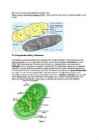
1. Suhkrute lühiiseloomustus. (CH2On)e süsivesikud on org.ühendid
: koostis süsinik, vesinik, hapnik.
Lihtsuhkrud –
monosahhariidid .
Liitsuhkrud – *
oligosahhariidid (2-10 kovalentselt seotud monosahhariidi jääki);
*polüsahhariidid
(sadu kuni tuhandeid monosahhariidi jääke).
Monosahhariidid jagunevad:
*C-aatomite arvu järgi (
trioos , tetroos);
*
funk .ühma
järgi (
aldoosid , ketoonid);
*tsüklilise
struktuuri alusel (püranoosid, furanoosid).
Polüsahhariidid:
*homopolüsahhariidid (ühe monosahhariidi jäägid);
*heteropolüsahhariidid
(mitme monosahhariidi jäägid);
*hargnenud
või lineaarse ahelaga.
Bioloogiline roll: *väga
mitmekesine ja looduses laialt levinud org.molekulide klass;
*päikese
energia salvestatakse fotosünteetiliste organismide poolt
süsivesikutesse;
*paljude
biomolekulide eelühendid;
*struktuuriline
roll;
(
*molekulaarsed ja
rakk -rakk äratundmismehhanismid.)
Mono:süsinike
arv (enamasti) 3-6;
5-e süsinikulised
monosahariididest on olulised
riboos
ja
desoksüriboos6-e süsinikulised
suhkrud (C6H12O6)
glükoos(viinamarja
suhkur) &
fruktoos (puuviljasuhkur)
Nii glükoos kui fruktoos on
organismis põhilised energiaallikad. Roh.taimedes valmib glükoos
fotosünteesi tulemusena. Glükoosi järkjärgulisel oksüdatsioonil
CO2
ja H2O´ks
vabaneb energia (17,6KJ/g)
Oligo sahhariidid on madalmolekulaarsed ühendid, mis on enamasti mood. 2-3
monosahariidi seostumisel. Ntks glükoos + fruktoos =
sahharoos (roo
ja peedisuhkru põhiosa)
Sahharoos
Laktoos(
piimasuhkur )
kuulub ka oligosahhariidide hulka
Polüsahhariidid
– taimed säilitavad oma glükoosi varusid
tärklisena.
Vajadusel muundavad taimed tärklise glükoosiks.
Loomorg säil glükoosivarusid
peamiselt maksas ja lihastes
glükogeeni(koosneb
sammuti glükoosi jääkidest) kujul.
Lülijalgsete välisskeletis
ja ka seente rakukestas esineb
kitiin Selle monomeerideks
on lämmastikku sisaldav suhkur.
2. Lipiidide lühiiseloomustus.Lihtlipiidideks ehk
rasvadeks nim propaamdiooli(glütserooli) ja rasvhapete estreid.
Rasvad erinevad nende koostisse kuuluvate rasvhappejääkide poolest. Mida
rohkem on rasvhappe jääkides
kaksiksidemeid , seda vedelam on rasv.
Lipiidid on org üh klass,
kuhu kuuluvad rasvad, õlid, vahad, fosfo- ja glükolipiidid,
rasvhapped ja nende derivaadid,
terpenoidid (ka steroidid) - vees
(enamasti)
mittelahustuvad ühendid.(lipiidid lah erinevates
apolaarsetes üh, ntks. benseen ja
eeter )
Lipiidid täidavad org
põhilise energiaallika osa. Osküdeerimisel vabaneb 2x rohkem
energeiat(38,9KJ/g) kui samakoguse sahhariidide või valkude lag.
Lipiidid(rasvakiht) võivad omada ka kaitse- funktsioone(ümbritseb
siseorganeid), kaitseb liigse
jahtumise ees(jääkarul naha all).
Taimedes on suurim lipiidide
sisaldus seemnetes(pähklid).
Lihtlipiidide
ühinemisel teise keem.ühenditega mood.
liitlipiidid .
Ntks kuuluvad rakumembraani koostisesse
fosfolipiidid.Vahades
on glütserooli asemel mõni teine
alkohol .
Steroidid
on ülejäänud lipiididega võrreldes teistsuguse ehitusega.
Tsüklilised ühendid, mis vees (praktiliselt) ei lahustu. Steroidide
hulka kuuluvad mitmesugused
hormoonid(
suguhormoonid ja neerupealse hormoonid) ning ka
vitamiin D(kaltsiferool).
Hormoonid
on bioaktiivsed ained, mis põh mood loomorganismide
sisekretsiooninäärmetes. Väga väikestes kogustes mõjuvad.
Reguleerivad ainevahetust ja kogu org talitlust.
Kolesterool moodustub
organismis, kui toit sisaldab palju rasva ja suhkrut. Kolesteroool on
samuti üks steroididest. Kolesterool kuulub kõigi membraanide
koostisesse(muudab elastseks??!?!). Liigne kolesterool võib koos
rasvhapete ja kaltsiumisooladega ladestuda arterite seintele ja põh
veresoonte lupjumist e ateroskleroosi.
Funktsionaalsuse järgi jaotatakse: *varulipiidid;
*struktuurilipiidid;
*signaalmolekulid (hormoonid);
*kofaktorid
ja
rasvlahustuvad vitamiinid (D,A,E,K);
*
pigmendid .
(vesilahustuvad vitamiinid on B,H,C)
3. Valkude lühiiseloomustus. Valgud ehk proteiinid
on biopolümeerid, mille monomeerideks on aminohappejäägid.
Organismis on 20 erinevat
aminohapet. Väheste valkude koostisest leiame aga kõik 20
aminohappe jääki.
Aminohapped on amforteersed
ühendid, sest iga aminohappe koostisesse kuulub aluseliste
omadustega aminorühm (-NH2)
ja happeliste omadustega karboksüülrühm (-COOH)
Aminohappejäägid on valgu
molekulis omavahel ühendatud peptiidsidemetega.
Peptiidside moodustub ühe
aminohappe karboksüülrühma ja teise aminohappe aminorühma vahel:
Peptiidside moodustub valgu
sünteesi käigus ribosoomides. Sünteesi tulemusena tekib
polüpeptiid.Vähem kui kümnest
aminohappejäägist
koosnevat ühendit nimetatakse oligipeptiidiks.
Erinevatel peptiididel on erinevad
struktuurid .
Valgu aminohappelist
järjestust nimetatakse eselle esimest järku struktuuriks, ehk
primaarstruktuuriks.
Primaarstruktuur on kõigil valkudel ja see määrab ära kõik valgu
omadused, samas ei väljenda see otseselt valgumolekuli kuju.
Valgu teist järku struktuur
ehk
sekundaarstruktuur
tekib polüpeptiidi keerdumisel kruvikujuliseks α-heeliksiks või
kõrvuti asetsevate
ahelate voltumisel β-struktuuriks.
Valgu molekuli edasisel
kokkukeerdumisel tekib kolmandat järku struktuur e
tertsiaalstruktuur
(kera sarnane kuju, nimetatakse
gloobuliks)
Mitte kõik valgud pole
globulaarsed : mõned võivad jääda
väljavenitatult niitjateks ehk
fibrillaarseteks.Kahe või enama polüpeptiidi
ühinemisel moodustub liitvalk, mida nimetatakse
neljandat järku
struktuuriks ehk
kvaternaarstruktuuriks.
(hemoglobiin).
Kui valgulahust kuumutada,
siis soojusenergia toimel nõrgad keem sid
katkevad ning valk kaotab
oma kõrgemat järku struktuurid, seda nimetatakse
denaturatsiooniks.
Lisaks temperatuurile võivad valke denatureerida ka mehhaanilised
tegurid,
happed , raskmetalliühendid,
ioniseeriv ja UV kiirgus.
Aminohappejääkidevahelised
peptiidsidemed
denaturatsiooni käigus ei katke.
Denatureeriva mõju lakkamisel
võivad valgud taastada sekundaar – ja tertsiaalstruktuuri, seda
pöördprotsesse nim
renaturasiooniks.
Valgud võivad ühineda ka
teiste org ainetega, mood
liitvalgud :
Valgukompleks nukleiinhapetega
nim – nukleoproteiinideks (eukarüootide
kromosoomid )
Lipiidedega –
lipoproteiionid(rakumembraani koostis)
Oligosahhariididega –
glükoproteiinid(
membraan )
Funktsioon:1) Valkude ensümaatiline
funktsioon:
Ensüümis reageerivad
biokeemiliste reaktsioonide kiirust, ensüümid on VÄGA
spetsiifilised . Mõnede ensüümide aktiveerumiseks on vajalik
vitamiinide olemasolu rakus.
2) Ehituslik funktsioon:
Rakuorganellide koostis,
karvad , suled, sõrad jne.
3)
Transportfunktsioon :
Hemoglobiin transpordib
hapniku ka rakumembraani koostises asuvad transportvalgud, mis
juhivad molekule selektiivselt nii rakku sisse kui välja.
4)Info:
Rakumembraanis esineb valgu
molekule , mis edastavad rakuvälist infot raku sisemusse. Tänu
retseptor valkudele liigub amööb toiduosakeste suunas.
5) Regulatoorne:
Valgulised hormoonid,
(
insuliin , tekib kõhunäärmes).
6)Kaitse:
Antikehad .
7)Liikumine:
Kontraktsiooni
valgud(
lihasvalgud ), algloomade
vibur (valgulised torukesed –
mikrotuublid)
8)Energeetriline:
17,6kJ/g
Enamasti algab valkude
lagundamine alles peale sahhariidide ja lipiidide
tagavara lõppemist,
ehk nälgimisel.
4. Nukleiinhapete lühiiseloomustus.Nukleiinhapped on
biopolümeerid, mille monomeerideks on
nukleotiidid .
DNA –
desoksüribonukleiinhape
RNA –
ribonukleiinhape DNA on biopolümeer, mille
monomeerideks on desoksüribonukleotiidid mis on moodustunud kolme
ühendi – lämmastikaluse, desoksüriboosi ja fosfaatrühma
liitumisel.
DNA
monomeerid erinevad
lämmastikaluste(4) poolest:
adeniin (A), guaniin(G), tümiin(T) ja
tsütosiin(C)
Seetõttu nimetatakse
nukleotiide lämmastikaluste järgi.
Komplementaarsus printsiip –
A ja T, G ja C.
DNA´l on ka
sekundaarstruktuur, biheeliks.
RNA on biopolümeer,
monomeerideks ribonukleotiidid, tümiini asemel on
uratsiil (U). RNA
omadused sõltuvad monomeeride järjestusest molekulis.
Ribonukleotiidide järjestust molekulis nimetatakse RNA
prim.struktuuriks.
Primaarstruktuur on ka RNA-l nukleotiidide ahel,
kusjuures ühinemine toimub fosfodiestersidemetega. Nukleotiidse järjestuse määrab RNA sünteesil eeskujuks olev DNA ahel.
Kuigi RNA on peamiselt üheahelaline, võivad selle molekulis esineda ka pikad kaheahelalised lõigud, kus
ahelad on ühinenud komplementaarsete nukleotiidide vaheliste vesiniksidemete abil. Komplementaarsed paarid on siin A — U ja C — G. Sellised lõigud esinevad näiteks transport-RNA-s.
Nagu iga teine nukleiinhapegi koosneb mRNA nukleotiididest, mida tihti kutsutakse ka
alusteks . RNAs sisalduvad nukleotiidid on: adeniin (
A), guaniin (
G), tsütosiin (
C) ja uratsiil (
U). DNA ahelas vastab uratsiilile tümiin (
T). Nukleotiidide kolmikuid e. triplette
mRNAs nimetatakse koodoniteks, mis moodustavad geneetilise koodi põhialuse. Igale koodonile mRNAs vastab mingi konkreetne
aminohape proteiiniahelas või juhis valgusünteesi alustamiseks või lõpetamiseks.
Ühtekokku on olemas 43 = 64 erinevat
koodonit . Kuna kodeeritavaid
aminohappeid on ainult 20, siis on paljud
koodonid "sünonüümsed", st. ühele
aminohappele võib vastata mitu erinevat koodonit. Milliseid koodoneid genoomis eelistatakse, sõltub aga konkreetsest organismist endast. Otsene seos on aga aminohappele vastavate
koodonite hulga ja selle aminohappe esinemissageduse vahel valkudes.
Mõnele koodonile (UGA, UUA ja UAG) ei vasta ühtegi aminohapet, mistõttu neid mõnikord
nimetetakse ka nonsensskoodoniteks. See nimetus ei ole aga päris õige, kuna nimetatud koodonid
toimivad märgina valgusünteesi lõpetamiseks, umbes nagu punkt lause lõpus. Stoppkoodoneid e terminaatorkoodoneid
tunnevad ära teatud ensüümid, nn. terminatsioonifaktorid, mis osalevad valgusünteesi lõpetamisel.
5. Millised on peamised erinevused DNA ja RNA vahel?DNA sisaldab endas pärilikku informatsiooni ning RNA osaleb päriliku info realiseerimises — valgu biosünteesis.
Kuna DNA primaarstruktuuris võivad nukleotiidid paikneda suvalises järjestuses, võimaldab DNA nende järjestuste kaudu talletada bioloogilist informatsiooni (seda võimaldab ka RNA, kuid DNA on oma suurema keemilise stabiilsuse tõttu pikaajalisemaks info säilitamiseks märksa
sobivam ühend)
6. Kolm põhilist RNA-de klassi rakkudes, nende funktsioonid.mRNA(informatsioon) – RNA, mille molekulilt toimub
translatsioon : informatsiooni-RNA nukleotiidijärjestuse põhjal sünteesitakse polüpeptiid.
tRNA(transport) – tegeleb rakus aminohapete transpordiga ribosoomi, kus geneetilise koodi alusel lisatakse aminohape sünteesitavasse valguahelasse.
rRNA(ribosoomis) – seostub valkudega kompleksideks – ribosoomideks (osaleb valgu sünteesis).
7. Prokarüootsete ja eukarüootsete rakkude peamised erinevused.Prokarüoodid on väiksemad, neil puuduvad
histoonvalgud , DNA on nukleoidis (eukarüoodis DNA kromosoomidesse pakituna tuumas), prokarüoot pooldub või pungub, eukarüoodil
mitoos , membraaniga ümbritsetud organelle prokarüoodil pole (eukarüoodil
mitokondrid ,
kloroplastid jne.), prokarüoodil puudub nii tsütoskelett kui ka rakusisene liikumine (eukarüoodil
kompleks mikrotuubulitest ja filamentidest ning liikumine: endotsütoos, mitoos, vesiikulite transport).
8. Membraanide struktuuri lühiiseloomustus.Membraanide koostisse kuuluvad peamiselt fosfolipiidid (moodustavad kaksikkihi – pikad hüdrofoobsed sabad sissepoole pööratud) ja valgud (on seotud lipiidi kaksikkihi pinnaga –
perifeerne valk, ulatuvad kaksikkihi sisse või läbivad seda – integraalne valk). Membraan võib
sisaldada ka steroole ja süsivesikuid.
Rakusisesed
membraanid on oma ehituselt sarnased raku välismembraaniga.
Endoplasmaatilise retiikulumi lühiiseloomustus. Eukarüootse raku tsütoplasmat läbib membraanse ehitusega kanalikeste ja tsistrenikeste süsteem, mis moodustab tsütoplasmavõrgustiku ehk endoplasmaatilise retiikulumi.(ER)
karedapinnalisel ER`il paiknevad ribosoomid .
siledapinnalisel ER´i membraanidel paiknevad ensüümid, mis võtavad osa lipiidide ning sahhariidide sünteesist. Sünteesiproduktid liiguvad kanalikeste ja tsisternikeste süsteemi kaudu raku erinevatesse osadesse.
Funktsioon: lipiidide süntees, biosünteesitud ühendite suunamine nende lõplikku paikka.
10. Ribosoomide ehitus ja funktsioon.
Iga ribosoom koosneb kahest osast, mõlemad osad omakorda rRNA´st ja valgu molekulidest.
Ribosoomid moodustuvad rakutuumas olevates tuumakestes.
Ribosoomides toimub valkude süntees.(mujal elusorganismis ei sünteesita)
Ribosoome võib leida ka suuremates rakuorganellides(motokonder, kloroplast), kus need sünteesivad vastavale organellile vajalikke valke.
11. Lüsosoomide funktsioon.
Lüsosüümid on ühekordse membraaniga ümbritsetud põiekesed, kus lagundatakse erinevaid makromolekule ja rakustruktuure.
Primaarsed lüsosüümid sisaldavad üksnes ensüümivalke, sekundaarsed lüsosüümid sisaldavad lagundavaid aineid ning neid lõhustavaid ensüüme. Hüdrolüütiline lõhustamine.
12. Golgi kompleksi funktsioon.
Golgi kompleksis jõuab lõpule valkude ümbertöötlemine ninge nende pakkimine sekreedi põiekestesse ja lüsosoomidesse.
Osaleb ka rakumembraani ning taimerakkudes ka rakukesta moodustamises.
13. Mitokondrite funktsioon.
Ümbritsetud kahe membraaniga, sisemembraan varustatud arvukate kudrude ja sopistustega( harjakesed ). Nende vahel leiab mitokondriaalset DNA´d ja RNA´d ja ribosoome.
DNA sisaldab geneetilist infot mitokondrile omaste RNA ja valkude sünteesiks.
Ribosoomid sünteesivad organellile vajalikke valke.
Põhiül on raku varustamine energiaga (ATP). Samuti osalevad mitokondrid signaaliülekandes ja raku surmas.
14. Kloroplastide ehitus ja funktsioon.
Kloroplastid sarnanevad mitmeti mitokondritega. Neil on hästi läbilaskev välismembraan ja kitsas intermembraanne ruum. Sisemembraan ümbritseb ruumi, mida nimetatakse stroomaks. Seal asub DNA, RNA, ribosoomid jne. Erinevalt mitokondritest on kloroplastidel üks lisakompartment - tülakoidid. Ka on kloroplastid suuremad kui mitokondrid. Eri tülakoidide valendikud on ühendatud omavahel. Seal paiknevad fotosünteetiline valguse absorbeerimise süsteem, elektrontranspotahel ja ATP süntetaasi kompleks.Tülakoidides toimuvaid reaktsioone nimetatakse ka valgusreaktsioonideks, sest valgus on seal otseseks energiaallikaks (elektroni võtmine vee molekulilt ja hapniku teke). Kloroplasti stroomas toimub süsiniku fikseerimise reaktsioon e. pimeduse reaktsioon (sest seal pole otsest valgusenergiat vaja, seal kasutatakse ATP energiat, mis on saadud valgusreaktsioonist. See reaktsioon jätkub tsütoplasmas. Seega valgus- ja pimedusreaktsioon on ruumiliselt eraldatud.
15. Tsütoskeleti funktsioonid.
Tsütoskelett (tsütoplasmas) hoiab raku kuju, kaitseb rakku ja võimaldab raku liikumist. Mängib suurt rolli rakusiseses transpordis , raku jagunemises ja organellide ümberpaiknemises.
Tsütoskelett koosneb valgulistest fiibritest, mis ühendavad omavahel rakumembraani, tuumamembraane, tsütoplasmavõrgstiku ja enamikku raguorganelle.
16. Rakutuuma osised.
...?!
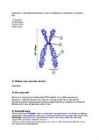
17. Kromosoomide struktuur.
Eukarüoodi DNA on jaotunud mitmeks individuaalseks elemendiks e. kromosoomiks - seda tõenäoliselt selleks, et genoom oleks rakus lihtsamini ja efektiivsemalt manipuleeritav. Kromosoomis on DNA püsivalt seotud valkudega, mis pakivad DNA kaksikahela ning loovad rakupõlvkondades säilivaid struktuurseid seisundeid, kus geenid on kas püsivalt inaktiveeritud, püsivalt ekspresseeruvad või saavad alluda jooksvale regulatsioonile. Kromosoomide spetsialiseerunud piirkonnad nagu näiteks tsentromeer , telomeerid , satelliidid või kromomeerid on naaberaladest visuaalselt eristatavad ja täidavad erinevaid funktsioone. Taolise lineaarse diferentseerituse taga on unikaalse ja kordus-DNA alade paiknemine kromosoomis ning eu- ja heterokromatiini erinev kokkupakkimine, mida saab in situ hübridisatsiooni ja diferentsiaalvärvimise meetodite abil kromosoomitasandil nähtavale tuua.
Prokarüootide (Archaebacteria ja Eubacteria incl . Cyanobacteria) kromosoom on rõngas-DNA molekul . Kogu prokarüdoodi genoom on pakitud ühte kromosoomi, mille koostisesse kuuluvad peale DNA kaksikahela veel valgud ja RNA-d. DNA, mille pikkuseks on keskmiselt 1 mm, on kokku pakitud mitmekümneks linguks, mida hoitakse koos RNA abil. Transkribeeritavad lingualad despiraliseeruvad. Valkude abil kinnituvad DNA lingud oma basaalse alaga bakteriraku sisemembraanile, moodustades nukleoidi piirkonna.
Kõigil eukarüootidel on nukleosoomse struktuuriga kromosoomid, mis paiknevad rakutuumas ning on tsütoplasmast eraldatud kahekihilise tuumamembraaniga. Lisaks tuumagenoomile on eukarüoodi rakus tuumaväline mitokondrigenoom, millele taimede ja mõnede vetikate puhul lisandub kloroplastigenoom. Erinevalt prokarüoodi genoomist on eukarüoodi genoom jagatud kromosoomide vahel. DNA molekulide arv kromosoomis võib olla üks või kaks, vastavalt sellele, kas on tegemist ühe- või kahekromatiidilise kromosoomiga. Ühekromatiidiline e. G1 kromosoom on rakutsükli G1 või G0 faasis olev kromosoom, milles üks DNA kaksikahel ulatub pidevana ühest kromosoomi otsast teise. Kahekromatiidiline e. G2 kromosoom on rakutsükli S faasi läbinud kromosoom, milles on 2 lineaarset DNA molekuli. Viimaste hulka kuuluvad ka näiteks mitoosi pro- ja metafaasi kromosoomid. Iga kromosoom koosneb seega ühest või kahest lineaarsest DNA molekulist, mis on seostunud struktuursete ja regulatoorsete valkudega. Interfaasi tuumas on valgusmikroskoobis näha vaid kromatiini kogumikud, elektronmikroskoobi lahutuse tasemel võib saada aga kinnituse , et kromosoomid on omaette struktuurina olemas raku tuumas kogu rakutsükli vältel. Kromosoomid kinnituvad tuuma sisemembraanile tuuma lamiinide vahendusel. Mitoosi ja meioosi kromosoomid on tugevasti kokkupakitud struktuurid ja valgusmikroskoobis analüüsitavad. Niisiis , võrreldes prokarüoodiga on eukarüoodi kromosoomide arv suurem, kromosoomid ise on palju suuremad ning nende ehitus keerulisem. Kromosoomi eri piirkonnad sisaldavad erinevaid DNA klasse ja täidavad kindlaid funktsioone. Eriti olulised kromosoomi kui terviku säilimisel on tsentromeeri ja telomeeri alad.
1.Kromatiid
2.Tsentromeer
3.Lühike „käsi“
4.Pikk „käsi“
18. Milliseid rakke ümbritseb rakukest ?
Taimerakke.
19. Mis on plasmiid ?
Plasmiid on rõngakujuline kaheahelaline DNA molekul, mis sisaldab autonoomset paljunemist võimaldavaid geneetilisi elemente (eelkõige replikatsiooni alguspunkti), selektiivset markerit (ampitsilliinile resistentsust tagavat geeni) ja unikaalseid restriktaaside lõikamiskohti (esinevad plasmiidis ainult 1 kord).
20. Rakutsükli etapid.
Rakutsükkel koosneb M-faasist (mitoos e. karüokinees(tuuma jagunemine) + tsütokinees(tsütoplasma jagunemine) e. raku pooldumine, mille puhul kromosoomid jaotuvad tütarrakkude vahel võrdselt) ning interfaasist e. päristuumse raku kahe jagunemise (mitoosi või meioosi) vahele jääv eluperiood (ajaliselt 90% vôi rohkem rakutsükli kestusest).
Interfaasis toimub kõikide rakukomponentide sünteesimine, et tekkivatel tütarrakkudel oleks olemas kõik vajalik uue tsükli alustamiseks.
M-faas: tuuma jagunemine ja tsütoplasmaatiline jagunemine.
Mitoosi all mõeldakse raku tuuma jagunemist koos tavaliselt sellega kaasneva tsütoplasma jagunemise e. tsütokineesiga. Mitoosi vaadeldi juba rohkem kui 100 aastat tagasi ning jõuti arusaamisele, et see kujutab endast mehanismi raku geneetilise materjali ning tsütoplasma jaotamiseks tütarrakkude vahel. Mitoos e. M-faas jaotatakse tavaliselt kuueks alafaasiks. 5 esimest moodustavad tuumajagunemise e. mitoosi kitsas mõttes. Kuues faas, mis ajaliselt kattub mitoosi lõpuga, kujutab tsütoplasma jagunemist e. tsütokineesi. Need 6 faasi moodustavad dünaamilise järjestuse, mille keerukust ja ilu on ainult kirjelduse abil edasiantuna raske tunnetada.
Üleminek G2-st M-faasi pole mikroskoobis võimalik täpselt fikseerida . Kromatiin , mis interfaasis on difuusne, kondenseerub aeglaselt kompaktseteks kromosoomideks. Kaob tuumake, sest kondenseerunud kromatiinilt ei toimu enam RNA transkriptsiooni. Profaasis lagunevad ka tsütoplasmaatilised mikrotuubulid ning hakkab moodustuma mitoosiaparaadi peamine komponent - kääviniidistik. See on bipolaarne struktuur, mis koosneb mikrotuubulitest ja nendega seotud valkudest. Kääv moodustub alguses tuumaväliselt, lahknevate tsentrosoomide vahele. 2. Prometafaas
Algab järsku tuumamebraani lagunemisega vesiikuliteks. Need vesiikulid jäävad nähtavaks kääviniidistiku ümbruses kogu mitoosi ajal. Kääviniidid, mis alguses olid väljaspool tuuma, hõlmavad nüüd ka selle ruumi, kus enne oli tuum. Kromosoomide tsentromeeride külge moodustuvad kinetohoorid, mis seovad enda külge osa mikrotuubulitest - neid nim. nüüd kinetohoorseteks mikrotuubuliteks. Käävi ülejäänud mikrotuubuleid nim. polaarseteks, väljapoole käävi jäävaid mikrotuubuleid aga astraalseteks. 3. Metafaas
Kinetohoorsete mikrotuubulite abil kromosoomid reastatakse ühele tasapinnale kahe pooluse vahel. Sel hetkel on kromosoomid juba liikumiseks valmis, nende kinetohoorid on aktiivsed ning kumbki kinetohoor püüab liikuda poolusele. Liikumist aga veel ei toimu, sest kinetohooride poolt tekitatav jõud tasakaalustatakse vastastikku tänu sellele, et tütarkromatiidid on tsentromeeri abil ühendatud. Algab järsku, tütarkromatiidid alustavad liikumist pooluste suunas. Liikumise kiirus on ca 1µm/min. Anafaas kestab tüüpiliselt mõne minuti.
Lahknevad tütarkromatiidid (kromosoomid) jõuavad poolustele, kinetohoorsed mikrotuubulid kaovad. Tütartuumade ümber moodustub uus tuumaümbris. Kromatiin dekondenseerub, ilmuvad uuesti tuumakesed.
6. Tsütokinees
Algab tihti juba anafaasis. Aktiinist ja müosiinist moodustub kontraktiilne struktuur aktiini rõngas, mis paigutub raku keskele käävi teljega risti. See struktuur tekitab jõu, mis on vajalik plasmamebraani sissenöördumiseks. See lõpeb kahe uue tütarraku eraldumisega teineteisest.
Interfaas : G1- R – S – G2 – R - G0
S-faas: algab DNA sünteesiga, S-faasi vältel DNA on kopeeritud. RNA ja valkude süntees on väga aeglased selles faasis.
G1 ja G2 faas annavad rakule vajaliku aja kasvamiseks: kui interfaas oleks ainult nii pikk, mis oleks hädavajalik DNA replikatsiooniks, siis ei jätkuks aega muude rakukomponentide kordistamiseks. G1-faasi ajal rakk seirab oma ümbrust ning omaenda suurust, ja kui aeg on küps, siis rakk alustab DNA replikatsiooni. G1 faasi lõpul on kontrollpunkt , kus rakutsükkel vajadusel peatatakse. Kõrgematel eukarüootidel nimetatatakse seda R-punktiks (restriction point) või ka lihtsalt G1-faasi kontrollpunktiks. Selle punkti läbimisel käivitatakse rakus DNA replikatsioon ja algab seega S-faas. G2-faas annab rakule vajaliku aja, mis võimaldab tal kontrollida, kas DNA replikatsioon on lõpetatud. Läbides G2-faasi kontrollpunkti, alustab rakk M-faasi. Kõrgematel eukarüootidel toimivad väliskeskonnast tulevad rakutsüklit seiskavad signaalid valdavalt G1-kontrollpunktile, seetõttu G1-faasi pikkus võib eri rakkudel olla väga erinev.
G0-faas: puhkefaas.
Imetaja rakkude puhul, mida on kasvatatud koekultuuris, kestab rakutsükkel umbes 24 tundi. G1 faas kestab 10 tundi ning sel ajal toimub rakus normaalne metabolism , rakk kasvab suuremaks , temas sisalduvate organellide arv kahekordistub ja toimub ettevalmistus DNA replikatsiooniks. S faas algab DNA replikatsiooniga ning kestab ligikaudu 9 tundi. S faasi lõpuks koosnevad kromosoomid kahest tütarkromatiidist.
Kui replikatsioon on lõppenud, läheb rakk faasi G2, mis kestab neli tundi. Selles faasis jätkub raku kasvamine ja ta valmistub mitoosiks. Mitoos (M faas) e. raku jagunemine kestab ligikaudu tunni.
21. Milleks on gameetide küpsemisel vaja meioosi?
Gameetide keharakkudes on diploidne kromosoomistik , et sugulisel paljunemisel kromosoomide arv viljastumise tulemusena ei kahekordistuks, peab kromosoomide arv vähenema 2x.
22. Mis on moorula, blastula, gastrula ?
Moorula( kobarloode ): peale viljastumist jagunema hakanud sügoot. Totipotentsed rakud mis võivad regenereerida terviklikku organismi. Kõrgematel loomorganismidel on totipotentne rakk viljastatud munarakk ning väga varajase embrüo(moorula) rakud.
Blastula: moorula rakud paigutuvad ümber ja moodustab blastotsüst, see vorm vastab blastulale. Blastotsüst on seest õõnes põisloode, mille sein koosneb ühest rakukuhist. Selle ühel poolusel on rakukobar(embrüoblast), millest areneb loode.
Gastrula e. karikloode : (järgneb blastotsüsti staadiumile)
eristub 2 rakukihti (lootelehed)
a) väline e. ektoderm
b) sisemine e endoderm
- need muutused toimuvad varases gastrula staadiumis.
- hiljem eristub ka kolmas e. c)mesoderm
Igast lootelehes arenevad välja kindlad elundid ja elundkonnad . Ektoterm paneb aluse närvisüsteemile, meeleelunditele ning naha ja suu epiteelkoele, küüned, karvad, hammaste vaap . Mesoderm moodustab tugi, liikumiseldundkonna (luud, lihased) vereringe, eritus ja sigimiselundkonna, Entotermist seede ja hingamiselundkond .
23. Mis on alleel , homosügootsus, heterosügootsus?
Alleel: ühe geeni erinevad vormid on alleelid. (ntks kollast seemne värvust määrav geen A ja rohelist värvust määrav a on sama geeni erinevad vormid)
Homosügootsus: Geenipaari seisund, millepuhul mõlemas homoloogilises kromosoomis paikneb vaadeldava tunnuse suhtes sama alleel. (AA või aa)
Heterosügootsus: geenipaari seisund, mille puhul mõlemas homoloogilises kromosoomis paikneb vaadeldava tunnuse suhtes erinev alleel. (Aa)
24. Mis põhjustab geenide ahelduse ?
Geeniaheldus (gene linkage) -- geneetilise ahelduse kitsam, klassikaline mõiste, mis tähendab piiratud (mittevaba) rekombineerumist geenide vahel genotüüpide pärandumisel, st. Mendeli III (sõltumatu lahknemise) seaduse rikkumist. Geeniaheldus on tingitud geenilookuste lähestikusest paiknemisest piki kromosoomi. Aheldunud geenid rekombineeruvad ristsiirde kaudu, mille sagedus oleneb geenidevahelisest kaugusest kromosoomis ( Morgani seadus).
Geneetiline aheldus on tingitud geneetiliste markerite lähestikusest paiknemisest ühes lineaarses struktuuris, ühes kromosoomis või muus genoomses nukleiinhappemolekulis (NH).
25. Mis on replikatsioon?
Nukleiinhappe (DNA või RNA) ahel kasvab 5’→3’ suunas. Nukleiinhappe sünteesil lülituvad sünteesitavasse ahelasse nukleotiidid, mis on komplementaarsed matriitsahela (informatsiooni-allikas ehk kopeeritav ahel) nukleotiididega. Matriitsina toimib üksikahelaline nukleiinhape . Seega peab kaksikahelaline DNA replikatsiooni initsiatsioonisaidist olema eelnevalt viidud üksikahelaliseks. DNA ahelate eraldumist teineteisest viib läbi ensüüm DNA helikaas . Sünteesi viivad läbi ensüümid, mida nimetatakse polümeraasideks. DNA replikatsiooni käigus lülitatakse kasvavasse DNA ahelasse inimesel 3000 nukleotiidi minutis , bakteril 30,000 nukleotiidi minutis. Replikatsioonil käituvad matriitsina mõlemad DNA ahelad (mõlema ahela kopeerimine toimub paralleelselt) ning replikatsiooni lõpp- produktideks on kaksikheeliksid, milles üks ahel on uus ja teine vana. Seetõttu nimetatakse sellist DNA replikatsiooni mudelit semikonservatiivseks (1 uus/1 vana ahel + 1 uus/1 vana ahel) (joonis 3). DNA replikatsioon toimub kindlas rakutsükli faasis- S faasis, mis koos G1(raku normaalne metabolism ja kasvamine;temas sisalduvate organellide kahekordistumine, ettevalmistus replikatsiooniks) ja G2(raku kasvmine, ettevalmistus mitoosiks) faasidega moodustab interfaasi) ning rakk hakkab peale replikatsiooni jagunema ainult tingimusel, et DNA replikatsioon on toimunud veatult.
Semikonservatiivne replikatsiooni mudel.
26.Mis on geen?
Geen on kromosoomi kindlas lookuses (geeni asukoht kromosoomis) paiknev pärivustegur, mis määrab otse või kaudselt (tihti koostoimes teiste geenidega) ühe või mitme tunnuse arengu; DNA-molekuli funktsionaalne lõik, mis tavaliselt sisaldab informatsiooni (mRNA vahendusel) ühe valgu (või polüpeptiidi) sünteesiks (kuid on ka nt rRNA ja tRNA geenid, mis valgumolekule ei kodeeri). Eristatakse struktuur- ja regulaatorgeene. Geenid eksisteerivad struktuurilt ja tihti ka funktsioonilt erinevate alleelide kujul. Geenide oma- ja keskkonnavahelisest mõjust on tingitud organismi areng ja fenotüüp.
27. Transkriptsioon
Informatsioon valgu primaarstruktuuri kohta on salvestatud rakutuuma DNA-s.
Info ülakanne tuumast ribosoomidesse toimub mRNA e informatsiooni RNA vahendusel.
Transkriptsioon on geeni struktuuris sisalduva info ümberkirjutamine mRNA-le rakutuumas interfaasi ajal RNA- polümeraasi toimel.
RNA- polümeraas seostub promootoriga( kinnitumise piirkond DNA-l) ja keerab järk- järgult lahti DNA biheeliksi.
RNA- polümeraas sünteesib DNA ahelaga komplementaarse mRNA molekuli.
Terminaator on koht DNA-l, kus lõpeb transkriptsioon ja DNA omandab endise biheeliksi kuju.
Transkriptsioon toimub rakutuumas.
28. Geneetiline kood.
Geneetilisele koodile on iseloomulikud järgmised omadused:
- Geneetiline kood põhineb nukleotiidide tripletitel: kolm nukleotiidi, mis moodustavad koodoni, määravad ära ühe aminohappe polüpeptiidis;
- Geneetiline kood ei ole kattuv : iga nukleotiid mRNA molekulis kuulub ainult ühte koodonisse(v.a. erandjuhtumid, kus geenide nukleotiidne järjestus DNA-s kattub ning tulemusena kodeeritakse erinevaid valke).
3) Geneetiline kood on komavaba: pole väljajäetavaid nukleotiide, kõik mRNA-s asuvad koodonid loetakse translatsioonil järjest, ühes lugemisraamis (reading frame ); Lugemisraame saab põhimõtteliselt olla 3 erinevat, edasi hakkab minema nagu 1. lugemisraam, kuna kood koosneb tripletitest (joonis 21).
4) Geneetiline kood on degenereerunud e. kõdunud: peaaegu kõigile aminohapetele vastab enam kui üks koodon (v.a. metioniin/Met ja trüptofaan/Trp, joonis 20);
5) Geneetiline kood on seaduspärane (ingl. k. ordered): ühte aminohapet määravad koodonid ning samuti sarnaseid aminohappeid määravad koodonid on oma nukleotiidselt järjestuselt sarnased ja erinevad sageli üksteisest vaid ühe nukleotiidi poolest (näiteks Phe ja Leu, joonis 20);
6) Geneetilisse koodi kuuluvad spetsiifilised koodonid, mis on signaaliks translatsiooni initsiatsioonile ja terminatsioonile;
7) Geneetiline kood on (v.a. mõned erinevused mitokondrites) universaalne kõigile elusorganismidele.
29.Translatsioon
Translatsioon on mRNA- s nukelotiidide järjestusena salvestatud informatsiooni ülekandumine aminohapete järjestuseks sünteesitava valgu molekulis.
Translatsioon toimub ribosoomides.
Protsess algab mRNA ühinemisest ribosoomiga.
mRNA initsiaatorkoodoniga seostub esimene tRNA e transpordi RNA molekul, mis tunneb antikoodoni abil ära mRNA vastava koodoni.
tRNA molekul on aminohappespetsiifiline, st igat aminohapet transpordib kindel tRNA.
Aminohapete vahele sünteesitakse peptiidside.
Kasutatakse ATP ja GTP energiat.
30. Valgusünteesi regulatsioon.
Aminohape liidetakse kasvavasse polüpeptiidahelasse tRNA abil. Aminohape seotakse otse tRNA molekuliga ning moodustub aminoatsüül-tRNA. Vastavat protsessi nimetatakse tRNA aktiveerimiseks e. aminohappega laadimiseks ning seda viivad läbi ensüümid, mida nimetatakse aminoatsüül-tRNA süntetaasideks (näit. alaniiniga aktiveeritud tRNA on alanüül-tRNAAla). Kõigile 20-le erinevale aminohappele on rakus vähemalt üks spetsiifiline aminoatsüül-tRNA süntetaas. Samuti iga spetsiifiline aminoatsüül-tRNA-süntetaas tunneb ära teatud aminohappele vastavaid tRNA molekule, st. et iga aminoatsüül-tRNA süntetaas seob omavahel kokku ühe aminohappe ja sellele vastava tRNA. Igale aminohappele vastab rakus aga üks kuni neli erinevat tRNA molekuli.
Valgu süntees toimub vesikeskkonnas .
31. Ribosüümid.
1980. aastal uuris T.Cech alglooma DNA-struktuuri ja avastas ribosüümid. Ribosüüm koosneb ribonukleiinhappest ja ensüümist. Need on RNA ensüümid.
Esimestel avastatud RNA ensüümidel oli katkilõikamise võime(RNA ahela?). Hiljem avastati, et neil on ka sünteesimisvõime.
Cech sõnastas RNA word’, mis püüdis sõnastada elu teket maal.
32. Geen- ja kromosoommutatsioonid.
Geenmutatsioon - hälve on mõnes nukleotiidis geeni sees.
Nukleotiidid on rplikatsioonil: kaduma läinud; juurde tulnud; asendunud teise nukleotiidiga.
Ei ole kahjulikud kuni selleni , millal mutatsioonid kuhjuvad ühte piirkonda. Selle tulemusena võib tekkida suhkruhaigus .
Kromosoommutatsioonid- muutused kromosoomides- kromosoomide pikkuse või struktuuri muutused:
Deletsioon - kromosoomilõik on kaduma läinud.
Duplikatsioon- kromosoomilõik on kahekordistunud.
Iversioon- geenide järjestus on muutunud ümberpaiknemise tõttu.
Translokatsioon - geenide asukoht on muutund ümberpaiknemise tõttu.
Nii võib tekkida näiteks skisofreenia .
33. Viiruste ehitus
Viiruste osakesed - virionid (üksik viiruse isend )- omavad hoolimata oma erinevast päritolust mõningast ehituslikku sarnasust . Enamasti sisaldab virion valkudest koosnevas kestas (nukleokapsiidis) ühe või mitu molekuli nukleiinhapet. Nukleokapsiid võib koosneda ühesugustest valgu molekulidest, või olla üsna keerulise ehitusega. Mõnikord ümbritseb nukleokapsiidi veel peremeesraku rakumembraanist lipiidkest. Mõnede viiruste pinnavalkude või polüsahhariidide ülesandeks on seonduda peremeesraku pinnaretseptoritega. On ka viirusi, mis omavad virionis paljunemiseks või rakku tungimiseks vajalikke ensüüme, isegi ribosoome (nt Ebola viirus ). Neil 5 erinevat ehitustüüpi: keraviirus, silinderviirus, kristalliline viirus ehk tubaka mosaiigiviirus, kinnitusniitidega viirus ehk sabaga viirus, saabasviirus ehk kahekordse kapsiidiga viirus.NB! virion- viiruse rakuväline vorm, mis on ette nähtud levimiseks uuele peremehele.
Erinevaid viiruste ehitustüüpe: ikosaeedriline, spiraalne, faagikujuline
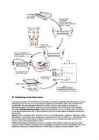
34. Viiruste paljunemine.
Kõik viirused on elusorganismide siseparasiidid. Nad sisenevad peremeesorganismi rakku, või sisestavad sinna oma nukleiinhappe. Viiruslik nukleiinhape (DNA) integreerub peremeesraku kromosoomi, või ka transleeritakse otse, mõnede RNA-viiruste RNA põhjal valmistatakse rakus sellele vastav DNA, mis kromosoomi integreerub (pöördtranskriptsioon; vt retroviirused).
Nakatumise esimest staadiumit, kus viirus end nö. rakus "sisse seab" ja oluliselt viimase normaalset talitlust ei mõjuta, nimetatakse lüsogeenseks faasiks. See võib sõltuvalt tingimustest ja viirusest kesta mõnest minutist paljude aastateni.
Nakatumise teine staadium kannab nime lüütiline faas, mille käigus raku normaalne elutegevus katkestatakse, ning kujundatakse ümber uute viirusosakeste tootmiseks. Uued virionid väljuvad rakust (sellega võib kaasneda raku surm) ning üritavad nakatada uusi rakke. Selline viiruste paljunemine ja organismi normaalse talitluse häirimine kutsub enamasti esile vähem- või rohkem tõsise viirushaiguse. Viiruste sisenemine peremeesraku genoomi võib esile kutsuda ka muid raku talitluse häireid, millest tähtsaimaks võib pidada raku muutumist kasvajarakuks. Raku talitluse häirimine ja raku või peremeesorganismi tapmine pole viirusele tegelikult enamasti kasulikud, kuna see võtaks viiruselt võimaluse edasi paljuneda. Nii ongi levinuimad ja ökoloogiliselt edukaimad need viirused, mis peremeesorganismi normaalset elutegevust vähe mõjutavad.
35. Bakteri-, putuka-, ja taimeviiruste eripärad.
Iga viirus on võimeline nakatama ainult teatud tüüpi rakke. Vastavalt sellele klassifitseeritakse viiruseid loomaviirusteks, taimeviirusteks ning bakteriviirusteks e. bakteriofaagideks. Viirusega nakatumiseks on vaja, et viiruse välispind interakteeruks rakupinna spetsiifiliste retseptoritega. Loomaviiruse retseptorid paiknevad plasmamembraanil, faagide retseptorid bakteriraku rakukestal või kiududel e. piilidel.
36. Kuidas imetaja organism kaitseb ennast viirusnakkuste eest?
Viirushaigusi ehk viroose esineb kõikidel rakulise ehitusega olenditel. Inimesel tuntakse umbes viissada levinumat viirushaigust, kuid nende arv on tegelikult arvatavasti märksa suurem ( inimeselt isoleeritud viiruste liike on kokku üle tuhande). Kõige tõhusamaks viirushaigusi tõrjuvaks vahendiks on seni organismi enda immuunsüsteem, mille võimet viirustega võidelda suurendatakse vaktsineerimise abil. Viimasel ajal on teatud edu saavutatud ka viiruste kemoteraapias, kuid ravimite väljatöötamine viiruste vastu on raske (selle peamiseks põhjuseks on asjaolu, et viirused kasutavad oma elutegevusel raku enda vahendeid).
Lisaks viiruste kahjulikkusele haiguste tekitajana on viirused oluliseks tööriistaks geeniteraapias, kuna võimaldavad rakkudesse viia ka kasulikke geene, mis rakus muidu puuduvad või on kahjustatud (vt viirusvektor )
37. Prokarüootide kaks peamist rühma (riiki). Mis neid iseloomustab?
Bakterid , tsüanobakterid,aktinomütseedid.
Puudub piiritletud tuum,esineb vähem organelle ja membraanseid struktuure,suurus 0,1-25 mikromeetrit.
38. Bakteriraku ehitus.
Bakterid on prokarüootsed organismid-DNA ei ole eraldatud tuumamembraaniga ja asub tütoplasmas tuumapiirkonnas. Haploidne kromosoomistik; 1 rõngakujuline kromosoom.Genoomi suuru ehk DNA hulk on väiksem kui eukarüootidel. Tuumapiirkonda nim.nukleoidiks.Plasmiididel ehk DNA rõngasmolekulidel on geenid, mis pole bakterile tavaolukorras vajalikud.Antibiootilises keskkonnas võivad olla need geenid olulisteks resistentsusfaktoriteks. Tsütoplasmat katab rakumembraan ehk plasmamembraan. Membraani ümbritseb rakukest. Selle pinnal on limakapsel , mis kaitseb bakterit kuivamise eest. Tsütoplasmas on mitmesugused sisaldised:varuainete (tärklis, glükogeen, polüfosvaadid, väävel jne) terakesed.
Bakteritel puuduvad mitokondrid, plastiidid , tsentrioolid, tsütoplasma võrgustik, Golgi kompleks, lüsosoomid.
Bakterirakke isloomustavad:
viburid, mis aitavad liikuda
karvakesed ehk piilid , raku pinnal olevad valgulised karvakesed, mis on vajalikud kinnitumiseks
aerosoomid ehk gaasivakuoolid, mis reguleerivad vees elavatel tsüanobakteritel raku erikaalu
spoorid , mis moodusuvad bakteritel ebasoodsate tingimuste üleelamiseks:rakukest pakseneb, veesisaldus väheneb ja ainevahetus aeglustub.Ühes rakus moodustub üks spoor . Spoorid võivad säilida idanemisvõimelistena väga pikka aega.
39. Bakterite kasv ja paljunemine.
Bakterite paljunemine toimub pooldumise teel.Taoimub ainete biosüntees, DNA ja plasmiidide replikatsioon. Rakukest ja plasmamembraan sopistuvad sisse. Moodustub rakuvahesein. Tekib kaks tütarrakku.
Generatsiooniaeg on ajavahemik , mis kulub bakterite populatioonis rakkude arvu kahekordistumiseks.
Koloonia on bakterite populatsioon , mis moodustub tardsöötmel ühe raku järglaskonnana.(koloonias umbes 109 rakku)
40. Eukarüootide riigid ja nende peamised tunnused.
Taimed, loomad, proistid, seened
Rakul on olemas tuum;ainu-ja hulkraksed organismid. Rakkude kuju on väga erinev;suurus 10-20 mikromeetrit.
41. Seeneraku ehitus.
Eukarüootsed rakud, eristatakse ühe-ja hulkrakseid seeni.
Üherakulised:ümarad päristuumsed rakud(pärmseened) või hulgatuumalised vaheseinteta hüüfid(must täpphallik)
Hulkraksed:hüüfides esinevad vaheseinad( kottseened ). Seened, va pärmseened, moodustuvad torujatest rakkudest- seeneniitidest ehk hüüfidest, mis on liigist sõltuvalt kokku pakitud seeneniidistikuks ehk mütseeliks.
Seeneraku rakumembraani katab kitiinist ja teistest süsivesikutest koosnev rakukest.
Rakumembraan ja –kest on õhukeseseinalised , et ensüümide abil lagundatud ühendid saaksid imenduda seenerakku . Kõige aktiivsem imendumine toimub hüüfi tippudes.
( Heterotroofne , piiramatu rakkude jagunemisvõime, varusüsivesinikuks on glükogeen, tsentrosoom on osades rakkudes, plastiidid puuduvad, on väikesed lipiidivakuoolid.)
42.Taimeraku ehitus.
Rakukest koosneb tselluloosist, ligniinist ja pektiinist.
Noorel rakul on kest õhuke, raku vananedes see pakseneb.
Raku kestades on poorid, milel kaudu toimub ainevahetus. Naaberrakkude tsütoplasmad ühenduvad tsütoplasmaväätide ehk plasmodesmide kaudu.
Taimerakus on vakuoolid, plastiidid(proplastid, kloroplastid, kromoplastid, leukoplastid), ribosoomid, karedapinnaline ja siledapinnaline tsütoplasma võrgustik, tuum, mitokonder, Golgi kompleks, tsütoplasma, lüsosoomid.
43. Restriktaasid.
Ehk endonukleaasid – lagundavad nukleiinhapet, lõhkudes suhkur-fosfaat selgroos nukleotiidide vahelist sidet ehk fosfodiestersidet ahelasiseselt, mitte otstest.
Restriktaase toodavad bakterid. Restriktaasid on järjestusspetsiifilised.
44. DNA kloneerimise etapid.
Lõigatakse nii vektorit B-saidis(vektoriteks kasutatakse tihti plasmiide) kui ka uuritavat DNA-d retriktaasiga, mis genereerib “kleepuvad” otsad . (kui nii kloneerimisvektori DNA-d kui ka kloneeritavat DNA-d on lõigatud ühe ja sama restriktaasiga, saame klonerimisvektoi otste vahele “kleepida” uuritava DNA lõigu. Fosfodiestersidemete moodustamiseks kloneerimisvektori DNA ja uuritava DNA lõigu otse vahele kasutatakse ensüümi DNA ligaas )
DNA fragment kleebitakse vekroti B-saiti
Uus konstrukt viiakse bakterirakkudesse.
45.DNA sekveneerimise põhimõte.
DNA nukleotiidse järjestuse määramine.Põhineb DNA fragmentide populatsiooni analüüsil, kus kõik fragmendid algavad samast nukleotiidist, kuid lõppevad sellest statistiliselt erineval kaugusel.Korraga analüüsitakse nelja fragmentide segu, millest ühest lõppevad kõik fragmendid suvalisest G nukleotiidist, teises A nukleotiidist, kolmandas T ja neljandas C. DNA sekveneerimisel kasutatakse nii keemilist kui ensümaatilist meetodit.Keemiline põhineb keemilistel reaktsioonidel, mis võimaldavad DNA ahelat spetsiifiliselt katkestada kas A, G, C või C + T kohalt. See oli kasutusel DNA sekveneerimise algaastatel.
Ensümaatilise meetodi töötas välja Sanger. Viiakse läbi DNA in vitro süntees, kus reaktsioonisegusse on lisatud DNA polümeraas, 4 erinevat desoksüribonukleotiidi ja 4 spetsiifilist terminaatornukleotiidi(ddNTP), millest on igaüks märggitud erineva flourestseeruva märgisega. Selle tulemusena tekivad DNA fragmendid, mis algavad kõik samast kohast, kuid nende süntees on termineerunud erinevatest kohtadest . Sünteesitud DNA fragmendid lahutatakse polüakrüülamiidgeeli. Geelelektroforeesi tingimused on sellised, et sünteesitud DNA fragmendid eristuvad geelist üksteisest ühenukleotiidse täpsusega. Mida ühem onfragment, seda kiiremini liigub ta geelis. See võimaldab DNA fragmentide mustrist lugeda välja DNA lõigu nukleotiidset järjestust.
46. Polümeraasi ahelreaktsioon.
PRC meetod võimaldab spetsiifiliselt amplifitseerida(saada DNA paljundamisel koopiaid) suvalist DNA järjestust. PRC rakendamiseks on vaja teada amplifitseeritavast DNAst primaarjärjestust paljundatava DNA piirkonna mõlemast otsast.PRC toimub kolmeetapiliselt:
Amlifitseeritava DNA denatureerimine emperatuuri toimel (92-95 kraadi juures)
DNA renatureerimine temperatuuri langetamisel ülehulgas lisatud sünteetiliste oligonukleotiidsete praimerite juuresolekul, millest üks on komplementaarne amplifitseeritava DNA ühele ahelale, teine selle vastasahelale.Selle toimel hübridiseeruvad praimerid amplifitseeritava DNA ahelatega.
DNA replikatsioon ahelate kinnitunud
47.Kromatograafia
Keemiliste ühendite füüsikalis-keemiline ükteisest lahutamine, mis põhineb aineosakeste korduval ümberjaotumisel liikuva ja liikumatu faasi vahel. Liikuvas faasis olevas aineosakesed kanduvad selle vooluga kaasa järjekordse siirdumiseni liikumatusse faasi. Viimase põhjustatud liikumispauside kestus ja aineosakeste edasikandumise keskmine kiirus sõltuvad igale keemilisele ühendile eripärasest jaotumissuhteks nimetatavast suurusest .Sellest tulenev ühendite erisugune liikumiskiirus faaside kokkupuute piirkonnas põhjustab ühendite eraldumise ükteisest. Jaotumisprotsessi olemuse järgi eristatakse kuute erinevat kromatograafiat.Liikuva faasi agregaatoleku põhjal eristatakse gaasi- ja vedelikukromatograafiat. Liikumatu faasi paigutuse põhjal eristatakse kolonn - ja kihtkromatograafiat.
Esimesena võttis kasutusele M.Tswett.
Rakendatakse keemilises analüüsisja eriti puhastekeemiliste ühendite saamisel.
48. Elektroforees
Makromolekulide separeerimine elektriväljas.
Kolloidosakeste ja makroioonide liikumine elektrvälja mõjul katoodile(kataforees) või anoodile (anaforees). Seda põhjustab osakeste elekrilaeng, mis kolloidosakeste puhul tuleneb osakeste tuumadel absorbeerunud ioonidest. Elektroforees võib toimuda ka lahustiga immutatd kandmaterjalil-filterpaberil(paberelektroforees) või geelil(geelelektroforees).
Elektroforeesi rakendatakse metallide pinnale kolloidosakestest kattekihi tekitamiseks, kõrgmolekulaarsete ühendite (nt valkude) segude analüütiliseks lahutamiseks, meditsiinis jms.
49. Nukleiinhapete hübridiseerimine.
Kui segada üheahelalisi DNA või RNA järjestusi, millel on omavahel komplemen-taarseid järjestusi, siis need piirkonnad paarduvad vastavalt komplementaarsuse printsiibile, moodustades DNA-DNA või DNA-RNA kaksikheeliksi. Kui nukleiinhapped pärinevad eri-nevatest allikatest, siis nimetatakse neid duplekse hübriidmolekulideks. Meetodeid , mis põhinevad komplementaarsete NH-te piirkondade võimel paarduda, nimetatakse hübridiseerimismeetoditeks. Hübridiseerimismeetoditel on väga lai kasutusspekter. Eelkõige kasuta-takse hübridiseerimist NH-te identifitseerimisel, näit DNA raamatukogude skriiningul ja kloneerimise abivahendina.
Hübridiseerimise kasutamise NH-te identifitseerimisel teeb võimalikuks NH-te märkimine. Märgitakse tuntud NH fragment e sünteesitakse proov . Kasutatakse nii radioaktiivseid (32P, 33P, 35S) kui ka mitteradioaktiivseid (biotiin, digoksigeniin) NH-te märkeid. Mär- gitud proovide abil saab visualiseerida väga väikeseid DNA koguseid,


























































Kõik kommentaarid