UUED TEADUSLIKUD FAKTID HÜPOTEES TÕESTATAKSE või LÜKATAKSE ÜMBER (e HÜPOTEES PEAB OLEMA FALTSIFITEERITAV) PÄDEVA TEADUSLIKU TEOORIA ALUSEL ON VÕIMALIK ENNUSTADA NÄHTUSI/FAKTE, MILLE OLEMASOLU HILJEM EKSPERIMENTAALSELT TÕESTATAKSE 2. Elu organiseerituse tasemed - MOLEKULAARNE tase molekulaarbioloogia, geenitehnoloogia, süsteemibioloogia (BIOMOLEKULID ainult ELUSlooduses). Esmane organiseerituse tase. Kõikjal, kus on elu, esinevad biomolekulid: sahhariidid, lipiidid, valgud, nukleiinhapped. - ORGANELLI tase (molekulaarne) rakubioloogia. Uuritakse raku organelle: tuum, ribosoomid, mitokondrid jne. Kui need rakkudest eraldada, ei kanna nad enam elu tunnuseid. Organellide koostööst tulenevad rakkude omadused. - RAKU tase rakubioloogia
(võrdlev anatoomi, geenijärjestuse võrdlus), katse (kui muudetakse üht tingimust ja võrreldakse tulemusi nii muudetud kui muutmata tingimustega katse puhul) 1)Probleemi püstitamine 2)Taustinfo kogunemine 3)Hüpoteesi sõnastamine 4)Hüpoteesi kontrollimine 5)Tulemuste analüüs ja järelduste tegemine 2. Eluslooduse organiseerituse tasemed 1) MOLEKULAARNE tase – molekulaarbioloogia, geenitehnoloogia, süsteemibioloogia . Esmane organiseerituse tase. Kõikjal, kus on elu, esinevad biomolekulid: sahhariidid, lipiidid, valgud, nukleiinhapped. 2) ORGANELLI tase – (molekulaarne) rakubioloogia. Uuritakse raku organelle: tuum, ribosoomid, mitokondrid, lüsoosoomid jne. Kui need rakkudest eraldada, ei kanna nad enam elu tunnuseid. Organellide koostööst tulenevad rakkude omadused. 3) 3)RAKU tase – rakubioloogia, tsütoloogia
tulemusi nii muudetud kui muutmata (st kontroll) tingimustega katse puhul Biokeemilised meetodid Biofüüsikalised meetodid (nt valkude struktuuri analüüs) Mikroskoopia (valgus- ja elektronmikroskoopia) Geneetilised meetodid (mutatsioonanalüüs koos molekulaargeneetikaga) Eluslooduse organiseerituse tasemed MOLEKULAARNE tase – molekulaarbioloogia, geenitehnoloogia, süsteemibioloogia (BIOMOLEKULID ainult ELUSlooduses) ORGANELLI tase – (molekulaarne) rakubioloogia RAKU tase - rakubioloogia KOE tase - histoloogia, arengubioloogia/embrüoloogia. Inimesel põhikoed: epiteel-, lihas-, närvi- ja sidekude ELUNDI tase – ERI KOED (Tissues) moodustavad ELUNDID e. ORGANEID (anatoomia, füsioloogia). Organitest moodustuvad ELUNDKONNAD e. ORGANSÜSTEEMID (füsioloogia)
1) Tuum 2) Tuumake 3) Ribosoomid 4) Mitokoondrid 5) Lüsosoomid 6) Golgi kompleks 7) Tsütoplasmavõrgustik (kareda- ja siledapinnalist (kareda-pinnaline ER)) 8) Tsütoplaasm 9) Tsütoskeleet 10) Rakukest (tselluloosist, ligniinist, pektiinist.) 11) Vakuool 12) Platiidid (Leukoplastod, hloroplastis ja hromoplastid) 13) Rakumembraan 18. Rakukesta ehitus ja funktsioon Sarapuu õpik Rakuehitus ja talitus lk 18-20(64-66) 19. Geenitehnoloogia mudelorganismid *soolekepike *pärm *poolduv pärm *ümaruss *äädikakärbes *sebrakala *koduhiir *rändrott *müürlook *riis täpsemalt on powerpointis igast liigist juttu Selle leiate loengud 2011 alt ja see on pealkirjaga Geenitehnoloogia I 2011 -Rakk - rakutüübid-koed-ECM ja MUDELORGANISMID 20. DNA pakkimine, kromosoomide ehitus Sarapuu õpik Rakuehitus ja talitus lk 8-9(54-55) 21. Replikatsioon
toimu. Nõgus- ja kumerplasmolüüs erinevad plasmolüüsunud osa kuju poolest. Rakke liidab rakuvaheaine (vahelamell), vanemate rakkude nurkade vahel võib olla ka rakuvaheruume ehk intertsellulaare. Raku ehitusest paremaks arusaamiseks pidage silmas, et see pole mitte jäik, staatiline moodustis, vaid raku ehitus ja koostis muutuvad pidevalt ja küllaltki kiiresti: organellid jagunevad või kujunevad ümber, keemiline koostis muutub jne. 19. Geenitehnoloogia mudelorganismid *soolekepike *pärm *poolduv pärm *ümaruss *äädikakärbes *sebrakala *koduhiir *rändrott *müürlook *riis täpsemalt on powerpointis igast liigist juttu Selle leiate loengud 2011 alt ja see on pealkirjaga Geenitehnoloogia I 2011 Rakk rakutüübidkoedECM ja MUDELORGANISMID 20. DNA pakkimine, kromosoomide ehitus Sarapuu õpik Rakuehitus ja talitus lk 8-9(54-55) 21. Replikatsioon
Geenitehnoloogia eksam 1. Suhkrute lühiiseloomustus. Süsivesikud=sahhariidid. On orgaanilised ühendid, mille koostises esinevad süsinik, vesinik ja hapnik. Süsivesikud säilitavad rakusiseselt keemilist energiat. Rakk saab energiat suhkrumolekulide lagunemisel lihtsateks ühenditeks, aeroobidel veeks ja süsihappegaasiks. I Monosahhariidid ehk lihtsuhkrud on madalamolekulaarsed ühendid, milles süsinike arv on enamasti kolmest kuueni- riboos ja desoküriboos (5 süsinikulised). Glükoos ehk viinamarjasuhkur- kiire energiaallikas, näitab veresuhkrutaset. Funktsioon- energeetiline, DNAs ja RNAs ehituslik (6 süsinikuline). Rohelistes taimedes moodustub glükoos fotosünteesi tulemusena, loomorganismid saavad seda toidust. Fruktoos ehk puuviljasuhkur. II Polüsahhariidid on kõrgmolekulaarsed orgaanilised ühendid (polümeerid), mille ehituslikeks lülideks (monomeerideks) on monosahhariidid. Neil on energee
Kordamisküsimused Geenitehnoloogia I 1. Millised molekulid on polümeerid? Polümeerid ehk kõrgmolekulaarsed ühendid on ained, mille molekulid koosnevad kovalentsete sidemetega seotud korduvatest struktuuriühikutest elementaarlülidest. Looduslikud polümeerid: polüsahhariidid (tselluloos, kitiin, tärklis), valgud, nukleiinhapped (DNA, RNA). Polümeerid on väga suured molekulid, moodustunud kui sajad monomeerid liituvad pikkadeks ahelateks. 2. Nukleotiidide lühiiseloomustus. Nukleotiidid on orgaanilised molekulid, mis moodustavad suuri biopolümeere- nukleiinhappeid, näiteks DNA ja RNA. Nukleotiidid on DNA ja RNA molekuli alaüksused, mis koosnevad lämmastikalusest (N-alus), suhkrust (riboos või desoksüriboos) ja fosfaatrühmast. Lämmastikalused on kas puriini või pürimidiini derivaadid. Puriinid: kahte lämmatikku sisaldava tsükliga ühendid, aden
Molekulaarbioloogia Molekulaarbioloogia – tegeleb päriliku info kodeerimise, säilitamise ja ülekande mehhanismi uurimisega, samuti päriliku info realiseerumise molekulaarsete mehhanismidega (kuidas info geenides määrab elusorganismi ehituse ja tema funktsioneerimise. Uurib füüsikalis-keemiliste struktuuride ja biokeemilis-füsioloogiliste funktsioonide vastavust. Teadussuund hakkas arenema pärast makromolekulide ruumilise struktuuri kindlakstegemist (DNA 3-ruumiline struktuur). Molekulaarbioloogia dimensioon – 1 A – 300 A (üle 500 – rakubioloogia, alla 1 - biofüüsika) 1 A (ongström) = 10 -10 m 1nm = 10 A 2-ahelalise DNA läbimõõt – 20 A kovalentne side – 1,5 A globulaarse valgu d – 50 A dsDNA (double stranded) d – 50 A ribosoomide, valgumolekulide d – 200-300 A DNA aluspaaride vahe – 3,4 A vesiniksideme pikkus – 3 A nukleosoom – 60x110x110 A bakteri ribosoom – 200x200x230 A tuumapoorid – 120x120x75 A bakteriaalne RNA polümeraas – 90x90x60







































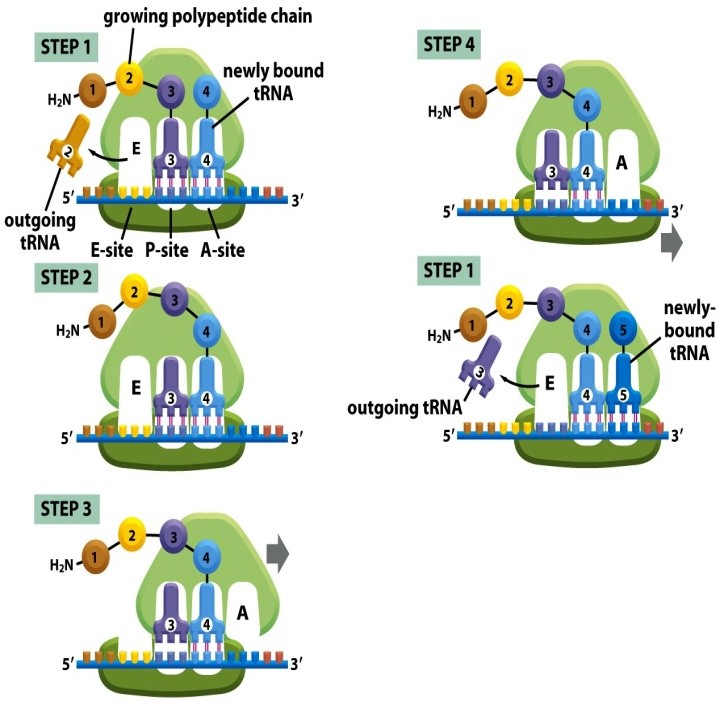




Kõik kommentaarid