Sissejuhatus1.
Gram + ja Gram- bakterite rakuseina ehitus ja esindajad
Gram+
- peptidoglükaanide kiht, teihoiinhape (ioonide liikumine, kaitse,
antigeenne
spetsiifilisus);
1
membraan +paks sein,
Bacillus polymyxaLearn
moreGram-
- peptidoglükaanide kiht, teihoiinhape puudub; välismembraanil on
LPS (lipopolüsahhariidid)
(endotoksiin),
poriinid ja see kaitseb ksea; 2 membraani+õhuke sein,
E.
coli2.
Prokarüoodi raku ja genoomi suurus
Prokarüoodi
rakk on 1μm
-
10μm.
400-4000 geeni
3.
Eukarüoodi raku ja genoomi suurus
Eukarüoodi
rakk on 5μm
- 100μm.10000-
40000 geeni
4.
Nimetage prokarüoodi (eubakter) ja eukarüoodi raku peamised
erinevused
Prokarüoot (
Bakterid +arhed)
Eukarüoot (Taimed, loomad, seened, protistid)
Raku suurus
Organellid Tuum
Rakumembraan Mitokondrid Ribosoomid Tsütoskelett (
mikrofilamendid ,
mikrotorukesed )
Mitoos +meioos
DNA struktuur
RNA ja valk
Metabolism
Rakuline organiseeritus
1-10 μm
Puuduvad või vähe
Puudub
Esineb (ei sisalda steroole, vaid hepanoide)
Puuduvad (oksüdeerumist katalüüsivad ensüümid seotud rakumembraaniga)
Esinevad (70S)
Puudub
Puuduvad
Rõngas, (
kromosoom ja plasmiidid) tsütoplasmas
Süntees samas kohas
Anaeroobne+
aeroobne Peamiselt üherakuline
5-100 μm
Tuum, mitokonder,
kloroplast Esineb
Esineb
Esinevad
Esinevad (80S)
Esineb
Esinevad
Lineaarne, erinevad
kromosoomid ,
histoonid , paiknevad tuumas
RNA tuumas,
valgud tsütosoolis
Aeroobne
Peamiselt
hulkraksed 5.
Arhede ja eubakterite peamised erinevused
Arhede
membraanilipiidides on eetersidemed, eubakteritel estersidemed ning
rakuseinas
peptidoglükaanid
ka.
Arhedel on
intronid , eubakteritel neid pole. rRNA ja ribosoomide
valgud erineva koostisega (eubakterite ribosoomid on tundlikud
klooramfenikooli suhtes). Valkude sünteesil esimene n-
terminaalne aminohape arhedel metioniin, eubakteritel formüülmetioniin. Arhede
viburid paneb liikuma ATP hüdrolüüs, eubakteritel [H+] erinevus
membraani külgedel.
6.
Esimesed prokarüootsed organismid tekkisid ~ aastat tagasi
3.5miljardit
7.
Esimesed eukarüootsed organismid tekkisid ~aastat tagasi
1-1.5miljardit
8.
Millised sümbiontsed bakterid on seotud õhulämmastiku
assimileerimisega?
Nt
tsüanobakterid, Rhizobium
9.
Millised tunnused näitavad et mitokondrid on tekkinud endosümbioosi
teel
Kahekordne
membraan, sisaldab mitokondriaalset DNA’d mis on koostiselt sarnane
bakteri DNA’le. Sisemise membraani koostis on sarnane
bakteriraku membraani koostisega. Paljunevad jagunemise teel, mitoos puudub. DNA
on rõngaskromosoomi kujul, histoonid puuduvad. Operonide esinemine
DNAs. Esinevad ribosoomid, mis ehituselt ja koostiselt sarnanevad
bakteri ribosoomidega.
10.
Millised tunnused näitavad et
kloroplastid on tekkinud endosümbioosi
teel
Sama
mis mitokondri puhul.
Membraanid 11.Raku
membraani paksus 6~10
nm
12.Millise
membraani komponendi struktuurvalem on esitatud (fosfoglütseriid,
sfingolipiid, etanoolamiin, koliin,
inositool )
13.
Nimetage vähemalt kolm tegurit mis mõjutavad membraanide
dünaamilisust (voolavust)
1.
lipiidide küllastatus - küllastamata
rasvhapped muudavad
paindlikumaks.
2.
kolesterool membraani koostises
3.
temperatuur
14.
Milliseid rasvu nimetatakse trans-
rasvadeks Trans-
rasvad on küllastumata
rasvad , mille rasvhapete jääkides esinevad
kaksiksidemed on trans asendis ehk E-
isomeerid .
15.
Mis on
lateraalne difusioon Ühe
membraani kihi piires toimuv aktiivne lipiidide molekulide
difusiooniline liikumine
16.
Mis on hübridoom ja kuidas ning milleks neid tekitatakse
Hübridoom
on
antikeha sünteesiva b-lümfotsüüdi ja müeloomiraku hübriid,
mille abil saab toota suurtes
kogustes antigeeni spetsiifilisi
monokloonseid antikehi.Säilinud mõlemad eellasraku tunnused.
1)
katseloom immuniseeritakse vajaliku antigeeniga
2)
immuniseeritud katselooma
viidud antigeenile vastavaid antikehasid
tootvad b-lümfotsüüdid liidetakse müeloomirakkudega
3)
selektiivsöötmel eraldatakse hübridiseerunud
rakud 4)
hübridoomi kasvatatakse masskultuuris või katseloomas
5)
antikeha eraldamine, puhastamine ja kasutamine
17.
Mida nim membraani sulamistemperatuuriks
Temperatuuri,
mille juures toimub
faaside üleminek(näiteks geeljas struktuur
->vedel strukt. või vastupidi)
18.
Millised on valkude membraanidesse kinnitumise viisid?
Transmembraanselt,
kovalentne side rasvhappe molekuli v isoprenoidse ühendiga,
kovalentne
side fosfatidüülinositooliga (GPI
ankur ), mittekov. side teiste
membraanivalkudega.
19. Nimetage membraanivalkude funktsioone.
Ainete
transport rakkude vahel, membraani struktuuri hoidmine, signaalide
vastuvõtt keskkonnast ja teistelt rakkudelt, rakkude
seostamine rakuvälise maatriksiga ja teiste rakkudega, tsütoskeleti valkude
seostumine membraaniga.
20.
Kirjutage lahustunud ühendi elektrokeemilise potentsiaali võrrand
ja milliseid suurusi võrrandi liikmed tähistavad. Millistes
ühikutes elektrokeemilist potentsiaali mõõdetakse.
Elektrokeemiline
potensiaal :
μ=μ0
+ 2.3 RT log a + zFE. Mõõdetakse J/mol
μ0
- aine standartne keemiline potensiaal
a
- aine kontsentratsioon (aktiivsus)
E
- aine elektriline potensiaal
R
-
gaasikonstant (8.3 J/mol * K)
F
-
Faraday konstant (96 kJ/V*mol)
z
- aine osakese laeng
T
- temp Kelvinites(!)
21.
Defineerige
membraanipotentsiaal , millistes ühikutes mõõdetakse.
pindade
laengud ? lahuste laengud?
Membraanipotentsiaal
- ioonide kontsentratsioonide erinevus raku sise- ja väliskeskkonna
vahel. Mõõdetakse voltides. Tsütoplasmapoolne külg on negatiivsem
väliskülje suhtes. Loomsete/
taimsete rakkude tsütoplasmas on palju
K+,
vähe Na+
ja Ca+2
ioone. Veres Na+
kontsentratsioon suht kõrge.
22.
Nimetage membraanipotentsiaali tekkimise põhjusi
1.
ainete elektrokeemiliste potensiaalide erinevus rakus ja rakuvälises
ruumis
2.
vastasnimeliselt laetud ioonide erinev
liikumiskiirus läbi membraani
- moodustub elektriline potensiaal e difusioonipotensiaal.
3.
jätkuvalt säiluva difusioonipotensiaali põhjuseks on nn “
pumpade ”
funktsioneerimine rakumembraanis . (Pumpadeks nim membraanide
transportsüsteeme, mis ainete
transpordiks kasutavad vahetult ATP
energiat - osalevad aktiivses
transpordis )
23.
Rakumembraani, kloroplasti tülakoidi, mitokondri sisemembraani
membraanipotentsiaali väärtused.
Rakumembraan
-70mV, kloroplastil -30mV, . Mitokondril -180 mV,
24.Defineerige
aktiivne ja passiivne transport elektrokeemilise potentsiaali
vahendusel ja nimetage passiivset ja aktiivset transporti teostavad
valgud.
Aktiivne
transport kasutab energiat kas otseselt näiteks ATP kujul või
kaudselt membraanipotentsiaali kujul, mis on esialgselt loodud ATP
energia arvelt.
Passiivne
transport ei kasuta mingit välist energiat.
Aktiivse
näide: ATP-
aasid Passiivse
näide:
Ioonkanalid 25.Kirjutage
Nernsti võrrand ja selgitage selle
tähtsus.E
= E0
- RT/nF ln a(oks)/a(red)
E
- elektroodi potentsiaal,
E0
- elektroodi standardpotentsiaal,
R
- universaalne gaasikonstant (8.314 J/(K mol)),
F
- Faraday arv (96485 C/mol),
T
– temperatuur kelvinites,
n
– määratava iooni laengu absoluutväärtus või reaktsioonis
osalevate elektronide arv
a
- potentsiaali määrava iooni aktiivsus.
Tasakaaluseisund kui E=0, Nernsti potentsiaal. Näitab kas energiat on vaja juurde või
mitte.
Nernsti
võrrand näitab, et konts. erinevus rakus sees ja rakus väljas on
tasakaalustatud elektriliste potentsiaalide erinevusega rakus sees ja
väljas. Ehk siis põhimõtteliselt võtab Nernsti võrrand arvesse
seda, et erinevatel ioonidel on erinev mõju (elektrilisele)
potentsiaalile.
26.
Kirjeldage akvaporiinide ehitust, millise aine transpordiks vajalikud
Koosneb
kuuest transmembraansest alfaheeliksist,
kusjuures nii amino kui
karboksü ots jäävad tsütosooli poole. Moodustavad kanali läbi
mille liigub kas ainult vesi või ka selles lahustunud ained,
sõltuvalt akvaporiinist.
Reguleerivad
veevoolu (aquaporins are "the plumbing system for cells").
27.
Nimetage 2 tegurit mis mõjutavad kanalivalkude avatust.
Elektriline
signaal (membraanipotentsiaal), keemiline signaal. (teatud ühendite
seostumine kanali valguga) Lisaks ka temperatuur,
mehaaniline jõud
ja fosforüleerimine.
28.
Mida mõistetakse sekundaaraktiivse transpordina. Nimetage
transportvalgu tüüp, tooge näiteid milliste ainete transpordiks
kasutatakse?
Sekundaaraktiivne
transport ei kasuta otseselt ATP energiat, et aineid transportida,
vaid elektrokeemiliste potentsiaalide erinevust. Antipordid ja
sümpordid. Nt Glükoosi transporditakse
niimoodi rakku koos Na
ioonidega.
29.
Iseloomustage K/Na-
ATPaas -i ja nimetage mõni protsess, mille läbiviimisel on
oluline.
Aktiivtransport ,
mille käigus liigutatakse ATP hüdrolüüsi energia arvelt 3 Na
iooni välja ja 2 K iooni raku sisse. Oluline membraanipotentsiaali
hoidmisel, koos Na ioonidega glükoosi sisseveol rakku
sekundaaraktiivtranspordil,
osmoosi kontroll.
30.
Millise membraanipotentsiaali juures
avanevad pingesõltuvad K
kanalid närvirakkudes?
+40mV
juures avanevad lisa K kanalid ja K liigub välja, tekitades kiirelt
negatiivse membraanipotentsiaali (
repolarisatsioon ).
Aga
jah.. konspektis on ta kirjutanud tõesti -50 aga loengus ta rõhutas,
just +40...((Positiivse.
umbes
-50mV pole? arvan ka, et -50mV))
31.
Loetlege närviimpulsside tekkimises ja edasiliikumises osalevad
transportvalgud .
Na
kanalid, K kanalid. Na/K
pump . Veel
midagi? Ligandiseoselised
kanalid
32.
Milline on närviimpulsi edasiliikumise kiirus?
100-120
m/s
33.
Milline on aktsioonipotentsiaali kestvus?
umbes 4ms
34.
Mida nimetatakse aktsioonipotentsiaaliks?
Aktsioonipotentsiaal on raku membraanipotentsiaali kiire järsk langus (membr pot on
negatiivne) ja tõus kindla reeglipära alusel.
35.
Kuidas tagatakse närvi-impulsi ühesuunaline liikumine?
Pärast
AP-d on rakk refraktoorses
perioodis , mil teda ei ole võimalik enam
erutada niimoodi. Seepärast ei saa
impulss liikuda tagasi vaid
ainult edasi selles sünapsis. Refraktoorses perioodis on Na kanalid
suletud ja ainult K saab liikuda rakust välja.
36.
Milleks on vajalik müeliinikiht aksonite ümber ja kuidas see
moodustub?
Müeliin
suurendab kiirust, millega impulss saab liikuda mööda närvikiudu.
Signaal n-ö “hüppab” ühest müeliinita kohast järgmisele.
Käitub natuke nagu
isolaator juhtmel . Seda toodavad Schwanni rakud
(perifeerses närvisüsteemis) ja oligodendrotsüüdid (tsentraalses
närvisüsteemis).
37.
Defineerige signaalmolekul (sünonüümid - neuromediaator,
neurotransmitter,
virgatsaine ).
Neurotransmitter
ehk neuromediaator ehk virgatsaine on
keemiline aine, mille abil neuron (närvirakk) edastab keemilise
sünapsi kaudu
närviimpulsi teisele (närvi)rakule38.
Kuidas toimub signaalmolekuli sekreteerimine sünapsisse ja kuidas
indutseeritakse sünapsijärgses neuronis närvi-impulsi teke.
AP
liigub mööda presünaptilise raku membraani kuni sünapsini. Seal
avanevad depolarisatsiooni tõttu Ca ioone rakku laskvad kanalid. Ca
kontsentratsioonile reageerivad mediaatorainega vesiikulid, mis
liituvad rakumembraaniga.
Mediaatoraine lastakse sünapsisse, osa
sellest hajub, osa satub vastasraku retseptoritele, mis reageerivad
mediaatorainele kas avades ioonkanalid, mille tagajärjel muutuks
membraanipotentsiaal või panevad raku
tootma signaali
kandjaid molekule, mis muudavad raku talitlust mingil viisil.
39.
Nimetage ajutegevust toetavaid signaalmolekule
Serotoniin ,
dopamiin , endorfiinid.
40.
Nimetage lihaste kokkutõmbeid reguleerivaid signaalmolekule
Atsetüülkoliin,
(nor)
epinefriin (südametegevus).
41.
Nimetage sclerosis
multiplex (hulgiskleroos) tekkimise põhjusi
Kahjustatud
müeliiniga piirkondades esineb makrofaagide infiltratsioon, mis
viitab põletikulisele geneesile. Lisaks on leitud ka laialdast
oligodendotsüütide
apoptoosi ning
mikrogliia aktivatsiooni.
Immuunsussüteemi ebanormaane B- rakuline vastus (oligklonaalsete
antikehade tootmine) 2/3 patsientidest. Aga tegelt see on kalane
teema ja keegi ei oska päris seletada kus ja mida ja miks.
42.
Nimetage ja iseloomustage transportvalkude häiretest sõltuvaid
haigusi
Mootorneuronite probleemid
Na+ kanalid
Tsüstiline
fibroos Cl- kanalid
Bipolaarsed häired
Na+/K+-ATPaas
Südametegevuse häired
Na+/K+-ATPaas
Resistentsus ravimite suhtes ja kemoteraapiale
ABC pump (multi-drug resistance)
Värvipimedus
[H+]
gradient kui pump (rodopsiin)
43.
Rakumembraan on läbitav K+ jaoks. [K+] raku sees on .....M ja
väljaspool rakku ......M. Kui suur ja millise märgiga on
tasakaaluline membraanipotentsiaal? R - gaasikonstant (8.3 J
mool -1
K-1);F - Faraday konstant (96 kJ V-1 mool-1); z - aine osakese laeng
(
valents ),T- temperatuur Kelvini
kraadides (toatemperatuur
20oC=293K).
Sain
vastuseks 25,33 mV * ln (Kväljas/Ksees)
44.
Kui membraanipotentsiaal on .......V ja [K+] väljaspool on
.........M, siis milline on [K+] rakkude sees tasakaaluseisundis ? R
- gaasikonstant (8.3 J mool-1 K
-1);F - Faraday konstant (96 kJ
V
-1 mool
-1); z - aine osakese laeng (valents),T-
temperatuur Kelvini kraadides (toatemperatuur 20oC).
45.
Millest on tingitud membraani depolariseerumine aktsioonipotentsiaali
tekkel?
Ioonkanalite
positiivse tagasiside süsteemist. Kui kuskilt sattub raku sisse Na
ioone, siis need indutseerivad
kanalite avamise ja veel rohkemate Na
ioonide rakku
voolamise . Võivad tulla retseptoritelt või sünapsist
jne.
46.
Millest on tingitud membraani repolariseerumine esialgsele
membraanipotentsiaali väärtusele (~ - 60 mV) närvi-impulsi tekkel?
Kui
membraanipotentsiaal on piisavalt kõrge, siis see sulgeb Na kanalid
ja samas avab K kanalid ja K voolab rakust välja, normaliseerides
potentsiaali.
47.
Kui rakumembraan on hüperpolariseerunud, siis milliste
protsesside/ainete liikumise vahendusel võiks toimuda membraani
depolariseerumine?
nt.
Na või Ca sissevooluga vastavatest kanalitest.
Endomembraanid
(ER/Golgi)48.
Nimetage vähemalt 5 rakkudest ER/Golgi vahendusel sekreteeritavat
valku
Proteaasid,
glükoproteiinid, proteoglükaanid, mutsiinid, fibromoduliin,
laktoferriin.
49.
Kirjeldage SRP
(signaali äratundja
partikkel ) struktuuri ja milleks vajalik.
Proteiin -RNA
kompleks . Eukarüootides kuus polüpeptiidi + RNA. Prokarüootides 1
polüpeptiid + RNA. Seostub valgu signaaljärjestusele ja tegeleb
selle liigutamisega membraanile.
50.
Kirjeldage signaaljärjestust valgu liikumiseks tsütosoolist ER-i,
milliste teiste valkudega signaaljärjestus komplekseerub? Kus
paikneb selle järjestuse
retseptor ?
ER
- Sekreteeritavad valgud sisaldavad N-terminaalses otsas nn
ER
signaal- e liiderjärjestuse. Signaaljärjestused on näiteks NLS ja
NES.(tõin
selle 48. juurest ära, sest ei vastanud eriti küsimusele, pigem
sellele, aga mitte eriti hästi...)
Signaalpeptiidi
mõlemis otsas on mõned pos laetud AH-d, keskel 8-10 hürdofoobset
AH-d. Hüdrofoobne piirkond on oluline just valgu seostumisel ER
pinnal paiknevate retseptoritega.
51.
Nimetage pöördtranskriptaasi (RNA sõltuv DNA polümeraas) osavõttu
vajavaid protsesse eukarüoodi rakus.
Retrotransposonite
liikumine
genoomis ühest punktist teise RNA vahendusel.
Telomeeride
pikenemist läbiviiv
telomeraas kannab oma RNA molekuli, mille alusel
pikendatakse telomeere.
52.
Nimetage ER-is ja Golgis sekreteeritavate valkudega toimuvad
modifikatsioonid
ER
- disufliidsideme tekke, valkude kokkukeerdumine ja oligomeeride
teke.
Golgi
- glükosüleerimine, osaline proteolüüs st eemaldatakse
teatud osa polüpeptiidahelast, et muuta näiteks seedeensüümid
aktiivseteks .
53.
Kirjeldage antikehade struktuuri. Milliste sidemete vahendusel
selline struktuur moodustub ja millises raku piirkonnas? Kuidas
antikehasid kasutatakse kindla valgu lokalisatsiooni määramiseks
rakus?
Antikehad
koosnevad neljast disulfiidsildadega ühendet valguahelast (2
suuremat identset H-
ahelat ning 2 väiksemat identset L-ahelat)(tekib
Y kuju).
Suured
globulaarsed
proteiinid . Glükoproteiinid. Funkts. üksus on
immunoglobuliini
monomeer . Neid võib olla ka rohkem kui 1. Moodustub
kahe identse raske ja kahe identse kerge ahelda omavahelisel
seostumisel disulfiidsidemetega. B
plasma rakud toodavad
karedapinnalisel ER-l? Toodetakse flourestseerumisvõimeline
antikeha, mis kinnitub otsitava valgu külge ja fluorestseerudes on
see valgusmikroskoobis kohe näha.
Kõigile antikehadele on omane sarnane
üldstruktuur – nad koosnevad neljast
disulfiidsildadega
ühendatud
valguahelast.
Kaht suuremat ja omavahel identset ahelat nimetatakse rasketeks ehk
H-ahelateks (
ingl.k
heavy chains),
kaht väiksemat ning samuti omavahel identset ahelat nimetatakse
kergeteks ehk L-ahelateks (
ingl.k
light chains).
54.
Kuidas pannakse kokku valgu Asn jäägile seotav oligosahhariidne
kompleks
UDP-GlcNAc
seostub ER- membraanis dolihoolfosfaadi külge. Seal liidetakse
sellele 5 mannoosijääki. Dolihool liigub membraani luumenipoolsele
küljele flipaasi toimel. Liituvad veel 4 Man ja 3 Glc jääki.
Oligosahhariid on valmis ja oligosahhariid-valk transferaas toimetab
selle Asn külge.
55.
Miks on sekreteeritavate valkude glükosüülimine vajalik.
Õige konformatsiooni omandamine, rakkude
vastastikune
identifikatsioon ja adhesioon, tagab organellidevahelise
liikumise, suurendab resistentsust proteeaside suhtes. Epiteelirakud
produtseerivad mutsiine, mis on olulised epiteelirakke katva lima
tekkes .
Kodeeriv osa pikeneb ja lisajuppe jääb
vähemaks? mutsiin vs mütsiin
56.
Mis määrab A, B, O vererühmad.
Neid
määravad A v B aglutinogeenide olemasolu v puudumine.
57.
Tähistage tabelis milliste
doonor ja aktseptorvererühmade vahel
võib vereülekandeid teha.
Vastavasse
kohta lisada +
58.
Mida tuleb ette võtta, et reesuskonflikti puhul (Rh
negatiivne
ema, Rh positiivne laps) vältida teise lapse sündimisel loote
kahjustamist reesusvalgu antikehadega?
Reesuskonflikti
ärahoidmiseks süstitakse Rh- negatiivsetele naistele
profülaktiliselt Rh-immunoglobuliine. Need ained vähendavad
antikehade kontsentratsiooni tõusu riski sünnituseelses perioodis
ja sünnituse ajal.
Raseduse
ajal saab väga tugeva reesuskonflikti tagajärjel tekkinud
aneemia raviks teha emakasisest vereülekannet.
Ultraheli kontrolli all
kantakse lootele nabaveeni kaudu verd. (Seda tehakse alates 22.-24.
Rasedusnädalast). Loote raske aneemia korral ollakse sunnitud
vereproove võtma ja vereülekandeid tegema vajadusel korduvalt.
Reesuskonflikti
saab ära hoida Rh-neg emale profülaktiliselt Rh-immunoglobuliine
süstides ja aneemia vältimiseks teha in
utero vereülekandeid.
59.
Millisesse raku piirkonda liikuvad valgud korjavad kokku vesiikuli
kesta valgud COPI, COP II ja klatriin?
COP
I - Golgi → ER
COP
II - ER → Golgi
Klatriin
- 1) transGolgi → endosoom
2) rakumembraan → endosoom
60.
Milline tähtsus on järjestustel
KDEL ja Man-6-P sekreteeritavatel
valkudel.
KDEL
on järjestus, mis hoiab valku ER-st sekreteerumast. Kui selles on
mutatsioon , valk sekreteerub. KDEL järjestus tagab ka nende
residentsete valkude transpordi tagasi Golgist ERi, millel on
õnnestunud KDEL retseptorit vältida.
Man-6-P
markeeritud valgud liiguvad Golgist lüsosoomi. Samuti vastutab see
järjestus selle eest, et lüsosoomi valgud jõuaksid oma määratud
organellidesse.
61.
Millised valgud ja milleks on vajalikud tagamaks vesiikuli membraani
ja õige märklaudmembraani ühildumist.
RAB
(seovad märklaua efektoriga, mis transpordib vesiikuli
märklaudmembraanini). SNARE (aktiviseeruvad pärast vesiikuli ja
märklaudmembraani seondumist; vesiikuli V-SNARE seostub
märklaudmembraani T-SNARE valguga). RAB ja V-SNARE
paljusus ning
vesiikulite spetsiifika.
RAB
valgud - vesiikuli membraanis, seostuvad märklaua RAB efektoriga,
mis võib transportida vesiikulit mikrotorukesi mööda
märklaudmembraanini. Pärast seondumist hakkavad tööle SNARE
valgud. Vesiikuli V-SNARE seostub märklaudmembraani T-SNARE valguga.
Spetsiifilisuse tagab esiteks RAB valkude paljusus (~70) ja V-SNARE
valkude paljusus (~35). Kuna igal vesiikulil on need
spetsiifilised ,
siis tagab see õigete membraanide sulandumise.
62.
Millistesse membraanidesse võib sekretoorne rada vesiikuleid
toimetada?
ER,
Golgi kompleksi erinevad osad, rakumembraan. Tegelt
lisandused on oodatud. lüsosoom,
vakuool taimedes
63.
Nimetage organelle mis on ümbritsetud poolega lipiidsest
kaksikkihist.
Kuidas ja kus sellised
kompleksid tekivad, mis on nende ülesanded?
Lipoproteiinsed
kompleksid,mis transpordivad kolesterooli ja
rasvhappeid (
VLDL , HDL,
IDL, LDL), sünteesitakse siledapinnalises
ERis . Oleosoomid taimedes.
64.
Kuidas tagatakse valkude õige konformatsiooni teke ER-is. Kuidas
märgistatakse ja parandatakse/kõrvaldatakse
vales konformatsioonis
valgud? Kalneksiini ja kalretikuliini osa.
Glükosüleerimisega.
Chaperone’
idega . Glükosüülitransferaas seob oligosahhariidse
jäägi otsa glükoosijäägi, mille alusel chaperone’id selle ära
tunnevad . Glükoosijääk eemaldub kui
konformatsioon on õige. Kui
mitte, tehakse kogu tsükkel uuesti või seostub valk
translokaatoriga, liigub tsütosooli ja seal
seostatakse ubikvitiiniga, mis on märgistuseks, et see valk tuleb tõmmata
lagundamiseks proteasoomi. Kalneksiin ja kalretikuliin on
chaperone’id, mis seostuvad mittetäielikult struktureerunud
valkudega ja hoiavad neid ERis
65.
Nimetage millised reaktsioonid (milliste ensüümide vahendusel)
peavad toimuma membraanide lipiidide sünteesil. Kus toimub selliste
lipiidide süntees?
1)Vajalike
molekulide süntees.
2)Rasvhapete
liitmine glütseroolile või sfingosiinile (atsüülitransferaasid)
3)Hüdrofiilse
pea liitmine
Siledapinnalises
ERis
66.
Järjestage VLDL, HDL, LDL partiklid vastavalt neutraalrasvade
kontsentratsioonile (
suuremalt kontsentratsioonilt väiksema suunas).
Millest on tingitud neutraalrasvade kontsentratsiooni muutus nendes
partiklites.
VLDL
50%, LDL 10%, HDL 8%
Kontsentratsioon
muutub, kuna veresoonte seinad sisaldavad
lipaasi , mis hüdrolüüsib
triatsüülglütserooli glütserooliks ja rasvhapeteks, rasvhapped
seonduvad kolesterooliga.
67.
Milline haigus on hüperkolesteroleemia ja millest on tavaliselt
tingitud haiguse avaldumine
noores eas.
Pärilik
haigus, mis tekib häirete tõttu kolesterooli transpordil
rakkudesse. Sümptomiteks kõrge kolesterooli tase veres ja
ateroskleroosist tingitud infarktid noores eas.
68.
Kuidas toimub LDL partiklite
sisenemine lipiide vajavatesse
rakkudesse?
Rakud,
mille pinnal on
retseptorid LDL osakeste jaoks
neelavad need
klatriiniga kaetud vesiikulitesse. Klatriini eraldumisel moodustuvad
endosoomid . Happelise keskkonna toimel retseptorid eralduvad ja
liiguvad vesiikulite membraani koostises tagasi rakumembraani, kus
toimub LDL sisu kasutamine membraanisünteesil.
69.
Mis on
androgeenne tundetus ja millest põhjustatud?
Testikulaarne
feminisatsioon, mis esineb XY genotüübiga inimestel, kelle fenotüüp
on XX. Põhjuseks mutatsioon X kromosoomi
AR-geenis,
mis
kodeerib testosterooni retseptori.
70.
Millised võimalused on lipiidide liikumiseks rakus ühest
membraanist teise, nimetage lipiide transportivaid valke.
Vesiikulitega,
monomeertransport (võib olla spontaanne või proteiini
assisteeritud) - fosfolipiidide üksikute molekulide liikumine neid
transportivate valkude abil. Flipaas liigutab membraani ühelt
küljelt teisele
71.
Milline transpordisüsteem peaks endosoomi membraanis olema, et pH
endosoomis muutuks
happeliseks ?
H+-ATPaas.
Esinevad ka Cl- kanalid, mis võimaldavad prootonite konts.
gradiendi teket ilma, et membraanipotentsiaal oluliselt muutuks
72.
Kuidas lüsosoomi madal pH tagab tsütosooli komponentide kaitse
hüdrolaaside eest lüsosoomi lõhkemisel?
Lüsosoomid
sisaldavad ~50 erinevat hüdrolaasi, mille ph optimum on 4.5-5. ph 7
juures on nad inaktiivsed. Seega kui lüsosoomidest hüdrolaasid
vabanevadki tsütosooli, on nad seal inaktiivsed.
73.
Nimetage ja kirjeldage endotsütoosi
variante Endotsütoos-
rakumembraani teatud osa sopistub sisse koos väliskeskkonna
materjaliga ja moodustab uue membraaniga ümbritsetud
organelli ,
endosoomi
- pinotsütoos- rakumembraan ebaspetsiifiliselt ümbritseb väliskeskkonnas paikneva vesilahuse osa
- retseptorseoseline selektiivne endotsütoos- rakumbembraanis paikneva retseptoriga seostub väliskeskkonna teatud aine (ligand), tekib ligand-retseptor kompleks, mis assimileeritakse, moodustub transportvesiikul
74.
Mis on transtsütoos?
Protsess,
mille käigus membraani ühel küljel moodustunud endotsütootilised
vesiikulid liiguvad raku vastasmembraanini ja sealt eksotsütoosi
abil välja. Ainete (makromolekulide) liikumine läbi raku.
Antikehade liikumine emalt lootele.
75.
Millised on erinevused rakumembraani ja tuumamembraani vahel?
Milline raku
organell on tuumaümbrisega ühendatud?
Rakumembraani
fosfolipiidkihid vahelduvad (1-2), tuumal on neid alati 2.
ER
Rakutsükkel76.
Defineerige rakutsükkel, nimetage ja iseloomustage peamised
rakutsükli etapid.
Rakutsükkel
on sündmuste ahel, mis algab ühe raku jagunemisest ja lõppeb
mitoosi ja jagunemisega kaheks tütarrakuks. RT’d saab ajaliselt
jagada 3. Esimene on
puhkeaeg (gap1), teine kolmandik ajast kulub DNA
sünteesiks ning viimane kolmandik on enamjaolt puhkeaeg (gap2), kuid
3. kolmandiku viimane kolmandik on mitoos.
Raku
eluperiood ühest jagunemisest teiseni.
Rakutsükkel
jagatakse
faasideks :
1)
S faas – DNA kahekordistumine, histoonide süntees
2)
G2 faas – vahemik S faasi ja mitoosi vahel
3)
M faas – mitoos
4)
G1 faas – vahemik M ja S faasi vahel
Rakutsükkel
on periood ühest rakujagunemisest teiseni. (G0). Interfaas - G1
kasvufaas, S DNA süntees, G2 mikrotuubulid, Mitoos - profaas kromosoomide kondenseerumine , metafaas kromosoomid joonduvad, anafaas kromatiidid lahknevad, telofaas uute rakutuumade teke, tsütokinees.77.
Mis tüüpi valgud on tsükliinid? Mis on nende funktsiooniks?
Millist tüüpi tsükliine võite nimetada?
Tsükliinid
- rakutsüklit käivitav masinavärk kujutab endast tsükliliselt
toimivat biokeemilist süsteemi. Seostuvad CDK-dega ja kontrollivad
nende aktiivsust. CDK – tsükliin-sõltuvad
kinaasid .
Tsükliine
on 2 põhilist klassi: mitootilised
tsükliinid , mis seostuvad CDK-dega
G2- faasis ja mis on vajalikud rakkudes M-faasi käivitamiseks, ning
G1-tsükliinid, mis seostuvad
CDK-dega G1-faasis ja on vajalikud S-faasi käivitamiseks.
78.
Nimetage tsükliinidest sõltuvate kinaaside omadusi, nimetage
vähemalt kolm viisi nende aktiivsuse regulatsiooniks.
aminohappeline järjestus säilinud
N-terminaalses otsas piirkond ATP sidumiseks
Katalüütilise tsentri AH-d laiali üle kogu ahela. Satuvad kõrvuti tertsiaarstruktuuri moodustumisel.
Tsükliiniga
seondumine, CDK aminohappeliste jääkide fosforüleerimine ja
komplekside valgulised inhibiitorid .
79.
Nimetage tsükliinidest sõltuvate kinaaside substraate rakutsüklis.
· Erinevad CDKd tagavad erinevate rakutsükli faaside
läbimise:
CDK-1 – üleminek G2 faasist mitoosifaasi
CDK-2 – üleminek S faasist G2 faasi
CDK-4 ja 6 – üleminek G1 faasist S faasi
80.
Mis on MPF (maturation promoting factor , sün mitose promoting
factor), kuidas tekib, millest koosneb, kuidas laguneb.
Heterodimeerne
proteiiin tsükliin B ja CDK1.
kuidas tekib?
Mitootilise tsükliini kontsentratsioon tõuseb
järk-järgult ning seostub CDK-ga, moodustab MPF kompleksi
Millest koosneb?
mitootilisest tsükliinist ja CDK-st
(tsükliin-sõltuvad kinaasid)
Kuidas laguneb?
MPF ise aga laguneb metafaasi- anafaasi jooksul. MPF laguneb seetõttu, et ta käivitab proteolüütilise
aktiivsuse, mis teda ennast lagundab . MPF-i lagunemine on vajalik, et
rakk saaks mitoosist väljuda.
81.
Milline on valk p53 osa rakutsükli kontrollis , kuidas see
realiseerub?
*tumor repressor e märgistab vigase DNA (võib tekitada kasvajaid),
tingides apoptoosi või rakutsükli peatamise ning vigase DNA
parandamise.
Aktiveerib
CDK inhibiitorvalgu p21
82.
APC (anaphase promoting complex ) roll metafaasist anafaasi üleminekul
(M pärssimine).
Märgistab
ubikvitiiniga proteolüüsiks valgud ning aktiveerib valgu Cut1p, mis
lagundab märgistatud valgulised sidemed õdekromatiidide vahel,
toimub kromosoomide eraldamine. Kui ei funktsioneeri, ei hakka
anafaas pihta.
83.
Nimetage 4n kromosoomistikuga rakutsükli faasid .
S-faasi
lõpust kuni mitoosi anafaasini (G2 faas, mitoosi profaas, metafaas,
anafaas)
84.
Nimetage vähemalt kolm protsessi raku/organismi elutegevuses, mis
vajavad apoptoosi toimumist .
*Elimineerib
ebavajalikke rakke, kontrollib mitoosi kiirust, hoiab ära tõsisemate
mutatsioonide teket organismis, kontrollib rakkude arvu homöostaasis?
nt
loote areng, luude kasv, valesti arenenud rakkude kõrvaldamine
kasvajate vältimiseks?
85.
Iseloomustage apoptoosi etappe
Apoptoosi
alguses toimub raku kokkutõmbumine, kromatiin pakitakse kokku ja
viiakse tuumamembraani äärealadele (püknoos), tsütoplasma
kondenseerub. Seejärel tuum mureneb (karüoreksis) ja rakk
fragmenteerub surnud rakuks ja apoptootilisteks osakesteks. Lõpuks
fagotsüüt teeb pacmani.
Alternatiivne variant sama seletamiseks :): 1. aktiveeritakse kaspaasid 2.
kromatiin kondenseerub (pakitakse kokku), aktiveeritakse nukleaase
(kromatiin lagundatakse fragmentideks), tuum fragmenteerub 3.
tsütoplasma kondenseerub, raku membraani muutused markeerivad
apoptootilise raku fagotsüütidele (rakk laguneb vesiikuliteks)
86.
Iseloomustage kaspaase ja kaspaaside kaskaadi
kaspaas
- (spetsiifilised proteaasid) apoptoosile iseloomulikud valke
lagundavad ensüümid. Aktiivtsentris asub tsüsteiin. Peptiidsidemed
lagundatakse Asp jäägi kohalt. Kaspaase on palju - esmalt aktiveeritakse prokaspaas, mis oma korda aktiveerib järgmise
kaspaasi -> aktiveerub kaspaaside kaskaad .
87.
Kirjeldage mitokondritega seotud apoptoosi (Apaf valgud, tsütokroom
c, Bcl valgud)
Mitokondrid
osalevad rakusisesel apoptoosi rajal, mille käivitavad Bcl valgud.
Kui rakk läheb apoptoosi, muutub mitokondri membraan õhemaks ja
välja pääseb valk CytC ning seostub Apaf 1-ga, mis aktiveerib
profaas-9, sealt eraldub jupp ning sama rada jätkub, kuni saabub
raku surm.
88.
Nimetage kasvufaktoreid (GF) (välisfaktoreid) mis olulised
apoptoosi pärssimises
NGF
– närvikasvu faktor, IGF-id, (insulin like GF), EGF (epidermal
GF), adrenomedulliin
89.
Nimetage apoptootiliste rakkude avastamise meetodeid
Hoechsti
värvumine, trüpaansininsega värvimine, DNA fragmentatsioon, mitokondriaalse transmembraanipotentsiaali vähenemise uurimine ,
90.
Mõõdeti suure hulga rakkude DNA sisaldus. See
kõikus vahemikus............ pikogrammi tuuma kohta. Ühe
konkreetse raku tuumas oli DNA sisaldus ...........pikogrammi.
Millises rakutsükli faasis see rakk on?
91.
Kirjutage, milline on kromosoomistik (n, 2n, 3n või 4n) rakutsükli
erinevates faasides .
G1
- 2n
S
- 2n >> 4n
G2
- 4n
M
- 4n >> 2x2n
92.
Rakk sisaldab mitoosi alguses .........kromatiidi. Mitoosi lõppedes
moodustub 2 rakku, milles kromosoomide arv on.............
4v8,
2 Mitoosi alguses on kromatiide 2 korda
nii palju kui kromosoome. Mitoosi lõppedes on kromosoome sama palju
kui alguses (diploidsed rakud)
93.
Raku keskosas algab fragmoplasti moodustumine ja raku vastasotstes
on moodustunud tuumad . Millise rakuga on tegemist?
1. loomarakk metafaasis
2. loomarakk telofaasis
3. loomarakk tsütokineesis
4. taimerakk metafaasis
5. taimerakk tsütokineesis
http://en.wikipedia.org/wiki/Phragmoplast
Mitokondrid
94.
Mitokondrite suurus
~0.5
- 1μm
95.
Iseloomustage mitokondrite DNA-d (% raku DNAst,
molekuli suurus, struktuur, geneetiline kood, geenide paiknemine ,
millised valgud kodeeritud)
16.5kbp
e 15k-17k aluspaari .
Koosneb kergest ja raskest ahelast .
Kodeerib kokku 37 geeni, millest 13 on valkudele , peamiselt
elektrontrasportahela valgud. Kodeeritavad
valgud: mtETA, ATP sünteesi valgud, translatsioonivalgud.
16,5
kbp e 15k-17k aluspaari. 100-10.000 mitokondrit, igas oma DNA, kerge
(C-rikas, 9geeni) ja raske ahel (G-rikas, 28 geeni). kodeerib 37
geeni, 13 valku, 22 tRNA'd, 2 rRNA. Elektrontransportahela valgud:
NADH dehüdrogenaasid, koensüüm q - tsütokroom c
reduktaas/tsütokroom b, tsütokroom c oksüdaas, ATP süntaas.
96.
Kuidas mitokondrid liiguvad eellasrakkudelt tütarrakkudele?
Mitokondrid
poolduvad rakkudega sarnaselt, mõlemale tütarrakule umbes sama
palju mitokondreid
97.
Mis on tsütoplasmaatiline pärilikkus?
Sugulisel
paljunemisel pärinevad raku mitokondrid emasliinist, kuna
viljastatud munarakul on vaid munarakult saadud tsütoplasma ja seega
ainult munarakult saadud mitokondrid. Tsütoplasmaatilise pärilikkuse
tõttu levivad mitmed mitokondriaalsete mutatsioonidega seotud
haigused- haigus pärandub, kui emal on mutatsioonid mitokondrites,
edasi pärandub vaid naisliini pidi, kuid avaldub ka meestel.
98.
Nimetage vähemalt viis madalmolekulaarseid ühendeid transportivaid
valke mitokondrite membraanis ja mille jaoks nad on vajalikud.
alfa-ketoglutaraat
- tagab malaadi liikumise mitokondri maatriksisse
CoA
- rasvhapete transport mitokondritesse
püruvaat/OH
-antiport - püruvaadi importsüsteem
karnitiin - lipiidide transport
tsütokroom
C - elektronide transport
poriin
- see moodustab kanaleid, mis lasevad läbi valke molekulmassiga kuni
10 kD
99.
Mis on vahetu sama energiaallikas ATP sünteesil nii mitokondrites
kui kloroplastides elektronide transpordi ahelas.
NADH
100.
Defineerige pmf ja kirjutage valem. Millises mitok piirkonnas on
prootoneid rohkem, millises vähem, kust kuhu prootonid liiguvad ATP
sünteesil.
Proton
motive force. Elektrokeemilise potentsiaali erinevus membraani
külgedel. Mitokondris pumbatakse prootonid membraanide vahelisse
alasse. ATP sünteesil liiguvad nad läbi ATPsüntaasi tagasi
tsütosooli.
Prootonite
liikumapanev jõud on avaldatav valemiga
pmf
= Y
- [(RT/F) * ΔpH] = Y
- 59 ΔpH.
101.
Kuidas mitokondrites pmf tekib? Kui palju prootoneid transporditakse
2 elektroni liikumisel hapnikule?
Elektrontranspordiga
pumbatakse prootonid kahe membraani vahele. 4 prootonit.
102.
Milline valguline kompleks mitokondrites kasutab prootonite
kontsentratsiooni erinevuses peituvat energiat ATP sünteesiks? Kus
see kompleks mitokondris paikneb? Millises mitokondri piirkonnas ATP
sünteesitakse? Kuidas ATP transporditakse mitokondrist tsütosooli?
ATP-süntaas. Koosneb kahest valgulisest
subühikute kompleksist - F0 paikneb sisemembraanis ja F1 membraani
maatriksipoolses osas. ADP/ATP-transportija abil - adeniinnukleotiid
kandja.
103.
Mitu prootonit kannab suktsinaat -CoQ reduktaas membraani ühelt
küljelt teisele?
Prootonite
transport läbi sisemembraani elektronide liikumise suktsinaat-CoQ
reduktaasi toimel ei toimu. Katalüüsib kahe elektroni liikumist
suktsinaadilt FAD-le ja lõpuks CoQ-le
104.
Milline erinevus on substraatsel ja pmf põhisel ATP sünteesil?
Substraatsel
fosforüleerimisel tekib 2 ATPd, pmf põhisel ATP sünteesil 28 ATPd
Substraatne
ei kasuta prootongradiendi energiat.
105.
Mis põhjustab elektronide liikumise mitokondriaalses ETA-s, nimetage
peamised elektronide transpordis osalevad valgulised kompleksid.
mtETAs
toimub elektronide liikumine negatiivse redokspotentsiaaliga
redokspaaridelt positiivsema redokspotentsiaaliga paaridele.
Peamised
elektronide transpordis osalevad valgud: NADCoQ-reduktaasne kompleks,
suktsinaat-CoQ reduktaasne kompleks, CoQH2-cyt c reduktaasne kompleks
ja tsütokroomi c oksüdaasne kompleks.
106.
Mis toimub elektronide transpordi ja oksüdatiivse fosforüleerumise
lahutamisel? Tooge näiteid.
Kaob
prootonite gradient, sisemembraan muutub prootonite suhtes
läbitavaks. ATP-d ei sünteesita. elektronide ülekandel vabanev
energia eraldub soojusena
nt.
ainevahetushaiguste korral, türeotoksikoos- kilpnäärme ületalitlus
107.
Kuidas toimub tsütosoolis sünteesitud valkude liikumine
mitokondrisse, milline on mitokondrisse liikuva valgu
signaaljärjestus?
Transport
toimub translatsiooni järgselt. Mitokondritesse viidavate valkude
signaaljärjestused tunnevad ära mitokondri välismembraanis asuvad
retseptorid ning signaaljärjestusteks võivad olla N-terminaalses
otsas paiknevad 3-5 Arg või Lys jääki(mitte järjest) või Ser ja
Thr, mitte kunagi Glu ja Asp. Signaalpiirkondi nim.
transiitjärjestusteks. Erinevatesse mitokondri piirkondadesse
liikuvad valgud omavad lisaks transiitjärjestustele ka
lisasignaaljärjestusi, mis sihtpunkti jõudes proteaaside toimel
lagundatakse, kuna need ei kuulu valmis valgu koosseisu.
Etapid:
Signaaljärjestusega valk seostub chaperonidega, mis kasutavad ATP
energiat, et hoida valk lahtikeerdununa, ainult sellise struktuuriga
valgud saavad mitokondrisse siseneda. Sisenev valk liigub läbi
välismembraani ja sisemembraani spetsiaalsete kanali kaudu ning see
saab toimuda vaid sellises kohas, kus kaks membraani kokku puutuvad.
Transpordiks on vajalik ka pmf. Ka maatriksis säilitatakse valgu
lahtikeerdunud strukt. ning N-terminaalsest otsast eemaldatakse signaaljärjestused. Pärast seda valgud konformeeruvad.
108.
Eeldades, et ühe ATP tekkimiseks peab läbi ATP süntaasi minema
mitokondrites 4 prootonit, mitu ATPd on võimalik saada steaarhappe
(18 süsinikku molekulis) oksüdeerumisel.
Põhjendada
arvutuskäik. 144?
Oksüdeerub
9 AcCoA, 8 NADH ja 8 FADH2 molekuli. AcCoA omakorda 3NADH ja 1 FADH
molekuli peale. Kokku on siis 35NADH ja 17FADH2. NADH oksüdeerumisel
liigub 10 prootonit, FADH2 puhul 6. Kokku 350 + 102 = 452 prootonit.
452/4=113 ATP-d
Vaadake
igaks juhuks seda, ma olen natuke primitiivne olnud siin.
http://www.elmhurst.edu/~chm/vchembook/622overview.html ← siin arvestatakse ~3 prootonit ATP peale. Lahendaks
siis mitteprimitiivselt ka?
Jagades süsinike arvu kahega, saab AcCoA arvu, lahutad sealt ühe ja saad
NADH ja FADH2 arvu. Igast AcCoA’st tekib omakorda 3 NADH’d ja 1
FADH2. Iga NADH arvelt saad üle kanda 10 prootonit, iga FADH2 arvelt
6 prootonit. Prootonite arvu jagad ühe ATP sünteesimiseks vajaliku
prootonite arvuga ja vastus käes. Sain ka 113.
109.
Mitokondriaalses elektronide transpordi ahelas ( mETA ) liigub NADH
oksüdeerumisel vabanenud 2 elektroni. Kui palju prootoneid liigub
läbi mitokondri sisemembraani 2 elektroni liikumisel? 4
prootonit Põhjendage.NADHCoQ
seob mitokondri maatriksist 2 prootonit ja kaks elektroni, tekib
CoQH2, mis annab erinevate komplekside vahendusel 2 prootonit ja kaks
elektroni membraanide vahelisse alasse, ühest elektronist saab nö
ringlev elektron mis tekitab CoQ- kompleksi. Selle kompleksiga seondub veel üks ringlev elektron ( vabanenud teiselt CoQh2lt, sest
protsess toimub ju pidevalt), saab tekkinud kompleks veel kaks
prootonit membraanidevahelisse alasse toimetada. Üks elektron muutub
jälle ringlevaks elektroniks ja teine elektron seotakse Cyt c-le,
kust edasi oksüdeerub nende elektronide arvelt hapnik.
Põhimõtteliselt jääb membraanidevahelisse
alasse 2 elektroni, kuna iga transporditava elektronpaari kohta on
üks ringlev elektron. Milliste mitokondri piirkondade vahel
ja kust kuhu prootonid liiguvad? Liiguvad
maatriksist kahe membraanivahelisse alasse. Miks tehakse
prootonite liikumise arvutused kahe, mitte ühe elektroni liikumise
kohta? Sest Q-tsükkel on üks tervik ning vaid
ühe elektroniga arvutamisel ei oleks sisuline taust õige???
Ma pakun, et kuna poolikute hapnikumolekulidega
ei ole esteetiliselt kaunis tegutseda, siis võetakse 2 elektroni
tasakaalustamaks reaktsiooni. Sest elektronid esinevad vaheldumisi oksüdeerunud ja redutseerunud vormis - redokspaarina.
110.
Loomaraku hingamisel neelduvat hapnikku kasutatakse otseselt (Õige
variant alla kriipsutada).
a) Glükolüüsis
b) elektronide aktsepteerimiseks mitokondriaalse ETA
lõpus
c) tsitraaditsüklis, NADPH genereerimise reaktsioonides
d) tsitraaditsüklis, püruvaadi oksüdeerimisel
atsetüülCoA-ks.
111.
Tsitraaditsükkel produtseerib kõrge energiasisaldusega ühendeid,
mis transpordivad energia mitokondriaalsesse elektronide transpordi
ahelasse (mETA), sest neid oksüdeeritakse mETAs. Need ühendid on:
(õige variant alla kriipsutada)
a. ATP ja CO2
b. CO2 ja FADH2
c. FADH2 ja NADH
d. NADH ja ATP
e. FADH2, NADH ja ATP
Kloroplastid
112.
Kloroplastide suurus. 2 - 10 micrometers in diameter
113.
Kloroplastide peamised kompartmendid ja membraanid.
Membraanid
- Välismembraan, sisemembraan ja tülakoidide ( lamellide )membraanid.
Kompartmendid - Tülakoidid (väikesed lamedad vesiikulid), graanid (tülakoidide virnad), graanitülakoid (üksikud tülakoid graanis),
stroomatülakoid (ühendab graanid üksikute hõredamalt paiknevate
tülakoididega), strooma (kloroplastide sees väljaspool tülakoide
paiknev piirkond), luumen (tülakoidide siseruum ).
114.
Fotosünteesi valgusstaadiumi reaktsioonid ja produktid , millises
kloroplasti osas toimuvad.
Lähteained:
süsihappegaas ja vesi
Produktid:
glükoos ja hapnik
Reaktsioonid
toimuvad kloroplastide sisemembraanidel (tülakoidmembraanidel).
Saab
alguse klorofülli ergastamisest valgusenergia abil. Edasised
reaktsioonid toimuvad ergastatud klorofülli energia arvel. Peamised
protsessid: klorofülli ergastamine valguse toimel; fotooksüdatsioon
ehk vee fotolüüs; elektronide energia salvestamine
eletrontranspordiahelas ATP energiaks; prootonite liitmine
vaheühendile (2NADP + 2H+ -> NADPH2)
Lähteained
vesi ja 2 NADP, produktid hapnik ja 2 NADPH'd, 3ATP'd.
valgusstaadiumi lõpuks glükoosi veel valmis pole.
115.
Vee fotooksüdatsiooni valem, millises kloroplasti piirkonnas toimub,
tekkinud produktide kasutamine.
2H2O
+ 4hV + = 4H+ + O2
+ 4elektroni, seega neli valguskvanti on vaja, et eemaldada 4
elektroni kahest vee molekulit, vabaneb hapnik. Toimub tülakoidide
membraanides. Tekkinud prootonid liiguvad luumenisse, pmf. Elektronid
liiguvad kinoonile ja sealt NADP-le, tekib NADPH.
Tekkivad hapnikuaatomid liidetakse molekuliks ning suunatakse atmosfääri.
116.
Fotosüsteemi ehitus, mis toimub fotosüsteemis, kus paikneb. Milline
protsess toimub fotosüsteemi tsentris ?
Fotosüsteemiks
nimetatakse pigment -valk kompleksi, paiknevad tülakoidide
membraanides. Iga fotosüsteem koosneb antennist(valgust neelavad pigmendid ) ja reaktsioonitsentrist(valkude kompleks+2 klorofüll a
molekuli+seos elektroni doonor-akseptoriga). Antenn sisaldab ühe või
mitu valgust koguvat kompleksi, mille pigmendi molekulid
absorbeerivad valguskvante. Valgusenergia liigub antennist tsentris
paiknevatele klorofüll a molekulidele, mis muudavad valgusenergia
keemiliseks energiaks. Kui klorofüll neelab valguskvandi , elektronid
ergastuvad ja muutuvad nõrgemalt seotuks. Reaktsioonitsentris toimub
ka laengute lahutamine e. elektroni eraldamine klorofülli molekulilt
ning selle kandumine kinoonile(elektroni akseptorile). Klorofüllile
jääb seega + laeng ning on tugev oksüdeerija, võtab elektroni
luuumenipoolsel küljel paiknevat elektronidoonorilt. (H2O)
117.
Kuidas tagatakse elektronide liikumine fotosünteetilises ETA-s,
millised valgulised kompleksid osalevad.
Elektronide
liikumiseks fETAs on vajalik PSI ja PSII koostöö, kuna PSII
eraldunud elektronid ei saavut piisavat energiat NADP-le liitumiseks
ning see saab toimuda ainult PSI-st saadud täiendava energia arvelt.
Valgud: Cytb/f-kompleks, plastostüaniin, ferrodoksiin, OEC,
feredoksiin NADP reduktaas jne.
118.
Kuidas ja kus valgusenergia muutub keemiliseks energiaks fotosünteesi
protsessis?
Valgusenergia neeldumine ja selle muutmine keemiliseks energiaks toimub
pigment-valk kompleksides, mida nimetatakse fotosüsteemideks ja mis
paiknevad tülakoidide membraanides. Valgusenergia (mitte
elektronid!) liigub antennist reaktsioonitsentrumis paiknevatele
klorofüll a molekulidele, millede vahendusel toimub valgusenergia konverteerimine keemiliseks energiaks
119.
Mitu H+ transporditakse 2H2O oksüdeerumisel vabanenud
elektronide liikumisel fs ETA-s, kui palju ATP-d on võimalik selle
arvel sünteesida?
8
prootonit. 2-3 ATP-d.
Kui
H2O oksüdeerumisel tekib kaks elektroni ja vabaneb 8 elektroni, kas
siis kahe H20 (H2Od on ju kaks molekuli) redutseerumisel ei
transpordita 16 prootonit?
Stroomast
luumenisse kokku 12 H+ -i, ATPaas sünteesib ATP u 3-5 H+ ga.
Sõltuvalt subühikute arvust promootoris. Seega 12 H+ -st umbes 3
ATP-d + O2
120.Pmf
suurus kloroplastides.~8 stroomas ja ~5
luumenis –> ΔpH = 3
-0,03-0,06*3=-0,21V=-210mV
121.
Fotosünteesi biokeemiline staadium, millised faasid eristatakse.
Millises faasis on vajalik ATP ja NADPH, milleks?
?
Biokeemiline staadium: CO2
ainevahetuses.
Faasid:
Süsihappegaasi seostumine ja redutseerimine sahhariidideks –
toimub ainult valguses (valgusstaadium) ja Calvini tsükkel( Calvin -
Benson – Bassham tsükkel)
(fotosünteesi
pimereaktsioonid).
Milleks
ATP ja NADPH: valguses tekib ATP ja
NADPH mis on vajalikud Calvini tsükli toimumiseks.?
122.
Millises kloroplastide osas CO2 sidumise ja redutseerumise
reaktsioonid toimuvad?
Stroomas
123.
Millist reaktsiooni katalüüsib ensüüm Rubisko .
Sahhariidide
sünteesil
124.
Kloroplasti DNA ehitus, suurus, milliseid valke kodeerib.
Rõngas
DNA molekulid, sarnaneb tsüanobakterite DNAle. 120k-160k bp. ~120
geeni, millest 20 kodeerivad fETA valke ja kodeeritakse ka rubisko
suur subühik.
125.
Fotosünteesi protsessis ..............................oksüdeeritakse
ja ..........................redutseeritakse. Kirjutada sisse õige
variant.
a) Sahharoos -hapnik
b) Süsihappegaas-vesi
c) Vesi-süsihappegaas Fotooksüdatsioon ja
süsihappegaas redutseeritakse suhkruks
d) Vesi-hapnik
e) Sahharoos-süsihappegaas
126.
Nimetage kloroplastis funktsioneerivaid subühikutest koosnevaid valke, mille mõned subühikuid on kodeeritud tuuma genoomis, mõned
kloroplasti genoomis.
rubisco
Mikrofilamendid
127.
Aktiini omadused. Aktiinimolekuli polaarsus ja seotus ATP/ADP-ga, +
ja – ots.
Aktiin moodustab 1-5% koguvalgust; kodeeritud geenide perekonna poolt;
selgroogsetes aktiinide geenide produktid erinevad ainult 4-5
aminohappe poolest, kuid erinevate aktiinimolekulide funktsioonid on
erinevad; geenid on konserveerunud; molekulis ~375 aminohappe jääki;
Iga aktiini molekul sisaldab Mg iooni ja ATP või ADP
esineb
kahel kujul: globulaarne monomeer (molekul on polaarne - molekuli jaotab kaheks osaks vagumus , millesse seostuvad ATP ja Mg); niitjas polümeer G- aktiinist (Polümeer on samuti polaarne. (-) otsana
tähistatakse ATPd siduvuva vagumusega otsa, vastaspiirkonda
tähistatakse (+) otsana. Subühikute liitumine toimub eelkõige (+)
otsa)
128. Kriitilise kontsentratsiooni mõiste ja suurus.
Polümeriseerumine
käigus väheneb keskkonnas olevate monomeeride hulk kuni konstantse
väärtuseni mida nim kriitiliseks kontsentratsiooniks (Kk). Selle
kontsentratsiooni juures polümeriseerumisel liituvate subühikute
arv võrdub dissotsieerumisel vabanevate subühikute arvuga. Kk on in vivo tingimustes ~ 0.1µM
129.
Nimetage tegureid, millest sõltub aktiini polümeriseerumine.
(Ioonide
kontsentratsioonist lahuses), vabade subühikute kontsentratsioonist,
polümeriseerumise kiiruskonstandist, aktiini polümeriseerumist
reguleerivad valgud.
130.
Nimetage peamised valkude klassid mis osalevad aktiinifilamentidest
moodustuvate struktuuride tekkes. Nimetage aktiini filamentidest
moodustuvaid struktuure.
Aktiiniseoselised
valgud e ARP, tekib võrgustik
nukleatsiooniseoselised
valgud, tekivad mikrohatud , filopoodid , lamellipoodid või
fagotsüteerijatel tassisarnased moodustised
Struktuurid :
1) kimbud -filamendid paralleelselt ( fibriin , villiin, müosiin)
2) võrgustik-filamendid ristuvad (a-aktiniin, filamiin,
spektriin)
131.
Aktiini mootorvalkude – müosiinide - struktuur
Keemilist
energiat motoorseks muutev valk e mehhanokeemiline ensüüm. Koosneb
rasketest ja kergetest ahelatest.
- Raskeid ahelaid 1-2, sõltuvalt tüübist. Koosneb kolmest domeenist:
Pea, N-terminaalne, seostumiskohaks aktiinile nim ka P- ling . Seostumiskohaks ATPle, omab ATPaasi aktiivsust, toodab mehhaanilist jõudu
Kael , ümbritsetud kergete ahelatega ja võib olla seotud regulaatorvalkudega
Saba, C-terminaalne, sisaldab seostumiskohti mis annavad spetsiifilisuse
132.
Peamised etapid müosiin-aktiin interaktsioonis.
Ensümaatiliselt
aktiivne müosiini pea teostab ATP hüdrolüüsi.
Konformatsioonilised muutused müosiinimolekulis, mis libistavad
aktiini müosiini suhtes edasi poole(alati - ots ees aktiinil ja
müosiin liigub seega + otsa poole).
Etapid:
- nukleotiidi seostumine, müosiin ja aktiin seotud, ATP seostub ATP- seostumiskohta , aktiini vagumus avaneb ja pea dissotseerub .
- hüdrolüüs, pea pöördub, kaela konformatsioon muutub, müosiini pea seostub uuesti aktiinile, kuid ühe subühiku võtta + otsa poole.
- Pi vabanemine , pea pöördub ja liigutab filamenti edasi, - ots ees
- ADP vabanemine, esialgse konformatsiooni e kangestusseisundi taastamine
133.
Lihasrakkude ehitus ja kontraktiilsuse printsiip
Vöötlihased:
koosnevad
lihasrakkude kimpudest , lihasrakk koosneb müofibrillidest e.
aktiinikimpudest, ja jaotuvad tumedateks ja heledateks ribadeks piki
lihasrakku. filamentide + otsad kinnituvad valgulistele Z ketastele.
silindrilist osa kahe Z joone vahel nimetatakse sarkomeeriks =>
müofibrill koosneb sarkomeeride ahelast. Iga sarkomeer koosneb kahte
tüüpi filamentidest. Paksud filamendid koosnevad ainult müosiinist
ja peened filamendid aktiinist. Sarkomeeride tumeda osa moodutavad
paksud müosiinimolekulid ja aktiin ja heleda osa ainult aktiin.
Aktiini filamentide + ots on seotud valkudega CapZ ja a-aktiniin, -
ots tropomoduliiniga, et takistada filamentide lagunemist.
Silelihased :
Koosnevad
värtnakujulistest rakkudest, mis sisaladavad ühte tuuma. Peened ja
paksud filamendid. Puuduvad sarkomeerid ja müofibrillid.Peened ja
paksud filamendid on seotud tihedate piirkondadega, analoogiliselt Z
ketastele.Teine ots on kaetud plaatidega, mis asuvad membraanis =>
membraanide seostumine fibrillidega.
Lihaste
kokkutõmbumisel peened ja paksud filamendid libisevad üksteise
suhtes. Muutub sarkomeeri pikkus. ATP ja Ca2+ juuresolekul
müosiinipead pöörduvad ja lükkavad aktiinifilamendid sakromeeri keskossa . Kokkutõmbumine toimub tsoonis, kus aktiin ja müosiin
kattuvad.
134.
Millistes protsessides lisaks lihasrakkude kontraktsioonile veel
osaleb aktiin/müosiin kompleks.
- Tsütoplasma voolamine
- Tsütokineesis osalemine- kahe tütarraku moodustumine
- raku komponentide liigutamine
- vesiikulite liikumine
- rakkude edasiliikumine tahkel pinnal
135.
Kuidas närviimpulsi jõudmine mootorneuronini põhjustab
sarkomeeride kokkutõmbe. Ca2+ osa.
Ca
konts. kasv tsütosoolis põhjustab lihaste kokkutõmbeid. Ca konts.
on puhkeseisundis madal ja see tagatkse Ca-ATPaasi abil, mis Ca
pidevalt rakust välja pumpab. Kui närviimpulss jõuab lihasrakuni,
muutub EP ja toimub depolarisatsioon . Signaal toimetatakse mööda
spetsiifilisi signaaliülekande ahelaid sakroplastilise retiikulumi
Ca pumpadeni, mis avanevad ja Ca kontsentr. suureneb. Selle mõjul
aktiiniga seondunud valgud tropomüosiin ja troponiin muudavad oma
asetust aktiini filamentidel, vabaneb aktiini müosiiniga seostumise
piirkond ja müosiin saab mööda aktiinifilamenti liikuda.
136.
Mootorvalk liigub tsütoskeleti + otsa suunas. kinesiin
137.
Mootorvalk liigub tsütoskeleti valgu – otsa suunas. düneiin
138.
10 ATP hüdrolüüs võimaldab müosiinil II liikuda ..50-100
nm
. kaugusele
Mikrotorukesed
139.
Aktiini ja tubuliini võrdlus (mille poolest erinevad ja mille
poolest sarnanevad)
Aktiin
seob ATP’d, müosiinirada, reguleeritud kokkupanek paljudes
kohtades. Tubuliin seob GTP’d, kinesiinide ja düneiinide rada,
reguleeritud kokupanek vähestes kohtades.. Mõlemad dünaamilised, polariseeritud , üsna jäigad.
140.
Mis on treadmilling ja millistes tingimustes toimub
Treadmilling
on ühe filamendi otsa kasv, samas kui teine ots lüheneb. See
tekitab efekti, justkui filament liiguks. Toimub tingimustes, kus
ühest otsast proteiinisubühikuid pidevalt eemaldatakse samas kui
teise otsa neid lisatakse.
141.
Nimetage vähemalt 4 tegurit, mis mõjutavad mikrotorukeste
polümeriseerumist ja depolümeriseerumist.
Dimeeride
kriitiline konsentratsioon, GTP tubuliini subühikute seostumise
kiirus, taimedest isoleeritud alkaloidide lisandumine, MAPi
fosforüleerimine.
tsentrosoomide
olemasolu? Kriitiline kontsentratsioon e alfa ja beeta dimeeride
konts mis on tasakaalus mikrotorukestega. Gtp tubuliini subühikute
seostumise kiirus.taimedest isoleeritud alkaloidid mis seostuvad
dimeeriga pöördumatult ja takistab polüm või depolüm. MAP kui
fosforüülida siis mikrotorukesed lagunevad. Ainult sellised
mikrotorukesed mille + ots on seotud GTP tubuliiniga on stabiilsed ja
võivad siduda tubuliini subühikuid.
142.
Millistes protsessides osalevad mikrotorukesed ja nende mootorvalgud .
Vesiikulite
transport rakkudes. Et mootorvalkude saba kinnituks vesiikuli külge
peavad vesiikulite membraanis esinema spetsiifilised valgud,
retseptorid.
143.
Millistes kohtades rakus toimub mikrofilamentide ja millistes
mikrotorukeste polümeriseerumine, millised valgud vajalikud?
Mikrofilamentide
polümeriseerumine toimub rakumembraani läheduses, vajalikud
tümosiin ja profiliin. Mikrotorukeste polümeriseerumine toimub
tsentrosoomis rakutuuma lähedal, vajalikud ilmselt alfa- ja
beetatubuliinid...
144.
Kus ja kuidas toimub loomarakus mikrotorukeste moodustumine?
Mikrotorukesed on selle struktuuriga seotud kas (-) või (+) otsa
vahendusel? (alla kriipsutada õige variant).
Taimede
tsentrosoomides ja teatud tüüpi loomarakkudes (mõned
epiteelirakkude tüübid) tsentrioolid puuduvad ja järelikult pole
vajalikud mikrotorukeste kokkupanekuks. Katsed puhastatud tsentrosoomidega on näidanud, et nende lisamisel tubuliini
subühikute segule toimub kiire mikrotorukeste moodustumine – seega
tsentrosoomid
on kohaks kus algab mikrotorukeste moodustumine
nii, et (-) otsad jäävad seotuks tsentrosoomiga. Seega
mikrotorukeste orientatsioon rakkudes on polaarne. Subühikute
lisandumine toimub mikrotorukeste (+) otsale.
145.
Kirjeldage mikrotorukeste mootorvalke ja iseloomustage (sammu
pikkus, liikumise suund mikrotorukesel)
Mikrotorukeste
mootorvalkude klassid
Klass
Transporditav kompleks
Liikumissuund
Tsütosooli kinesiinid
Tsütosooli vesiikulid
(+)
Mitoosivärtna kinesiinid
Värtna mikrotorukesed, kinetohoorid (kromosoomid)
(+) ja (-)
Tsütosooli düneiinid
Tsütosooli vesiikulid, kinetohoorid (kromosoomid)
(-)
Aksonema düneiinid
Mikrotorukesed viburites ja ripsmetes
(-)
146.
Kirjeldage lühidalt aksoneemi ehitust ja nimetage piirkondi inimese
organismis, kus sellised struktuurid paiknevad. Kuidas aksoneemi
struktuur tagab liikumise?
Aksoneem
on membraani väljakasv mis sisaldab mikrotorukestest koosnevat struktuuri. See koosneb 9 välimisest mikrotorukestepaarist, mis
ümbritsevad kahte ühekaupa paiknevat mikrotorukest, kusjuures iga
paar koosneb A ja B torukesest. Kõikide mikrotorukeste + ots paikneb
aksoneemi raku tsentrist kaugemas osas. Seondub rakuga basaalkehakese
abil.
Liikumises
osaleb düneiin, mille vahendusel toimub ühe paari A torukese
liikumine teise paari B torukese - otsa poole. Teiste sidemete
esinemise tõttu mikrotorukeste paaride vahel tekib lainetav
liikumine.
A toruke sisaldab 13 protofilamenti, B toruke 10. Aksoneem on seotud
rakuga basaalkeakese abil. Selle strktuur on sarnane tsentrioolide
struktuuriga ja sisaldab 0 kolmest mikrotorukesest koosnevat rühma
milles 13 protofilamendist koosnev A toru on seotud ebatäieliku B toruga ja ebatäieliku C toruga. Basaalkehakese A ja B torud
jätkuvad viburis aga C tor lõpeb basaalkehakese ja viburite vahelises üleminekutsoonis. Inimkehas on näiteks inimese hingamisteed epiteelrakud ja spermatosoidide vibur . Aksoneemis
dimeersete torukeste A toru on soetud väliste ja sisemiste düneiini
molekulidega ja aksoneemi struktur säilib mitmete valkude abil.
147.
Millised haigused milliste tunnustega on seotud aksoneemi
häiretega.(Kartageneri sündroom)
Mutatsioonid viburite v ripsmete düneiinis
võivad põhjustada seda sündroomi mida iseloomustab isasteriilsus
kuna spermatosoidid ei liigu. Samuti vastuvõtlikkus
kopsuinfektsioonidele kuna liikumatud ripsmed ei eemalda hingmisteede
pinnalt baktereid.
148.
Kinetohoorsed, polaarsed ja astraalsed mikrotorukesed, nende
funktsioonid mitoosis, milliste mootorvalkude abil need
realiseeruvad?
Kinetohoorsed
mikrotorud seostuvad kromosoomidega ja osalevad kromatiidide
eraldamises üksteisest ja jaguneva raku poolustele liikumises.
Polaarsed
mikrotorukesed on seotud vasastsentrosoomist lähtuvate polaarsete
mikrotorukestega ja anafaas B toimumisega. Nende + otstega seostuvad
kinesiinid. Toimub erinevatelt poolustelt lähtuvate polaarsete
mikrotorukeste liikumine üksteise suhtes.
Astraalsed
mikrotorukesed paiknevad kiirjalt ümber tsentrosoomi ja osalevad
mitoosivärtna ruumilises fikseerimises ja selle pikemaks venitamisel
anafaasis. - suunalised mootorvalgud düneiinid rakkude kooreosas
membraani all tõmbavad astraalseid mikrotorukesi kaugemale (pooluste
kaugenemine).
Anafaas
A’s toimub liikumine mikrotorukeste – otsa poole siis võivad
osaleda düneiinid.
149.
Tsentrioolide funktsioonid rakutsüklis, tsentrioolide tsükkel.
Tsentrioolid
on seotud mitootilise värtna organiseerimisega ning tsütokineesi
lõpetamisega.
Tsentriool on väike silindrikujuline organell mille seina moodustavad 9
kolmekaupa agregeerunud mikrotorukest.
150.
Defineerige tsentrosoom , tsentromeer , kinetohoor.
Tsentrosoom
on valgulist materjali sisaldav piirkond, mis ümbritseb tsentrioole
ja tagab interfaasis mikrotorukeste moodustumise.
Tsentromeer
on kromosoomi DNA järjestus, millele seostub kinetohoor. Seob
omavahel õdekromatiidid.
Kinetohoor
on valguline kompleks, mis seostub kromatiidi tsentromeeri
piirkonnaga ja tagab mikrotorukeste seostumise kromatiidi külge
mitoosivärtna moodustumisel.
151.
Nimetage tsentrioolide funktsioone rakkudes lisaks mitoosile.
Osalevad viburite ja ripsmete moodustumises,
152.
Nimetage 3 taimeraku mitoosi iseärasust võrreldes loomarakuga
Tsentrioolid
puuduvad, rakuseina tõttu erinev tsütoplasma pooldumine . Raku
jagunemistasapinda moodustub Golgi kompleksi vesiikulitest ja neid
transportivatest mikrotorukestest fragmoplast. Vesiikulid sisaldavad
rakuseina materjali, nende kokkusulamisel tekibki uus rakusein .
153.
Kirjeldage kinetohoori ehitust.
Kinetohoor
on kihiline - võib eristada sisemist e kromosoomipoolset ja välimist
mikrotorukese poolset plaati, mis on omavahel valkudega ühendet.
Kinetohoor
on kihilise ehitusega ja võib eristada sisemist ehk kromosoomi
poolset ja välimist mikrotorukese poolset plaati mis omavahel teatud
valkudega ühendatud. Imetajate sisemises plaadis on leitud 4
erinevat valku, välimises plaadis on leitud düneiin ja kinesiini
sarnane valk CNP-E. On ka tsentromeeriga seostuvad faktorid .
154.
Kromosoomide liikumine anafaasis a ja b
--A:
kinetohoorsed mikrotorukesed lühenevad ja kromosoomid tõmmatakse
poolustele.
--B:
poolused kaugenevad üksteisest, mitoosivärten pikeneb.
155.
Millistes mitoosi protsessides osalevad kinesiinid ja millistes
düneiinid, millistes tubuliini polümeriseerumine ja
depolümeriseerumine
-Kinesiin:
anafaas B.
-Düneiin:
prometafaas, metafaas, anafaas B.
Polümeriseerub
prometafaasis; depolümeriseerub anafaasides A ja B.
156.
Mootorvalk liigub tsütoskeleti (-) otsa suunas. Millise
mootorvalguga võiks olla tegemist?
Mingi düneiin või mitoosivärtna kinesiinid.
157.
Millises mitoosiprotsessi osas sellised mootorvalgud
funktsioneerivad?
Metafaasis
kromosoomise pendeldamise ajal (mõlemad)
158.
Milliste mootorvalkude ja milliste tsütoskeleti valkude interaktsioon toimub loomaraku tsütokineesis?
a. DNA replikatsiooni
b. Mitoosivärtna teket
c. Kahe tütarraku eraldumist tsütokineesis - aktiini
rõngas
d. Viburite liikumist
159.
Tsütohalasiin B on mikrofilamentide moodustumist takistav ühend.
See ühend seetõttu takistab
.....................................(kirjutada, millise protsessi
toimumist mitoosis)
See ühend seetõttu takistab tsütokineesi
toimumist mitoosiprotsessis.
Meetodid
rakubioloogias
160.
Milliseid geenitehnoloogilisi meetodeid kasutatakse valgu
lokalisatsiooni muutuse jälgimiseks elusas rakus?
Bioluminestseeruvad/fluorentsed
lisandused rakku vastavale valgule.
161.
Kuidas toimite, kui tahate näha ühe spetsiifilise valgu
lokalisatsiooni rakus?
162.
Mille poolest erineb uuritavast objektist tavalise
fluorestsentsmikroskoobi abil saadav kujutis konfokaalse
fluorestsentsmikroskoobi abil saadavast kujutisest?
163.
Kuidas tuleb uuritavat preparaati töödelda, et see oleks vaadeldav
skanneeriva elektronmikroskoobiga?
164.
Milliste meetoditega on võimalik apoptoosi näidata.
Peroksüsoomid,
lüsosoomid, proteasoomid
165.
Lüsosoomide struktuur, suurus, ehitus, toimuvad protsessid
Lüsosoomid
on loomarakkudes üheks piirkonnaks, kus toimub kõige erinevamate
ühendite lagundamine. Lüsosoomidesse liiguvad ja lagundatakse seal
ka endotsütoosi ja fagotsütoosi teel rakkudesse sattuvad ühendid
ja organismid, aga samuti teatud ERst punguvates vesiikulites olevad
ained. Lüsosoomid on seotud ka vananenud ja mittefunktsioneerivate
organellide lagundamisega.
Lüsosoomid
on ühe membraanikihiga ümbritsetud organellid diameetriga~0.2-0.5
µm. Nende luumeni pH on happeline (4,5-5). Happelise pH tagab
V-tüüpi H+-ATPaas lüsosoomide membraanis, pumbates prootoneid
tsütoplasmast lüsosoomi.
166.
Kuidas kaitseb lüsosoom enda membraani hüdrolaaside lagundava mõju
eest?
Lüsosoomide
membraanis esinevad valgud on tugevasti glükosüülitud. Arvatakse,
et see kaitseb neid lüsosoomisiseste proteaaside lagundava toime
eest.
167.
Nimetage vähemalt kaks lüsosomaalset ladestushaigust ja kirjeldage mehhanism , mis haiguse tingib.
1)Pompe
haigus – puudub a-glükosidaas, mis vajalik glükogeeni
lagundamiseks
2)Harvaesineva
I rakulise haiguse (ingl. Inclusion-cell disease) korral puuduvad
praktiliselt kõik lüsosomaalsed ensüümid lüsosoomi transportiva
süsteemi häirete tõttu. Defektne on ensüüm, mis katalüüsib
lüsosoomi liikuvate valkude fosforüülumist. Seetõttu Golgi
kompleksi retseptorid ei tunne vastavaid valke ära ja need
transporditakse rakkudest välja, mitte lüsosoomidesse. Esinevad
veres. Lagundamata ained kogunevad rakkudes lüsosoomidesse
inklusioonidena
168.
Peroksüsoomide struktuur, suurus, ehitus.
Need
on väikesed ühekordse membraaniga organellid (0,2 – 1 µm diameeter ).
Peroksüsoomid
sisaldavad mitmesuguseid flavoproteiinseid oksüdaase (ensüüme,
mille kofaktoriteks on FAD või FMN), mis oksüdeerivad orgaanilisi
ühendeid hapniku kasutamisega. Sellisteks ensüümideks on näiteks
D-aminohapete oksüdaas, rasvhapete oksüdeerumises osalevad
ensüümid, lutsiferaas jt.
Peroksüsoomides
DNA ja ribosoomid puuduvad, seega kõik peroksüsoomide valgud peavad
saama imporditud tsütoplasmast
169.
Loetlege, millised biokeemilised protsessid toimuvad peroksüsoomides?
1)Rasvhapete
lagunemine toimub nii mitokondrites kui ka peroksüsoomides.
2)Eriti
pikaahelalised rasvhapped (süsiniku aatomeid> 20) lagundatakse
ainult peroksüsoomides.
3)Maksa
ja neerude rakkude peroksüsoomides lagundatakse mitmesugused
vereringes esinevad toksilised ühendid ( alkohol , formaldehüüd,
aromaatsed ühendid, ravimid ) vesinikperoksiidi kasutamisega näiteks
peroksüdaaside poolt.
170.
Kuidas peroksüsomaalsed valgud liiguvad tsütosoolist
peroksüsoomidesse?
Kuidas
satuvad ensüümid peroksüsoomidesse?
Need
ei satu sinna mitte Golgi kompleksi kaudu, nagu lüsosomaalsed
ensüümid, vaid peroksüsoomides vajaminevad ensüümid korjatakse
kokku tsütoplasmast. Äratundmissignaaliks on siin tripeptiid
Ser-Lys-Leu (SKL). Kui selline järjestus eksperimentaalselt mingile
valgule lisada, siis satub ta peroksüsoomi.
171.
Kirjeldage peroksüsoomide funktsioneerimise häiretega kaasnevaid
haigusi inimesel
Zellwegeri
sündroom on tingitud ensüümide
peroksüsoomidesse impordi häirest. Neil haigetel on aju, maksa ning
neerude talitluse häired ning nad surevad varsti peale sündi. Üks
selle haiguse vormidest on tingitud mutatsioonist peroksüsoomi
membraanivalku määravas geenis, mis ei võimalda ensüüme
korralikult importida. Selle tulemusel nende patsientide
peroksüsoomides pole peaaegu üldse vajalikke ensüüme ning
tagajärjeks on raske ainevahetuse häire.
X-liiteline
adrenoleukodüstroofia (ALD). Selle
haiguse puhul puudub peroksüsoomidest üks kindel ensüüm, mis seob
peroksüsoomis ko-ensüüm A (CoA) väga pika ahelaga rasvhapetele.
Ka selle haiguse primaarne põhjus peitub selles, et peroksüsoomi
membraanis ei funktsioneeri korralikult üks transportvalk, mis
vastutab selle ensüümi impordi eest. Tagajärjeks aga on see, et
rakk ei suuda lagundada pika ahelga rasvhappeid. Sellised lapsed on
pealtnäha terved kuni keskmise lapseeani, siis aga arenevad rasked neuroloogilised häired ning surevad paari aasta jooksul.
172.
Kirjeldage proteasoomide
ehitust, lokalisatsiooni rakus ja nendes toimuvaid protsesse.
Proteasoom
on ehituselt silindriline, koosneb valgulistest beeta- ja
alfa-subühikutest. Esineb kindlasti tuumas,
tsütosoolis,
vakuoolis, kloroplastis. Lagundab valke, mis on kovalentselt
ubikvitiiniga seotud. Proteasoom moodustub ATP abil (mitmest
kompleksist ühtseks) ning ADP ja Pi lahkumisel siseneb sinna
ubikvitiinijupp, mille taga on termineerimisele kuuluv valk. Valgud
harutatakse tagasi lahti (polüpeptiidid) ning ubikvitiin ja
proteasoom lagunevad.
173.
Iseloomustage chaperon’id ja nimetage protsesse milles osalevad.
Chaperon’id
on spetsiifilised saatjavalgud, mis stabiliseerivad ebastabiilsed
konformatsioonid, kiirendavad oligomeeride teket, valkude lagunemist
ja tagavad lokalisatsiooni kindlates raku piirkondades. Chaperon’ide
toime tsütosooli ribosoomidel toimub kotranslatsiooniliselt,
kloroplastide ribosoomidel translatsioonijärgselt.
174.
Iseloomustage chaperonin’id ja nimetage protsesse milles osalevad.
Chaperonin’id–
tagavad kokkukeerdumise. Mõjuvad struktuuri tekke hilisematel
staadiumitel. On oligomeersed valgud, mis moodustavad sisemusse kanali, kus valgu konformatsiooni teke toimub. Vajalik ATP. Esinevad
pro- ja eukarüootides.
Rakubioloogia I,
Kordamisküsimused:
rakutuum, geeniregulatsiooni sissejuhatus.
1.
Nimeta protsesse,
mis toimuvad eukarüootse raku tuumas.
DNA replikatsioon ,
reparatsioon(parandamine) ja RNA transkriptsioon , tuumatransport, RNA
subühikute kokkupanek
2.
Kus ja kuidas
paiknevad lamiinid ja mis on nende ülesanne?
Lamiinid paiknevad tuumamembraani
sisepoolusel ning ühendavad membraani kromatiiniga. Toetavad sisemembraani seespoolt.
3.
Milline on valkude
transpordi erinevus tsütoplasma-ER-i ja tsütoplasma-tuuma vahel?
Tsütoplasma-tuuma
vahel toimub transport läbi spetsiifiliste tuumapoooride, mis
lasevad NLS’i ja NES’i abil endast läbi kõrgmolekulaarseid
ühendeid. Valk kokkukeerdunud.
Tsütoplasma-ER’i
vahel toimub transport läbi translookonite, SRP(signaaliäratundja
partikkel)
determineerib, kas valk saab ER’i v mitte. Valk lahtikeerdunud.
Tsütoplasma-tuuma vahel toimub
transport läbi tuumapoori kompleksi, ehk läbi spetsiifiliste
pooride, mis on valmis läbi laskma kõrgmolekulaarseid ühendeid. (või pigem vaata küsimus number 5. vaja on signaale NLS ja NES +
tuuma transpordi retseptorid.)Tsütoplasma ja ER-i vahel toimub
transport aga läbi translookonite. Osa võtab SRP signal recognition particle , ehk signaali äratundev partikkel, mis on RNA
signaaljärjestus, mis ütleb, kas valk saab ER-i või mitte. Pmst
vaja on GTPd, SRP-d ja selle retseptorit moodustamas kompleksi ning
translookonit, kui signaaljärjestus seostub translookoniga siis
selle värav avaneb ja valk liigub ER-i sisse.
Erinevuseks
on veel see, et ER-i liikudes peab valk lahtikeerduma, tuuma minekuks aga ei pea.
4.
Kui suur on
tuumapoori difusioonipiir? Kirjelda lühidalt tuumapoori kompleksi
ehitust.
Tuumapoori difusioonipiir on 40kD –
ehk kuni 40kD suuruste molekulide difusioon toimib läbi NPC ilma
takistuseta, ei vaja spetsiaalseid kandjaid ega energiat.
Tuumapoori kompleks koosneb umbes
3000-5000 poorist, iga poor koosneb u 100st valgust, millest enamik
on tähistatud erineva numbriga nukleoporiinina. Tsütoplasmapoolsel
osal poorist jäävad väljaulatuvad filamendid, sissepoole jääb
tuuma korv . Poori moodustavad valgukompleksid on seotud ka tuuma
lamiinidega.
5.
Millised
valgumolekuli osad tagavad tuumaimpordi ja -ekspordi valkudele, mille
suurus ületab tuumapoori difusioonipiiri? Kirjelda neid lühidalt.
Tuumaimpordi tagab valkudele NLS
signaaljärjestus ( nuclear localization signal) ja ekspordi NES ehk
nuclear export signal. Mõlemad on lühikesed aminohappelised
järjestused. Ei lõigata kohe ära, sest võib korduvkasutuseks vaja
minna. Valgud ei pea olema lahti keerdunud .
6.
Kuidas
liigitatakse karüoferriine ning mis on rakus nende ülesandeks?
Karüoferriinid ehk tuuma
transpordi retseptorid, jagatakse eksportiinideks ja importiinideks.
Rakus on nende ülesandeks valke tuuma ja tuumast välja
transportida.
7.
Milline
makroergiline ühend tagab energia transpordil tuuma ja tsütoplasma
vahel ning milliste valkude vahendusel see transport teoks saab (3
valku)?
Energia transpordi tuuma ja
tsütoplasma vahel tagab GTPaase aktiivsusega Ran-valk. Tuuma
sisenedes on seotud GDPga, tuumas paikneb (Ran-)GEF-valk(guanine exchange factor), mis seostub Ran-valguga ja muudab selle GTP vormi.
Tuumast väljudes seondub Ran-valguga aga hoopis (Ran-)GAP(GTPase
activating protein ), mis viib valgu jälle GDP vormi. Transport
toimub GDP-GTP gradendi toimel. Ran-GTP vabastab import -retseptori
ehk importiini tema koormast.
8.
Kuidas paiknevad
tuumas kromosoomid ning kuidas paiknevad tuumas rohkem geene
sisaldavad kromosoomid?
Kromosoomidel on tuumas igal ühel
oma piirkond, territoorium . Rohkem geene sisaldavad kromosoomid
paiknevad tuumakeses tsentraalsemalt ja vähem geene sisaldavad
perifeersemalt.
9.
Mis eristab tuuma
organelle tsütoplasma organellidest?
Pole eraldatud membraaniga, ning
ained, mis asuvad mujal karüoplasmas võivad sinna vabalt
difundeeruda.
10.
Milline on kõige
tuntum tuuma organell/reaktsioonikeskus?
Tuumake ehk transkriptsioonivabrik
11.
Mis toimub
tuumakeses ja mitu tuumakest võib maksimaalselt inimese rakutuumas olla? Mis selle numbri määrab?
Tuumakeses toimib rRNA süntees ja
ribosoomide subühikute moodustumine. Inimese rakutuumas võib
maksimaalselt olla 10 tuumakest, sest diploidses kromosoomistikus on
10 NORi ehk tuuma organisatoorset regiooni, kus paiknevad rRNA
sünteesiks vajalikud geenid. Tavaliselt neid ei transkribeerita
korraga ning erinevad NORid koonduvad ühte piirkonda, seega
tuumakesi on enamasti palju vähem.
12.
Miks on ribosoomi
RNA geene eukarüootides palju?
Sest ribosoom koosneb kahest
subühikust ja need omakorda koosnevad erinevatest rRNAdest.
Eukarüootidel on 4 erinevat rRNAd, prokarüootidel 3. Prokarüoodid
toodavad umbes 55 erinevat valku, eukarüoodid 80.
13.
Miks ribosoomide
koostises olevat kolme RNA molekuli ühise pre-RNA-na sünteesitakse?
Tagab nende võrdse koguse.
14.
Mis on hetero - ja eukromatiin ? Kus paikneb rakutuumas enamasti heterokromatiin?
Millised kromosoomi osad liigitatakse konstitutiivseks
heterokromatiiniks? Miks?
Heterokromatiin
on kokkupakitud kromosoomide kogum, mille geene eriti ei
transkribeerita. Paikneb tuumas enamasti perifeerselt, lamiinidega
seotud tuumakeste ümber.
Eukromatiin
on rohkemkasutatav ja vähem kokkupakitud, paikneb tsentraalselt.
Konstitutiivne
heterokromatiin ei sisalda eriti geene, vaid kõrgelt korduvaid
tandeemseid järjestusi, mis enamus ajast tugevalt kokku pakitud.
konstitutiivne
heterokromatiin – pakitakse lahti ainult DNA kordistamise ajaks.
Heterokromatiin on kromosoomid,
mille geene vähe transkripteeritakse, rohkem kokkupakitud.
Eukromatiini kasutatakse rohkem, on vähem kokkupakitud.
Heterokromatiin paikneb enamasti tuumamembraani siseküljel olevate
lamiinidega seotult tuumakeste ümber. Eukromatiin on tsentraalsem.
Konstitutiivseks heterokromatiiniks nimetatakse sellist kromosoomi
osa, mis praktiliselt ei sisaldagi geene vaid kõrgelt korduvaid
tandeemseid järjestusi, mis enamus ajast tugevalt kokku pakitud.
15.
Milles seisneb
gefenide positsiooniefekt?
Kui heterokromatiin viia
eukromatiini alasse, hakatakse seda suure tõenäosusega
transkripteerima, kui eukromatiin viia heterokromatiini alasse
vaigistatakse selle avaldumine.
16.
Millised on kolm
kromosoomi säilitamiseks vajalikku struktuurielementi?
Telomeerid , tsentromeerid ja
replikatsiooni alguspunkt.
17.
Replikatsiooni alguspunkt
– milleks vajalik, erinevused pro- ja eukarüootidel.
Prokarüootidel ja lihtsamatel
eukarüootidel on kindlad replikatsiooni alguspunktid, DNA
järjestused, kuhu DNA polümeraas kinnitub. Prokarüootidel piisab ühest alguspunktist , originist. Kõrgematel eurkarüootidel on
replikatsiooni algus stohhastilisem, kuid reeglina replitseerub enne
eukromatiin, siis heterokromatiin.
18.
Tsentromeer – milleks
vajalik, kirjeldada lühidalt üldist ehitust. Kinetohoori ülesanne.
Tsentromeer on kromosoomi piirkond,
mis reguleerib kromosoomide liikumist mitoosi ja meioosi ajal. Kindla
DNA järjestusega(100 kuni miljoneid aluspaare tandeemselt korduvates
järjestustes) ja sinna seonduvate valkudega. On heterokromatiin, mis
pakitakse lahti ainult replikatsiooni ajal. Hoiab õdekromatiide kuni
mitoosi anafaasini koos. Tsentromeeri külge seondub omakorda
kinetohoor, kuhu seonduvad mikrotuubulid, mis kromatiidid üksteisest
eemale tütarrakkude erinevatele poolustele tõmbavad.
19.
Telomeer –
milleks vajalik, kirjeldada lühidalt üldist ehitust.
Telomeer on kromosoomi otsa kaitsev
spetsiifiline kordusjärjestus. Kindlustab kromosoomi otste
replikatsiooni, kaitseb nukleaaside eest ning hoiab ära kromosoomide
„kokkukleepumise’’. Koosnev u 600-2500 aluspaarist, 3’ ots on
200bp ulatuses üleulatuv, lassotaoline, moodustab T- lingu .
20.
Miks telomeerid
lühenevad?
Oksüdatiivne stress ; DNA lüheneb iga replikatsioonitsükliga (RNA praimeri
seondumiskohas pole enam nukleotiide ).
Telomeeride otsi pikendab 3’
otsast telomeraas. Enamus somaatilistes rakkudes on telomeraas
inaktiivne ning rakud saavad jaguneda vaid kindla arvu kordi , sest
iga replikatsioonitsükliga jääb DNA lühemaks. See tuleb sellest,
et DNA sünteesil mahajäävale ahelale RNA praimeri seondumiskohta
ei täida nukleotiididega enam miski. Telomeerid
on väga vastuvõtlikud ka oksüdatiivsele stressile. Põhjustab
isegi suuremat kadu kui see viimase RNA praimeri värk.
21.
Mis juhtub rakuga,
kui telomeerid on raku põlvkondade vältel täielikult kadunud?
Hakkab kaotsi minema geneetiline
materjal, DNA kahjustuse korral peatub rakutsükkel, rakk ei jagune
enam ning sureb .
22.
Milline ensüüm
pidurdab osades rakkudes telomeeridel lühenemast? Mil viisil see pidurdamine toimub?
Telomeraasi
RNA on telomeerse DNA’ga komplementaarne ja toimib telomeeri pikendamisel matriitsina.
Telomeraas- selle RNA komponent on
telomeerse DNAga komplementaarne ja toimib telomeeri pikendamisel
matriitsina.
23.
Millistes rakkudes
on vaja aktiivset telomeraasi?
Erinevate organite tüvirakud,
naha, soolestiku, vere tüvirakud, sugurakud .
24.
Miks prokarüoodi
DNA pole nii tihedalt kokku pakitud kui eukarüoodi DNA?
Prokarüootide geenid on tihedamalt koos, rohkem ekspresseeritud, kogusummas on neid aga vähem.
Kokkupakkimine pole nii otstarbekas kui eukarüoodil.
25.
Millises
rakutsükli faasis on eukarüoodi kromosoomid kõige rohkem kokku
pakitud? Mitu korda on DNA selles struktuuris kondenseerunud? Kuidas
erineb kromatiini kokkupakitus nn G- ja R-bändides?
Mitoosi
metafaasis on kromosoomid 10 000x lühemad kui DNA kaksikheeliksis.
G-bändis on rohkem kokku pakitud, R-bändis vähem (Giemsa värvid).
Kromosoomid on kõige rohkem kokku
pakitud mitoosi metafaasis. Nad on 10 000x lühemad kui DNA
kaksikheeliks. G- ja R- bändid tulenevad sellest, kuidas need Gimsa
värvidega värvides värvuvad, tumedamad on G ja heledamad R,
tumedamalt värvuvad rohkem kokku pakitud kromatiinid.
26.
Kuidas ühe liigi
mitoosi metafaasi kromosoomidest koostatud kogumit nimetatakse ning
mida selle abil teha saab?
Karüotüüp, selle abil saab
määrata kromosoomide ümberpaiknevusest tingitud haigusi.
27.
Eukarüoodi DNA
esimene ja teine pakkimistasand – kuidas neid nimetatakse? Mitu
korda need DNAd lühendavad?
I
pakkimistasand – pakkimine nukleosoomi – lühendab 7x
II
pakkimistasand – 30 nm fiiber / solenoid - lühendab 40x
28.
Millistest
histoonidest moodustub nukleosoom (mitu histooni ja millised
histoonid)? Milline histoon sellesse struktuuri ei kuulu ja mis on
tema teadaolevad ülesanded tuumas?
Nukleosoomi
moodustavad histoonid H2A, H2B, H3 ja H4 - kõiki 2x. Histoon H1
stabiliseerib ja hoiab nukleosoomi sidemeid püsivatena ning vastutab
kõrgemate pakkimistasandite stabiilsuse eest.
Nukleosoom moodustub histoonidest
H2A,H2B, H3,H4, kõiki neid on kahekordselt. On olemas ka histoon H1,
mis vastutab nukleosoomide omavaheliste sidemete püsivuse eest,
stabiliseerib nukleosoomi ja osaleb kõrgemate pakkimistasandite
stabiilsuse eest. On väga liikuv ja teised funktsioonid teadmata.
29.
Millised
mittehistoonsed valgud aitavad kromatiini kõrgemat järku
struktuuridesse pakkida ja millised piirkonnad on selle jaoks DNAs?
Kromatiini
aitavad kõrgemat järku struktuuridesse pakkida kondensiinid ja
kohesiinid. DNAs seonduvad mittehistoonsete valkudega piirkonnad SAR
ja MAR.
Mittehistoonsed valgud on näiteks
kondensiinid ja kohesiinid (dimeersed, vabad otsad seotud
kromatiiniga, hüdrolüüsivad ATPd DNA liigutamiseks). Osaleb ka DNA topoisomeraas II. DNAs on mittehistoonsete valkudega seondumiseks
piirkonnad SAR(toesega seotud piirkond) ja MAR(maatriksiga seotud
piirkond).
30.
Milles seisneb
C-väärtuse paradoks ?
Genoomi suurus ei määra organismi
keerukustaset.
31.
Kirjeldage geeni
ehitust.
Geen on DNA lõik, mis
transkripteeritakse ühe üksusena ja kannab infot ühe valgu kohta.
Geeni hulka loetakse ka intronid ja transkriptsiooni
regulaatorpiirkonnad. Promootorpiirkond, kuhu seondub RNA polümeraas
transkriptsiooni algatamisel ja terminaatorpiirkond mis lõpetab
transkriptsiooni.
32.
Mida mõeldakse
eukarüoodi geenide segmenteerituse all ning mis toimub splaissingu
käigus?
Segmenteerituse all mõeldakse
seda, et DNAs on intronid ja eksonid , eksonite geneetiline materjal
jõuab valgu kujule, intronid lõigatakse RNAst splaissingu käigus
välja.
33.
Milliseid
ülesandeid võib täita rakus RNA (mRNA-d mitte arvestades)?
Nimetage selle kursuse raames käsitletud
funktsioone/organelle/ensüümikomplekse, kus RNA on osaline.
RNA
aitab moodustada sekundaarstruktuure, transportida valke ER-i,
osalevad X kromosoomi inaktivatsioonil ja telomeeride sünteesis.
rRNA
– ribosoomide struktuuriüksus, katalüüsib valkude sünteesi
tRNA
– transpordib RNA ribosoomile.
snRNA
– osaleb splaissingus
siRNA – võimelised geeniekspressiooni välja lülitama.
34.
Kui suure osa
inimese genoomist moodustavad valke kodeerivad järjestused ehk
eksonid? Mis on pseudogeen ?
Eksonid moodustavad 1,5%.
Pseudogeen on introniteta ja promootoriteta, ei ekspresseeru. (mRNA
pöördtranskripteeritud molekul)
Pseudogeen on
mRNA pöördtranskripteeritud molekul, ei ekspresseeru, puuduvad
intronid ja promootorid.
35.
Millised
kromosoomi osad liigitatakse kõrgelt korduva DNA hulka? Kust tuleneb
nende nimetus „ satelliit -DNA“?
Tsentromeerid ja telomeerid.
Nimetatakse nii, sest need piirkonnad eralduvad muust DNAst
tihedusgradiendist tsentrifuugimisel, kuna sisaldavad rohkem A-T paare .
36.
Millisesse kahte
rühma liigitatakse mobiilsed DNA elemendid ja kuidas võib nende
kahe rühma liikumise mehhanismi põhimõtteliselt ja väga lühidalt
kirjeldada?
Transposonid ja retrotransposonid.
Transposonid on ’’hüppavad’’ DNA elemendid, liiguvad
cut- paste meetodil genoomis ühest kohast teise, enamasti ühe
kromosoomi piiris . Retrotransposonid liiguvad aga copy -paste
meetodil, geen jääb vanasse kohta alles. DNAlt sünteesitakse mRNA, sellelt omakorda pöördtranskriptaas, RNA+pöördtranskriptaas
otsivad DNAs uue koha, RNA pöörtranskripteerib end tagasi DNAks ja
integraasi toimel liigub uude kohta.
37.
Kust tuleneb
retrotransposoni „ retro “ nimetus?
Retrotransposonide
liikumismehhanism sarnaneb retroviiruste omadega.
38.
Millised ensüümid
vahendavad transposonide (1) ja retrotransposonide (2)
transponeerumist? Kui suure protsendi inimese genoomist moodustavad
transponeeruvad elemendid?
Transposonitel
transposaas, retrotransposonil pöördtranskriptaas ja integraas.
45%.
Transponeerumiseks on vaja
liikumist tagav geen, mis tuleb transkipteerida valguks nimega
transposaas. Transposonid moodustavad inimese genoomist 3%.
Retrotansposonil on sees pöördtranskriptaasi ja integraasi geen.
Moodustavad 8+34% Kokku siis transponeeruvad elemendid 45% genoomist.
39.
Kuidas liigitatakse
retrotransposone ning milline neist kahest alarühmast on praegugi
inimpopulatsioonis aktiivne? Nimeta selle alarühma kolm enim
kirjeldatud mobiilset elementi.
Retrotransposone liigitatakse
viiruslikeks ja mitte viiruslikeks, mitteviiruslikud on praegugi
aktiivsed. Suurimad rühmad neist – LINE, Alu, Sva elemendid.
40.
Kas
transponeeruvate elementide aktiivsus on hea või halb? Põhjendage
vastust.
Puudub ühene vastus. Aktiivsed
transponeeruvad elemendid põhjustavad erinevaid mutatsioone , mis
võivad olla kahjulikud, kui mõnikord ka kasulikud, sest võisid
evolutsiooniliselt tuua kaasa kasulikke kohastumisi (DNA
duplitseerumis ja ümberkorraldusi).LINEd võivad olla eellasteks
telomeraasile ning võisid mängida suurt rolli immuunvastuse
spetsiifika välja kujunemisel.
41.
Milleks rakutuuma
olemasolu hea võiks olla?
Rakutuuma olemasolu võib hea olla
selleks, et transkriptsioon ja replikatsioon toimuks muust rakust
eraldi, et transkriptsioonifaktorid saaks kiirelt reageerida
väliskeskonnale. Võimaldab paremat kontrolli, kas faktor saab üldse
tuuma sisse või mitte.
42.
Tooge näiteid
valkudest, mis peavad igas rakutüübis ekspresseerunud olema. Kuidas
neid valke ühiselt nimetada võiks?
nn. Koduhoidjavalgud - hoolitsevad
ekspresseerumise eest ehk nii replikatsiooni, transkriptsiooni kui
translatsiooni eest. DNA polümeraas, histoonivalgud,
ribosoomivalgud, energia ja ainevahetuse valgud.
43.
Kui palju
arvatakse (hetkel) inimesel geene olevat ning kui suur osa neist
korraga ekspresseeruda võib? Kui suur arvatakse olevat geenide
protsent, mille produktid geeniregulatsioonis osalevad?
Inimesel arvatakse olevat umbes
23-25 000 geeni, milleks korraga ekspresseeruda võib 30-60%. Geeniregulatsioonis osalevate geenide protsent arvatakse olevat
10-15% kõikidest geenidest .
44.
Millised
cis-elemendid
on vajalikud, et RNA polümeraas ühe geeni transkribeerimise
alustaks ja lõpetaks?
Promootor, enhanser, terminaator.
(cis-elemendid asuvad DNAs, mõjutavad transkriptsiooni.
Trans-faktorid asuvad RNAs ja mõjutavad transkripsiooni,
trans-faktorid on näiteks kõik transkriptsioonifaktorid)
45.
Mis tagab enamike
valguliste transkriptsioonifaktorite DNA-ga seondumise
spetsiifilisuse?
Tõmbejõud,
mis tekivad hüdrofoobsestest vastasmõjudest, elektrilisest laengust
või teistest molekulaarsetest äratundmismehhanismidest.
Transkriptsioonifaktoride DNAga
seondumise tagavad tõmbejõud, mis tekivad hüdrofoobsetest
vastasmõjudest, elektrilisest laengust või teistest
molekulaarsetest äratundmismehhanismidest – näiteks heeliks -ling-heeliks, heeliks-pööre-heeliks, leutsiini tõmblukk,
tsinksõrmed, peptiidiling, beeta-leht.
46.
Kuidas saab RNA
polümeraasi ligipääsu geeni ees asetsevale promootorile takistada?
Kuidas saab seda soodustada?
Nt
repressoritega. E.coli näitel - keskkonnas leiduv trüptofaan
seondub repressorvalguga ja tekitab seega konformatsioonimuutuse.
Suundudes promootorile takistab translatsiooni algust.
Eukarüootidel
tähtis mediaator , mis takistab v soodustab RNA polümeeri
seondumist.
Saab kasutada repressoreid. Näiteks
kui E.coli keskkonnas on trüptofaani, seondub trüptofaan
repressorvalguga, mis tekitab konformatsioonimuutuse, repressor
seondub seepeale promootorile ja takistab translatsiooni algust.
47.
Milliste
vahenditega tagatakse võrreldes prokarüootidega eukarüootide
geeniregulatsiooni paindlikum kontroll?
Eukarüootide nukleosoomi koostises
on histoonid,
konserveerunud valgud, mille N-terminaalses otsas asuvast
aminohappest ja selle modifitseerumisest sõltub, kas geen
ekspresseerub või mitte.
48.
Nimeta histoonide
kovalentseid modifikatsioone. Milline neist põhjustab DNA ja
histoonide vaheliste elektrostaatiliste jõudude nõrgenemise?
Atsetüleerimine, metüleerimine,
fosforüleerimine, ubikvitineerimine. Atsetüleerimine põhjustab
histoonide ja DNA vahelise elektrostaatilise jõu nõrgenemise, mis laseb RNA polümeraasil kergemini DNA molekuli vahel liikuda
promootorini.
49.
Miks on imetajate
emasorganismis vajalik ühe X-kromosoomi heterokromatiiniks
kondenseerimine?
Et vältida olukorda, kus
emasorganism hakkab geeni korraga kahelt kromosoomilt ekspresseerima,
põhjustades geeniproduktide ebavõrdsuse emas-ja isasorganismide
vahel.
50.
Miks nimetatakse
inaktiveerunud X-kromosoomi fakultatiivseks heterokromatiiniks?
Mis juhtub tunnustega, mis
on määratud X-kromosoomil olevate geenidega ?
Inaktiveerunud X-kromosoomi
nimetatakse fakultatiivseks heterokromatiiniks(Barri kehake )
see-pärast, et tema geenid on vaigistatud, kromosoom on
kondenseerunud ja asub nagu heterokromatiinile kohane – tuumas
perifeersel, membraanilähedases kohas. Siiski 10% geenidest
ekspresseeruvad siiski.
Tunnused
avalduvad, ent mitte täielikult.
51.
Mis vahendab ühe
X-kromosoomi inaktivatsiooni? Millisel hetkel inaktivatsioon toimub
ja kui kaua kestab?
Inaktivatsiooni indutseerib RNA. X
kromosoom transkribeerib RNA, mis jääb tuuma ja seondub
inaktiveeritava X kromosoomi DNAga, indutseerib
heterokromatiseerumise. Inaktivatsioon algab varases loote arengus
suvalisel ajal ning kestab elu lõpuni.
52.
Milles seisneb
alternatiivne splaissing ja kuidas võiks see kaasa aidata
organismide paremale kohastumusele keskkonnamõjude suhtes?
Alternatiivne splaissing tähendab
geeni võimet eksonitest erinevate mRNAde kokkupanekut. Võimaldab
erinevate geeniproduktide teket, ehk reageerida erinevatele
keskkonnamõjudele, kasutades sama algmaterjali. Valgud on võimalik
vastavalt vajadusele eri moodulitest kokku monteerida.
53.
Millise kahe
protsessi toimumine on peamiseks märguandjaks, et mRNA on valmis
tuumast välja transportimiseks?
Splaissing, 3’ ja 5’ otste
märgistamine- 5’ otsa lisandub cap-struktuur, 3’ otsa
modifitseeritakse adeniini nukleotiididega.
54.
Millises mRNA
molekuli osas asub info tema rakusisese suunamise kohta? Too üks
näide, kus mRNA suunamine/gradient rakus olulist rolli mängib.
mRNA 3’ otsas asub
mittetransleeritav piirkond.
Näiteks
munarakus on olemas teatud kohtades teatud mRNA, mis omab
arengubioloogilist rolli. Näiteks pea arenguks vajalik mRNA asub
raku asukohas , kus see just tähtis on.
Või
närvirakkudes toimib ennetamine, kus signaalikandeks vajalik mRNA
asub aksonite piirkonnas, et info kanduks kiiremini.
55.
Mida mõeldakse
selle all, et geeniekspressioon on mürarikas? Mis on selle tulemus?
Ekspressioonil on suur roll
juhuslikkusel. Molekule küll vähe, aga reaktsioonide toimumise
sagedus varieerub palju. Muudab bioloogilised protsessid kõikuvaks,
raskesti uuritavaks.
32













































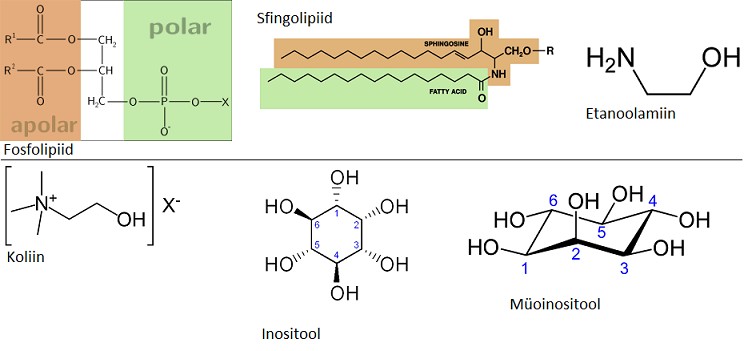
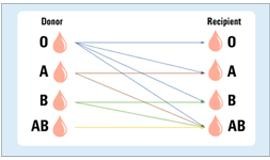
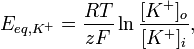




Kõik kommentaarid