RAKUBIOLOOGIA Prokarüoot Eukarüoot Raku suurus1-10 μm
5-100 μm
Organellid Puuduvad või vähe
Tuum,
mitokonder ,
kloroplast TuumPuudub
Esineb
Rakumembraan Esineb (ei sisalda steroole,
vaid hepanoide)
Esineb
Mitokondrid Puuduvad (oksüdeerumist
katalüüsivad
ensüümid seotud rakumembraaniga)
Esineb
Ribosoomid Esinevad (70S)
Esinevad (S80)
Tsütoskelett Puudub
Esineb
Mitoos , meioosPuudub
Esineb
DNA struktuurRõngas, (
kromosoom ja
plasmiidid) tsütoplasmas
Lineaarne, erinevad
kromosoomid ,
histoonid ,
paiknevad tuumas
RNA ja valkSüntees samas kohas
RNA tuumas,
valgud tsütosoolis
MetabolismAnaeroobne+
aeroobne Aeroobne
Rakuline organiseeritusPeamiselt
üherakuline Peamiselt
hulkraksed - Esimesed prokarüootsed organismid tekkisid ~ 3 -3,5 miljardit aastat tagasi
- Esimesed eukarüootsed organismid tekkisid ~ 1-1,5 miljardit aastat tagasi
- Sümbiontsed bakterid , mis on seotud õhulämmastiku assimileerimisegaon näiteks tsüanobakterid ja Rhizobium
Millised
tunnused näitavad et mitokondrid ja kloroplastid on tekkinud
endosümbioosi teel?Kahekordne
membraan sisemise membraani koostis on sarnane
bakteriraku membraani
koostisega.
Paljunevad
jagunemise teel, mitoos puudub.
DNA
on rõngaskromosoomi kujul nagu bakteritel, histoonid puuduvad.
Operonide
esinemine DNAs.
Esinevad
ribosoomid, mis ehituselt ja koostiselt sarnanevad bakteri
ribosoomidega.
Membraanid ÜlesandedAinete
transport
Metaboolsete jääkide eraldamine rakust
Tsütoplasma pH säilitamine
Tsütoplasmas teatud osmootse jõu
hoidmine
Organellides spetsiifiliste tingimuste hoidmine
Membraanid
koosnevad lipiidide
kaksikkihist, mida
läbistavad integraalsed valgud.
Lipiidid takistavad rakus esinevatel
polaarsetel ühenditel kiiresti rakust välja difundeeruda.
Raku
membraani paksus 6~10 nm
Membraanide
valgudIntegraalsed
e transmembraansed-
läbivad ühe või mõlemad lipiidsed kaksikkihid
Perifeersed-
paiknevad lipiidse kaksikkihi tsütoplasma või väliskeskonna
poolsel pinnal.
Väliskeskonna
poolsed valgud osalevad
rakk -rakk äratundmises ja signaalide
ülekandmises, taimedel ka tselluloosi sünteesiks
rakuseinas.
Tsütoplasma poolsed osalevad tsütoskeleti sidumises
valkudega, signaalide ülekandes.
Integraalsed osalevad
kanalite moodustamises ja ainete
transpordis .
Integraalsete membraanide valgud omavad vähemalt ühte segmenti, mis koosneb
hüdrofoobsest või laenguta aminohapetest ja on
seetõttuhüdrofoobsete vastasmõjudega kinnitatud lipiidide kihti.
Hüdrofoobne piirkond on ümbritsetud + laenguga aminohappega, et
takistada piki membraani libisemist (+ laenguga aminohappe seotud
fosfolipiidi – laenguga peaga)
Perifeersed
valgud kinnituvad
vesinik - või
ioonsete sidemetega integraalsete
valkude külge aga ka lipiidide
polaarsete peade külge.
Membraanivalkude
funktsioone:Ainete
transport rakkude vahel
Membraani struktuuri hoidmine
Signaalide
vastuvõtt keskkonnast ja teistelt rakkudelt
Rakkude seostamine
rakuvälise maatriksiga ja teiste rakkudega
Tsütoskeleti valkude
seostumine membraaniga.
Membraanide
lipiididTrans-rasvad
on küllastumata
rasvad, mille rasvhapete jääkides esinevad kaksiksidemed on
trans
asendis ehk E-
isomeerid .
Fosfolipiidid
on membraani põhiliseks lipiidseks komponendiks ja neil on
struktuuris
polaarne pea ja kaks hüdrofoobset saba.
Steroidid
(kollesterool
puudub prokarüoodi plasmamembraanis.)
Sahhariidid
Võivad
olla seotud nii lipiidide kui valkudega
Olulised rakk-rakk
äratundmisreaktsioonides
Membraanikomponendid:Lateraalne difusioon on ühe
membraani kihi piires toimuv aktiivne lipiidide molekulide
difusiooniline liikumine.
Membraanide
dünaamilisust (voolavust) mõjutavad:Lipiidide
küllastatus - küllastamata
rasvhapped muudavad paindlikumaks (mida
rohkem seda voolavam)
Kolesterool membraani koostises (alandab
sulamispunkti )
Temperatuur
Membraani
sulamistemperatuuriks
nimetatakse temperatuuri, mille juures toimub faaside üleminek (St.
membraan läheb liposoomide lahuse kuumutamisel geelisarnasest
struktuuri faasist üle mobiilsemasse vedelasse faasi).
Hübridoom
on
antikeha sünteesiva b-lümfotsüüdi ja müeloomiraku
hübriid ,
mille abil saab toota
suurtes
kogustes antigeeni spetsiifilisi monokloonseid antikehi.Säilinud
mõlemad eellasraku
tunnused.
1)
katseloom immuniseeritakse vajaliku
antigeeniga 2)
immuniseeritud katselooma
viidud antigeenile vastavaid antikehasid
tootvad b-
lümfotsüüdid liidetakse
müeloomirakkudega
3)
selektiivsöötmel eraldatakse hübridiseerunud
rakud 4)
hübridoomi kasvatatakse masskultuuris või katseloomas
5)
antikeha eraldamine, puhastamine ja kasutamine
ELEKTROKEEMILINE POTENSIAAL VIHIKUST!Aktiivse
transpordiga,
mis vajab täiendavat (ATP) energiat on tegemist kui ioonide (ainete)
liikumine toimub
piirkonnast kus ainete elektrokeemiline potensiaal
on madalam, piirkonda, kus aine elektrokeemiline potensiaal on kõrgem
(nt ATP-
aasid ).
Passiivse
transpordi
korral pole vaja täiendavat (ATP) energiat, sest ainete liikumine
toimub kõrgema elektrokeemilise potensiaaliga piirkonnast madalama
potensiaali suunas (nt
ioonkanalid ).
1.Difusioon läbi lipiidide
(Peamiselt
hüdrofoobsed ja väikese molekulmassiga
polaarsed(
etanool ) ühendid, gaasid)
2.
Difusioon valkude vahendusel
Membraanipotentsiaal
- ioonide kontsentratsioonide erinevus/ elektrokeemiliste
potensiaalide erinevus raku sise- ja väliskeskkonna vahel.
Reeglite
järgi võrreldakse tsütosooli väliskeskonnaga
Mõõdetakse
voltides. Membraanipotensiaal on negatiivne, st tsütoplasma poolne
külg on negatiivsem väliskülje suhtes/ tsütosoolis on
negatiivsete ioonide ülekaal.
Membraanipotentsiaali
tekkimise põhjusi:Ainete
elektrokeemiliste potensiaalide erinevus rakus ja rakuvälises ruumis
Vastasnimeliselt
laetud ioonide erinev
liikumiskiirus läbi membraani - moodustub
elektriline
potensiaal
e difusioonipotensiaal.
Jätkuvalt
säiluva difusioonipotensiaali põhjuseks on nn “pumpade”
funktsioneerimine rakumembraanis .
(Pumpadeks nim membraanide transportsüsteeme, mis ainete
transpordiks kasutavad vahetult ATP energiat - osalevad aktiivses
transpordis)
Membraanipotensiaali
tekitavat transporti nimetatakse elektrogeenseks
transpordiks.
Membraanipotensiaalide
väärtused: Rakumembraan -70mV, kloroplastil -30mV, mitokondril -180
mV
NERNSTI VÕRRAND VIHIKUST
Vee
liikumine toimub
osmoosi teel.
Vesi
liigub väiksema lahuse
kontsentratsiooniga piirkonnast suuremasse
Rakud
peavad paiknema isotoonilises
lahuses, et rakk ei
paisuks või kuivaks osmoosi tõttu.
Sise-
ja välislahuse võrdse kontsentratsiooni hoidmiseks on olulised
membraanides paiknevad transpordisüsteemid (Na+/ K+ -
ATPaas )
K/Na-ATPaas
on aktiivtransport ,
mille käigus liigutatakse ATP hüdrolüüsi energia arvelt 3 Na
iooni välja ja 2 K iooni raku sisse. Oluline membraanipotentsiaali
hoidmisel, koos Na ioonidega glükoosi sisseveol rakku
sekundaaraktiivtranspordil, osmoosi kontroll.
Akvaporiin
koosneb kuuest transmembraansest alfaheeliksist,
kusjuures nii amino
kui
karboksü ots jäävad tsütosooli poole. Moodustavad kanali läbi
mille liigub kas ainult vesi või ka selles lahustunud
ained,
sõltuvalt akvaporiinist.
Reguleerivad
veevoolu
Kanalivalkude
avatust mõjutavad:Elektriline
signaal (membraanipotentsiaal), keemiline signaal. (teatud ühendite
seostumine
kanali
valguga) Lisaks ka temperatuur,
mehaaniline jõud ja
fosforüleerimine.
Sekundaaraktiivne
transport
ei kasuta otseselt
ATP energiat, et aineid transportida, vaid
elektrokeemiliste
potentsiaalide erinevust. Transportvalkudeks antipordid ja sümpordid.
Nt Glükoosi transporditakse
niimoodi rakku koos Na ioonidega.
Aktsioonipotentsiaal
on aksonite omane kiire membraani depolariseerimine ja selle järel
repolariseerimine elektrilise signaali/ impulsi ülekandel (moodustud
aksoni ja neuroni
kokkupuutekohas).
Aktsioonipotentsiaali kestvus on umbes 1-2 ms.
Sünaps on kahe neuroni
ühenduskoht, kus toimub impulsi
edasiliikumine ühelt rakult teisele.
Neurotransmitter ehk neuromediaator ehk
virgatsaine on keemiline aine,
mille abil
neuron
(
närvirakk ) edastab keemilise sünapsi kaudu närviimpulsi teisele
(närvi)rakule
Närviimpulsi
teke ja liikumineStiimuli
toimel vabanevad neurotransmitterid ja seostuvad jörgneva neuroni
pinnaga
Lokaalne neuroni membraani depolarisatsioon
Membraanipot
sõltuvate Na kanalite avanemine, Na+
sisenemine ,
aktsioonipotensiaali teke, Na kanalite sulgumine
Avanevad membraanipot sõltuvad K+ kanalid, K+ väljub,
taastub puhkeseisundi
membraanipotensiaal, K kanalid sulguvad
Membraanipotensiaalist
sõltuvatel Na kanalitel esineb refraktsiooniperiood,
mille jooksul sulgunud Na kanalid ei avane ka stiimuli toimel, seega
impulss liigub ainult ühes suuas.
+40mV
juures avanevad pingesõltuvad K+ kanalid ja K liigub välja,
tekitades kiirelt negatiivse
membraanipotentsiaali
(repolarisatsioon).
Membraani
depolariseerumine aktsioonipotensiaali toimel
on tingitud ioonkanalite positiivse tagasiside süsteemist. Kui
kuskilt sattub raku sisse Na ioone, siis need indutseerivad kanalite
avamise ja veel rohkemate Na ioonide rakku voolamise. Võivad tulla
retseptoritelt või sünapsist jne.
Kui
membraanipotentsiaal on piisavalt kõrge, siis see sulgeb Na kanalid
ja samas avab K
kanalid
ja K voolab rakust välja, toimub membraani repolariseerumine
esialgsele membraanipotentsiaali väärtusele (~ - 60 mV)
närviimpulsi tekkel, normaliseerides potentsiaali.
Kui
rakumembraan on hüperpolariseerunud,
siis membraani depolariseerumine võib toimuda
nt.
Na või Ca sissevooluga vastavatest kanalitest.
Närviimpulsside
tekkimises ja edasiliikumises osalevad transportvalgud :Na+
kanalid, K+ kanalid, Na+/K+
pump , ligandiseoselised kanalid
Närviimpulsi
edasiliikumise kiirus on 100-120 m/s
Müeliinikihti
on aksonite ümber vaja, sest suurendab kiirust, millega impulss saab
liikuda mööda närvikiudu. Signaal n-ö “hüppab” ühest
müeliinita kohast järgmisele. Käitub natuke nagu isolaator
juhtmel. Seda toodavad Schwanni rakud (perifeerses närvisüsteemis)
ja oligodendrotsüüdi(tsentraalses närvisüsteemis).
Impulsi
liikumine läbi sünapsi
Aktsioonipotensiaal
jõuab neuroni terminaalini
Neurotransmitterit
sisaldavad vesiikulid sulavad eksotsütoosi protsessis kokku
rakumembraaniga ja virgatsaine vabaneb ning difundeerub järgmise
raku pinnal paikneva retseptorini
Avanevad
ligandiseoselised kanalid, Na siseneb, membraan depolariseerub ja
tekib Ap
Ajutegevust
toetavaid signaalmolekule:Serotoniin ,
dopamiin,
endorfiinid .
Lihaste
kokkutõmbeid reguleerivaid signaalmolekuleAtsetüülkoliin,
(nor)
epinefriin (südametegevus).
Sclerosis multiplex ( hulgiskleroos ) tekkimise põhjusiMüeliini
kadumine pea- ja
seljaajust Autoantikehade tootmine müeliini
vastu või vastavate proteaaside tootmine
Transportvalkude
häiretest sõltuvaid haigusiMootorneuronite probleemid
Na+ kanalid
Tsüstiline fibroos
Cl – kanalid
Bipolaarsed häired
Na+ /K+ -ATPaas
Südametegevuse häired
Na+ /K+ -ATPaas
Resistentsus ravimite suhtes ja kemoteraapiale
ABC pump (multi-drug resistance)
Värvipimedus [H+]
gradient kui pump (rodopsiin)
ENDOMEMBRAANID
Tsütoplasmavõrgustik
= ER ja sekreetoorne rada
ER/Golgi
vahendusel sekreteeritavad valgudProteaasid ,
glükoproteiinid, proteoglükaanid, mutsiinid, fibromoduliin,
laktoferriin.
SRP
(signaali äratundja partikkel )
on
proteiin-RNA
kompleks . Eukarüootides kuus polüpeptiidi + RNA.
Prokarüootides 1 polüpeptiid +RNA.
Vajalik
valgu signaaljärjestuse äratundmiseks
Kirjeldage
signaaljärjestust valgu liikumiseks tsütosoolist ER-i, milliste
teiste valkudega signaaljärjestus komplekseerub? Kus paikneb selle
järjestuse retseptor ?N-terminaalses
otsas korduvad leutsiini jäägid ning ka lüsiini jäägid.
Signaaljärjestus komplekseerub SRP-ga, translokaatoriga. Selle
järjestuse retseptor on hüdrofoobne Met jääkidega ümbritsetud
tasku SRP P54 subühikus. Ning translokaatorid paiknevad ER-s.
Nimetage
pöördtranskriptaasi (RNA sõltuv DNA polümeraas ) osavõttu
vajavaid protsesse eukarüoodi rakusRetrotransposonite
teke, DNA liikumine mitokondro ja tuuma vahel
Nimetage
ER-is ja Golgis sekreteeritavate valkudega toimuvad modifikatsioonidÕige
konformatsiooni teke, osaline
proteolüüs , oligosahhariidse
komponendi lisamine, disulfiidsidemete teke, multimeeride teke
Kirjeldage
antikehade struktuuri. Milliste sidemete vahendusel selline struktuur
moodustub ja millises raku piirkonnas? Kuidas antikehasid kasutatakse
kindla valgu lokalisatsiooni määramiseks rakus?Koosnevad
neljast disulfiidsildadega ühendatud valguahelast. Kaht suuremat ja
omavahel identset
ahelat nimetatakse rasketeks ehk H-ahelateks
(ingl.k
heavy chains), kaht väiksemat ning samuti omavahel identset
ahelat nimetatakse kergeteks ehk L-ahelateks (ingl.k
light chains).
Moodustub tsütosoolis. Antikeha tunneb ära võõrkeha antigeeni.
Iga antikeha tipp sisaldab paratoopi (lukku), mis on spetsiifiline
antigeeni epitoobile (võtmele). See lubab neil struktuuridel
ühineda. Selle mehhanismi abil saab antikeha märkida nakatunud
rakku teistele immuunsüsteemi osadele ründamiseks või
neutraliseerib ise.
Kuidas
pannakse kokku valgu Asn jäägile seotav oligosahhariidne kompleksER
membraani tsütoplasma poolsel küljel toimub
kõigepealt kahe
N-atsetüülglükoosamiini (GlcNAc) seostumine membraanis paikneva
dolihoolfosfaadi külge. Järgneb viie mannoosi jäägi liitumine.
Seejärel toimub dolihhooli liikumine ER luumeni poolsele küljele
fosfolipiidi translokaasi e flipaasi toimel. Luumenis lisatakse
täiendavad 4 Man ja 3 Glc jääki ja kompleks Glc3Man9GlcNAc2
on moodustunud dolihhooli küljes. Glükoosi jäägid on signaaliks,
et oligosahhariid on ‘valmis’.
Miks
on sekreteeritavate valkude glükosüülimine vajalikVajalik
N-seoselist sahhariidset jääki sisaldavate valkude õige
konformatsiooni moodustumisel. Ainult õigesti voltunud valgud
liiguvad ER-st Golgi kompleksi.
Mis
määrab A, B, O vererühmad .
Vererühmad
on määratud sahhariidsete jääkide poolt vere glükolipiidides ja
glükoproteiinides erütrotsüütide ja mõnede teiste rakkude
pinnal.
Tähistage
tabelis milliste doonor ja aktseptorvererühmade vahel võib
vereülekandeid teha.Vastavasse
kohta lisada +DOONOR A
B
AB
O
AKTSEPTOR A
B
AB
O
Mida
tuleb ette võtta, et reesuskonflikti puhul (Rh
negatiivne
ema, Rh positiivne laps) vältida teise lapse sündimisel loote
kahjustamist reesusvalgu antikehadega?Nn
’reesusvaksineerimine’ – reesusnegatiivsele rasedale naisele
antakse
hiljemalt 2-3 ööpäeva pärast reesuspositiivse lapse sündi
antikehakontsentraati (anti-D-immunoglobuliini). See takitsab ema
vereringesse sattunud lapse punaliblesi ema immuniseerimast.
Millisesse raku piirkonda liikuvad valgud korjavad kokku vesiikuli kesta valgud
COPI, COP II ja klatriin. - COP I : Golgi ER (retrograadne transport)
- COP II : ER Golgi (anterograadne transport)
- Klatriin : Rakumembraan lüsosoom (entotsütoos)
Milline
tähtsus on järjestustel KDEL ja Man-6-P sekreteeritavatel valkudel. - KDEL järjestus tagab, et ER luumenis esinevad valgud jäävad sinna püsima ja ei sekreteerita raku pinnale või lüsosoomidesse.
- Man-6-P järjestus tagab lüsosoomi valkude transpordi lüsosoomidesse.
Millised
valgud ja milleks on vajalikud tagamaks vesiikuli membraani ja õige
märklaudmembraani ühildumist.
Igal
endomembraani vesiikuli tüübil on oma spetsiifiline Rab valk
(monomeerne G-valk). Rab
valkusid on tuntud ~70 erinevat. Kui on
seotud GDP-ga, on inaktiivne ja paikneb tsütosoolis. GTPga seotud
kujul on seotud
organelli või transportvesiikuli membraaniga GPI
ankruga ja aktiivne. Rab valgud seostuvad märklaudmembraani Rab
efektoriga, mis on vajalik membraanide kokkusulamiseks. Rab
efektoriks võivad olla
mootorvalgud , mis transpordivad vesiikuleid
piki mikrotorukesi või filamente märklaudmembraanini. Rab valgu
seostumisel Rab efektoriga saab võimalikuks vesiikuli membraani
integraalse valgu V-SNARE seostumine märklaudmembraani T-SNARE
valguga. V-SNARE tüüp on iga transportvesiikuli tüübi puhul
spetsiifiline ja tagab kokkusulamise kõige märklaudorganelliga,
millel esineb üldine ühildumist
tagav valk SNAP25 ja üks või mitu
spetsiifilisust tagavat valku T-SNARE. Seega spetsiifilisus on
kindlustatud ka V-SNARE – T-SNARE vastastikuse toimega.
Millistesse
membraanidesse võib sekretoorne rada vesiikuleid toimetada?Rakumembraan,
lüsosoom,
vakuool , peroksüsoomid, Golgi kompleks, ER, endosoom
Nimetage
organelle mis on ümbritsetud poolega lipiidsest kaksikkihist. Kuidas
ja kus sellised kompleksid tekivad, mis on nende ülesanded? - VLDL – sekreteeritakse maksarakkudest ja kannavad sünteesitud triglütseriidid rasvarakkudesse
- IDL – veres tavaliselt puuduvad, VLDL ja LDL vahepealsed
- LDL – transpordivad kolesterooli maksast keharakkudenid.
- HDL – koguvad kolesterooli keharakkudest ja viivad tagasi maksa
Kuidas
tagatakse valkude õige konformatsiooni teke ER-is. Kuidas
märgistatakse ja parandatakse/kõrvaldatakse vales konformatsioonis
valgud? Kalneksiini ja kalretikuliini osa. - Esineb suur hulk valkude õige konformatsiooni omandamiseks vajalike valke (chaperone)
- Mutantsed ja vales konformatsioonis valgud suunatakse ER-st tagasi tsütosooli või jäävad seotuks chaperonidega, mis vähendab nende konsentratsiooni.
- Kalneksiin ja kalretikuliin on lektiinisarnased chaperonid , mis seostuvad valkude sahhariidsete jääkidega. Seostuvad mittetäielikult struktureerunud valkudega ja hoiavad neid ER-is(on ER membraaniseoselised) kuni tekib õige struktuur.
- Ebatäielikult struktureerunud valgud tunneb ära glükosüülitransferaas (UGGT) , mis seob oligosahhariidse jäägi otsa glükoosijäägi. Selle ühe glükoosijäägi alusel chaperon’id tunnevad ära vales konformatsioonis valgu. Kui valk lõpuks omandab õige struktuuri, glükoosi jääk eemaldub, chaperon’id dissotsieeruvad ja valk võib ER-ist edasi liikuda.
- Kui valk ei omanda õiget struktuuri, ta märgitakse UGGT poolt uuesti glükoosijäägiga ja läbib kalneksiiniga tsükli uuesti. Või vigane valk liigub läbi translokaatori tagasi tsütoplasmasse. Vales konformatsiooonis valgud seostuvad translokaatoriga. Translokaatoriga on tsütosooli poolselt küljelt seotud valkude ubikvitiiniga seostamise ensüümid. Ubikvitiiniga seostumisel toimuv ATP hüdrolüüs aitab valke tõmmata tsütosooli proteasoomi lagundamiseks. Tsütosoolis eemaldatakse sahhariidne kompleks, valk lagundatakse proteasoomis pärast märkimist ubikvitiiniga.
Nimetage
millised reaktsioonid (milliste ensüümide vahendusel) peavad
toimuma membraanide lipiidide sünteesil. Kus toimub selliste
lipiidide süntees?Fosfolipiidide
moodustumiseks on vajalik:
koostisesse kuuluvate molekulide ( glütserool , sfingosiin, rasvhapped) süntees.
rasvhapete liitmine glütseroolile või sfingosiinile (atsüülitransferaasid)
hüdrofiilse ‘pea’ liitmine
Rasvhapete biosüntees toimub AcCoA jääkide lisandumisel kasvavale ahelale erinevates raku osades:
- tsütoplasmas (küllastunud rasvhapped süsiniku aatomite arvuga kuni 16 ), taimedes ka kloroplastides
- ER-is (küllastatud rasvhapetest küllastamatute teke ja ahela pikenemine )
Glütseroolfosfaat
tekib
dihüdroksüatsetoonfosfaadi redutseerumisel glütserool 3-fosfaadiks
tsütosoolis vastava dehüdrogenaasi toimel.
Rasvhappejääkide
sidumine glütserool 3-fosfaadile toimub atsüülitransferaaside
vahendusel, mis paiknevad
ER tsütoplasma poolsel küljel.
Rasvahappejäägid
seostuvad rasvatsüül CoA koostises.
Järjestage
VLDL, HDL, LDL partiklid vastavalt neutraalrasvade
kontsentratsioonile ( suuremalt kontsentratsioonilt väiksema
suunas).Millest on tingitud neutraalrasvade kontsentratsiooni muutus
nendes partiklites.
VLDL,
LDL, HDL. VLDL modifitseeritakse IDL-iks (ingl intermediate- density lipoproteins) ja siis LDL-iks. Modifitseerumise käigus väheneb TAG
sisaldus, sest veresoonte seinad sisaldavad lipaasi, mis hüdrolüüsib
triatsüülglütseroolid glütserooliks ja rasvhapeteks ja seega
partiklite tihedus kasvab. HDL
partiklid
adsorbeerivad kolesterooli perifeersetest kudedest ja muudavad selle
estriteks.
Milline
haigus on hüperkolesteroleemia ja millest on tavaliselt tingitud
haiguse avaldumine noores eas.
Esineb
kolesterooli rakkudesse transpordi häiretega inimestel, pärilik.
Süptoomiks on kõrge kolesteroolitase veres ja ateroskleroosist
tingitud infarktid varases nooruses.
Kuidas
toimub LDL partiklite sisenemine lipiide vajavatesse rakkudesse?
Kui
rakk vajab kolesterooli ta sünteesib vajalikud LDL retseptorid ja
väljutab need plasma membraanile. LDL retseptorid diffuseeruvad
vabalt kuni nad seostuvad klatriiniga kaetud osakestega. LDL osakesed
veres seostuvad nende rakuväliste LDL retseptoritega. Klatriiniga
kaetud osakesed moodustavad seejärel vesiikulid, mis
endotsüteeritakse rakku.
Mis
on androgeenne tundetus ja millest põhjustatud?
Suguliiteline retsessiivne haigus. Esineb genotüübiga XY inimestel, fenotüüp on
XX. Põhjuseks on mutatsioon X kromosoomi geenis, mis kodeerib testosterooni retseptorit.
Testosteroon
kontrollib nii primaarsete kui ka puberteedis avalduvate sekundaarsete sugutunnuste arengut. Kui ei seostu muteerunud
retseptoriga, need tunnused ei teki.
Millised
võimalused on lipiidide liikumiseks rakus ühest membraanist teise,
nimetage lipiide transportivaid valke.
Membraanis
esinevad valgud nn fosfolipiidide translokaatorid e skramblaasid, mis
sageli tagavad minutite jooksul ER membraani tsütosooli poolse ja
luumenipoolse lipiidikihi koostise ühtlustumise.
Esineb
ka ABC tüüpi transportvalk lipiididele nn flipaas, mis peamiselt
transpordib luumenipoolsest kihist fosfatidüülseriini ja
fosfatidüületanoolamiini tsütosooli poolsesse külge, tekitades
ebasümmeeetria kahe lipiidikihi koostises.
Fosfolipiidide
üksikute molekulide liikumine võib toimuda ka fosfolipiide
transportivate valkude (ingl phospholipid exchange proteins)
abil, mis seovad teatud lipiidimolekuli ühest membraanist (ER) ja
vabastavad selle molekuli teise membraani (oletatavasti eelkõige
mitokondritesse ja peroksüsoomidesse).
Milline
transpordisüsteem peaks endosoomi membraanis olema, et pH endosoomis
muutuks happeliseks ?
V
tüüpi H+-ATPaas
Kuidas
lüsosoomi madal pH tagab tsütosooli komponentide kaitse
hüdrolaaside eest lüsosoomi lõhkemisel.
Lüsosoomi
hüdrolaasid on aktiivsed vaid lüsosoomi madala pH juures,
aluselises tsütosoolis nad inaktiveeruvad.
Nimetage
ja kirjeldage endotsütoosi variante
Endotsütoos – rakumembraani teatud osa sopistub sisse koos väliskeskkonna materjaliga ja moodustab uue membraaniga ümbritsetud organelli,
endosoomi (0,05 –0,1 µm).
- Pinotsütoos - rakumembraan ebapetsiifiliselt ümbritseb väliskeskkonnas paikneva vesilahuse osa
- Reteptorseoseline selektiivne endotsütoos – raku membraanis paikneva retseptoriga seostub väliskeskkonna teatud aine (ligand) ja retseptor-ligand kompleks assimileeritakse endotsütoosi teel ja moodustub transportvesiikul.
Mis
on transtsütoos?
Protsess,
mille puhul endotsütoosiga rakku sisenenud ained sekreteeritakse
teisele
poole rakku, liiguvad raku vastasmembraanile.
Millised
on erinevused rakumembraani ja tuumamembraani vahel? Milline raku organell on tuumaümbrisega ühendatud?
Nimeta
protsesse, mis toimuvad eukarüootse raku tuumas.
DNA replikatsioon , DNA parandamine, RNA transkriptsioon , ribosoomi
subühikute kokkupanek, DNA molekulide kokku pakkimine ja lahti arutamine ,
Kus
ja kuidas paiknevad lamiinid ja mis on nende ülesanne?
Tuuma
sisemise membraani sisepinnal on õhuke kiht tuuma
lamiine.
Lamiinid on valgud, mis kuuluvad raku tsütoskeleti valkude kolmest
tüübist intermediaarsete filamentide klassi (ülejäänud kaks
klassi: aktiini filamendid ja mikrotuubulid ). Lamiinid toetavad tuuma
sisemist membraani seestpoolt. Lisaks on rakutuumas paiknev kromatiin lamiinide vahendusel tuumaümbrise sisepinnaga seotud.
Milline
on valkude transpordi erinevus tsütoplasma-ER-i ja tsütoplasma-tuuma
vahel?
- Transport tuuma toimub tuumapooride vahendusel, kuid ER-i translokaatorite vahendusel.
- Tsütoplasmas ribosoomidel sünteesitud valkudest liiguvad tuuma need, mis sisaldavad lühikest aminohappelist järjestust, mida nimetatakse tuuma lokalisatsiooni signaaliks ehk NLS-ks ( nuclear localization signal ), mis võib valgus paikneda suvalises kohas. Valgu tranpordil ER-i aga esineb signaaljärjestus ehk liiderjärjestus valgumolekuli N-terminaalses otsas.
- Pärast tuuma sisenemist ei lõigata valkude küljest ära NLS-i, sest neid võib korduvalt vaja olla. ER-is aga valkude töötlemise käigus liiderjärjestus eemaldatakse (signalaas).
- Tuumast välja transportimiseks sisaldavad valgud vastavalt ekspordisignaali: NES-i (nuclear export signal). Enamik valke, mis sisaldavad NLSi, sisaldavad ka NES-i, sest tuumas ei toimu valkude lagundamist.
- Veel üks tuumatranspordi eripära on, et tuuma liikumisel ei pea valk olema lahti keerdunud, nagu on nt tavaline nende valkude puhul, mis liiguvad nt ER-i värskelt sünteesituna selle pinnal olevalt ribosoomilt.
- Tuuma importi ja eksporti vahendavad spetsialiseerunud transportvalgud e. tuuma transpordi retseptorid. Neid nimetatakse ka karüoferriinideks (vrd. i.k ferry) ja vastavalt transpordi suunale saab neid jagada importiinideks ja eksportiinideks. Tihti on vajalik ka veel lisaretseptor ehk adaptorvalk, mis ühendab transportija ja transporditava.
Tuuma
transpordi saab jagada kolme etappi :
1.tuuma
viidavad molekulid tuntakse ära tsütoplasmas oleva lahustuva
retseptori poolt
2.retseptori
ja tuuma viidava valgu kompleks seondub tuumapoori filamentidele ja
liigub tuuma
3.tuumas
toimub koorma ja retseptori dissotsieerumine
Kui
suur on tuumapoori difusioonipiir? Kirjelda lühidalt tuumapoori
kompleksi ehitust.
40kDa
Poor
(avause läbimõõt ~9Å) moodustub ligi 100 erinevast valgust,
millest enamik on tähistatud erineva numbriga nukleoporiinina
(NUP). Poore on tuumamembraanis 3000-5000. Elektronmikroskoobiga saadud
tulemustele toetudes on leitud, et tuuma poori tsütoplasma poolsel
küljel asuvad välja sirutunud filamendid ning tuuma sisemuse poole
jääb nn tuuma korv (korvi seina moodustava filamendi pikkus ca 100
nm). Poori moodustavad erinevad valgukompleksid on lisaks seotud
tuuma
lamiinidega.
Millised
valgumolekuli osad tagavad tuumaimpordi ja -ekspordi valkudele , mille
suurus ületab tuumapoori difusioonipiiri? Kirjelda neid lühidalt.
Tuuma
poori avaust täidavad tuumapoori valkude sabad, kus paiknevad vaheldumisi hüdrofoobsed a-happejäägid (FG – fenüülalaniin ja
glütsiin) ja hüdrofiilsed aminohappejäägid, moodustades seeläbi
geelja struktuuri. On postuleeritud, et see keskkond on sobitunud
just tuuma transportvalkudele, mis tänu oma keemilistele omadustele
suudavad geeljat keskkonda kergelt läbida. Vastavat
seondumisspetsiifikat mitte omavad ühendid tuumapoori läbida ei
saa.
Kuidas
liigitatakse karüoferriine ning mis on rakus nende ülesandeks?
Tuuma
importi ja eksporti vahendavad spetsialiseerunud transportvalgud e.
tuuma
transpordi retseptorid.
Neid nimetatakse ka karüoferriinideks (vrd. i.k ferry)
ja vastavalt transpordi suunale saab neid jagada importiinideks
ja eksportiinideks.
Tihti on vajalik ka veel lisaretseptor ehk adaptorvalk, mis ühendab
transportija ja transporditava.
Tuuma
transpordi saab jagada kolme etappi:
tuuma viidavad molekulid tuntakse ära tsütoplasmas oleva lahustuva retseptori poolt
retseptori ja tuuma viidava valgu kompleks seondub tuumapoori filamentidele ja liigub tuuma
tuumas toimub koorma ja retseptori dissotsieerumine
Milline
makroergiline ühend tagab energia transpordil tuuma ja tsütoplasma
vahel ning milliste valkude vahendusel see transport teoks saab (3
valku)?
GTP
GTPaasse
aktiivsusega valk Ran
Ran-GEF
Ran-GAP
Tuuma
transpordi füsioloogiast
Energiat
mõlemasuunaliseks transpordiks ehk impordi- ja ekspordiretseptorite
koormast vabastamiseks annab GTPaasse aktiivsusega valk Ran. See
valk esineb kahes konformatsioonis, milles ühes on ta seondunud
makroergilise ühendi GTP ja teises GDP vormiga . Tuumas on rohkem
Rani, mis on GTP-ga seotud, sest tuumas alaliselt paiknev
Ran-GEF-valk (guanine
exchange factor )
viib tuuma saabuva GDP-ga seondunud Ran-i kohe GTP-ga seondunud
vormi. Tsütoplasmas aga on rohkem GDP-ga seotud Rani, sest seal
katalüüsib nukleotiidivahetust (GTPst saab GDP) valk nimega Ran-GAP
(i.k GTPase
activating
protein). Seega tekib tuuma ja tsütoplasma vahel Ran valgu kahe vormi
gradient ning pideva nukleotiidivahetuse reaktsiooniga on seotud ka
tuumaretseptoride kooremvalguga seondumine/dissotsioeerumine: Ran-GTP
vabastab tuumas sinna jõudnud impordiretseptori tema koormast –
koorem
on tuumas. Samal
ajal tõstab Ran-GTP tuumas olles tuuma ekspordisignaali omavate molekulide afiinsust ekspordiretseptorite suhtes. Kui need on
omavahel seondunud ja tsütoplasmasse transporditud, lahutatakse
NES-i sisaldav valk ekspordiretseptorist selle energiaga, mida
saadakse GTP hüdrolüüsist GDPks.
Kuidas
paiknevad tuumas kromosoomid ning kuidas paiknevad tuumas rohkem
geene sisaldavad kromosoomid?
Üldiselt
kehtib seaduspära , et rohkem geene sisaldavad kromosoomid paiknevad
tuumas rohkem tsentraalselt, ning kromosoomi need osad, milles on
rohkem geene, paiknevad kromosoomi territooriumi äärealadel.
Mis
eristab tuuma organelle tsütoplasma organellidest?
Nad
pole membraaniga ümbritsetud, ja molekulid, mis asuvad mujal
tuumaplasmas, saavad neisse piirkondadesse vabalt difundeeruda.
Milline
on kõige tuntum tuuma organell/reaktsioonikeskus?
Tuumake .
Mis
toimub tuumakeses ja mitu tuumakest võib maksimaalselt inimese
rakutuumas olla? Mis selle numbri määrab?
Selles
piirkonnas toimub rRNA geene sisaldavalt DNAlt rRNA süntees ja
protsessimine ning rRNA-le ribosomaalsete valkude liitmine, ehk siis
ribosoomi subühikute kokkupakkimine. Lisaks on leitud, et lisaks
rRNA geenide transkriptsioonile toimub tuumakeses ka ribosoomi
valkude geenide transkriptsioon – kinnitav näide, et ühe
funktsionaalse üksuse (ribosoom) varuosade tootmine on koondatud
tuumas ühte keskusesse.
Kromosoomi
osa, milles paiknevad rRNA geenid , nim tuumakese
organisaatori piirkonnaks
(i. k nucleolus
organizer region ,
NOR). Inimese genoomis (23 kromosoomi) esineb NOR viies erinevas kromosoomis. Diploidses rakus on seega 10 NOR-i ja neis paikneb
tandeemselt kokku ~200 rRNA geeni. Need 10 NOR-i pole tavaliselt
eraldi nähtavad ja moodustavad tihti kokku ühe suure tuumakese.
Kuid teoreetiliselt võib ühes tuumas olla maksimaalselt 10
tuumakest. Tavaliselt kõikidel kromosoomidel olevaid rRNA geene
korraga ei transkribeerita ja tuumakesi on tuumas vähem.
Miks
on ribosoomi RNA geene eukarüootides palju?
Raku
suur nõudlus ribosoomide järgi?
Miks
ribosoomide koostises olevat kolme RNA molekuli ühise pre-RNA-na
sünteesitakse?
See
tagab nende saamise võrdsetes kogustes, nagu neid ka vaja on.
Mis
on hetero - ja eukromatiin? Kus paikneb rakutuumas enamasti
heterokromatiin? Millised kromosoomi osad liigitatakse
konstitutiivseks heterokromatiiniks? Miks?
- Kondenseerunud DNA-ga piirkondi nimetatakse heterokromatiiniks.
- Dekondenseerunud DNA-ga piirkondi nimetatakse eukromatiiniks.
- Imetajate rakkudes seostub heterokromatiin tuumamembraaniga lamiinide vahendusel ning paikneb tuumakeste ümber.
- On leitud, et heterokromatiin ei sisalda praktiliselt üldse geene, vaid hoopis kõrgelt/mõõdukalt korduvaid DNA järjestusi, mis on enamiku raku eluajast tugevalt kokku pakitud. Seetõttu nimetatakse seda kromatiini konstitutiivseks heterokromatiiniks (nt tsentromeeride ja telomeeride DNA).
Milles
seisneb geenide positsiooniefekt?
Geenide
kohasõltuv avaldumine. Kui heterokromatiini piirkonnas olev DNA
tuumas oma asukohta vahetab ja liigub eukromatiini alasse, võib see
algatada selle DNA transkribeerimise. Sama efekt esineb ka
eukromatiini puhul – kui ekspresseeruv geen viia eukromatiini
piirkonnast üle heterokromatiini piirkonda, geen vaigistatakse.
Millised
on kolm kromosoomi säilitamiseks vajalikku struktuurielementi?
Et
DNA molekul oleks funktsionaalne ehk võimeline edukalt
tütarrakkudesse kanduma, vajab ta kolme strukturaalset ühikut –
telomeeri, tsentromeeri ja replikatsiooni alguspunkte
Replikatsiooni
alguspunkt – milleks vajalik, erinevused pro- ja eukarüootidel.
Struktuur,
mis oma järjestuselt pole küll korduselement, aga on vajalik
kromosoomi kui terviku toimimiseks.
Prokarüootidel esinevad kindlad DNA järjestused, kuhu DNA polümeraas seostub.
Eukarüootidel kindlad alguspunktid puuduvad, replikatsioon algab
stohhastiliselt. Ajaliselt varem replitseerub neil eukromatiin.
Tsentromeer – milleks vajalik, kirjeldada lühidalt üldist ehitust.
Kinetohoori ülesanne.
Kromosoomi
piirkond, mis reguleerib kromosoomide liikumist mitoosi ja meioosi
ajal. Kindla DNA järjestusega ja kindlate sinna seonduvate
valkudega. Tsentromeeri ülesanne on hoida koos õdekromatiide kuni
mitoosi anafaasini (või meioosi II jagunemise anafaasini).
DNA lõigud , mis koosnevad tandeemselt korduvatest monomeersetest
järjestustest, mis on omakorda organiseerunud kõrgemat järku
kordusteks.
Tsentromeeri
külge moodustub jagunevas rakus spetsiaalne ligi 90 erinevast
valgust koosnev struktuur - kinetohoor.
Selle külge kinnituvad omakorda mikrotuubulid, mis õdekromatiidid
tütarrakkude poolustele tõmbavad.
Telomeer – milleks vajalik, kirjeldada lühidalt üldist ehitust.
Kromosoomi
otsa kaitsev spetsiifiline kordusjärjestus, kindlustab kromosoomide
otste replikatsiooni ja kaitse raku elu vältel. Üks hüpotees telomeeride funktsioneerimise kohta: nad katavad (koos sinna
seondunud valkudega) kromosoomi otsi, et raku kontrollmehhanism ei
tõlgendaks kromosoomi otsi kui DNA ahela katkemiskohti.
Üldiselt
pole telomeerid kindla pikkusega. Inimesel 650 kuni 2500 kordust (eri
kudedes erinev). 3´ots on alati G-rikas ja ca 200 bp üleulatuv, mis
arvatakse tagasi pöörduvat ja lassotaolise T- lingu moodustavat.
Miks
telomeerid lühenevad?
DNA
molekul jääb aga iga replikatsioonitsükliga otstest järjest
lühemaks, sest DNA sünteesil nn mahajääva ahela iga jupi sünteesi
alustamiseks kasutatava RNA praimeri kõige viimast enda
seondumiskohta ei täida DNA nukleotiididega enam miski. Seega
sünteesitud mahajääv ahel on 5’ otsast lühem kui ahel, mille
järgi ta sünteesiti.
Mis
juhtub rakuga, kui telomeerid on raku põlvkondade vältel täielikult
kadunud?
Sellised
kromosoomid muutuvad ebastabiilseteks – kui katkiste otstega
kromosoome on mitu, võivad nad omavahel liituda. Lisaks toimib
kõikides rakkudes rakujagunemise kontrollsüsteem, mille käivitab
DNA kahjustus - kui DNA ahel ( kromosoom ) mingil põhjusel katkeb ja
tekib vaba DNA ots, tekitab see signaali, mis peatab rakujagunemise.
Lisaks DNA katke hüpoteesile on välja pakutud, et rakujagunemise
kontrollimisel osalevad geenid paiknevad vahetult telomeeride järel.
Kuna on teada, et telomeeride kõrval olevad geenid on tavaliselt
inaktiivsed, siis on arusaadav, et telomeeride lühenemisel või
kadumisel aktiveeritakse need rakujagunemist kontrollivad regulaatorgeenid .
Milline ensüüm pidurdab osades rakkudes telomeeridel lühenemast? Mil
viisil see pidurdamine toimub?
Telomeraas uuendab pidevalt telomeere.
Millistes
rakkudes on vaja aktiivset telomeraasi?
Need
rakud, mis peavad koguaeg paljunema. Soolerakud, vereloomerakud,
naharakud.
Miks
prokarüoodi DNA pole nii tihedalt kokku pakitud kui eukarüoodi DNA?
Proarüoodi
DNA-l on geene tihedamalt kui eukarüootidel ja samas kogusummas
vähem kui eukarüootidel, ning selle tulemusena on mingi konkreetne
DNA piirkond ilmselt suhteliselt sagedamini ekspresseeritud. Seega
võiks eeldada, et bakteri kromosoomi väga tugev kokkupakkimine pole
ilmselt nii otstarbekas kui eukarüoodi puhul.
Millises rakutsükli faasis on eukarüoodi kromosoomid kõige rohkem kokku
pakitud? Mitu korda on DNA selles struktuuris kondenseerunud? Kuidas
erineb kromatiini kokkupakitus nn G- ja R-bändides?
Mitoosi metafaas .
~10 000x.
Tugevamalt
värvunud G- bändid on rohkem kokkupakitud/kondenseerunum. Heledalt
värvunud R-bändid vähem.
Kuidas
ühe liigi mitoosi metafaasi kromosoomidest koostatud kogumit
nimetatakse ning mida selle abil teha saab?
Karüotüüp .
Nende
põhjal saab kromosoome ning nende ümberkorraldustest tulenevaid
haigusi määrata.
Eukarüoodi
DNA esimene ja teine pakkimistasand – kuidas neid nimetatakse? Mitu
korda need DNAd lühendavad?
- Esimene tasand : pakkimine nukleosoomi, lühendab 7x
- Teine tasand : 30nm fiiber/ solenoid , lühendab 40x
Millistest
histoonidest moodustub nukleosoom (mitu histooni ja millised
histoonid)? Milline histoon sellesse struktuuri ei kuulu ja mis on
tema teadaolevad ülesanded tuumas?
Kromatiini
koosseisus on histoone viit tüüpi – H1, H2A, H2B, H3, H4.
Nukleosoom
koosneb histoonidest ja nende ümber keerdunud DNA-st. Ühes
nukleosoomis on kokku kaheksa histooni molekuli: iga nelja histooni
(H2A, H2B, H3, H4) on kahes korduses. Nukleosoomide vahel paiknevale
DNA ahelale seondub H1 histoon, mida nimetatakse ka linkerhistooniks.
H1 ülesandeks on nukleosoomi struktuuri stabiliseerida, aga samuti
tagada kõrgemate DNA kondenseerumistasandite stabiilsus.
Millised
mittehistoonsed valgud aitavad kromatiini kõrgemat järku
struktuuridesse pakkida ja millised piirkonnad on selle jaoks DNAs?
Need
moodustavad kromosoomikujulised tellingud, millele kinnituvad 30 nm
fiibrid, arvatavasti lingudena. DNA-s asuvad nende valkudega
seondumiseks kindlad piirkonnad, mida nimetatakse toese/tellinguga
seotud piirkondadeks SAR (i.k, scaffold-associated regions )
või maatriksiga seotud piirkondadeks (i.k, matrix
attachment regions MAR).
S/ MARs paiknevad transkriptsiooniühikute vahel, st geenid paiknevad
kromatiini lingudes ning geen võiks teoreetiliselt avalduda ka nii,
et DNA jääb toesevalkudega seondunuks.
Kromosoomide
mitootilise struktuuri tekkes osalevad lisaks veel üht tüüpi
valgulised kompleksid, mida nim kondensiinideks
ja kohesiinideks, mis kuuluvad
SMC valkude perekonda (structural
maintenance of chromosomes).Tegu
on dimeersete (funktsionaalne on kahest identsest valguahelast
moodustunud struktuur) molekulidega, mille vabad otsad osalevad
kromatiiniga seostumises ja hüdrolüüsivad ATP-d, et DNA
liigutamiseks energiat saada.
Milles
seisneb C-väärtuse paradoks ?
Seaduspära
organismi keerukuse ja genoomi suuruse vahel alati ei kehti ja seda
nimetatakse C-paradoksiks.
Kirjeldage
geeni ehitust.
Geeniks
loetakse lisaks kodeerivatele piirkondadele ka regulaatorpiirkonnad,
mis võivad kodeeerivast osast küllalt kaugel paikneda, ja ka osad,
mis mRNA eellasest välja lõigatakse - intronid .
Geeni RNAd kodeerivale alale eelneb vahetult piirkond, kuhu kinnitub RNA
polümeraas ning seda piirkonda nimetatakse promootoriks.
Geeni
koosseisu arvatud regulaatorpiirkonnad võivad RNA-d kodeerivast
osast (vähemalt eukarüootides päris kaugel paikneda – kuni 50
000 aluspaari )
RNA
transkriptsioon lõpeb piirkonnas, mida nimetatakse terminaatoriks.
Mida
mõeldakse eukarüoodi geenide segmenteerituse all ning mis toimub
splaissingu käigus?
Geenide
segmenteeritus
tähendab eksonite
ja intronite
olemasolu.
Eksonid
(i.k - expressed
sequences)
on piirkonnad DNAs ja RNAs, milles olev info jõuab valgu kujule, ja
intronid
(i.k
- intervening
sequences)
lõigatakse splaissingu
käigus RNA molekulist välja.
Milliseid
ülesandeid võib täita rakus RNA (mRNA-d mitte arvestades)?
Nimetage selle kursuse raames käsitletud
funktsioone/organelle/ensüümikomplekse, kus RNA on osaline.
Ribosoomide
koostises rRNA, vastutab mRNA lugemise mehhanismi eest.
mRNA
sisaldab geneetilise informatsiooni jõudmine rakutuumast ribosoomini
translatsiooni protsessiks.
tRNA
transpordib ribosoomidele spetsiifiliselt aminohappeid valgu
sünteesiks.
Geneetilise
informatsiooni vahetus mitokondrite DNA ja tuuma vahel käib RNA
kaudu.
Transposonite
teke.
snRNA
– pre-mRNA splaissimine
snoRNA
– rRNA keemiline modifitseerimine
Kui
suure osa inimese genoomist moodustavad valke kodeerivad järjestused
ehk eksonid? Mis on pseudogeen ?
1,5%, Pseudogeen on normaalse geeni analoog, mis on kaotanud oma
funktsiooni.
Millised
kromosoomi osad liigitatakse kõrgelt korduva DNA hulka? Kust tuleneb
nende nimetus „ satelliit -DNA“?
Kõrgelt
korduv DNA koosneb enamasti
tandeemselt järjestatud identsetest 5-10 bp pikkustest
järjestustest. Mõned tandemkordused on ka pikemad, 20-200 bp. Nim
satelliit-DNAks, sest eraldub muust DNAst tihedusgradiendis
tsentrifuugimisel, kuna sisaldab rohkem AT paare (taimedes on
vastupidi, GC aluseid). Kõrgelt korduva DNA hulka arvatakse
tsentromeerid ja telomeerid,
kaks piirkonda, mis on kromosoomi kui iseseisva struktuuriüksuse
toimimiseks vajalikud.
Millisesse
kahte rühma liigitatakse mobiilsed DNA elemendid ja kuidas võib
nende kahe rühma liikumise mehhanismi põhimõtteliselt ja väga
lühidalt kirjeldada?
- Transposonid. Transposonid on DNA segmendid, mis liiguvad genoomis ühest piirkonnast teise „lõika ja kleebi “ põhimõtte alusel – kohta vahetab jupp DNA-d. Liikumine toimub sagedamini ühe kromosoomi piirides, harvem erinevate kromosoomide vahel.
- Retrotransposonid. Nende liikumismehhanismi võib hästi võrrelda „kopeeri ja kleebi“ põhimõttega, ehk transponeerumine tähendab tegelikult jupi DNA kordistamist ja selle DNAs uude kohta sisestamist. Protsess toimub RNA vaheühendi kaudu. Kõik seni uuritud eukarüoodid sisaldavad retrotransposone.
Kust
tuleneb retrotransposoni „retro“ nimetus?
Vanasse
kohta jääb retrotransposoni koopia alles.
Millised
ensüümid vahendavad transposonide (1) ja retrotransposonide (2)
transponeerumist? Kui suure protsendi inimese genoomist moodustavad
transponeeruvad elemendid?
- Transposoneid vahendavad :
Transposaas – valk, mis vahendab DNA lõikamist ja aitab sisendada uues kohas – vahendab transpositsiooni.
- Retrotransposoneid vahendavad :
Pöördtranskriptaas – sünteesib mRNA-l oleva info põhjal uue lõigu DNA-d.
Integraas – selle toimel lülitub sünteesitud DNA lõik uude kohta.
Inimese
genoomist moodustavad transponeeruvad elemendid ~3%.
Kuidas
liigitatakse retrotransposone ning milline neist kahest alarühmast
on praegugi inimpopulatsioonis aktiivne? Nimeta selle alarühma kolm
enim kirjeldatud mobiilset elementi.
- Retroviiruslikud
- Mitte-retroviiruslikud
Mitteviiruslikud
retrotransposonid on ainsad transponeeruvad DNA elemendid, mis
tänapäeval inimeses transponeerumise aktiivsuse on säilitanud.
LINE elemendid
Alu elemendid
Sva elemendid
Kas
transponeeruvate elementide aktiivsus on hea või halb? Põhjendage
vastust.
Kuigi
transpositsioon pole sage, võib sisestumine teatud kohta DNAs
toimuda mõnda geeni ja põhjustada seeläbi geeni funktsiooni
häirumise. Ent samas on mutatsioonid geenides evolutsiooni aluseks,
seega võib toimuda ka harvasid positiivseid mutatsioone .
Milleks
rakutuuma olemasolu hea võiks olla?
Tuuma
olemasolu tõttu on eukarüootidel DNA replikatsioon ja RNA
transkriptsioon tsütoplasmas olevast translatsiooni masinavärgist
ja teistest metaboolsetest protsessidest täiesti eraldatud. RNA ja
valkude selektiivne läbilaskvus pakub ühtlasi täiendavat kontrolli
geenide ekspresseerumise ning replikatsiooni üle.
Tooge
näiteid valkudest, mis peavad igas rakutüübis ekspresseerunud
olema. Kuidas neid valke ühiselt nimetada võiks?
Kõigis
rakkudes on vaja nt niinimetatud koduhoidjavalke: DNA
replikatsioonil, RNA transkriptsioonis jne osalevad valgud,
histoonid, ribosoomivalgud, energia- ja ainevahetust käigus hoidvad
valgud jne.
Kui
palju arvatakse (hetkel) inimesel geene olevat ning kui suur osa
neist korraga ekspresseeruda võib? Kui suur arvatakse olevat geenide
protsent, mille produktid geeniregulatsioonis osalevad?
20 000
– 25 000.
Inimese
rakus ekspresseerub korraga keskmiselt ~30-60% geenidest
10-15%
kõikidest geenidest arvatakse olema seotud geeniregulatsiooniga.
Millised
cis-elemendid
on vajalikud, et RNA polümeraas ühe geeni transkribeerimise
alustaks ja lõpetaks?
Promootor ja terminaator
Mis
tagab enamike valguliste transkriptsioonifaktorite DNA-ga seondumise
spetsiifilisuse?
Konkreetsete
cis-elementide
ja trans-faktorite
seostumise aluseks on tõmbejõud, mis tekivad lisaks H-sideme
doonorite või aktseptoritele ka hüdrofoobsete vastasmõjude,
elektriliste laengute ja teiste molekulaarset äratundmist tagavate
faktorite tõttu.
Kuidas
saab RNA polümeraasi ligipääsu geeni ees asetsevale promootorile
takistada? Kuidas saab seda soodustada?
- Takistada : repressorvalgu seondumine promootoriga, DNA ahela kokku pakkimine
Soodustada
: promootorite sidumisega mingi muu ühendi poolt, DNA ahela enda
konformatsiooni avatumaks muutumine
Milliste
vahenditega tagatakse võrreldes prokarüootidega eukarüootide
geeniregulatsiooni paindlikum kontroll?
Eukarüootides
on kõik geenid on üksikuina kontrollitavad, puuduvad mitme geeni
ekspressiooni kontrollivad operonid. Üht geeni reguleerivaid valke
võib olla mõnisada (võivad toimida nii transkriptsiooni
alguspunkti lähedal kui ka kaugemal asuvate DNA
regulaatorpiirkondade kaudu). Oluline on mediaatori
– suure valgukompleksi - osalus transkriptsiooni algatamises. See
toimib vaheühendina RNA polümeraasi ja erinevate regulatoorsete
valkude vahel.
Nimeta
histoonide kovalentseid modifikatsioone. Milline neist põhjustab DNA
ja histoonide vaheliste elektrostaatiliste jõudude nõrgenemise?
Histoonide
N-otste a-hapete modifikatsioone erinevate funktsionaalsete rühmadega
võiks teada nelja:
- Atsetüleerimine
- Metüleerimine
- Fosforüleerimine
- Ubikvitineerimine (väike 8 kDa valk ubikvitiin, rohkem tuntud tsütoplasmas valkude lagundamisele suunava signaalina).
Atsetüleerimine
(äädikhappe jää CgiH3COO-
liitimine)
Miks
on imetajate emasorganismis vajalik ühe X-kromosoomi
heterokromatiiniks kondenseerimine?
Et
vältida olukorda, kus emasorganism hakkab mingit geeni korraga kahelt kromosoomilt ekspresseerima ja põhjustab seeläbi
geeniproduktide hulga ebavõrdsuse isas- ja emasorganismi vahel.
Miks
nimetatakse inaktiveerunud X-kromosoomi fakultatiivseks
heterokromatiiniks? Mis juhtub tunnustega, mis on määratud
X-kromosoomil olevate geenidega?
Ühe
X-kromosoomi valikulisest vaigistamisest lähtuvalt nimetatakse
kondenseeritud X-kromosoomi fakultatiivseks
heterokromatiiniks. Kuna
ühe X kromosoomi vaigistamine toimub embrüo varajasel arenguetapil,
siis organismis leidub rakke, millel on isasorganismilt pärit X
kromosoom vaigistatud ja ka neid, millel on emasorganismilt pärit X
kromosoom vaigistatud. Ekspressioon toimub vaid ühelt, seega võivad
eirnevates rakkudes olla ekspresseeritud X kromosoomi geeni eirnevad
alleelid. X-i inaktivatsioon pole siiski 100%-line, umbes 10%
geenidest ekpresseeruvad ka kondenseerunud kromosoomis.
Mis
vahendab ühe X-kromosoomi inaktivatsiooni? Millisel hetkel
inaktivatsioon toimub ja kui kaua kestab?
X
inaktivatsioon algatatakse teatud geeni (Xist) transkribeerimisel
RNA-ks selles kromosoomis. Vastavat RNAd hakkab ekspresseerima ainult
inaktiveeritav X kromosoom ja piirkonda, kus RNA geen asub,
nimetatakse X-i inaktivatsiooni keskuseks (X-inactivation
center).
Transkribeeritud RNA molekul jääb tuuma, seostub X-kromosoomi
DNA-ga ja indutseerib selle heterokromatiniseerumise. X kromosoomi
inaktiveerumine toimub embrüo arengu varajastel etappidel ning
kestab terve raku eluea .
Milles
seisneb alternatiivne splaissing ja kuidas võiks see kaasa aidata
organismide paremale kohastumusele keskkonnamõjude suhtes?
Alternatiivne
splaissing tähendab valgu sünteesiks mRNA kokkupanekut geeni
erinevatest eksonitest. Eluvormide poolt alternatiivse splaissingu
laialdane kasutamine lubab oletada, et valkude erinevad
splaissinguvariandid suurendavad organismi paindlikkust
keskkonnamõjudega kohandumisel – valgud on võimalik sõltuvalt
vajadusest eri moodulitest kokku monteerida.
Millise
kahe protsessi toimumine on peamiseks märguandjaks, et mRNA on
valmis tuumast välja transportimiseks?
- Splaissingu toimumine
- 5’ ja 3’ otste kovalentne modifitseerimine. 5’ otsa lisatakse spetsiifiline cap-struktuur. 3’-ots modifitseeritakse aga adeniini nukleotiididega ja sealt ka selle nimetus – polü-A-saba.
Millises
mRNA molekuli osas asub info tema rakusisese suunamise kohta? Too üks
näide, kus mRNA suunamine/gradient rakus olulist rolli mängib.
On
olemas suunav järjestus mRNA 3’ alas, mis ei sisalda valgu
kodeerimiseks vajalikku infot ning mida seetõttu nimetataksegi
mittetransleerivaks piirkonnaks (i.k - 3´
untranslated region, UTR).
mRNAde
rakusisest transporti kasutatakse ka närvirakkude ökonoomses
toimimises: närvirakud võivad olla väga pikad, kuid tihti peab
raku ühte ossa (sünapsisse) jõudnud signaalile vastama mingi geeni
ekspressiooni näol väga kiiresti. Üheks meetodiks selle
probleemiga hakkama saamisel on valitud ennetamine: rakku saabuvale
signaalile vastamiseks vajalike valkude mRNAd on juba eelnevalt
närviraku lõpmetesse transporditud.
Mida
mõeldakse selle all, et geeniekspressioon on mürarikas? Mis on
selle tulemus?
Rakkudes
toimuvates reaktsioonides on keemilise reaktsiooni parameetritest
lähtudes suhteliselt vähe molekule, siis mõjub juhuslikkus neile reaktsioonidele tugevamalt kui reaktsioonidele, kus osalevaid molekule on palju – reageerijate võimaliku kokkusaamise keskmine
sagedus võib küll võrreldes kangemate lahustega võrdne olla, aga
kuna molekule on vähe, varieerub see sagedus tegelikult suurtes
piirides. See teebki kõik bioloogilised protsessid rohkem või vähem
fluktueeruvaks/kõikuvaks. Tulemuseks on see, et paljude protsesside
toimumise tõenäosus sõltub juhuslikkusest.
Rakutsükkel
Defineerige
rakutsükkel, nimetage ja iseloomustage peamised rakutsükli etapid.
Sündmuste
rida (aeg) ühe raku jagunemisest kuni tekkinud tütarraku
jagunemiseni.
Rakutsükkel
on protsesside kogumik, mis tagab DNA kahekordistumise ja selle täpse
jaotumise kahe tütarraku tekkimisega .
Mis
tüüpi valgud on tsükliinid? Mis on nende funktsiooniks? Millist
tüüpi tsükliine võite nimetada?
Valkude
perekond, mis kontrollivad raku tsüklit tsükliinsõltuvate
kinaaside aktiveerimisega.
Tsükliinid
on väikesed valgud, mis on heterodimeeride tsükliinidest sõltuvate
proteiini kinaaside (CDK – cyclin dependet kinases) regulatoorseks
subühikuks. Rakutsükli erinevate faaide sisseülitamine on
kontrollitud nende valkude (CDK) poolt. (Teine subühik tsükliinist
sõltuv kinnaas).
CDK-d
teostavad mitmesuguste rakutsüklis osalevate valkude (DNA
replikatsiooni valgud, mitoosiprotsessis osalevad valgud jne)
fosforüülimist aktiveerides/inaktiveerides vastavad valgud. Nt CDK1
ja CDK2, CDK4, CDK6.
Nimetage
tsükliinidest sõltuvate kinaaside omadusi, nimetage vähemalt kolm
viisi nende aktiivsuse regulatsiooniks.
- Aminohappeline järjestus kõrgelt konserveerunud.
- N-terminaalsesotsas piirkond ATP sidumiseks.
- PSTAIRE regioon tsükliini sidumiseks.
- Katalüütilise tsentri aminohapped laiali üle kogu ahela ja satuvad kõrvuti tertsiaarstruktuuri moodustumisel
- Esineb nn. T- ling , aminohappeline järjestus, mis võib katta katalüütilise tsentri.
- CDK aktiivsus lisaks tsükliinidele on reguleeritud kinaaside, fosfataaside ja spetsiifiliste inhibiitorite (CKI) poolt.
- Erinevad CDK-d tagavad erinevate rakutsükli faaside läbimise
Aktiivsuse regulatsioon :
- Tsükliinide seostumine (katalüütiline tsentrum, mida katab muidu T-ling, on avatud kui Thr 161 on fosforüülitud, mis toimub pärast seostumist tsükliiniga)
- Teatud aminohappeliste jääkide fosforüülimine.
- Komplekside valgulised inhibiitorid (CKI). Teatud inhibiitorid pärsivad kõik CDK-tsükliin kompleksid, osa inhibiitoreid mõjuvad valikuliselt ainult teatud kompleksidele.k G
Nimetage
tsükliinidest sõltuvate kinaaside substraate rakutsüklis.
CDK1
– üleminek G2 faasist mitoosifaasi
CDK2
– üleminek S faasist G2 faasi
CDK4
ja CDK6 – üleminek G1 faasist S faasi
Mis
on MPF (maturation promoting factor, sün mitose promoting factor),
kuidas tekib, millest koosneb, kuidas laguneb.
Tsükliin
B-st ja tsükliinidest sõltuvast kinaasist koosnev heterodimeerne
valk, mis stimuleerib mitootilist ja meiootilist rakutsüklit. MPF
laguneb kui anafaasi soodustav kompleks (APC) märgistab tsükliin B
ubikvitiiniga ning määrab valgu lagundamisele.
Milline
on valk p53 osa rakutsükli kontrollis , kuidas see realiseerub?
P53
reguleerib rakutsüklit hulkraksetes organismides ja funktioneerib
kui kasvaja supressor.
- Aktiveerib DNA paranduse
- Peatab rakutsükli G1 – S faasis kui DNA kahjustus
- Indutseerib apoptoosi kui DNA-d pole võimalik parandada
APC
(anaphase promoting complex ) roll metafaasist anafaasi üleminekul (M
pärssimine).
Metafaasist
anafaasi üleminek tagatakse APC fosforüülimisega CDK-de poolt.
Märgistab ubikvitiiniga valgud proteolüüsiks. Kui ei
funktsioneeri, ei toimu mitootiliste tsükliinide lagunemist ja
anafaasi.
Nimetage
4n kromosoomistikuga rakutsükli faasid .
- S faas – DNA kahekordistumine, histoonide süntees
- G2 faas – vahemik S faasi ja mitoosi vahel
- M faas – mitoos
- G1 faas – vahemik M ja S faasi vahel
G1,
S ja G2 moodustavad kokku interfaasi.
Nimetage
vähemalt kolm protsessi raku/organismi elutegevuses mis vajavad
apoptoosi toimumist .
- Vajalik organismi normaalse arengu toimumiseks ( embrüogenees on fülogeneesi lühike ja kiire kordumine – vaja elimineerida inimestel nt lõpuste algmeid, sõrmi ühendavad rakud jne)
- Kontrollib rakkude arvu homeostaasi organismis. Homeostaas saavutatakse kui mitoosi kiirus on tasakaalus apoptoosiga. Kui rakud jagunevad kiiremini kui surevad, tekivad kasvajad .
- Elimineerib mittevajalikud ja muutunud rakud ( kahjustunud , genoomi mutatsioone sisaldavad, viirusega nakatunud rakud, autoreaktiivsed lümfotsüüdid jne)
Iseloomustage
apoptoosi etappe
Spetsiifiliste proteaaside – kaspaaside aktiveerumine. Peptiidsidemeteb lagundamine nende poolt. Kaspaase on palju, kõigepealt aktiveerub prokaspaas, mis osalise proteolüüsi teel aktiveerib järgmise prokaspaasi jne. Akiveerib kaspaaside kaskaad .
Kromatiini kondenseerumine
DNA fragmentatsioon – kaspaaside toimel lagundatakse endonukleaasi inaktiveeriv valk ja aktiveerub endonukleaas, mis hüdrolüüsib sidemed nukleosoomide vahel. Tuumamembraan (lamiinid) laguneb.
Valkude fragmenteerumine kaspaaside aktiveerumise tõttu.
Rakumembraanis toimuvad muutused, mis märgistavad apoptootilise raku fagotsüütidele.
Tsütoplasma fragmenteerub – rakk jaguneb väikesteks vesiikuliteks nn apoptoosi kehakesteks.
Vesiikulid endotsüteeritakse fagotsüütide poolt.
Iseloomustage
kaspaase ja kaspaaside kaskaadi
Kaspaasid
on apoptoosile iseloomulikud valke lagundavad ensüümid.
Aktiivtsentris on tsüsteiin. Kaspaasid aktiveeruvad kaskaadselt,
kõigepealt aktiveerub kaspaas 8, see omakorda aktiveerib teisi.
Kaspaaside toimel aktiveeruvad ka nukleaasid, mis asuvad lõikama
DNA-d. Kaspaaside kaskaadi käivitumisel toimub rakustruktuuride
süstemaatiline purustamine, sündmused toimuvad kindlas järjekorras
ja ette-ennustatavalt. Rakk hävitatakse kiiresti, 30-60min jooksul.
Kirjeldage
mitokondritega seotud apoptoosi (Apaf valgud, tsütokroom c, Bcl
valgud)
Mitokondritest
vabaneb tsütokroom c, mis seostub adaptervalguga Apaf1 (apoptotic
protease activating factor). Apaf1 polümeriseerub rattasarnase
heptameeri moodustumisega – apaptosoom. Aktiveerub prokaspaas 9
ning toimub kaspaaside kaskaad.
Bcl
2 – anti-apoptootilised valgud, mis pidurdavad apoptoosi seesmist
rada. Evolutsioonis konserveerunud. Pidurdavad tsütokroom c
vabanemist mitokondritest.
Nimetage
kasvufaktoreid (välisfaktoreid) mis olulised apoptoosi pärssimises
Loomarakud
vajavad enamasti apoptoosi vältimiseks pidevat teiste rakkude
singaliseerimist. Kui rakk ei saa eluspüsimise või kasvusignaali,
siis ta sureb , sest teda pole vaja. Signaalmolekulid seostuvad
märklaudraku pinnal paiknevate retseptoritega. Seostumine lülitab sisse signaali liikumise ahela, mis päris apoptoosi ahelat.
- Faktorid , mis suurendavad Bcl2 valgu sünteesi
- Faktorid, mis vähendavad proapatootiliste valkude sünteesi
Nimetage
apoptootiliste rakkude avastamise meetodeid
DNA fragmenteerumisel DNA ahelasse tekkivaid katkemiskohti on võimalik tuvastada spetsiaalse ensüümi – terminaalse desoksünukleotidüül transferaasi (TdT) abil (nn TUNEL reaksioon ). See ensüüm tunneb ära vabad 3’-OH otsad ning pikendab neid nukleotiididega. Kui reaksioonikeskkonda lisada radioaktiivse või fluorestsentsmärgistusega nukleotiide , siis sisestuvad need märgistatud nukleotiidid sünteesitavasse DNA-sse.
Normaalsetes rakkudes on plasmamembraan ebasümmeetriline fosfolipiidide suhtes. Fosfatidüülseriin eksponeerub ka membraani tsütoplasmaatilises pooles . Apoptootilises rakus fosfatidüülseriin eksponeerub ka memrbaani ekstratsellulaarsel poolel. On olemas valk (anneksiin V), mis seostub fosfatidüülseriiniga. Kasutades märgistatud anneksiini, võib apoptootilise raku nähtavaks muuta.
Kuna apoptootilises rakus DNA fragmenteerub, siis rakust eraldatud DNA analüüsimisel geel-elektroforeesil annab spetsiifilie redelitaolise mustri, mis on tingitud paljudest erineva pikkusega fragmentidest.
Mitokondrite sisemembraani membraanipotentsiaal muutub positiivsemaks. Seetõttu teatud negatiivse laenguga fluorestseeruvad värvid, mis kogunevad mitokondrites muudavad need mikroskoobis nähtavaks.
Mõõdeti
suure hulga rakkude DNA sisaldus. See kõikus vahemikus 3-6
pikogrammi tuuma kohta. Ühe konkreetse raku tuumas oli DNA sisaldus
5 pikogrammi. Millises rakutsükli faasis see rakk on? S faasis.
Kirjutage,
milline on kromosoomistik (n, 2n, 3n või 4n) rakutsükli erinevates faasides .
G1
faasis 2n. S faasi lõpust kuni mitoosi anafaasini on rakus
kromosoomide arv kahekordistunud (4n), see vahemik hõlmab S, G2, profaas , metafaas, anafaas . Telofaasis uuesti 2n.
Rakk
sisaldab mitoosi alguses 92 kromatiidi. Mitoosi lõppedes moodustub 2
rakku, milles kromosoomide arv on 46
Raku
keskosas algab fragmoplasti moodustumine ja raku vastasotstes on
moodustunud tuumad . Millise rakuga on tegemist?
1. loomarakk metafaasis
2. loomarakk
telofaasis
3. loomarakk
tsütokineesis
4. taimerakk metafaasis
5. taimerakk
tsütokineesis
Mitokondrid
Mitokondrite
suurus
Pikkus
1-2 mikromeetrit
Laius
0,1-0,2 mikromeetrit
Iseloomustage
mitokondrite DNA-d (% raku DNAst, molekuli suurus, struktuur,
geneetiline kood, geenide paiknemine , millised valgud kodeeritud)
Rõngasjas
struktuur, histoonide puudumine, operonide sarnaste struktuuride
esinemine (ühe ainevahetusrea struktuursed geenid tandeemselt). MtDNA suurus, tema poolt kodeeritud valkude arv ja tüüp, samuti
geneetiline kood varieerub erinevates organismides. DNA liigub
mirokondro ja tuuma vahel. Erinevates organismides on mitokondrite
genoomi suurus väga erinev (6000-300 000bp). Imetajates
kontrastina on mitokondrite genoom väga kompaktne ~16kb ja
mittekodeerivad geenide vahelised piirkonnad praktiliselt puuduvad.
Mitokondrite genoomi poolt kodeeritavad valgud on enamasti mtETA ja
ATP sünteesi valgud, samuti translatsiooni valgud ja ribosoomide RNA
ning tRNA. Eukarüootsete rakkude iga mitokonder sisaldab mitmeid
(1-50) rõngas mitokondriaalse DNA molekule. Mitokondrite
geneetiline kood erineb standartsest geneetilisest koodist, mida
kasutavad prokarüootsed ja eukarüootsed rakud (4 koodonit 64-st on
erinevad).
Kuidas
mitokondrid liiguvad eellasrakkudelt tütarrakkudele?
Sugulisel
paljunemisel organismi mitokondri pärinevad põhiliselt
emsasorgismilt ja väga väike osa (alla 1%) isasorganismilt. mtDNA
replitseerub nagu tuuma DNA interfaasis ja mitoosis tütarrakud
saavad ~võrdse arvu mitokondreid. Kuid mitokondrite jagunemine
toimub ka teistel rakutsükli etappidel, mitte ainult S faasis.
Mis
on tsütoplasmaatiline pärilikkus?
Sugulisel
paljunemisel organismi mitokondri pärinevad põhiliselt
emsasorgismilt ja väga väike osa (alla 1%) isasorganismilt.
Nimetage
vähemalt viis madalmolekulaarseid ühendeid transportivaid valke
mitokondrite membraanis ja mille jaoks nad on vajalikud.
- Poriinid. Moodustavad kanaleid, millest saavad vabalt läbi difuseeruda madalmolekulaarsed ühendid.
- Tsütokroomi c oksüdaasne kompleks transpordib elektrone ühekaupa redusteeritud tsütokroom c-lt hapnikule.
- Karnitiin /atsüülkarnitiin translokaas. Antsüülkarnitiini transport maatriksisse.
- Atsüülkarnitiini transferaas – atsüülkarnitiini transport läbi membraani.
- Püruvaat /OH-antiport – püruvaadi importsüsteem
- ADP/ATP-antiport süsteem
- Pi/OH-antiport süsteem
- Malaat /alfa-ketoglutaraat transportsüsteem (antiport) – tagab malaadi liikumise mitokondri maatriksisse.
- Enamik valkudest seostuvad Hsc70- nega , mis seostub mitokondri välismembraani retseptorite Tom20 ja Tom22-ga. Need resteptorid on seotud Tom40-ga, mis moodustab mitokondrisse sisenemise kanali. (TOM – translocator outer membrane)
Mis
on vahetu sama energiaallikas ATP sünteesil nii mitokondrites kui
kloroplastides elektronide transpordi ahelas.
PMF
– proton motive force . ATP sünteesiks kasutatakse elektronide
liikumisel tekkinud prootonite liikumapanevat jõudu.
Defineerige
pmf ja kirjutage valem. Millises mitok piirkonnas on prootoneid
rohkem, millises vähem, kust kuhu prootonid liiguvad ATP sünteesil.
Prootonite
liikumapanev jõud on prootonite elektrokeemilise potentsiaali
erinevus kahel pool mitokondrite sisemembraani. Prootoneid on rohkem
mitokondri membraanide vahelises ruumis (sisemembraani seinal ), mis
ATP sünteesil liiguvad sisemembraani seinast läbi maatriksisse.
pmf
=
- [(RT/F) * ΔpH] =
- 59 ΔpH
R
– gaasikonstant 8,315 J/( kraad mool )
T
– temperatuur Kelvini kraadides
F
– Faraday konstant (96.5 kJV-1mol-1)
-
membraanipotentsiaal
Kuidas
mitokondrites pmf tekib? Kui palju prootoneid transporditakse 2
elektroni liikumisel hapnikule?
Kõik
mtETA kompleksid va suktsinaat -CoQ reduktaasne kompleks,
funktsioneerivad ka kui prootonite transportijad mitokondri maatriksist membraanidevahelisse ruumi. Elektroni
seostumine/eraldumine muudab valguliste komplekside konformatsiooni
rohkem/vähem afiinseks prootonite suhtes.
Iga
elektronpaari liikumisel hapnikule transporditakse 4 prootonit
membraanide vahelisse ruumi.
Milline
valguline kompleks mitokondrites kasutab prootonite kontsentratsiooni
erinevuses peituvat energiat ATP sünteesiks?
ATP
süntaas (F-tüüpi H-ATPaas). Koosneb kahest subühikust Fo ja F1.
Kus
see kompleks mitokondris paikneb? Millises mitokondri piirkonnas ATP
sünteesitakse?
Paikneb mitokondri sisemembraanis. Fo subühik paikneb membraanis, F1
membraani maatriksi poolsel küljel.
Kuidas
ATP transporditakse mitokondrist tsütosooli?
ATP
transporditakse ATP/ADP translokaasi abil välja
Mitu
prootonit kannab suktsinaat-CoQ reduktaas membraani ühelt küljelt
teisele?
Mitte
ühtegi.
Milline
erinevus on substraatsel ja pmf põhisel ATP sünteesil?
Pmf
põhiline ATP süntees põhineb prootonite gradiendil kahel pool
membraani, substraatne ATP süntees aga kasutab substraadi
oksüdeerumist ja kõrge energiaga sidet omava vaheühendi
kasutamist.
Mis
põhjustab elektronide liikumise mitokondriaalses ETA-s, nimetage
peamised elektronide transpordis osalevad valgulised kompleksid.
Elektronide
liikumisel NADH/FADH2-lt
hapnikule iga vaheülekandja kõigepealt redutseerub omandades
elektroni ja seejärel oksüdeerub, andes elektroni järgmisele
vaheühendile. Seega iga ülekandja esineb vaheldumisi oksüdeerunud
ja redutseerunud vormis, mida nimetatakse redokspaariks. Mis määrab
redokspaaride järjekorra elektronide ülekandes, sõltub redokspaari
afiinsusest elektroni suhtes. Väiksema afiinsusega redokspaar annab
elektronid suurema afiinsusega redokspaarile. mtETA-s toimub seega
elektronide liikumine negatiivsema redokspotentsiaaliga
redokspaaridelt positiivsema redokspotentsiaaliga paaridele.
- NADH-CoQ reduktaasne kompleks (nn kompleks I)
- Suktsinaat-CoQ reduktaasne kompleks (kompleks II)
- CoQH2-cyt c reduktaasne kompleks (kompleks III)
- Cyt c oksüdaasne kompleks (kompleks IV)
Mis
toimub elektronide transpordi ja oksüdatiivse fosforüleerumise
lahutamisel?
Tooge
näiteid.
Siis
NADH oksüdeerumine ja hapniku redutseerumine toimuvad suure
kiirusega, aga ATP-d ei sünteesita. Elektronide ülekandel vabanev
energia hajub soojusena. Toimub teatud ainevahetus haiguste korral.
Teatud ained, mida nimetatakse lahutajateks (ingl uncouplers)
kaotavad prootonite gradiendi, muutes sisemembraani prootonite suhtes
läbitavaks
Kuidas
toimub tsütosoolis sünteesitud valkude liikumine mitokondrisse,
milline on mitokondrisse liikuva valgu signaaljärjestus?
Valk,
mis omab mitokondrisse suunamise signaali, seostub chaperon- idega ,
mis kasutavad ATP energiat valgu hoidmiseks kindlas lahtikeerdunud
seisunis. Ainult lahtikeerdunud valk saab siseneda. Enamik valkudest
seostuvad Hsc70-ga, mis soestub mitokondri välismembraanil
retseptoritega Tom20 ja Tom22. Need resteptorid on seotud Tom40-ga,
mis moodustab mitokondrisse sisenemise kanali. Sisenev valk liigub
läbi Tom40 kanali välismembraanis ja läbi täiendava kanali
sisemembraanis, mis moodustub Tim valkude vahendusel. Translokatsioon toimub ainult sellistes kohtades, kus välis- ja sisemembraan
puutuvad kokku. Transpordiks on vajalik pmf.
Mitokondrite
signaal paikneb ahela N-terminaalses otsas 3-5 Arg või Lys jääki
(mitte üksteise kõrval), sageli Ser ja Thr, mitte kunagi Glu ja
Asp, tavaliselt 20-60 aminohapet sisaldav piirkond, aga võib olla ka
lühem.
Mitokondriaalses
elektronide transpordi ahelas ( mETA ) liigub NADH oksüdeerumisel
vabanenud 2 elektroni. Kui palju prootoneid liigub läbi mitokondri
sisemembraani 2 elektroni liikumisel? Põhjendage.
4NADH-CoQ
reduktaasse kompleksi CoQ sidumistsentrisse (paikneb maatriksi
poolsel küljel) seostub CoQ molekul ja seob maatriksist 2 elektroni
ja 2 prootoni.
Redutseerunud
CoQH2
difundeerub membraanis kuni seostub CoQH2-cyt
c reduktaase kompleksiga ja annab ära 2 prootonit mitokondri
membraanidevahelisse ruumi (ja 2 elektroni). Elektronidest üks
liigub Fe-S valgu ja cyt c1 vahendusel tsütokroomile c, teine elektron (nn ringlev elektron) liigub cyt b566 ja b562 vahendusel
teise CoQ sidumistsentrini redutseerides seotud CoQ (moodustub
semikioon). Kui teine ringlev elektron, mis vabaneb teiselt CoQH2-lt
liigub semikioonile, siis veel 2 prootonit liiguvad maatriksist
membraanidevahelisse ruumi ja tekib CoQH2.
Redusteerunud CoQ vabastab täiendavalt 2 prootonit
membraanidevahelisse ruumi, üks liigub cyt c-le, teine tsükliseerub
jne.
Milliste
mitokondri piirkondade vahel ja kust kuhu prootonid liiguvad? Miks
tehakse prootonite liikumise arvutused kahe, mitte ühe elektroni
liikumise kohta?
Elektronid
liiguvad maatriksist läbi sisemembraani membraanidevahelisse ruumi.
Kahe
prootoni kohta, sest NADH-CoQ reduktaasne kompleks katalüüsib kahe
elektroni liikumist NADH-lt CoQ-le.
Loomaraku
hingamisel neelduvat hapnikku kasutatakse otseselt (Õige variant
alla kriipsutada).
a) Glükolüüsis
b) elektronide
aktsepteerimiseks mitokondriaalse ETA lõpus
c) tsitraaditsüklis, NADPH genereerimise reaktsioonides
d) tsitraaditsüklis,
püruvaadi oksüdeerimisel atsetüülCoA-ks.
Tsitraaditsükkel
produtseerib kõrge energiasisaldusega ühendeid, mis transpordivad
energia mitokondriaalsesse elektronide transpordi ahelasse (mETA),
sest neid oksüdeeritakse mETAs. Need ühendid on: (õige variant
alla kriipsutada)
a. ATP
ja CO2
b. CO2
ja FADH2
c. FADH2
ja NADH
d. NADH
ja ATP
e. FADH2,
NADH ja ATP
Kloroplastid
Kloroplastide
suurus.
Diameeter : 2-10 mikromeetrit
Paksus
: 1 mikromeeter
Kloroplastide
peamised kompartmendid ja membraanid.
Ümbritsetud
kahe membraaniga – sisemembraan ja välismembraan.
Lisaks
ka kolmandat tüüpi membraan : tülakoidide e. Lamellide membraan.
Strooma
– kloroplastide sees väljaspool tülakoide paiknev piirkond
Luumen – tülakoidide siseruum .
Fotosünteesi
valgusstaadiumi reaktsioonid ja produktid, millises kloroplasti osas
toimuvad.
- Valguse absorptsioon . Valgusenergia neeldumine ja selle muutmine keemiliseks energiaks toimub pigment -valk kompleksides, mida nimetatakse fotosüsteemideks ja mis paiknevad tülakoidide membraanides.
- Elektronide transport, millega kaasneb NADP redusteerumine NADPH-ks. Toimub tülakoidide membraanis vist .
- ATP süntees, mis toimub pmf arvelt stroomas .
Vee
fotooksüdatsiooni valem, millises kloroplasti piirkonnas toimub,
tekkinud produktide kasutamine.
2H2O
+ 4P+
4H+
+ O2
+ 4P
Vee
fotooksüdatsioonil vabanevad prootonid liiguvad tülakoidide
luumenisse, elektronid liiguvad oksüdeerunud klorofülli kaudu
kinoonile ja lõpuks NADP-le NADPH tekkega.
Fotosüsteemi
ehitus, mis toimub fotosüsteemis, kus paikneb. Milline protsess
toimub fotosüsteemi tsentris?
Fotosüsteem on pigment-valk kompleks, mis paikneb tülakoidide membraanides. Iga
fotosüsteem koosneb kahest osast – antennist (valkudega seotud
valgust neelavad pigmendid) ja reaksioonitsentrumist (valkude
kompleks + 2 klorofüll a molekuli + seos elektroni doonori ja
akspetoriga). Antenn sisaldab ühte või mitut valgust koguvat
kompleksi (LHC), mille pigmendi molekulid absorbeerivad valguskvante.
Valgusenergia liigub antennist reaktsioonitsentrumis paiknevatele
klorofüll a molekulidele, millede vahendusel toimub valgusenergia
konverteerimine keemiliseks energiaks.
Kuidas
tagatakse elektronide liikumine fotosünteetilises ETA-s, millised
valgulised kompleksid osalevad.
Elektronide
liikumine fsETA-s on tagatud kahe fotosüsteemi (PSI ja PSII ) töö
kooskõlastatusega. Et toimuks elektronide liikumine veelt NADP-le,
on vajalikud mõlemad fotosüsteemid. Ainult PSII ergastumisel
footoni energiast suur osa kulub süsteemisisesteks muutusteks ja
elektroni energiast ei piisa NADP-le liikumiseks. Täiendava footoni neelamine PSI poolt tõstab elektroni energia nii kõrgeks, et NADP
redusteerumine muutub võimalikuks.
Kuidas
ja kus valgusenergia muutub keemiliseks energiaks fotosünteesi
protsessis?
Reaksioonitsentris
toimub ülekantud valgusenergia toimel laengute lahutamine. Selle all
mõistetakse ergastunud elektroni eraldumist klorofülli molekulist
ja liikumist reaksioonitsentri kõrval paiknevasse elektroni
akseptori, milleks fotosüsteem II-s on kinoonimolekul, koostisesse
st toimub kinoonimolekuli redutseerumine. Ergastamata klorofülli
molekul ei ole piisavalt tugev redutseerija , et anda elektron
kinoonile. Klorofülli molekulile jääb elektroni eemaldumise tõttu
positiivse laenuga auk, selline klorofüll on hea oksüdeerija ja
võtab elektroni ära tülakoidi luumenipoolsel küljel paiknevalt
elektroni doonorilt. Neli valguskvanti on vaja, et eemaldada neli
elektroni kahest vee molekulist hapniku vabanemisega.
Mitu
H+
transporditakse 2H20
oksüdeerumisel vabanenud elektronide liikumisel fs ETA-s, kui palju
ATP-d on võimalik selle arvel sünteesida?
Transporditakse
12 prootonit (kahelt vee molekulilt saadud neli elektroni
transpordivad 8 ning sellele lisanuvad kahelt vee molekulilt
vabanenenud neli prootonit) ning sünteesitakse selle arvel 3 ATP-d.
Pmf
suurus kloroplastides.
30mV
Fotosünteesi
biokeemiline staadium, millised faasid eristatakse. Millises faasis
on vajalik ATP ja NADPH, milleks?
Fotosünteesi
biokeemililiseks staadiumiks on Calvini tsükkel .
I
faas, II faas, III regenereerimisfaas.
II
faasis on vaja ATP-d 1,3-bifosfoglütseraadi sünteesiks. III faasis
on vaja NADPH-d 1,3-bifosfoglütseraadi redutseerumiseks.
Millises
kloroplastide osas CO2 sidumise ja redutseerumise reaktsioonid toimuvad?
Millist
reaktsiooni katalüüsib ensüüm Rubisko .
Kloroplastide
stroomas, Calvini tsüklis .
Rubisco e. Ribuloos 1,5-difosfaadi karboksülaas katalüüsib süsihappegaasi
seostumist ribuloos 1,5-difosfaadile (RubP)
Kloroplasti
DNA ehitus, suurus, milliseid valke kodeerib.
Kloroplastid
sisaldavad mitmeid kuni mitmeid kümneid rõngas-DNA molekule
suurusega 120 000 – 160 000 bp. Sisaldab ~120 geeni, 60
neist on seotud RNA transkriptsiooni ja translatsiooniga (rRNA, tRNA,
RNA polümeraasi ja ribosoomi valkude geenid). ~20 geeni kodeerivad
fsETA valke. Kloroplastides on kodeeritud ka Rubisco suur subühik.
Esineb
~40
geeni, mille funktsioon pole teada.
Fotosünteesi
protsessis vesi oksüdeeritakse ja süsinik ( süsihappegaas )
redutseeritakse. Kirjutada sisse õige variant.
Nimetage
kloroplastis funktsioneerivaid subühikutest koosnevaid valke, mille
mõned subühikuid on kodeeritud tuuma genoomis, mõned kloroplasti
genoomis.
Ribosoomi
valgud, Rubisco väike subühik, Calvini tsükli ensüümid.
Mikrofilamendid
Aktiini
omadused. Aktiinimolekuli polaarsus ja seotus ATP/ADP-ga, + ja –
ots.
Aktiin on eukarüootse raku kõige levinum valk.
Kodeeritud
geenide perekonna poolt (inimesel nt 6 geeni)
Konserveerunud
geenid
Molekulis
~375 aminohappe jääki.
Esineb
kahel kujul :
Globulaarne monomeer (G-aktiin)
Niitjas polümeer G- aktiinist (F-aktiin)
Aktiinimolekul
ja polümeer on polaarsed, - otsana tähistatakse ATP-d siduva
vagumusega otsa ja vastaspiirkonda tähistatakse + otsana, kuhu
eelkõige toimub subühikute liitumine polümerisatsiooni korral.
Iga
aktiini molekul sisaldab Mg iooni ja ATP-d või ADP-d. Seega
eksisteerivad mitmesugused aktiini vormid : ATP-G-aktiin,
ADP-G-aktiin, ATP-F-aktiin ja ADP-F-aktiin. Neist levinumad on
ATP-G-aktiin ja ADP-F-aktiin. Mg, K, Na ioonide juuresolekul lahuses
G-aktiin polümeriseerub F-aktiini pikkadeks ahelateks ATP
hüdrolüüsumisega. Monomeerid asetsevad ahelates helikaalselt. Kui
lahuse ioonne jõud väheneb, siis F-aktiin depolümeriseerub.
Kriitilise
kontsentratsiooni mõiste ja suurus.
Kriitiliseks
kontsentratsiooniks nimetatakse kontsentratsiooni, mille juures
polümeriseerumisel liituvate subühikute arv võrdub
dissotsieerumisel vabanevate subühikute arvuga. In vivo tingimustes
~0,1 mikromolaarne.
Nimetage
tegureid, millest sõltub aktiini polümeriseerumine.
- Ioonne jõud
- Monomeeride kontsentratsioon
- Mitmesugused toksiinid (nt seenest pärit alkaloid tsütohalasiini toimel aktiini fibrillid depolümeriseeruvad ja latrunkuliin käsnadest seostub G-aktiiniga ja takistab subühikute polümeriseerumist. Toksiin falloidiin seenest seostub F-aktiiniga, ühendades naabermonomeerid nii tugevalt, et nende dissotsieerumist ei toimu isegi siis kui G aktiini kontsentratsioon väheneb alla Kk).
- Rakkudes on filamentide pikkus, stabiilsus, nende omavaheliste komplekside geomeetria reguleeritud mitmete täiendavate valkude poolt.
Profiliin (soodustab polümeriseerumist)
Tümosiin beeta4 (takistab polümeriseerumist)
Gelsolin ja kofilin (ahelat lagundava toimega)
CapZ – seostub + otsaga ja takistab uute monomeeride lisandumist ning vanade eemaldumist (stabiliseerib)
Tropomoduliin – sama, mis CapZ, aga seostub filamentide – otstega.
Nimetage
peamised valkude klassid mis osalevad aktiinifilamentidest
moodustuvate struktuuride tekkes.
- Aktiiniseoselised valgud (ARP – actin related proteins) – on olulised aktiini filamentide tekkes nukleatsiooni tsentrite loomises.
- Aktiini ristseoselised valgud – vajalikud nii kimpude kui ka võrgustiku püsimiseks.
Aktiini filamentide kimpude moodustumise tagavad lühikese ahelaga ristseoselised valgud (fimbriin filopoodides ja villiin mikrohattudes)
Võrgustiku moodustumise tagavad pika ahelaga valgud (alfa-aktiin tagab võrgustiku moodustumise stressi fiibrites, filamiin lamellipoodides).
Nimetage
aktiini filamentidest moodustuvaid struktuure.
- Kimbud (filamendid paiknevad paralleelsetes kimpudes)
- Võrgustik (filamendid ristuvad)
Aktiini
mootorvalkude – müosiinide - struktuur
- Rasked ahelad . Neid võib olla 1-2 sõltuvalt müosiini tüübist. Kahe ahela esinemisel need on alfa- heeliksi konformatsioonis üksteise ümber põimunud (koos hoiavad perioodiliselt esinevad hüdrofoobsed vastasmõjud). Koosnevad kolmest struktuurilt ja funktsioonilt erinevast domäänist :
Pea – globuraalse struktuuriga, N- terminaalne . Sisaldab seostumiskohta aktiinile (P-ling), seostumiskohta ATP jaoks.
Kael – ümbritsetud kergete ahelatega ja võib olla seotud müosiini regulaatorvalkudega.
Saba – C-terminaalne, sisaldab seostumiskohti, mis tagavad spetsiifilisuse.
- Kerged ahelad. Ca sidumine.
Peamised
etapid müosiin -aktiin interaktsioonis.
Nukleotiidi seostumine – müosiin on setud aktiiniga, ATP seostumiskoht vaba, toimub ATP seostumine, aktiini vagumus vabaneb ja pea dissotsieerub aktiini filamendist.
Hüdrolüüs – pea pöördub, kaela konformatsioon muutub (vagumus sulgub) ja müosiini molekul seostub uuesti aktiini filamendiga, aga subühikuga, mis paikneb –otsast kaugemal.
Pi vabanemine – pea pöördub ja liigutab filamenti edasi – ots ees, sest filament on peaga seotud ja konformatsiooniline muutus kaelas liigutab filamenti.
ADP vabanemine – ja esialgse konformatsiooni nn kangestusseisnud taastumine .
Lihasrakkude
ehitus ja kontraktiilsuse printsiip
Lihasrakk
koosneb müofibrillidest, mis kujutavad endast aktiini filamentide
kimpe ja jaotuvad heledateks ja tumebdateks ribadeks piki lihasrakku.
Filamentide + otsad kinnituvad valgulisele kettale, mida tähistatakse
kui Z ketast . Silindrilist osa müofibrillis kahe Z joone vahel
nimetatakse sarkomeeriks. Seega müofibrill koosneb sarkomeeride ahelast . Sarkomeerid on skeletilihase struktuurseks ja
funktsionaalseks ühikuks. Iga sarkomeer koosneb kahte tüüpi
filamentidest :
- Paksud filamendid – müosiin II
- Peened filamendid – aktiin
Lihaste
kokkutümbel müosiini pead astuvad aktiini fibrillide + otsa poole
(Z ketta poole), samal ajal müosiini ega aktiini ahelate pikkus ei
muutu.
Millistes
protsessides lisaks lihasrakkude kontraktsioonile veel osaleb
aktiin/müosiin kompleks.
Aktiini
filamendid funktsioneerivad rakus kui rööpad, mida mööda ATP-st
sõltuvad mootorvalgud (müosiinid) saavad liigutada raku komponente.
Tsütokineesis
osalemine – moodustavad jagunevas rakus ekvatoriaaltasapinnas
kontraktiilse ringi.
Kuidas
närviimpulsi jõudmine mootorneuronini põhjustab sarkomeeride
kokkutõmbe. Ca2+
osa. Kui
närviimpluss jõuab lihasrakuni, muutub rakumembraani elektriline
potentsiaal (toimub depolariseerumine). Signaal liigub spetsiifilises
signaali ülekande ahelas SR-i Ca kanalite valkudeni, mille
tagajärjel kanalid avanevad ja Ca kontsentratsioon tsütosoolis
suureneb. Ca toimet vahendavad aktiiniga seostuvad valgud
tropomüosiin ja troponiin . Ca kontsentratsiooni kasvades muutub
nende valkude asetus aktiini filamentidel ja vabanevad aktiini
müosiiniga seostuvad piirkonnad, mis võimaldab müosiini liikumist
piki aktiini filamenti.
Mootorvalk
liigub tsütoskeleti + otsa suunas. Millise mootorvalguga võiks olla
tegemist.
Kinesiin .
Mootorvalk
liigub tsütoskeleti valgu – otsa suunas. Millise mootorvalguga
võiks olla tegemist.
Düeniin.
10
ATP hüdrolüüs võimaldab müosiinil II liikuda 50-100nm kaugusele
Mikrotorukesed
Aktiini
ja tubuliini võrdlus (mille poolest erinevad ja mille poolest
sarnanevad)
- Mõlemate filamentsed vormid koosnevad monomeeridest
- Tubuliini polümeer on torujas, aktiini polümeer alfa-heeliksi sarnase struktuuriga
- Tubuliini on heterodimeer, subühikuks alfa ja beeta tubuliin. Aktiini monomeer koosneb ühest subühikust.
- Tubuliini iga subühik seob ühe GTP, kokku siis kaks. Aktiin seob ATP-d.
- Mõlemad on polaarse ehitusega. Aktiinil + ja – ots, tubuliini puhul alfa ja beeta ots.
Mis
on treadmilling ja millistes tingimustes toimub
On
ühe konkreetse subühiku liikumine +otsast –otsa poole läbi
filamendi.Toimub juhul kui subühikud liituvad + otsale sama kiiresti
kui eemalduvad – otsalt.
Nimetage
vähemalt 4 tegurit, mis mõjutavad mikrotorukeste polümeriseerumist
ja depolümeriseerumist.
- Nukleatsiooni tsentrite esinemine
- Mikrotorukeste ja subühikute kontsentratsiooni tasakaal
- Kolhitsiin (alkaloid sügislillest) ja vinblastiin (igihali) soestuvad pöördumatult alfa-beeta dimeeriga ja takistavad mikrotorukeste moodustumist
- Taksool (jugapuu) takistab mikrotorukeste depolümeriseerumist.
- MAP (microtubule associated proteins) valkude olemasolu.
Millistes
protsessides osalevad mikrotorukesed ja nende mootorvalgud.
- Rakusisene transport, vesiikulite
- Kromosoomide eraldumine mitoosis ja meioosis
- Viburite liikumine
Millistes
kohtades rakus toimub mikrofilamentide ja millistes mikrotorukeste
polümeriseerumine, millised valgud vajalikud?
Mikrotorukeste
polümeriseerumine toimub rakus mikrotorukesi organiseerivates
tsentrites (MTOC) rakutuuma lähedal. Nendes esineb nt gamma
tubuliini.
Aktiini
filamentide kokkupanek, nukleatsiooni tsentrite valkude seotuse tõttu
membraaniga, toimub sageli rakumembraani läheduses. Aktiini
filamentide tekkes on nukleatsioonitsentrite loomises olulised nn
aktiiniseoselised valgud (ARP).
Kus
ja kuidas toimub loomarakus mikrotorukeste moodustumine?
Mikrotorukesed on selle struktuuriga seotud kas (-) või (+) otsa
vahendusel? (alla kriipsutada õige variant).
Tsentrosoomid
on kohaks, kus algab mikrotorukeste moodustumine nii, et –otsad
jäävad seotuks tsentrosoomiga. Seega subühikute lisandumine toimub
mikrotorukeste +otsale.
Kirjeldage
mikrotorukeste mootorvalke ja iseloomustage (sammu pikkus, liikumise
suund mikrotorukesel)
Mikrotorukeste
mootorvalgud saab jagada kinesiinideks (+suunalised) ja düeniinideks
(-suunalised).
Kinesiini
molekul on dimeer kahest raskest ja koahest kergest ahelast. Molekul
koosneb kolmest domäänist : 1. Globulaarsed pead mõlemil raskel ahelal (seostumine mikrotorukestega) ; 2. Raskete ahelate
superspiraliseerunud osa ; 3. Rakete ahelate terminaalne osa, mis
seostub kergete ahelatega ja transporditava vesiikuliga. Sammu pikkus
8nm.
Düeniinid
koosnevad 2-3 raskest ahelast, mis on komplekseerunud kergete
ahelatega. Tavaliselt kaks pead. Düeniini molekul üksi ei ole
suuteline liikumist tekitama. Et toimuks düeniini seostumine
vesiikuliga, on lisaks vajalikud teised mikrotorukesi siduvad valgud,
mis seovad vesiikulid ja kromosoomid mikrotorukestega. Ilmselt sama
pikk samm umbes.
Kirjeldage
lühidalt aksoneemi ehitust ja nimetage piirkondi inimese organismis,
kus sellised struktuurid paiknevad. Kuidas aksoneemi struktuur tagab
liikumise?
Aksoneemid
on mikrotorukestest koosnevad struktuurid, mis moodustavad membraani
väljakasve. Aksoneem koosneb üheksast välimisest mikrotorukeste
paarist, mis ümbritsevad kahte ühekaupa paiknevat mikrotorukest.
Kõikide mikrotorukeste + ots paikneb aksoneemi raku tsentrist
kaugemas osas. Iga välimiste torukeste paar koosneb A ja B
torukesest.
Inimese
organismis nt hingamisteede epiteelirakkude ripsmed .
Liikumises
osaleb düeniin ja tema vahendusel toimub ühe paari A torukese
liikumine teise paari B torukese aluse, - otsa poole. Kuna esinevad
ka teised sidemed mikrotorukeste paaride vahel, siis sirgjooneline
liikumine muutub lainetavaks.
Millised
haigused milliste tunnustega on seotud aksoneemi
häiretega.(Kartageneri sündroom )
Inimesel
mutatsioonid viburite/ ripsmete düneiinis võivad põhjustada nn
Kartagener’i sündroomi, mida iseloomustab isasteriilsus
(spermatosoidid ei liigu) ja samuti vastuvõtlikkus
kopsuinfektsioonidele, sest liikumatud ripsmed ei eemalda
hingamisteede pinnalt baktereid
Kinetohoorsed,
polaarsed ja astraalsed mikrotorukesed, nende funktsioonid mitoosis,
milliste mootorvalkude abil need realiseeruvad?
- Kinetohoorsed mikrotorukesed seostuvad kromosoomidega ja osalevad kromatiidide eraldamises üksteises ja jaguneva raku poolustele liikumises. Mootorvalkudeks kinesiinid ja düneiinid
- Polaaresd mikrotorukesed on seotud vastastsentrosoomist lähtuvate polaarsete mikrotorukestega. On seotud pooluste üksteisest kaugemale liikumisega anafaas B-s.
- Astraalsed mikrotorukesed, mis paiknevad kiirjalt ümber tsentrosoomi, osalevad mitoosivärtna pikemaks venitamises anafaasis. Mootorvalkudeks düeniinid.
Tsentrioolide
funktsioonid rakutsüklis, tsentrioolide tsükkel.
Kahest
tsentrioolist tekib neli enne profaasi. Mitoosis liiguvad tsentrioolid raku poolustele. Osaleb ka tsütokineesi lõpule
viimises. Rakutsüklis uus tsentriool kasvab iga ematsentriooli
küljest. Nad jäävad ühendatuks mitoosi alguseni. Tsentriooli
duplikatsioon algab G1 ja S faasi vahel. Osaleb raku jagunemises
mitoosi värtna moodustumises ja funktsioneerimises
Defineerige tsentrosoom , tsentromeer, kinetohoor.
- Tsentrosoom – valgulist materjali sisaldav piirkond, mis ümbritseb tsentrioole ja mis interfaasis tagab mikrotorukeste moodustumise.
- Tsentromeer – kromosoomi DNA järjestus, millele seostub valguline kompleks kinetohoor. Tsentromeer seob õdekromatiidid omavahel.
- Kinetohoor – valguline kompleks, mis soestub kromatiidi tsentromeeri piirkonnaga ja tagab mikrotorukeste seostumise kromatiidi külge mitoosivärtna moodustumisel.
Nimetage
tsentrioolide funktsioone rakkudes lisaks mitoosile .
- Osalevad eukarüootides ripsmete ja viburite moodustamises
Nimetage
3 taimeraku mitoosi iseärasust võrreldes loomarakuga
- Taimerakus moodustub raku jagunemistasapina fragmoplast Glogi kompleksi vesiikulitest ja neid transportivatest mikrotorukestest, mille kokkusulamisel tekib uus rakusein .
- Taimedes tsentrioolid puuduvad.
- Taimedes esineb preprofaasi riba.
Kirjeldage
kinetohoori ehitust.
Kinetohoor
on kihilise ehitusega, võib eristada sisemist (kromosoomi poolset)
ja välimis (mikrotorukeste poolset) plaati , mis on omavahel teatud
valkudega ühendatud.
Kromosoomide
liikumine anafaasis a ja b
- Anafaas A (varajane anafaas) – kinetohoorsed mikrotorukesed lühenevad ja kromosoomid tõmmatakse poolustele (mitoosivärtna pikkus konstante )
- Anafaas B ( hiline anafaas) – poolused liiguvad üksteisest kaugemale (mitoosivärten pikeneb)
Millistes
mitoosi protsessides osalevad kinesiinid ja millistes düneiinid,
millistes tubuliini polümeriseerumine ja depolümeriseerumine
- Kinesiinid osalevad anafaasis B mitoosivärtna pikenemises
- Düeniinid osalevad metafaasis toimuva kromosoomide pendeldamises. Anafaasis toimuvas kromosoomide liikumises ekvaatorilt poolustele. Anafaasis B toimuvas värtna pikenemises (astraalsete mikrotorukeste tõmbamises)
- Tubuliini depolümeriseerumine osaleb anafaasis B toimuvas värtna pikenemises.
Mootorvalk
liigub tsütoskeleti (-) otsa suunas. Millise mootorvalguga võiks
olla tegemist?
Düeniin
Millises
mitoosiprotsessi osas sellised mootorvalgud funktsioneerivad?
Düeniinid
osalevad metafaasis toimuva kromosoomide pendeldamises. Anafaasis
toimuvas kromosoomide liikumises ekvaatorilt poolustele. Anafaasis B
toimuvas värtna pikenemises (astraalsete mikrotorukeste tõmbamises)
29. Milliste
mootorvalkude ja milliste tsütoskeleti valkude interaktsioon toimub
loomaraku tsütokineesis?
Müosiin
ja aktiin – kontraktiilne rõngas.
Tsütohalasiin
B on mikrofilamentide moodustumist takistav ühend. See ühend
seetõttu takistab tsütokineesi.
Meetodid
rakubioloogias
Milliseid
geenitehnoloogilisi meetodeid kasutatakse valgu lokalisatsiooni
muutuse jälgimiseks elusas rakus?
Valk
märgistatakse fluorestseeruva aminohappe järjestusega, mida valk
endaga kaasas veab ja mida on fluorestsentsmikroskoobis näha.
Kuidas
toimite, kui tahate näha ühe spetsiifilise valgu lokalisatsiooni
rakus?
Monoklonaalsete
antikehade kasutamine.
Mille
poolest erineb uuritavast objektist tavalise fluorestsentsmikroskoobi
abil saadav kujutis konfokaalse fluorestsentsmikroskoobi abil
saadavast kujutisest?
Võimaldab
kõrgkvaliteedilist kujutist ka suhteliselt paksust koematerjalist,
ei ole vaja teha üliõhukesi lõike. Kujutis tekib ainult sellest
tasapinnast, mis on fookuses . Fookusest väljas olevad piirkonnad
jäävad mustaks, neid ei näe. Kui teha uuritavast objektit suur
hulk opitilisi lõike, saab arvuti abil need sünteesida
kolmemõõtmeliseks kujutiseks.
Kuidas
tuleb uuritavat preparaati töödelda, et see oleks vaadeldav
skanneeriva elektronmikroskoobiga?
Proovid kaetakse üliõhukeste elektrir juhtiva materjalikihiga, kasutades
madal- vaakum pinnakatmist või kõrg-vaakum aurufaassadestust.
Juhtivate kattematerjalidena on tänapäeval kasutusel kuld , kulla ja
palladiumi sulam , plaatina jne.
Milliste
meetoditega on võimalik apoptoosi näidata.
DNA fragmenteerumisel DNA ahelasse tekkivaid katkemiskohti on võimalik tuvastada spetsiaalse ensüümi – terminaalse desoksünukleotidüül transferaasi (TdT) abil (nn TUNEL reaksioon). See ensüüm tunneb ära vabad 3’-OH otsad ning pikendab neid nukleotiididega. Kui reaksioonikeskkonda lisada radioaktiivse või fluorestsentsmärgistusega nukleotiide, siis sisestuvad need märgistatud nukleotiidid sünteesitavasse DNA-sse.
Normaalsetes rakkudes on plasmamembraan ebasümmeetriline fosfolipiidide suhtes. Fosfatidüülseriin eksponeerub ka membraani tsütoplasmaatilises pooles. Apoptootilises rakus fosfatidüülseriin eksponeerub ka memrbaani ekstratsellulaarsel
Peroksüsoomid, lüsosoomid , proteasoomid
Lüsosoomide
struktuur, suurus, ehitus, toimuvad protsessid
Ühe
membraanikihiga ümbritsetud organellid
Diameeter
: 0.2-0.5 mikromeetrit
Luumeni
pH on happeline (4-4,5). Happelise pH tagab V-tüüpi H/ATPaas
lüsosoomide membraanis, pumbates prootoneid tsütoplasmast
lüsosoomi.
Lüsosoomid
on loomarakkudes üheks piirkonnaks, kus toimub kõige erinevamate
ühendite lagundamine. Lüsosoomidesse liiguvad ja lagundatakse seal
ka endotsütoosi ja fagotsütoosi teel rakkudesse sattuvad ühendid
ja organismid, aga samuti teatud ERst punguvates vesiikulites olevad
ained. Lüsosoomid on seotud ka vananenud ja mittefunktsioneerivate
organellide lagundamisega.
Kuidas
kaitseb lüsosoom enda membraani hüdrolaaside lagundava mõju eest?
Lüsosoomide
membraanis esinevad valgud on tugevasti glükosüleeritud.
Nimetage
vähemalt kaks lüsosomaalset ladestushaigust ja kirjeldage
mehhanism, mis haiguse tingib.
Harvaesineva
I rakulise haiguse (ingl. Inclusion-cell disease ) korral puuduvad
praktiliselt kõik lüsosomaalsed ensüümid lüsosoomi transportiva
süsteemi häirete tõttu. Defektne on ensüüm, mis katalüüsib
lüsosoomi liikuvate valkude fosforüülumist. Seetõttu Golgi
kompleksi retseptorid ei tunne vastavaid valke ära ja need
transporditakse rakkudest välja, mitte lüsosoomidesse. Esinevad
veres. Lagundamata ained kogunevad rakkudes lüsosoomidesse
inklusioonidena. Lõpeb surmaga varases lapseeas
Tay- Sachs haigus on põhjustatud gangliosiide (glükolipiidid) lagundavate
ensüümide puudusest (inaktiivsusest).
Toimub gangliosiidide kuhjumine lüsosoomides. Selle haigusega vastsündinud
on esimese aasta jooksul normaalsed, kuid siis ilmnevad haigusele
iseloomulikud sümptomid – nõrkus, üldise ja vaimse arengu
aeglustumine, nägemishäired. Vanuses 3-4 a lapsed surevad. Haigus
on pärilik.
Pompe
haigus – puudub -gükosidaas,
mis vajalik glükogeeni lagundamiseks.
Peroksüsoomide
struktuur, suurus, ehitus.
Väikesed
ühekordse membraaniga organellid (0,2-1 mikromeeter diameeter)
Loetlege,
millised biokeemilised protsessid toimuvad peroksüsoomides?
- Pikaahelaliste rasvhapete oksüdeerimine
- Puriinide , prostaglandiinide katabolism
- Hapniku radikaalide metaboliseerumine
- Alkoholi jt toksiliste ainete lagunemine
- Sialdavad flavoproteiinseid oksüdaase, mis oksüdeerivad orgaanilisi ühendeid hapniku kasutamisega
- Vesinikperoksiidi lagundamine (katalaas)
Kuidas
peroksüsomaalsed valgud liiguvad tsütosoolist peroksüsoomidesse?
Tsütoplasmaatilistel
ribosoomidel sünteesitud peroksüsoomide valgud sisaldavad
peroksüsoomi transpordi signaali, mis võib paikneda C-terminaalses
aga harvem ka N-terminaalses otsas. Peroksüsoomi transpordi signaal
(PTS) seostub tsütosoolis esineva retseptorvalguga. Moodustunud
kompleks seostub peroksüsoomi membraani retseptoriga ja
transporditakse peroksüsoomi.
Kirjeldage
peroksüsoomide funktsioneerimise häiretega kaasnevaid haigusi
inimesel
Zellwegeri
sündroomi korral on takistatud enamiku valkude liikumine
tsütosoolist peroksüsoomidesse, seega peroksüsoomid on tühjad. On
leitud, et mutatsioonid kaheksas geenis (seotud impordivalkudega)
võivad sellise sündroomi põhjustada. Haigetel esinevad maksa ja
neerude talituse häired ja ajukahjustused, mis põhjustab surma
varases lapseeas.
Inimesel
esineb pärilik haigus nimega X-seoseline adrenoleukodüstroofia
(ALD). Selle haiguse puhul on häiritud eriti pikaahelaliste
rasvhapete oksüdeerumine peroksüsoomides. Toimub küll selliste
rasvhapete transport peroksüsoomidesse, kuid seostumine CoA-ga
mitte, sest ensüüm, mis katalüüsib rasvhappe esterifitseerumist
CoA-ga (rasvatsüül – CoA süntaas) transpordisüsteemi defektsuse
tõttu ei liigu peroksüsoomidesse. Ensüümi tugevalt väljendunud
puuduse korral lapsepõlve keskel tekivad tõsised neuroloogilised
häired, maksa ja neerude kahjustused, millele järgneb surm mõne
aasta möödudes .
Kirjeldage
proteasoomide ehitust, lokalisatsiooni rakus ja nendes toimuvaid
protsesse.
Koosnevad
tsentraalsest 20S südamikust, mis omakorda koosneb neljast
valguliste subühkite rõngast. Südamiku mõlemis otsas paiknevad
19S ’mütsikesed’.
Iseloomustage
chaperon’id(Hsp70 rühm) ja nimetage protsesse milles osalevad.
Valgud,
mis stabiliseerivad ebastabiilsed konformatsioonid, kiirendavad
oligomeeride teket, valkude lagunemist ja tagavad lokalisatsiooni
kindlates raku piirkondades. Hoiavad valke lahustunud kujul, mis on
valmis kokkukeerdumiseks. Kaitsevad proteolüüsi eest. Vajalik ATP.
On olulised rakkude liikumisel läbi membraanide. Paljud valgud
omandavad natiivse konformatsiooni ainult seda tüüpi valkude abil.
Iseloomustage
chaperonin´id ja nimetage protsesse milles osalevad.
Tagavad
valkude kokkukeerdumise. Mõjuvad struktuuri tekke hilisematel
staadiumitel. On oligomeersed valgud, mis moodustavad sisemusse
kanali, kus valgu konformatsiooni teke toimub. Vajalik ATP.
Reguleerib oligomeersete komplekside teket.












































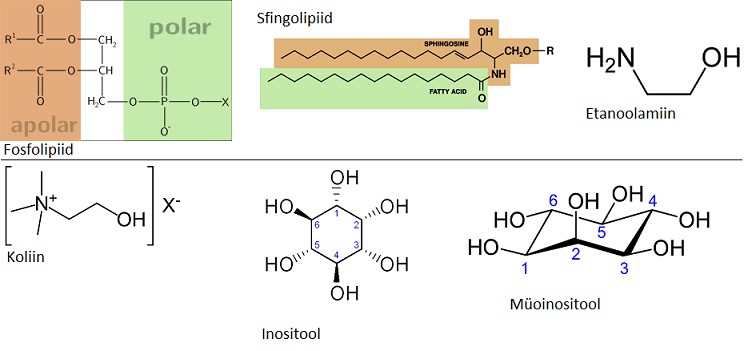




Kõik kommentaarid