1) Tuum 2) Tuumake 3) Ribosoomid 4) Mitokoondrid 5) Lüsosoomid 6) Golgi kompleks 7) Tsütoplasmavõrgustik (kareda- ja siledapinnalist (kareda-pinnaline ER)) 8) Tsütoplaasm 9) Tsütoskeleet 10) Rakukest (tselluloosist, ligniinist, pektiinist.) 11) Vakuool 12) Platiidid (Leukoplastod, hloroplastis ja hromoplastid) 13) Rakumembraan 18. Rakukesta ehitus ja funktsioon Sarapuu õpik Rakuehitus ja talitus lk 18-20(64-66) 19. Geenitehnoloogia mudelorganismid *soolekepike *pärm *poolduv pärm *ümaruss *äädikakärbes *sebrakala *koduhiir *rändrott *müürlook *riis täpsemalt on powerpointis igast liigist juttu Selle leiate loengud 2011 alt ja see on pealkirjaga Geenitehnoloogia I 2011 -Rakk - rakutüübid-koed-ECM ja MUDELORGANISMID 20. DNA pakkimine, kromosoomide ehitus Sarapuu õpik Rakuehitus ja talitus lk 8-9(54-55) 21. Replikatsioon
UUED TEADUSLIKUD FAKTID HÜPOTEES TÕESTATAKSE või LÜKATAKSE ÜMBER (e HÜPOTEES PEAB OLEMA FALTSIFITEERITAV) ∨ PÄDEVA TEADUSLIKU TEOORIA ALUSEL ON VÕIMALIK ENNUSTADA NÄHTUSI/FAKTE, MILLE OLEMASOLU HILJEM EKSPERIMENTAALSELT TÕESTATAKSE 2. Elu organiseerituse tasemed - MOLEKULAARNE tase – molekulaarbioloogia, geenitehnoloogia, süsteemibioloogia (BIOMOLEKULID ainult ELUSlooduses). Esmane organiseerituse tase. Kõikjal, kus on elu, esinevad biomolekulid: sahhariidid, lipiidid, valgud, nukleiinhapped. - ORGANELLI tase – (molekulaarne) rakubioloogia. Uuritakse raku organelle: tuum, ribosoomid, mitokondrid jne. Kui need rakkudest eraldada, ei kanna nad enam elu tunnuseid. Organellide koostööst tulenevad rakkude omadused. - RAKU tase – rakubioloogia
UUED TEADUSLIKUD FAKTID HÜPOTEES TÕESTATAKSE või LÜKATAKSE ÜMBER (e HÜPOTEES PEAB OLEMA FALTSIFITEERITAV) PÄDEVA TEADUSLIKU TEOORIA ALUSEL ON VÕIMALIK ENNUSTADA NÄHTUSI/FAKTE, MILLE OLEMASOLU HILJEM EKSPERIMENTAALSELT TÕESTATAKSE 2. Elu organiseerituse tasemed - MOLEKULAARNE tase molekulaarbioloogia, geenitehnoloogia, süsteemibioloogia (BIOMOLEKULID ainult ELUSlooduses). Esmane organiseerituse tase. Kõikjal, kus on elu, esinevad biomolekulid: sahhariidid, lipiidid, valgud, nukleiinhapped. - ORGANELLI tase (molekulaarne) rakubioloogia. Uuritakse raku organelle: tuum, ribosoomid, mitokondrid jne. Kui need rakkudest eraldada, ei kanna nad enam elu tunnuseid. Organellide koostööst tulenevad rakkude omadused. - RAKU tase rakubioloogia
(võrdlev anatoomi, geenijärjestuse võrdlus), katse (kui muudetakse üht tingimust ja võrreldakse tulemusi nii muudetud kui muutmata tingimustega katse puhul) 1)Probleemi püstitamine 2)Taustinfo kogunemine 3)Hüpoteesi sõnastamine 4)Hüpoteesi kontrollimine 5)Tulemuste analüüs ja järelduste tegemine 2. Eluslooduse organiseerituse tasemed 1) MOLEKULAARNE tase – molekulaarbioloogia, geenitehnoloogia, süsteemibioloogia . Esmane organiseerituse tase. Kõikjal, kus on elu, esinevad biomolekulid: sahhariidid, lipiidid, valgud, nukleiinhapped. 2) ORGANELLI tase – (molekulaarne) rakubioloogia. Uuritakse raku organelle: tuum, ribosoomid, mitokondrid, lüsoosoomid jne. Kui need rakkudest eraldada, ei kanna nad enam elu tunnuseid. Organellide koostööst tulenevad rakkude omadused. 3) 3)RAKU tase – rakubioloogia, tsütoloogia
tulemusi nii muudetud kui muutmata (st kontroll) tingimustega katse puhul Biokeemilised meetodid Biofüüsikalised meetodid (nt valkude struktuuri analüüs) Mikroskoopia (valgus- ja elektronmikroskoopia) Geneetilised meetodid (mutatsioonanalüüs koos molekulaargeneetikaga) Eluslooduse organiseerituse tasemed MOLEKULAARNE tase – molekulaarbioloogia, geenitehnoloogia, süsteemibioloogia (BIOMOLEKULID ainult ELUSlooduses) ORGANELLI tase – (molekulaarne) rakubioloogia RAKU tase - rakubioloogia KOE tase - histoloogia, arengubioloogia/embrüoloogia. Inimesel põhikoed: epiteel-, lihas-, närvi- ja sidekude ELUNDI tase – ERI KOED (Tissues) moodustavad ELUNDID e. ORGANEID (anatoomia, füsioloogia). Organitest moodustuvad ELUNDKONNAD e. ORGANSÜSTEEMID (füsioloogia)
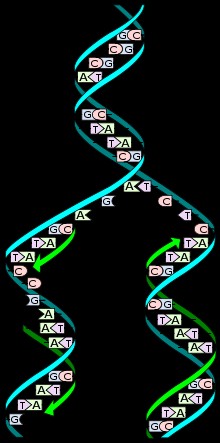
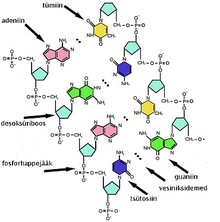
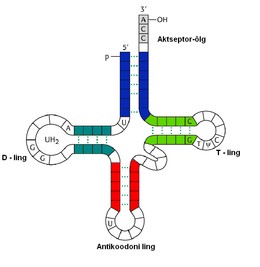
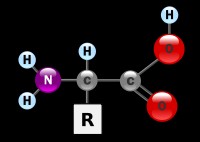
Kordamisküsimused Geenitehnoloogia I 1. Millised molekulid on polümeerid? Polümeerid ehk kõrgmolekulaarsed ühendid on ained, mille molekulid koosnevad kovalentsete sidemetega seotud korduvatest struktuuriühikutest elementaarlülidest. Looduslikud polümeerid: polüsahhariidid (tselluloos, kitiin, tärklis), valgud, nukleiinhapped (DNA, RNA). Polümeerid on väga suured molekulid, moodustunud kui sajad monomeerid liituvad pikkadeks ahelateks. 2. Nukleotiidide lühiiseloomustus. Nukleotiidid on orgaanilised molekulid, mis moodustavad suuri biopolümeere- nukleiinhappeid, näiteks DNA ja RNA. Nukleotiidid on DNA ja RNA molekuli alaüksused, mis koosnevad lämmastikalusest (N-alus), suhkrust (riboos või desoksüriboos) ja fosfaatrühmast. Lämmastikalused on kas puriini või pürimidiini derivaadid. Puriinid: kahte lämmatikku sisaldava tsükliga ühendid, aden
1. Millised molekulid on polümeerid? Molekulid, kus üks struktuuriüksus esineb palju kordi, see üksus võib koosneda ühest või mitmest erinevast monomeerist. 2. Millised biopolümeerid esinevad rakkudes? Nukleiinhapped, valgud, polüsahhariidid, ligniin (moodustab suure osa taimse materjali rakukestadest). 3. Nukleotiidide suhkrujääkide lühiiseloomustus. Riboos ja desoksüriboos, 5-süsinikulised suhkrud ehk pentoosid, erinevus seisneb selles, et desoksüriboosil on 2. süsiniku juures hüdroksüülrühma asemel vesinik 4. Nukleotiidide lämmastikaluste lühiiseloomustus. Dna nukleotiidide lämmastikalused on Adeniin, Guaniin, Tsütosiin ja Tümiin, Rna nukleotiidides on 3 lämmastikalust samad, kui Tümiini asemel on Uratsiil. Lämmastikalused võivad olla suuremad, kahetsüklilised puriinid (A ja G) või väiksemad, ühetsüklilised pürimidiinid (C, T ja U). Lämmastikalused on omavahel komplementaarsed ja komplementaarsete lämmastikaluste vahel moodustuvad vesiniksidem
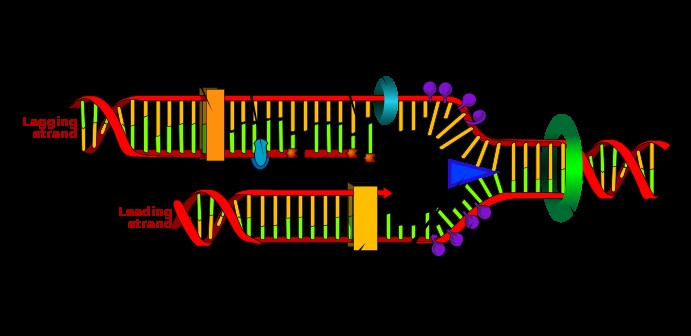
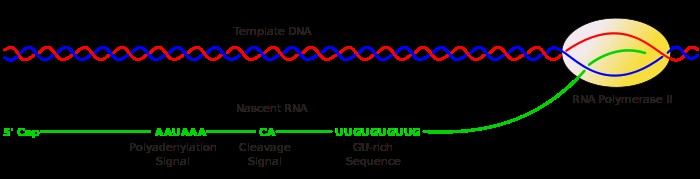
TRANSLATSIOON e valgusüntees, toimub tsütoplasmas, ribosoomides. Aminohapetest sünteesitakse polüpeptiidahel. Protsessi viib läbi ribosoomikompleks, mis koosneb: · 30S subühikust (eukar: 40S) ja 50S subühikust (prokar: 60S); või vastavalt väike ja suur subühik · mRNA (sisaldab geneetilist koodi) · initsiaator-tRNA · initsiatsiooni- või elongatsioonifaktor (oleneb faasist). Protsessis on kolm faasi: initstiatsioon -> elongatsioon -> terminatsioon. Ribosoom läbib selle käigus valgusünteesi ribosoomi tsükli. Vastavalt faasidele toimub: funktsionaalse ribosoomi moodustumine -> aminohapete lisamine peptiidahelasse -> sünteesitud valgu vabastamine ribosoomist. Tegu on kahe-astmelise dekodeerimisprotsessiga: 1. preribosomaalne etapp -> aminoatsüül-tRNA süntees 2. ribosomaalne etapp -> koodon-antikoodon translatsioon ja peptidsideme süntees ribosoomil. Avatud lugemisraam e valkukodeeriv järjestus - nukleiinhappe järjestus, mis













































Kõik kommentaarid