Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
1. Seedeelundite ehitus ja funktsioon
Mao ja soolestiku peaülesanne on muuta söödud toit resorbeeritavateks
koostisosadeks ja need kehasse vastu võtta. See algab mehhaaniliste
protsessidega (peenestamine,
segamine , transport) ning ensüüme sisaldavate
seedenõrede sekretsiooniga, mis lõhustavad hüdrolüütiliselt valgud,
rasvad ja
süsivesikud imendatavateks fragmentideks (seedimine). Vesi,
vitamiinid ja
mineraalained võetakse soolevalendikust läbi soolelimaskesra verre
(resoptsioon).
Seedimine algab
suus , kus hammastega mäludes toit mehhaaniliselt
purustatakse ning süljega segatakse (lisaks limaskestasisestele
süljenäärmerakkudele suubuvad suhu
gl. sublingualis’e,
gl. submandibularis’e ja
gl. parotis’e juhad). Neelamisega transporditakse toit söögitorru (
oesophagus ) ja
makku.
Maos (
gaster s. ventriculus) seguneb toit maonõrega ja transporditakse
maoliigutustega distaalsesse mao-
ossa . Sülje ja maonõrega segunenud
toidumassi nimetatakse küümuseks, mis transporditakse läbi maolukuti (
pylorus)
duodenum’ isse .
Duodeenumisse sekreteerub soolenäärmete sekreet ning siia suubuvad ka
pankrease ja sapijuhad (ühissapijuha tuleb duodenumi
pars descendens’isse ja
avaneb papilla duodeni majori’il koos pankreasejuhaga-
ductus pancreaticus’ega.
Juhade lõpposad võivad olla ka ühinenud
ampulla hepatopancreatica’ks).
Duodeenumi limaskesta näärmerakkudes sünteesitakse palju
seedekulgla hormoone, mis reguleerivad seedenäärmete tegevust. Peensoole lihaskihid
kindlustavad küümuse
segamise ja transpordi. Enamik imendumisprotsessidest
toimub
peensooles .
Jämesooles imenduvad vesi ja elektrolüüdid lõplikult ning bakterite
ensüümide toimel lõhustatakse
kiudained . Jämesoole lõpposas moodustub
väljaheide, mis sisaldab imendumata jääkaineid, irdunud sooleepiteeli rakke,
mikroobe ja vähesel hulgal vett. Defekatsiooniga eemaldatakse
pärasoole kaudu
1
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
roojamassid.
Maks osaleb bilirubiini moodustamises,
sapi abil lipiidide seedimises,
toksiinide kahjutustamises. Maksal on
tsentraalne roll süsivesikute, rasvade,
valkude ja hormoonide ainevahetuses.
Pankrease välissekretoorne osa sekreteerib rikkalikult HCO -
3 , valkude,
rasvade ja süsivesikute seedimiseks toodetakse kõhunäärmes vastavaid
ensüüme.
2.Seedeensüümid ja nende toimeks vajalikud tingimused
Seedeensüümidest on süljes keelenäärmete
lipaas (keele seroossetest
näärmetest), mille toimeoptimum on pH 2,0…6,5. See on happe- ja pepsiinikindel
ja toimekohaks on
magu .
α- amülaasi (isoensüümide segu) toimeoptimum on
pH 6,8…7,2 ja ta alustab tärklise seedimist suus (pisut ka maos, seni kuni
maohape ta denatureerib).
Mao limaskesta pearakkudes sünteesitakse peptiidsidemeid
lõhustavaid
pepsiine, mille isoensüümide 1-5 (ka isoensüüm A) pH toime
optimum on
1,5…2,2 ja isoensüümide 6-7 (ka isoensüüm C) toime optimum on 3,0…3,2).
Maos sünteesitakse ka
lipaasi pH optimumiga 7,0…8,8, mis lõhustab imikul
piimarasvu (täiskasvanul vähetähtis).
Peensooles, enterotsüüdi membraanil tekib tärklise, glükogeeni ja maltoosi
lõhustamiseks vajalik
γ- amülaas, pH optimum ~6,8…7,2. Samas tekivad ka
süsivesikuid lõhustavad
isomaltaas,
maltaas (pH optimum 5,5…6,5),
sahharaas (pH optimum 5,5…6,0),
laktaas (pH optimum 5,0…5,5). Peensoole
limaskestas tekib ka
enteropeptidaas (vajalik trüpsiini aktivatsiooniks, pH
optimum 6…9),
tripeptidaas,
dipeptidaas ja
endopeptidaas (lõhustavad
peptiidsidemeid). Peensooles ja kõhunäärme aatsinusrakkudes tekib ka
fosfolipaase ja
aminopeptidaase.
Pankrease eksokriinses osas tekib
trüpsiin (pH optimum 7,5…8,5, toimib
peptiidsidemesse Lys ja Arg vahel),
kümotrüpsiin (pH optimum 7,5…8,5) ja
pankreatopeptidaas E ehk
elastaas , mis hüdrolüüsivad valgumolekuli
ahelasiseseid peptiidsidemeid ning ahela otsmisi
sidemeid hüdrolüüsivad
karboksüpeptidaas A ja
B ning
aminopeptidaas. Ensüümide
aktivatsiooniprotsessis on oluline trüpsinogeeni aktiveerumine trüpsiiniks
peensoole seinas happelises keskkonnas (happeline keskkond tekib sooles
lühiajaliselt maosisaldise üleminekul soolde). Tee toimub peensooles vabaneva
enterokinaasi (enteropeptidaasi) toimel. Enterokinaasi toimel eraldub
trüpsinogeenist inhibeeriv valk ja vabaneb trüpsiin, mis ka ise jätkab edasist
trüpsinogeeni aktivatsiooni trüpsiiniks. Trüpsiini toimel aktiveeritakse ka teised
ensüümid (prokarboksüpeptidaas => karboksüpeptidaas, elastaasi aktivatsioon
jt.).
Pankreases tekivad ka nukleiinhappeid lõhustavad
ribonukleaas ja
desoksüribonukleaas (pH optimum 7,0…7,5), emulgeeritud
lipiide lõhustav
lipaas (pH optimum 6,5) ja
kolesteraas, mis lõhustab kolesterooli estreid (pH
optimum 6,6…8,0). Süsivesikuid (polüsahhariide) lõhustab pankrease
α- amülaas (toimeoptimum pH 6,8…7,2).
3. Seedeelundite talitluse regulatsioon
3.1. Seedeelundite innervatsioon: sümpaatiline, parasümpaatiline, enteraalne.
Seedekulglal on
enteraalne närvisüsteem, mis võimaldab juhtida mao ja
soolestiku motoorset ja sekretoorset aktiivsust, sõltumata välisest autonoomsest
närvisüsteemist. Enteraalne närvisüsteem asub neuronaalse võrgustikuna piki ja
ringlihaskihi vahel (
plexus myentericus s. Aurebachi ) ning ringlihaskihi ja
2
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
submukoosa muskulatuuri vahel (
plexus submucosus s. Meissneri). Plexus
myentericus’e eferentsed kiud lõpevad piki- ja ringlihaskihi silelihasrakkudel,
mõjustavad lihastoonust, kontraktsioonide rütmi. Submukoospõimik juhib
peamiselt epiteelirakkude sekretoorset funktsiooni. Mõlema põimiku aferentsed
kiud juhivad sensoorseid impulsse
mehhano ja notsiretseptoritest KNS-i.
Enteraalsel närvisüsteemil on olemas sünapsid parasümpaatilise ja
sümpaatilise närvisüsteemi neuronitega.
Parasümpaatiline N. vagus innerveerib
ösofagust, magu, peensoolt, proksimaalset jämesoole osa, sapipõit ja pankreast.
Parasümpaatilised kiud
seljaaju sakraalosast (
nn. splanchici)
innerveerivad sigmasoolt, rektumit ja päraku piirkonda. Parasümpaatiliste närvikiudude
preganglionaarne kiud lülitub ümber seinasiseses (intramuraalses) ganglionis,
seega
postganglionaarne kiud on lühike. Mediaatoriks on siin atsetüülkoliin, aga
postganglionaarsetel kiududel on mediaatorina leitud ka VIP,
substants P ja
serotoniin . Atsetüülkoliin reageerib muskariintundlike retseptoritega.
Sümpaatikuse preganglionaarsed kiud seedekulglale väljuvad Th5…Th12,
L1…L3, lülituvad ümber ganglion coeliacum’is (söögitoru, maks, kaksteistsõrmik,
maks, pankreas), ganglion
mesentericum superius’es (
peensool , jämesoole
algusosa ), ja ganglion mesentericum inferius’es (alumine käärsool, pärak).
Preganglionaarsete kiudude erutuse ülekande substants on atsetüülkoliin,
postganglionaarsetel
noradrenaliin . Aferendid kulgevad nii uitnärvi kui
sümpaatilise närvi
koosseisus , juhivad KNS-i impulsse, mis
kutsuvad esile
teadliku taju või autonoomsete
reflekside vallandumise. Sensorid võivad paikneda
epiteelirakkudes, lihaskihis (olles tundlikud
venitusele ) või
seedekulglas (olles
tundlikud sisu konsistentsile,
osmootse rõhu või pH suhtes).
Aferentsed autonoomsed kiud kulgevad nii sümpaatiliste kui
parasümpaatiliste närvide koosseisus kesknärvisüsteemi.
3.2. Difuusne neuroendokriinne süsteem seedekulglas. Seedeelundite
endokriinne funktsioon.
Seedekulgla kuulub organismi kõige hormooniderikkamate ja
hormoonaktiivsemate elundkondade hulka. Seedekulgla limaskestas on
diferentseeritud 18 rakutüüpi, milles on tõestatud gastrointestinaalse toimega
hormoonide või peptiidide olemasolu. Klassikalised GI hormoonid on
gastriin ,
sekretiin , koletsüstokiniin, mis vastava ärrituse puhul lähevad verre ning
avaldavad oma toimet lõpporganile. Bioloogiliselt aktiivsed
peptiidid ei vasta
kõigile hormooni
kriteeriumidele , kuid avaldavad GI hormoonide
omaga sarnast
toimet. Mõned neist difundeeruvad neid moodustavast rakust otse lähedal
olevasse efektorisse ilma vere abita (parakriinsus), teised vabanevad
närvilõpmetes,
toimivad samuti otsesel teel (neurokriinsus). Mõnedele
neuropeptiididele, mida peeti ainult ajus olevateks, nagu enkefaliinid ja
endorfiinid, on ka sooles olemas opiaadiretseptorid. Näärmerakke on eriti rohkelt
maos ja peensooles.
Endokriinsed rakud toodavad hormoone, mis toimivad kas
vere kaudu või parakriinselt. Parakriinselt toimib näiteks
histamiin (mao
limaskesta ECL-rakkudest parietaalrakkudele). Endokriinrakud paiknevad
seedekulglas difuusselt, kuuluvad difuussesse neuroendokriinsesse süsteemi,
mille rakke seedekulglas paarkümmend. Stiimul hormooni või
peptiidi vabanemiseks toimib uitnärviärrituse või GI endokriinsetel rakkudel
soolevalendikus olevate retseptorite toimel. Hormoonide regulatsioon toimub
seega eelkõige toidu koostisosade otsese kontakti kaudu endokriinselt aktiivsete
rakkudega vastavas soolelõigus, vähem hormooni (peptiidi) veretaseme kaudu.
GI hormoone võib vastavalt nende aminohapete järjestusele ja toimele jaotada
rühmadeks. Üks rühm moodustub gastriinist ja koletsüstokiniinist, mis omavad
ühes otsas samasugust aminohappelist järjestust, toimivad samadesse
3
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
rakuretseptoritesse, on sama toimega, kuid retseptorite tundlikkus ja toime
tugevus on erinev. Gastriin toimib mao parietaalrakkudele ja ECL-rakkudele
CCKB-retseptori vahendusel tugevamini kui koletsüstokiniin, viimane aga
põhjustab CCKA-retseptori kaudu sapipõiekontraktsiooni ja kõhunäärme
ensüümide väljutuse. Teise rühma moodustavad sekretiin, temaga sarnased VIP,
glükagoon, GIP; ka neil on samasugune järjestus polüpeptiidahelas. Need kaks
rühma võivad toimida antagonistlikult, aga lõppelundil on näidatud ka
sünergistlikke efekte. Pankrease näärmerakul on tõestatud ensüümide
sekretsiooni stimulatsiooni nii
gastriini ja koletsüstokiniini kui sekretiini ja VIPi
toimel; esimesed kaks toimivad üle intratsellulaarse Ca2+ kontsentratsiooni
suurenemise, teised üle
cAMP -i.
Sekretsioon järgneb mõlemal puhul. Peale
nimetatute on ka teisi rõhmi.
3.3. Reflektoorse ja humoraalse regulatsiooni üldised seaduspärasused. Tagasiside printsiip. Otsesed ja kaudsed mõjud efektorrakkudele.
Tagasiside on
regulatoorse funktsiooni tulemuse otsene või kaudne
ülekanne reguleerivasse ossa tagasiinformatsioonina. Negatiivse tagasiside
mehhanismi korral vastus ise tingib enda eemaldamise või inhibeerimise.
Positiivse tagasiside korral põhjustab initsieeriv stiimul sama protsessi
intensiivistumist.
Autonoomne närvisüsteem ja endokriinne/ parakriinne süsteem
seedekulglas interakteeruvad ja kontrollivad sekretsiooni, absorptsiooni ja
motoorikat . Sihtmärkrakkudel, olgu nad sekretoorsed, absorptiivsed või
silelihasrakud, on
retseptorid , millede kaudu saab nende rakkude tööd kontrollida
nii
neuraalset (toimub neuronite vahendusel.
Aksonit mööda leviv
aktsioonipotentsiaal põhjustab neurotransmitteri vabenemise) kui humoraalselt
(Hormoone
tootev näärmerakk salvestab toodetu granulaarsel kujul, ühes
graanulis on palju hormoonimolekule) vallandatud keemiliste ühendite abil.
Hormonaalne regulatsioon on aeglasem, võtab minuteid ja tunde.
Need neurokriinid, endokriinid ja parakriinid vabastatakse närvilõpmetest ja
näärmerakkudest
vastuseks erinevatele stiimulitele, mis on omakorda üle
retseptorite
toiminud . Nende
stiimulite allikas võib pärineda nii väliskeskkonnast,
kui ka organismist endast (hea näide on maosekretsiooni ajufaas, kus lisaks
lõhnale on ka ettekujutusel, st kortikaalsel mõjul toime näärmerakkudele). Paljudel
juhtudel on
stiimulid sihtmärkrakkude enda sekretsiooni tulemus. Olenemata
allikast, indutseerivad need stiimulid signaale, mis integreeritakse närvi- ja
endokriinsüsteemi kaudu nii, et sihtmärkrakkude funktsioon saab vastavalt
vajadusele reguleeritud. Neuraalne aktiivsus võib põhjustada hormoonide
vallandumist, hormoonid moduleerivad neuraalset aktiivsust ja efektorraku
aktiivsus saab mõjutatud nii neuraalsete kui hormonaalsete teede kaudu.
4.Seedeelundkonna talitluse uurimismeetodid
4.1. Eksperimentaalsed uurimismeetodid Eksperimentaalsete uurimismeetodite korral kogutakse andmeid katse
käigus, katsed viiakse läbi katseloomadel.
Akuutne katse, et uurida konkreetse organi funktsiooni
tervikuna ja
detailsena, ilma teiste süsteemide mõjuta. Muudetakse operatiivsel teel kas
elundite omavahelist normaalset talitlust või isoleeritakse teistest elunditest
osaliselt või täielikult. Elundi osaline isoleerimine ja perfuseerimine: elundil on
mingi kontakt organismiga olemas, ta võib olla toodud kõhuõõnest välja või on
organil kõhuõõnes
kamber ümber. Alles on jäetud üks toov ja üks
viiv veresoon ,
mida saab kontrollida ja nendesse saab manustada aineid, ilma et need läheks
süsteemsesse ringesse ja maksa. Isoleeritud soolelingude moodustamine, mille
4
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
käigus jäetakse mingi peensoole osa vahelt välja, kuid sellele saab alles jätta
verevarustuse mesenteeriumi kaudu. Soolelõigu ühe või mõlemad
otsad võib keha
pinnale välja tuua. Nii saab uurida imendumise regulatsiooni. Täielikult isoleeritud
elunditega töötamiseks peab organ asetsema toitelahuses, mille pH, osmootne
rõhk ja keemiline koostis on sarnane vereplasmale ja see peab sisaldama glükoosi.
Preparaat peab saama hapnikku, püsisoojase organi puhul peab temperatuur
olema 37˚C.
Kroonilise katse käigus
luuakse operatiivselt teatud mudel. Looma saab
korduvalt kasutada. Näiteks
fistul (uuris keha välispinna ja õõne vahel või ka
välispinna ja näärmejuha vahel, kus näärmejuha tuuakse välispinnale),
ösofagotoomia (ösofaaguse lõige, söögitoru otsad on kaelale välja toodud, suu
kaudu söödud toit makku ei
satu , vaid tuleb ösofagotoomia ava kaudu välja. Selle
abil uuritakse mao sekretsiooni ajufaasi), väikese mao
operatsioonid – maost
eraldatakse osa, kuid ei eemaldata (*
Pavlovi järgi: lõige läbi kõikide kihtide suurel
kurvatuuril, ühendus suure maoga jäetakse alles seal, kus maole tuleb vaaguse
haru.
Limaskest lõigatakse suurel kurvatuuril läbi, toit väiksesse makku ei jõua,
sellel on oma õõs ja limaskest.Väiksele maole lisatakse ka fistul; nii saab uurida
maosekretsiooni närviregulatsiooni ja on välditud kontakt mao limaskestaga.
*Heidenhaini järgi: väike magu on suurest maost eemaldatud. Parasümpaatiline
regulatsioon elimineeritakse ehk närvid on läbi lõigatud. Alles on veresooned,
millest sümpaatilist saab innervatsiooni, kuid see on näärmeregulatsioonis
tagasihoidlik. Saab uurida humoraalset mõju.). Gastroenteraalne anastomoos,
kunstlik ühenduse loomine mao ja peensoole/ söögitoru ja peensoole/ peensoole
erinevate osade vahel, nii et ei esine enam normaalset toidu passaaži.
Selgitatakse, milliseid muutusi seedekulglas passaaži muutus esile kutsub. Nende
katsete tulemusi saab kasutada inimeste sooletrakti
operatsioonide planeerimisel
(vähi või
haavandi tõttu). Gastroenteraalanastomoosid võivad olla ka akuutsed.
Eksperimentaalsete uurimismeetoditena kasutatakse ka seedenäärmete
stimulatsiooni, sest puhkeseisundis paljud näärmed üldse ei tööta. Stimuleeritakse
näiteks toiduga (see on nõrk ärritaja). Kui soovitakse maksimaalset stimulatsiooni,
kasutatakse mediaatoreid, elektristimulatsiooni, hormoone. Võib kasutada
organismis endas olevaid mediaatoreid ja nende sünteetilisi analooge, närvi
ärritamist alalisvooluga (soovitatav on ärritada vaaguse perifeerset könti, selleks
tuleks vaaguspõimik läbi lõigata => akuutne katse). Kasutatakse ka histokeemilisi
uurimismeetodeid. Ning kaudseid
meetodeid , et hinnata raku talitlust: Elundi/ raku
verevarustuse ja hapniku kasutamise ning ka glükoosi kasutamise hindamine.
Töötavas
elundis on nende kasutamine ja ka
verevarustus intensiivsem.
4.2. Kliinilis- funktsionaalsed uurimismeetodid.
Neid kasutatakse inimese seisundi muutuste iselomu ja
ulatuse selgitamiseks ning diagnoosi püstitamiseks. Röntgen, sageli kontrastainega, mis
röntgenkiiri läbi ei lase ja selle tõttu joonistuvad õõne kontuurid. Röntgeniga saab
üldise pildi, selle sügavust on raske hinnata, kompuutertomograafiga saab palju
võtteid eri tasapindadelt. Ultraheli võimaldab määrata kudede
tihedust ,
diagnoosida kasvajaid, tsüste, kive, võõrkehi. Samas, kui kudede tihedus pole
eriti muutunud, pole need meetodid eriti informatiivsed näiteks funktsioonihäirete
korral, mil suuremat koekahjustust pole veel kujunenud. Visuaalseks
diagnostikaks kasutatakse ka endoskoopiat, mille korral viiakse õõnde optilise
aparatuuuriga ühendatud sond. See võimaldab võtta ka biopsiamaterjali.
Seedeelundite talitluse hindamiseks kasutatakse
funktsionaalseid proove. Mao
ja kõhunäärme sekretsiooni hindamiseks tuleb näärmete tööd stimuleerida.
'Maonõre sekretsiooni hindamine
histamiini ja pentagastriini testide abil koos
maonõre happesuse ja peptilise aktiivsuse määramisega. Maonäärmete
töölepanekuks on vaja piisavalt tugevat stimulatsiooni, et kõik näärmed tööle
5
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
hakkaks (toidu peale kõik ei hakka näiteks), muidu ei tea kas mitte tööle
hakanud näärmed ei tööta mingi häire tõttu või nõrga stiimuli tõttu.
Kõhunäärme sekretsiooni hindamine sekretiini-, CCK/sekretiini- või Lundhi testi
abil. Viimasel juhul stimuleeritakse kõhunäärme talitlust proovieinega, milles on
5% valku, 6% lipiide, 15% süsivesikuid ja 74% mittesöödavat kiudainet. Siis on
vaja nõret koguda ja hinnata ensüümide aktiivsust.
Funktsionaalsed proovid kuuluvad enamuses otseste meetodite alla, mille käigus
võidakse määrata stimulatsiooni tulemusel sekreteeritud:
a)
pankreasenõre või duodenaalnõre kogust, kõhunäärmenõrega väljutatud
ensüümide ning HCO -
3 kontsentratsiooni ja üldist väljutust;
b)
pankreasenõre ensüümide kontsentratsiooni veres. Ägeda pankreatiidi korral
on pankrease ensüümide kontsentratsioon veres tõusnud.
c)
seedeensüümide kontsentratsiooni roojas. Üks tundlikkumaid meetodeid on
pankrease elastaas 1 (PE1) määramine roojas. PE1 ei lagune peensoole ega
mikroobide ensüümide mõjul ja selle sisaldust saab määrata
ELISA meetodiga
spetsiifilise antikeha abil. Meetod on pankrease välissekretoorse funktsiooni
puudulikkuse korral vastavalt raskusastmele 63...100% tundlikkusega.
Pankrease talitluse hindamise kaudsed meetodid põhinevad suu
kaudu manustatud ensüümsubstraatide lõhustusproduktide määramisel uriinis.
Ensüümpuudulikkuse korral on lõhustusproduktide sisaldus uriinis alanenud.
Bentiromiidtest põhineb ensüümi kümotrüpsiini võimel lõhustada peensooles
sünteetilist peptiidi N-bensoüül-L-türosiin/paraaminobensoehapet (bentiromiidi)
vabaks PABA-ks. PABA väljutust uriiniga mõõdetakse 6 h jooksul pärast 500 mg
bentiromiidi manustamist. Pankrease funktsiooni languse korral on PABA väljutus
vähenenud.
Üksikute toitainete lõhustumise ja imendumise hindamise testid:
Pankreolaurüültest kus suu kaudu manustatav sünteetiline fluorestsiindilauraat
lõhustatakse pankrease laurüülesteraasi toimel laurüülhappeks ja fluorestsiiniks,
mis imenduvad verre ja mida saab määrata uriinis.
Kaudsete proovide puuduseks on piiratud tundlikkus kõhunäärme kerge ja
mõõduka puudulikkuse korral., tulemuste mõjutatavus
kasutatavatest ravimitest,
pH-st, kõhulahtisusest ja seedekulgla operatsioonidest.
Peensoole funktsiooni uurimine : Ksüloositest peensoole (kaskteistsõrmiksoole ja tühisoole) funktsiooni
hindamiseks. Selleks manustatakse suu kaudu 25g ksüloosi. Normaalselt peaks
uriiniga väljutatama 5h jooksul enam kui 4g ksüloosi. Sellest väiksem kogus
viitab imendumishäiretele. Laktoositest laktoositalumatuse (laktaasi
defitsiidi )
kindlaks tegemiseks: suu kaudu manustatakse 50g laktoosi, mis lõhustub laktaasi
toimel galaktoosiks ja glükoosiks ja seega viimase tase peaks veres tõusma, kuid
seda ei juhtu laktoositalumatuse korral. Schillingi test imendumisfunktsiooni
hindamiseks niudesooles. Siin manustatakse
vereloome seesmist
faktorit koos
radioaktiivselt märgistatud
57Co -
vitamiin B12. Kaks tundi hiljem manustatakse
suu kaudu kobalamiini, mis peaks esile kutsuma radioaktiivselt märgistatud B12
väljutuse neerude kaudu ja kobalamiini puudujäägi kompenseerima. 57Co-
kobalamiini väljutuse langus alla 6% isotoobi suu kaudu manustatud kogusest on
tõendiks imendumishäiretest ileumis.
Süljenäärmete talitluse hindamiseks kasutatakse järgnevaid
mooduseid:
A) sülje kogumine katsealuselt katseklaasi. See on lihtne, kuid ei
võimalda hinnata sülje sekretsiooni kiirust.
B) Katsealune loputab suu,
kallutab pea
ettepoole ja
laseb süljel suunurgast välja tilkuda. See sobib süjle sekretsiooni
kiiruse hindamiseks, kuna suu
liigutustega süljenõristust ei mõjutata.
C) Sülje
kogumine süljenäärmest spetsiaalse kapsli abil, mis asetatakse vaakumi abil
6
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
süljenäärmejuha väljumiskohale ja on ühendatud katseklaasiga. Nii saab sülge
koguda söömise ajal ühest suurest süljenäärmest.
D) süljenäärme
juha kanüülimine. Inimesel kasutatakse seda meetodit harva, küll aga narkoosis
katseloomadel akuutses katses.
5. Seedimine suuõõnes
Suus toimuvad protsessid: 6 põhifunktsiooni
mälumine
niisutamine: teeb toidukämbu libedaks
neelamise algatamine
kaitsefunktsioon: tänu süljes sisalduvatele bakteritsiidsetele
ainetele ; ka
hambakaariese profülaktika
maitsemine: keelel 4 tüüpi maitseretseptoreid, sellele lisaks suu limaskestal
ka
termo - ja puuteretseptorid
rääkimisel suu limaskesta niisutamine
Mälumisel tahke toit purustatakse – selles tegevuses osalevad üla- ja alalõualuu
koos hammastega, vöötlihaseline mälumismuskulatuur, keel ja põsed, suupõhi ja
suulagi. Toiduosakeste puuteärritus suulael ja hammastel juhib reflektoorselt
mälumisliigutusi külgedele, ette ja taha, üles ja alla. Keel ja põsed hoiavad
toidupala mälumispindade vahel ja sees. Mälumise poolt stimuleeritud süljevoolus
valmistab toidupala kontsistentsi neelamiseks ette. Suus toimub ka maitsetaju, mis
omakorda põhjustab reflektoorselt süljevoolust ja maosekretsiooni.
5.1. Sülje koostis ja omadused Ööpäevas moodustub ~1...1,5L sülge. Sülge sekreteerivad väikesed
seinasisesed süljenäärmed:
gll. labiales et buccales, gll. palatinae et linguales ja
suured seinavälised süljenäärmed
gl. parotis (seroosnäärne),
gl. submandibularis (seroosnääre) ja
gl. sublingualis (ülekaalukalt mukoosnääre). Sülg moodustub
süljenäärmete aatsinusrakkudes, kus tekib esmassülg (vereplasmaga isotooniline),
millest liikumisel läbi määrmejuhade Na+ ja Cl- resorbeeritakse, K+ ja HCO -
3
sekreteeritakse.
Kiirema süljesekretsiooni korral ei jõua need protsessid väga
intensiivselt toimuda, mistõttu on sülg siis Na+, Cl- ja K+ kontsentratsiooni ja
osmolaarsuse poolest vereplasmale sarnane. HCO -
3 aktiivset sekretsiooni verest
sülge soodustab karboanhüdraas. Sülje pH on
puhkeolekus 5,45...6,06,
stimulatsiooni korral kuni 7,8. Seedeensüümidest esineb süljes lingvaallipaasi
(keele seroossetest näärmetest; tungib läbi piimarasva gloobuli membraani ja
hüdrolüüsib seal triglütseriidi kolmandas asendis olevat rasvhapet) ja α- amülaasi
(põhikogus
gl. parotisest, hüdrolüüsib α-1,4-glükosiidsidemeid). Lisaks on süljes
veel makromolekule: glükoproteiinid, mukopolüsahhariidid, lüsosüüm, IgA ja
veregrupi substantsid.
5.2. süljenäärmete talitluse regulatsioon Impulsid süljenäärmetele lähtuvad
piklikaju sekretoorsetest keskustest
(
nucl. salivatorius sup =>
n. facialis => harud süljenäärmetele va.
gl. parotis et
gll. buccales.
Nucl. salivatorius inf =>
n. glossopharyngeus => harud
gl. parotis’ele
ja
gll. buccales’le), aferentsed
signaalid tulevad
suust ja suulaelt (maitse,
puutumine) ning ninast (lõhn) ja kõrgematest keskustest (
ettekujutus ).
Puhkeolekus sekreteerib
gl. submandibularis 71% (limajas sülg),
gl. parotis 25%
(vedel sülg) ja
gl. sublingualis 4% sekreteeritud süljest, pärast stimulatsiooni
suureneb
gl. parotis’e osatähtsus (34%).
Veerikka sülje eritumist mõjutab kolinergiline, α- adrenergiline ja substants P
stimulatsioon , mis käivitab aktiivsema Ca2+ sisenemise tsütoplasmasse.
Kolinergiline stimulatsioon vabastab ensüüm kallikreiini, mis formeerib
vereplasma kininogeenidest vasodilatatoorse toimega bradükiniini. Vasodilatatsioon on põhiline
7
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
maksimaalse süljevooluse põhjustaja. Süljenäärmete β- adrenergiline stimulatsioon
põhjustab viskoosse sülje
eritumise , mis sisaldab rohkesti mutsiini (
gl. submandibularis’est ja
gl. sublingualis’est, aga mitte
gl. parotis’est). See esineb
inimesel kuiva toidu mälumisel ja stress-situatsioonis.
6. Söögitoru ehitus ja talitlus. Neelamine Söögitoru proksimaalne 1/3 on vöötlihaseline,
distaalsed 2/3 on silelihaseline ja
allub autonoomse närvisüsteemi
kontrollile .
Neelamisel eristatakse oraalset,
farüngeaalset ja ösofageaalset faasi,
kusjuures vaid esimene on tahtele alluv.
Toidukämp suus lükatakse keele ja suulihaste abil suuõõnes taha, neelupära
piirkonda. See ärritab sealseid retseptoreid ja vallandub reflektoorne neelamisakt.
Suulagi ja kontraheerunud palatofarüngeaallihased moodustavad suuõõne ja
ninaneeluruumi vahele seina. Kui keel toidupala tahapoole lükkab, tõuseb
kõrisõlm ja katkeb lühikeseks ajaks
hingamine . Epiglottise poolt suletakse
trahhea ja toidukämp libiseb üle epiglottise söögitorru.
Söögitoru (
oesophagus,
25...35cm) ülemine sfinkter avaneb ja toidukämp liigub edasi peristaltiliste
liigutustega.
Silelihaste kontraktsioonilaine (
peristaltika ) liigub lõõgastus ees,
kontraktsioon taga piki söögitoru. Seda juhitakse piklikajust. Söögitoru keskkosal on peale
elundivälise neuronaalse kontrolli ka
endogeenne regulatsioon. Alumisse
söögitoru piirkonda jõudes avaneb alumine sfinkter (kõrgrõhutsoon, mis tagab
makkupääsu sulgumise) ja kämp liigub edasi makku. Sulgurlihased sulguvad.
Söögitoru alumine sfinkter ja mao
kardia silelihased takistavad maosisaldise
refluksi. Et suurem osa söögitorust asub rindkereõõnes, on seal rõhk
atmosfärirõhust 4...6 mmHg madalam, ülemine söögitorusinkter takistab õhu
pidevat pääsu söögitorru. Söögitoru kontrollib põhiliselt
n. vagus.
7 Seedimine maos Maol on 3 põhifunktsiooni:
1)
söödud toidu salvestamine,
2)
toidu segamine ja peeneks jahvatamine,
3)
õigeaegne tühjenemine,
4)
Toitekäitumise regulatsioon.
Elundisisesed närvipõimikud paiknevad:
plexus submucosuse s. Meinssneri, see on submukoosas
plexus myentericus s. Auerbachi seedetraktis ring- ja pikilihaskihi vahel.
7.1 Maolimaskesta ehitus Magu on serooskihiga kaetud, maol on kolm lihaskihti: longitudinaalne,
tsirkulaarne ja põikikiht. Mao sisepinda moodustab silinderepiteel. Maonäärmetel,
mis suubuvad maolohukestesse on
topograafilised erinevused. Eristatakse:
kardia regiooni:
kitsas 1...4 cm laiune viirg allpool
ösofaguse sisenemiskohta. Pikad, tugevasti hargnenud
tubulaarsed näärmed.
Maopõhja (
fundus ) ja maokeha (korpus) regioon: ¾
maost. Näärmed on
sirged või vähe looklevad ja
voodertatud lima produtseerivate kõrvalrakkudega,
parietaalrakkudega (produtseerivad HCl ja
Castle ’i
faktorit) ja pearakkudega (ensüüme).
8
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
Endokriinsetest rakkudest esinevad ECL (enterokromatofiil-
like), EC
(enterokromatofiil), X /A-
like rakud, D- rakud.
Püüloruse (ja antrumi) regioon: võtab alla 15...10% maost ja sisaldab lihtsalt
hargnevaid tubulaarseid näärmeid, mis produtseerivad lima. Hapet
produtseerivaid rakke siin eriti ei ole.
Endokriinsed rakud on EC, G ja D rakud
7.2 Maonõre koostis ja omadused Parietaalrakud sekreteerivad HCl, K+ ja Castle’i faktorit, limarakud sekreteerivad
pidevalt Na+, K+, Cl-, HCO -
3 ,. Koos sekretsioonimahu tõusuga väheneb Na+
kontsentratsioon ja HCO -
3 kaob. Maomahla üks
komponent on ka kogu magu
kattev lima, mis kaitseb keemilise ja
mehhaanilise ärrituse eest ja mao enda
seedimise eest. Samuti sisaldab maomahl ensüüme.
Funduse piirkonnas produtseerivad parietaalrakud
HCl, mis
a) tekitab maos happelise keskkonna, vajalik valke lõhustavate
pepsinogeenide aktivatsiooniks,
b) omab
bakteritsiidset toimet
c) tekitab üleminekul
peensoolde seal happelise keskkonna, mille toimel
vabaneb peensoole limaskestas
sekretiin –
hormoon , mis stimuleerib
pankrease nõre eritumist ja hakkab pidurdama maohappe edasist
sekretsiooni.
Happeline keskkond peensooles vabastab peensoole ensüümi:
enterokinaasi (enteropeptidaas), mis muudab kõhunäärme
trüpsinogeeni aktiivseks
trüpsiiniks.
d) HCl osaleb toidu (maosisaldise - küümuse) üleminekus maost
peensoolde: kui peensooles tekib HCl mõjul happeline keskkond, pyloruse
sfinkter aheneb ja sulgub ja takistab järjekordse portsjoni üleminekut
peensoolde. (Õigeaegne tühjendamine)
ECL ja G-rakkude funktsioon HCl sekretsiooni regulatsioonis: G-rakke leidub
maos antrumis ja ECL rakke funduses. G-rakku stimuleerib ajufaasi vaagusärritus
ning maofaasis erinevad toidud-joogid (valkude hüdrolüsaadid, kofeiin,
lahjad alkohoolsed joogid). G-raku stimulatsioonil vabaneb gastriin, mis mõjub
parietaalraku HCl väljutusele stimuleerivalt. ECL-
rakk vabastab histmaiini, mis
H3-histamiintundliku retseptori kaudu enda produktsiooni negatiivselt
tagasisidestab ja H2-retseptori kaudu stimuleerib parietaalrakkude HCl
produktsiooni. Gastriin stimuleerib ECL-rakke. D-rakkudest vabanev
somatostatiin pärssib HCl produktsiooni.
Ensüümid maos => pepsinogeenid
=> lipaas
Pepsinogeenid on valkude lõhustamist algatavad ensüümid maos, mis pole aga
pearakkudes veel aktiivsed. Pepsinogeene on
7 isoensüümi (
pepsinogeen 1...7).
Pepsinogeenid 1...5 => molekulmass 42 000...43 000 D, toimeoptimum on pH
1,5...2,2. Kõik isoensüümid aktiveeruvad pH alla 7 korral, kuid
esmalt aktiveeruvad (mitte nii happelise pH juures) isoensüümid 6 ja 7. Aktiveerumisel
lagunevad pepsinogeenid pepsiinideks (35 000 D), väikesteks pepsiinideks
(~5000 D) ja inhibeerivaks valguks (2000...3000 D) = 42 000...43000.
Pepsinogeenid 1...5 tuntud ja pepsinogeen 1 või
pepsinogeen A nime all,
toimeoptimum, nagu
eelpool mainitud , pH 1,5…2,2.
9
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
Pepsinogeenid 6 ja 7 omavad optimaalset toimet pH 3...3,2 juures, tuntud ka kui
pepsinogeen C või
gastriksiin. Osa pepsinogeene võib pärineda ka duodenumi
Brunneri näärmetest, suurem osa aga antrumi
piirkonnast , veidi ka funduses.
Pepsinogeen B all mõeldakse pepsinogeene, mis kuuluvad pH optimumi poolest
6 ja 7 hulka, aga pärinevad peensoolest.
Mao
lipaasid ei vaja sapi olemasolu, selleks et rasvu emulgeerida. Toimib
piimarasvadele maos, seetõttu on tema tähtsus
lapseeas suurem, kui
sappi veel
ei produtseerita; sapi aktiivsus tekib esimeste aastate vältel.
Mutsiin (lima glükoproteiin), mida produtseerivad limarakud omab mao limaskesta suhtes
kaitsefunktsiooni. Lima
tekkes on oluline roll
prostaglandiinidel (PGE2). Ained (näit.
Aspiriin ), mis selle teket pärsivad, pärsivad ka lima teket. Mao limaskesta kaitseb H+ ioonide
vastu aktiivne HCO -
3 sekretsioon. Prostaglandiinid soodustavad
HCO3 sekretsiooni. Kui lima
ei piisa, siis võivad tekkida maos
haavandid ülehappesuse tõttu.
Vereloome sisemine faktor e. Castle’i faktor on vajalik
vitamiin B12 imendumiseks
peensooles. Vereloome väline faktor vabaneb lihavalkudest maohappe mõjul ning see peab
liituma maost pärineva sisemise vereloomefaktoriga ja kitsas peensoole osas on vastav
retseptor , mis seob kompleksi. See on vajalik vitamiin B12 imendumiseks enterotsüüti, kus
kompleks laguneb ja vitamiin B12 läheb vereringesse. Parietaalrakkude kahjustuse korral
(esineb täielik alahappesus) sisemist vereloomefaktorit ei toodeta ja tekib pernitsioosne e.
pahaloomuline aneemia.
Ghreliin. Põhiliseks ghreliini tekkekohaks on mao limaskesta endokriinsed X/A-sarnased
rakud. Ghreliini vallandumine
mao limaskestas suureneb nälgimisel ja väheneb söömisel.
Tühja kõhuga on inimesel vereplasma ghreliinisisaldus kõrgem, söönult madalam.
Rasvarikas
dieet langetab maos ghreliini teket ja sekretsiooni, mis kinnitab ghreliini
tundlikkust toiduga saadud energiahulgale. Ghreliin suurendab isu ja kehakaalu.
7.3 Maonäärmete talitluse regulatsioon 7.3.1. Stimulatsiooni ja pidurduse mehhanismid Funduse piirkonnas
histamiini produtseerivad ECL rakud, antrumi piirkonnas on
somatostatiini produtseerivad D-rakud ja
gastriini produtseerivad G-rakud.
Regulatsioonis on oluline parasümpaatiline närvisüsteem.
Sihtmärk: parietaalrakk.
Stimuleerivad retseptorid
Kolinoretseptor (M3 muskariintundlik), mida stimuleerib atsetüülkoliin (ACh),
mis vabaneb parasümpaatiliselt närvilõpmelt.
Histamiinitundlik H2 retseptor. Histamiin vabaneb parietaalraku
naabruses asuvast ECL rakust, parakriinne toime läbi ekstratsellulaarruumi. H3 retseptor
asub ECL raku peal ja on autoretseptor, mille kaudu histamiin hakkab iseenda
vabanemist pidurdama.
CCK-B retseptor on gastriinile tundlik retseptor ECL rakul ja avaldab selle kaudu hapet
stimuleerivat mõju, sest ECL rakk vabastab siis histamiini.
(Koletsüstokiniin on keemiliselt gastriinile väga sarnane. Tegemist on ühise retseptoriga,
mille alaliigid on erineva tundlikkusega. B-retseptor on tundlik gastriinile. Peensooles, ajus,
vaagusel olev CCK-A- retseptor on koletsüstokiniinile tundlikum.)
Pidurdavad retseptorid
Somatostatiin- retseptor
Prostaglandiin E2 retseptor Somatostatiin ja prostaglandiin E2 pidurdavad happeproduktsiooni. Happe üleproduktsioon
hakkab pärssima gatriini ning suureneb somatostatiini sekretsioon.
MILLAL JA MILLE MÕJUL VABANEB ACh, HISTAMIIN JA GASTRIIN?
10
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
Protsessi käivitaja on tavaliselt
n. vagus’e ärritus, mis tekib juba toidu nägemisel,
lõhna tundmisel, toidule mõtlemisel (
ajufaas). Uitnärvi ärritus tugevneb, kui toit
on suus, ärritus läheb
aferentseid kiude mööda piklikajusse. Vaaguse ärritus
tugevneb ka mao limaskesta ärrituse mõjul (maos on kemo- ja
mehhanoretseptorid). Piklikust
ajust toimub regulatsioon ja impulss läheb
eferentseid kiude mööda tagasi makku (Maofaas).
Uitnärvi kaudu ärritatakse
parietaalraku M3 retseptorit (stimuleerib happe sekretsiooni). Ärritatakse ka
ECL rakku, mis hakkab
histamiini sekreteeerima ja histamiin mõjub
parakriinselt parietaalraku H2 retseptorile ja sellega happe produktsioonile
stimuleerivalt. ACh pärsib D- rakke, ka antrumi D- rakkudes pärsitakse
somatosatiini sekretsiooni. D- ja G- rakud on vastastikkuses
retsitprookses
suhtes: ühe
erutus surub teist alla. Vaaguse kaudu reguleeritakse G- rakke
gastriini produtseerima, see toimub järgmiselt: vaaguse lõpmetelt vabaneva
ACh mõjul vabaneb GRP-ergiliselt vaheneuronilt gastriin-riliising
peptiid GRP, mis
mõjub parakriinselt G-rakule ja vabaneb gastriin. Gastriini teket vallandavad ka
teatud toidukomponendid (valgud ja rasvad). Gastriin soodustab üle
parietaalraku CCK-B retseptori happe sekretsiooni.
Endokriinsed rakud võivad olla lahtist tüüpi ja kinnist tüüpi, antrumi piirkonna G-
rakud on
lahtist tüüpi rakud, neid saab erutada ka ilma mediaatorita, mõjuvad
osaliselt lõhustatud valkude
produktid => lihapuljong. Ka kofeiini sisaldavad
joogid, kohvi, tee ja õlu ning vein (valge ja punane).
Gastriin läheb verre, sealt ka funduse piirkonda, kus stimuleerib histamiini teket,
mis mõjub parietaalrakule, aga gastriin mõjub ka otse parietaalraku
CCK-B retseptorile.
(vt skeem konspekti lõpus)
Sekretsioonifaasid:
Maol esineb ka basaalsekretsioon 2...3 mmol H+ 1 tunni joolsul. Maksimaalse
stimulatsiooni korral võidakse väljutada aga 10...35 mmol H+/h.
Ajufaas: ärritajad on toidu lõhn, maitse, nägemine, mõtlemine toidust. Mao
sekretsiooni stimuleeritakse uitnärvi kaudu.
Maofaas: ärritajaks on
venitus , mida vahendavad pikk leviku tee aju kaudu ja ka
lokaalsed refleksid enteraalse närvisüsteemi kaudu, mis soodustavad mao
sekretsiooni. Happe sekretsiooni pidurdab antrumi piirkonna pH öökimised.
Oksendamine võib ette tulla, kui mao kardia ja ülemine söögitoru sfinkter
lõõgastuvad ilma piklikaju erutuseta. Maosisaldis tavaliselt sel juhul suhu ei jõua
ja tegemist on
refluksiga.
Oksendamisega kaasneb peensoole ülaosa kontraktsioon (kuigi seda ei pruugi
alati olla). Osa soolesisust satub sel juhul makku. Kestva
oksendamise korral
satub makku ka
sapp ,
sapi lisandumine on seisundi raskuse näitaja.
Oksendamisega kaasneb tõsine
happe ja vee kaotus. Veekaotusega väheneb plasma maht
ja vere
viskoossus suureneb, mis võib põhjustada täiendava lisakoormuse südamele, kuna
takistuse vere liikumisele on suurem. pH ja plasma vedeliku hulk
taastuvad tavaliselt küllalt
kiiresti neerude ja kopsude töö tulemusel. Vedelikku tuuakse verre teistest kudedest selleks,
et viskoossust taastada. Selleks et vedeliku hulka taastada, tuleb oksendamise korral juua,
raskematel juhtudel tuleb rakendada tilkinfusiooni, et
korrigeerida vee ja elektrolüütide
tasakaalu.
8. Seedimine peensooles
Peensool on väljastpoolt kaetud peritoneumiga ja serooskihiga (
tunica serosa),
järgneb lihaskest (
tunica muscularis), kus on longitudinaalne (
stratum longitudinale) ja tsirkulaarne (
stratum circulare)
lihaskiht ja mukoosa e.
limaskest (
tunica mucosa), milles on limaskesta lihaskiht (
lamina muscularis mucosae). Piki- ja tsirkulaarlihaskihi vahel paikneb
plexus myentericus ja
ringlihaskihi ning submukoosa vahel paikneb
plexus submucosus. Limaskesta
pinda suurendavad mukoos- ja submukoosvoldid,
hatud ja
mikrohatud .
Viimased moodustavad enterotsüütide valendikupoolsel välispinnal harjasäärise. Peensoole
limaskestas esinevad äärisrakud, mis on kõige arvukamad hattudel ja
funktsioneerivad absorbeerivate rakkudena;
karikrakud , mis on krüptide ülaosas,
raku ülaosas on limatilk; Panethi rakud krüptide põhjas; Endokriinrakud
peamiselt krüptide alumises kolmandikus, aga ka hattudel. Endokriinrakkude
retseptorid on valendiku pool, oma
peptiidhormoonid (sekretiin, CCK, motiliin,
13
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
GIP, serotoniin jt) vabastavad nad verre, samas CCK toimib ka uitnärvi CCK-A
retseptorile ning esineb ka lokaalseid toimeid. Peensool jagatakse kolme ossa:
duodenum (20...30 cm),
jejunum (1,5...2,5 m) ja
ileum (2...3 m). Peensoole
kogupikkus
in vivo on 4 m,
post mortem 6-8 m. Peensoole imendamisfunktsiooni
arvestades vajalik hea verevarustus pärineb
a. mesenterica sup-st, duodenumil
a. ceoliaca, iileumi lõpposas ka
a. mesenterica inf.
8.1 Peensooles toimuvad protsessid, selles osalevad nõred. Peensoole osa seedeprotsessi regulatsioonis. Peensooles toimub:
1) küümuse segamine pankrease, maksa ja peensoolelimaskesta sekreetidega.
2) toidu koostisosade seedimine.
3) seeditud sisaldise
resorptsioon .
4) järelejäänud sisaldise edasitransport distaalsemale.
5) mitmesuguste hormoonide sekretsioon.
6) immunoloogiline kaitsefunktsioon (M- rakud, mis kuuluvad limaskestaga
assotsieerunud koe hulka (GALT)).
Peensoole nõre sisaldab elektrolüüte (HCO -
3 ). Ensüüme peensoole
epiteelrakkude sekreteeritavas nõres pole. Toidu seedimiseks vajalikud ensüümid
asetsevad enterotsüütide mikrohattude pinnal (aminopeptidaas, enteropeptidaas,
dipeptidaasid, maltaas, laktaas, sahharaas). Peensoolde (duodeenumisse)
avanevad pankrease- ja sapijuha. Pankreasenõre sisaldab ohtralt seedeensüüme:
proteaase, α- amülaasi, lipaasi, samuti HCO -
3 ja teisi elektrolüüte. Sapp aga on
vajalik lipiidide emulgeerimiseks, sisaldades vett, elektrolüüte, bilirubiini,
steroidhormoone, sapphappeid, letsitiini jt. aineid.
Peensoolel on oluline osa ka
seedeprotsessi regulatsioonis. Duodenumi (ka
mao) limaskestas on osmo- ja
kemoretseptorid , mis on tundlikud
soolasisaldusele, pH-le ja osmootsele rõhule. Samuti on seal toidu
hüdrolüsaatidele tundlikkud retseptorid. Maosisaldis on tavaliselt hüpertooniline ja
muutub duodeenumis veelgi hüpertoonilisemaks tänu ensüümidele, mis
peensooles lõhustamist läbi viivad. Kui maost saabub uus küümuse ports,
muutub pH duodenumis tugevalt happeliseks.
Mao tühjenemist pärsivad:
hüpertooniline keskkond
duodenumis; CCK,
GIPi ja sekretiini kaudu
(jejunumi ja iileumi piirkonnas on toidu hüdrolüsaatidele tundlikud retseptorid).
duodenaal-pH langemine alla 3 põhjustab püüloruse lihaste kontraktsiooni.
Kui
hape on neutraliseeritud ja pH liigub aluselisuse suunas, avatakse taas
duodenaalsfinkter.
aminohapped ja peptiidid
peensooles => CCK kaudu, mis vabaneb eriti
valgu hüdrolüsaatide toimel. (Toimib I rakkudest).
rasvhapped ja monogütseriidid, toime GIP ja CCK kaudu.
Kui peensooles on liiga palju valgu või lipiidide lõhustumisprodukte, siis
maost uut portsjonit juurde ei lasta ja peensooles jõutakse eelmised lammutada,
imendada ja edasi transportida.
HCl mõjul vallandub peensoole limaskesta S- rakkudest sekretiin, mis stimuleerib
pankrease sekretsiooni (rohkem juha rakkudest elektrolüütide sekretsioon), kuid
soodustab mao D- rakkudest somatostatiini vabanemist ja sellega pidurdab
happesekretsiooni maos. Toidu hüdrolüsaatide mõjul vabaneb peensoole I-
rakkudest koletsüstokiniin, mis stimuleerib samuti pankrease sekretsiooni (CCK-A
retseptorite kaudu), kuid mõju on suurem aatsinuse rakkudele (ensüümide
produktsioon ). Samuti põhjustab peensoolest vabanev koletsüstokiniin (üle CCK-
A retseptorite) sapipõie kontraktsiooni ja
sphincter Oddi lõõgastuse, et sapp
pääseks peensoolde. Peensoolest vabanev CCK on ka küllastustunde
tekitaja (lühiajalise toimega signaalmolekul), seega osaleb toitekäitumise regulatsioonis.
14
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
Siin vabaneb toidu hüdrolüsaatide mõjul ka GIP, mis samuti soodustab
somatostatiini produktsiooni ja vabastab ka insuliini. Peensoole GI rakud
produtseerivad gastriini intestinaalses faasis ja mis soodustab püüloruse
kontraktsiooni ehk pärsib toidu üleminekut duodeenumisse. Seega on peensoolel
oluline roll seedeprotsessi regulatsioonis, aga ka süsteemsemad toimed, mis
realiseeruvad tänu peensoole limaskestast vabanevatele hormoonidele.
8.2. Kõhunäärme välissekretsioon Kõhunäärme välissekretoorne osa produtseerib pankrease nõret, mis osaleb
seedeprotsessis. Eksokriinne pankrease nõre sisaldab orgaanilisi ja anorgaanilisi
aineid. Pankreasenõre tekkekohaks on
aatsinuserakud, kus tekivad ensüümid,
ning pankreasejuhade
epiteelrakud , kust vabanevad
ioonid ja vesi. Valke
lõhustavad ensüümid toodetakse inaktiivsena, sellega on välditud pankrease
iseseedimine. Pankreasenõre pH 7,8...8,4.
8.2.1. Kõhunäärmenõre koostis ja omadused, ensüümide aktivatsioon Pankrease nõre anorgaaniline osa koosneb veest ja ioonidest HCO -
3 , mis on
olulised peensoole sisaldise neutraliseerimisel. Oluline on ka Ca2+, seda näiteks
amülaasi aktivatsiooniks peensooles ja valke lõhustavate ensüümide
aktivatsioonis. Pankreasenõre orgaanilise osa ensüümid on valdavas enamikus
hüdrolaasid. Ülekaalus on proteolüütilised ensüümid (peptidaasid), mida võib
kaheks jagada:
1) Valgumolekuli ahelasiseseid peptiidsidemeid lõhustavad endopeptidaasid
(trüpsiin, kümotrüpsiin, elastaas, kollagenaas).
2) Ahela otsmisi sidemeid lõhustavad eksopeptidaasid (karboksüpeptidaasid,
aminopeptidaas).
Pankrease α- amülaas lõhustab tärklise ja glükogeeni tri- ja disahhariidideks.
Pankrease lipaas on üks võimsaim neutraalrasvu hüdrolüüsiv ensüüm, mis
lõhustab triglütseriidid monoglütseriidideks ja vabadeks rasvhapeteks.
Pankreasenõre nukleaasid, fosfodiesteraasid, nukleotidaasid ja nukleosidaasid
lõhustavad nukleiinhappeid.
Ensüümide aktivatsiooniprotsessis on oluline
trüpsinogeeni aktiveerumine trüpsiiniks peensoole seinast happelises keskkonnas
(maohappe mõju) vabaneva enterokinaasi (enteropeptidaasi) toimel. Tänu
toimele peptiidsidemetele eraldub trüpsinogeenist inhibeeriv valk ja tekib trüpsiin,
mis ka ise jätkab edasist trüpsinogeeni aktivatsiooni trüpsiiniks. Trüpsiini toimel
aktiveeritakse ka teised ensüümid (prokarboksüpeptidaas => karboksüpeptidaas,
elastaasi aktivatsioon jt, pms valke lõhustavad ensüümid). Lipaas, amülaas ja
ribonukleaas sekreteeritakse juba aktiivsel kujul.
8.2.2 Kõhunäärme välissekretsiooni regulatsioon
Aatsinuserakud: neid mõjutatakse otse tänu retseptoritele nende pinnal (on ka
kaudseid mõjutusi). Esineb kolinoretseptoreid, CCK- retseptorid (CCK-A), mida ka
gastriin ärritab, kuid vähem; GRP retseptor, substants P retseptor. Nendele
neljale retseptorile
ligandi seostumine käivitab intratsellulaarse IP3 ja Ca2+
vabanemisega vahendatud protsessid. (rakumembraanil aktiveeritakse fosfolipaas
C => IP3 => Ca2+ => ensüümi sekretsioon) ja see soodustab ensüümide
produktsiooni. On ka sekretiini ja VIP’i retseptor, mida on kaks alatüüpi, üks
rohkem tundlikum sekretiinile, teine rohkem VIP suhtes. Sekretiin ja VIP
käivitavad reaktsiooni cAMP alusel. Sekretiin ja VIP potenseerivad eelpool
nimetatud ensüümide sünteesi protsesse, kuid samas mõjuvad pankrease juha
epiteeli rakkudele ja stimuleerivad nõre vedela osa ja HCO -
3 sekretsiooni.
Pankrease aatsinuserakkudele toimib
n. vaguse lõpmete kaudu atsetüülkoliin,
15
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
kuid vaaguse närvilõpmetes on transmitterina kindlaks tehtud ka VIP. Uitnärvi
ärrituse vastuseks produtseeritakse ensüümiderikast sekreeti.
Sekretsioonifaasid:
Pankreasel esineb ka puhkeolekus basaalsekretsioon (vesinikkarbonaadi väljutus 2...3 ja ensüümide
väljutus 10...15% maksimaalsest).
Ajufaas Pankrease puhul pole nii määrav, kuid käivitavaks faktoriks on toidu lõhn,
nägemine, söömine. See põhjustab vaaguse kaudu ACh vabanemise ja
aatsinuserakkudes on selle tarvis
kolinoretseptorid . Vaaguse ärritumisel vabaneb
maos gastriin, mis jõuab ka pankreasse. Aatsinuserakkudel on CCK ja gastriini
tarvis ühine retseptor, nii stimuleerib ka gastriin ensüümide vabanemist (kuigi
toime CCK omast nõrgem). Vaaguse lõpmetelt vabaneb ka VIP.
Maofaas Stiimuliks on toidu mõjul mao venitus, samuti toidu hüdrolüsaatides olevad
aminohapped. Pankreast stimuleerivad gastriin ning mao ja pankrease vahelised
vagovagaalsed refleksid (parasümpaatikuse osavõtul
tekkivad refleksid. Vastava
ärrituse tagajärjel edastatakse parasümpaatiliste aferentsete närvikiudude ärritus
mao/pankrease parasümpaatilise närviraku kehale, kust lähevad eferentsed kiud
mao-pankrease suuna puhul näiteks aatsinuse rakku ja stimuleeritakse
pankreast).
Soolefaas Käivitub peensooles aminohapete, peptiidide, rasvhapete, monoglütseriidide, HCl,
Ca2+ mõjul. Erinevad toidu hüdrolüsaadid käivitavad erinevate hormoonide
vallandumise. Peensoole S-rakkudest vabaneb HCl mõjul sekretiin, mis
stimuleerib pankreasenõre anorgaanilise komponendi sekretsiooni. I- rakkudest
vabaneb hüdrolüsaatide toimel CCK => pankreasenõre ensüümide sekretsiooni
stimulatsioon. Esinevad vagovagaalsed refleksid: peensoole sein → pankreas ning
enteropankreaatilised refleksid.
8.2.3 Kõhunäärme sise- ja välissekretoorse osa morfoloogiline ja funktsionaalne seos
Kõhunäärme sise- ja välissekretoorne osa omavad omavahel
seoseid . Selle
seose olemasolule
viitavad mitmesugused funktsiooni muutused insuliini
alaproduktsiooni korral pankrease välissekretoorses osas. On teada
morfoloogiline ja füsioloogiline seos. Morfoloogiline seos:
vereringe omapära.
Osadelt pankrease saarekestelt (endokriinne osa) lähtuv kapillaaristik viib
venoosse vere vahetult eksokriinsetele rakkudele, seda nimetatakse
insuloatsinaarseks portaalsüsteemiks. Selline süsteem lähtub väikestelt
saarekestelt, suurtelt saarekestelt lähevad hormoonid otse verre.
Eksperimentaalse diabeediga loomadel stimuleerib
insuliin aatsinuserakus
valgusünteesi ja suurendab rakust sümogeensete graanulite (seal sees asuvad
hormoonid) tühjenemist. Eriti stimuleerib insuliin amülaasi väljutust.
Eksperimentaalse diabeediga loomadel oli just amülaasi süntees alanenud.
Insuliini korrapärane manustamine taastas amülaasi sünteesi ja sekretsiooni.
Pankrease eksokriinsetel aatsinusrakkudel on
spetsiaalsed insuliini ja CCK
retseptorid. CCK ja insuliin stimuleerivad aatsimuserakus valgusünteesi.
Insuliinipuudulikkusega diabeedihaigel on
häiritud amülaasi süntees.
8.3 Maksa funktsioonid
1. Detoksikatsioon . Organismis tekkivate mitmesuguste ainevahetusproduktide
kahjutuks tegemine, samuti sissevõetavate võõrainete kahjutustamine.
Mürkained jõuavad maksa tänu portaalvereringele: seedekulglast tuleb veri
v. 16
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
porta kaudu maksa, sealt
v. hepatica kaudu
v. cava inferior’isse.
2. Glükogeeni süntees ja deponeerimine 3. Eksokriinne funktsioon. Sapi süntees ja sekretsioon. See toimub maksa
rakkudes.
Maksas sünteesitakse primaarsed
sapphapped : koolhape ja
kenodesoksükoolhape.
4. Valkude desamiinimine. 5. Valkude transamiinimine. 6. Vereplasma valkude süntees. Fibrinogeeni, protrombiini ja albumiinide
süntees. Albumiinide sünteesi languse korral (tsirroos) langeb onkootne rõhk ja
vesi tõmmatakse rakkudest rakuvahelisse ruumi, sageli ka kõhuõõnde (
astsiit tsirrooside ja kasvajate korral).
7. Vitamiinide deponeerimine ja süntees. Maksas deponeeritakse A, B12, D,
E, K. D- vitamiini muutmine hormoonvormiks leiab aset maksas.
8. Ammoniaagi kahjutustamine. NH4 muudetakse: a) CO2 ühinemisel
karbamiidiks ja viikase uriiniga välja, b) tehakse glutamiini, mis on lämmastiku
depoo maksas ja ajus, c) NH4 seotakse α-ketoglutaarhappega ja tekib
glutamiinhape.
9. Rasvade metabolism. Maksas sünteesitakse kolesterooli ja fosfolipiide.
Organismis ringlevad rasvhapped lähevad maksa, kus nad muudetakse
triglütseriidideks ja need lipoproteiinideks, sel kujul saab neid vereringes
transportida ja neid saab rasvkoerakkude membraanide juures uuesti
rasvhapeteks muuta ja lipoproteiini lipaasi toimel rasvkoesse viia. Rasvhapetest
saab maksas ka CoA-SH kaudu energiat toota.
8.4. Sapi koostis, ülesanded, teke ja väljutuse regulatsioon
Maksas sünteesitakse primaarsed sapphapped: koolhape ja
kenodesoksükoolhape.
Peensooles sünteesitakse sekundaarsed sapphapped: desoksükool- ja
litokoolhape. Sapis on sapphapped konjugeeritud glükokooli ja tauriiniga. 4
sapphapet on liitunud: glükokoolhape, glükodesoksükoolhape, taurakoolhape,
tauradesoksükoolhape. Sapp sisaldab sapipigmente, mis annavad sapile
iseloomuliku värvuse.
Bilirubiin annab kollaka värvuse, kontsentreerumisel
sapipõies muutub sapi värv rohekaks. Uriiniga tuleb välja sapipigment urobiliin, mis
annab kusele kollaka värvuse. Sapp annab roojale kollakas– pruunika värvuse.
Patoloogiate korral, kui sapp ei jõua sapiteedesse (probleem maksa juures), on
kusel tume värvus, roe aga savikarva. Sapis on veel kolesterooli, mida
sünteesitakse maksas ja läheb sapi koosseisu. Samuti on sapis letsitiini. Sapi pH on
maksasapil 7,3...8 ja sapipõies 6...7, Ööpäevas tekib 500...1500 ml sappi. Teke on
pidev, teket stimuleerib sekretiin ja gastriin. Sapi väljutust sapipõest stimuleerib
CCK, mis vabaneb, kui rasvarikas toit on jõudnud peensoolde. Sapp aktiveerib
lipaasi, mis on vajalik rasvade emulgeerimiseks ja mitsellide moodustamiseks.
8.5. Peensoolenõre koostis ja toime. Valendikusisene ja
kontaktseedimine
Peensool produtseerib pidevalt bikarbonaadi ja mustiinirikast sekreeti.
Karikrakud (duodenumis Brunneri näärmed) produtseerivad mutsiini, mis katab
geelitaoliselt epiteeli. Mutsiin kaitseb peensoole epiteeli proteaaside toime eest,
duodeenumis maomahla toime eest ja lihtsustavad toidukördi läbiminekut
peensoolest. Peensoolekrüptide pearakud sekreteerivad plasmaga isotoonilist
NaCl lahust. Duodenumi Brunneri näärmed produtseerivad mutsiini ja
bikarbonaadirikast aluselist sekreeti, kusjuures ka siin toimub HCO -
3 sekretsioon
luumenisse tänu HCO -
3 vahetusele Cl- vastu. Soolesekreedis ensüüme ei esine,
17
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
küll aga on enterotsüüdi pinnal lokaliseeritud ensüümid, mis valendikus oma
toime kaotaksid. Luumenisse võib sekundaarselt ensüüme sattuda
mukoosarakkude uuenemise käigus, kui irdunud mukoosarakud koos mikrohatu
pinnal olevate ensüümidega luumenisse langevad. Peensooles on
valendikusisene seedimine, mis toimub keset soolevalendikku kõhunäärme
ensüümide ja anorgaaniliste komponentide osavõtul.
Kontakt(membraan-)seedimine toimub peensoole seinaga vahetus
kontaktis olevate ensüümidega, küümus peab olema vahetus kontaktis peensoole seina
ensüümidega hariäärisel. Laktaas, maltaas sahharaas, enteropeptidaas- need
ensüümid ei lahku hariääriselt (vastasel korral kaotavad oma aktiivsuse). Et
küümus ei oleks pidevalt sama ensüümiga kontaktis, ongi vajalik küümuse
liikumine.
8.6 Peensoole motoorika liigid ja regulatsioon
Esineb kahte liiki motoorikat:
Propulsiivne => küümuse transport,
mittepropulsiivne => küümuse
segamine. Peensoole motoorikat juhib basaalne müogeenne rütm: aeglased
lained. Aeglaste lainete ostsillaatoritel on peensoole algusosas suurem sagedus
(12 tüklit/min), lõpposas väiksem (8 tsüklit/min). Nii toimub soolesisaldise
aeglane edasiviimine. Interdigestiivses
perioodis esineb peensooles propulsiivne
migreeruv müoelektriline kompleks (MMK). Selle faasid:
täielik rahu 40...60 min
korrapäratu motoorika 30 min
korrapäraste elektriimpulsside vahendatud motoorika. 15 min
MMK algab antrumist ja peensoolest liigub järjest edasi jämesooleni, kus
kindlaid faasidevahelisi erinevusi pole. Elektriline aktiivsus on igal juhul kogu aeg olemas,
aga võib olla nii nõrk, et igale impulsile ei järgne kokkutõmmet. Motoorse rahu
korral ei järgne impulsile üldse kokkutõmmet. Korrapäratu motoorika korral
järgnevad ka impulssidele kokkuõtmbed, kuid on ka nõrku impulsse, millele
kokkutõmbeid ei järgne. Korrapärase motoorika faasis järgneb igale impulsile
kokkutõmme.
Migreeruvat müoelektrilist kompleksi reguleerib hormoon
motiliin ja sellele
lähedane aine
ghreliin, mis avastati alles 1999. Motiliinil on põhitoime
motoorikale, ghreliinil mõju aga toitekäitumisele. Motiliin toimib teatud
parasümpaatilise fooni taustal. Kui inimene sööb kasvõi lühiajaliselt
interdigestiivses perioodis, siis sõltumata faasist, MMK kohe katkeb.
Söömise ajal, digestiivses faasis esinevad järmised liigutused:
Rütmiline segmentatsioon : lähedaste soolesegmentide ringlihased tõmbuvad
kokku ja küümus liigutatakse kahele poole. Tomub küümuse segamine.
Pendelliigutused: pikikiudude kontratktsioon kutsub esile soolelõigu pikkuse
muutuse (8...
15cm pikkune lõik), samas on kõrval pikikiudude lõõgastus. Nii on
soolesisu soole suhtes liikunud.
Funktsiooni muutus toimub
gastriini ja
koletsüstokiniini vahendusel.
Peensooles toimuvad liigutused ka hattude tasandil. Küümuse keskmine
liikumiskiirus peensooles on 1...4 cm/min. 2...10 h jooksul jõuab küümus
caecum ’isse.
Rasvasem toit liigub aeglasemalt. Kõige kiiremini liigub
süsivesikuterikas toit.
Peensoole motoorse aktiivsuse seisukohalt on oluline
plexus Auerbachi.
Atsetüülkoliin avaldab ringlihaskihile pidurdavat toimet, samas kui pikilihaskihile
avaldab see stimuleerivat toimet. Sümpaatikus põhjustab
plexus myentericus’es
motoorika pidurdumise, samas parasümpaatikus (
n. vagus) põhjustab selle
18
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
aktiveerumise.
Söömisvaheaegses perioodis on kontraktsioonid tugevamad, kui söömise ajal.
Refleksid:
Intestino-intestinaalne
refleks : ühe peensoole osa liigne venitus lõõgastab
ülejäänud osa peensoolest.
Gastro- ileaalrefleks: mao suurenenud motoorika ja sekretoorne aktiivsus
suurendavad iileumi lõpposa motoorset aktiivsust ja kiirendavad küümuse
üleminekut jämesoolde läbi ileotsökaalsfinkteri.
Iileumi venitus vallandab peptiid YY iileumist, mille mõjul (vere kaudu maole ja
sfinkteritele) aeglustub mao tühjenemine. Nende reflekside käivitaja on
mao/peensoole venitus.
Ileotsökaalüleminek Ileumi lõpposas on 4cm pikkune segment: sfinkter, mis on tavaliselt suletud. See
lõõgastub
ileumi lõpposa venitusel ja laseb järjekordse portsioni jämesoolde.
Tema toonus tõuseb, kui
caecum’i rõhk tõuseb ja ei lase
ileumi sisaldist
caecumisse üle minna.
Ileumi ja
caecumi vahel on ka ileotsökaalklapp (
valva ileocaecalis s Bauhini).
9. Imendumine 9.1. Imendumisprotsessi üldised seaduspärasused Imendumine on protsess, milles seedimise lõpp-produktid ja vesi, vitamiinid,
mineraalained,
mikroelemendid võetakse soolevalendikust läbi soolelimaskesta
verre ja lümfi. Seedekulgla põhifunktsioon on vee, elektrolüütide, toitainete
resorptsioon. Peensoole põhiliseks funktsiooniks on energiakandjate, vee,
elektrolüütide,
sapphapete , vitamiinide transport. Jämesool on oluline, lisaks
väljaheite reservuaariks olemisele, vedeliku- ja veeresorptsiooni lõplikus
regulatsioonis. Küümuse transport toimub kahes suunas: (a) oraalselt
aboraalsele, kusjuures liikumapanevaks jõuks on soolemotoorika; (b)
mukosaalselt serosaalsele (soolevalendikust supepiteliaalsetesse kapillaaridesse,
lümfisoontesse). Mingi aine transport läbi sooleepiteeli võib põhimõtteliselt
toimuda kas soolevalendikust serosaalsele või vastupidi. Kui ülekaalus on
ühesuunaline vool serosaalsele, on tegemist resorptsiooniga; kui
domineerib vool
mukosaalsele, on tegemist sekretsiooniga. Imendumine toimub peamiselt
soolelimaskesta hattude
tipus . Imendumiseks peavad molekulid, ühendid läbima
membraane. Resorptsiooniprotsessiks vajaliku suure pinna tagavad peensooles
kurrud, hatud, mikrohatud, mis muudavad pindala 600-kordseks.
Peensooleepiteel on keha kõige kõrgema jagunemis- ja käibekiirusega kudede
seas. Kogu enterotsüüt on nii luminaalselt kui basolateraalselt kaetud
kolmekihilise elementaarmembraaniga. Rasvlahustuvate substantside transport
läbi luminaalse membraani on lihtne difusiooniprotsess. Basolateraalselt
membraanilt on nende transport aga energiat nõudev, seotud spetsiaalsete
mehhanismidega. Polaarsete ja elektriliselt laetud substantside transport läbi
lipiidmembraanide on aga väga aeglane. Tuleb oletada pooride või
kanalite olemasolu membraanides. Enterotsüütide pinna negatiivne laeng on oluline laetud
osakeste passiivseks
transpordiks . Na+ on peamiselt
vastutav potentsiaali- ja
osmootse
gradiendi tekke eest, osaleb teiste substantside sidestatud transpordil.
Na+-sümporti kasutavad D-
heksoosid , L-aminohapped,
vesilahustuvad vitamiinid, niudesooles ka sapphapped.
Sõltuvalt intestinaalse epiteeli lokalisatsioonist, kulgeb kuni 90% ainete
transportdist paratsellulaarselt. Substantside võimet läbida epiteeli
paratsellulaarse tee kaudu osmootse, hüdrostaatilise, keemilise või elektrilise
19
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
gradiendi tõttu nimetatakse passiivseks permeaabluseks. Tiheühendused on
vabalt läbitavad kuni 0,8 nm läbimõõduga substantsidele. Proksimaalselt
distaalsele tiheühenduste pooride või kanalite läbimõõt väheneb, epiteeli
läbilaskvus ka vee suhtes on seega regiooniti erinev.
9.2. .
Süsivesikute imendumine Süsivesikud saavad imenduda ainult monosahhariididena, nende lõplik hüdrolüüs
toimub hariäärise oligosahharidaaside toimel. Oligosahharidaase on rohkem
tühisooles, vähem duodeenumis, iileumis. Hüdrolüütilise lõhustumise lõpp-produktid
on glükoos, galaktoos ja
fruktoos . Glükoos,
mannoos ja galaktoos resorbeeritakse läbi
valendikupoolse hariäärise membraani aktiivselt sümpordis Na+-ga. Transporteriks on
seejuures
SGLT1 (
sodium glucose transport protein 1). SGLT1 transpordib korraga 2
Na+ iooni ja 1 glükoosi või galaktoosi. Fruktoos
imendub läbi sama membraani
passiivselt kergendatud difusioonil transporteri
GLUT 5 abil kontsentatsiooni
gradiendi tõttu.
GLUT5 paikneb ainult sole hariäärisel ning teised
suhkrud teda ei
blokeeri.
Kui glükoosi ja galaktoosikontsentratsioonid soolevalendikus on väga suured (pärast
süsivesikuterikka toidu seedimist), toimub ka nende transport passiivselt.
Teise, so. basolateraalse membraani läbivad kõik 3 monosahhariidi kergenadtud
transpordil
GLUT2 transporteri abil seetõttu, et nende kontsentratsioon enterotsüüdis
on kõrgem kui basolateraalse membraani taga plasmas. Sama glükoosi transporter
töötab veel maksa, neerude ja pankrease saarekeste rakkudes.
Heksoosid imenduvad suhteliselt kiiresti, resorptsioon lõpeb peaaegu peensoole
algusosas. See väldib hüpertoonilisuse teket soolevalendikus, kuna tärklise
laguproduktid (vastandina tärklisele endale) on osmootselt aktiivsed. Imendunud
monoosid satuvad värativeeni kaudu maksa.
9.3. Valkude imendumine Täiskasvanu tarvitab päevas ligikaudu 70...90g valku. Intaktsed valgumolekulid imenduvad
vähesel määral pinotsütoosi teel, seda eelkõige vastsündinul. Sellel imendumisviisil toiteline
tähtsus puudub, küll aga omab ta immunoloogilist rolli, võides põhjustada nii sensibilisee-
rumist kui allergiat.
Valkude seedimine algab
maos, kus
pepsiin lõhustab 10...15% toiduvalgust: pankrease
peptidaaside produktsioon algab 10...20 min peale sööki ja kestab
niikaua , kui sooles on
valku. Pankrease ensüümid jõuavad
duodeenumisse inaktiivsena, kus trüpsinogeen
aktiveeritakse enteropeptidaasi toimel trüpsiiniks, mis omakorda aktiveerib teisi ensüüme.
Endopeptidaasid (trüpsiin, kümotrüpsiin, elastaas) lõhustavad valkudest oligopeptiide
molekuli tsentraalses osas, eksopeptidaasid (karboksüpeptidaas A ja B, aminopeptidaasid)
eraldavad
aminohappeid ahela lõpust. Peptidaasid asuvad ka harjasäärise membraanis ja
raku sisemuses, kus 90% di- ja tripeptiide lõhustataksegi, kui nad on rakku viidud
spetsiaalse transporteriga. 4...8- aminohappelised oligopeptiidid lõhustatakse harjasäärisel
hüdrolaaside poolt.
Duodenumis resorbeeritakse 50...60% toiduvalgust, kuni iileumini on
80...90% toiduvalgust resorbeeritud. Intaktsed valgumolekulid resorbeeritakse vähesel
määral pinotsütoosi teel, kuid see ei oma nutritiivset tähtsust. Peptiidid resorbeeritakse di-
ja tripeptiididena kandja abil aktiivselt või passiivselt. Aminohapete transporterid on Na+
sõltuvad ja Na+sõltumatud ning jagunevad omakorda:
a) neutraalsetele aminohapetele,
b) kaksik- aluselistele aminohapetele,
c) dikarboksüülsetele aminohapetele,
d) iminohapetele,
e) glütsiinile
Kolm esimest transpordivad aminohappeid soolerakku sarnaselt glükoosi transpordile Na+-
sümpordi mehhanismina (
sekundaarne aktiivne transport). Struktuurselt sarnased
20
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
aminohapped konkureerivad omavahel sama kandja pärast. Basolateraalse membraani
läbivad aminohapped kergendatud difusiooni teel.
Jämesoolde jõuab vaid kuni 10% valku, mis mikroobide poolt lõhustatakse. Vähene hulk
irdunud epiteelrakkudest pärit valku väljutatakse roojaga.
9.4. Lipiidide imendumine Päevas tarvitatakse umbes 60...
100g rasva, millest 90% on triglütseriidid ja sellest
enamus on pika ahelaga rasvhapped (16 või 18 C- aatomit). 10% moodustavad
fosfolipiidid (letsitiin), kolesteroolestrid ja
rasvlahustuvad vitamiinid. Rasv
vallandab
peensoole limaskestast CCK (koletsüstokiniin), mis põhjustab
pankrease nõre sekretsiooni ja sapipõie kontraktsiooni. Pankreas sekreteerib lipaasi
(ka kolipaas tema aktivatsiooniks). Lipaas lõhustab triglütseriide. Hüdrolüüsi
tulemusel tekivad 2-monoglütseriidid. Pealse selle sekreteerib kõhunääre ka
fosfolipaas A (fosfolipiidide lõhustamine) ja kolesterooli esteraasi (lõhustab
kolesteriide).
Üle 95% triglütseriididest (kuid vaid 20...50% kolesteroolist)
imendub duodenumis ja jejunumi algusosas. CCK toimel vabaneb ka sapp,
mis emulgeerib rasvu – muudab neid rohkem seedeensüümidele
kättesaadavamateks. Emulgeerimisprotsessis osalevad sapphaped, nende solad
ning ka 2-monoglütseriidid. Resorptsioonil peab segamitsell läbima järgmised
barjäärid:
a) liikumatu veekiht raku pealispinnal (suur takistus pikaahelalistele rasvhapetele ja
monoglütseriididele)
b) limakiht harjasäärise membraanil (transporditakistus)
c) enterotsüütide lipiidmembraan (rasvhapped ja monoglütseriidid lähevad enterotsüüti,
mitsellide koostisosised, mis ei tungi soolerakku, vabanevad ja on valmis uusi rasv-
koostisosiseid vastu võtma).
Pärast rakumembraani läbimist toimetatakse monoglütseriidid ja rasvhapped
madalmolekulaarse valgu poolt endoplasmaatilisse retiikulumi, kus toimub triglütseriidide ja
lipiidide resüntees. Selleks tuleb rasvhapped aktiveerida ja esterifitseerida (esterdatakse ka
fosfolipiidid,
kolesterool re- esterifitseeritakse ning iileumis toimub ka kolesterooli
resüntees). Uuesti sünteesitud triglütseriidid ja lipiidid ümbritsetakse spetsiaalse kestaga,
mis sisaldab peale kolesterooli ja fosfolipiidide ja glükoproteiine: moodustuvad külomikronid,
mis jõuavad sekretoorsesse põiekesse, mis lateraalse rakumembraaniga kokku sulab ja
külomikron vabaneb intertsellulaarruumi, kust külomikron liigub soolehatu
tsentraalse lümfisoone kaudu lümfisüsteemi ja lõpuks
ductus thoracicus’e kaudu ülemisse õõnesveeni.
Kuni 30% keskmise ja lühikese ahelaga rasvhappeid jõuab lõhustamatuna soolerakku, kus
toimub esteraaside toimel nende lõhustamine rasvhapeteks, mis koos soolt otse läbivate
difusiooni teel vastu võetud keskmise ja lühikese ahelaga rasvhapetega värativeeni jõuavad.
9.5. Vee ja mineraalainete imendumine Ööpäevas läbib peensoolt 9L vedelikku, millest 2L pärineb toidust ja 7L näärmete
endogeensetest ja soolesekreetidest. Üle 80% sellest resorbeerub peensooles, ülejäänud
jämesooles ja roojaga väljub vaid ~100 ml vett.
Vee liikumine läbi limaskesta toimub vaid
koos lahustuvate ainetega (elektrolüüdid ja mitteelektrolüüdid).
Peensoole algusosa läbilaskvus veele on suur, mistõttu muutub duodenumis küümuse
osmolaarsus verega isotooniliseks mõne minuti jooksul. Vesi liigub läbi peensoole seina
osmoosi teel, nii et kui osmootselt aktiivsed ioonid (Na+, Cl-) absorbeeritakse, järgneb neile
vesi. Nii samuti võib vesi ka sekreteeruda läbi rakkude kui ka läbi rakkudevahelise ruumi.
Jämesooles resorbeerub vesi osmoosi teel.
Na+ resorptsioon on väga ulatuslik.
Peensooles on aktiivsed ja passiivsed Na+
resorptsiooni mehhanismid:
a) Elektrogeenne Na+ transport: Na+ toimetatakse läbi basolateraalse membraani
intertsellulaarruumi Na+- pumba abil, mis saab energiat Na+- K+ATPaasi tööst.
21
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
Gradiendid põhjustavad vee liikumise valendikust rakku. Kuna tiheühendused on
suhteliselt läbilaskvad, voolab Na+ pärast resorptsiooni osaliselt
valendikku tagasi.
b) Na+- sümpordi abil liiguvad koos Na+-ga ühise kandja abil rakku ka D-heksoosid
(glükoos), L- aminohapped, veeslahustuvad vitamiinid ja sapphapped.
c) Neutraalse NaCl transpordi korral liiguvad Na+ ja Cl- rakku sümpordi teel; protsess
kulgeb elektriliselt neutraalselt.
d) Teine mudel NaCl neutraalse transpordi jaoks oletab kahekordset vahetust: Na+ H+
vastu ja Cl- HCO -
3 vastu, kusjuures H+ ja HCO3 moodustuvad karboanhüdraasi toimel
H2O-st ja CO2-st ja
vahetatakse välja Na+ ja Cl- vastu. Käivitavaks jõuks on Na+
aktiivne transport läbi basolateraalse membraani.
e) Passiivne konvektiivne Na+ transport on oluline Na+ resorptsioonil peensoole
lagusosas. Epiteeli suhtelise läbilaskvuse juures toimub 85% Na+ resorptsioonist
transportimisega lahusti abil (
solvent drag).
Na+ resorptsioon jämesoole valendikust toimub difusiooni teel, kuna basolateraalselt
toimub Na+ aktiivne transport verre.
K+ transport peensooles toimub peamiselt passiivselt vastavalt
kontsentratsioonidiferentsile.
Cl- resorptsioon toimub osaliselt koos Na+ resorptsiooniga, mida soodustab
potentsiaalidiferents (seroosa on valendiku suhtes positiivse laenguga).
HCO -3 sekreteeritakse valendikku peensoole algusosas Brunneri näärmete poolt ja
niudesooles punkt d) kirjeldatud
vahetuse teel.
Jejunumis toimub aga HCO -
3 resorptsioon,
kusjuures HCO -
3 võib karboanhüdraasi toimel osaliselt muutuda CO2 –ks, mis difundeerub
rakku ning HCO -
3 läheb üle plasmasse.
Jämesooles resorbeeritakse ulatuslikult Cl- ja HCO -
3 , millede resorptsioon on seal
sidestatud.
Ca2+ soolad muutuvad maohappe toimel veeslahustuvaks ja resorbeeritakse
40% ulatuses
peensoole algusosas. Kaltsiumi imendumist reguleerib vitamiin D
hormoonvorm. Aktiivsele transpordile madalate kontsentratsioonide korral lisandub ka
passiivne transport suurte kontsentratsioonide korral. Rakku sisenemiseks on Ca2+ tarvis
siduv
proteiin ja väljumine basolateraalse membraani kaudu toimub Ca2+-ATPaasi
aktiveeritava pumba abil. Toiduga saadavast 10...20mg
rauast resorbeerub umbes 10%.,
kusjuures heemne raud resorbeerub tunduvalt paremini.
Punaliha on parim rauaalllikas!
Raud imendub peensoole algusosas kahevalentsena, kui ka kolmevalentsena (aeglasemalt).
Raua sisenemine ja väljumine enterotsüütidest on aktiivne protsess. Seerumis võetakse
raud vastu ja transporditakse transferriini poolt. Ülemäärane raud seotakse enterotsüüdis
ferritiiniga ja rakkude irdumisel see eritatakse.
9.6. Vitamiinide imendumine (ei ole vaja nii täpselt teada) Vesilahustuvad vitamiinid toimivad peamiselt ensümaatilistes reaktsioonides
koensüümidena. Kõik peale ühe – vitamiin B12 – imenduvad difusioonil. Vitamiin C on
samuti
erandlik , imendub aktiivsel transpordil Na+ gradiendi arvel. Rasvlahustuvad
vitamiinid imenduvad segamitsellides ja ka sapphapetega konjugeerunult. Enterotsüütides
inkorporeeritakse nad külomikronitesse.
Tiamiin (B1) imendub jejunumist kandja vahendatud aktiivse
transportmehhanismiga, edasi läheb tiamiin maksa, kus üks osa temast
fosforüülub, teine osa aga transporditakse kudedesse, kus ta samuti fosforüülub.
Riboflaviin (B2) on toidus valguga seotud. Pankrease ensüümid lõhustavad
riboflaviin– valk kompleksi ja B2 imendub peensoolest aktiivse
transportmehhanismiga. Toidus sisalduvast
pantoteenhappest hävitab töötlemine
peaaegu poole, kuid seedekulglasse jõudnud osa vabaneb koensüüm A koostisest
hüdrolüüsireakstiooni tagajärjel ja imendub peensoolest.
Niatsiini (B3) on toidus
22
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
vabana vähe, enamus on NAD ja NADH koosseisus, need aga hüdrolüüsuvad
seedekulglas ja vabanev
niatsiin imendub peensoolest põhiliselt difusiooni teel,
kuid ka aktiivse transpordiga.
Vitamiin B6 (püridoksiin) imendub peensoolest
difusiooni teel.
Askorbiinhape (vitamiin C) hakkab
imenduma juba suus ja
maos, kuid põhiline imendumine toimub iileumis passiivse difusiooniga, kusjuures
imendumine sõltub askorbiinhappe hulgast (kõrgema kontsentratsiooni korral on
imendumisprotsent väiksem).
Kobalamiinide (B12) imendumine vajab vereloome
sisemist faktorit e. Castle’i faktorit. Vereloome väline faktor vabaneb lihavalkudest
maohappe mõjul ning see peab liituma sisemise vereloomefaktoriga ja kitsas
peensoole osas on vastav retseptor, mis seob kompleksi. See on vajalik vitamiin
B12 imendumiseks enterotsüüti, kus kompleks laguneb ja vitamiin B12 läheb
vereringesse. Vaid siis kui toidus on palju B12, võib väike osa imenduda ilma
seesmise faktorita peensoole proksimaalsetest lingudest.
Foolhappe (B10) molekul koosneb pteridiini-, paraaminobensoehappe- ja glutamiinhappe (neid
1...7) jäägist. Selleks et vaid üks pteroüülglutamiinhape saaks peensoole
valendikust (ülemisest jejunumist) imenduda, peab polüglutamüülahel lühenema.
See toimub ensüüm pteroüülglutamaadi hüdrolaasi toimel enne imendumist ning
see ensüüm asub harjasäärisel. Vabanenud pteroüülglutamiinhape imendub
spetsiaalse aktiivse transpordimehhanismiga.
Rasvlahustuvad vitamiinid (A- retinoid, D- kaltsiferool, E-
tokoferoolid , K-
naftokinoonid, F-
linool - ja linoleenhape) imenduvad nagu pikaahelalised
rasvhapped mitsellidesse sisestatult. Imendumismehhanism on küllastatav ja
aktiivne.
Vitamiin A imendub mitsellidesse sisestatult. Imendunud vitamiin A
esterifitseerub peensoole limaskestas
palmit - ja stearhappega ning tekkinud
retinüülstearaat ja –
palmitaat transporditakse külomikronites maksa.
Vitamiin D imendumine peensooles sõltub sapphapetest (mitsellide moodustumisest).
Peensoole limaskestas moodustunud ergokaltsiferool (D2) ja kolekaltsiferool (D3)
transporditakse külomikronites maksa.
Vitamiin E (tokoferoolid) esinevad toidus
estritena, mida hüdrolüüsib kõhunäärme esteraas ja vabanenud tokoferoolid
imendatakse peensoole keskosas sapphapetest ning triglütseriidide seedimisest ja
imendumisest sõltuvana. Toidu tokoferoolidest imendub vaid 1/3.
Vitamiin K: nii
toiduga seedekulglasse sattunud füllokinoon (K1), kui ka soole mikrofloora
toodetud farnokinoon (K2) imenduvad peensooles. Imendumine sõltub sapphapete
hulgast ja pankrease lipaasi aktiivsusest. Vitamiin K liigub külomikronite koostises
maksa. Samas ei saa märkimata jätta jämesoole osa vitamiin K ja ka B- rühma
vitamiinide imendumisel.
10. Seedimine jämesooles. Toidu kiudained: nende lõhustamine ja tähtsus. Defekatsioon. Jämesooles toimub:
1.
Vee resorptsioon osmoosi teel. Tavaliselt resorbeeritakse 1...2L vett. Na+
resorptsioon valendikust toimub difusiooni teel, kuna basolateraalselt toimub
Na+ aktiivne transport verre. Cl- ja HCO -
3 resorbeeritakse siin ulatuslikult. Cl-
resorptsioon on sidestatud HCO -
3 sekretsiooniga. K+ setserneeritakse
jämesoolevalendikku elektrilise gradiendi mõjul.
2.
Vitamiinide, näit B- rühm, K, süntees ja
absorptsioon . See toimub
bakterite mõjul.
3.
Valkude lõhustamine bakterite ensüümide poolt. Lõhustumise lõpp-
produktid
indool ja skatool on mürgised, ning annavad roojale iseloomulikku
lõhna.
4.
Kiudainete lõhustamine bakterite ensüümide poolt. kiudainete
lõhustamisel tekib rohkesti
gaase : 400...700 ml. CO2, H2, N2, H2S,
metaan (CH4)
Peale gaaside tekib bakterite lõhustamise tulemusel ka piimhapet, äädikhapet ja
alkoholi. Piimhape kasuatatkse ära limaskesta ainevahetuses energia saamiseks.
23
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
Valkude lõhustumisel tekivad indool ja skatool, histamiin ja türamiinid lähevad
venoossesse verre ja
v. porta kaudu maksa, kus need kahjutuks tehakse.
Jämesooles on rohkesti
baktereid: roisu- ja käärimisbaktereid, mille poolt
põhjustatud roiskumis- ja käärimisprotsessid peavad olema tasakaalus.
Ravimid võivad mõjutada mikrofloorat ja muuta olukorda roisubakterite kasuks.
Käärimisel tekib happeline keskkond, mis takistab roiskumist. Valdav E. coli.
Jämesoole motoorika aeglaste lainete sagedus on muutlikum, kui peensooles.
Küümus liigub jämesoolde 2...3 päeva jooksul, seda tüüpilise europlase toidu
korral, loodusrahvastel on see periood lühem. Parasümpaatikus stimuleerib
jämesoole motoorikat, samas sümpaatikus ja noradrenaliin mõjub motoorikat
pidurdavalt. Gastriin ja koletsüstokiniin soodustavad ja sekretiin ja glükagoon
pidurdavad jämesoole motoorikat.
Jämesoole motoorika on rohkem
mittepropulsiivne peristaltika. Laine on
harvem ja ilmneb ebareeglipäraselt ning võib
liikuda mõlemas suunas.
Peristaltiline laine ei haara tavaliselt rohkem, kui 10...20 cm pikkust lõiku. See
segab jämesoole sisaldist ja annab jämesoole protsessidele aega. Vastupidine
peristaltika soodustab vee ja soolade tagasiimendumist, sest peensoolest tuleb
üsna vedel küümus. Roojaga väljub päevas umbes 100...150ml vett. 1...3x
päevas tekib
massi liikumine e. Holzknechti liikumine, millega haaratakse
pikem osa jämesoolest ja suunatakse jämesoole
sisaldis sigmasoolde.
Gastrokooliline refleks algab 10 minutit peale toidu sisenemist makku, see on
kolinergilise
iseloomuga . Toidu makku
sattumine aktiveerib mao motoorikat ja ka
sigmasool saab erutatud korduvate massiliikumistega ja gastrokoolilise refleksiga
ning see võib vallandada defekatsiooni.
Defektasioonil osaleb pärasoole sisemine ja tahtele alluv väline vöötlihaseline
sfinkter. Sisemine sfinkter on silelihaseline ja vegetatiivsele närvisüsteemile
allumatu. Osalevad ka kõhupressi lihased ja hingamislihased ning soole
silelihased. Keskus asub seljaaju sakraalosas ja on tahtele allumatu. Samas on
see allutatud
peaaju keskuse kontrollile. Kui seljaaju on vigastatud, kontroll kaob.
Sigmasoole täitumine põhjustab rektumi täitumise. Sisemine pärasoole sfinkter
lõõgastub, samas tõmbub väline tugevamini kokku. Tekib venitustunne. Selline
sfinkterite
reaktsioon on mööduv, refleks võib edasi lükkuda. Selleks, et
defekatsioon saaks toimuda, peab peaaju vastav keskus andma käsu lõõgastada
väline sfinkter, siis lõõgastub ka sisemine. Defekatsiooni ajal toimub alaneva
kooloni ja sigmasoole tugev propulsiivne kontraktsioon (lõõgastus ees,
kontraktsioon järel). Tahteline ja reflektoorne tegevus toimuvad samaaegselt.
Glottis sulgub ja diafragma ja kõhulihased kontraheeruvad (
prelum abdominale)
ja toimub rooja väljutamine.
Toidu kiudained
Inimene peab tarbima 20-30g kiudaineid ööpäevas. Nende tunnuseks on
seedumatus peensooles. Eristatakse polüoosseid ja mittepolüoosseid kiudained.
Polüoossed kiudained jagunevad vees lahustuvateks (näit. pektiinid, need
punduvad kergesti ja nende
lahused tarretuvad kergesti) ja vees lahustumatuteks
(
tselluloos , sisuliselt ka
hemitselluloos ). Mittepolüoossetest kiudainetest on
sagedaseim
ligniin . Kuigi kiudained peensooles ei seedu, on neil siiski olulised
ülesanded:
a) tselluloos ja hemitselluloos suurendavad toidukördi mahtu ja viskkoossust
(soolesisaldis on seetõttu pooltahke) ―> roojamassi
formeerumine ;
b) kiirendavad/ soodustavad toidukördi edasiliikumist peensooles (kiirendavad
motoorikat, parandavad seedimist);
c) soodustavad lima eritumist jämesooles;
d) reguleerivad biomolekulide imendumist peensooles;
e) tekitavad küllastustunnet.
24
Kaur Kersalu, Grigory Vasiliev
Koostatud: Mõmmi baasil 2008/2009
Kiudained suurendavad mehhaaniliselt mutsiini sekretsiooni- see on seedekulgla
limaskesta kaitsev. Kiudained
alluvad jämesooles sealsete mikroobide toimele.
MAO HAPPESEKRETSIOONI REGULATSIOON
25







































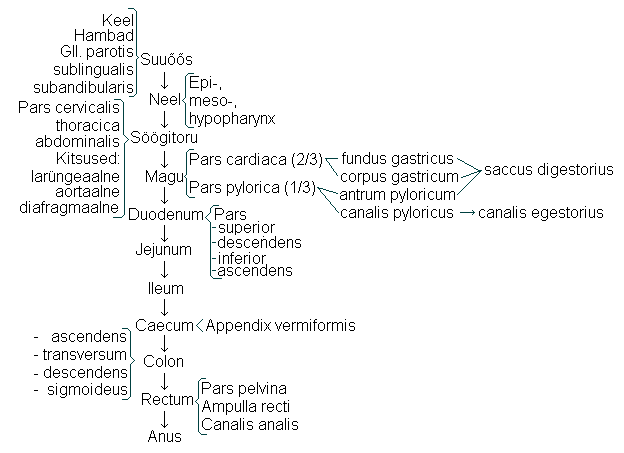
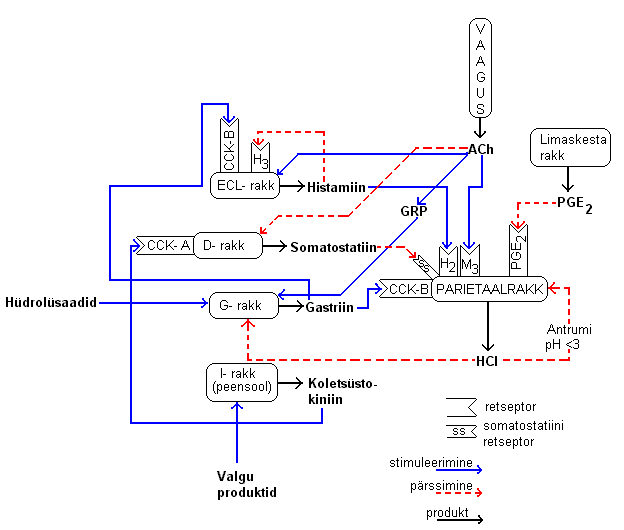
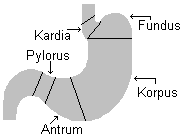




Kõik kommentaarid