1.
DNA
replikatsioon *
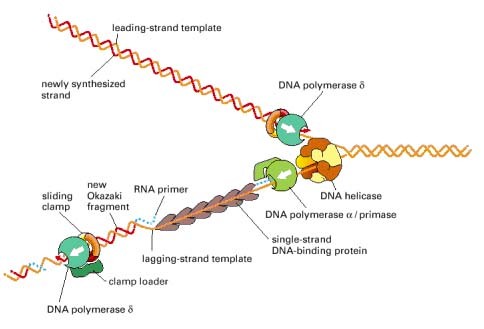
DNA replikatsioonikahvli struktuur
Replikatsioonikahvel
on Y-kujuline aktiivne struktuur, mis moodustub sünteesilookuse
juures, kus 2-
ahelaline DNA läheb üle 1-ahelaliseks. See
tekib rakutuumas DNA replikatsiooni ajal. Selle loovad helikaasid,
mis lõhuvad kahte DNA
ahelat koos hoidvaid vesiniksidemeid. Selle
tulemusena tekib kaks üksikahelat, mis moodustavadki kahvli harud.
Need üheahelalised harud on aluseks juhtiva ja mahajääva ahela
tekkele.
*
Imetaja DNA replikatsiooni
kahvel (vt ka
seminari materjali) 3`
Klaambri laadur
Keerab ahela lahti
Topoisomeraas I
3`
liudklamber
3`
5`
5`
5`
liugklamber
Inimese
rakutuumas sünteesitakse juhtiv ja mahajääv ahel Pol α
ja Pol δ abil ning
mitokondris Pol γ
abil. keerab ahela lahti.
Juhtivalt
ahelalt liigub replikatsiooni kahvel 3’-5’ suunas. See võimaldab
komplementaarse ahela sünteesi 5’-3’ suunas.
Mahajääval
ahelal liigub replikatsioonikahvel 5’-3’ suunas, mistõttu ei saa
mahajäävat ahelat replikatsioonikahvli liikumise suunas pidevalt
sünteesida. Mahajääv ahel sünteesitakse katkendlikult -
fragmentide kaupa.
*
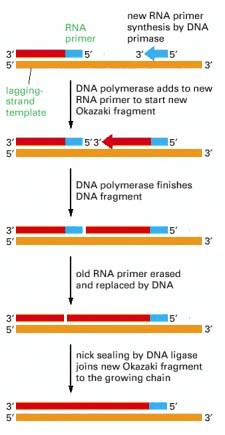
DNA fragmentide süntees mahajääval õlal
Mahajääv
ahel on DNA kaksikheeliksi ahel, millel replikatsioonikahvel liigub
5’-3’ suunas. Selle tõttu ei saa mahajäävat ahelat
replikatsioonikahvli liikumise suunas pidevalt sünteesida. Mahajääv
ahel sünteesitakse fragmentide kaupa. Algsele DNA
ahelale liidetakse
RNA
praimer ning uut ahelat sünteesitakse vastupidiselt
replikatsioonikahvli liikumise suunale. Praimer eemaldatakse
(prokarüootides DNA polümeraas I poolt) ning RNA molekulid
asendatakse DNA molekulidega. Toimub uue RNA praimeri liitumine ning
järgmise fragmendi süntees. Neid lõike nimetatakse Okazaki
fragmentideks ning need liidetakse DNA
ligaasi poolt, et saada
terviklik DNA ahel.
Algsele
DNA ahelale liidetakse RNA praimer ning uut ahelat sünteesitakse
vastupidiselt replikatsioonikahvli liikumise suunale. RNA primer
is erased by a
special DNA
repair enzyme (an RNAse H) that recognizes
an RNA
strand in an RNA/DNA
helix and fragments it; this leaves gaps
that are fiļled in by DNA polymerase and DNA
ligase .
Mahajääval
ahelal on ahela väikese lõigu ühes otsas RNA praimer ja lõigu
teises otsas sünteesib DNA primaas (mis on sisuliselt RNA
polümeraas) samuti uue RNA praimeri. DNA plümeraas lisandub uuele
RNA praimerile ning sünteesib okazaki fragmendi, kuni vana RNA
praimerini, siis eksonukleaasidega eemaldatakse RNA praimer ning asendatakse okazakiga.
RNA
primer sünteesitakse DNA primaasi poolt (see on oma
olemuselt RNA
polümeraas). DNA polümeraas lisandub RNA praimerile, et sünteesida
okazaki fragment.
Mahajääval
ahelal liigub replikatsiooni kahvel 5’--- 3’suunal seepärast
peabki toimuma süntees katkendlikult
*
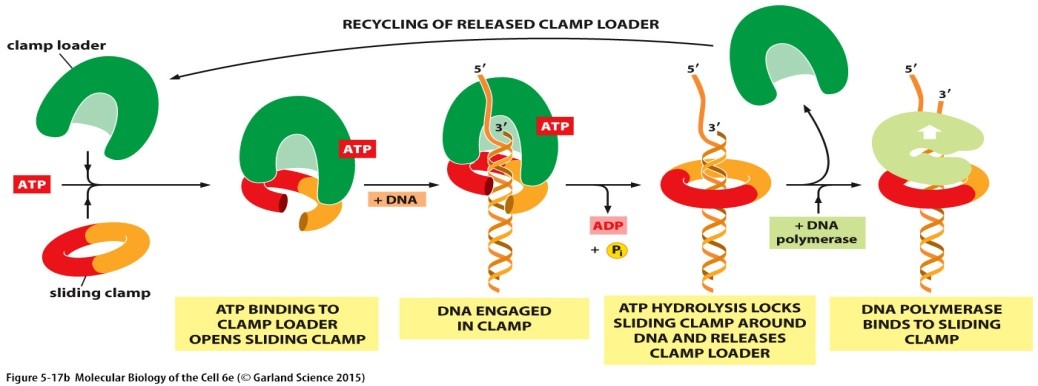
DNA polümeraasi ja klambervalgu seostumine ja vabanemine
viivisahelal
(Liug) klamber hoiab DNA
polümeraasi kopeeritava ahelaga tugevalt seotuna ning ei lase sel
dissotseeruda ehk eralduda.
Klambri
asetab DNA-le libiseva klambri
laadija,
valgukompleks, mis katalüüsib klambri avamist ja asetamist DNA-le,
kasutades ATP hüdrolüüsi energiat.
Liugklamber
hoiab DNA polümeraasi kopeeritava ahelaga tugevalt seotuna ning ei
lase sel dissotsieeruda ehk eralduda. Klambri asetab DNA-le libiseva
klambri laadija, see on valgukompleks, mis katalüüsib klambri
avamist ja asetamist DNA-le, kasutades ATP hüdrolüüsi energiat.
*
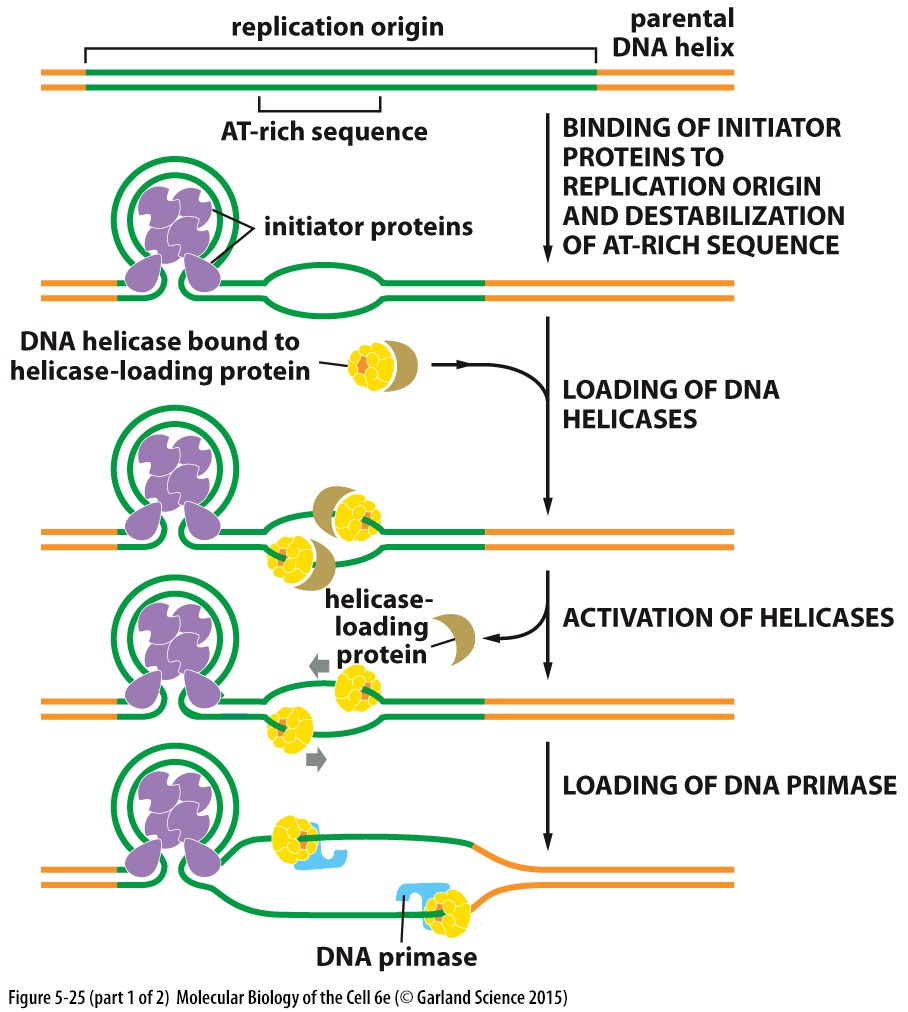
DNA replikatsiooni algatamine bakterite rakus
Valgud ,
mis algatavad bakterites DNA replikatsiooni
2
replikatsiooni kahvlit lähtuvad originist ning liiguvad
vastassuundades.
Initsiaator proteiinid seostuvad spetsiifilisele DNA järjestusele
replikatsiooni alguspunktis (replication
origin ) ja destabiliseerib
kaksikheeliksi, moodustades struktuuri, mille puhul DNA on tugevalt
pakitud ümber valgu.
Kaks
helikaasi seostuvad helicase-loading (DNA C valk) valkudega, mis
inhibeerivad helikaasi, kuni see on täielikult replikatsiooni
alguspunkti jõudnud.
SSB
valkude kaasabil helikaasid avavad DNA. Võimaldades seeläbi
primaasil sisenda ja sünteesida initsiaalne praimer.
Initsiaator
proteiinid eemalduvad kui vasakpoolne replikatsiooni kahvel liigub
neist läbi.
*
DNA replikatsiooni algatamine eukarüootide rakus
See
mehhanism kindlustab, et
igat replikatsiooni alguspunkti
aktiveeritakse ainult üks kord rakutsükli jooksul.
Uut
replikatiivset kompleksi ei saa enne tekkida kui
rakk on jõudnud
uude G1
faasi ja alguspunktiga seotud
kompleks (ORC) on defosforüleeritud.
*
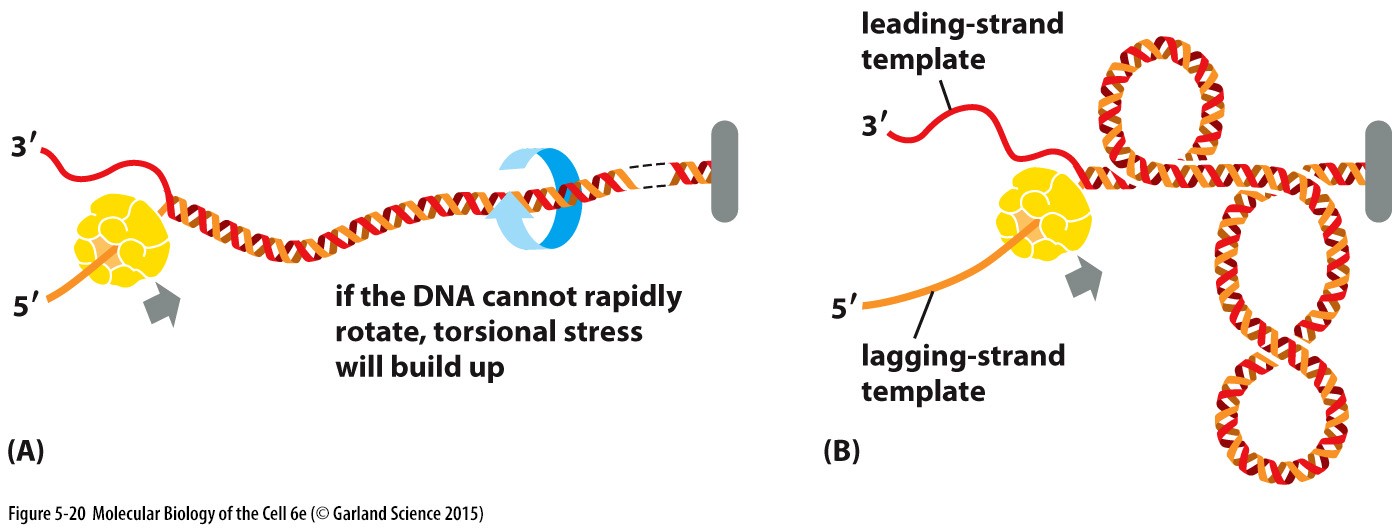
DNA ahela keerdumise probleemid, mis tekivad DNA replikatsiooni
korral
Helikaasi
toimel keerdub kaksikheeliks lahti. Lahtikeerdumisel tekib DNA
torsionaalne pingestumine. Torsioonjõudude leevendamiseks tekitavad
topoisomeraas valgud DNA ahelal katked.
*
DNA topoisomeraas I funktsioon rakus
DNA
topoisomeraas I katkestab eukarüootide DNA ühe ahela
ajutiselt ,
selleks et vältida ahela keerdumist.
*
Kirjelda eksonukleolüütilist korrektuuri (ingl.k. proofreading)
DNA polümeraas kontrollib kas õige nukleotiidiga on tegemist kohe selle lisamisel. Vale nukleotiidi korral on tugeva sideme tekke tõenäosus väiksem.
Eksonukleolüütiline korrektuur – DNA polümeraasi kompleksi üks ensüümidest käitub kui eksonukleaas, mis eemaldab vale nukleotiidi.
2.
DNA-kahjustused
*
DNA-d kahjustavad tegurid
DNA replikatsiooni vead
Kiirgused : ioniseeriv kiirgus (gamma- ja röntgenkiirgus) ja mitteioiniseeriv kiirgus (UV-kiirgus)
Kemikaalid (bensopüreenid) ja keskkonnategurid
Oksüdatiivne stress (superoksiid, O2-)
*
DNA kahjustuste tüübid
Lämmastikaluste eemaldamine DNA-st
Nukleotiidide desamiinimine (aminogrupi eemaldamine lämmastikalusest) ja nukleotiidide valesti paardumine [DNA polümeraas viib läbi ebakorrektse DNA korrektuuri (ingl. k. proofreading)]
DNA- ahelate katkemine
Kovalentsete ristsidemete teke (DNA ahelasiseselt või ahelate vaheliselt)
*
DNA kahjustuste kõrvaldamise viisid/ mehhanismid sh kaks põhilist
DNA parandamise viisi
DNA
kahjustuste kõrvaldamise viisid:
Kahjustatud või valede lämmastikaluste asendamine
DNA-ahelate katkemiskohtade parandamine
DNA
kahjustuste kõrvaldamise mehhanismid:
Otsesed keemilised pöördreaktsioonid (kahjustatud koha kõrvaldamine
ja algse oleku taastamine, näitek T-T
dimeeri kõrvaldamine)
2) Kahjustuste kõrvaldamine väljalõikega (ingl.k. excision
repair)
a)
lämmastikaluste väljalõige
b)
nukleotiidide väljalõige
c)
valepaardumise parandamine
3) Rekombinatsioonist sõltuv parandamine
a)
homoloogne rekombinatsioon
b)
mittehomoloogne DNA otste ühendamine
3.
Kaks viisi kuidas parandada mõlema DNA ahela samaaegseid katkeid
*
Mittehomoloogne- ja homoloogne DNA otste ühendamine’
Mittehomoloogne
DNA otse ühendamine toimub enne DNA replikatsiooni. Kui
heterodimeerid unnevad DNA katkised otsad ära. Katkised otsad
liidetakse, kuid esineb deletsioon, osa geneetilist materjali läheb
kaduma.
Homoloogne
rekombinatsioon toimub vahetult peale DNA replikatsiooni ja enne raku
jagunemist. Otsi ei liideta vaid katkemiskoht parandatakse kasutades
teist homoloogse kromosoomi komplementaarset ahelat. Katkenud kohta
lõigetakse 5` otsast lühemaks ja ahelaid vahetakse
komplementaarsuse alusel. Katkenud kohale pikendatakse ahel kasutades
homoloogse kromosoomi tervet ahealt.
4.
Mis on DNA hübridiseerimine?
Hübridiseerimine on
meetod, mis võimaldab nähtavaks teha, enda jaoks, in vivo olevaid
molekule – molekule, mis pole sünteeside käigus
märgistatud. Palju
DNA ahelaid, mis
juhuslikult leiavad endale komplementaarse paarilise ja toimub nii
paardumine. Paardumine on nõrk ja lühikese ulatusega.
5. Ristsiire ehk krossingover
Ristsiire ehk krossingover (homoloogiline
rekombinatsioon) on protsess, mille käigus toimub
homoloogiliste kromosoomide põimumine,
mille jooksul nad vahetavad võrdsetes kogustes pärilikkusainet.
Ristsiire
toimub meioosi profaas.
Meioos
on protsess, kus diploidne rakk jaguneb 4 haploidseks rakuks, milles
on emalt ja isalt saadud geneetiline info. Geenikonversiooni
puhul toimub ainult lühikese osa DNA ülekanne ühest homoloogsest
kromosoomist teise ja tihti muutub ainult osa geenist.
*
Kiasmid
Kiasm
- Esimese meiootilise jagunemise profaasi diploteeni staadiumis kahe
homoloogse kromosoomi neljast kromatiidist koosnevas grupis nähtav
kahe kromosoomi vaheline DNA-lõikude vahetuskoht, kus toimub
ristsiire. Diploteenis sünaptonemaalne kompleks laguneb, kromosoomid eemalduvad teineteisest, kuid jäävad seotuks sealt, kus toimus
kromosoomide ristsiire ehk kiasmide
koha pealt.
*
Sünaptonemaalne kompleks
Sünaptonemaalne
kompleks –
sügoteenis moodustuv valguline struktuur.
Sünaptonemaalne
kompleks laguneb, homoloogid eemalduvad veidi teineteisest, kuid jäävad siiski veel
seotuks nendest kohtadest , kus toimus krossingover.
*
Holliday ühendus (ingl. k. Holliday junction )
Holliday
ühendus (ingl. Holliday
junction)
on DNA nelja
ahela vaheline liikuv ühendus, mis ilmub, kui
toimub kromosoomide konjugatsioon meioosis. Need
ahelad võivad võtta palju erinevaid konformatsioone sõltuvalt
soolakontsentratsiooni puhverlahusest ja ümbritsevate nukleotiidide
järjestustest, mis asuvad ühenduse lähedal.
6.
Geenikonversiooni ja ristsiirde erinevus
Geenikonversiooni
puhul üks DNA
annab enda info teisele, vastu ei taha midagi
Geenikonversiooni
teevad läbi ainult väiksed DNA osad või isegi ainult osa geenist
Krossing
over – kaks DNA-d vahetavad omavahel
geneetiist infot.
7. Mobiilsed geneetilised elemendid – transposoonid - mis need on ja
mida nad teevad?
Mobiilsed
geneetilised elemendid ehk transposoonid,
mis „hüppavad” iseseisvalt genoomi ühest kohast teise
(protsess, mida kutsutakse transpositsiooniks),
koosnevad mittekodeeriva DNA kordusjärjestustest, mis moodustab
inimese tuumagenoomist umbes 45%.
*
Konservatiivse kohaspetsiifilise rekombinatsiooni erinevus
transposoonidest
Konservatiivse kohaspetsiifilise rekombinatsiooni puhul on vaja spetsiaalseid DNA järjestusi nii doonori kui ka retsipient DNA osas
Transposoonide puhul peab ainult nende endi järjestus olema spetsiifiline, mitte aga märklaud-DNA oma
Konservatiivse kohaspetsiifilise rekombinatsiooni puhul taastatakse kõik katkenud fosfaatsillad
Transposoonide puhul jäävad DNA ahelasse augud, mille parandab DNA polümeraas
*
Transposoonide põhiklassid, nende kirjeldus/erinevus (sh millised
ensüümid vahendavad transposoonide transponeerimist)?
Transposoonide tüübid:
Lühikesed vahelelükitud tuuma elemendid (SINE)
SINE klassifitseeritakse kolme perekonda:
Alu
MIR
MIR3
On
olemas 3 liiki transposoone:
Ainult DNA kujul esinevad transposoonid, mis toodavad ise endale ensüümi TRANSPOSAAS. Sisenevad retsipient DNAsse lõika ja kleebi põhimõttel.
Retroviirusesarnased retrotransposoonid – ensüümideks on pöördtranskriptaas ja integraas. Levivivad vahendaja RNA kaudu rakkude jagunemisel.
Mitteretroviiruselised retrotransposoonid – pöördtranskriptaas ja endonukleaas – Levivad vahendaja RNA kaudu. Sarnanevad retroviirustele, kuid neil puudub valguline kate.
*
Mis on transposoom (ingl. k. transposome)?
mingid
DNA järjestused, mis hakkavad transpositsioneeruma, nende otstes on
lühikesed inverteeritavad kordusjärjestused. Transposaasid tekivad
rõnga ehk loopi ning lõikavad doonorahelast transposoomi
välja. Transposoomi rõngasstruktuur läheb aktseptor -ahelasse.
Sisenemist katalüüsivad transposaasid.
8.
Kuidas saab konservatiivset kohaspetsiifilist rekombinatsioonis
toimivat bakteriaalset ensüümi kasutada koespetsiifiliste geenide
väljalülitamiseks hiires (vt ka seminari materjali)
9.
RNA ja transkriptsioon
Transkriptsioon on matriitssüntees,
mille käigus sünteesitakse DNA molekuli
ühe ahela nukleotiidse järjestusega komplementaarne RNA molekul .
*
Bakterite ja eukarüootide polümeraasid.
Bakterites
on ainult ühte tüüpi RNA polümeraasi, selle poolt sünteesitakse
kõik RNA tüübid.
1.
Transkriptsioon algab DNA promootorkpiirkonnast, kus RNA polümeraasi
holoensüüm – faktor sigma ja RNA plümeraas, asetuvad DNA promootor alale.
2.
Plümeraas avab DNA aehla ning transkirptsioon algab.
3.
Kui RNA plümeraas on sünteesinud 10 nukelotiidse RNA, siis murduvad interaktsiooni promootorpiirkonnaga DNA’l,
4.
Sigma faktor vabaneb.
Polümeraas läheb tugevamini DNA külge, toimub elongatsioon, liigub
mööda DNA ahelat. Sigmafaktor liitub hiljem uuesti RNA
polümeraasiga, et hakata sünteesima uut ahelat.
5.
Elongatsioonis on transkriptsioon väga produktiivne, polümeraas
lahkub DNA ahelalt ja vabastab vast-sünteesitud RNA, kui kohtab terminatsiooni signaali.
6.
Terminatsiooni signaal tuleb DNAlt , selle tulemusel moodustub RNA
juuksenõela struktuur, mis destabiliseerib polümeraasi hoidmist RNA
küljest.
RNA
polümeraasid eukarüootide rakkudes
RNA
polümeraas I – transkribeerib 5,8S,
18S ja 28S rRNA geene
RNA
polümeraas II – transkribeerib kõiki
valke kodeerivaid geene,
aga lisaks ka snoRNA , miRNA , siRNA , lncRNA ja enamust snRNA geene
RNA
polümeraas III – transkribeerib
tRNA, 5S
rRNA, mõnesid snRNA ja teiste väikeste RNA-de geene
*
Nimeta rakkude poolt toodetavad RNA-d ja nende funktsioon.
Informatsiooni-mRNA kannab
informatsiooni valgujärjestuse kohta ribosoomi,
mis on valgusünteesi masinavärgiks rakus. mRNA on kodeeritud
niimoodi, et järjestikused kolm nukleotiidi (koodon)
vastavad ühele aminohappele .
Transpordi-RNA (tRNA)
on väike RNA ahel, mis kannab kindlaid aminohappeid ribosoomi
valgusünteesi aktiivtsentrisse, kus aminohapped liidetakse kasvavale
polüpeptiidahelale. tRNA-l on piirkonnad aminohapete seondumiseks
ja antikoodonregioon koodonite äratundmiseks
mRNA ahelal
snRNA-d
- Väiksed tuuma RNA-d, osalevad paljudes tuumas toimuvates
protsessides, k.a. pre-mRNA splaissing
snoRNA-d
- Väiksed tuumakese RNA-d, mis aitavad töödelda ja keemiliselt
muuta rRNA-sid
miRNA-d
- MikroRNA-d, reguleerivad geeniekspressiooni blokeerides
spetsiifiliste mRNA-de translatsiooni ja põhjustavad nende
lagundamist
siRNA-d
- Väiksed segavad RNA-d, lülitavad välja geeniekspressiooni
juhtides mRNA-de lagundamist ja moodustades kompaktse kromatiini struktuuri
piRNA-d
- Piwi-ga seostuvad RNA-d, seostuvad piwi
valkudega ja kaitsevad sugurakke transposoonsete elementide eest
lncRNA-d
- Pikad mittekodeerivad RNA-d, paljud neist toimivad kui tellingud;
nad reguleerivad mitmeid erinevaid protsesse rakus, k.a. X-kromosoomi inaktivatsioon
*
Prokarüootide ja eukarüootide mRNA üldise struktuuri võrdlus.
Prokarüootide
mRNA 5’ ja 3’ otsas on polümeraasi poolt sünteesitud mitte modifitseeritud otsad.
Eukarüoodil
on lisatud 5’ cap ja 3’ otsa on lisatud poly A saba. Samuti on
bakteril mRNA sisaldab mite valgu järjestusi, eukarüoodil sisaldab
üldiselt ainutl ühe valgukohta infot. Eukarüoodi 5’ G---CH3.
lisatud on metüül rühm cap.
10.
Transkriptsiooni alustamine RNA polümeraas II toimel eukarüoodi
rakus.
*
Nimeta üldised transkriptsioonifaktorid
Eukarüootides on
olemas oluline klass transkriptsioonifaktoreid, mida nimetatakse
üldisteks transkriptsioonifaktoriteks (GTF)
Kõige
tavalisemad GTF-id on TFIIA, TFIIB, TFIID, TFIIE, TFIIF
1.
Nad aitavad paigutada eukarüootide RNA polümeraasi täpselt
promootorile.
2.
Nad aitavad DNA ahelaid teineteisest lahti tõmmata, selleks et
transkriptsioon saaks alata.
3.
Nad aitavad vabastada RNA polümeraasi promootori küljest, selleks
et viia transkriptsioon n.ö. pikendamise (ingl.k. elongation )
etappi.
*
Mis on TBP funktsioon initisiatsiooni kompleksi tekkes?
TBP
on DNA ahela neljanukleotiidine järjestus Promootorist 25
nukleotiidi eespool Eukarüootse
RNAP II-st sõltuv promootor sisaldab tavaliselt TATA-elementi,
mille nukleotiidseks järjestuseks
on TATAAA . Selle järjestuse tunneb ära TBP (TATA-binding protein ).
11.
Pre-mRNA põhilised modifitseerimise protsessid.
Alternatiivne RNA splaissimine – pre-mRNA võib saada splaissitud mitmetest
erinevatest ühenduskohtadest, nii et tekivad erinevad küpsed
mRNA-d, millel on erinevad eksonite kombinatsioonid
Prokarüootidel on geeni poolt
määratav esmane transkript võrdne mRNA-ga
ning ta on ka kohe transleeritav. Eukarüootides toimub
aga esmalt eellas- ehk pre-mRNA süntees,
misjärel toimub nn. eellas-mRNA protsessing küpseks mRNA-molekuliks.
12.
Pre-mRNA kokkupõime ehk splaissingu reaktsioon
*
Kirjelda mehhanismi, mille abil pre-mRNAst kõrvaldatakse intronid .
splaissing (ka splaising ; inglise splicing)
protsess, mille käigus lõigatakse rakutuumas asuvast RNA molekulist
välja intronjärjestused ning
allesjäänud eksonite otsad
ühendatakse. Splaissingu tulemusena tekib mRNA,
mida kasutatakse translatsioonil korrektse proteiini
sünteesiks. Eukarüootsete intronite
puhul katalüüsib splaissimisreaktsioone splaissosoom,
kuid olemas on ka isesplaissuvaid
introneid.
Splaissosoom on väikeste tuuma ribonukleoproteiinide (inglise small nuclear ribonucleoprotein, snRNP)
kompleks.
13.
mRNA molekuli transport läbi tuumapoori kompleksi
*
Millised valgud osalevad selles protsessis?
EJC
– eksoni ühendus kompleks (ingl.k.
exon junction complex )
seostub mRNA-le kohtadesse kus varem pre-mRNA-l paiknesid intronid
CBC
– cap-iga
seostuv kompleks
hnRNPs
- Heterogeneous nuclear ribonucleoproteins
SR
valgud (SR proteins) - reguleerivad mRNA stabiilsust, eksporti ja
translatsiooni. Nende C-terminuses on RS
domäänid
( seriini –arginiini
kordusjärjestused,
mis vahendavad valk-valk ja valkRNA interaktsioone.
14.
Transkriptsiooni kontroll järjestusspetsiifiliselt DNA-ga seonduvate
valkute abil
*
Mis on cis-regulatoorsed järjestused?
Cis
järjestused –
samal molekulil , mida reguleeritakse trans faktorid – valgud.
Transkriptsiooni regulaatorid sisaldavad struktuurseid motiive , mis n.ö. loevad DNA
nukleotiidseid järjestusi. Need on mittekodeerivad cis-regulatoorsed
järjestused, mis peavad asuma samas kromosoomis (in
cis)
kus asuvad geenid, mida nad reguleerivad. Cis-regulatoorsetele
järjestustele seostuvad transkriptsiooni regulaatorid. Iga
transkriptsiooni regulaator tunneb ära oma cis- regulatoorse järjestuse.
*
Nimeta ja kirjelda transkriptsiooni regulaatorite põhilisi
struktuurilisi motive
Transkriptsiooni
regulaatorid sisaldavad struktuurseid motiive, mis n.ö. loevad DNA
nukleotiidseid järjestusi.
1.
Heeli-pööre-heeliks - koosneb kahes heeliksist, mida seob oma
vahel lühike AH ahel.
Heeliksid
on fikseeritud nurga all.
2.
homeodomeeni valgud. koosnevad
kolmest
alfa-heeliksist, mis on ühendatud omavahel hüdrofoobsete sidemete
abil.
3.
Leutsiini tõmbluku valgud – koosnevad
kahest alfa-heeliksist, mida hoiavad koos hüdrofoobsed AH ( leutsiin )
külgmised sidemed.
4.
β-lehe
tüüpi DNA-d äratundvad valgud
5. Tsink -sõrme valgud – struktuuris üks või rohkem tsingi aatomit.
*
Milleks on oluline transkriptsiooni regulaatorite dimeriseerumine?
Transkriptsiooni
regulaatorite dimeriseerumine suurendab nende afiinsust ja
spetsiifilisust DNA suhtes.
15.
Nukleosoomide mõju transkriptsiooni regulaatorite seondumisele
*
Nukleosoomide ’ hingamine ’
Nukleosoomi
äärealal DNA n.ö. „ hingab “ st ajutiselt eksponeerib DNA-d, mis
võimaldades transkriptsiooni regulaatoritel kergemini seostuda
Seetõttu
võimaldab nukleosoomi hingamine transkr. regulaatoril seostuda;
16.
Transkriptsiooni repressorid.
Transkriptsiooni
regulaatorid:
transkriptsiooni aktivaator lülitab geenid sisse ja repressor välja.
Mõlemad
seostuvad cis-regulatoorsele
järjestusele DNA-
Ühel
transkriptsiooni faktoril, DNA
helikaasil,
on helikaasne aktiivsus ning on seetõttu seotud kaheahelalise DNA
ahelate eraldamisega, tagamaks ligipääsu üheahelalisele matriits
DNA-le. Teised
valgud, aktivaatorid ja repressorid vastutavad koos erinevate seotud
koaktivaatorite või korepressoritega transkriptsiooni moduleerimise
taseme eest.
17.
Geenide transkriptsiooni erinevused eukarüootides ja prokarüootides.
Prokarüootides seostuvad transkriptsiooni regulaatorid (aktivaatorid ja repressorid) otse DNA-le või RNA polümeraasile.
Eukarüootides seostuvad transkriptsiooni regulaatorid (aktivaatorid ja repressorid) DNA-le või RNA polümeraasile paljude vahevalkude kaudu.
Eukarüootides kontrollivad paljud transkriptsiooni regulaatorid cis-regulaator piirkondade kaudu ühte geeni, mille ulatus on tuhandeid nukleotiidi paare.
Eukarüootides tekivad transkripsioonil DNA lingud.
Eukarüootide DNA on pakitud nukleosoomidesse ja veel keerulisematesse struktuuridesse.
18.
Transkriptsioonijärgne geeniekspressiooni kontroll
*
Ribolülitid ja nende roll geeniekspressiooni kontrollis
Ribolülitid
on lühikesed RNA järjestused, mille kuju muutub kui nendele
seostuvad väiksed molekulid (näit. metaboliidid). Nad paiknevad
vastsünteesitud mRNA 5´-otsa läheduses.
*
RNA alternatiivse splaissingu negatiivne ja positiivne kontroll
Negatiivne
kontroll-repressorvalk seostub pre-mRNA-l kindlala järjestusele ning
blokeerib splaissingu valkudenligipääsu.
Positiivne
kontroll- splaissingu valgud ei suuda eraldada kindlat intronit ilma
aktivaatorvalguabita RNA toimetamine A-st l-ks-adeniini
desaineerimine inosiiniks (aDAR ensüüm) C-st U-ks tsütosiiini
deamineerimine uratsiiliks mRNA transport tuumast tsütoplasmasse on
rakus rangelt reguleeritud.
*
Kontroll translatsiooni tasemel sh Shine- Dalgarno järjestus,
translatsiooni alustamise kaks mehhanismi (nt rakkudes ja viirustes)
Bakterites
kontrollib mRNA translatsiooni konserveerunud nukleotiidne järjestus
(Shine-Dalgarno järjestus),
mis asub alustava AUG koodoni ees
1.
Cap- dependent mechanism-
Rakkudes algab translatsioon mRNA 5´-otsast AUG koodonist Cap ja polüA saba
stimuleerivad komplekti initsiatsiooni faktorite seondumist.
2. IRES -dependent mechanism-
Viiruste puhul algab
translatsioon tihti mRNA 5´-otsast kaugel, kus asub sisemine
ribosoomi sisenemise koht (ingl.k. internal ribosome entry site, IRES),
spetsiifiline RNA järjestus
*
Geeniekspressiooni reguleerimine mRNA stabiilsuse tasemel
Geenide
ekspressioonil eristatakse kahte geneetilise kontrolli mehhanismi:
positiivne ja negatiivne geneetiline kontroll. Mõlema korral
kasutatakse geenide sisse- ja väljalülitamist vajaliku kontrolli
saavutamiseks. Mõlema kontrollmehhanismi juures esinevad
indutseeritud ja pidurdatud süsteemid.
Positiivse
kontrolli puhul on süsteem algolekus suletud. Regulaatorgeeni
produktiks on aktivaator, mida on vaja struktuurgeenide
sisselülitamiseks. Positiivse geneetilise kontrolli puhul regulatsioon toimub promootori tasemel, aktivaator seondub promootori
ees oleva regulaatorvalgu seondumissaidiga.
Negatiivse
kontrolli puhul on süsteem algolekus suletud. Regulaatorgeeni
produktiks on repressor, mis on vajalik geenide väljalülitamiseks.
Negatiivse geneetilise kontrolli puhul on regulatsioon operaatori
tasemel ja repressor seondub promootori järel olevasse
regulaatorgeeni seondumissaiti.
19.
Mittekodeerivad RNA-d geeniekspressiooni regulatsioonis
*
RNA interferents (RNAi) eukarüootides
RNA
interferents e. RNA häiring e. RNAi eukarüootides
Üheahelalised
interfereerivad RNA-d moodustuvad kaheahelaliselt RNAlt (20-30
nukleotiidi).
Nad
paarduvad sihtmärk RNAle ja võivad nii põhjustad mitmeid
kahjustusi:
1.
lõikavad,
2.
represseerivad translatsiooni ja sihtmärk RNA destruktsiooni,
3.
moodustavad heterokromatiinseid alasid DNAl, millelt sihtmärk RNA
transkribeeritakse.
Siia
klassi kuuluvad:
mikroRNA-d
(miRNA-d), väiksed segavad RNA-d (siRNA-d) ja piwi-seoselised RNA-d
(piRNA-d).
Kõik
nad on lühikesed, algselt kaheahelalised, seostuvad RNA-le ja
vähendavad geeniekspressiooni.
*
miRNA-d
miRNA-d
reguleerivad mRNA translatsiooni ja stabiilsust
miRNA-de protsessing ja toime.
miRNA-sid
sünteesitakse RNA polümeraas II poolt, lisatakse cap-struktuur
ja polüadenüleeritakse.
Prekursor miRNA moodustab endaga kaheahelalise struktuuri. Tuumas kärbitakse
teda ja saadetakse tsütosooli, kus see lõigatakse dicer ensüümiga,
nii moodustub miRNA.
*
Mis vahe on miRNA-l ja siRNA-l?
RNA
interferents (RNAi)
on süsteem elavates rakkudes,
mis osaleb geeniaktiivsuste määramisel. RNA interferentsiks on
olulised kaks tüüpi väikseid RNA molekule
– mikroRNA (miRNA)
ja väike interfereeriv RNA (siRNA)
Lühikesed
üheahelalised RNA-d, kuid pikemad kui miRNA-d ja siRNA-d.
Taimedes
RNAi levib siRNA transpordi teel läbi rakuseintes
olevate plasmodesmide.
Taimede
ja loomade vaheline suurem üldine erinevus peitub endogeensete
miRNA-de sihtmärgiks määramises. Taimedes miRNA-d on täiuslikult
või peaaegu täiuslikult komplementaarsed oma sihtmärk-geeniga ja
indutseerivad otse RISC -ilt mRNA lõikamise, kuid loomades on miRNA-d
järjestuselt pigem divergeerunud ning indutseerivad
translatsioonilise repressiooni.































Kõik kommentaarid