Geenitehnoloogia kordamisküsimused
1.Suhkrute
lühiiseloomustusSuhkrud ehk
sahhariidid on orgaanilised ained, mille koostisse kuuluvad
süsinik, vesinik ja hapnik. Sahhariidid jaotatakse kolme rühma
mono -, oligo- ja
polüsahhariidid . Monosahhariidid ehk
lihtsuhkrud koosnevad enamasti kolmest kuni kuuest süsinikust. Neist tähtsamad
on viiesüsinikulised
riboos ja desoksüriboos, mis kuuluvad
nukleiinhapete koostisesse. Lisaks on olulised kuuesüsinikulised
glükoos ehk
viinamarjasuhkur ja
fruktoos ehk puuviljasuhkur, mis
mõlemad on olulised makroenergilised molekulid, mida organismid
kasutavad oma elutegevuseks.
Oligosahhariidid on orgaanilised
ühendid, mis on
enamuses moodustunud kahe- kolme monosahhariidi
(disahhariidid) ühinemisel. Näiteks võib tuua sahharoosi (roo-ja
peedisuhkur), mis on moodustunud glükoosi ja fruktoosi ühinemisel,
maltoosi ehk linnasesuhkru, mis on moodustunud kahest glükoosijäägist
ja laktoosi ehk piimasuhkru, mis on moodustunud glükoosist ja
galaktoosist. Neid suhkruid kasutatakse samuti peamiselt energia
saamiseks. Polüsahhariidid on kõrgmolekulaarsed orgaanilised
ühendid, mis koosnevad monosahhariidide lülidest ehk monomeeridest.
Tuntumad polüsahhariidid on tärklis, fotosünteesi käigus
sünteesitud glükoos, mis on talletatud taime sälitusorganitesse,
tselluloos, samuti sünteesitud glükoosist ning mis on taime
rakukesta ja tugikoe rakkude peamine
koostisosa ,
kitiin , lülijalgsete
välisskeleti ja seente rakukesta peamine koostisosa, ja glükogeen,
loomne tärklis, mida sälitatakse glükoosivarudena
maksas ja
lihastes.
Sahhariide kasutatakse peamiselt energia saamiseks,
varuainena ning organismis struktuuride koostisosadena.
2.
Lipiidide iseloomustusLipiidid on vees mittelahustuvad orgaanilised ühendid, mille alla kuuluvad
rasvad ehk
lihtlipiidid , õlid, vahad ja
steroidid .
Lipiide kasutavad
organismid energiaallikana, nende oksüdeerumisel vabaneb 2 korda
rohkem energiat ehk 38,9 kJ/g kui sama koguse sahhariidide või
valkude lagunemisel. Samuti kaitseb lipiididest moodustunud rasvkude
kõhuõõnes paiknevaid organeid kahjulike välismõjude eest ja
aitab välitada keha liigset jahtumist. Rasvad ehk lihtlipiidid on
glütseroolist ja rasvhappejääkidest koosnevad estrid.
Lihtlipiidide ühinemisel teiste keemiliste ühenditega moodustuvad
liitlipiidid , näiteks rakumembraani koostisesse kuuluvad
fosfolipiidid. Õlid on vedelad rasvad, mille rasvhappejäägid
sisaldavad ohtralt
kaksiksidemeid . Vahad on lipiidid, mille
molekulides esinevad glütserooli asemel muud alkoholid, näitek
taimsed vahad puuviljadel, okastel, mis täidavad kaitsefunktsiooni;
loomsed vahad näiteks mesilasvaha (
mesilaste kärjed). Steroidid
madalmolekulaarsed tsüklilised ühendid, mille hulka kuuluvad
kolesterool , hormoonid ja
vitamiin D. Hormoonid on bioaktiivsed
ained, mis reguleerivad ja koordineerivad samaaegselt mitme
elundkonna talitusi, näiteks neerupealised hormoonid ja
suguhormoonid
testosteroon ning östrogeen.
Kokkuvõte-
lipiidide funktsioonid
- Energeetiline funktsioon. Lipiidide koostises olevad rasvhapped on olulised energia saamise seisukohast – lipiidid on kõige energiarikkamad inimtoidu komponendid: 1g annab 38,9 kJ, so 9,3 kcal
- Ehituslik funktsioon. Fosfolipiidid ja kolesterool kuuluvad rakumembraani koostisse.
- Varuaine funktsioon. Loomadel varurasv , taimedel õlid seemnetes, viljades ja mesilaskärjed (vahad).
- Ainevahetuslik funktsioon. Metaboolse vee teke - lipiidide lõplikul lõhustumisel moodustuvad vesi ja süsihappegaas. Omane kõrbeloomadele nagu kaamel , kes üldse ei joo.
- Kaitsefunktsioon.
- Nahaalune lipiidide kiht, kui ka siseorganite ümber olevad lipiidid kaitsevad mehhaaniliste põrutuste eest.
- Nahaalune lipiidide kiht kaitseb keha mahajahtumise eest.
- Veelindudel kaitseks märgumise eest.
- Rasvkoes võivad talletuda kehavõõrad ained (mürgid).
- Pruun rasvkude, kus toimub aktiivne rasvhapete lõhustumine on oluline imikute soojusregulatsioonis, samuti talveunest ärkavatel loomadel aga ka talisuplejatel.
- Lahusti funktsioon. Veres olevad lipoproteiinid kannavad rasvlahustuvaid vitamiine organismi kõikidesse kudedesse.
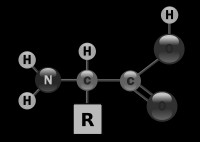
3.Aminohapete
ja valkude lühiiseloomustusValgud ehk
proteiinid on aminohapetest moodustunud polümeerid. Erinevaid
aminohappeid võib olla
kuni
20 valkude ehituses.
Aminohapped koosnevad aluseliste omadustega
aminorühmast (NH2),
happeliste omadustega karboksüülrühmast (COOH) ning molekuli
ülejäänud osa on erinev kõigil aminohapetel. Selletõttu ongi
neil mitmesugused keemilised omadused. Aminohappeid tähistatakse
kolmetäheliste lühenditega. Valke sünteesitakse raku tsütoplasmas
paiknevates ribosoomides. Kahe aminohappe reageerimisel moodustub
nende vahele
kovalentne side ehk peptiidside. Valgu molekulis on
peptiidsidemega ühendatud sadu või isegi tuhandeid aminohappejääke.
Enamus valke koosneb ühest peptiidsidemest. Valkudel on palju
erinevaid struktuure. Valgu aminohappelist järjetust nimetatakse
valgu esimeseks struktuuriks. Nt insuliinil on see Phe- Val- Asn-
Gly....jne.
3.
Aminohapete lühiiseloomustusAminohapped
(aminokarboksüülhapped)
on keemilised
ühendid,
mis sisaldavad funktsionaalsete
rühmadena
nii aminorühmi(NH2)
kui ka karboksüülrühmi(C+).
Aminohapped on karboksüülhapped,
mille alküülradikaalis
on üks või mitu vesiniku(H)
aatomit
asendunud aminorühmaga.
Kakskümmend
peamist (standartset) aminohapet moodustavad enamiku
elusorganismide valgud.
Valkude
lühiiseloomustusValgud
(proteiinid)- on polümeerid, mille monomeerideks on aminohapped. On
20 erinevat aminohapet (neist 8 asendamatud ja 12 , mida
rakud saavad
ise sünteesida), mis võivad
kuuluda valkude koostisesse.
Aminohappeid iseloomustavad amino- ja karboksüülrühmad. Valgu
molekulis aminohapete vahel on peptiidsidemed:
N-H
ja
karboksüülrühma vaheline kovalentne side. Peptiidsideme
moodustamisel eraldub üks
molekul vett.
Valkudes
on kolm osa: N-terminaalosa, peptiidsidet
moodustav osa ja C-terminaalosa.
Peptiidsidemete
süntees toimub alati kindlas suunas: N- terminus→C-terminus.
Valkude
omadused sõltuvad:
aminohapete järjestusest valgu molekulis
aminohapete arvust (DNA→valk→tunnus)
Oluline
on, et valgumolekul on lineaarne, ei hargne ega ei ole tsülklis.
Valke
jagatakse:
lihtvalgud-koosnevad aminohappejääkidest;
liitvalgud- koosnevad valgulisest ja mittevalgulisest osast.
Valkudel
on 4 struktuuri:
1) primaarstruktuur -
on kõikidel valkudel. Selle aminohapete järjestuse järgi on näidatud valkude omadused.
Aminohapped
on ühendatud peptiidsidemetega.
2)sekundaarstruktuur-
tekib aminohappeahela keerdumisel spiraaliks -α-heeliks-
või kõrvalahelate kokkuvoltimisel-
b
– struktuur.
Seda struktuuri hoiab koos vesiniksidemed (O ja H vahel). (kõõluste,
kõhrede, juuste, küünkarvade valgud, soomuste, ämblikuniidi
valgud )
3) tertsiaalstruktuur -
moodustub aminohappeahela edasisel kokkukeerdumisel. Seotud
vesiniksidemetega.Sellise struktuuriga valku nimetatakse gloobuliks.
( ensüümid , antikehad , vereplasma valgud)
4)kvaternaalstruktuur-
tekib mitme
gloobuli on ühinemisel. On ühendatud vesiniksidemetega.
( hemoglobiin )
Valkude
struktuur võib muutuda järgmiste protsesside tulemusena:
denaturatsioon - hävitatakse valgu kõrgemat järku struktuur, mille tulemusena aminohappeahel muutub sirgeks . See võib toimuda
Mehaanilisel
teel
Kõrge temperatuuriga
Keemilisel teel
Kiirguse toimel
renaturatsioon - valkude kõrgemat järku struktuuri taastamine.
Valgu
süntees toimub ribosoomides .
Valkude
ülesanned:
- ensümaatiline - ensüümid kiirendavad reaktsioone. Ensüümi tööks on vaja kindla vitamiini juuresolek.
- struktuurne - rakumembraanide ehitus, karvad , küüned, suled, kabjad, sarved , viiruste kapslid .
- transport- hemoglobiin transpordib hapnikku, membraanides valgulised transportijad.
- regulatoorne- hormoonid- bioaktiivsed ained, mis vere kaudu reguleerivad elundite tegevust. (nt.insuliin).
- retseptoorne- rakumembraani pinnaretseptorid annavad välissignaale edasi.
- liikumine- algloomade viburid , ripsmed, lihaskoe valgud ( aktiin , müosiin), mitoosi kääviniidid .
- kaitse- valgud on antikehade koostises; toodavad antikehasid. Antigeen - võõras aine organismis. Antikehad-seostuvad kindlate antikehadega. Ka verehüübimisvalgud, kattevalgud.
- energeetiline- väga madal -1g valkude lõhustumisel vabaneb 17,6 kJ energiat (nagu glükoosil). Valke lagundatakse ainult pärast sahhariide ja lipiide.
4.Nukleiinhapete
lühiiseloomustus
Nukleiinhapped- on polümeerid, mille monomeerideks on nukleotiidid (omavahel
seotud fosfordiester sidemetega).
Nukleiinhappeteks
on :
RNA ( ribonukleiinhape ) – geneetilise informatsiooni vahendaja, mis koosneb ribonukleotiididest.
DNA (desoksüribonukleiinhape)- geenetilise informatsiooni kandja, mis koosneb desoksüribonukleotiididest.
Nukleotiidid
koosnevad kolmest komponendist :
viiesüsinikuline suhkur e pentoos :
- RNA´s – riboos,
- DNA´s- desoksüriboos (teise süsiniku juures OH asemel on H)
lämmastikalus :
fosfaatrühm.
Biofunktsioonid
1.
kolme fosforhappe jäägiga nukleotiidid osalevad
energiasalvestamises (ATP ja GTP osalevad energia salvestamises,
neil on makroenergilised sidemed)
2. ühe fosforhappe jäägiga
nukleotiidid nt AMP ja GMP on nukleiinhapete ehitusüksusteks, mitmed
nukleotiidid on liitensüümides mittevalguliseks osaks (tavaliselt
kohaks, kus toimub reaktsioon ) osad nukleotiidid on antibiootilise
toimega (tapavad baktereid)
3. tsüklilise ehitusega nukleotiidid
nt cAMP on biosignaalide vahendajad (virgatsühendid ehk käskjalad)
4. disainitud ehitusega nukleotiidid on vähirakkude vastased ravimid (keemiaravi ehk kemoteraapia )
(Nukleotiidid
on
a) substraadiks nukleiinhapete sünteesil
b) energiakandjad
c)Tsüklilised nukleotiidid on signaalimolekulid ja regulaatorid
raku metabolismis ja reproduktsioonis)
DNA
ehitus:
DNA on lineaarne polümeer. Seda moodustavate nukleotiidide vahel on fosfordiester side. See side moodustub ühe nukleotiidi 3. süsiniku juures oleva hüdroksüülrühma ja teise nukleotiidi 5. süsiniku juures oleva fosfaatrühma vahel. Nukleiinhapete sünteesil on kindel suund: 5´ (prim) ots + 3´ (prim) ots.
Kaksikahelaline, nn biheeliks . Ahelad on antiparalleelsed: üks ahel:-5´ ots, teine-3´ots.
Nukleotiidide vahel on vesiniksidemed: A jaT vahel 2 ja G ja C vahel 3 vesiniksidet.Leiab aset komplementaarsusprintsiip: A=T, G=C.
DNA´l on kolm struktuuri:
- DNA esmane struktuur - nukleotiidijääkide hulk ja järjestus DNA üksikahelas. Üksikahelaline DNA esineb rakus sünteesiprotsessides ja teatud viirustes .
- 2) DNA sekundaarstruktuur - DNA levinuim esinemisvorm. (biheeliks ja kaksikspiraal)
- 3) DNA tertsiaalstruktuur - tekib DNA ja valkude koosmõjul. DNA + valgud = kromosoomid .
RNA
ehitus:
RNA esmane struktuur - primaarstruktuur. Nukleotiidijääkide hulk ja järjestus RNAs. Tekib sünteesijärgselt.
Teisene struktuur. Molekul , milles üksikahelalised lõigud vahelduvad kaksikahelaliste lõikudega. Omavahel paarduvad (tRNA)
RNA
on ebastabiilne, et seda saaks vajaduse korral lagundada. See
võimaldab rakul kasutada uusi RNA järgi sünteesitud valke. Ilma
selleta ei oleks rakk võimeline kiiresti reageerima väliskeskkonnast
tulenevatele signaalidele.
5.Millised
on peamised erinevused DNA ja RNA vahel:
Peamisteks
erinevusteks on 3, 1 ja 4
Tunnus
DNA
RNA
1) monomeer
desoksüribonukleotiid
ribonukleotiid
2)pentoos
desoksüriboos
riboos
3)N-alused
A=T, G=C
A=U, G=C
4)struktuur
biheeliks , so kaks ahelat , mis on kruvikujuliselt keerdunud
üks ahel
5) klassid
tRNA, rRNA, mRNA jt
6)ülesanne
päriliku informatsiooni säilitamine ja edasiandmine
pärilikkuse realiseerimine
7)leidmine
tuumas, mitokondrites, kloroplastides
tuumas, mitokondrites, kloroplastides, ribosoomis, tsütoplasmas
6.
Kolm põhilist RNA-de klassi rakkudes, nende funktsioonid
On
kolm põhilist RNA-de klassi:
informatsiooni RNA (mRNA)- sünteesitakse rakutuumas DNA ühe ahela järgi. See toob geneeetilise info rakutuumas asuvatest kromosoomides tsütoplasmas olevatesse ribosoomidesse.
transport RNA (tRNA) ülesandeks on mRNA molekuliga ribosoomidesse saabunud geneetilise info lahtimõtestamine. Vastavalt sellele toovad tRNA molekulid kohale ”õiged” aminohapped ja lülitavad need sünteesitava valgu ahelasse. Selle lülitamise koha tunneb ära tRNA antikoodon . Iga tRNA suudab siduda ainult üht kindlat aminohappet.
ribosoomi RNA (rRNA)- kuulub ribosoomi koostisesse ja sünteesib peptiidsidemeid aminohapete vahel.
Kujundlikult öeldes mRNA ”ütleb, kuidas valku teha”,tRNA toob selleks ”ehituskive” ning rRNA on ” tootmishoone ” üheks moodustajaks.
7.
Prokarüootsete ja eukarüootsete rakkude peamised erinevused
Prokarüootsed e eeltumsed rakud
Eukarüootsed e päristuumsed rakud
1)tüübid
bakterid
taimede, loomade, seente rakud ja protistid
2)tuum
puudub, selle asemel on tuumapiirkond
on kahemembraaniga ümbritsetud tuum
3)tuumamebraan
puudub
on olemas
4)DNA
DNA hulk on väiksem, on üks rõngaskromosoom
DNA´d on rohkem, on lineaarsed (alguse ja lõpuga) kromosoomid
5)sisemembraanistik(tsütopalsma-võrgustik, organellid )
puudub
on arenenud
6)tsütoplasma
jäik ja liikumatu
vedelam ja liikuv
8. Rakumembraan ja rakutuum , lisaks rakutuuma membraan
vaata
konspekti
http://et.wikipedia.org/wiki/Rakumembraan
http://et.wikipedia.org/wiki/Rakutuu m
9.
Membraansete organellide ehitus ja funktsioon
Endoplasmaatilise
retiikulumi (e tsütoplasmavõrgustiku) lühiiseloomustus
Raku
tsütoplasmat läbib endoplasmaatiline retiikulum e tsütoplasmavõrgustik. Eristatakse siledapinnalist ja
karedapinnalist tsütolasmavõrstikku.
siledapinnaline tsütoplasmavõrgustik e sile endoplasmaatiline retiikulum (sER) on membraanse ehitusega kanalikeste ja tsisternite süsteem. sER ülesanneks on lipiidide ja süsivesikute süntees ning ainete transport.
karedapinnaline tsütoplasmavõrgustik e karedapinnaline endoplasmaatiline retiikulum (rER) on membraanse ehitusega kanalikeste ja tsisternite süsteem. rER pinnal paiknevad ribosoomid , mis sünteesivad valku. Peale seda rER ülesannek on ainete transport raku sees.
Lüsosoomide
funktsioon
Lüsosoomid
on membraaniga ümbritsetud põiekesed, mis sisaldavad ensüüme.
Ensüümid lõhustavad rakkudesse transporditud toitaineid,
jääkprodukte ja surnud organelle.
Ühed
lüsoomid sisaldavad üksnes ensüümvalke, teised laguntatavaid
aineid ja neid lõhustavaid ensüüme.
Golgi
kompleksi funktsioon
Golgi kompleks on lamedate kohakuti paiknevate membraansete tsisternikeste
ja põiekeste süsteem.
Golgi
kompleksi ülesanneks on valkude kõrgemat järki struktuuride
kujundamine ja pakkkimine sekreedipõiekestesse ja lüsosoomidesse
ning ainete pakendamine. Golgi kompleks osaleb ka rakumembraani
uuendamises ja taimerakkudes ka rakukesta moodustamises.
10.
Mitokondrite
ja kloroplastide ehitus ja funktsioon
Kloroplastide
ehitus ja funktsioon
Kloropalstid-
on plastiidide liik, mis sisaldavad rohelist pigmenti klorofülli,
mis on oluline fotosünteesiprotsessis. Kloroplastid paiknevad
peamiselt lehtede rakkudes.
Kloroplast on ümbritsetud kahe membraaniga. Kloroplasti sisemuses paiknevad
membraanidest moodustunud kotjad moodustised- lamellid. Üksteisega
kohakuti painknevad lamellid moodustavad lamellide kogumikke.
Lamellide membraanides on klorofülli molekulid. Lisaks sellele on
kloroplasti sisemuses DNA, RNA ja valgu molekule. DNA omab pärilikku
infot kloroplastile omaste RNA ja valkude sünteesiks. Kloroplast
sisaldab ka ribosoome, mis sünteesivad sellele organellile vajalikke
valke. Kloroplastides toimub fotosüntees - suhkrute moodustamine
süsihappegaasist ja veest valgusenergia abil.
Mitokondrite
funktsioon
Mitokonder
on rakuorganell, mis on ümbritsetud kahe
membraaniga.
Sisemembraan moodustab arvukaid kurde ja sopistusi, mida nimetatakse
harjakesteks. Mitokondri vedelat sisekeskkonda nimetatakse
maatriksiks. Seal leidub mitokondrile omaseid DNA ja RNA molekule.
Mitukondri DNA sisaldab geneetilist infot organellille vajalike RNA
ja rakkude sünteesiks. Valke sünteesitakse mitokondri sees olevates ribosoomides.
Mitukondri
ülesanneks on rakuhingamine , energiarikaste ühendite(ATP) süntees,
raku varustamine energiaga.
11.
Tsütoskeleti funktsioonid
Tsütoskeleti
funktsioonid
Tsütoskelett-
on raku tsütoplasmas asuv valguliste niidikeste ja kanalikeste
võrgustik. Tsütoskelett on raku tugi- ja liikumissüsteem.
Tsütoskeleti
ülesandeks on organellide liigutamine või teatud asendis hoidmine,
see on raku toes, mis annab kuju ja vormi.
12.
Kromosoomide struktuur
Kromosoomide
struktuur
Kromosoom - DNA ja valgu molekulide kompleks (nukleoproteiin), milles sisalduvad geenid määravad pärilikke tunnuseid. Kromosoomid koosnevad DNA´st
ja sellele kinnitunud valgumolekulidest. Valgu molekule nimetatakse
histoonideks. Kromosoomides asuvad geenid.
Kromosoomis
asuvad lineaarses järgnevuses ja kindla paiknevusega geenid
ning mitmesugused mittegeensed nukleotiidijärjestused
( lookused ).
Kromosoom sisaldab ühe DNA
molekuli
massivõrdse koguse aluselisi valke
– histoone,
varieeruvas hulgas mittehistoonseid (happelisi) valke
ja vähesel hulgal RNAd .
Seda kromosoomi koostisainete kompleksi nim. kromatiiniks.
http://et.wikipedia.org/wiki/Kromosoo m
13. Bakteriraku ehitus
Kuna
bakteritel puudub tuum on nad eeltuumsed e.prokarüoodid. Tuuma
asemel on neil tuumapiirkond e. nukleoid . Bakteritel on ainult üks
rõngakujuline kromosoom. Bakterirakk on kaetud kapsliga, mis kaitseb
neid keskonna mõjude eest. Veel esineb bakterirakus DNA
rõngasmolekul e. plasmiid . Bakterid paljunevad pooldudes.Vees
elavatel bakteritel on gaasivakuoolid.
Plasmiid
on väike rõngas DNA. Selles on geenid, mis aitavad bakteril elada
ekstreemsetes oludes. Ilma plasmiidita on bakter
elujõuetu.Antibiootikumide suhtes võivad bakterid muututa resistentseks. Plasmiide võib olla mitu bakteril.
Bakterirakk
kooseneb:
Rakuseinast
Bakteri
väline kuju oleneb rakuseinast, mis kaitseb teda kahjulike
välismõjude eest ja kindlustab bakterile suhteliselt püsiva kuju.
Plasmamembraanist
Plasmamembraan
paikneb tihedalt vastu tsütoplasmat ja rakuseina ning muutub hästi
nähtavaks plasmolüüsi korral.
Plasmamembraan
koosneb kahest tihedast valgukihist, mille vahel on lipiidide kiht.
Ta koosneb põhiliselt fosfolipiididest, mis on ensüümsed valgud.
Mitokondritest
Mitokondrid on seenekujulised struktuurid . Nad paiknevad plasmamembraanil ning
võtavad osa ATP ( adenosiintrifosfaat ) sünteesist.
Golgi kompleksi analoogitest
Nad
teostavad raku ainevahetust, lõpp-produktide ja glükoproteiidide
väljutamist. Nende kanalite kaudu juhitakse väliskeskkonda näiteks
eksotoksiinid.
Basaalgraanulitest
Basaalgraanulid
kujutavad endast isetaastuvat (iseprodutseeritavat) bakteri osakest
(organelli). Basaalgraanulitele kinnituvad viburid. Struktuurilt kujutavad agranulaarsed võrgud tsisterne, kanaleid , põiekesi,
milledel ei ole spetsiifilisi alaosakesi. Nad vastutavad süsivesikute
ja lipiidide sünteesi eest. Arvatakse, et granulaarne
tsütoplasmaatiline võrgustik esineb kanali kujul, millele
kinnituvad ribosoomid ning milles toimub valgu süntees.
Mesosoomidest
Mesosoomid
tulevad nähtavale paljunevatel bakteritel vaheseina vööndis.
Nad
võivad olla tsisterni-, kanali-, või põiekesekujulised. Nad
vastutavad süsivesikute lipiidide jt. rakuseina koostisosade
sünteesi eest kui ka raku kasvu ja paljunemise eest. Jagunemise
lõppemisel need struktuurid eemaldatakse bakterist ja tekivad uuesti
rakus enne tuumaaine ja bakteri uut jagunemist.
Tsütoplasmast
See
on bakteri osa ilma rakuseina ja plasmamembraanita. Ta on
kolloidsüsteem. Tsütoplasmas eristatakse ribosoome, mitmesuguseid
erinevate funktsioonidega teralisi moodustisi, vakuoole, nukleoide.
Ribosoomid
vastutavad bakteri rakus valgu sünteesi eest.
Nukleoid
Geneetiline
materjal paikneb bakteritel DNA molekulis, mis esineb tsütoplasmas
enam-vähem kompaktse moodustisena . Nukleoid ei ole piiritletud tuumamembraaniga ning seda nimetatakse ka genofooriks. Ta sisaldab
DNA niiti. See niit kujutab endast suletud ringi ja seda nimetatakse
bakteriaalseks kromosoomiks .
Viburitest
Viburid
on bakteri liikumisorganiteks. Nad on niidikujulised, valgulise
koostisega ning kinnituvad tsütoplasmas paiknevale basaalgraanulile.
Arvatakse, et basaalgraanul on viburite ATP-aasi energiaallikaks.
Viburite
kuju võib olla silinderjas või lindikujuline
Viburid
paiknevad otstes või ümber rakukeha. Neid võib olla üksikult või
arvukalt.
Narmastest
Narmad
ehk karvad on torujad moodustised, mis esinevad paljudel bakteritel.
Narmad ei põhjusta raku liikumist. Narmaste abil kleepub mikroorganism seente, taimede, loomsete rakkude külge, seedekulgla hingamisteede jne epiteelile.
Kapslist
Kapsel on limaaine, mida produtseerib ja kihistab rakk oma pinnale.
Kapsel
koosneb peamiselt lämmastikku sisaldavatest polüsahhariididest.
Kapsel
kaitseb bakterit kuivamise ja teiste kahjulike välismõjude eest.
Eosed
Osa
baktereid moodustavad pärast vegetatsioonitsüklit eoseid. Et eosed
paiknevad raku sees siis nim. neid endospoorideks. Eose kuju on kas ümar, ovaalne või veidi välja venitatud, peaaegu
kepikujuline.
Eos
võib batsillis paikneda ekvatoriaalselt raku keskosas, terminaalselt
– raku otstes, subterminaalselt – raku keskosa ja otste vahel.
14.
Eukarüootide riigid ja nende peamised tunnused
Eukarüootide
riigid ja nende peamised tunnused
- autotroofsed organismid
-olemas
plastiidid (koroplastid)
- rakukest koosneb tselulloosist
-
rakukest puudub
-
heterotroofse toitumisega organismid
- hulkraksed päristuumsed organismid
- tavaliselt üherakkulised organismid (puudub liitsruktuur)
- võivad olla nii heterotroofsete kui ka autotroofsete organismidena
- mõnedel protistidel puuduvad mitokondrid
- tavaliselt elavad vees, niisketes paikades ja loomadekehas
- erinevad kehavormid
-rakukest
koosneb kitiinist
- heterotroofsed organiismid
-
osmotroofiline toitumine
15.
Loomaraku
ehitus. Põhilised loomsed koed .
Sarapuu
õpik Raku ehitus ja talitus
lk
3(49) all nurgas lühidalt kirjas(see käib siis „põhilised
loomsed koed“ kohta)
lk
7- 11 (53- 63)
16.
Seeneraku ehitus
Seeneraku
tsütoplasmas on samad organellid, mis on loomarakuehituses (tuum,
ribosoomid, mitokondriid,
lüsosoom,
Golgi kompleks, tsütoplasma,
tsütoplasmavõrgustik).
Kuna seened on heterotroofse ehitusega, siis puuduvad neil
taimerakule omased plastiidid ja vakuoolid . Üherakulised pärmseened
on ümarad, aga hulkraksete seente hüüfe moodustavad rakud on pikad
ja silindrikujulised. Rakkude otstes on avad, mille kaudu liiguvad
tsütoplasma, organellid ja rakutuum teise rakku. Mõnel seeneliigi
rakul need avad puuduvad, ja siis koosneb seeneniit ühest
hulktuumsest rakust.
Seeneraku
ehitus:
Seenerakk
on ümbritsetud membraaniga (sarnaneb looma ja taimeraku omaga ).
Membraanist
väljapoole jääb rakukest koosneb kitiinist. Rakukest kaitseb,
toestab rakku ja annab talle kindla kuju. Kuna enamik seeni toitub
kogu keha pinnaga, siis liiguvad vesi ja selles lahustunud ained läbi
rakukesta ja membraani tsütoplasmasse osmoosi teel. Seeneraku
keskosas asub kahe membraaniga ümbritsetud rakutuum.Tsütoplasmas
paiknevad mitokondrid varustavad rakku energiaga.
Tuum
Tuumake
Ribosoomid
Mitokoondrid
Lüsosoomod
Golgi kompleks
Tsütoplasmavõrgustik (kareda- ja siledapinnalist)
Tsütoplaasm
Tsütoskeleet
Rakukest (kitiinist)
Mõnedel on vakuool gaasiga
Rakumebraan
17.
Taimeraku ehitus:
http://www.zbi.ee/~tomkukk/anatoom/anato1.ht m
Taimeraku
ehitus.
Mitokondrid varustavad rakku energiaga, mida on vaja tema
elutegevuseks ja olemasolevate rakustruktuuride säilitamiseks.
Hapnikku tarbides muundavad nad süsivesikutes ja rasvades peituva
energia rakkudele kättesaadavaks . ON KA LOOMRAKUS.
Tsütoplasma
on raku sees. See sisaldab rohkesti vett ning selles on lahustunud
orgaanilisi ja anorgaanilisi aineid. ON KA LOOMARAKUS.
Rakumembraan
eraldab rakku teistest rakkudest kui ka ümbritsevast keskkonnast.
Vakuool on õhukese membraaniga ümbritsetud vee ja selles
lahustunud ainete mahuti.
Kloroplastid on plastiidid, milles
toimub fotosüntees. Neis valmistatakse orgaanilisi aineid, kasutades
päikese energiat.
Rakukest (koosneb tselulloosist) annab
taimerakule tugevuse ja kuju. Läbi rakukesta ja rakumembraani
pooride toimub aine- ja energiavahetus.
Tuum
Tuumake
Ribosoomid
Mitokoondrid
Lüsosoomid
Golgi kompleks
Tsütoplasmavõrgustik (kareda- ja siledapinnalist (kareda-pinnaline ER))
Tsütoplaasm
Tsütoskeleet
Rakukest (tselluloosist, ligniinist, pektiinist.)
Vakuool
Platiidid (Leukoplastod, hloroplastis ja hromoplastid)
Rakumembraan
18.
Rakukesta ehitus ja funktsioon
Sarapuu
õpik Rakuehitus ja talitus
lk
18-20(64-66)
19.
Geenitehnoloogia mudelorganismid
* soolekepike
*pärm
*poolduv
pärm
*ümaruss
*äädikakärbes
*sebrakala
* koduhiir
*rändrott
*müürlook
*riis
täpsemalt
on powerpointis igast liigist juttu
Selle
leiate loengud 2011 alt ja see on pealkirjaga Geenitehnoloogia
I 2011 -Rakk - rakutüübid-koed-ECM ja MUDELORGANISMID
20.
DNA pakkimine, kromosoomide ehitus
Sarapuu
õpik Rakuehitus ja talitus
lk
8-9(54-55)
21. Replikatsioon
Replikatsioon- matriitssüntees, mille tulemusena saadakse ühest DNA molekulist kaks ühesuguse nukleotiidse järjestusega DNA molekuli.
Päristuumsetel rakkudel toimub see enne mitoosi ja meioosi.
Matriitssüntees- st, et DNA , RNA ja valgud sünteesitakse
olemasolevate molekulide (DNA või RNA) ahelate alusel, mis määravad
sünteesitavate molekulide monomeeride järjestuse. Sel teel
tagatakse geeneetilise info ülekanne.
http://et.wikipedia.org/wiki/DNA_replikatsioon
22.
Geen ja genoom
Geen-
DNA lõik, mis määrab ära ühe RNA molekuli sünteesi.
Genoom-
liigiomases ühekordese kromosoomikomplektis sisalduv geneetiline
materjal. Inimese genoom koosneb 24 kromosoomist.
23. Transkriptsioon
http://et.wikipedia.org/wiki/Transkriptsioon_(geneetika)
Transkriptsioon-
matriitssüntees, mille käigus saadakse DNA molekuli ühe ahela
nukleotiidse järjestusega komplementaarne RNA molekul.
Transkriptsioonil saadakse mRNA, tRNA ja rRNA molekulid.
Kui
geenilt toimub transkriptsioon, nim seda geeni avaldumiseks.
Avaldumise järgi jaotatakse geenid 4 gruppi: 1)Samaaegselt kõigis
rakkudes avalduvad geenid- rRNA, tRNA. 2)Kindla koe rakkudes
avalduvad geenid- insuliini geen kõhunäärmerakk udes. 3)Rakkude
mingil kindlal elutegevuse etapil avalduvad geenid, näiteks: loote
elundkondade väljaarenemine. 4)Geenid, mis ei avaldu mitte kunagi evolutsioonis , kaotanud oma tähtsuse. Geenide aktiivsust
reguleerivad : struktuurgeenid , mis määravad raku ehituses ja
ainevahetuses osalevate valkude, tRNA ja rRNA sünteesi, ning regulaatorgeenid kontrollivad struktuurgeenide avaldumist .
TRANSKRPITSIOON
I.
Koht:
Eeltuumsed- tsütoplasmas
Päristuumsed tuumas,
mitokondrites, kloroplastides .
II. Aeg:
Eeltuumsetel - kogu
raku eluea jooksul
Päristuumsetel enamuse rakutsükli ajast va mitoos ja meioos (siis ei toimu)
III. Eeldused:
1.
Üksikahelaline DNA lõik
2.
Ensüümid: RNA polümeraas
3. Energeetilised faktorid ATP
4.
Nukleotiidid RNA koostises
5. Protsessi kontrollivad ja
reguleerivad valgud
IV: Olemus:
Kopeerimistüüp ehk
matriitssüntees
V:
Komplementaars us
VI: Tulemus:
Transkriptsioonil moodustuvad
kõik 3 tüüpi RNA molekulid
Tekivad RNA eelmolekulid, mis
vajavad täiendavat töötlust:
eelmolekulist kas eemaldatakse
teatud lõigud
jaguneb teatud fragmentideks
lisatakse teatud
järjestused
muudetakse keemilist koostist (N-aluseid)
24.
mRNA struktuur prokarüootidel ja eukarüootidel.
POWERPOINT:
L3 DNA-valk
http://en.wikipedia.org/wiki/Messenger_RNA#Prokaryotic_mRNA_degradation
Translatsioon toimub ribosoomidel.
RNA molekuli, millelt toimub translatsioon,
nimetatakse mRNA-ks (inglise
keelest messenger
RNA).
Prokarüootsetes rakkudes on primaarne transkript üldjuhul ka
koheselt transleeritav. Eukarüootses rakus toimub aga primaarse transkripti,pre-RNA,
protsessimine transleeritavaks mRNA molekuliks. Enamus eukarüootseid
geene rakutuumas sisaldavad endis mittekodeerivaid alasid
– introneid,
mis vahelduvad kodeerivate piirkondadega – eksonitega.
Intronjärjestusi sisaldavatelt geenidelt sünteesitakse
transkriptsiooni käigus pre-mRNA, millest seejärel kõrvaldatakse
mittekodeerivad järjestused splaissingu (splicing)
teel. Slaissingureaktsiooni läbiviimiseks moodustub
makromolekulaarne struktuur, mida nimetatakse splaissosoomiks.
Enne splaissimist lisatakse pre-mRNA-le 5´-otsa 7-metüülguanosiin
ja 3´- otsa polü-A järjestus – polüA
saba,
mis on 20-200 nukleotiidi pikk. Kõik need protsessid toimuvad
rakutuumas. Protsessitud mRNA transporditakse tsütoplasmasse ning
alles seal toimub translatsioon. Erinevalt eukarüootidest, kus
transkriptsioon ja translatsioon on ajaliselt ja ruumiliselt
teineteisest lahutatud, toimuvad prokarüootsetes rakkudes mõlemad
protsessid korraga: parasjagu sünteesitavalt mRNA molekulilt algab
kohe ka translatsioon.
RNA
molekulide tüübid
Eukarüootses
rakus on põhiliselt nelja tüüpi RNA molekule: mRNA, tRNA, rRNA ja snRNA . Eelnevalt oli juttu mRNA molekulidest, mis kodeerivad valke.
Nende ülesandeks on vahendada DNA nukleotiidses järjestuses salvestatud geneetilist informatsiooni
translatsiooniaparaadile. tRNA molekulid
( transfer RNAs)
on väikesed RNA molekulid, mis toimivad adapteritena translatsioonil polüpeptiidahelasse lülitatavate aminohapete ja mRNA molekulis
asuvate aminohappeid määravate koodonite vahel. Ribosomaalsed RNA
molekulid (rRNA)
kuuluvad ribosoomide koostisesse. Väikesed tuuma RNA-d
(snRNA molekulid
– small
nuclear RNAs)
osalevad intronite splaissingul.
25.
mRNA protsessing
mRNA
processing
The
pre-mRNA molecule undergoes three main modifications. These modifications are 5'
capping,
3' polyadenylation,
and RNA
splicing,
which occur in the cell
nucleus before the RNA is translated .[2]
Main
article: 5'
cap
[edit]Capping
Capping
of the pre-mRNA involves the addition of 7-methylguanosine (m7G)
to the 5' end. To achieve this, the terminal 5' phosphate requires removal, which is done with the aid of aphosphatase enzyme .
The enzyme guanosyl
transferase then
catalyses the reaction, which produces the diphosphate 5'
end. The diphosphate 5' prime end then attacks the gamma phosphorus atom of a GTP molecule
in order to add the guanine residue
in a 5'5' triphosphate link . The enzyme
(guanine-N7-)-methyltransferase
("cap MTase") transfers a methyl group from S-adenosyl
methionine to
the guanine ring.[3] This
type of cap, with just the (m7G)
in position is called a cap 0 structure. The ribose of
the adjacent nucleotidemay
also be methylated to give a cap 1. Methylation of nucleotides
downstream of the RNA molecule produce cap 2, cap 3 structures and so
on. In these cases the methyl groups are added to the 2' OH groups of
the ribose sugar. The cap protects the 5' end of the primary RNA
transcript from attack by ribonucleases that
have specificity to the 3'5'phosphodiester
bonds.[4]
[edit]3'
Processing
Main
article: Polyadenylation
[edit]Cleavage
and Polyadenylation
The
pre-mRNA processing at the 3' end of the RNA molecule involves
cleavage of its 3' end and then the addition of about
250 adenine residues
to form a poly (A) tail .
The cleavage and adenylation reactions occur if a polyadenylation signal sequence (5'-
AAUAAA-3') is located near the 3' end of the pre-mRNA molecule, which
is followed by another sequence, which is usually (5'-CA-3').
The second signal is the site of cleavage. A GU- rich sequence is
also usually present further downstream on the pre-mRNA molecule.
After the synthesis of the sequence elements, two
multisubunit proteins called cleavage
and polyadenylation specificity factor (CPSF)
and cleavage
stimulation factor (CStF)
are transferred from RNA
Polymerase II to
the RNA molecule. The two factors bind to the sequence elements. A protein complex forms that contains additional cleavage factors and
the enzymePolyadenylate
Polymerase (PAP).
This complex cleaves the RNA between the polyadenylation sequence and
the GU-rich sequence at the cleavage site marked by the (5'-CA-3')
sequences. Poly(A) polymerase then adds about 200 adenine units to
the new 3' end of the RNA molecule using ATP as
a precursor. As the poly(A) tail is synthesised, it binds multiple
copies of poly(A) binding protein, which protects the 3'end from
ribonuclease digestion.[4]
[edit]Splicing
Main
article: RNA
splicing
RNA
splicing is the process by which introns,
regions of RNA that do not code for protein, are removed from the
pre-mRNA and the remaining exons connected
to re-form a single continuous molecule. Although most RNA splicing
occurs after the complete synthesis and end-capping of the pre-mRNA,
transcripts with many exons can be spliced
co-transcriptionally.[5] The
splicing reaction is catalyzed by a large protein complex called
the spliceosome assembled
from proteins and small
nuclear RNA molecules
that recognizesplice
sites in
the pre-mRNA sequence. Many pre- mRNAs , including those
encoding antibodies,
can be spliced in multiple ways to produce different mature mRNAs
that encode different protein
sequences.
This process is known as alternative splicing,
and allows production of a large variety of proteins from a limited amount of DNA.
26.
Geneetiline kood
http://et.wikipedia.org/wiki/Geneetiline_kood
Geneetiline
kood- on vastavus, kus mRNA kolm järjestikust nukleotiidi (st. koodon ) määravad ära ühe aminohappe paigutuse valgu molekulis.
27.
Translatsioon, tRNAde ja ribosoomide ehitus
Translatsioon
Translatsioon-
valgu süntees toimub
tsütoplasmas, ribosoomides.
Translatsioon-
protsess, mille käigus sünteesitakse liigiomaseid valke kõikide
talitluste läbiviimiseks organismis. Valgud on liigiomased, st
sünteesitud päriliku info alusel. Selle protsessi aluseks on mRNA
molekulid.
Translatsiooni
protsess toimub ribosoomides. mRNA molekulis on geneetiline kood, mis
määrab valgu molekuli aminohapete järjekorra. Geneetiline kood
seisneb kolmes järjestikulises nukleotiidis ehk ühes koodonis,
millele vastab kindel aminohape .
Geneetiline
kood on looduses universaalne, st kehtib kõikide elusorganismid e
valkude ülesehituses. Koodonite ja aminohapete seoseid kujutatakse
koodipäikese abil. Ühele aminohappele võib vastata kuni 6 erinevat koodonit , kuid üks koodon vastab alati ühele kindlale aminohappele.
Üks ja sama nukleotiid mRNA ahelas ei kuulu üheaegselt
kõrvutiasetsevatesse koodonitesse.
Esimeseks
koodoniks mRNA molekulis on alati initsiaatorkoodon A-U-G
(meteoniin). Viimaseks koodoniks on stopp - ehk nonsenskoodon (3 varianti ), millele ei vasta ühtegi aminohape
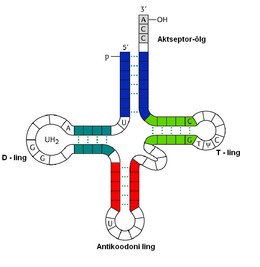
tRNAde
struktuur ja funktsioon.
tRNA
on ribonukleiinhape,
mis tegeleb rakus
aminohapete
transpordiga ribosoomi,
kus geneetilise
koodi
alusel lisatakse aminohape
sünteesitavasse valguahelasse.
tRNA
molekulide sekundaarstruktuuri
iseloomustatakse "ristikheinalehe" kujuga. tRNA
sekundaarstruktuuri
moodustavad 4 kaksikahelalist osa - õlga ja 4 üksikahelalist
piirkonda - lingu , mis paiknevad vastavate õlgade otstes. tRNA
molekuli otsad asuvad lähestikku, nende paardumisel tekkiv
kaksikahelaline osa kannab nimetust " aktseptor -õlg", selle
3'
otsa paardumata nukleotiididele liidetakse estersidemega
aminohape.
Aktseptor-õlg on 7 aluspaari
pikk. T-õlg on saanud oma nime tänu modifitseerunud
nukleotiididele,
mis asuvad T- lingus . Need nukleotidid on ribosüültümidiin
T
ja pseudouridiin
ψ.
T-õla pikkus on 5 aluspaari,
T-lingu pikkus varieerub harilikult 7-9 nukleotiidi
ulatuses. D-õlg on saanud nime temas leiduva modifitseeritud
nukleotiidi dihüdrouridiini
D
järgi. D-õla pikkus on harilikult 4 aluspaari,
D-lingu pikkus on varieeruv . Antikoodon-õlg on alati 5 aluspaari
pikk ja nagu nimestki on näha, sisaldab antikoodoni ling kolmest
nukleotiidist antikoodonit mis vastab mRNA
koodonile.
Antikoodoni lingus on alati 7 nukleotiidi.
Lisaks neile võivad paljud tRNA molekulid sisaldada lisalingu ja
lisaõlga, mille pikkus võib olla suuresti varieeruv.
Funktsionaalsus
tRNA
molekuli tasapinnaline struktuur
Funktsionaalselt
on tRNA molekuli tähtsaim osa antikoodon, mille kolm nukleotiidi
paarduvad komplementaarsusprintsiibi
alusel mRNA koodoniga,
mis on geneetilise translatsiooni
ja geneetilise
koodi strukturaalseks aluseks. Seetõttu on antikoodoni lingu
ruumiline konformatsioon oluline täpse koodon-antikoodon seondumise
toimumiseks. Antikoodoni ees paikneb konserveerunud
U33 nukleotiid,
mille järel teeb tRNA ahel järsu pöörde. Nukleotiid
34, esimene antikoodoni nukleotiid
paardub mRNA koodoni
viimase 3. nukleotiidiga
ja seetõttu on oluline, et koodon-antikoodon seondumine lõppeks
just 34. nukleotiidiga ega jätkuks U33 ga, niiviisi määrab tRNA
ahelas toimuv järsk pööre ära koodoni pikkuse (3 aluspaari).
Antikoodon on sobivas ruumilises struktuuris mRNA
koodoniga
paardumiseks. Antikoodoni järel paiknevad nn. hüpermodifitseeritud
nukleotiidid, mis pole võimelised aluspaare moodustama, nii on
tagatud translatsiooni
täpsus.
tRNA
teine oluline piirkond, 3'
ots, paikneb aktseptor-õla otsas. tRNA kolm viimast nukleotiidi
on CCA järjestus, mis ribosoomis seondub valguahela peptiidsideme
moodustumist katalüüsiva
piirkonnaga. tRNA 3' otsa seotakse ka ensüüm
aminoatsüül-tRNA ligaasi vahendusel estersidemega
vastav aminohape.
Olgugi, et paljudele aminohapetele
vastab mitu erinevat tRNAd, vastab igale erinevale tRNAle vaid üks
konkreetne aminoatsüül-tRNA ligaas . Aminoatsüül-tRNA
ligaas tunneb tRNA ära vastavate tRNA identsuse elementide
järgi, milleks on kindlad, erinevatele tRNA molekulidele ainuomased
nukleotiidid
antikoodoni lingus ja aktseptoorses õlas. Valgusünteesi käigus
ribosoomis
lahutatakse tRNA antikoodoni ja mRNA koodoni paardumisel aminohape
tRNAst ja lisatakse valguahelasse.
Ribosoomide
ehitus ja funktsioon
http://et.wikipedia.org/wiki/Rakutuu m
Ribosoomide
ülesanneks on valkude süntees.
Ribosoomid
moodustuvad tuumakestes. Sünteesijärgselt liiguvad nad mööda
tuumamembraanide pooride tsütoplasmasse. Seal kinnitub osa neist
tsütoplasmavõrgustikule. Ribosoome võib leida ka mitokondrites ja
kloroplastides.
28.
Valgusünteesi regulatsioon
- Toimub ribosoomis.
- Osalevad mRNA, tRNA ja rRNA.
- mRNA seostub ribosoomiga.
- Translatsioon algab alguskoodoniga AUG.
- Komplementaarse antikoodoniga tRNA toob esimese (kindla) aminohappe ribosoomi. Teine tRNA mahub ka koos oma aminohappega ribosoomi.
- Kahe aminohappe vahele tekib tugev peptiidside
Translatsioon
lõpeb stoppkoodonitega
http://et.wikipedia.org/wiki/Valgus%C3%BCntees
29.
Ribosüümid ja RNA maailm
Ribosüüm on ribonukleiinhape,
millel on katalüütilised omadused.
Ribosüümid on ensüümid,
mis aga ei koosne polüpeptiididest,
nagu enamik ensüüme, vaid polünukleotiididest.Ribosüüm-
ensüüm, mis ei ole valguline, aga koosneb RNA ahelatest. Seda
peetakse ensüümiks, sest ribosoomi koosseisu kuuluv rRNA sünteesib
valgusünteesi käigus peptiididemeid aminohapete vahel.Arvatakse
et RNA oli DNA eellane. Arvatakse, et olles üheahelaline ja
võimeline moodustama mitmesuguseid tertsiaarstruktuure, võis kunagi
olla ensümaatilise aktiivsusega ja katalüüsida ise enda
replikatsiooni. Esmase info kandjaks olid RNA molekulid. 1982 a
leiti, et on olemas RNA, millel on katalüüsi võime, mida nimetati
ribosüümiks.
30. Geeniekspressioon , selle regulatsioon
Rakkude diferentseerumine on üldjuhul hoopis geenide valikulise
ekspressiooni tulemus. Hulkrakses organismis esinev rakutüüpide
mitmekesisus on põhjustatud sellest, et rakkude samasuguselt DNA-lt
sünteesitakse erinevates rakkudes erinevaid RNA ja valgu molekule.
Informatsioon
kandub DNA-lt valguni mitme etapina , kõiki neid etappe on võimalik
reguleerida. Rakk võib oma aktiivsete valkude tootmist kontrollida
järgmistel viisidel:
*
kontrollides, kui sageli ja millal transkripeeritakse vastavat geeni
(kontroll transkriptsiooni tasemel)
*
kontrollides, kuidas toimub primaarse transkripti splaising või mõni
muu modifikatsioon (Kontroll RNA protsessingu tasemel)
*
kontrollides, milliseid tuumas toodetud mRNA molekule viiakse
tsütoplasmasse, kus toimub valkude süntees (kontroll RNA transpordi
tasemel)
*
kontrollides, milliste tsütoplasmas leiduvate mRNA molekulide
kaasabil toodetakse ribosoomides valke (kontroll translatsiooni
tasemel)
*
selektiivselt lagundades mõnesid mRNA molekule tsütoplasmas
(kontroll mRNA degradatsiooni tasemel)
*
selektiivselt aktiveerides või inaktiveerides toodetud valke või
transportides neid erinevatesse raku osadesse (posttranslatsiooniline
kontroll)
Geeni
ekspressiooni reguleerimine annab rakule kontrolli struktuuri ja
funktsiooni üle, on raku diferentseerumise, morfogeneesi,
paindlikkuse ja kohastumise aluseks.
31.Geeniekspressiooni
regulatsioon, sarnasused ja erinevused prokarüootidel ja
eukarüootidel.
http://et.wikipedia.org/wiki/Geeniekspressioon
Eukarüootidel:
- Regulatoorsed valgud on transkriptsioonifaktorid (TF).
- TF seostudes teiste geenide cis elementidega reguleerivad nende geenide ekspressiooni: “trans- acting ” faktorid.
Ühisjooned:
- Polütsistroonsed geenid, st mitmed sarnase funktsiooniga geenid on paiknevad koos ja reguleeritakse ühiselt --- operon.
- Geeni regulatsioon on peamiselt negatiivne, vahendatud repressorvalkude kaudu. Alles induktori seostumine repressoriga inaktiveerib viimase ja võimaldab ekspressiooni.
POWERPOINT:
regulatsioon
32.
Lac
ja Trp operonid, nende reguleerimise sarnasused ja erinevused
POWERPOINT:
regulatsioon
33.
Cis-elemendid, trans-faktorid
POWERPOINT:
regulatsioon
34.
DNA-valk
interaktsioonid
- regulatsioon toimib läbi paljude cis-acting elementide ja trans-acting faktorite vaheliste seoste.
- mitte-kovalentsed
POWERPOINT:
regulatsioon
35.
Rekombinantse DNA metoodika alused
POWERPOINT:
rekombinantne DNA
36.
Restriktaasid
Restriktaasid-
restriktaasid on ensüümid, mia lõikavad DNA katki. On olemas
järjestusspetsiifilised restriktaasid ja restriktaasid, mis lõikavad
DNA ahelat suvalistest kohtadest .
Restriktaasidel
on
omadus lõikata DNA topeltahel läbi kindlas piirkonnas
(lõikepiirkond- ingl. k. cleavage
site),
mille määrab ära antud piirkonna DNA nukleiinhappeline-järjestus
(äratundmis-järjestused;
ingl. k. recognition
sequences-
koosnevad 4-8 nukleotiidipaarist), kusjuures iga ensüümi jaoks on
see erinev.
Kasutades
erinevaid restriktaase, võime saada DNA fragmente, millel on kas
tömbid (Hpa I) või siduvad otsad (ingl. k. cohesive
ends). Viimased kujutavad endast lühikesi ühekordse ahela juppe. Siduvate
otsadega fragmente võib omavahel taas liita. Seega vôib
teoreetiliselt mistahes geene omavahel liita.
Restriktaasid
on ensüümid, mida toodavad bakterid enesekaitseks – need lõikavad
DNA lõikudeks, aga nii, et tekivad üheahelalised otsad –
“kleepuvad otsad”.
Selliste
otstega DNA juppe on komplementaarsuse tõttu võimalik mugavalt liita.
Erinevate
DNA-de liitmisel saame rekombinantse
DNA.
37.
DNA kloneerimise etapid
DNA kloneerimine - ühesuguste plasmiidide koopiate tegemine bakteri
paljunemise tulemusena.
Geeni
paljundamise põhietapid plasmiide abiga on järgmised:
1)
plasmiidi isoleerimine bakterirakust (tavaliselt kasutatakse E. coli
plasmiide);
2)
plasmiidi "lõikamine" spetsiifilise restriktaasiga;
3)
paljundatava geeni vôi DNA-lôigu "väljalõikamine"
kromosoomist sama restriktaasiga- s.o. geeni
isoleerimine;
4)
isoleeritud geeni "istutamine" plasmiidi
5)
plasmiidi viimine bakterirakku ja bakteri kasvatamine , mille käigus
paljuneb ka vastav plasmiid.
6)
paljundatud geeni isoleerimine plasmiididest
38.
DNA
sekveneerimise põhimõte
DNA sekveneerimine - DNA nukleotiidse järjestuse kindlaks tegemine.
Ensümaatilise
meetodi puhul kasutatakse DNA-polümeraasi abil toimuva topeltahela
sünteesi blokeerimist kindla nukleotiidi kohal. Tulemuseks on
erineva pikkusega fragmendid , mille elektroforeesil joonistub välja
DNA molekuli NH järjestus.
39.
Polümeraasi ahelreaktsioon
PCR
viiakse
läbi biokeemilise reaktsioonina ja selle puhul ei vajata
elusorganisme DNA kopeerimiseks. Reaktsioon pôhineb ensüümi-
DNA-polümeraas kasutamisel ,
mis katalüüsib DNA komplementaarse ahela sünteesi. PCR on
DNA-molekuli paljundamine kunstlikes tingimustes.
Reaktsiooni
läbiviimiseks on vajalik teada uuritava DNA lõigu otste
nukleotiidset järjestust.
Reaktsiooni
käivitamiseks kasutatakse kahte oligonukleotiidset (väiksest arvust
nukleotiididest koosnevat (8..30) praimerit (ingl. k.
primer),
mis kumbki vastavad ühe komplementaarse DNA ahela alguse
nukleotiidsele järjestusele ja talitlevad kui ensüümi substraat ,
kuna neil on vabad otsad uute nukleotiidide sidumiseks.
PCR
pôhietapid on
järgmised:
topeltahelalise DNA denaturatsioon kaheks üksikahelaks kôrge temperatuuriga (90-95 °C; 40-60 sek);
praimerite hübridiseerimine e. "istutatamine" kummalegi üksikahelale, milleks temperatuur viiakse alla ca 50 °C juurde 30 sekundiks;
komplementaarse DNA ahela süntees DNA-polümeraasi toimel (72 °C juures aeg sôltub lôigu pikkusest, kuid ca 1-3 min). Ensüüm on termostabiilne ja on isoleeritud kuumavee allikates elavatest bakteritest (näit. Thermus aquaticus ja temalt saadud ensüümi nimetatakse TaqI).
Reaktsioonis
kasutatavad komponendid (maatriks DNA, praimerid, ensüüm ja vabad
nukkleotiidid- viiakse kohe algselt ühte katsutisse. Kuna
ahelreaktsiooni etapid toimuvad erineva temperatuuri juures, siis on
kogu protsess juhitav temperatuuri abil ja vastavaid tsükleid vôib korrata kümneid kordi . Igas tsüklis DNA hulk teoreetiliselt
kahekordistub. Praktiliselt hilisemates tsüklites on reaktsiooni
efektiivsus väiksem, kuna ensüümi aktiivsus langeb, samuti lôpevad
otsa vabad nukleotiidid. Neid juurde pole aga vôimalik lisada.
Saadud koopiate arv ulatub aga 30 tsükli järel juba miljonitesse,
mis on rohkem kui küll olenematra, mis eesmärgil reaktsioon läbi
viidi.
PCR produkt eraldatakse lahusest elektroforeesiga agargeelis.
PCR-i
saab kasutada ka diagnostilisel eesmärgil: teatud DNA vôi RNA
järjestuse (viiruste vôi bakterite) avastamiseks uuritavas materjalis vôi näiteks geenidefektide avastamiseks genoomis. Tema
tundlikkus on teoreetiliselt selline, et kui uuritavas materjalis on
üks DNA-molekul, mille järjestus ühtub materjalile lisatava
praimeriga, siis me selle ka avastame. PCR- on kôrvaletôrjumas DNA
sondide meetodit geneetiliste haiguste uurimisel ,
viirusinfektsioonide tuvastamisel ja mikroobide tüpiseerimisel.
40. Elektroforees
Elektroforees
on elektriliselt laetud osakeste liikumine vedelikus
elektrivälja
mõjul: positiivsed osakesed katoodile
ja negatiivsed osakesed anoodile.
Elektroforees
on meetod, kus laetud osakesed liiguvad elektrijuhtivust omavas
vedelas keskkonnas elektrivälja mõjul.
Geel -elektroforees
- DNA kaksikahel liigub geelis elektriväljas kiirusega, mis on
pöördvõrdeline log10–ga
tema massist ( suurusest ).
1. maatriks on agaroos või polüakrüülamiid
2. puhver tagab, et DNA on laetud negatiivselt
3.
DNA liigub suunas - + (katoodilt anoodile).
40.
Nukleiinhapete hübridiseerimine
NH
hübridiseerimine pôhineb denatureerunud DNA ja RNA renatureerumise
fenomenil, mis seisneb selles, et teatud tingimustel denatureeritud NH ahelad on vôimelised uuesti renatureeruma ja moodustama ka vabade
komplementaarsete NH ahelatega topeltahelaid.
See
on vôimaldanud luua kôrge tundlikkusega meetodid spetsiifiliste NH
järjestuste avastamiseks uuritavas materjalis.
Selleks
kasutatakse puhastatud vôi kloonitud NH ahelate fragmente, millel on
kindlaksmääratud NH järjestus ja mis on märgistatud kas keemilise
markeri vôi radioaktiivse isotoobiga ( signaal ). Selliselt töödeldud
DNA fragmente nimetatakse DNA
sondideks (ingl.
k. probes).
Sondide
abil on vôimalik määrata geenide lokalisatsiooni kromosoomides,
defektgeenide olemasolu, geenide talitluslikku aktiivsust määrates
nende poolt produtseeritava informatsiooni RNA hulka tsütoplasmas,
aga ka näiteks viirusliku RNA vôi DNA olemasolu ja lokalisatsiooni
kudedes ning rakkudes.
NH
hübridiseerimise eri juhuks on ka sellised meetodid nagu Southern blotting ja Northern blotting ("lõuna" ja "pôhja"
märgistamine).
E.M. Southern töötas välja meetodi DNA fragmentide kindlakstegemiseks agaroosgeelis.
Restrikataasi
abil lôhustatud kaksikahelalise DNA fragmentide elektroforeetilise lahutamise järel denatureeritakse DNA topeltahelad üksikahelateks NaOH abil. Denatureeritud ahelad kantakse üle nitrotselluloos kilele. Kile viiakse sondi sisaldavasse lahusesse. Sond hübridiseerub
vastava üksikahelise DNA fragmendiga ja tema asukoht
elektroforegrammil on vôimalik kindlaks määrata, kuna sond on
märgistatud.
Northern
blotting'iks nimetatakse RNA fragmentide määramist samal meetodil.
42. Kromatograafia .
Kromatograafia
on üldmõiste
mitmesuguste laboratoorsete füüsikalis-keemiliste meetodite kohta,
mida kasutatakse ainete segu komponentide lahutamiseks paljukordse
sorptsiooni ja
desorptsiooni
tingimustes.
Lihtsustatult:
praktikas kantakse ainete segu läbi sorbendi (liikumatu faas) sobiva
vedeliku või gaasi vooluga (liikuv faas). Segu komponentide
spetsiifilise sorptsiooni ja desorptsiooni tulemusena toimub nende
jaotumine liikumatu ja liikuva faasi vahel vastavalt
jaotuskoefitsientidele ning nende aktide paljude korduste tagajärjel
komponendid liiguvad edasi erinevate kiirustega. See viib ainete
lahutumisele ning moodustuvad kiiremini ja aeglasemalt liikunud
komponentide tsoonid . Protsess teostatakse kas kolonnis , kapillaaris, paberil või plaadil. Lahutunud komponendid detekteeritakse
füüsikaliste või keemiliste meetoditega.
43.Valkude
struktuuri uurimise meetodid
- Valgu molekulaarse struktuuri võib määrata tuumamagnetresonantsi abil
- Valgu funktsioonide üle võib otsustada tema järjestuse võrdlemisel mõne tuntud funktsiooniga valgu järjestusega
- Affiinsuskromatograafia ja immunosadestamine võimaldavad leida seostunud valke
Valgu
funktsiooni uurimisel võib kasutada liitvalke ( fusion proteins),
samuti võib nende abil valku rakus lokaliseerida.
44.Antikehade
ja liitvalkude kasutamisvõimalusi molekulaarbioloogias
Antikehade
kasutamine molekulaarbioloogias Flourentsmikroskoopia: Väga laialt
on levinud rakubioloogias nn. immuunfluorestsentsi meetod, mis
põhineb fluorestseeruvat värvainetega konjugeeritud antikehade
kasutamisele. Enamlevinud fluorokroomidena antikehade märgistamiseks
kasutatakse FITC (fluorestsiin-isotiotsüanaat), TRITC
(tetrametüül-rodamiin- isotiotsüanaat ) PE (fükoerütriin) jt.
Fluorestsentsmikros-koobis on valgusallikaks elavhõbedalamp, mis
annab lühilainelist kiirgust (paljudel
lainepikkustel).
Immuunfluorestsents: Elavhõbedalambiga
valgustatakse helendavate värvainetega märgistatud antikehasid.
-
45.
Viirused .
HIV
Viirused
on eluta ja elusa looduse piirimail olevad rakulise ehituseta ainult
elusrakkudes paljunevad bioloogilised objektid.
Viirus on rakuta moodustis, tema koostises on vähemalt:
- genoom ( nukleiinhape - DNA või RNA)- nikleiinhaped säilitavad pärilikku info. Viirusel peab olema vähemalt kolm geeni
- kapsiid (valgud)- kaitseb genoomi keskkonnamõjutuste eest ja aitab viiruse genoomi peremeesrakku.
Nendele
võib lisanduda ümbris,
mille viirus rakust väljudes kaasa võtab. St. viiruse ümbris
koosneb rakumembraani koostisosadest : lipiitidest ja valkudest.
Kapsiid ja ümbriis on genoomi kaitseks, aga ka viiruse rakku
tungimise tagamiseks ( taku äratundmiseks). Viiruse ümbrise pinnal
on valgus, mis käituvad signaalidena. Kui raku membraani pinnal
plevad valgud seonduvad viiruse valkudena, siis rakk arvab , et see on
mingi signaal ja viirus viiakse raku sisse..Siis viirus alustab raku
sees oma tegevust.
Viiruste
paljumine jagatatakse nelja etappiks:
1. Viirusosake kinnitub fibrillidega rakumembraanile;
2.
Viirusosake vabaneb ümbrisest ja lagundab rakumembraani;
3.
Viirusosakese nukleiinhape koos kapsiidiga siseneb rakku;
4.
Viirusosake vabaneb kapsiidist;
http://en.wikipedia.org/wiki/HIV



































![[edit]Cleavage and Polyadenylation](./mpic/161179/Backup-15_1.jpg)





Kõik kommentaarid