Kordamisküsimused,
Genoomi Struktuur ja Funktsioon 2014/2015.NB! Need ei
ole
eksamiküsimused . Need on märksõnad, mille
tausta teada.
Eksamiküsimused (3 suuremat, 3 väiksemat) on pigem laiemat tüüpi
ja integreeritud
teadmistele
suunatud. Põhinevad loengumaterjalidel ja üldisemas kontekstis
seminarides
käsitletud teemadel . Seminariartiklite üksikuid detaile eksamil ei
küsita.
Genoomi arhitektuur :Kromosoomistik on liigispetsiifiline, igal liigil on kindel arv, kindla kujuga
kromosoome. Näiteks inimestel on 46 kromosoomi,
koertel 78,
kassidel 38, tubakal ja
kartulil 48 jne.
- Mõisted Eu- ja heterokromatiin: omadused, funktsioon.
Kromatiin jaguneb eu- ja heterokromatiiniks.
Eukromatiin
on valdavalt dekondenseerunud olekus ning seal toimub geenide
transkriptsioon ehk on transkriptsiooniliselt aktiivne
interfaasis .
On geenirikas. Eukromatiini iseloomustab histoonide kõrge
atsetüleerituse tase (ning samuti on transkriptsiooniliselt
aktiivsele kromatiinile
omased histoonide metüleerimised). Tema
replikatsiooni aeg on
varieeruv .
Heterokromatiin
on transkriptsiooniliselt inaktiivne. Esineb tsentromeerides ja
telomeerides. Heterokromatiini iseloomustab võime
levida : kui on
juba tekkinud hakkab toimuma
ahelreaktsioon , mis viib ühe suurema
osa
kromatiini heterokromatiniseerumisele.
Rakud peavad vaeva nägema
selle tõkestamiseks. Heterokromatiini iseloomustavad
deatsetüleeritud histoonid.
Replikatsioon toimub hiljem.
Heterokromatiin:
- on oluline tsentromeerides, heterokromatiini valgud seonduvad DNA kordustele mis on tsentromeeride ümbruses ning on vajalikud õigeks tütarkromatiidide seondumiseks ja kromosoomi segregatsioonis.
- stabiliseerib korduvaid DNA järjestusi tsentromeerides ja mujal genoomis inhibeerides rekombinatsiooni homoloogsete korduste vahel.
- heterokromatiin reguleerib geeniekspressiooni arengu ja rakkude diferentseerumise ajal.
- X-kromosoomi inaktivatsioon emaste imetajate somaatilistes rakkudes.
- Heterokromatiini olek päritakse epigeneetiliselt rakkude jagunemise käigus
Heterokromatiin
jaguneb omakorda konstitutiivseks ja fakultatiivseks
heterokromatiiniks.
Konstitutiivse
heterokromatiini puhul pole teada transkriptsioonilist funktsiooni.
On tavaline DNA, ei esine Mendeliaalseid geene, lokaliseeruvad
C-vöötides, strukturaalselt polümorfsed kromosoomide 1, 9, 16,Y
tsentromeerides, ei toimu ristsiiret meioosis.
Fakultatiivne
heterokromatiini funktsioon on geenide
regulatsioon . On
inaktiveeritud eukromaatiline DNA, esineb geene, G- ja R-vöötides,
dünaamiline heterokromatinisatsioon: nt X-kromosoomi inaktivatsioon,
meioosis toimub
ristsiire .
- Metafaasi kromosoomide ‘vöödistamine’, metoodika(te) põhimõte
Kromosoomide
vöödistamine on oluline tehnika kromosoomide segmentide
värvimiseks, et identifitseerida individuaalseid kromosoome ja
kromosoomi osasid.
- G- banding – AT spetsiifiline värving Giemsa värviga. Heterokromatiinsed regioonid , mis on AT-rikkad ja geenivaesed, värvuvad tumedaks. GT rikkad transkriptsiooniliset rohkem aktiivsed alad värvuvad kehvemini ning jäävad heledateks.
- R-banding – reverse Giemsa, vastupidine vöödistus G-banding’ule. GC spetsiifiline. Värvib geenirikkad alad, avatud kromatiini.
- Q-banding – sama mustriga kui G-banding, fluorestseeruv vöödistus.
- T-banding – kasutatakse telomeersete regioonide analüüsiks/värvimiseks. GC-spetsiifiline.
- ‘avatud kromatiin’: mõiste, paigutus , omadused
Avatud
kromatiin on eukromatiin, dekondenseerunud kromatiin, mis paikneb
peamiselt transkriptsiooni algussaitides või selle läheduses ja
kattub CTCF seondumissaitidega. Avatud kromatiin on GC-rikas, asub T
ja R kromosoomi vöötides, geenirikas, Alu-elemendirikas, vähem
kondenseerunud, kõrge histoonide atsetüleeritusega, replitseerub
varases faasis ja ning transkriptsiooniliselt aktiivne.
- Genoomi ‘pakkumine’ kromosoomidesse – klassikaline mudel ja selle kriitika.
2nm DNA
kaksikheeliks keerdub ümber histoonide. DNA pakkimise väikseim ühik
on nukleosoom, mis koosneb 8-st histoonist ja 147-st aluspaarist
DNAst . Nukleosoomi tuumiku moodustavad histoonid H3, H4, H2A ja H2B,
mida kõiki on kahes korduses.
Nendest H3 ja H4 moodustavad
tetrameeri, millele omakorda kinnituvad kaks H2A-H2B dimeeri. Ümber
histoonide oktameeri on
keerdunud DNA. Nukleosoomide rida DNAl
moodustab pärlikee struktuuri, so 10 nm
fiiber . Järgmises
etapis pakitakse nukleosoomid omavahel kokku, mille tulemusena moodustub 30
nm kromatiini fiiber. Nukleosoomide vahele jäävale DNA-le
seondub histoon H1 ja toob nukleosoomid omavahel kokku. 30 nm fiiber kinnitub
lingudena kromosoomi tugivalkudele (chromosome scaffold),
tugivalkudega
seonduvaid DNA piirkondi nim SAR-iks (scaffold
associated
region ). 30 nm fiiber moodustab
loop ’e, mis on umbes 300
nm pikad. 300 nm
fiibrid surutakse kokku tihedalt pakitud spiraaliks
ning moodustub 700 nm fiiber. Ning 700 nm fiiber keritakse tihedalt
kokku metafaasi kromosoomi kromatiidiks (
1400 nm).
Kriitika –
kas 30 nm fiibrid eksisteerivad ka in
vivo ? On mitmeid töid 30 nm
fiibri eksisteerimise kohta in
vitro katsetes, kuid mitu tööd ka,
mis kummutavad 30 nm fiibri olemasolu in vivo. Krüo-EMga
külmutatud inimese mitootilisi rakke vaadates ei ole näha 30 nm kromatiini
fiibreid kromosoomis.
- Genoomi ‘pakkumine’ kromosoomidesse – uus ‘polümeeri sulam’ (‘polymer melt’) mudel, selle tõendusmaterjal ja funktsionaalsed eelised.
Mitootiliste
kromosoomide nukleosoomi fiibrid ja aktiivsed interfaasi
tuumad ei
moodusta 30 nm kromatiini fiibreid, vaid eksisteerivad
ebakorrapärases segunenud olekus, mis on sarnane dünaamilise
liikuvusega polümeerile. Tõendusmaterjal – Krüo-EM katsed
külmutatud imetajate mitootiliste rakkudega ja Xenopus 3D
kromosoomide kokkupanek in vitro näitas 30 nm kromatiini fiibri
puudumist täielikult kondenseerunud kromatiinis. Kõrgemate
eukarüootide ja pärmi interfaasi tuumade
uurimine krüo-EM ja 3C
meetodiga ei näidanud samuti 30 nm fiibri olemasolu. Mudeli eelised:
kuna nukleosoomi fiibri voltumine määrab DNA kättesaadavuse on
nendel dünaamilistel protsessidel mitmeid
eeliseid transkriptsiooni
regulatsioonil ja DNA replikatsioonil, mudelil on eeliseid
transkriptsiooni komplekside seondumisel, 30 nm fiibri puhul on
ennustatavalt need märklaudjärjestused peidus. Dünaamiline
ebaregulaarne voltumine võib tekitada loop’e, mis hõlbustab
promootor - ja enhanserjärjestuste vahelisi interaktsioone. Samuti
madala energeetilise tasuga geeniekspressiooni vaigistamine.
- Boveri–Sutton kromosoomi theooria; Boveri kromosomaalsete territooriumite mudel, omaaegsed põhjused selles kahtlemiseks
Teooria,
mille kohaselt kromosoomid on geneetilise materjali
kandjaks . Teooria
selgitab, et mendellik
pärandumine on seotud kromosoomidega. Samuti
väidab, et kromosoomid on
lineaarsed struktuurid , kus
geenid paiknevad
konkreetsetes saitides
pikki kromosoome. Ehk jagunevates
rakkudes olevad kromosoomid on geneetilise pärandumise aluseks.
Näitasid, et kromosoomid esinevad
paaridena ema ja isa
kromosoommidest, mis eralduvad meioosis ning võivad olla aluseks
mendellikule pärandumisele. Kromosomaalsete territooriumite mudel –
kromosoomid ei paikne tuumas hajusalt, vaid neil kõigil on oma
territoorium . Boveri väitis, et kromosoomi poolt hõivatud ala sees
on kromatiini
kimpudest kanalite võrgustik. Kromosoomi
territooriumite
paiknemine säilis stabiilsena interfaasi ajal ning
kromosoomide kaugused muutusid prometafaasi ajal. Ehk interfaasi ajal
hõivavad kromosoomid konkreetse territooriumi tuumas.
Kõhklused
selle teooria kohta tekkisid, kuna tol ajal ei olnud EM nii arenenud,
et tuvastada kromosoomi territooriume. Sellel ajal oli
populaarne arvamus, et ainult konstitutiivne heterokromatiin jääb
kondenseerunuks interfaasi ajal ning eukromatiin dekondenseerub ja
seguneb. Pollisteri teooria kohaselt oli tuum nagu kromosoomide
lõngakera, mis on sõlmes.
- FISH metoodikate põhimõte; plussid ja miinused teadusuuringutes.
FISH –
fluorecent in situ hybridisation – kasutatakse kromosoomidel DNA
järjestuste kaardistamiseks ning nende puudumise või olemasolu
kinnitamiseks. Kasutatakse geenide positsioonide
identifitseerimiseks, kromosomaalsete hälvete diagnoosimisel,
tervete kromosoomide värvimisel, interfaasi kromosoomide
analüüsimisel jne. Kromosomaalne sihtmärk-DNA denatureeritakse
ning hübridiseeritakse märgistatud prooviga/sondiga, mis on samuti
eelnevalt
denatureeritud ja märgistatud fluorofoori või hapteeniga.
Fluorestseeruvat reportermolekuli on näha fluorestsentsmikroskoobiga
ning saadakse infot teatud geneetilise aberratsiooni olemasolust või
puudumisest.
Plussid –
saab uurida kromosomaalseid aberratsioone mittejagunevates rakkudes.
On kõrge tundlikkuse ja spetsiifilisusega. Ulatusliku
detekteerimisvõimega – saab mitmeid erinevaid teadaolevaid
aberratsioone uurida.
Miinused –
saab uurida ainult teadaolevaid geneetilisi aberratsioone, kuna on
vaja spetsiifilise järjestusega sondi hübridisatsiooniks. On vaja
teada millist piirkonda vaja uurida, kus
kromosoomil paigutub jne.
- Cremer’ite uuesti esile toodud ja edasiarendatud interfaasi kromosoomide territooriumite (CT) mudel, sobivus uue kromatiini ‘pakkumise’ mudeliga , mille aluseks on 10 nm kromatiini fiiber.
Näitasid,
et interfaasi tuum on kompartmentaliseeritud kindlateks kromosoomi
territooriumiteks (CT). Kromosoomide territooriumite pinnad ja nende
vahelised alad moodustavad
võrgustiku -laadse 3D tuuma sektsiooni
geeniekspressiooniks, mRNA splaissinguks ja
transpordiks mida nim
interkromatiini kompartmendiks (IC).
10 nm
fiiber ebakorrapärases olekus, mis on dünaamilise liikumisega või
klasterdunud CT-tes avatud ja suletud kompartmentite gloobulites.
Mudeli kohaselt asuvad aktiivsed
geenid territooriumite pindadel ning
mRNA
transkriptid vabanevad IC ruumi.
Kromatiini
domeenid on kromosoomide territooriumite komponendid.
Interkromatiinne
kompartment esineb kromatiini domeenide vahel, see
on DNA-vaba piirkond ning sisaldab splaissimise spekleid.
Perikromatiinses ruumis toimub perikromatiini fibrillide tootmine.
Iga
fibrill kannab
vastset transkripti erinevatelt geenidelt.
Splaissimise speklid varustavad fibrille splaissimise faktoritega.
- CT-de ruumiline paigutus tuumas – üldised tendentsid
Iga
kromosoom hõivad kindla territooriumi interfaasi tuumas ning need
territooriumid ei kattu üksteisega. Geenirikkad kromosoomid
paiknevad rohkem tuuma keskel ning suuremad geenivaesed kromosoomid
paiknevad pigem perifeerias. Kromosoomi territooriumid on
dünaamilised struktuurid, kus geenid saavad sisselülitamisel
perifeeriast ümberpaigutuda tuuma
keskele poole.
- Kromosoomide territooriumite (CT)-interkromatiin kompartment (IC) mudel: kromatiini ja IC paigutus; ‘avatud’ ja suletud kromatiini paigutus; tuumapoorid ja IC; laguunid
CT-IC
mudelis moodustavad kromosoomi territooriumid ühendatud kromatiini
võrgustiku, mis on seotud külgneva 3D ruumiga, mida kutsutakse
interkromatiini kompartmentiks. Ehk tuum on ülesehitatud kahe
komponendi poolt – kromosoomi territooriumitest ja interkromatiini
kompartmentist. Üksikud kromatiini territooriumid moodustavad
ühendatud võrgustiku ning see võrgustik on ruumiliselt seotud
teise 3D võrgustikuga, milleks on IC. IC on nagu DNA-vaba kanalite
ruum. Geenirikkad kromosoomid (ehk avatud kromatiin) paikneb rohkem
tuuma keskel ning suuremad geenivaesed kromosoomid (suletud
kromatiin) paikneb pigem perifeerias. Kromosoomi territooriumid on
dünaamilised struktuurid, kus geenid saavad sisselülitamisel
perifeeriast ümberpaigutuda tuuma keskele poole. Interkromatiini
kanalid algavad tuumapooridest ning laienevad kromatiini
territooriumite võrgustiku vahel nagu suuremad kanalid ja laguunid.
Ehk laieneva suurusega mittekromatiinsed domeenid lükkavad
ümbritsevad kromatiini fiibrid kõrvale ja suurendavad
interkromatiini ruumi laguunideks. Interkromatiini kompartmendis
liiguvad vabalt transkriptsiooni- ja splaissingu
faktorid .
DNA-vaba
võrgustik
laieneb kromatiini võrgustiku vahel, interkromatiini
kanalid algavad tuumapooridest ning laienevad nii CT-de perifeeriasse
kui ka nende sisemusse. Sisemised laiemad IC laguunid sisaldavad
splaissingu faktoreid.
- CT-de detailsem struktuur: kromatiini domäänid (CD)=fraktaalsed gloobulid= ‘chromatin liquid drops’; periromatiinne piirkond.
Kromatiini
gloobulid on tihedalt pakitud kromatiini domeenid. Fraktaalsed
gloobulid moodustavad kromosoomi territooriumeid. Gloobuli tuumikus
paiknevad aktiivselt transkribeeritavad geenid ning
mittetranskribeeritav kromatiin paikneb gloobuli perifeerias. Iga
piirkond fraktaalsest gloobulist on võimeline kiiresti lahti
voltuma, translokeeruma ning muutuma transkriptsioonifaktoritele ja
teistele
valkudele kättesaadavateks.
Perikromatiinne
piirkond on funktsionaalne ruum tuumas, mis määrab kromatiini
voltumise. Selles piirkonnas esinevad replikatsiooni,
transkriptsiooni ja RNA prosessingu faktorid. Perikromatiinne
piirkonnas toimuvad transkriptsioon, kotransktiptsiooniline
vaigistamine, DNA replikatsioon ja tõenäoliselt ka
reparatsioon .
Nii RNA kui DNA süntees toimub perikromatiinses piirkonnas, kuid
mitte sisemistes interkromatiini kompartmendi laguunides.
Perikromatiinne
piirkond
jaotab kromosoomide territooriumi kromatiini domeenideks.
- CT-de (ja CD-de) funktsioneerimine: kromosoomide ja kromatiini dünaamika mudelid genoomi funktsiooni tagamisel ja geenide koordineeritud ekspressiooniprofiili tagamisel.
Geeniregulatsioonil
erinevate kromosoomide osad suhtlevad teine
teisega ning toovad
seotud geenid kokku, et koordineerida nende ekspressiooni. Seda
geenide kokkupuudet nimetatakse gene kissing ehk geenisuudluseks.
Selle tagajärjel toimub transkriptsiooniline
aktivatsioon ,
vaigistamine või koekspressioon. Geenisuudlus võib toimuda ühe
kromosoomi territooriumi siseselt (cis) või kahe või enama
kromosoomi territooriumi geenide vahel (trans). Võib toimuda naaber
CT-te
piiril , ühe CT silmuse penetratsioonil naaber CT-sse, või
mitme erineva CT suurte
silmuste interaktsioonil.
Mudelid: Kompartmentid on juba olemasolevad struktuurid mis sisaldavad molekulaarset masinavärki, mis on pühendunud spetsiifilistele tuumafunktsioonidele.
Kompartmentid on tekkivad ja kaduvad iseornagiseeruvad üksused – geeni aktivatsiooniga kaasneb vaigistamiskompartmentide likvideerimine, muutused geenipositsioonis ja regulatoorsete kompartmentite iseassambleerumise aktivatsioon.
- Transkriptsiooni ‘vabrikud’: paigutus, arv tuumas, tekke ja funktsioneerimise mudel(id).
Äsja
sünteesitud transkriptid ja aktiivne RNA polümeraas II ei ole üle
tuuma jagunenud, vaid on kontsentreeritud kindlates piirkondades mida
nimetatakse transkriptsioonivabrikuteks ehk aktivatsiooni
kompartmentiteks. Ühes tuumas esineb mõni sada kuni mõni tuhat transkriptsioonivabrikut olenevalt raku tüübist ja
diferentseerumisolekust, ning paiknevad üle tuuma.
Transkriptsioonivabrikute diameeter on 45-100 nm. Transkriptsiooniga
hõivatud RNA polümeraas II fraktsioon vabrikutes on seotud mingi
alamstruktuuriga. Ehk vabrikud on immobiliseeritud mingi struktuuri
külge. Vabrikud on valgurikkad – sisaldavad aktiivset või
inaktiivset RNA polümeraasi, transkriptsioonifaktoreid,
ribonukleoproteiine, ko-aktivaatoreid, RNA helikaasi ja splaissingu
ja protsessingu ensüüme. Vabrik sisaldab ainult ühte tüüpi
polümeraase. Ühes vabrikus võidakse transkribeerida samadelt või
erinevatelt kromosoomidelt pärit geene.
Teke – on
vastuolulisi arvamusi selle kohta kas transkriptsiooni vabrikud
pannakse kokku de
novo
vastusena transkriptsiooni nõudlusele või kas nad on stabiilsed
struktuurid millede number raku tuumas püsib konstantsena. Ehk kas
RNAP II transporditakse vabrikusse geenide juurde või geenid
transporditakse vabrikusse, kus juba on olemas immobiliseeritud RNAP
II. Eelisatakse teist mudelit, mille korral arvatakse et RNAP II on
immobiliseeritud.
- Splaissingu ‘vabrikud’ (=tuuma speklid, splaisingu speklid)
Splaissingu
vabrikud ehk tuuma speklid on väikesed membraanitud organellid või
struktuurid, mis sisaldavad valku. See valk on faktorite
reservuaariks, mis osaleb transkriptsioonil ja pre-mRNA
protsessingul. Splaissingu vabrikud on irregulaarsed intervallidena
esinevad sturktuurid tuumas, mis varieeruvad suuruse ja kuju poolest.
Umbes 25-50 speklit esineb ühes imetaja interfaasi tuumas. Speklid
koosnevad interkromatiini graanulite klastritest. Splaissingu
vabrikud on dünaamilised struktuurid – nii nende valgulised kui
RNA-valk komponendid saavad ringelda pidevalt spektite ja teiste
tuuma piirkondade vahel olenevalt raku transkriptsioonilisest
olekust. Nad asuvad kromosoomi territooriumites, kromatiini domeenide
vahel.
Speklid
moodustavad klastreid interkromatiinse kompartmenti laguunides.
Laguunid on ainult osaliselt tidetud speklitega.
- 3C (Chromosome conformation capture) metoodilisel lähenemise põhimõte ja andmete interpreteerimine genoomi struktuuri ja funktsiooni uurimisel ; eelised (e.g. FISH ees), miinused.
3C
meetodiga uuritakse kromosoomide organiseeritust raku loomulikus
olekus. Põhimõte seisneb lähedal asuvate lookuste paaride sageduse
kindlaksmääramisel nende ristsidumisega. Toimub kolme sammuliselt:
Ristsidumine – formaldehüüdi lisamisel üksteisele lähedal asuvad kromatiini segmendid ristseotakse. DNA segmendid ristseotakse valkudega ja valgud ristseotakse üksteisega.
Kromatiin fragmenteeritakse ja ligeeritakse
DNA puhastatakse ja analüüsitakse ( qPCR , sekveneerimine )
3C: one by
one analüüs – kasutab 3C lookus -spetsiifilisi praimereid. Tulemus
nt interaktsiooniprofiil valitud geenist näitab promootori ja
ümbritseva kromatiini interaktsiooni .
4C:
tsirkulariseeritud 3C, one by all analüüs – üle genoomne
interaktsiooniprofiil üksikule lookusele.
5C: many by
many analüüs – analüüsib interaktsioone kahe suurema lookuste
kogumi vahel, nt promootorite kogumi ja distaalsete regulatoorsete
elementide kogumi vahel. Ehk võimaldab analüüsida interaktsioone
mitme valitud lookuse vahel.
Hi-C –
all by all analüüs
Andmete
interpreteerimine – 3C ja 4C annab lineaarsed profiilid piki
kromosoomi. 3C meetod annab andmeid lookuste lähedal paiknemise
kohta, kuid ei erista funktsionaalseid mittefunktsionaalsetest. 3D
andmed esindavad suuremat interaktsioonide summat üle raku
populatsiooni. 5C ja HiC interaktsioonikaardid – mida tumedam värv,
seda tugevam interaktsioon .
Eelised
- Võimaldab detekteerida genoomseid interaktsioone.
- Need interaktsioonid võivad paljastada geeniekspressiooni regulatsiooni detaile.
- 5C võimaldab konstrueerida komplekseid interaktsioone spetsiifilistes lookustes.
Miinused
- Produkti detekteerimine ei tähenda alati, et on toimunud spetsiifiline interaktsioon kahe regiooni vahel. spetsiifiline interaktsioon toimub kui interaktsioon on kõrgema sagedusega kui naaber-DNAdel.
- Ei tuvasta funktsionaalseid mittefunktsionaalsetest interaktsioonidest.
- 5C ei sobi ülegenoomsete interaktsioonide uurimisel kuna vajaks miljoneid 5C praimereid.
- Suureks miinuseks kõikide puhul on see et nad vajavad palju rakke.
- Interaktsiooni sagedused saadakse juhuslike üksustena ja tegelikud interaktsioonide sagedused raku populatsioonis jäävad teadmata.
- 3C (ja selle metoodiliste edasiarenduste, nagu HiC) põhiste uuringute olulisemad leiud genoomi erinevate piirkondade interaktsiooni kohta.
- Interfaasi kromosoomid hõivavad individuaalsed territooriumid tuumas - kromosoomi territooriumid.
- Interfaasi kromosomaalsed territooriumid on kompartmentaliseeritud:
- suured aktiivsed A ja inaktiivsed B kompartmendid (fraktaalsed gloobulid)
- väiksemad topoloogiliselt assotseerunud domeenid (TAD)
- kromatiini silmused ühendavad geene kaugemate regulatoorsete elementidega, vahendades pikamaalisi geeniinteraktsioone.
- Interfaasis A/B kompartmentite ja TADide mustrid on raku tüübi spetsiifilised .
- Interfaas ja metafaas esindavad kahte funktsionaalselt erinevat genoomi 3D organisatsiooni.
- Mitootilised kromosoomid säilitavad mõned, kui üldse, strukturaalsed omadeused mis iseloomustavad interfaasikromosoome.
- Metafaasi kromosoomid omandavad sarnase organisatsiooni erinevates rakutüüpides.
- Metafaasi kromosoomides kaob A/B kompartmentitesse segregatsioon ja puuduvad TADid.
- HiC põhiste uuringute põhjal tuletatud uued genoomi struktuuri üksused: (i) A & B kompartmendid, seos vastava genoomse piirkonna üldomadustega; (ii) topologically associated domains (TADs); (iii) ‘fraktaalne gloobul’ (FC)
A/B kompartmendid – vahelduvad piki kromosoomi. A kompartmendid interakteeruvad eelistatult teiste A kompartmentidega genoomis. B kompartmendid interakteeruvad teiste B kompartmentidega. A kompartmentid on suuresti eukromaatilised, transkriptsiooniliselt aktiivsed. A ja B kompartmendid moodustavad kromosoomidel aktiivse ja inaktiivse kromatiini mustereid. A ja B kompartmendid on koespetsiifilised. A ja B kompartmendid vastavad fraktaalsetele gloobulitele polümeeri sulami mudelis.
TADid – interfaasi kromosoomid koosnevad topoloogiliselt seotud domeenidest. TADid on piirnevad kromosomaalsed regioonid, mis interakteeruvad iseendaga ning on isoleeritud naabritest. TADid on universaalsed kromosoomide ehituskivid. TADid on defineeritud geneetiliselt kodeeritud piiri elementide poolt, mis on rikastatud CTCF-seotud lookustega. Arvatakse et silmus -interaktsioonid on piiratud sama TADi kuuluvate elementide vahel. Sama TADi geenid kipuvad olema koordineeritult ekspresseeritud.
Fraktaalne gloobul – ilmneb 10 nm fiibri polümeeri kondensatsiooni tagajärjel, mille kestel topoloogilised piirded takistavad sõlmumist ja aeglustavad polümeeri tasakaalu. Gloobulid võivad esindada kas A või B kompartmente. Umbes 10 TADi moodustavad ühe fraktaalse gloobuli.
G1, S
faasis esinevad kompartmendid ja TAD struktuur, mitoosis
kompartmendid ja TAD strukutuur kaovad. Tekib fiiber mille aluseks on
silmused.
Esimene
faas – lineaarne kokkupakkumine järjestikuste kromosomaalsete
silmuste moodustumisega.
Teine faas
– fiibri selgroo aksiaalne kokkusurumine , moodustub tihedalt
pakitud kromosoom
- FISH metoodikate põhistel uuringute alusel loodud CT-IC mudeli ja 3C metoodikate põhistel uuringute alusel loodud FC-TAD mudeli järelduste kokkulangevus.
FISH
3C
CT põhiliseks strukturaalseks elemendiks on kromatiini domeenid.
CT põhiliseks strukturaalseks elemendiks fraktaalne gloobul.
Nii CD kui FG sarnased suuruse poolest.
Kromatiini domeenid on ehitatud mitmetest kromatiini silmustest.
Kromosoomid koosnevad diskreetsetest dopoloogiliselt seotud domeenidest (TADidest). Ühes gloobulis u 10 TADi.
Dünaamiline iseloom võimaldab CD pindade lühiajalist kontakti in cis ja in trans, mis võimaldab kromosomaalseid korraldusi ja mitte-suvaliste kromatiini interaktsioonide moodustumist.
Silmus-interaktsioonid on piiratud ühte TADi ja tihti jätavad vahele ühe või rohkem geene lineaarsesse dimensiooni. Samas TADis olevad geenid võivad koekspresseeruda.
Perikromatiinne regioon on piirkond kus toimub transkriptsioon, ko-transkriptsiooniline vaigistamine, DNA replikatsioon ja ilmselt ka reparatsioon.
2 kompartmenti A ja B – mis vahelduvalt esinevad piki kromosoomi moodustades mustri.
- Genoomi strukturaalne varieeruvus ja selle dünaamika:
- Suured sturktuursed genoomi varieeruvused ( mikroskoobi tasemel resolutsioon ), nende detekteerimise võimalused.
- Aneuploidiad – üks või rohkem ekstra kromosoomi või kromosoomide puudumine
- Isokromosoomid – kaks identset kromosomaalset õlga
- Õrnad saidid – kordused mis on võimelised võtma ebatavalise mitte B-DNA vormi, mis võib häirida DNA replikatsiooni. Nt fragiilse X sündroom – detekteeritakse X kromosoomi muutusi vaadeldes, esineb CGG trinukleotiidse korduste arvu suurenemine FMR1 geenis.
- Marker kromosoomid – strukturaalselt ebanormaalsed kromosoomid, kus ei ole võimalik ühtegi osa identifitseerida
- Peritsentriline heterokromatiinne variatsioon kromosoomides ChrY , Chr9 ja Chr15
- Satelliitregioonide variandid akrotsentrilistes kromosoomides
-
- Segmentaalsed duplikatsiooonid: definitsioon, põhiomadused, rikastatus ja jaotuvus genoomis (intra- ja interkromosomaalsed), ebastabiilsus , olulisus eukarüootsete genoomide evolutsioonis .
- Segmentaalsed duplikatsioonid on järjestuste ülekanne genoomi ühest regioonist teise. Segmentaalsed duplikatsioonid on DNA segmendid, millel on peaaegu identne järjestus. Segmentaalsed duplikatsioonid on kromosoomi-spetsiifilised madala koopiaarvuga kordused (LCR), duplikonid, millel on suur homoloogia. Segmentaalsed duplikatsioonid on kas peritsentromeetrsed (peamiselt interkromosomaalsed), subtelomeersed (nii inter - kui intrakromosomaalsed) või interstitsiaalne (kromosoomi õlgades).
- Erinevatel kromosoomidel on erinev segmentaalse duplikatsiooni muster – kromosoomid 7, 9, 15, 16, 17, 19, 22 ja Y on rikastatud duplikatsioonidega ning kromosoomid 2, 3, 4, 5, 8, 14 ja 20 on vähendatud duplikatsioonidega.
- Intrakromosomaalsete duplikatsioonide ja geenide tiheduse vahel esineb positiivne korrelatsioon . Segmentaalsed duplikatsioonid ei ole suvaliselt jaotunud – geenid, mis on seotud immuunsuse ja kaitsega , membraani pinna interaktsioonidega, ravimite detoksifikatsiooniga, kasvu ja arenguga on rikastatud. Segmentaalsed duplikatsioonid on aluseks evolutsioonile duplikatsioonide kaudu – Susumu Ohno teooria – Ohno pakkus välja, et geenide duplikatsioon on peamine tegur uute geenide arengul. Mis on praeguseks hetkeks teada üks olulisimaid mehanisme uute geenide tekkel. On tavaline evolutsiooni mehanim ning seega esineb paljudes liikides. Segmentaalsed duplikatsioonid kolokaliseeruvad tihti koos evolutsiooniliste kromosomaalsete katkete kohtadega.
- SD on ebastabiilsed regioonid ning võivad olla CNV-de tekke allikaks mitte-alleelse homoloogilise rekombinatsiooni kaudu.
-
-
-
-
-
- Segmentaalsed duplikatsioonid kui genoomi nn. stuktuursete genoomsete varieeruvuste ( Copy number variation ’, CNV) tekke allikad, mehhanismid CNVde tekkeks.
- Segmentaalsed duplikatsioonid on ebastabiilsed regioonid, mis võivad läbi mitte-alleelse homoloogilise rekombinatsiooni või konversiooni tekitada koopiaarvu variatioone CNV-sid (deletsioonid, duplikatsioonid, inversioonid).
-
NAHR – suure identsusega SD-de vahel võib põhjustada edasisi ümberkorraldusi olenevalt SD-de koopiate paiknemisest ja orientatsioonist. NAHR on DNA homoloogsete piirkondade vaheline ristsiire.
Külgnevate duplitseeritud järjestuse vahel toimuva NAHR tagajärjel võivad tekkida tandemduplikatsioonid ja deletsioonid (Aa).
Translokatsioonid võivad tekkida mittehomoloogiliste kromosoomide SD-de vahelise vahetuse tagajärjel (Ab).
- Mõlemal juhul on tekkinud koopiad sama orientatsiooniga kui originaalne SD.
Inversioonid võivad tekkida pööratud intrakromosomaalsete duplikatsioonide vahelise rekombinatsiooni tagajärjel (Ac).
Konversioon kahe SD vahel toimub koopiate vahelise järjestuse informatsiooni vahetuse tagajärjel. See võib suurendada alleelset mitmekesisust konverteeritud koopias ja põhjustada SD koopiate homogenisatsiooni.
-
-
-
- CNV-de määramine genoomis: põhimeetodid (CGH, SNP- kiibid , ‘ paired -end’ sekveneerimine), plussid ja miinused.
- CGH – comparative genomic hybridization – kasutatakse uuritavas DNA proovis teatud geenide või DNA lõikude koopiaarvu muutuste või deletsioonide leidmiseks.
- Referents -DNA ja test-DNA märgistatakse erinevate fluorestseeruvate märgistega, segatakse kokku ning hübridiseeritakse kiibile. Fluorestsentsi suhteid kasutatakse piirkondade detekteerimiseks test ja referents proovide vahel, mis erinevad koopiaarvult.
- Saab visualiseerida deletsioone ja duplikatsioone väga väikestes DNA segmentides
- Saab uurida tervet genoomi ilma varasema teadmiseta kromosomaalsete aberratsioonide kohta
- Ei vaja spetsiifilisi sonde
- Saab detekteerida amplifitseeritud geenide olemasolu vähis ja kaardistada nende asukoht
- Ebatäpsused teatud kromosoomide regioonides (kus on palju kordusi, akrotsentriliste kromosoomide tsentromeerides ja enamus kromosoomide telomeerides)
- Koopiaarvu muutusi saab tuvastada ainult siis, kui rohkem kui 50% rakkudest sisaldavad kromosomaalseid muutusi.
- Ei saa identifitseerida kromosomaalseid ebanormaalsusi mis on tasakaalus.
- Langenud tundlikkus tänu test rakkude kontaminatsioonile normaalsete rakkude poolt.
- SNP-kiibid – kasutatakse populatsiooni siseste polümorfismide detekteerimiseks. Kiip sisaldab immobiliseeritud alleel -spetsiifilisi oligonukleotiidi sonde. Fragmenteeritud proov märgistatakse fluorestseeruva märgisega ning detekteeritakse hübridisatsiooni signaali alusel.
- Saab töödelda tuhandeid proove madala kuluga
- Saab detekteerida haruldasi ja levinud CNVsid madala valede tulemuste tasemega kasutades viit sondi regiooni kohta.
- Kiire
- Vähi genoomi mosaiikne struktuur muudab CNVde tuvastamise keerulisemaks
- Sondide jaotus ei ole homogeenne – spetsiifiline regioon jääb sidumata , piiratud resolutsioon.
- Ei võimalda tuvastada tasakaalustatud kromosomaalseid translokatsioone, inversioone ja ülegenoomset ploidsuse muutust ilma teiste meetoditega kombineerimist.
- Paired-end sekveneerimine – võimaldab sekveneerida fragmendi mõlemad otsad ja saada kõrge kvaliteediga, joondatavad järjestuse andmed. DNA proov fragmenteeritakse ja spetsiifilise suurusega fragmentide otstesse ligeeritakse adaptorid. Fragmendid tsirkulariseeritakse adaptorite vahendusel, lõigatakse ning selekteeritakse adaptoreid sisaldavad fragmendid. Fragmentide otsad sekveneeritakse kaardistatakse võrrelduna referents järjestusega. Kui otsad on oodatava kaugusega ei esine variatsiooni referents-järjestuse ja proovi vahel. Kui otsad lähemal kui oodatud, on tegemist insertsiooniga proovis (duplikatsioon). Kui otsad on kaugemal lahus kui oodatult, on tegemist deletsiooniga proovis. Kui otsad on mitteoodatud orientatsiooniga on tegu inversiooniga proovis.
- Võimaldab paljastada seosed kahe otsa vahel.
- Võimaldab leida strukturaalseid variatsioon ning isegi molekulaarseid interaktsioone kaugete genoomsete elementide vahel
- Odav
-
- CNVde osakaal ja jaotuvus genoomis
- Ligikaudu 10% inimese genoomist on seotud CNVdega. Kohtades, kus toimub sagedasem replikatsioon, tekib ka variatsioone rohkem. Deletsioonide ja duplikatsioonide suhe on 2:1. Ehk deletsioone on rohkem kui duplikatsioone. Deletsioonid on lühemad, duplikatsioonid pikemad . CNV regioonid on jaotunud ebaühtlaselt genoomis. Duplikatsioonid on rikastatud kromosoomides Chr22, ChrY, Chr 13-16. Deletsioone esineb kõige rohkem kromosoomides 19, 22 ja Y. CNVd on ebaühtlaselt jaotunud piki kromosoome. Peritsentromeersed ja subtelomeersed regioonid sisaldavad rohkem CNVsid. Suurem osa CNVsid esineb mittekodeerivates geenide eksonites, promootorite ja CpG saartes. Kõige vähem CNVsid konserveerunud elementides. Duplikatsioonid on vähem tõenäoliselt patogeensed ning on tihedamini positiivse selektsiooni all, mis juhib evolutsiooni. Deletsioonid on sagedamini patogeensed ning negatiivse selektsiooni all.
-
- Üle 40% kõikidest teadaolevatest koopiaarvu varieeruvustest kattuvad teiste geenidega. See näitab, et neil on olulie roll geeniekspressioonis.
- 21 trisoomia – downi sündroom, X monosoomia – Turneri sündroom, XXY – Klinefelteri sündroom – kindel kliiniline tagajärg, kuid paljudel siiski seda pole. Koopiaarvult stabiilsed regioonid on tundlikud ning nedes on CNVde esinemine tõenäoliselt seotud haigustega.
-
- CNV-de effekt indiviidi genoomi funktsioonile (s.t. ka populatsiooni fenotüübilisele diversiteedile): polümorfsed CNVd ja normaalne fenotüübiline varieeruvus; patoloogilised struktuursed ümberkorraldused (tüüpiliselt >1 Mb) ja genoomsed haigused
-
- Genoomsed haigused – suured CNVd, jäävad 1-100 Mb vahemikku. Nt Downi sündroom, 21 kromosoomi trisoomia. Cri du Chat sündroom 5 kromosoomis deletsioon (u 10 Mb).
- Normaalse fenotüübi CNV-d – CNVd, mis on normaalse fenotüübilise variatsiooni aluseks jäävad vahemikku 10 kb – 1Mb. Nt reesus veregrupp - RhD negatiivsust põhjustab RHD geeni deletsioon. Sülje amülaasi geen on duplitseerunud – AMY1 geeni koopiate arv on indiviiditi varieeruv.
- CNVd mis on seotud normaalse fenotüübilise variatsiooniga ja immuun -funktsioonidega – jäävad alla 1Mb. Nt α-globiini geenide duplikatsioon.
-
- Regulatoorne RNA:
- RNA omadused
Omab erinevaid olulisi rolle raku ainevahetuses
Evolutsiooniliselt varajane
Seondub RNA, DNA, valguga
Lineaarstruktuur võimaldab kodeerivat funktsiooni DNA-RNA-valk.
Sekundaarstruktuur varjab või toob esile valkude seondumiseks olulised motiivid ja tertsiaarstruktuur loob katalüütiliseks reaktsiooniks sobiva mikrokeskkonna
Keemiliselt ja bioloogiliselt labiilne
- mRNA – informatsiooni RNAd , kodeerib valku
- rRNA – osaleb translatsioonil , moodustades mikrokeskkonna katalüütiliseks reaktsiooniks.
- tRNA – osaleb translatsioonil, transpordib aminohappeid ribosoomidesse
- tmRNA – transfer messenger RNA, päästab stoppama jäänud ribosoomid
- snRNA – small nuclear RNA ehk U-RNAd, paiknevad tuumas Cajal kehades , osalevad splaissimisel
- snoRNA – small tuumakese RNA, modifitseerivad teisi RNAsid
- miRNA – mikro RNA, osaleb geeniregulatsioonis
- siRNA – small interfering RNA, osaleb geeniregulatsioonis
- piRNA – piwi-interacting RNA, peamiseks funktsiooniks on transopsoonide kaitse
- SRP RNA – signaali äratundmis kompleksi RNA komponent . Funktsiooniks ko-translatsiooniline translokatsioon ja post-translatsiooniline transport.
- IncRNA – pikk mittekodeeriv RNA, reguleerib geenide transkriptsiooni ning osaleb epigeneetilises regulatsioonis
- TERC – telomeraasi RNA komponent
- Rnaaside RNA komponent
- scaRNA – väikesed cajal keha RNAd
miRNA – mikro RNAsid leidub eukarüootsetes rakkudes. miRNAd toimivad läbi RNA interferentsi, kus miRNA efektorkompleks ja ensüümid saavad seonduda komplementaarsele RNAle ja blokeerida mRNA transleerimist või kiirendada mRNA degradatsiooni
siRNA – väike interfereeriv RNA on lühike kaheahelaline RNA. Neid tekib nii viriaalsete RNAde lagundamisel kuid on ka endogeenseid siRNAde allikaid .
piRNA – osaleb transposoonide kaitsel
Antisenss-RNA – enamus surub maha geene, kuid mõned võivad olla transkriptsiooni aktivaatorid . Seondub mRNAle, moodustub dsRNA, mille lagundavad ensüümid.
CRISPR RNA – paljudel prokarüootidel, moodustavad RNA interferentsile sarnase süsteemi
lncRNA
- “ Molekulaarbioloogia keskne dogma”
- DNA ↔ RNA valk. Ehk DNAst transkribeeritakse RNA ja RNAst transleeritakse valku. Genoomist transkribeeritakse pre-mRNA, mida snoRNA ja snRNAde vahendusel splaissitakse ja protsessitakse, küpsest mRNA transleeritakse ribosoomide vahendusel valku, kus osaleb ka tRNA.
-
- lncRNAd (biogenees, funktsioonid)
- lncRNAd võivad rakendada oma funktsiooni seostudes DNA või RNAga järjestusspetsiifiliselt või seostudes valkudega. Osad lncRNAd on prekursoriks väiksematele regulatoorsetele RNAdele nagu nt miRNAdele või piRNAdele. LncRNAd võivad reguleerida geeniekspressiooni ja valgu sünteesi erinevatel viisidel . Reguleerivad geeniekspressiooni kromatiini tasemel, mõjutavad transkriptsiooni ja pre-mRNA protsessingut, mRNA stabiilsust ja translatsiooni. Geeniregulatsioon võib toimuda in cis – transkribeeritud lncRNA lähedal või in trans – transkriptsioonisaidist kaugemal. Posttranskriptsiooniline funktsioon hõlmab endas protsesse nagu splaissing , likalisatsioon, translatsioon ja degradatsioon .
Transkriptsiooni regulatsioon
- Reguleerivad geenispetsiifilist transkriptsiooni – ncRNAde märklaudadeks võivad olla aktivaatorid või repressorid, erinevad transkriptsiooni komponendid nt RNAP II ja siegi DNA. Moduleerivad transkriptsioonifaktorite funktsiooni erinevate mehhanismide kaudu, nt funktsioneerides koregulaatoritena, modifitseerides transkriptsioonifaktorite aktiivsust või reguleerides koregulaatorite aktiivsust.
- Reguleerivad transkriptsiooni masinavärki – märklauaks RNAP II vahendatud transkriptsiooniks vajalikud transkriptsioonifaktorid. Nt initsiatsioonikompleksi komponendid.
- RNAP III poolt transkribeeritud lncRNAd
Posttranskriptsiooniline regulatsioon
- Splaissimine
- translatsioon ja
- siRNA juhitud geeniregulatsioon – substraadiks endogeensetele siRNAde tekkele
Epigeneetiline regulatsioon –
- Imprinting
- Xist ja X-kromosoomi inaktivatsioon – X inaktivatsiooni viib läbi Xist lncRNA
- Telomeersed mittekodeerivad RNAd – lncRNA, mis moodustab telomeerse heterokromatiinse osa koos telomeersete valkudega.
-
- LncRNAde biogenees – transkriptsioon ja protsessimine on väga sarnane mRNAle. Enamus lncRNAsid transkribeeritakse RNA polümeraas II vahendusel, kuid osa lncRNAsid transkribeeritakse RNAP III vahendusel. Suur osa lncRNAdest splaissitakse, polüadenüleeritakse ja lisatakse 5’cap. lncRNA geenid asuvad eelistatult valku kodeerivates geenides ning võivad olla mRNA biogeneesi kõrvalsaaduseks.
- Regulatoorsed RNAd ja kromatiini modelleerimine
- lncRNAd võivad vahendada epigeneetilisi muutusi pannes tööle kromatiini remodelleerivad kompeksid spetsiifilistes lookustes. Osadel kromatiini remodelleerivatel kompleksidel on RNA seostumisdomeen kuid väike DNA järjestuse spetsiifilisus ning tänu lncRNAdele on nad võimelised spetsiifiliselt modifikatsioone läbi viima.
- ncRNAd nagu HOTAIR, Xist/RepA seostuvad kromatiini modifitseeriva polycombi kompleksiga, mis viib läbi metüleerimist HoxD lookuses või X kromosoomis. Indutseeritakse heterokromatiini moodustumine ja geeniekspressiooni repressioon.
- X-kromosoomi inaktiveerimine on vahendatud lncRNA poolt mida nim Xist. RepA paneb tööle PRC2, mis vaigistab ühe X-kromosoomi. Xist omab antisense partnerit Tsix. Tsix toimib aktiivsele X kromosoomile titreerides sellelt PRC2 kompleksi. Tsix hoiab ära PRC2-RepA laadimist kromatiinile ja blokeerib X kromosoomi inaktivatsiooni. Kui Tsix tootmine lõppeb, muutub PRC2 kompleks kättesaadavaks. PRC-RepA seondub kromatiniga ja PRC2 metüleerib tulevase inaktiveeritud X-kromosoomi. Jpx RNA tootmine ja Tsix sünteesi peatumine soodustab Xist RNA ekspressiooni. Xist seondub PRC2-ga. PRC2-Xist levib piki X kromosoomi ja metüleerib terve kromosoomi.
-
-
-
- HOTAIR RNA (HOX transkripti antisense RNA) vaigistab in trans HoxD lookuses transkriptsiooni indutseerides repressiivse kromatiini oleku, mis toimub polycombi kromatiini remodelleeriva kompleksi PRC2 tööle panemisega. HOTAIRi 5’ots interakteerub polycombi valguga PRC2 ning selle tulemusena reguleerib kromatiini olekut. HOTAIRi 3’ots interakteerub histooni demetülaasiga LSD1. Tulemusena toimub HOXD geenide epigeneetiline vaigistamine. Seda geeni ekspresseeritakse kõrgel tasemel kasvajates.
- Antisense RNA
-
- RNA interferents – liigid ja üldpõhimõtted
- miRNA – on vajalikud et geeniekspressiooni sihipäraselt reguleerida. Argonaudiga moodustub RISC kompleks. Neil on omad geenid, sünteesitakse RNAP II poolt. miRNAdel on osaline komplementaarsus märklaud-RNAga 3’mittetransleerivas osas. Dicer lõikab pre-miRNA lühikesteks RNAdeks. Lõikamise järgselt tekib dupleks , millest üks ahel on valmis miRNA ja teine temaga osaliselt komplementaarne vastasahel. Tagajärjeks on translatsiooniline repressioon või märklaud mRNA degradatsioon ja geeni vaigistamine.
- siRNA – Pakitakse samuti RISC kompleksi. siRNAd paarduvad sihtmärgiga täieliku komplementaarsuse alusel ja indutseerivad RNAi – märklaudgeeni lõikamist ja degradatsiooni. On väga levinud taimedes kus kaitsevad taime erinevate patogeenide eest. Tekivad pikkadest kaheahelalistest RNA lõikudest.
- piRNA – suuremad kui miRNAd ja siRNAd. Seonduvad PIWI perekonna valkudega, millede välja lülitamisel väljendub kordusjärjestuste kontrollimatu transkriptsioon. Esineb kahte tüüpi piRNAsid: varasemad pärinevad kordusjärjestustelt, hilisemad suurtest genoomsetest piRNA klastritest. Dicer ei osale piRNAde sünteesis. PIWI tüüpi valgud lõikavad märklaud-RNAd. piRNAd on vajalikud, et kaitsta genoomi mobiilsete elementide ja korduselementide plahvatusliku paljunemise eest.
- AGO valgud ja RISC kompleks
- Argonaut valgud jaotatakse kolme rühma:
Argonaut sarnased valgud, mis sarnanevad arabidopsis thaliana AGO1 valguga
Piwi sarnased valgud, mis sarnanevad D.melanogaster PIWI valguga
C.elegansi spetsiifilised grupp 3 argonaudid.
-
- miRNAd, siRNAd ja piRNAd pakitakse koos Argonaut perekonna valkudega RISC kompleksi. Argonaut on RISC kompleksi katalütiline komponent. Väikesed RNAd juhivad argonaudi märklauani tundes ära neil ekomplementaarse järjestuse. Argonaut sisaldab PIWI domeeni, mis viib läbi kaheahelalise RNA lõikamist. Argonaut sisaldab PAZ ja PIWI domeene – PAZ seondub küpse miRNA üheahelalise 3’otsaga, PIWI domeen interakteerub juhtahela 5’otsaga. Nad seovad küpse miRNA ja orienteerivad seda interaktsiooniks märklaud mRNAga.
-
- siRNAd (omadused, funktsioonid)
- siRNAd võivad olla eksogeensed kui endogeenset päritolu. Võõras RNA tuntakse ära, sünteeritakse komplementaarne ahel, Dicer seondub dupleksiga ja lõikab selle lühikesteks juppideks. Need fragmendid on sobilikud argonaut valkudele adaptoriteks. Endogeensete siRNAde tekkimine – vajalik 2ahelalise RNA olemasolu, et dicer saaks ära tunda selle.
- siRNAd pakitakse RISC kompleksi. siRNAd paarduvad sihtmärgiga täieliku komplementaarsuse alusel ja indutseerivad RNAi – märklaudgeeni lõikamist ja degradatsiooni. On väga levinud taimedes kus kaitsevad taime erinevate patogeenide eest. Tekivad pikkadest kaheahelalistest RNA lõikudest.
- piRNAd (omadused, funktsioonid)
- piRNA on suuremad kui miRNAd ja siRNAd. Seonduvad PIWI perekonna valkudega, millede välja lülitamisel väljendub kordusjärjestuste kontrollimatu transkriptsioon. Esineb kahte tüüpi piRNAsid: varasemad pärinevad kordusjärjestustelt, hilisemad suurtest genoomsetest piRNA klastritest. Dicer ei osale piRNAde sünteesis. PIWI tüüpi valgud lõikavad märklaud-RNAd. piRNAd on vajalikud, et kaitsta genoomi mobiilsete elementide ja korduselementide plahvatusliku paljunemise eest. Tekivad kordusjärjestusterikastes piirkondades genoomis, mis ei ole liikide vahel konserveerunud. Mingil hetkel algab selle aktiivne transkriptsioon ja protsessimine ja moodustavad ka argonaudiga RISC kompleksi. Valmis piRNAd võivad olla ise ka matriitsiks uutele piRNAdele. piRNAd juhivad piwi valgud märklaud-transposoonideni
-
- MikroRNAde biogenees
- miRNAdel on oma geenid, kust transkribeeritakse RNAP II vahendusel pri-miRNA. Primaarselt transkriptil pri-miRNAl 5’cap ja 3’ polyA . Pri-miRNA moodustab juuksenõelastruktuuri, kus ahel paardub iseendaga, moodustades lingu . Juuksenõelastruktuur lõigatakse tuumas paikeva DROSHA poolt. seejuures on abiks RNAd siduv valk DGCR8. Lõikamise tulemusel moodustub pre-miRNA ehk prekursor miRNA, mis transporditakse tuumast tsütoplasmasse valgu eksportiin-5 abil. Eksportiin-5 tunneb ära Drosha tekitatud 3’ üleuletuva otsa. Tsütoplasmas lõikab pre-miRNA juuksenõela Dicer ning vabaneb küps miRNA. Tekib miRNA-miRNA dupleks – üks ahel läheb lagundamisele, teine ühineb RISC kompleksiga. Küps miRNA on osa aktiivsest RISC kompleksist, mis sisaldab veel Dicerit ning mitmeid lisavalke. Argonaut sisaldab PAZ ja PIWI domeene – PAZ seondub küpse miRNA üheahelalise 3’otsaga, PIWI domeen interakteerub juhtahela 5’otsaga. Nad seovad küpse miRNA ja orienteerivad seda interaktsiooniks märklaud mRNAga.
- MikroRNAde geenid
- miRNAde geenid paiknevad genoomis kas:
- Eraldi geenidena
- Geeniklastritena ja transkribeeritakse polütsistroonse primaarse transkriptina mis lõigatakse mitmeteks miRNAdeks.
- Valke kodeerivate geenide intronites ja transkribeeritakse kodeeriva geeniga samalt ahelalt kui ka vastasahelalt. Kui miRNA on orienteeritud samas suunas kui ümbritsev geen, transkribeeritakse see juhuslikult koos peremeesgeeniga ja lõigatakse splaissimisel suuremast transkriptist, kus nad ennast peidavad.
- Või ka mittekodeerivate geenide eksonites
- miRNA transkriptsiooni viib läbi enamasti RNAP II, kuid harvemini ka RNAP III.
-
- MikroRNAde seondumiskohad – paiknemine, omadused, ennustamine
- Paiknevad praktiliselt alati valku kodeerivate geenide mittekodeerivates alades, 3’-UTR-des, võivad asuda ka nt kodeerivas alas , aga see on väga erandlik. Võivad paikneda ka mittekodeerivate geenide transkriptides, pseudogeenide või lncRNAde järjestustes. miRNAde seondumiskohad on peamiselt intron kodeeritavas alas, kuna degradatsioon takistab miRNAde seondumist mRNAl, sp on need seondumiskohad 3’UTR, kuna ribosoom ei läbi seda. Küpse miRNA järjestuse nukleotiidid 2-7 moodustavad seed regiooni, mis paarduvad komplementaarsuse alusel sihtmärk mRNA 3’UTR-ga ning selle piirkonnaga on määratud mRNAde seondumise spetsiifilisus. Kui seed ja 3’UTR vaheline seondumine on täielikult komplementaarne, toimub mRNA lagundamine, kui aga osaliselt komplementaarne, toimub mRNA translatsiooni inhibeerimine . Seondumiskohal on lisaks seed regioonist tagapoolt 3’otsa poole lisa järjestus, aga ei pruugi olla miRNAga seondunud.
- miRNAde seondumiskohad jagunevad seondumise tugevuse järgi:
- Kanoonilisteks kus seondumine on vajalik ainult seed regioonis
- 3’kompensatoorsed seondumiskohad kus seed regioonis võib olla nt GU paar ja mittetäielik paardumine , seda kompenseerib tugevam seondumine miRNA tagumises osas. Ehk kompenseerib seed mismatchi.
- Täielikult paardunud miRNA seondumiskohad – komplementaarsus kogu miRNA pikkuses . Tavaline taimedes, haruldane loomades , mRNA lõigatakse katki selle tulemusel.
- miRNAde seondumiskohtade ennustamine toimub põhiliselt seed regiooni järjestuse alusel mis peab olema täielikult komplementaarne miRNAga. Lisaks kasutavad algoritmid lisakriteeriume spetsiifilisemaks muutmiseks – lisakriteeriumid võivad hõlmata seondumiskoha järjestuse mustreid , mis soodustavad miRNA seondumist, erinevad sekundaarstruktuurid võivad mõjutada, kõige olulisemaks on konserveeritus lähedaste ja kaguete liikide vahel. Mida rohkem seondumiskoht on konserveerunud seda spetsiifilisemalt langeb see miRNA seondumiskohta ja seda parem on tõendusmaterjal et seondumiskoht on funktsionaalne.
- MikroRNAde toimemehhanismid
- miRNA toimib RISC kompleksis giidmolekulina, seondudes sihtmärk-mRNAle. Selle seondumise tulemusena toimub geeni ekspressiooni reguleerimine translatsiooni repressiooni või mRNA lagundamise kaudu. See millise mehhanismi kaudu regulatsioon toimub, sõltub miRNA seed järjestuse ja mRNA 3’UTR vahelise komplementaarsuse astmest . Täieliku komplementaarsuse korral toimub mRNA lagunamine, osalise komplementaarsuse puhul aga mRNA translatsiooni inhibeerimine. Taimedel on enamasti mRNA lagundamine, loomade puhul on lagundamine pigem harvem ning toimib enamasti repressioon. Translatsiooniline inhibeerimine võib toimuda:
- Elongatsiooni inhibeerimise kaudu
- Kotranslatsioonilise valgu degradeerimise kaudu – polüpeptiidahelat degradeeritakse kotranslatsiooniliselt
- Initsiatsiooni inhibeerimise kaudu:
- Argonaut valgud konkureerivad initsiatsioonifaktoriga eIF4E cap struktuurile seondumise pärast
- Argonaut paneb tööle initsiatsioonifaktori eIF6, mis hoiab ära ribosoomi suure subühiku seondumise väikese subühikuga
- Argonaut valgud takistavad mRNA suletud silmuse tekkimist ning põhjustavad mRNA deadenüleerimist (lagundamise alla kuulub)
- mRNA lagundamise puhul tingib miRNA seondumine sihtmärgi deadenüleerimise ja selle cap-struktuuri eemaldamise. See vähendab mRNA stabiilsust ja suurendab tema lagundamist eksonukleaaside poolt.
-
- MikroRNAde funktsioonid
- miRNAd mängivad olulist rolli transkriptsiooni järgses regulatsioonis. miRNAd reguleerivad mitmete geenide avaldumist posttranskriptsioonilisel tasemel. Nad seonduvad komplementaarsuse alusel mRNA 3’UTR järjestusega ja see võib viia mRNA lagundamiseni või translatsiooni represioonini. miRNAd on olulised koespetsiifilisuse määramisel – osad on aju spetsiifilised osad lihase spetsiifilised jne.
- miRNAd osalevad mitmetes bioloogilistes protsessides, nt metabolismis, rakkude jagunemises, diferentseerumises ja apoptoosis, organismi arengus, omavad ka mõju vähi tekkel jt. miRNAde ekspressioonimuster muutub vähi rakkudes ning neid saab kasutada vähi diagnoosimisel markeritena ning ka võimalike ravimisihtmärkidena. miRNAd võivad käituda nii tuumorsupressorina kui ka onkogeenina. miRNAde onkogeenne või tuumorsupressori funktsioon avaldub nende võimes reguleerida sihtmärke, mis on ise onkogeenid või tuumorsupressorid.
- miRNAd osalevad RNA interferentsis – mõjutavad geenide avaldumist. Nad seonduvad RISC kompleksiga, kus miRNA seondub mRNAga komplementaarsuse alusel.
- ceRNA hüpotees
- ceRNA ehk competing endogenous RNA – seovad miRNAsid ning seega derepresseerivad kõik sihtmärkgeenid, mille ekspressiooni miRNA oleks represseerinud. ceRNA hüpoteesi alusel erinevat tüüpi RNA transkriptid suhtlevad läbi miRNA response elemendi MRE. mRNAde ja miRNAde suhe võib olla retsiprookne, ühe mRNA tase mõjutab teise mRNA taset ja aktiivsust. Seega RNA molekulid võivad suhelda teineteisega läbi miRNAde ja MRE elementide. Mida suurem ja jagatud MREde tase, seda suurem on suhtlemise tase ja koregulatsioon. RNA molekulide 3’UTR-id sisaldavad MREsid, mis võivad funktsioneerida in cis, et ise reguleerida RNA molekuli või in trans, et reguleerida miRNAde ja teiste RNAde taset.
-
- CRISPR-Cas süsteem bakterites
- CRISPR-cas süsteem on bakteri immuunsüsteemi aluseks. See annab resistentsuse võõrastele geneetilistele elementidele nagu plasmiidid ja faagid ja annab omandatud immuunsuse. CRISPR-cas süsteem on eukarüootide RNAi analoog. Cas1 ja Cas2 tunnevad võõra DNA ära ja toimub protospeisseri lõikamine . Protospeisser ligeeritakse liiderjärjestusega külgnevale kordusele ja üheahelaline pikendamine parandab CRISPRi ja duplitseerib järjestust. crRNA protsessimine ja interferents toimub erinevaid teid pidi. Primaarset CRISPR transkripti lõigatakse cas geenide poolt et toota crRNAd. Küpsed crRNAd seonduvad cas valkudega ja moodustavad interferentsi kompleksi mille kaudu toimub sissetunginud DNA degradeerimine.
- RNP ehk ribonukleoproteiin on kombinatsioon RNAst ja valgust – RNA-valk kompleks. Nt ribosoomid, telomeraas, hnRNP-d ja snRNP-d. RNP-d on olulised geeniekspressioonil ja selle regulatsioonil. Peale transkriptsiooni seotakse pre-mRNAd erinevate valkude poolt, erinevas staadiumis mRNAga seotud erinevad valgud määravad mRNA saatuse . Pre-mRNA läbib protsessingu mida viivad läbi erinevad RNPd, nt U-rikas väike tuuma RNP (U snRNP), mis moodustavad splaissosoomi. Väikesed tuumakese RNPd (snoRNP) ja väikesed Cajal keha RNPd (scaRNPd) vahendavad RNPde (rRNA ja snRNA) komponentide küpsemist. Väikesed RNAd võivad moodustada mikroRNPd mille funktsiooniks on translatsiooni regulatsioon. Tsütoplasmas on peamiseks RNP-ks ribosoom, mis viib läbi translatsiooni. Ribosoomid assotsieeruvad ka signaali äratundvate partiklitega (SRP) RNP mis suunab valgud ERi. tRNAd moodustavad tsütoplasmas komplekse aminoatsüül-tRNA süntetaasidega, mis aminoatsüleerivad tRNAd.
-
-
-
-
-
-
-
- Genoomi varieeruvuse klassikalised allikad, mutatsioon ja rekombinatsioon :
- Somaatilised ja idurakkudes toimuvad DNA nukleotiidsed asendused: olulisus evolutsioonis; nende ‘saatust’ mõjutavad faktorid (indiviidi, populatsiooni ja evolutsiooni tasandil)
- Kui somaatilises rakus toimub asendus, mõjutab see ainult mutatsiooni kandva indiviidi fenotüüpi ning seega ei ole evolutsiooniliselt oluline. Kui aga idurakkudes toimub asendus, võib see päranduda ja levida edasi populatsioonis. Levik ja fikseerumine sõltub populatsiooni ajaloost, evolutsioonilistest jõududest nagu looduslik valik ja geenitriiv , mutatsiooni letaalsusest ning geenikonversioonist. Asendused võivad tekkida replikatsiooni või rekombinatsiooni vigade tõttu. Kui viga tekib kodeerivas alas, on suurem osa sellistest geneetilistest muutustest letaalsed. Harva võivad sellised asendused ka säilida ning fikseeruda uue variandina. Mittekodeerivates alades tekkivatel vigadel on väike efekt genoomi funktsioonile või genoomil pole vahel sellest asendusest ning tulemuseks on uus polümorfism .
-
-
- Mutatsiooni ja polümorfismi mõiste
- Mutatsioon on organismi kindlas kromosoomilookuses toimuv DNA muutus. Võivad olla nii punkt- kui ka kromosoommutatsioonid (kromosoomide struktuuri muutus).
- Polümorfism on kahe või enama geneetilise variandi olemasolu populatsiooni isendite hulgas, kusjuures vähemalt kahe variandi sagedus on suurem kui 1%.
-
- Negatiivne ja positiivne looduslik valik
- Negatiivne ehk puhastav valik eemaldab kahjulikke mutatsioone ja vähendab geneetilist varieeruvust populatsioonis. Ehk negatiivne valik liigub tunnuse kaotamise suunas.
- Positiivne valik on jõud mis juhib kasulike tunnuste leviku suurenemist . Positiivne valik viib uue tunnuse tekkeni, uue mutatsiooni kinnistumisele populatsioonis.
-
- Geenitriiv; positiivse valiku ja geenitriivi eristamine
- Geenitriiv on geeni alleelisageduste muutus väikese ristuva populatsiooni põlvkondades juhuslike kõrvalekallete tõttu.
- Geenitriivi näide: b on valge karvavärvus jänestel, B on pruun karvavärvus. Geenitriivi tagajärjel b-alleel kaob ning B-alleel triivib ainsa populatsiooni alleeli positsiooni suunas. Esimeses generatsioonis esinevad alleelid võrdsete sagedustega. Ainult pooled isenditest paarituvad ning teises generatsioonis on B alleelisagedus palju suurem kui b alleelisagedus. Teises generatsioonis paarduvad juhuslikult just need isendid kes on homosügootse dominantse genotüübiga ning pruunikarvavärvusega. Tulemuseks on kolmandas generatsioonis vaid pruunikarvaga dominantse homosügootse BB genotüübiga isendid.
- Esimesel pilgul võib see tunduda kui BB homosügootide positiivne valik, või vastupidiselt bb homosügootide negatiivne valik, kuid reaalsuses b-alleeli kadumine populatsioonis on siiski geenitriivi poolt põhjustatud. Nende kahe erinevus seisneb selles, et geenitriiv toimub juhuslikult, kuid valik toimub otsese vastusena keskonnatingimustele. Nt oleks positiivse valikuga tegemist kui kk tingimustest olenevalt on nt pruun karvavärvus eelistatum , nt rebased märkavad pigem valgeid ning pistavad need nahka ning pruun karvavärvus suurendab ellujäämist ja paljunemist ning rohkem pruune sünnib – nad on looduslikult valitud.
-
- Motoo Kimura neutraalsusteooria
- Enamus evolutsioonilisi muutusi on tingitud geneetilisest triivist, mis rakendub neutraalsetele alleelidele. Uus alleel tekib läbi spontaanse mutatsiooni.
-
- Mutatsioonikiirus (Mutation rate ) ja nukleotiidsete asenduste kiirus (substitution rate); nukleotiidsete asenduste kiiruse jaotuvus genoomis
- Mutatsioonikiirus on keskmine mutatsioonide esinemissagedus liigi kohta antud ajaühikus. Nt mtDNA oma kõrget mutatsioonikiirust. Aeglasem mutatsiooni kiirus näitab, et mutatsioon leidis aset kaugemas minevikus. Asenduste kiirus = mutatsioonikiiruse ja valiku kombinatsioon. Nukleotiidsete asenduste kiiruse jaotuvus genoomis ei ole juhuslik. Ehk asenduste kiirus on varieeruv üle genoomi. Asenduste kiirus vaikivate saitide eksonites on madalam kui intronites. Mutatsioonikiiruse hindamisel on oluline arvestada ka replikatsioonide arvu, mille lookus läbib. Nt isaste imetajate gameetide tekkel on mutatsioonid palju sagedasemad kui emastel, sest spermatogeneesi käigus toimub palju rohkem rakujagunemisi kui oogeneesis. Mutatsioonide absoluutarv sõltub lookuse pikkusest. Kõrge lokaalne varieeruvus autosoomides. CpG dinukleotiidides on suurem asenduste kiirus. Väiksem divergents telomeerides ja väiksemates kromosoomides. SNPide mutatsioonikiirus tõuseb insertsioonide/deletsioonide läheduses. Lühidalt – kromosoomi tasemel – Y kromosoom areneb kiiremini kui autosoomid, mis arenevad kiiremini kui X kromosoomid. See on tänu sellele et meestel on kõrgem mutatsioonikiirus kui naistel.
-
- Genoomse lookuse mutatatsioonikiiruse mõjutajad : CpG sisaldus, mutatsiooni hotspot ’id, indelite jaotuvus, kiiresti evolutsioneeruvad eksonid jne.
- GpC dinukleotiidide puhul on näidatud suurenenud mutatsioonikiirus. Kuna CpG dinukleotiide metüleeritakse ja metüültsütosiin on ebastabiilne viib see olukorrani kus C asendub A või T-ga suurema kiirusega kui mujal saitides. Mutatsiooni hotspotid on mutatsiooni-vastuvõtlikud regioonid. Ehk piirkonnad kus võib mutatsioone näha suurema sagedusega. On saidid kus esinevad SNPid nii inimestel kui šimpansitel – nagu konserveerunud või midagi sellist – piirkonnad kus on suurem mutatsioonikiirus.
- Samuti on näidatud, et mutatsioonikiirus on suurem insertsioonide ja deletsioonide läheduses. Indelid tõstavad mutatsioonikiirust kuid mitte spetsiifilistes saitides, pigem mutatsioonikiirus on suurem indeli läheduses ja see mutatsioonikiirus langeb peale mitmeid sadu nukleotiide .
- Kiiresti evolutsioneeruvad eksonid sisaldavad AT-to-GC asenduste klastreid. Ehk AT muteerumine GC-ks – toimub GC sisalduse suurenemine. GC alleelide fikseerumist juhib kallutatud geenikonversioon.
-
- Duonid
- Duon on topelt tähendusega koodon genoomis. Duonid on koodonid , millele seostuvad transkriptsioonifaktorid ning mis lisaks kodeerivad valgu kooseisu minevaid aminohappeid.
- Osadel koodonitel – duonitel – võib olla kaks tähendust, üks seotud valgu järjestusega ja teine seotud geeniregulatsiooniga. 15% inimese koodonitest on dual -use koodonit ehk duonid. Duonid määratlevad samaaegselt aminohappeid ja transkriptsioonifaktorite äratundmissaite. Duonid on väga konserveerunud ja on kujundanud valkude evolutsiooni. Duonite puhul aminohapete jaoks alternatiivsete koodonite kasutamine ei ole juhuslik ja kallutatud. Koodonkasutus sõltub parima sobivusega transkriptsioonifaktorit siduvast motiivist.
- Meioosi ‘eesmärgid’ võrreldes mitoosiga
- Meioosi eesmärgiks on segada ja kokkupanna kahe vahema genoomi variandid ja toota varieeruvust järgjaste hulgas. Mitoosi eesmärgiks on toota 2 tütarrakku, millel on täpselt sama genoom kui vanemrakul.
-
- Rekombinatsioon: meiootiline ristsiire ja geenikonversioon, mehhanismid, tagajärjed; geenikonversiooni toimumise eeldused ja ulatus.
- Ristsiire on peamiseks varieeruvuse genereerimise mehhanismiks populatsioonis indiviidide vahel. rekombinatsiooni tulemusena toimub meioosis homoloogiliste kromosoomide vahel ristsiire, mille tulemusena tekivad kas vanemlike kromosoomidega sarnased või uued rekombinatsed DNA molekulid. Rekombinatsiooni toimumise eeltingimuseks on kaheahelalise katke teke, mis initsieerib DSB paranduse raja, mille käigus toimub homoloogiliste DNA ahelate vahel ristsiire ning moodustuvad tütarkromosoomid. Mehhanism : homoloogilised kromosoomid paarduvad. Homoloogsete segmentide vahel toimub sünaps . Paardunud osade vahele moodustub valguline struktuur – sünaptonemaalne kompleks, mis hoiab kromosoome koos. Meioosi I profaasi pahhüteenis toimub ristsiire. Homoloogid eemalduvad veidi teineteisest, kuid jäävad siiski veel seotuks nendest kohtadest , kus on toimunud ristsiire. Ristsiirede kohad ehk kiasmid hoiavad kromosoome koos kuni anafaasini, täidavad sama funktsiooni nagu tsentromeerid mitoosis.
- Geenikonversioon on nähtus, kus kahe kõrgelt homoloogse järjestuse vahel (nt duplitseerunud geenid) toimub geneetilise info vahetamine ühesuunaliselt ilma ristsiirdeta. Ehk lühidalt vahetatakse aktseptor segment doonorsegmendi vastu. Geenikonversioon võib toimuda koos või ilma ristsiirdeta. Mehhanism – moodustub kaheahelaline katke DSB, millele järgneb 5’ otste lühenemine. Seejärel toimub ahela invasioon ning DNA süntees. Edasi võib minna kahte erinevat radapidi: geenikonversioon ristsiirdeta – ahela seondumine, süntees ja ligeerimine, mismatch parandus ning tagajärjeks konverteerunud ala. Geenikonversioon ristsiirdega – toimub Holliday struktuuri lahendamine, mismatch parandus ning ristsiire ja geenikonversioon. Konverteeritava ala pikkus on keskmiselt 300 bp. Lühidalt – rekombinatsiooniline protsess, kus üks alleel replitseeritakse teise alleeli järjestuse põhjal. Esineb nii kromosoomidevahelist kui ka kromosoomisisest geenikonversiooni.
-
- Rekombinatsiooni eeldus: DNA kaksikahelaline katke (DSB) ja selle mehhanismi konserveeritus
- Rekombinatsiooni toimumise eelduseks on kaheahelise katke teke Spo11 poolt, mis initsieerib DSB reparatsiooni raja. Ehk ühel homoloogsetest kromosoomidest tekib kaheahelaline katke millele järgneb 5’ otsade lühenemine. Seejärel toimub invasioon, kus üleulatuv 3’ots suundub homoloogilise DNA osani kus ei ole toimunud katkestust. Tekivad X-kujulised Holliday struktuurid, mis ühendavad kahte DNA molekuli. Toimub ristsiire. On väga konserveerunud mitmetes organismides nagu faagid, bakterid , pärmid ja inimesed.
-
- Rekombinatsiooni kaks lahendust ja nende omavaheline proportsioon: DSBR, SDSA
- DSBR – double - strand break repair – ehk kaheahelalise katke reparatsiooni rada. Alustuseks toimub kaheahelaline katkestus Spo11 poolt, millele järgneb 5’otste lõikamine. Seejärel toimub ahela invasioon, kus üleulatuv 3’ots tungib homoloogilise DNA osa vahele kus ei ole toimunud katkestust. Tekivad X kujulised Holliday struktuurid, mis ühendavad kahte DNA molekuli. Toimub mismatch parandus. DSBR mudeli järgi võivad tekkida nii ristsiire kui konversioon. Toimub 10% sagedusega.
- SDSA – synthesis -dependent strand annealing – ehk süntees-sõltuva ahela seondumine. Esimesed sammud on samad kui DSBR puhulgi: DSB, 5’otste lõikamine, ahela invasioon. Toimub 3’üleulatuva ahela paardumine homoloogilise järjestusega – invasioon. Siin ei teki Holliday struktuuri, moodustub D-loop. Toimub sissetunginud 3’otsa pikendamine piki homoloogset DNA dupleksit DNA polümeraasi vahendusel, nii et toimub D-loop’i translokatsioon. Nagu edasi liikuv mull. Toimub ligeerimine ja mismatch parandus ning tulemuseks on ainult geenikonversioon. Toimub 90% sagedusega.
-
- Rekombinatsioonipunktide üldisem lokalisatsioonimuster genoomis: pärm ja kõrgemad eukarüoodid ;
- Rekombinatsioonipunktid EI OLE kromosoomides ühtlaselt jaotunud, vaid on pigem klasterdunud spetsiifilistesse piirkondadesse, mida nimetatakse rekombinatsiooni hotspotideks. Rekombinatsiooni toimumiseks on vaja avatud kromatiini struktuuri ja kaheahelaliste katkete DSB initsiatsiooni. Tavaliselt esinevat DSB-d peamiselt kromatiini kergesti ligipääsetavates regioonides. Pärmis on kolme tüüpi hotspote:
- α – korreleerub transkriptsioonifaktorite seostumissaitidega ehk eeldab transkriptsioonifaktori seondumist.
- β – sisaldab nukleosoomidest vabat DNA järjestust ehk assotseerub nukleaasitundliku kromatiiniga
- γ – on kõrge C+G nukleotiide koostisega, kõrge GC %
- Pärmis rekombinatsiooni ja transkriptsiooni initsiatsioon on seotud 12-ne (CCGNN) tandeemse kordusega. See on väga tundlik DNaas I-le. Ainult transkriptsioon on seotud 48 CCGNN kordusega.
- Kõrgemate eukarüootide puhul on pakutud , et rekombinatsiooni toimumist tõstab kõrgem GC sisaldus, kõrgem Alu elementide ja geeni sisaldus. χ-motiiv stimuleerib rekombinatsioonilist aktiivsest. Samuti palindroomsed järjestused, mis soodustavad sekundaarstruktuuride moodustamist stimuleerivad DSB teket – loop osa on tundlik endonukleaasidele ja seega toimub selles piirkonnas kromatiini lõikamine millele järgneb rekombinatsiooniline aktiivsus.
- Alternatiivsed rekominatsiooni hotspotide mudelid imetajates:
- Cis elementide mudel – hiirtes NID1 hotspotis CCCCACCCC põhjustab rekombinatsiooni. Inimestes CCTCCCT on rikastatud hotspotides.
- Trans elementide mudel – hotspot aktiivsus on reguleeritud PRDM9 poolt
- DNA omaduste mudel – hotspoti aktiivsus peegeldub selle DNA- sidumise omaduses.
- Sekundaarstruktuuri mudel – palindroomsed järjestused indutseerivad DSB-sid – vt üles.
-
- Rekombinatsiooni ‘hotspot-id’: ulatus, piirid, jaotuvus genoomis ja indiviiditi, nende kaardistamise metoodilised võimalused ( otsesed ja kaudsed ); seos ‘ linkage disequilibrium’ ehk LD kaartidega; evolutsiooniline konserveeritus läbi liikide
- Hotspot on 1-3 kb pikkune kromosoomi lõik, kus toimub eriti palju ristsiirdeid. Ristsiirded ei ole jaotunud normaaljaotusega ümber hotspoti keskpunkti . Kõige esimeseks avastatud hotspotiks oli MHC kompleks hiire kromosoomis 17. Nt inimese MHC klass II regioonis on kuus rekombinatsiooni hotspoti. Rekombinatsioonisündmused erinevad meeste ja naiste vahel. X-kromosoomid on vähem kompaktsed ning seega pikemad, seega toimub neis suuremal hulgal rekombinatsioone kui Y-kromosoomis. Naistel on keskmiselt 25% rohkem kiasme kui meestel. Hotspotid on lühikesed genoomsed piirkonnad ning neil on selgesti eristatavad piirid.
- Rekombinatsioni aktiivsus on erinev indiviiditi.
- Kaardistamine:
- Analüüsides meioosi kasutades spermatosoidi DNA-d
- Detekteerides rekombinatsioonil osalevaid valke – nt SPO11, et tuvastada katkete kohti
- Analüüsides meioosi suurtes inimeste või hiirte sugupuudes
- Kasutades LD-blokkide ja populatsiooni variatsiooni andmeid
- Seos LD kaartidega – rekombinatsiooni hotspoti esinemist on võimalik hinnata kaudselt LD-analüüsi kaudu. Kuna LD-blokkide piirid kattuvad suuresti ajalooliste rekombinatsiooni sündmustega, siis võib alleelse aheldatuse mustrit kasutada vihjena hotspotide esinemise kohta genoomis.
- Rekombinatsiooni hotspotid ei ole konserveerunud liikide vahel ja ei ole kindlaksmääratud DNA primaarjärjestusega. Nt inimese ja šimpansi vahet on ainult 8% ühiseid hotspote. Kahe hotspoti vaheline ala on tugevas LD-s ja see moodustab ühe bloki.
- Inimestel on LD mustrite alusel identifitseeritud üle 30 000 hotspoti, mis enamasti asuvad väljaspool geene.
-
- PRDM9 kui rekombinatsiooni ‘hotspot-ide’ määraja; PRDM9 omadused, mis selgitavad liigispetsiifiliste rekombinatsiooni ‘hotspott-ide’ olemasolu; PRDM9 avastamise revolutsioonilisus
- PRDM9 ekspresseeritakse spetsiifiliselt varases meioosis nii naiste kui meeste puhul. PRDM9 puudumine põhjustab steriilsust mõlemates sugudes.
- Inimestel on 40 PRDM9 alleeli. Need erinevad alleelsed variandid tunnevad ära spetsiifilise hotspot motiivi. PRDM9 on on oluline hotspotide määraja. Ühine kromatiini omadus, mis kirjeldab kaheahelaliste katkete initsiatsioonisaite pärmis ja hiires on H3 neljanda lüsiini trimetülatsioon. Selles protsessis osaleb PRDM9 ning ta ekspresseerub vaid meioosis varastes etappides. PRDM9 tsingi sõrmed vahendavad DNA seondumise spetsiifikat.
- PRDM9 tsingi sõrmed seonduvad in vitro DNA motiividega, mis paiknevad hotspotide keskpaikades.
- DNAga järjestussõltuvalt kontakteeruvad tsingi sõrmed arenevad kiiresti ning on positiivse valiku all, mis viib rekombinatsiooni hotspotide paiknemise erinevusteni, polümorfne liikide vahel. PRDM9 seondumiskoht inimeses on täiesti erinev nt šimpansi PRDM9 seondumiskohast. Selle erinevuse põhjuseks on PRDM9 kodeeriva järjestuse kõrge mutatsioonikiirus – selles geenis toimub kiirem evolutsioon . Seletab mittekonserveerumist liikide vahel.
-
- Genoomi epigeneetiline komponent:
- Epigeneetika /epigenoomika mõiste; epigeneetilised mehhanismid
- Epigeneetika on teadus mis uurib rakulisi ja füsioloogilisi tunnuseid, mis päranduvad tütarrakkude kaudu, kuid ei ole seotud muutustega DNA enda nukleotiidses järjestuses . Funktsionaalsete geenide aktiveerimine ja inaktiveerimine. Epigeneetilised mehanismid on:
- DNA modifitseerimine (nt CpG metülatsioon),
- histoonide posttranslatsiooniline modifitseerimine,
- väikeste RNAdega seonduv vaigistamine
- kromatiini remodelleerimine – histoonide paigutamine piki DNAd ATP sõltuva nukleosoomi remodelleeriva kompleksi vahendusel. Kasutab ATP hüdrolüüsi energiat histoonide oktameeri ümberpaigutamiseks ja nukleosoomi-vaba kromatiini moodustamiseks.
- Epigenoomika on raku geneetilise materjali epigeneetiliste modifikatsioonide täieliku komplekti, mida nimetatakse epigenoomiks, uuring.
-
- DNA metülatsioon: levik ja metülatsioonimuster eukarüootsetes genoomides (mosaiikne, globaalne; geenikehad, kordus-elemendid jne); metüleeritava C-nukleotiidi kontekst; roll(id) genoomi(de) funktsioonis; CpG-saared
- Kõrge DNA metüleerituse tase iseloomustab transkriptsiooiliselt inaktiivset kromatiini. Samas transkriptsiooniliselt aktiivne kromatiin on praktiliselt metüleerimata, iseloomustab kõrge atsetüleerituse tase. Selgrootutes on peamiselt mosaiikne metülatsioonimuster, mis sisaldab kõrgelt metüleeritud DNA domeene, mis on eraldatud sama suurte mittemetüleeritud DNA domeenide poolt. stabiilsed metüleeritud ja mittemetüleeritud domeenid on segamini . Arabidopsisel on samuti mosaiikne metülatsioonimuster, mis on tekib tänu geenikeha metüleerimisele nagu selgrootuteski. Geenikeha metülatsioon mõjutab üle poolte kõikidest geenidest ja ülejäänud on varjul mittemetüleeritud DNAs. Transposoonid on sagedasti metüleerimata. Mosaiikse metülatsiooni puhul on metüleeritud ja mittemetüleeritud domeenide vahekord peaaegu võrdne. Selgroogsetel esineb globaalne DNA metülatsioon – metülatsioon on hajutatud üle genoomi, neil on kõige kõrgem m5C tase. Ainult CpG saared on metüleerimata. Geenide vahelised alad, transposoonid ja geenikehad metüleeritakse. Imetajate genoomis domineerivab metüleeritud DNA. Mittemetüleeritud DNA on kontsentreerunud CpG saartesse.
- Geenikehade metüleerimine – suur osa geenidest sisaldavad CpG metülatsiooni transkribeeritavates regioonides. DNA metülatsiooni ei esine geenide 5’ ega 3’ otstes . Geenidel mille geenikehad on metüleeritud esineb kõrgem ekspressioonitase kui nendel mille promootor on metüleeritud või terve geen metüleerimata. Geenisisene metüleerimine takistab transkriptsioonile vahelesegamist. DNA metüleerimine kaitseb genoomi transposoonide eest.
- CpG metülatsioonitase on erinev kudede ja indiviidide vahel populatsioonis. Erinev metüleerimine tagab erineva geeniekspressiooni. Vt järgmine C-nuk kontekst.
-
- DNA metüültransferaasid (DNMT); DNA demetülatsiooni võimalused
- DNMT1 metüleerib hemi-metüleeritud DNA, tagab replitseeritud tütarahela sama metülatsioonimustri kui vanemahelalgi. Represseerib transkriptsiooni kompleksis histoonide deatsetülaasidega.
- DNMT3a, DNMT3b – de novo metülaasid. Lisavad metüülrühma metüleerimata CpG dinukleotiididele, tulemuseks on uus hemi-metüleeritud ja siis täielikult metüleeritud CpG. Toimib piirkondades kus on vaja geeniekspressioon maha suruda. DNMTd katalüüsivad metüülrühma viimist S-adenüülmetioniinilt (SAM) tsütosiini jäägi viiendale süsinikule, et moodustada 5-metüültsütosiin 5mC.
- Demetülatsiooni võimalused – DNA demetülatsioon võib olla kas passiivne või aktiivne. Passiivne demetülatsioon toimib äsja sünteesitud DNA ahelale metülatsiooni puudumisel. Aktiivne DNA metülatsioon toimub läbi metüülrühma otse eemaldamise, DNA replikatsioonist olenemata. Imetajates toimub globaalne ehk ülegenoomne demetülatsioon. Esineb ka lookus-spetsiifilist demetüleerimist – nt genoomne imprinting jne. Aktiivne demetülatsioon võib toimuda läbi otsese 5mC eemaldamise või edasi modifitseeritud tsütosiini läbi. Otsene eemaldamine on nt aluspaari eemaldamine ja mittemetüleeritud aluse inserteerimine base excision repair mehanismi kaudu (BER). Või eemaldatakse terve DNA lõik mis täidetakse nukleotiididega nukleotide excision repair (NER) poolt. Edasi modifitseeritud tsütosiini puhul tekib oksüdatsiooni käigus hüdroksümetüültsütosiin, mis deaminaaside poolt deamineeritakse, nt AID/Apobec, TET. Deaminatsiooni ja oksüdatsiooni produktid parandatakse TDG glükosülaasi poolt mis on seotud BER mehanismiga.
-
- DNA hüdroksümetülatsioon: levik genoomides (mida tänaseks teada), roll(id) genoomide) funktsioonis.
- TET – ten- eleven translocation ensüüm oksüdeerib 5-metüültsütosiini 5-hüdroksümetüültsütosiiniks. Hüpoteesi järgi on 5hmC intermediaat 5mC eemaldamisel . TET üleekspressioon vähendab 5hC taset ja TET kaotamine viib 5mC akumuleerumisele. 5hmC geenikehades viitab aktiivsele geeniekspressioonile. 5-hmC eksisteerib peamiselt CG kontekstis. 5hmC on rikastatud aktiivselt transkribeeritavate geenide geenikehades ning 5mC on vastupidiselt seotud negatiivselt geeniekspressiooniga.
-
- Ten-eleven translocation (TET) ensüümid; nende avastamise revolutsioonilisus
- TET ensüümid viivad läbi 5-metüültsütosiini oksüdatsiooni ning tekib 5-hüdroksümetüültsütosiin. 5-hmC on intermediaat 5mC eemaldamisel. TET üleekspressioon vähendab 5hC taset ja TET kaotamine viib 5mC akumuleerumisele. 5hmC geenikehades viitab aktiivsele geeniekspressioonile. TET ensüüme kirjeldati juba 1972 roti, hiire ja konna ajus, seejärel unustati ning avastati uuesti 2009 aastal 5hmC tuvastamisel ESC-des. TET1 on seotud pluripotentsusega, TET3 ekspressioon on kõige kõrgem ootsüütides ja sügootides.
-
- Kõige enam levinud, bisultit-töötlusel põhinevate DNA metülatsiooni tuvastamise metoodikate oluline puudus
- Võimaldab eristada aluspaari tasemel mittemodifitseeritud ja modifitseeritud tsütosiinide vahel, kuid ei erista 5-metüültsütosiini 5-metüülhdroksütsütosiinist. Selle meetodi puhul mittemetüleeritud tsütosiin deamineeritakse U-ks, mis peale PCR reaktsiooni annab T, kuid 5-mC reageerib halvasti bisulfitiga ja annab C. 5-hmC annab 5-metüleensulfonaadi (CMS)
-
- DNA metülatsiooni loomulik ja patoloogiline dünaamika: epigeneetiliste signaalide kopeerimine replikatsioonis, epigeneetilise profiili stabiilsus vananemisel; vananedes ja epigeneetilise profiili seos haigustega; polümorfne ehk indiviidide genoomide DNA metülatsioonitaseteme vaheline erinevus ja selle põhjused – nii DNA järjestusest sõltuv ja kui sõltumatu.
-
-
- Epigeneetilise profiili kõrvaldamine ja reprogrammeerimine põlvkondade vahel
- Embrüonaalse arengu käigus idurakud demetüleeritakse, mis kustutab vanemspetsiifilise metülatsiooni märgised. Peale viljastumist, isapoolne genoom demetüleeritakse ja emapoolne genoom läbib passiivse demetülatsioon. Ülegenoomne remetülatsioon toimub mõlemas vanemgenoomis ennem implantatsiooni. Imprinditud geenid säilitavad oma metülatsiooni märgiseid läbi kogu reprogrammeerimise lubades vanema spetsiifilist ekspressiooni pärinemist somaatilistes kudedes.
-
- Geenidoosi kompensatsioon epigeneetiliste mehhanismide abil: X-inaktivatsioon.
- Geenidoosi kompensatsioon tagab X-liiteliste geenide võrdväärse avaldumise emastel ja isastel – emaste üks X kromosoom inaktiveeritakse ning tekib Barri kehake . X kromosoomi inaktivatsioon toimub läbi CpG metülatsiooni ja histoonide modifikatsioonide, need viivad kromatiini kondenseerumiseni, inaktivatsioon on juhuslik (kõik emased mosaiiksed). X inaktivatsioon toimub Xist transkripti kaudu, mis on lcnRNA – Xist katab terve inaktiveeritud X-kromosoomi.
-
- Geenide vanema-spetsiifiline ekspressioon ja vaigistamine (imprinting): levimine organismides, mõistete sisu ja mehhanismid.
- Imprintingu korral ekspresseerub vaid ema- või isapoolne geen. Teise vanema geen on inaktiveeritud. Imetajates ekspresseeruvad imprinditud geenid embrüogeneesi kestel ja platsentas. Imprinditud geenid moodustavad ko-reguleeritavad geeniklastrid. Väikesed mittekodeerivad RNAd lncRNAd takistavad ühe ahela ekspressiooni.
-
- Tüüpilisemad histoonide modifikatsioonid (atsetüleerimine, metüleerimine) ja nn. ‘histooni kood’ mõiste; teada mõned näited nn. ‘aktiivsele’ ja ‘inaktiivsele’ kromatiinile tüüpilis(t)est histoonide modifikatsioonidest (e.g. kui oleks oma uurimistöös vaja detekteerida ‘aktiivset’ ja ‘inaktiivset’ kromatiini, millised modifikatsioonid valida)
- Histoonide N terminaalsed sabad jäävad nukleosoomist väljapoole ning nendes nendes piirkondades toimub histoonide posttranslatsiooniline modifitserimine. Histoonide N- term sabad sisaldavad palju lüsiini ja arginiiinijääke. Lüsiinijäägid atsetüleeritakse või metüleeritakse, arginiinijäägid metüleeritakse. Histoonide atsetüleerimine on kõrge transkriptsiooniliselt aktiivses kromatiinis ja kõrgelt metüleeritud inaktiivses kromatiinis. Atsetüleerimist viivad läbi histoonide atsetüültransferaasid (HATid) ning atsetüleerimise tulemusena eemaldatakse lüsiinijäägilt positiivne laeng mistõttu nõrgenevad interaktsiooni DNA ja histoonide vahel ning samuti nõrgenevad naabernukleosoomide omavahelised interaktsioonid. Tekitatakse seondumiskoht bromodomeeni sisaldavatele valkudele. Lüsiini ja arginiinijääkide metüleerimisi viivad läbi histoonide metüültransferaasid. Metüleeritud lüsiinijääkidega seonduvad kromodomeeni sisaldavad valgud.
- Histoonide kood – histoonide modifikatsioonid moodustavad omalaadse mustri, mis on iseloomulik ühele või teisele kromatiini olekule (heterokromatiinile, aktiivselt transkribeeritavale kromatiinile jne).
-
-
-
- Genoomi korduselemendid ja roll genoomi evolutsioonis ja funktsioonis:
- Lihtsad kordusjärjestused ‘ Satelliit ’-DNA, ‘minisatelliit’- ja ‘mikrosatelliit’- kordused: jaotuvus genoomis; ülesanded (kui on) genoomis; evolutsiooniline konserveeritus; mutageensus ja vead replikatsioonil/rekombinatsioonil; olulisimad kasutamisvõimalused genoomi uuringutes ja geneetika -uuringute rakendundustes.
- Satelliit-DNA – suurtest tandeemselt korduva mittekodeeriva DNA blokkidest koosnev järjestus. Paikneb enamasti kromosoomi telomeerses või tsentromeerses piirkonnas. Moodustab heterokromatiini struktuuri põhi koostisosa . Stimuleerib ja säilitab epigeneetilisi heterokromatiniseerimise signaali nt H3K9 deatsetülerimine ja metüleerimine ning vastutab funktsionaalse tsentromeeri eest. Ei ole evolutsiooniliselt konserveerunud.
-
- Minisatelliit kordused – koosnevad keskmise suuruseda tandeemselt korduva mittekodeeriva DNA blokkidest. Esinevad telomeeride läheduses. Sageda muteerumisega inimestes, kõrge polümorfsusega. On rekombinatsiooni hotspotid. Kasutatakse geneetiliste markeritena inimeste tuvastamisel – DNA fingerprinting.
-
- Mikrosatelliitkordused – koosnevad väga väikese suurusega blokkidest tandeemselt korduvast mittekodeerivast DNAst. 1-4 bp lihtsad kordused. Esinevad hajusalt üle genoomi moodustades u 2% genoomist. Tekivad replikatsiooni libisemise tõttu. On mutatsioonilised hotspotid. Kasutatakse ülegenoomsete markeritena loomade ja taimede genoomide kaardistamisel. Inimese triplett kordused suurendavad haiguste esinemist – nt fragiilse X-sündroom, FMR1 geenis pikad CGG kordused. Või CAG Huntingtoni tõve korral. Vt järgmine.
- Nn. korduste ebastabiilsus (‘ Repeat -instability’); trinukleotiid-mikrosatelliit- kordustega seostatud haigused, (i) nn. polyQ (CAGn) või muude kodeeriva ala trinukleotiid-korduste ekspansioon (mõjutades transleeritavat valgustruktuuri) ja (ii) nn. mitte-polyQ haigused, mil trinukleotiid-korduste ekspansioonis mõjutab tranksriptsiooni, replikatsiooni, rekombinatsiooni, DNA parandamist
- CAG pikad kordused HTT geenis on seotud Huntingtoni tõvega. Mutatsiooni tulemusena tekib huntingtoni valgus polüglutamiinahela pikenemine . Mitte-polyQ haiguseks on nt fragiilse X-sündroom, kus pikeneb CGG kordus FMR1 geenis. Selle sündroomi puhul on FMR1 promootor hüpermetüleeritud mis suuresti vähendab FMRP valgu sünteesi. FMRP on oluline sünapside arengul.
-
- Genoomi transponeerivad elemendid, nende tüüpide võrdlus; osakaal genoomist.
- Transponeerivad elemendid on DNA järjestused, mis suudavad ühe raku piires genoomis ümber paigutuda ehk transponeeruda. Jagunevad retrotransposoonideks ja DNA transposoonideks. DNA transposoonid moodustavad u 3% inimese genoomist. Töötavad cut and paste mehhanismil, ehk ühest piirkonnast lõigatakse välja ja liiguvad teise piirkonda. Retrotransposoonid töötavad copy and paste mehanismil, kus pöördtranskribeeritakse RNA ja selle cDNA koopia inserteerub uude saiti genoomis. Esmalt transkribeeritakse DNAlt RNA, saadud vahevormi kasutatakse koopiaaru suurendamiseks . Seejärel moodustatakse pöördtranskriptaasiga RNAst tagasi DNA ning see siseneb uude positsiooni genoomis. Retrotransposoone on kahe tüüpi: pikkade otsmiste kordusjärjestustega (LTR) ja LTR puuduvad. LTR puudulikud jaotatakse LINEdeks ehk pikkadeks genoomi sisenenud elementideks ja SINEdeks ehk lühikesteks genoomi sisenenud elementideks. LINEsid umbes 17% ja SINEsid 11%. LINEd kodeerivad ise oma pöördtranskriptaasi, kuid SINEde transpositsioon sõltub teistest mobiilsetest elementidest.
-
- Genoomi transposon-elementide kompositsioon inimese genoomi näitel, rõhuasetusega retrotransposonidel Alu (SINE) ja LINE (L1) elementidel: päritolu, suurus, struktuur, omadused, jaotuvus genoomis, retropositsioneerumise mehhanismid, Alu-de roll primaatide evolutsioonis.
- U 45% inimese genoomist on transposoon-elementidega seotud. Suurem osa neist on mitteLTR retrotransposoonid: LINE1 (L1) ja SINE (Alu). L1 element koosneb kahest avatud lugemisraamist, mis külgnevad 5’ ja 3’ UTRdega. 5’UTR sisaldab RNA pol II promootorit. Alu element sisaldab kahte monomeeri, mis on eraldatud A-rikka linkerregiooniga. Üks monomeer RNA pol III promootorit. Transponeeruvad elementide jaotuvus üle genoomi ei ole juhuslik. SINEd esinevad eukromatiinis, geenirikastes R-vöötides ja LINEd paiknevad G-vöötides (AT rikkad alad, geenivaesed). SINEde korduste arv on väiksem kui LINEdel. SINEd ei ole viiruslikku päritolu, nad ei kodeeri oma pöördtranskriptaasi, on rekombinatsiooni hotspotideks. LINEd on viiruslikku päritolu. Kodeerivad oma pöördtranskriptaasi, pol II transkriptid. Mõlemad kasutavad copy paste mehanismi.
- Evolutsioon – inimgenoom on suurem ning sisaldab rohkem kordusi kui ükski teine looma genoom. Retrotransposoon-kordused on väga erinevad erinevate primaatide liinides. Need erinevused on viinud 15-20% suurema inimgenoomini viimase 50 milj aasta jooksul primaatide evolutsioonis, 90% sellest on tänu uutele retrotransposoonide insertsioonidele.
-
- Retrotransponeerumise erinevad mõjud genoomi struktuurile, stabiilsusele, evolutsioonile (e.g. Alu-de eksoniseerumine) ja funktsioonile, geeni ekspressioonile jne.
- Struktuur ja stabiilsus - võivad põhjustada mutageneesi inserteerudes kodeerivasse või regulatoorsesse piirkonda. Võivad tekitada ja parandada DNA kaheahelalisi katkeid. Põhjustada mikrosatelliitide tekkimist. Geenikonversiooni, moodustades homoloogse transponeeruva-elemendi koopia. Võivad põhjustada deletsioone insertseerudes. Tekitavad aktiivselt rekombinatsioone, mis on aluseks segmentaalsete duplikatsioonide ekspansioonile. Samuti transduktsioone.
- Geeniekspressioon – võivad tekitada insertsiooniga alternatiivseid transkriptsiooni algussaite, võivad katkestada promootori, sisestada antisense promootori jne. Transkripti tasemel võivad sisestada alternatiivse polüA saidi, miRNA sidumissaidi, tekitada alternatiivset splaissingut introni säilitamise või eksoni skippingu’ga.
- Alu-de eksoniseerumine – introonsed Alu-elemendid võivad tekitada mutatsiooni, mis tekitavad funktsionaalseid splaissimise saite. Võivad konverteeruda eksonitesse.
-
- Genoomi evolutsiooniline dünaamika, võrdlev genoomika :
- Genoomi funktsionaalse diversiteedi allikad
- Uute valgu domeenide ja struktuuride tekkimine
- Olemasolevate valgu domeenide ja valgu perekondade laienemine ja divergents
- Horisontaalne geeniülekanne
- Alternatiivne splaissing
- RNA editing
- Posttranskriptsioonilised modifikatsioonid
- Regulatoorsed võrgustikud
-
- Susumu Ohno: DNA duplikatsioon kui põhimehhanism genoomi evolutsioonis; seda teooriat toetavad teadmised
- Ohno pakkus välja, et geenide duplikatsioon on peamine tegur uute geenide arengul. Mis on praeguseks hetkeks teada üks olulisimaid mehanisme uute geenide tekkel. On tavaline evolutsiooni mehanim ning seega esineb paljudes liikides. Toetavad teadmised – selgroogsete genoomid sisaldavad palju geeniperekondasid, mida pole leitud selgrootutes. Geeniduplikatsioon on oluline sama funktsiooniga geenide koopiate tootmisel ning sellega mRNA ja valgusünteesi suurendamisel. Mitmetel valkudel on sarnased funktsioonid või struktuur.
-
- Genoomse materjali duplikatsiooni viisid; nende olulisus evolutsiooni kontektstis
Osa geenist duplitseeritakse
Ühe geeni duplitseerimine – on oluline mehanism evolutsioonis. Teine geenikoopia on tihti vaba valiku survest – mutatsioonidel ei ole kahjulikku effekti. Ehk teine koopia on just kui varuosa , millelt mutatsiooni puhul saab siiski funktsionaalset produkti toota. Üks koopiatest võib ülevõtta uue funktsiooni. Ehk on peamiseks allikaks uutele genoomi funktsioonidele.
Osa kromosoomist duplitseeritakse
Terve kromosoomi duplitseerimine – aneuploidia või polüsoomia. Võib Bloom ja Rett sündroome põhjustada.
Terve genoomi duplitseerimine – on olnud oluline keelikloomade evolutsioonis. Osad duplitseeritud geenid on muutunud pseudogeenideks, kuid paljud on funktsionaalsed
-
- Kogugenoomi duplikatsioonid selgroogsetel, taimedel; materjal ‘evolutsioonilisteks katsetusteks’
-
- Duplitseerunud geenide edasine areng, ‘saatus’; sarnased (invariant) ja divergeeruvad (variant) geenikoopiad - kasu genoomi funktsioonile
Geeni duplikatsioon üks koopia akumuleerib mutatsioone üleliigne koopia vaigistatakse või omandab uue funktsiooni
Bifunktsionaalse geeni duplikatsioon mõlemad koopiad akumuleerivad mutatsioone funktsioonide jagunemine, üks geen saab ühe ülesande teine teise
Unifunktsionaalse geeni duplitseerimine üks koopia vaigistatakse või mõlemad koopiad säilitavad ekspressiooni puhastava valiku all.
-
- Uute geenide või geenistruktuuride tekke molekulaarsed mehhanismid (8 alternatiivset)
Geeni duplikatsioon – klassikaline mudel. Põhiline allikas uute genoomi funktsioonide tekkel.
Geeni lühenemine või pikenemine – nt uue stop-koodoni tekkimine või vana lõhkumine – tekib pikem või lühem geen. Alternatiivse splaissingu võimalused, haaratakse mittekodeeriv ala, tekib alternatiivne splaissing ja uus ekson .
Ekson/domeen shuffling ja duplikatsioon – ühe või rohkema domeeni duplikatsioon geenis võib tõsta valgu funktsiooni. Strukturaalsete või funktsionaalsete domeenide vahetus valkude vahel võib muuta valgu mitmekesisemaks.
Retrotranspositsioon – kui esivanemlik geen transkribeeritakse, toimub pöördtranskriptsioon ja inserteerumisel võivad tekkida uued produktid.
Mobiilsed elemendid – moodustub uus ekson
De novo – moodustub täiesti uus geen. Mittekodeerivast genoomsest piirkonnast tekib kodeeriv ala.
Lateraalne geeniülekanne – omane prokarüootidele, avastatud ka taimedes. Inimestel u 7-8% genoomist on moodustunud retroviiruse järjestustest ehk uus geen on retroviiruse päritolu.
Fusion/fission – prokarüootides ja seentes – kaks naabergeeni ühinevad üheks geeniks või üks geen eraldub kaheks.
-
- Võrdlev genoomika, fülogeneetika – mis eesmärgid?
pikad fülogeneetilised distantsid – võrrelda organisme mis on kaua aega tagasi lahknenud
vahepealsed distantsid – võrrelda inimest hiirt , bakteri liike omavahel jne.
lähedasi liike positiivse valiku korral – nt inimene ja šimpans
-
- Ortoloogi ja paraloogi mõiste
- Ortoloogid on erinevates liikides, pärit samast esivanemast. Paraloogid on tekkinud duplikatsiooni tagajärjel, ühes liigis .
-
- • Homoloogsete genoomilõikude liikidevaheline võrdlus: etapid, väljakutsed
- • Konserveerunud genoomilõigud ja suured nn. ‘homoloogia blokid’ liikide vahel
-
- Klassikaliste mudelorganismid geneetikas/genoomikas (iseseisev materjali omandamine, sest 03.04 on lihavõttereede ja loengut ei ole) – teada ainult iga liigu põhieeliseid genoomi uuringutes:
- • Saccharomyces cerevisiae (pagaripärm)
- • Caenorhabditis elegans (varbuss, sireuss; nematood)
- • Drosophila melanogaster ( äädikakärbes )
- • Arabidopsis thaliana (harilik müürlook)
- • Danio rerio (sebrakala)
- • Mus musculus (koduhiir)
11


















































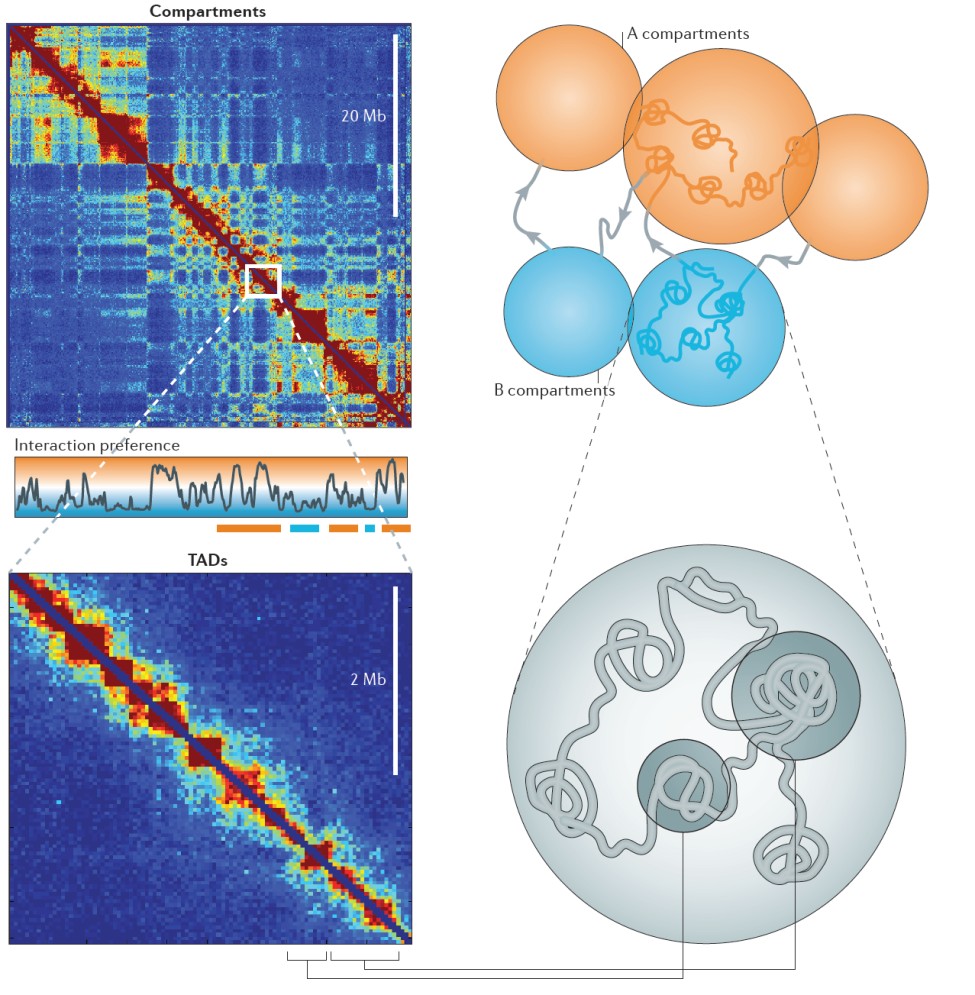
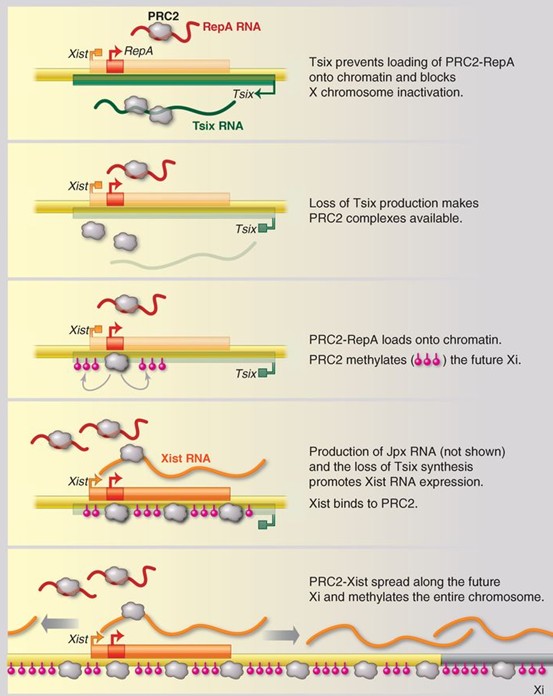
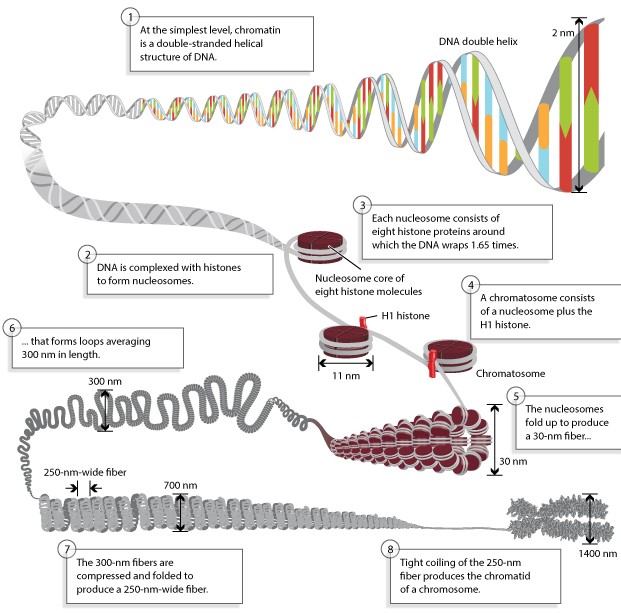
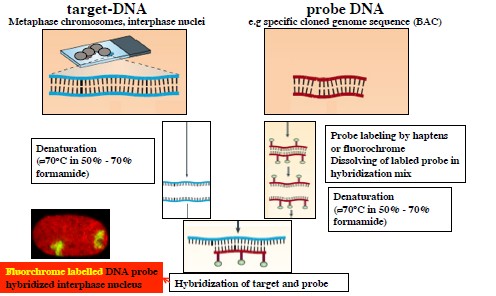
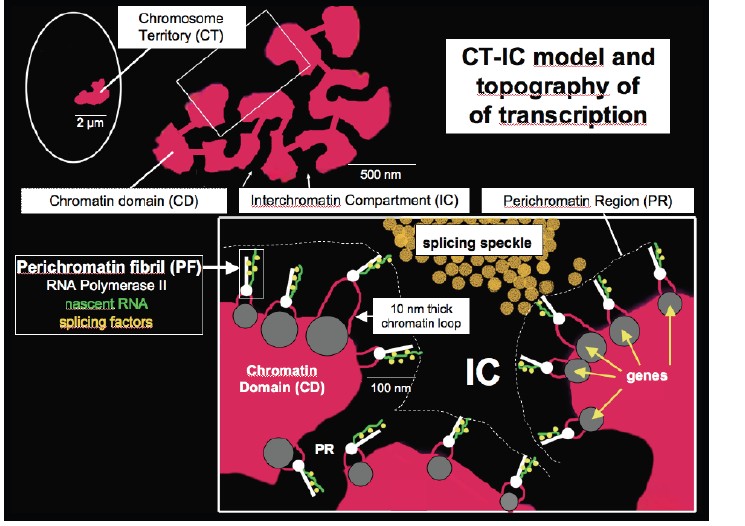
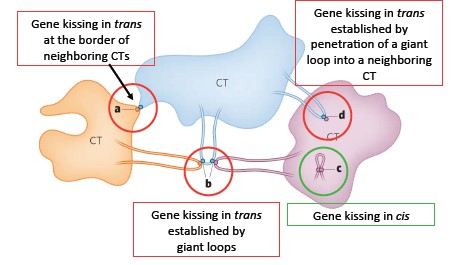
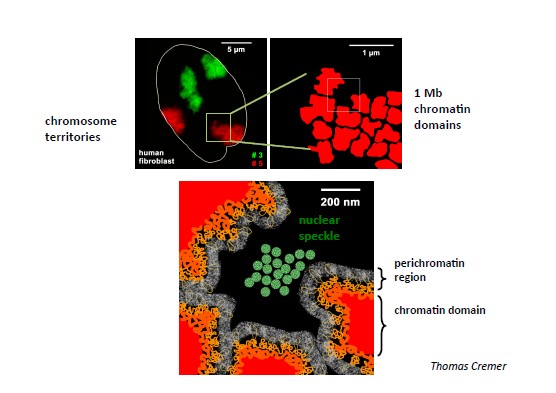
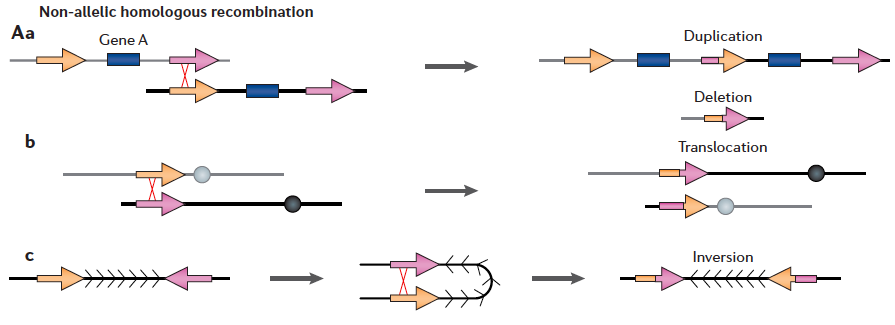

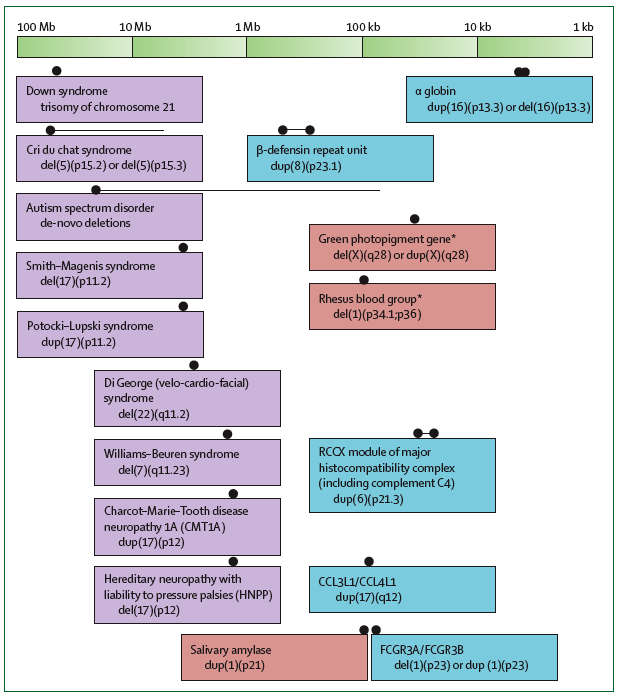
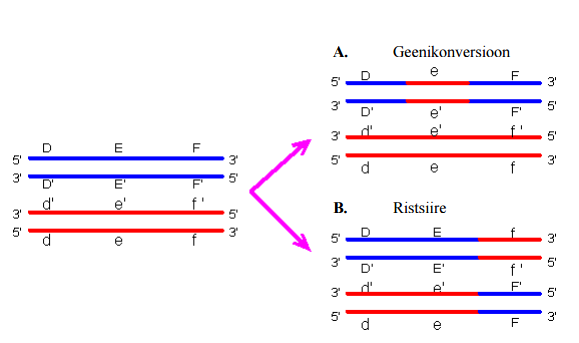




Kõik kommentaarid