjade ära tundmiseks.
Keeruline vahet teha aks seondumine oli spetsiifiline või sp et seal
olid vastavad retseptorid mis tundisd selle antikeha ära. Nt
vereloome rakkudel on olemas retseptorid mis tunnevad ära antikeha
(seda piirkonda millest enne rääkimine). Fc osa lõigatakse maha ja
alles jääb Fab osa. Oleneb ensüümist saab erinevaid fragmente
F(ab')2, silla piirkonnas jäävad kokku, või Fab või üks veel. On
olemas antikehade retseptorid mis tunnevad neid siis ära kui
antikehad on millegiga seondunud.
Silmud
Pole RAG
põhinevat rekombinatsiooni mehhanismi, kasutavad retseptorite
mitmekesisuse saamiseks geeni konversiooni. Lahknesime 500 miljonit
aastat tagasi. Potentsiaal 10 neljateistkümnendas astmes erinevat
retseptorit. Imetajatel 10 kaheksandas astmes erinevat T raku
tetseptorit.
Silmud
sekreteerivad antikehalaadseid struktuure, mis on stabiilsed pH
1,5-11.0 juures ja temperatuuril 56° umbes nädal. Need on nii
sekreteerituna kui B-lümfotsüüdi pinnal.
Kõrge
aviidsus- Spore agglutination by VLR4 was detected at a concentration
1,000- fold more dilute (5 pg/ml) than the mouse monoclonal antibody
(5 ng/ml).
Antikehade
korral peame rääkima aviidusest, mitte afiinsusest, sest tal on
kaks äratundvat piirkonda.
Alates luukaladest on antikehad.
Silmudel pole, aga neil on midagi muud. Pole
rekombinatsioonimehhanismi, retseptorid/antikehade laadsed molekulid
mitmekesisuse saamiseks kasutavad geenikonverisooni. Afiinsus ja
aviidsus. Afiinsus – korrektne olla siis ei saa me rääkida
antikehade afiinsusest, sest tal on kaks äratundmissaiti. Rohkem kui
kaks isegi. Aviidsus
midagi segast veel juurde. Silmude
antikehadel on kõrge aviidus.
- Pole RAG põhinevat rekombinatsiooni mehhanismi, kasutavad retseptorite mitmekesisuse saamiseks geeni konversiooni. Lahknesime 500 miljonit aastat tagasi.
- Potentsiaal 10 neljateistkümnendas astmes erinevat retseptorit
- Imetajatel 10 kaheksandas astmes erinevat T raku tetseptorit
- Stabiilsed pH 1,5-11.0 juures, temperatuuril 56° umbes nädal
- Kõrge aviidsus- Spore agglutination by VLR4 was detected at a concentration 1,000-fold more dilute (5 pg/ml) than the mouse monoclonal antibody (5 ng/ml).
Haide ja raide antikehad, i- body
- Antikehad kuni 10x väiksemad (14 Kda) ja eriti oluline, et nende antigeeniga seostumise ala on väiksem. Suudavad tuvastada piirkondi, mida imetajate antikehad ei erista.
- Võivad seostuda ka hüdrofoobsemate alade, „ pragude “ j a muude selliste piirkondadega („clefts and groves“) mida imetajate antikehad „ära ei tunne“.
- Ülimalt stabiilsed.
Haid ja
raid
Antikehad
kuni 10x väiksemad (14 Kda) ja eriti oluline, et nende antigeeniga
seostumise ala on väiksem. Valkude pinnal olevad praod või struktuurid on haige antikehadega tuvastatavad. Võivad seostuda ka
hüdrofoobsemate alade, „pragude“ ja muude selliste
piirkondadega („clefts and groves“) mida imetajate antikehad „ära
ei tunne“. Ülimalt stabiilsed.
Haidel
võime leida antikehasid, millel puuduvad konstantsed ahelad.
Luukalad
Luukaladel
on antikehade klasse palju rohkem kui imetajatel. Need on suht madala
afiinsusega. Afiinsusküpsemine nõrk või puudub. Tüümus on
olemas, lümfisõlmed puuduvad.
- IgM, IgX, IgR, IgW, NAR, IgM redox, klasse on rohkem kui imetajatel, evolutsioon on läinud lihtsustumise teel
- madal affiinsus, aeglane vastus
- afiinsusküpsemine nõrk või puudub
- tüümus olemas, lümfisõlmed puuduvad
Kasuatakse biotehnoloogias antigeenide immuniseerimiseks mis on väga
ürgsed ja konserveerunud.
Reptiilid
ja linnud
Roomajatel
kui osadel lindudel leidub trunkeeritud IgY tüüpi antikeha.
Imetajatel sellist pole. Seda sekreteeritakse nende munakollasesse,
kaitseks. Munavagesse sekreteeritakse IgA ja IgM. Võrreldes imetaja
IgG-ga on need natuke pikemad . Antikeha klassi tähistus tuleb sõnast
yolc. Lindude antikehadel on väiksem painduvus.
Kanal on
eelis, sest lindude Fc piirkond ei seondu imetajate Fc
retseptoritega, seepärast on lindude antikheade kasutamine
eelistatud.
Kaamlid,
alpacad
Osa nende antikehadest on normaalsed(rasked ja kerged ahelad), aga
teatud osa antikehadest(IgG2 IgG3) on ilma kergete ahelateta ja
täiesti funktsioneerivad. Aga sellist antikeha on lihtsam kloneerida
näiteks maisi või tubakasse. Sellisel viisil saab valmistada
söödavaid vaktsiine. Selliste antikehade kasutamine on palju
lihtsam. Sellepärast toodi need loomad vivaariumisse.
Trunkeeritud IgY ( ΔFc) osadel kilpkonnadel ja partidel, hanedel
(non-gallioformes (gallioformes – kanad ja nende sugulased)).
Imetajate IgG ga võrreldes IgY on pikemad.
Munakollane – sinna sekreteeritakse IgY tüüpi antikeha.
Kui on valk kas küülik või kana siis kana on eelis kui tahame
imetajate rakke uurida lindude Fc piirkond ei seondu imetajate Fc
retseptoridga.
Monoklonaalsed antikehad
Põhineb antikehade klonaalsel päritolul, lõpuni dferentseerunud
lümfotsüüt sünteesib ainult üht tüüpi antikehi, millel on üht
tüüpi spetsiifika. Tootvale B-lümfotsüüdile oleks vaja anda
igavene elu. Nad viiakse kokku sellise rakuga, millel ei ole stopi,
selleks sobivad B lümfotsüütide kasvajad – müeloomid, eriti
head on sellised mis antigeeni ei
sünteesi. Kuidas tagada ainult hübriidide ellujäämine hiljem?
Saavutatakse selektiivsete söötmetega. Müeloomi liinid erinevad
ainevahetuse sõltuvuste tõttu (või muu iseärasuse tõttu). Lisaks
on vaja, et müeloomi ja B lümfotsüütidel oleks samad MHC kompleksid , kasutatakse puhtaid liine, paljukordse tagasiristamisega
saadud. Van/pan hiired??
Mikrotiiterplaatidele külvatakse nii, et ühte auku satub
tõenäoliselt üks rakk, seda korratakse ja nii saadakse puhtad
liinid, mis toodavad ühte antikeha. Monoklonaalset antikeha
toodetakse rasedustestide tarbeks. Kõige enam kasutatakse hiiri, aga
ka rotte kanu. Ka inimesi, inimest maha ei lööda ja põrna välja
ei võeta, loodus ise immuniseerib, skriining peab teine olema,
puhastatakse verest B rakud, olemas on ka inimese müeloomi liinid,
pikaajaline geneetiline stabiilsus inimese hübriidsete rakkude puhul
on halb.
Kui tahame tappa ingit rakku mida antikeha ära tunneb, siis
kinnitame antikeha külge mingi toksiin nagu ritsiin. Või seotakse
külge väiksed magneetilised osakese. Paramagneetiline osake,
magneetilised omadused siis kui on tugevad magnetväljas, väga
väikesed.
Monoklonaalsed
antikehad-on pärit ühest konkreetsest b-lümfotsüüdist.
B-lümfotsüüdid
on viidud kokku teise sellise rakuga, millel pole sellist stoppi
(kasvaja analoog). Selleks sobivad müeloomid, mis ise ei suuda
antikeha sünteesida. Kuidas välja valida, et ellu jääks ainult
hübriidsed rakud, hübridoomid- müeloom , b-lümfotsüüt. Neid
kasvatatakse selektiivsöötmel ja nii saab rakke välja valida. Neil
peavad olema ühesugused MHC molekulid, need saadakse puhaste
katseloomade liinide kasutamisel.
Selleks,
et B-lümfotsüüte oleks piisavalt, immuniseeritakse katseloomi.
B-lümfotsüüte mõjutatakse mitmete antigeenidega ja siis tekib
plasma rakk. Mikrotiiterplaadile külvatakse üks rakk (ideaalis)
ning siis sünteesib ja sekreteerib ta lahusesse antikehi.
Kõige
rohkem sünteesitakse beeta-gonadotropiini vastast antikeha, seda
kasutatakse raseduse tuvastamisel.
Antikehale
seontakse külge paramagnetilise osakese abil mingi toksiini ahel.
See süstitakse organismi, anikeha liigub kindla märklauani, mis
asub näiteks vähkkasvajas. Seejärel kui antikeha on seodnunud,
rakendatakse tugevat magnetvälja ning seejärel see rakk sureb.
Paramagnetiline osake-magnetilised omadused on tal ainult siis,
kui on magnetväli. Hästi väikesed.
5. loeng
KOMPLEMENT
Charles Bordet –avastaja ,Nobeli preemia 1919. Täheldati, et
bakterite ja rakkude lüüsiks on antikehade seostumisel vaja veel
midagi – nn.komplementi . Rohkem kui 30 erinevat valku
Evolutsiooniliselt väga vana . Kolm erinevat käivitumisviisi---”teed”:
- klassikaline-märklaua äratundmine antikehadega – evol kõige noorem
- alternatiivne-bakterite ja muu võõra pinna eripärasused
- lektiinne-bakterite pinna mannoosi rikkad struktuurid – evol kõige vanem
Näitas värske seerumi lüütilist aktiivsust,mis kadus 56C juures 30 min jooksul nn. komplemendi inaktivatsioon. Kui seda teha
unustatakse laboris siis saadakse veidraid tulemusi, sest käivitub
komplemendi ahel.
Adaptive immunity – kõhrkaladest, merisiilikutest komplement.
Mis toimub? Millised on bioloogilised finktisoonid komplemendi
aktivatsioonil.
- Märklaud-rakkude tsütolüüs, võõrad bakteri, seenerakud või organismi enda rakud, misnon vaja kõrvaldada - pooride moodustamine ja osmootne lüüs
- Võõraks muutunud materjali opsoniseerimine - märgistamine fagotsütoosiks (teine varaint opsoniseerimikes oli antikehadega, näiteks tahma ei saa opsoniseerida antikehadega, aga on vaja kehast eemaldada - komplement)
- Opsoniinid - märgistamises osalevad komplemendi valgud
- Põletik reaktsiooni aktivatsioon - anafülotoksiinid, liiga tugev vastus
- Immuunkomplekside kõrvaldamine ringest - ?
- Humoraalse immuunvastuse toetamine - B rakkude mõjutamine germinaaltsentrites, mõningad signaalid lähevad B rakkudele, toetavad nende küpsemist lümfotsüütides
Zümogeenid – valgud, mis muutuvad proteaasideks spetsiifilise proteolüütilise toime järel. N'äiteks komplemendi kolmas valk C3,
millel ei ole ensümaatilist aktiivsust enne kui proteaas teda pole
püganud veits, siis C3 pügab järgmist jne – kaskaad .
Järjestikuline zümogeenne kaskaad (võrreldav hüübimis ja
kiniinse kaskaadiga).
Komplemendi süsteemi valgud (nomenklatuur)
- C1(qrs), C2, C3, C4, C5, C6, C7, C8, C9
- faktorid B, D, H ja I, properdiin (P)
- mannosi siduv lektiin (MBL), MBL seotud seriin proteaas (MASP-1 MASP-2)
- C1 inhibiitor (C1-INH, serpiin), C4-binding protein (C4-BP), decay accelerating factor (DAF), sellise protsessi reguleerimiseks on vajlikud inhibiitorid
- C1 receptor (CR1) – plaju sellistel rakkudel nagu erütrotsüüdid, on prügivedajad, kui antikeha on seostunud antigeeniga ja üles leitud komplemendi süsteemiga kogu see kompleks seotakse selle retseptori vahendusel retseptori külge ja viiakse põrna, protein-S (vitronectin)
C1 (C1qr2s2) 750kD
C1q 6A-24kD, 6B-23kD,6C-22kD
C1r 85kD 2x seriinesteraasne aktiivsus -varjatud
C1s 85kd 2x seriinesteraasne aktiivsus- varjatud
Valkude konformatsiooni muutused
1. antikehal mis on immuunkompleksis
2.C1 peale seostumist immuunkompleksile
- Kui antikeha seondub kasvaj takule, bakterirakule, siis tema konformatsioon muutub aj seda tunneb ära C1qr2s2. Üks IgM või vähemalt 2 IgG klassi antikeha.
- Kui C1q on seostunud, muutub ka tema konformatsioon ja see toob kaasa varjatud seriin - proteaasse aktiivuse avaldumise.
- Veres on olemas komplemendi faktor C4 ja C2, mis on selle proteaasi substraadid. Mida tähendab ülemine on valgu nime peal – varajtud proteaasne aktiivsus, tegu on zümogeeniga.
- Need lõigatud tükid moodustavad kompleksi C4b2a. C4a on signaalmolekul.
- C4b2a substraadiks n C3 faktor.
- Kui C3 lõigatakse, siis üks osa sellest seostub olemasoleva C4b2a-ga ja tekib suurem val C4b2a3b, millel on oma substraat ja proteaasne aktiivsus, selle substraat on C5.
- Osa sellest 3b-st ei seostu C4b2a-ga, vaid võib seostud amitmesgusutele pindadele, aminorühmade, hüdroksüülpindadega, kui kiirest seostuda ei saa siis reageerib mingi tioolesterside kuskil möllas jahm mhm jeh mnm
- C5 lõigatakse siis C5a-ks (tugevatoimeline anafülotoksiin), C5b teeb ka midgai
- Kui oleme jõudnud C5 konvertaasini läheb kolm rada samamoodi edasi, kuidas selleni jõuda on erinev
Edasi hakkame ehitama märklaud raku sisse auku. C5b seostub
komplemendi faktoritega C6, C7, C8. Lisaks on veel 9-11 9C faktorit,
mis moodustab poori seina, kõüike seda kokku nim membraani
atakeerivaks kompleksiks.
Sisemine tioester side
Aktiveerub C3 ja C4 spetsiifilisel lõikamisel (järgijäänud osa
konformatsioon muutub ja aktiveerub sisemine tioester side, mis on
väga reaktiivne). Peab väga kiiresti reageerima või hüdrolüüsub,
kui on midagi millele seostuda, siis seostub. Reaktsioon ruumiliselt lokaliseeritud. Ei erista oma ega võõrast. C3b esmajoones antikeha
seoselisel aktivatsioonil. C4b alternatiivsel aktivatsioonil. 200000
seostumist sekundite jooksul. Kuidas oma rakke kohe ära ei lüüsi,
oma rakudel on pinnal spetsiaalsed valgud, mis sinna seostunud C3b
või C4a sealt jupi ära lõikavad ja ei saa reakstioon edasi minna.
Bakteri seinale võõra raku seinale, tahmaosakele, kovalentselt
seostumine. Elusale materjalile tehakse auk sisse, kui on tahmaosake,
siis seostub mingi muu C3 mingi C4 ja C1 nii
et viiakse prügiga maksa?
On terve hulk baktereid, mis suudavad pääseda, kuidas nad kaitsevad
ennast komplemendi aktivatsiooni eest? Table 13-1. Malaaria ei peaida
ennast asjata erütrotsüüdi eest.
Alternatiuivne tee
6. loeng
Tsütokiinid
- Immuunreaktsioonides signaali ülekande ja rakkude diferentseerumises osalevad valgud.
- Väikesed (8-30 kD)
- piko.-nanomolaarsed konts.
- Hormoonide sarnased. Hormoonidele sarnesed, aga me ei saa neid ühte patta panna, sest tsütokiinid ei levi üle organismi, vaid kõrvalolebvatele rakkudele tihti samale rakule mis sünteesib.
- Väga lühikese elueaga (RNA ebastabiilne)
- Sünteesitakse vastusena mingile spetsiifilisele signaalile
- Toimivad ainult läbi retseptorite. Peab kiirelt retseptorile seondma, kui ei seondu siis hävib.
Leviku viisid
- Endorfiinse toime tulemusel tekib palavik. Levib verega.
- Kõige levinum on parakriinne toime, mõjub kõrval levatele rakkudele.
- Autokriinne ka levinud, miks selline jama vaja, mitmesugused mehhanismid käivituvad alles siis kui mingi osa nendest retseptoritest on hõivatud, või sünteesitakse kõrgema afiinsusega retseptoreid, kriitiline piir, protsess võtab teistsuguse ilme.
Toimed:
- Diferentseerumis mõjutamine, kõik diferentseerumise protsessid on tsütokiinide poolt mõjutatavad??
- pooldumine
- Inhibeerivad rakkude paljunemist
- võivad põhjustada märklaudrakkude apoptoosi, ka autokriinselt
- metaboolne aktiveereimine
- kemotaksis , rakkude liikumine mingis suunas, ka paigale jäämine
Tsütokiinide omadused:
- pleiotroopsus, üks konkreetne tsütokiin võib seostuda erinevatele märklaudrakkudele ja mõjub sarnase mõjuga.
- Reduntantsus – sama tulemus võidakse saada samade tsütokiinidega
- sünergia – kui IL 4 ja 5korraga võib atgajõrjesk olla allergiline reakstioon
- antagonism – kaks erinevat koos, võib tekkida blokeerimine
Kaskaad
IL2 ja IL4
– võiks meelde jätta omandatud immuunvastuse vahendajatena.
Kaasasündinudtest TNF,
IL1, aga mõlemas on
veel hästi palju teisi.
Kaasasündinud – põletiku reaktsiooni esilekutsumine ja rakud
lähevad kudedesse veenulites, kuhu minna ja kes lähevad –
tsütokiinide abil kontroll.
Tsütokiinide molekulaarsed struktuurid
Tsütokiine on jummala palju, ja ka retseptoreid ka. Neid on
klassifitseeritud nende struktuuri järgi. Viis põhilist klassi. Iga
retseptori tüübi kõrval siin toodud ka tsütokiinid, mis on
ligandiks.
Imuunoglobuliinide superperekond
klass I tsütokiinide retseptorid, konserneerunud tsüsteiinid, osad on retseptoriteks ka päris hormoonidele ja sp pole hormoonide ja tsütokiinide piir päris selge
klass II – valdavaltr interferoonide retseptorid
TNF retseptorid - monomeersed erinevalt klass I ja klass II, FAS ligand kasutab seda tüüpi retseptoreid
Kemokiini retseptorid – kompleks kahest valgust, G – valk ja membraane läbivad G-valguga seotud retseptorid. Maitsmistunne.
Erinevate tsütokiinide signaal võib käivitada sama signaalraja ka
raku sees, või põhjustada sama asjade sisselülitumine tuumas.
Signal transduction mediated by cytokine receptors = A series of
protein tyrosine phosphorylation
Signaali ülekanne tuuma
tüüpiline on et kui retseptorile seondub signaalmolekul siis see
toob kaasa retseptori dimeriseerumise ja retseptoriga seotud rakus
sisemuses olevate kinaaside aktivatsiooni. Enamasti kinaasid JAKid.
See toob kaasa dimeriseerunud STAT saab läbida tuumamambraani ja
osaleda transkriptsioonil, seondudes Tfdega. Meeles pidada türosiin
kinaasse aktiivsega molekulid, nende dimeriseerumine.
- Retseptorite ristsidumine ja oligomeriseerumine.
- JAK (Janus kinase) autofosforüleerumine –türosiinkinaasid.
- STAT (Signal Transducer and Activator of Transcription ).
- Imetajatel 7 erinevat STAT valku (geeni)-seostuvad kindlate geenide promootor alale-osa türosiinkinaasid, osa mitte.
NFκΒ activation by TNFR and TLR receptors. Mingi ubikvitineerimine.
Samadelt adaptormolekulidelt võidakse käivitada läbi
proteiinkinaaside teatud geenide aktivatsioon, või käivitatakse
teatud geenide aktivatsioon läbi NF-kappaB või läbi kaspaaside
apoptoor, siis on veel regulaatorid, mis midagi nendest kolmest
rajast võimednavad või peatvad. Signaal läheb küll väga kiirelt
tuuma, aga mitu rada on ja iga raja tugevust tuunitakse.
NF-kappaB
- nuclear factor kappa-light-chain-enhancer of activated B cells, nagu B rakkude kappa kerge ahela võimandav faktor,
- tegelikult on aga praktiliselt kõikide rakkudes, mitte ainult B rakkudes.
- Sisse lülitavad, käivitavad selle raja - Stress, tsütokiinid, vabad radikaalid, ultraviolettkiirgus, erinevad antigeenid, tol retseptorid, raku sees olevad retseporitd.
- Heterodimeer - Rel ja p50
TNF -toime
TNF- tapab kasvajarakke, sai sealt selle nime, aga organismis, et
kasvajaid tapaks siis vaja sellist kogust et tapab ka organismi.
Kui on vähe – mõned üksikud rakud sekreteerivad, plasmas konts
madal – muutused veresoonte läbilaskvuses, teatud adhesiooni
molkulide eksrprseerumine, tatud neutrofiilide aktiv makrofaagide
aktv – lokaalselt
Rohkem- jõuab ka teisetsse oragnitesse – luuüdis käivitab
vererakkude kiirema küpsemise, maksa akuutse faasi e põletikuga
seotud valkude sünteesi suurenemine.
Liiga palju – siis süda lakkab töötamast, trombid ja maksa kaudu
hüpoglükeemiline shokk.
Kui organism on rakelt haige siis enne surma näeme TNF kõrget
kontsi ja selle järgi ennustada kui kaa ja kas organism saab veel
elada.
Aktiivse faasi valgud, mida maksaas tehakse:
Fibrinogeen, C reaktiivne valk – selle järgi vaadatakse kas ja kui
tugeva põletikureaktsiooniga tegu on, käitub opsoniseerivalt.
Tsütokiinid ei mõju ainult teistele imuunrakkudele vaid mõjutatakse
veel lisaks näärmeid, mis sünteesivad hormoone. Mõju
organsüsteemidele.
Mis järjekorras ja mis kiirusega rakud signaalmolekule suudavad
sekreteerida?
Esimesena pärast mõne minuti pärast signaali saamist – tunni
järel on signaal maksimumis, loetud minutitega hakkab TNF süntees,
mis muudab kohalpeal olevate endoteeli rakkude läbilaskvust nii et
neutrofiilid ja makrofaagid pääsevad kahjustatud rakkude juurde,
IL-1, siis lülituvas sisse tsütokiinid mis hakkava immuunvastust
alla reguleerima. Samasugust asja näema ka miteete kasvajate puhul.
Veel mingi võrdlus, kell juba 45.
süsteemsete efektidega on vähesed tsütokiinid ja surnai shokini
viivad on esimesel kohal TNF.
IL-2 – tüüpiline lümfotsüütide ja T-helperitega seotdu
tsütokiin, mõjub nii tesitel T helperitele tsütolüütilistele ja
NK rakkudele
IL-4
IF-gamma ei jõudnud
Kemokiinide
Viirused – kavalpead
Kasutavad tsütokiinise mimikrit. On viirusi, mis suudavad
sekreteerida lahustuvast interferoon gamma retseptorid, inter gamma,
mille organism peaks kasutama...seotakse ära ja ei jõua
adressaadini.
Miks hea kui teatud homoloogid on, sest need mis on homoloogid on immuunsusüsteemi mahasuruvad
interleukiinid??
E. Metspalu:
Tsütokiinide toimemehhanismide põhiomadused.
Ei ole spetsiifilised, vajavad oma retseptorit ainul. Funktsioneerimise lausek son retseptorite hulk.
1.Tsütokiinid
ei ole antigeen ega MHC spetsiifilised.
Funktsioneerimise
aluseks ruumiline lähedus , spetsiifiliste retseptorite hulk ja
lisasignaalide olemasolu. Immunoloogilise sünapsi käigus toimub
tsütokiinide suunatud liikumine
2.Erinevad
tsütokiinid funktsioneerivad B-rakkude erinevates arengufaasides.
Mõjutavad proliferatsiooni, antikehade sekretsiooni, isotüüpset
ümberlülitumist. Koosmõju võib olla kas sünergistlik või
antagonistlik.
3.Erinevates
liikides võib ühe ja sama tsütokiini mõju olla erinev.
4.Toimemehhanismide
aluseks on seondumine B-rakkude vastavate pinnaretseptoritega.
Isotüüpsel ümberlülitusel antikeha spetsiifilisus ei muutu
kuid muutuvad effektofunktsioonid mida antikehad võimelised läbi
viima.
Tsütokiinide sekretsioon kasvad, aga mõjutatavate rakkude pinnal
kasvab ka nende retseptorite hulk. Immunoloogilise sünapsi
tekkimisel toimub tsütokiinide suunatud liikumine nende kahe raku
vahel. Tsütokiinide üleprodutseerumine, superantigeeni puhul, –
hakkvad seonduma ka neili kus on vähe resteptoreid. Rünntakse ka
keha enda rakke, mis ei ole infektsiooniga seotud.
Üleminek plasmarakkudeks toimub enamasti väljaspool lümfisõlmi.
Neid iseloomustab antikehade sekretsiooni kõrge tase. See tagatakse
kergete ning raskete ahelate transkriptsiooni kiiruse järsu tõusuga.
Ilmselt toimub ka vastavate mRNAde stabiliseerimine. Mehhanism pole
päris selge.
Erinevused ning sarnasused T ja B rakkude aktiveerimisel
Mõlemal juhul vajalik:
1.
antigeeni äratundmine
2.
tsütokiinide osalemine
3.
kontakt teise, eelnevalt aktiveeritud, immuunsüsteemi rakuga.
T rakkude puhul 1. ja 3. tingimus täidetakse koos lähtuvalt TCR
omadusest tunda ära peptiid -MHC kompleks. Autokriinset kasvufaktorit
IL2 ning teisi vajalikke tsütokiine toodavad ise.
B-rakkude korral aktivatsiooniks vajalikud signaalid on
füüsiliselt lahutatud. BCR abil toimub antigeeni äratundmine, MHC
kaudu kontakt TH rakkudega. Vajalikke tsütokiine produtseerivad TH
rakud.
7. loeng
Ülitundlikkus
Igasugune ülitundlikkus ei ole allergia.
Eristatakse nelaj ülitundlikkusreakstsioono ja nüüd ka viies on
välja toodud:
IgE seotud ülitundlikkus – allergiaga seotud
IgG seotud ülitundlikkus
Immuunkompleksidega sotud ülitundlikkus -
Rakulise immuunsusega seotud ülitundlikkus (DTH- delayed-type hypersensitivity) – suur rühm, aetakse allergiaga segamini, siia kuulub ka palju tuntud haigusi, nt tuberkuloos v leepra .
Retseptorite vastaste antikehadega seotud ülitundlikkus
1. ja 4. on väga palju erinevaid asju keskmised on väiksemad.
Miks meduusid põhjsutavad tugevaid põletusreaktsioone.
Vahetu ülitundlikkus—viivitusega ülitundlikkus
Õppimise skeem.
Mast rakud – kudedes olevad basolfiilid.
Tüüp I- ehk IgE tüüpi akehadega seotud
IgE tüüpi antikeha esineb ainult imetajatel. Papagoil ja kanal
sellist allergiat nagu meil ei ole saada. IgEd on vaja kehasiseste
parasiitide, ainuraksete kehasisese nematoodide vastu võitlemiseks,
selliste tugavte reakstioonidega saab neid katseid teha.
Allergeen – multideterminant, võimeline ristsiduma sama
tüüpi kõrvuti antikehi.
Algselt on IgM ja siis toimub tüübi ümebrlülitamine, see
määratakse lokaalsete tsütokiinide suhtega, kui on IL-4
esmajoones, siis see määrab selle, aks organism reageerib IgG, IgA,
IgE tüüpi. Sünteesitakse IgE tüüpi antikehi, need on plasmas,
eluiga lühike. Siis nad leiavad rakud nende spetsiifilised fc
retseptorid – basofiilid, mis on kudedes mast rakud, sinna jäävad
IgE-d kinni ja ootavad allergeeni, nädalad eluiga. Kiire allergilise
reakstiooni käivitavad – allergeeni sidumine ristseoseliselt
kahele retseptorile, need peaksid siduma sama antigeenset
determinanti, põiekestes on bioaktiivsed ained, mis vabanevad siis. Veres kui antigeen seodnub antikehaga ei tohiks midagi juhtuda.
- IgE –seotud ülitundlikkus (tüüp I) ehk allergia. Ainult imetajatel
- Allergeenid – multideterminantsed
- IgE spetiifiliste Fc retseptorite ristsidumine käivatab immuunreaktsiooni
- Evolutsiooniliselt tekkinud võitluseks parasiitidega
- Atoopia-IgE reaktsioon mitteparasiitsetele antigeenidele, kala või pähkli suhtes
- Pärilikk eelsoodumus – 5 kromosoomi, polümorfismid geenides q õlas (IL3,IL4,IL5,IL9, IL13 )
- 11kromosoomi q õlas (IgE retseptor), retseptori eri variantide olemasolu.
- 0,1-0,4 mikrogrammi ml veres - vähe
- Johansson ja Bennich 1967 IgE müeloom
- raskel ahelal lisa CH 4 domään
- IgE poolestusaeg vabana 2-3 päeva,seotuna nädalaid
- IgE spetsiifilised retseptorid basofiilidel ja nuumrakkudel
- Basofiilid-tsirkuleerivad veres
- Nuumrakud -kudedes:nahas, sooleümbruses, lümfisõlmede läheduses, hingamisteedes
- FcεRI väga kõrge affiinsusega 1x10¯9M, väikene hulk veres on, kiiirelt seotakse retseptori poolt ära. Seostub IgE CH4 domääniga
- Fcε RII madala affiinsusega 1x10 ¯6M, retseptor kaks, seda on ka tesitel rakutüüpidel, regulatoorse tähtsusega. Osa temast võib lahusesse vabastada ja lahustub ja seotakse Ige ära. Kas allergiline reakts on tugev v mitte, kui rohkem seotakse lahutuva poolt ära, siis pole nii hull. Seostub IgE CH3 domääniga. Lahuses cCD23.
IgE - FcεRI interaktsioon on teiste Fc retseptoritega võrreldes
tugevaim.
IgE seostumine FcεRI retseptoriIe pikendab IgE eluiga
(poolestusaega) oluliselt.
Kõrga afiinsusega retseptor koosneb kolmest ahelast.
IgE antikehade ristsidumisel võib kasutada teisi molekule, samuti
saame aktiveerida reaktsiooni tesite ühenditega või yesi pidi
antikehaga.
15 sekundit pärast ristsidumist fosfolipiidide metüleerimine, Ca²
kanalite teke
120 sek arahidoonhappe süntees, prostaglandinid, leukotrieenid
mikrofilamentide kontraktsioon
cAMP süntees (max 60sek)
PIP-fofatidüülinositool- 4,5bifosfonaat
DAG-diatsüülglütserool
IP3-inositooltrifosfaat
PC-fosfatidüülkoliin
PS –fosfatidüülseriin
PMT- fosfolipiid metüültransferaas I ja II
Degranulatsioon
Allergeeni poolt retseptorite ristsidumine
Anafülotoksiinid (C3a,C4a,C5a) – võivad samuti nuumrakkduele
basofiilidele mõjuda, et tekib degranulatsioon.
Mürgid
Fosforüleerimine – degranulatsiooni käigus toimuvad. Osalevad:
Proteiin türosiinkinaasid
Fosfolipaas C
Graanulites:
Palju – HISTAMIINI (närilistel serotoniin ), allergiaravimite
proovimisel hiir ja rott ei sobi, sest neil käib asi üle
serotoniini ja retseptorite vahekord on ka teine. Mesilasesuts on ka
siin. Histamiini vabanemise inhibeerimine- käitumisreakstioonide
mõjutamine, sest histamiin on ajus oluline.
10% graanulitest
Retseptorid H1 , H2, H3
H1-enamik bioloogilisi efekte
H2 –tagasiside,supressioon
PROSTAGLANDIINID JA LEUKOTRIEENID, LEUKOTRIEENID 1000X AKTIIVSEMAD
TSÜTOKIINID – IL-4,IL-5,IL-6,TNF
IgE vahendatud ülitunlikkuse jaoks ei ole universaalset ravimit,
sest reaktsioon liigub mitmeid eir radasid pidi.
Kui me leiame, mis kokreetsele inimesele allergiat põhjustab, saame
hakata teda immuniseerima nõrkade kogustega, siis hakkab rohkem IgG
tüüpi antikehasid kui IgE tüüpi. Aksutatakse tahkeid osakesi.
Mida rohkem interferoon gammat seda vähem IgE-d.
Tüüp II ülitundlikkus – antikehadega(IgG ,IgM ) seotud
ülitundlikkus
ADCC-antibody dependent cell-mediated cytotoxity
Antikeha kui opsoniin
Fagotsüüdid Fc või C3b retseptoriga hävitavad märklaud-raku
Transfusioonreaktsioonid
Veregrupid ABO,Rh,Kell,Kidd,MNSs,P,Lu,Duffy
Reesuskonflikt
Rh antigeenid 1940 Landsteiner ja Wiener
DdCcEe Rh negatiivne -dd
Reesuskonflikt
Teise raseduse ajal hakkab esimese raseduse ajal tekkinud antikehad
teise loote verd möllutama. Rhogam – ravim, atksitab mälurakkude
teket. Inimese IgG tüüpi reesuspos vastaste antikehdega.
Kui AB0 süsteemi järgi, siis kiirelt lüüsitakse vererakk, mis ema
kehassejõuab. Kui on sama, siis on reesuskonflikti tekke võimalus
suurem.
Vastsündinute reesuskonflikt –ema reesus negatiivne, kuid laps
reesus positiivne sagedamini siis, kui vastsündinu ja ema ABO
veregrupp on identsed-miks?
Tüüp III ülitundlikkus. Immuunkompleksidega sotud
ülitundlikkus.
Kui komplement ei tööta õieti. Immuunkomplekse ei koristata ära
ja nad võivad hakata häda tegea neerudes, liigestes, veresoontes.
Neutrofiilid kohale ja organismi enda reakstisoonid põhjustavad seal
kahjustusi. Võõra veregrupiga vere ülekanne nt, seeurmpalavik.
Mürkmaod – antisuurumi kasutamine aitab siduda. Esimesel korral
võib kasutada 8seerum saadaks nt hobuse käest) siis tekib
seerumpalavik teisel korral.
Põhjuseks immuunkomplekside kogunemine: neerud, veresooned , liigesed
jne
Komplemendi süsteemi aktivatsioon
C3a,C4a,C5a-anafülotoksiinid,kemotaktilised faktorid
- Neutrofiilide proteaasid
- Bakterite spoorid ,seened
- “Seerum palavik” - võõra liigi antikehad
- Autoimmuunhaigused, ravimid , nakkushaigused
Tüüp IV ülitundlikkus. Viivitusega ülitundlikkus
DTH-Delayed-Type Hypersensitivity.
Peab olema toimnud kontaks antigeeniga, sellel järgnenud T rakkude
klonaalne paljunemine, kui hiljem kontaks selle antigeeniga toimub,
siis Trakkude kaudu sekreteeritakse tsütokiine, mis aktiveeriuvad
amkrofaagid, mis omakorda tekitavad mitmeid reakstioone. Siia alla
lähevad libaallergiad - kontaktdermatiidid nt nikkel, formaliin ,
mürgised ühendid.
1890 Robert Koch - tuberkuliini reaktsioon
1.kontakt antigeeniga-T helper rakkude klonaalne paljunemine + makrofaagid ( MHCII)
2.kontakt antigeeniga -tsütokiinid
- makrofaagide aktiveerumine
tuberkuliini test 48-72 tundi 80-90% makrofaagid
Kontaktdermatiidid-Ni,formaliin,detergendid,mürgid
DTH mehhanismiga haigusi:
1. tüüpi diabeet
reumatroidartriit
multiskleroos
tsöliaaklia
tuberkuloos ja leepra – tuberkuloos saab juhtuda, sest bakterid
elavad happelises lüsosoomis, leepra ega tuberkuloositekitaja ei
sünteesi toksilisi ühendeid, ei suuda tappa organism ja tekitaba
organismile endale suuri kahjustusi, lümfotsüütide kaudu juhtitud
ülitundlikkus reakstioone
Niklist põhjustatud ülitundlikkus
toll4 retseptor seotud. Retseptorid ristseotakse nikli poolt ja üle
NfkappaB käivitatakse ülitundlikkusreaktsioonid.
Mürgitamm Ameerikas.
Ülitundlikkusreakstioon pentadeltakatehhool, seostub vere
albumiiniga ja kompleksis esitletakse lümfotsüütidele.
Retseptorspetsiifiliste autoantikehade poolt põhjustatud
ülitundlikkus
mõnedes käsitlustes klass V ülitundlikkus.
8. loeng
Rakuline immuunvastus
Märklauaks nakatunud
ja ebanormaalsed rakud :
viirused
rakusisesed
bakterid
kasvaja rakud
Efektor rakud - rakud mis osalevad
teiset oranismi rakkude likvideerimisel
antigeenspetsiifilised
CD8+
tsütotoksilised T lümfotsüüdid (TCL)
CD4+helper
T lümfotsüüdid (DTH)
mittespetsiifilised
NK
makrofaagid,neutrofiilid,eosinofiilid
Aktiivsus sõltub tsütokiinide efektiivsest lokaalsest
kontsentratsioonist, signaalmolekulide. Kas reaktsioon toimib kas on
aeglane või väga tugev.
Mittespetsiifilised abirakud võivad kasutada
antikehi “retseptormehhanismina”– Fc ja komplemendi
retseptoreid.
CTL, tsütotoksilised T rakud - põhiliselt CD8+,vähemal määral
CD4+T lümfotsüüdid
Praktiliselt kõik organismi rakud ekspresseerivad klass I MHC
molekule - st.võivad olla märklauad
Naiivsetest T lümfotsüütidest “tapjateni” kolm astet
1.
peab olema antigeenspetsiifiline signaal- TCR+ peptiid MHC I
märklaudraku poolt esitletud
2. CD28 + B7 interaktsioon
3.IL-2
seostumine kõrge affiinsusega IL-2 retseptorile, tavaliselt on
pinnal ainult madala aktiivsusega retseptor, aktivatsioon viib sinna,
et on olemas kõrge aktiivsusega
Mis rakkudega aktivatsioonikäigus juhtud – graanulid sünteesitakse
rakku, mis on tapsmiseks täis tsütotoksilisi ühendeid. Toimub
spetsiifiliste rakkude klonaalne proliferatsioon.
CTL tapmise mehhanism
1.seostumine
märklaud rakuga
2.membraanne
rünnak
3.lahknemine
märklauast ja selle häving
Ühendid mida kasutatakse: peab teadma
Perforiin
- 65kD pooride(aukude) 5-20 nm tekitamine, nagu komplemendi
reakstioonis tekkinud auk, seal aga tekkes osalevad mitmed eri
valgud.
+
Ca ioonid - polümeriseerumine
sarnane
komplemendi C9 faktoriga
granzymes
(fragmentiinid) seriin proteaasid, koos perforiiniga rakku
graanulites
1.läbi
pooride
2. mannoos 6 fosfaatretseptor - initsieerib DNA tükeldamise 200bp
oligomeerideks,
Mõlemad rajad viivad apoptoosini.
Tapjarakud ei kasuta ainult üht tapmismehhnismi. Mitu korraga.
CTL võivad tappa ka Fas ligandi (FasL) abil, seostudes märklaua Fas retseptorile. Viib samuti
apoptoosini. Fas ligandilt läheb signaal kaspaaside rajale.
Granzymes
viib ka läbi kaspaaside apoptoosile. Caspase
(cysteine aspartate protease).
Kiiresti toimub?
Interferoonid , siis NK rakkude maksimum ja viiruse tiiter on
maksimaalne viiendal päeval ja viiru spetsiifiliste CTLide maksimum
saabub pärast seda.
NK -(Natural Killer)
5-10% tsrkuleerivatest lümfotsüütidest, suur osa võib oll apaigal
lümfisõlmedes või organites
CD2 ( osa IL-2 retseptorist)
CD 16 – olemuselt IgG tüüpi komplekside püüdmiseks Fc retseptor
Fas L
Perforiin ja “granzymes”
CD94/NKG2- C tüüpi lektiin-inhibiitor retseptor
AR aktivatsiooni retseptor
Kui viiruse poolt nakatanud rakk ei näita mida tal sees tehakse, ei
päästa see midagi. NK rakule on kahtlane, see rakk läheb
hävitamisele.
Tapmismehhnismi võib käivitada ka opsoniseeritud rakk, millele on
kinnitunud mingi antikeha.
Märklaudrakkude tapmisega saavad hakkama veel peale CTLide (kes on
kiireimad ja efektiivseimad)
Neutrofiilif, eosinofiilid, NK
rakud, makrofaagid. Kasutavad
opsoniseeritud rakke – alati?.
Mehhanismie mida kasutavad on palju.
Antikehade poolt juhitud tsütotoksilisus reaktsioonid – CTLid
kasutasid spetsiifilisi retseptoreid, siis NK rakkkudel on ühek
retseptroriks antikeha.
Makrofaagide puhul oluline – millistes kudedes millised on. Ajus
mikrogliia, maksas kuplfleri vms rakud, nahas kopsus jne on eri
rakud.
Leukotsüütide migratsioon ja põletiku reaktsioon
Kuidas lekuotsüüdid kudedesse migreeruvad ja toimub seal
põletikureakstioon. Ka makrofaagid jne. Ei lähe lihtsalt läbi veresoone seina. Nii endoteeli kui läbivate rakkude vaheline
interakstioon. Teatud kohad veresoontes kus see toimuda saab, veenulid .
Protsessi alguses toimub raku pöörlev liikumine veresoone seinal ,
järjestikuline retseptorite seonumine.
Retseptorid, mis interakteeruvad omavahel
Selektiinid ja mutsiinid.
Ja need teised
Vaskulaarne endoteel-”teejuht”
(Extravasation)-
Ekstravasatsioon-rakkude
minek läbi veresoone seina
Rakkude adhesiooni molekulid –CAM(cell-adhesion molecules)
selektiinid-glükoproteiinide perekond-seostuvad
sialüleeritud mutsiinidega (L-leukotsüütidel,E –ja P- selektiin
endoteeli rakkudel)
mutsiinid- seriini ja treoniini rikkad ,väga, glükosüleeritud valgud (CD34,GlyCAM-1)
integriinid - heterodimeersed valgud, esinevad nt lümfotsüütides
ICAMS –immuunoglobuliinide superperekonda kuuluvad adhesiooni
molekulid
Ülejäänud on monomeerid , integriinid on dimeerid.
Kui kõik seostumised on ära olnud lastakse rakk läbi.
Adhesioonimolekulide spekter määrab ära, kus on selle raku kodu,
kas nahas limaskestades maksas jne.
Postkapillaarsed veenulid (HEV-high-endotelial venules)
1sekundis ühes lümfisõlmes 14x10³ lümfotsüüti läheb läbi
poskapillaarsete veenulite (HEV)
Vaskulaarsed adressiinid--Homing receptors, naiivsed lümfotsüüdid
liiguvad erinevalt võrreldes mälu-ja efektor lümfotsüütidega.
Vaskulaarne – juhtkudedega seotud
Kemokiinid (kemotaktilised tsütokiinid)
90-130 aminohapet,väikesed valgud
Osalevad ka mujal kui immuunreaktsioonides - aju,süda jne.
Üle 50 kemokiini ja 15 erineva retseptori on juba leitud
Retseptorid G-valgu seoselised
Selgroogsetel (+mõned viirused ja bakterid) -selgrootutel pole
leitud
Kemokiiinide retseptorite ekspresseerumine sõltub raku tüübist ja
tema aktivatsiooniseisunudst (akt v mitte).
Kemokiini gradiendi suunas liikumine – kemotaksis on põhiline mis
käivitub.
Kui on kahjustus toimunud (viirslik nakkus, bakteriaalne)
fagotsüteerivad jne rakud peavad sinna jõudma, suunavad sinna
komplemendi anafülotoksiinide ja kemokiinide gradient.
Põletiku reaksioonis osalevad teised mediaatorid
- kiniin süsteem - kallikreniin bradükiniin
- hüübimiskaskaad - fibriin
- fibrinolüütiline süsteem - plasmiin
- komplemendi süsteem
- prostaglandiinid,tromboksaanid
- tsütokiinid
Põletikureakstioonides osalevad kaskaadid.
Kiniin –kallikreiin süsteem
Inimesel 15 erinevat geeni, tüüpiliselt hulk erinevaid splaisingu
variante (eri spetsiifikaga)- olemuselt seriinproteaasid
Lõikavad kiniinie, üks levinuimad on – Bradükiniin, koosneb: Arg
- Pro - Pro - Gly - Phe - Ser - Pro - Phe – Arg. Bradükiniini
retseptor B1 avaldub ainult kudede kahjustuse korral, sellega kaasneb
krooniline valu.
Kiniinid määravad silelihaste pinge, verevarustusega seotud midagi.
Näiteid kallikreiinidest
Kallikreiin 1- vasoaktiivset (lisaks lüsiin - veel) Lys-bradükiniini
Kallikreiin 3 - prostata spetsiif.antigeen (PSA), kasutatakse
prostata vähi diagnostikas
Kallikrein 5 - epidermises (Desquamation), osaleb naha kestendumises,
et naharakud teatud arvu päevade pärast vabanevad, kui valesti
sellega siis kaasnevad mingid haigused ihtüoosi jne.
Kallikreiin 6-lõikab amüloid eellas valku, alzhaimer.
Kallikrein 7-mutatsioonide korra väga kuiv nahk
Kallikreiin 13-marker rinnavähi diagn.
Kallikreniinide aktivatsiuoon on immuunsüsteemi poolt juhtud tihti.
Nt endoteeli kahjustused toovad kaasa kalikreniinide sünteesi, valu
on retseptoritega seonudmisel, hüübimine. Füsioloogilised
kaskaadid on seotud.
Prostaglandiinide süntees,
leukotrieenide süntees on ka millegiga
seotud.
9. loeng
Immuunpuudulikkus
Haigused, kus immuunvastus ei funktsioneeri normaalselt.
Kaasasündinud -
immuunvastuses osalevate geenide defektid . Geenidefektid OMIM
andmebaasis olemas.
Omandatud
imuunpuudulikkus - haigustest põhjustatud, nakkushaigused nt
malaaria, HIV, HTLV-1,leetrid, aga ka halvaloomulised kasvajad, nälg,
mürgid, ravimid (doosid). Äärmusesse viidud organismid –
tippsportlased jäävad kergesti nakkushaigustesse.
Steriilne põletik – mehhaanilise vigastuse tagajärjel on kõrgem
interleukiinide, prostaglandiinide konts. Siis ei rasestu, sest
emakasein pole valmis. Kui võtad aspiriini, siis inhibeeritakse prostaglandiin süntetaase – saab seostuda vilj munarakk.
Diferentseerumisrada.
Müeloidsest reast mis rakud ja mis lümfoidsestreast. NK rakud ja
osa dendriitrakud tulevad kust reast.
Inimestel DiGeorge sündroom – tüümus puudub, võiks eksamil
teada. Karvade geen on lähestikku tüümuseta fenotüüpi määrava
geeniga. Tüümust ei arene, ei ole kohta kus T-lümfotsüüdid
saaksid areneda. Kaasasündinud immuunsussüsteem on normaalne (Tol
retseptorid), kõik mis on seotud rakulise imuunsusega – kasvajad,
viirused – vastuvõtlikud. Antikeha midagi on.
Leukotsüüdil puudvad teatud adhesioonimolekulid – ei saa
veenulist läbi, immuunpuudulikkus.
Ei suudeta märklauda fagotsüterida, lõhustada neutrofiilid.
Krooniline granulomatoosne haigus.
SCID (SEVERE COMBINED IMMUNODEFICIENCY)
Uuriti 141 PATSIENDIST kus on defektid, suur osa 64 tükki IL-2
retseptoridefektiga (aga ka IL-2,4,7,9,15), 9 inimesel oli JAK -3 defekt (paljud retseptorite signaaliülekande rajas, et signaal saaks
minna tuuma esimesed astmed), 22 inimesel ADA (adenosiindeaminaasi)
puudulikkus, ensüüm . Kasutai esimest korda geeniteraapias.
B lümfotsüütide küpsemisel suht levinud hypogammaglobulineemia - midagi on puudu ei sünteesita. Võib olla ka et ei suudeta teha
mingit klassi IgG-d nt IgG kaks klass vms.
Tabel, mis tüüpi pärandamisega on tegemist.
Võiks teada, et rida sündroome haigusi on X-liitelised, seda võiks
teada. Kust ja kuidas haigusi leida. Mitte ainult defektid
retseptorites, aga just signaalrajas ka JAK kinaasis jne, omavad
olulist rolli kas organism suudab õieti reageerida või mitte.
Mõned mutatsioonid on etnilistele rühmadele, piirkondadele omased .
Hiireliinid, kellel on mutatsioonid või puudused CTL surma
radades
Granzüümi puudus- granzüümiga viiakse märklaudrakud apoptoosi –
puudulikkus põhjustab viivitust apoptoosis märklaudrakkudes.
Probleemide uurimseks on mudelid hiirtel (inimestel on rake mudeleid
teha).
Inimese X kromosoom
Propedin puudulikkus – komplemendi aktivatsioonis, alternatiivses
rajas.
B ja T lümfotsüütdide vahelised retseptorid
Immuunpuudulikkus põhjused raku tasemel. JAK kinaasid, neid on
erinevaid. Lümfotsüütides võivad olla defektsed rekombinatsiooni
aktiveerivad geenid, RAK1 RAK2, kui need ei fn sisi
immuunpuudulikkus. MHC moelkulide ekspressiooniga seotud molekulid
defektsed nt.
Kui võtame ette ühe inidviidid (peale tõsiste juhtumite, kui
tõesti üks ei tööta üldse) siis eri indiviididel on eri molekuli
varaindid, polümorfsus, moelkulid töötavad eri efektiivsusega -
haigestuvad erinevalt, seal see kala peidus on.
Omandatud imuunpuudulikkus
Retroviirused –lentiviiruste perekond, visna viirus-lambad,veised,
SIV-simian imm.puudulikkuse viirus, HIV-1 (human immunodeficiency virus ), HIV-2, HTLV-1 (human T cell lymphothrophic virus).
Lähme HIVi ja AIDSi juurde.
AIDS – omandatud imuunpuudulikkus.
Kandjad ja kellel on välja arenenud imuunpuudulikkus tuleks vahet
teha. Kõige hullem on seis Sub Sahaara Aafrikas.
Võrdleme genotüüpe
Eri subtüübid Hiv-1'l , Hiv-2'l, Siv'il. Arenenud eri subklassideks
levinumi on HIV M alagrupp.
Üleminek ahvilt inimesele 1673 umbes – erinevate genoomide
võrdlusega saab öelda. M grupi teke 1920, umbes. Ei ole kandunud
ainult 1 korda üle vaid ilmselt 3 korda.
Pilt Põhja Ameerika kohta
Kuidas inimesed surevad.
HIV graafik alla – haakti kasutuma erinevaid ravimeid, ei ravinud
terveks, aga immuunpuudulikkuse etekt sai ajas edasi nihutada.
Viiruse pilt
gp – glükosüleeritud valk.
MHC valgud viiruse pinnal – miks seal ja kust ta sinna sai. Alati
ei pruugi olla, aga väljudes rakust siis võtavad pinnalt kaasa peremees raku MHC moelkule, aitab immuunsüsteemi eest kaitsta. Seda
võib näha ka mitmete teiste viiruste puhul.
Glükoproteiinid on vajalikud seondumiseks. Nende vastu tekivad
antikehad, siis pole suurt abi, sest makrofaage kes neid söööma
tulevad suudavad nakatada.
Hävitab ja põhikoduna peab T-helper rakke.
Immuunsüsteemi mälu hävib, rakuline imuunvastus kukub kokku. Arv
väheneb – kriitiline piir. Seostub CD4 molekulile, vaj aveel
abiretseptroid, kemokiini retseptorid, kas lümfokas või makrofaag –
eri retseptorid. Kemokiini retseptoriga tuleb mängu inimese
polümorfism. Üks jupp on puudu osadel inidviididel kui on
homosügootne – jääb sisenemine pidama . Ei suuda nakatada.
Kaasasündinud immuusususe valk
APOBEC - rakud
panevad kaasa selle viirusele, tekivad mutatsioonid. HIV viirusel selle protsessi blokeerivad vif geen.
Mehhnism kuidas APOBEC toimib. Miks viirus ei saa normaalselt
funktsioneerida.
Igal rakul on oma kemokiiinide spekter. Kemokiini reteptorite mille
korral ei nakatu HIVi seostatakse viikingite ekspansiooniga.
SDF-1, katab kemokiine miks?
AIDSi haigetel salmonella võib olla surmav ja toksikoos kahjsutada
aju.
Ravimid, mis kontrolli all hoiavad viiruse hulka
Rohkem T-lümfotsüüte alles kauem.
- Integraasi inhibiitorid.
- Proteaasi inhibiitorid.
- Pöördtranskriptaasi inhibiitorid.
10. loeng
KASVAJAD JA IMMUUNVASTUS
VÄHK (PAHALOOMULINE KASVAJA)--RAKUD, MIS ENAM EI DIFERENTSEERU JA
POOLDU TAVAPÄRASELT EGA ALLU TUUMORSUPRESSOR NING PROTOONKOGEENIDE
KONTROLLILE.
KARTSINOOM - EPITELIAALNE
SARKOOM - MESENHÜMAALNE
LÜMFOOM - LÜMFOIDNE
LEUKEEMIA - HEMATOPOEETILINE
MELANOOM- NEUROEKTODERMAALNE
SEMINOOM - TÜVIRAKULINE
GLIOOM- GLIIA RAKKUDE KASVAJA
Lisaks SEGATÜÜPI
Kõikidel kasvajatel on KLONAALNE PÄRITOLU, HETEROGEENSUSE
(EBASTABIILSUSE) TEKE
5X10 KAHEKSANDAS ASTMES RAKKU- 1 GRAMM KASVAJAT
2-20 AASTAT 1st MUUTUNUD RAKUST 1grammine KASVAJA
Kahekordistumine: 50-100 PÄEVA
Noortel ja vanadel indiviididel on erinevad kasvajate spektrid .
Lapseea kasvajad on seotud konkreetse fenotüübiga, võivad arenema
hakata juba looteeas. On valdavalt agressiivsed, kiire kasvaga,
alluvad ravilehalvasti.
Klassikaline kasvaja areng
Üks rakk – klonaalne areng. Kui pole läbinud basaalmembraani see
mass, siis võib immuunsussüsteem asjale piiri panna.
Metastaseerumine – lümfiteedesse, veresoontesse jõudmine.-
Kasutavad adhesioonimolekule. Ravi keeruline.
Geenid, mis osalevad kasvajate prostessil
Geenid, mis indutseerivad rakkude jagunemist: Muutused geenides, mis
on seoutud kasvufaktoritega, raku signaaliülekande molekulidega
seotud muutused. TF muutused. Kaob kontroll rakkude paljunemise üle.
Tuumorsupressorgeenid.
Apoptoosi reguleerijad.
Prote onkogeenide käivitumine
Saame eristada viiruslikku onkogeneesi.
Rakulist onkogeneesi – mutageenid, radiatsionid, geneetilised
eelsoodumused.
Krooniline müelodine leukeemia
Kasvaja rakkudes saame tuvastada tsütogeneetiliselt kromosoomi, mida
seal normaalselt ei ole. Nn Philadelphia kromosoomi. Suur osa
materjali on alles, aga seotud 9 kr-ga. 9 kromosoomis, saavad kokku
valed geenid ja tekib uus valk, millel on funktsionaalne aktiivsus
bcr-abl, suudab seostuda IL-3 retseptorile, alamühikule. bcr-abl
valk interakteerub interleukiin 3 beta(c) retseptori alaühikuga.
Aktiveeritakse valke, mida ei oleks vaja:
...genoomi instabiilsus
...aktiveerib rakutsüklit kontrollivaid valke
...inhibeerib DNA reparatsiooni mehanisme
...granülotsüütide ja g.blastide hulga tõus
Vereproovisnäeme granulötsüütide ja nende eellasrakkude kasvu.
Et leida sobivaid inhibiitoreid võidakse skriinida High-throughput
screening (HTS) -100 000 ühendit päevas.
Ravim – hübriide
kromosoomilt tekkinud valgust põhjustatud leukeemiale
Imatinib - müüakse Glivec® nime all, imatinib mesilate on päris
nimi, suudab blokeerida bcl-abl aktiivtsentri, kinaasse koha ja
vähirakud ei saa enam kontrollimatult paljuneda.
Burkitti lümfoom
Translokatsioon 8 ja 14 kr vahel. Immunoglobuliinide raskete ahelate
geenid ja c-myc.
Et asi jõuaks kasvajani, on vaja läbida pikk tee.
Immuunsussüsteemi roll kasvajate puhul
Mismoodi immuunsussüteem kontrollib ja kõrvaldab rakke, mis võiks
aluse anda kasvajatele.
Oncofetal geen, valk – geen, mid normaalselt ekspresseerub
looterakkudes, aga nüüd kasvajas.
Katse dendriit+ kasvaja
Kasvatatakse tassil, et tsütokiinid, ei saaks maha suruda
imuunvastust ja pärast sisse ja immuunsussüsteem suudab jälle
võidelda.
Vaja tsütolüütilisi T rakke ja T helpereid, et kasvajaid hävitada.
Immuunpuudulikkusega inimestel on kasvajaid rohkem. Enamike
klonaalselt tekkivate kasvataega saab immuunsüsteem hakkama.
Kui pinnal näidatatakse MHC-d aga väga vähe, on võimalik selliste
rakkude pääesemine imm süst poolt.
Monoklonaalsed antikehad
Ei ole igapäevases medistsiinipraktikas
Mingisuugsed tunnevad ära kasvajaid. Komplemendi käivitajad, mis
sisse viiakse, neid on palju, mida kasutatakse.
Miks kasvajad ikka tekivad siis? Sest lõpuks peab millesegi ära
usrema ja lapsi jääb rohkem ellu, kasvajatel võimalus tekkida 20 a
jooksul.
Immuunvastus kudede siirdamisel
Siirdamine -/transplantatsioon/transfusioon(kui pole tahke ülekanne – vereülekanne nt) — rakkude, kudede, organite (graft – ülekantav
materjal, kõik eelenvad) ülekanne
doonor - võetakse
retsipient / host / - antakse
lokalisatsioon –ortotoopne /heterotoopne (sõrmest varavas) –
sama või erinev koht
autoloogne-sama organismi piires nahaülekanne
süngeenne-geneetiliselt identne,
allogeenne-sama liigi piires
ksenogeenne-erinevate liikide vahel
tüvirakud
Vereülekanne kõige levinum. Järgmisena sarvkest , siirdamisedukus
väga suur, soovitav jälgida kokkusobivust AB0 veregrupi osas.
Sarvkestas pole veresooni ja lümfotsüüdid, ke ssaakks kontrollida
kas on oma või võõrad rakud ei pääse ligi üldse. Neerud ka suht
popid.
Miks nahka ei saa kanda teisele üle? Nahas on väga palju
dentriitrakke, hea verevarustus . Neeru või maksa veresooned on
võimalik läbipesta – oma rakud välja lümfotsüüdid. Nahast
neid kätte ei saa – peab olema autoloogne.
Mis juhtub kui nahka istutada teise kohta. Võõra nahatüki puhul –
kui erinevad MHC molekulid, siis revaskularsiatsiooni käigus jõuavad
kohale imm rakud ja saab selgeks et on võõras materjal.
Hävitatakse. Esimesel korral selline reakstiion. Uuesti proovides on
tekkinud imm mälu ja helperid ja antikehad on olemas ja protsess -
koe nekrotseerumine hakkab varem.
Koesobivuskompleks (MHC)
H-2 hiirel, HLA inimesel – palju polumorfseid ja mittepolümorfseid
lookusi.
Segatud lümfotsüütide reaktsioon?
Immuunosupressorid - Tsüklosporiin A-inhibeerib IL-2 retseptori,
kõik T lümfotsüütide aktivatsioon oli seotud IL2-ga retseptor
blokeerida – siis immuunvastus kinni panna, FK506,
rapamütsiin-blokeerib diferents. G1 faasi
Mõningatel kohtadel on
immunoloogiline priviligeritus aju, silm, emakas/monomorfne HLA –G,
kui üldse pole siis NK hävitaksid. Loote
immuunsus
Graft Versus Host haigus – ( akuutne , krooniline) –
krooniline on hea veel, võib lõppeda fataalselt. Eksamis. Suur oht
luuüdi ja naha siirdamisel. Luuüdi siirdamisel doonori omad
häviattakse, kui MHC-d pole õiged, siis hakkab graft hävitama
organismi, sest MHC-d on valed. Ülekantud materjal hävitab
organimi, kuhu ta üle kanti. Sp proovitakse lümfotsüüdid ikka
neerudest maksadest välja pesta. Aitab immuunosupressorite
kasutamine.
Krooniline – nt neeruülekande puhul.
Südame ja neeru ülekantudel on rohke kasvajaid, sest peavad
kasutama imuunosupressante.
Hla on üks kõige polümorfsemaid üldse inimesel lookusesüsteeme.













































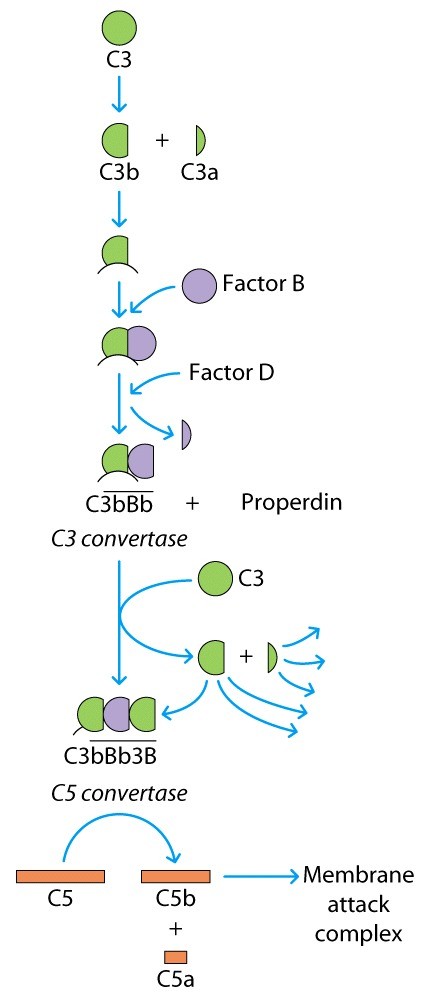




Kõik kommentaarid