1. Spermatogenees Imetajate
spermatogeneesi etapid:
(Kus ja millal toimuvad? Kirjelda igas
etapis toimuvaid protsesse.
Spermatogenees
toimub testises ehk munandis
(täpsemalt
väänilistes
seemnetorukestes),
luumeni osas ja verest eraldatult, et ei saaks tekkida immuunvastust.
Puberteedieas
kuni elu lõpuni.
- Paljunemine – ehk jagunemine (mitootiline); toimub spermatogoonide hulga suurendamine , toimub spermatogeenses epiteelis (seemnetorukese sees)
- Kasvamine – tsütoplasma hulga näol
- Küpsemine – (meiootiline) saavutatakse haploidne kromosoomistik ; tulemuseks sekund. Spermatotsüüdid; puberteedieas
- Transformatsioon – ehk spermiogenees; moodustub vibur (munaraku puhul ei toimu); toimub seemnetorukese valendikus; tulemuseks spermatiidid
- Spermatogoonid – meioosieelne isassuguraku (spermi) eellasrakk (2n), mis asub seemnetorukese perifeerses osas. Diferentseerumise käigus liiguvad perifeeriast seemnetorukese valendiku suunas.
- Spermatotsüüdid – rakud , mis teevad läbi 2 meiootilist jagunemist (spermatogeneesi) ja millest moodustuvad spermiidid. Esmased spermatotsüüdid teevad läbi esimese meiootilise jagunemise, sekundaarsed spermatotsüüdid tekitavad teise meiootilise jagunemise teel spermatiidid. Ühest spermatogoonist (2n) tekib meioosi tulemusena 4 spermatiidi.
- spermatiidid – moodustunud spermatogoonist ehk ürgsugurakust meioosi teel- haploidsed. Nendest küpsevad edasi spermatosoidid, kui omandavad transformatsiooniprotsessis liikuvuse .
- Spermid – küps/ väljaarenenud isassugurakk.
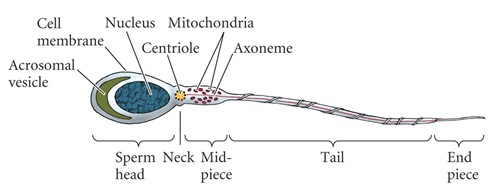
Spermi
ehitus:Keskosa
on mitokondritega; saba on mikrotorukestest koosnev vibur.
Akrosoom
on Golgi päritolu. Tuum on tugevamini kokku pakitud kui
som.rakkudes.
Aksoneemi
süntees algab tsentrioolist ja see on kõik 1
rakk .
Tuum
–
kaelaosa (tsentriool) – keskosa (mitokondrid) – saba –
lõpuosa
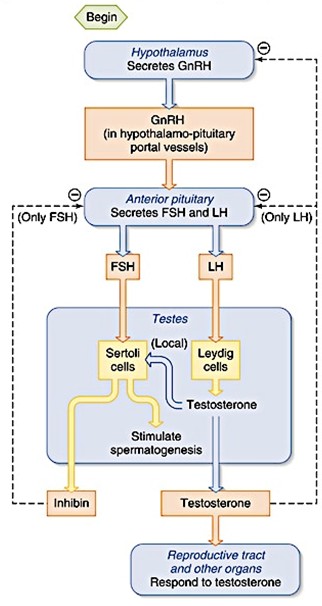
Leydigi
rakud
– toodavad testosterooni (seemnetorukeste vahel)
Sertoli rakud
– tugirakud; ümbritsevad küpsevaid sperme; toidavad ja suunavad
neid (spermatogeneesi); mööda nende külge toimub kogu
spermatogeneesi protsess. Väga harunenud kujuga. Ei jagune, tema
tuumas on näha tuumake. Moodustavad vahel tiheliiduseid ja jagavad
spermatogeense
epiteeli basaalseks (spermatogoonid) ja atluminaalseks
osaks (spermatotsüüdid ja spermid).
Spermatogeneesi hormonaalne regulatsioon
(GnRF, FSH, LH,
testosteroon ).
2. Oogenees Imetajate
oogeneesi etapid:
(Kus, millal toimuvad? Kirjelda igas etapis toimuvaid protsesse.)
Toimub
munasarjas;
meioos
peatub I profaasis diakeneesis
(enne sündi) ja
jätkub
puberteedieas.
Gonaadi jõudes on ootsüütide arv kõige suurem.
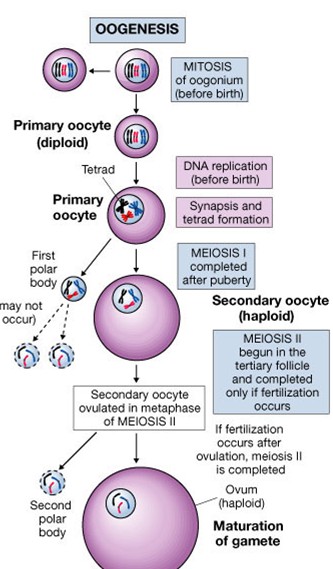
- Paljunemine – mitootiline; oogoonist saab (primaarne) ootsüüt
- Kasvamine - meiootiline; ootsüüdist munarakk ; rebumoodustumine; jätkub puberteedieas
- Küpsemine – sekund ootsüüdist munarakk (?)
- Oogoonid – emasloomade sugurakkude eellasrakud (2n) enne meioosi toimumist .
- Ootsüüdid – rakk, mis läbib 2 meiootilist jagunemist (oogenees), moodustab munaraku. Primaarne ootsüüt on rakk pärast meioosi 1.jagunemist ja sekund.ootsüüt kas meioosi II jagunemise mingis faasis või pärast meioosi teist jagunemist, aga enne munaraku lõplikku küpsemist. Oogeneesi käigus tekib phest oogoonist (2n) küps munarakk (n) ja 3 polaar - ehk reduktsioonkeha (n).
Munaraku
erinevus keharakkudest
–
munarakus on haploidne kromosoomistik; on mõõtmetelt suurem;
kaetud mitmete kestadega; tsütoplasma-tuuma suhe on erinev
(
keharakus 1:10, munarakus 1:1000, seemnerakus 1:1);
ainevahetus on
väheaktiivsem.
Munaraku
kestad, nende päritolu:
- Primaarsed munakestad – munaraku poolt tekitatud vitelliinkest e rebukest; imetajatel zona pellucida.
- Sekundaarsed munakestad – neid toodavad abirakud, nt putukate koorin
- Tertsiaarsed munakestad – lisanduvad munajuhas, nt munavalge
Kirjelda
imetajate follikulaarse oogeneesi etappe :
- Primaarne folliikul – esineb primaarseid ühekihilisi ja prim.mitmekihilisi folliikuleid . Esimesed neist on kaetud 1-kihilise kuupepiteeliga, keskel on munarakk ning hakkab moodustuma oolemm. Mitmekihilise folliikuli puhul on epiteel mitmekihiline , munarakk on ikka keskel ja on moodustunud ka zona pellucida.
- Sekundaarne folliikul – hakkab formeeruma teeka – sidekoeline kihn; munarakk liigub perifeeriasse; folliikuliõõne alged
- Tertsiaarne ehk Graafi folliikul – teeka on 2-kihiline (sise- ja välisteeka); folliikuliõõs on täidetud vedelikuga; munakühm, kus paikneb munarakk; folliikuliepiteeli nim granuloosaks; vahetult munarakku ümbritsevad epiteelirakud paiknevad kiirjalt ja mood kiirpärja ( corona raiata)
Kasvavate
ja jagunevate granuloosrakkude vahele moodustub vedelikku ning tekib
küps munasarjafolliikul ehk Graafi folliikul. Seda ümbritseb
sidekoeline munasarjafolliikulikihn. Selle kihnu rakud (teekarakud)
ning granuloosrakud valmistavad naissuguhormoone ehk östrogeene.
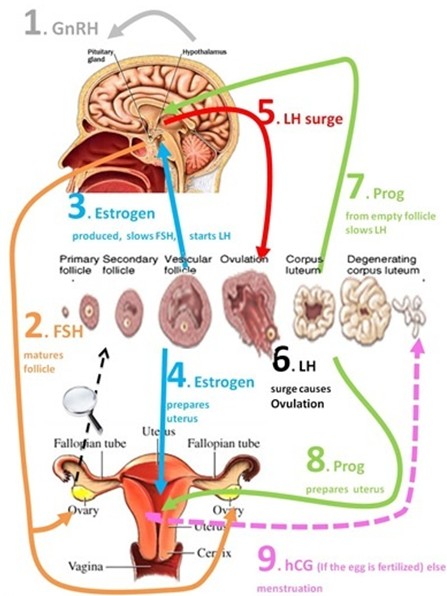
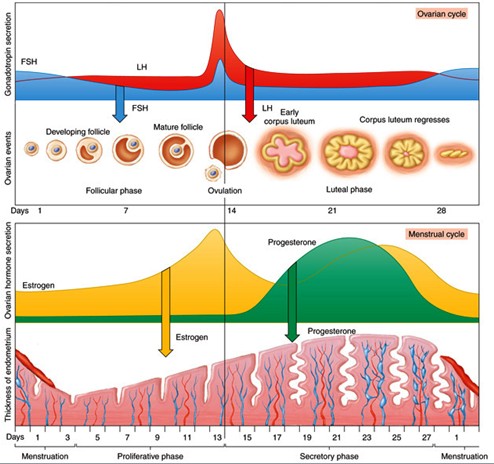
Oogeneesi
hormonaalne regulatsioon (GnRF, FSH, LH, östrogeen, progesteroon ).Gonadotropiinid:
FSH, LH, hCG - neid stimuleerib
ajuripats Steroidhormoonid:
östrogeen, progesteroon
1)
Hüpotalamuselt
tuleb
signaal ajuripatsile (gonadotropiini
hormoon )
2)
ajuripatsilt vabaneb
folliikuleid
stimuleeriv hormoon
– folliikulid hakkavad kasvama; üks folliikul valitakse välja
3)
folliikuli rakud toodavad
östrogeeni
– see omakorda mõjutab ajuripatsit nii, et see hakkab tootma
luteniseerivat
hormooni
(kantakse munasarja) (östrogeen stimuleerib ka maksa tootma
vitellogeniini)
4)
luteniseeriva hormooni toimel
munarakk
vabaneb,
vabaneb ka meioosi arrest ning meioos läheb veidi edasi ja seejärel
jälle arrest.
7)
kollaskeha hakkab tootma
progesterooni
– paneb vihama emakaseina ja inhibeerib luteniseeriva hormooni
sünteesi ajuripatsis (sest seda pole enam vaja, kuna munarakk on
vabanenud) et munarakke enam ei vabastataks.
*Kui
viljastumine toimub, siis sügoot kinnitub emakaseina – see hoiab
progesterooni ja östrogeeni taseme üleval, takistades nii uute
folliikulite moodustumise (östrogeen) ja hoiab all immuunreaktsiooni
(progesteroon). Progesteroon on oluline ka raseduse säilitamises,
ohjab emaka kontraktsioone ja soodustab piimanäärmete näärmerakkude
kasvu.
Östrogeen
inhibeerib folliikulite arengut.
Progesteroon
inhibeerib seda, et
sperm ei pääseks enam sisse- munaraku kest on
nii tugev.
Östrogeeni
tase tõuseb kuni ovulatsioonini, seejärel hakkab langema. Samal
ajal tõuseb progesterooni tase (peale ovulatsiooni) ja langeb alles
siis, kui
menstruatsioon algab.
3.
ViljastumineKehaväline
viljastumine
(
merisiilik ).
Viljastamine toimub merevees, nii
permid kui
munarakud on seal. Munarakk eraldab
peptiide, mida sperm ära tunneb (avab Ca kanali) ja hakkab liikuma
atraktantide toimel munaraku poole (
kemotaksis ); Spermi
välismembraanis olevad
retseptorid tunnevad ära glükoproteiine ja
spermid seonduvad
kallerkestaga
(kõige välimine kest)
munarakul .
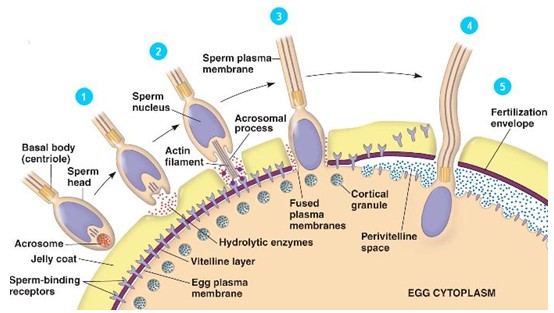
Järgneb
akrosomaalreaktsioon:
akrosoom moodustab akrosomaaljätke, mis tungib munaraku
seinani .
Akrosoomist moodustuvad ka aktiinifilamendid. Akrosomaaljätke peal
on
valgud (bindiin), mis seonduvad munaraku retseptoritele.
Vitelliinkest läheb katki ja spermi tuum tungib munarakku. Samal
ajal tekib munaraku pinnale viljastumiskoonus.
Spermi
sisenemisele järgneb
Ca sissevool .
Selle tagajärjel muutub
vitelliinkest
(rebukest)
viljastumiskestaks
ja teised spermid ei saa enam siseneda.
Spermi
sissetungimisel on meioosII arrest. Sperm siseneb otse, mitte lapiti
(nagu inimesel)
Pärast
tekitab munarakk enda peale veel paksu rebukesta.
Kuidas
tagatakse liigispetsiifilisus?
Kuidas
hoitakse ära polüspermia?
(membraanipotentsiaal, kortikaalreaktsioon)
- Kiire blokk – Na+ siseneb rakku põhjustades membraanipotentsiaali muutuse (-70 mV muutub positiivseks ), selle tulemusena membr depolariseerub. See kaitseb väga lühiajaliselt mitme spermi sisenemise eest.
- Aeglane blokk – Ca2+ sissevool kutsub esile kortikaalreaktsiooni (kortikaalgraanulid tühjendavad oma sisu rebukesta alla) ja vitelliinkest muutub permidele läbimatuks - tekib viljastumiskest.
Kehasisene viljastumine.
Spermid
paisatakse tupest ja emakast edasi munajuhasse
lihaste
kokkutõmmete
abiga. Munarakk ootab munajuhas
ampulla
piirkonnas
( munajuha kitsuses)- siin toimub viljastamine,
kus on umbes 2 kraadi soojem kui mujal munajuhas - spermid liiguvad termotaksise mõjul selles suunas. Kuid enne viljastamist peavad
spermid kinnituma munajuha seinale, et muutuda
viljastamisvõimeliseks, selleks toimub:
Kapatsitatsioon
– K iooonide väljumine põhjustab membr potentsiaali muutust;
albumiini mõjul võetakse spermi pinnalt kolesterool välja, mis
võimaldab Ca sissevoolu ja aluselisemaks muutumise – sperm muutub
viljastamisvõimeliseks.
Seejärel
hakkab sperm mööda temperatuurigradienti liikuma ja
kemotaksisena
mõjutab liikumist
progesteroon.
Munarakku
ümbritsevate barjääride läbimine
(
cumulus,
zona
pellucida,
akrosomaalreaktsioon, membraanide sulandumine).
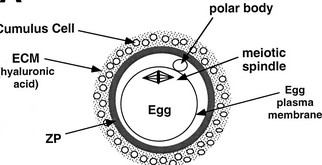
Cumulus’e
rakud ümbritsevad munarakku –
esmalt tuleb neist läbi pääseda.
Seejärel
tungib sperm läbi zona pellucida, kasutades selleks
akrosomaalreaktsiooni.
Peale
akrosomaalreaktsiooni seostub sperm akvaatortasapinnaga munarakuga
(siseneb lapiti) ja nende
membraanid sulanduvad- spermi tuum liigub
munarakku sisse. Mille järel Ca sissevool (aeglane blokk).
Polüspermia
vältimine
– ainult aeglane blokk! Ca sissevool põhjustab
kortikaalreaktsiooni, mis tekitab munaraku ümber viljastumiskesta
(zona pellucidast).
4.
Lõigustumine
Täpsemalt:
Kirjelda lõigustumist äädikakärbsel (sh. selgita, millise lõigustumistüübiga on tegu)
- Süntsüütium, süntsütiaalne ja rakuline blastoderm, energiidid, polaarrakud
Süntsüütium
– rakk ei jagune, kuid tuumad selle sees lõigustuvad – mood 1
rakk paljude tuumadega; 9.jagunemise stadiumini
//Vahetult
peale viljastamist on sügoodi jagunemine nii kiire, et tütarrakkude
vahele ei jõua membraani moodustuda ja tekib sünsüütsium. Pärast
9-ndat jagunemist sisaldab sünsüütsium 512 tuuma, mis seejärel
liiguvad embrüo äärealadesse ja jagunevad veel neli korda. Osa
rakke liigub veel spetsiaalselt embrüo tagumisele poolusele. Pärast
13-ndat jagunemist eralduvad rakud tsütoplasmaatiliste membraanidega
ning tekib rakuline blastoderm, kus on üle 4000 raku. Blastodermi
tagumisele poolele liikunud tuumadest moodustuvad polaarrakud,
mis on aluseks sugurakkudele.
Seega toimub somaatiliste ja sugurakkude liinide eristumine juba väga
varakult, blastula staadiumis .
Umbes
päevaga areneb äädikakärbse embrüost vastne . Vastne kasvab väga
kiiresti, vahetab 2 korda kesta ning moodustab viie päeva pärast
nuku. Nukustaadiumis (neli päeva) hävivad paljud
vastsespetsiifilised koed ning moodustuvad täiskasvanud kärbsele
iseloomulikud organid nagu tundlad , silmad, tiivad ja jalad. Kuna
täiskasvanud isendit nimetatakse valmikuks, imaagoks (ingl. k.
imago ),
nimetatakse vastse erinevates osades paiknevaid rakukogumikke,
millest arenevad organid, immaginaaldiskideks (imaginal
discs).//
Süntsütiaalne
blastoderm
on enne XIII lõigustumist; tuumade vahel pole membraani, ainult
munarakul on membraan; ühises tsütoplasmas jagunevad tuumad.
Rakuline
blastoderm
mood pärast XIII lõigustumist; siis munaraku plasmamembr voltub
sisse tuumade vahele ja jaotab iga tuuma üksikusse rakku.
Energiidid
– tsüntsütiaalse blastodermi tuumad, mida ümbritsevad
mikrotorukesed ja –filamendid.
Polaarrakud
- 9.jagunemisega liiguvad u 5 tuuma posterioorsesse ossa , nendest
saavad hiljem sugurakud .
Holoblastiline (täielik), mesoletsitaalne (keskm rebuhulk),
radiaalne
Kirjelda lõigustumist konnal (sh. selgita, millise lõigustumistüübiga on tegu)
- Hall sirp – dorsaalsel poolel, vastu spermi sisenemise piirkonda (sperm siseneb ventraalselt)
- kortikaalne rotatsioon – peale viljastamist kortikaalne tsütoplasma roteerub 30 kraadi sisemise tsütoplasma suhtes.
- Mikromeerid – animaalsel poolusel ekvatoriaalse (III)lõigustumise tagajärjel tekkivad väikesed rakud
- Makromeerid – vegetatiivsel poolusel ekvatoriaalse (III)lõigustumise tagajärjel tekkivad suured rakud
- Blastotsööl – õõnsus blastula sees, mis aitab vältida, et anim. ja veget. pooluse rakud omavahel liiga vara kokku ei puutuks. Võimaldab rakkude migratsiooni gastrulatsioonis.
- Moorula – 16-46 raku staadium
- Blastula – 128 raku staadium; eristunud on blastotsööl
- kesk-blastula transitsioon – aktiveeritakse embrüo enda genoom ; lisatakse G1 ja G2 faasid , sünkroonne rakujagunemine kaob.
Meroblastiline
(osaline), diskoidaalne e kettaline
Kirjelda lõigustumist kanal (sh. selgita, millise lõigustumistüübiga on tegu)
Lõigustumine
toimub ketta piirkonnas, kus rebu on vähem ja tsütoplasmat rohkem.
Rebuvaba osa lõigustumise tagajärjel kujuneb välja kettakujuline blastula e iduketas e blastodisk. Iduketta keskosa on rakuvälisest
rebust eraldatud subgerminaalse õõnega.
Kõigepealt
lõigustub pindmine osa – mood epiblast.
- Heleväli – area pellucida; iduketta tsoon, kust iduketas kumab subgermin õõne kohalt läbi
- Tumeväli – area opaca; piirkond, kus iduketas on tihedalt liibunud rakuvälisele rebule
- Blastoderm – rakkude kiht, mis on moodustunud munaraku ühele poolusele
- Epiblast – pealmine/ välimine iduketta osa, ühe rakukihi paksune; läbikumav
- subgerminaalne õõs – vaheruum rebu ja epiblasti vahel. ..tekib, kui blastodermi rakud absorbeerivad vett munavalgest, sekreteerides seda rebu ja endi vahele. Siia hakkab mood marginaaltsoon.
- marginaaltsoon – tume- ja helevälja piirialal.
Holoblastiline
(täielik), rotatsiooniline, isoletsitaalne
Kirjelda lõigustumist inimesel/hiirel (sh. selgita, millise lõigustumistüübiga on tegu)
- Kompakteerumine, moorula, blastotsüst, blastotsööl. Totipotentsus, pluripotentsus. (vih !)
- Trofoblasti ja ICM-sisemine rakumassi teke ja mis neist saab.
Trofoblast tekib blastotsüsti staadiumiks, asub blastotsööli ’ülaosas’.
Trofoblasti rakud aitavad embrüol emakaseinale kinnituda- lõhustavad
emaka kudesid ja poevad emakaseina sisse. Trofoblastist areneb
koorion .
ICM
tekib ka blastotsüsti ajaks. Need on pluripotentsed - võivad areneda
kõigiks rakkudeks, v.a. trofoblastideks. Neist
saab hiljem:
epiblast – embrüonaalne epiblast (embr endoderm ,
ektoderm
ja mesoderm )
- Blastotsüsti koorumine ja implantatsioon. Identsete kaksikute teke (moorula järgus jaguneb embrüo kaheks) +vih
5. Gastrulatsioon
Kirjelda
linnu gastrulatsiooni
- Hüpoblasti formeerumine (primaarne ja sekundaarne hüpoblast, hüpoblasti funktsioon).
Primaarne
hüpoblast
saab alguse rakkude delaminasiooniga epiblastist. Subgerminaalsesse
vaheruumi tekivad rakkude kogumikud/ saarekesed.
Sekund
hüpoblast
mood siis, kui Kolleri sirbi piirkonnast hakkavad rakud ettepoole liikuma.
Kogu
hüpoblast
areneb selle tulemusena, et sekund hüpoblast liigub järjest
ettepoole, haarates kaasa ka rakukogumikud (prim hüpoblast).
Funktsioon:
migreeruvatele
epiblasti rakkudele signaali-allikaks; ei anna ühtegi rakku
arenevasse embrüosse, kuid on eelduseks kogu edasisele arengule (kui
see eemaldada, siis arengut ei toimu).
- Ürgjuti teke ja funktsioon – tekib epiblasti paksendina Kolleri sirbi ees, mis venib ettepoole pikaks kitsaks jutiks. Fn: tähistab organismi anterio -posterioorset telge.
Kolleri
sirp
– määrab organismi anterio-posterioorse telje; see on helevälja
aposterioorses osas olev epiblasti rakkude ja MZ paksenenud osa.
Henseni sõlm
– ürgjuti laienenud eesmine osa; (asub ürgjuti kõige
peapoolsemas otsas)
Ürgjutt
(pea, keskosa, sabaosa – millised struktuurid tekivad läbi nende piirkondade):
Peaosa – Henseni sõlm: kordamesoderm, somiitide mesodermi mediaalne osa
Keskosa:
lateraalne osa somiitidest, süda, neerud
Sabaosa:
külgplaadi mesoderm, ekstra -embrüonaalne mesoderm
Ürgvagu
– ürgjuti keskele tekkiv süvend, kui epiblasti rakud hakkavad
laskuma epiblasti ja hüpoblasti vahele blastotsööli. // epiblasti
rakud migreeruvad ürgjutist embrüo sisse. //
mesodermi-endodermi
rakkude teke epiblastist
Germinatiivsirp.
Definitiivse
endodermi järk-järguline paigaldumine surub hüpoblasti koos selles
olevate esmaste sugurakkudega iduketta helevälja eesmisele piirile,
kus moodustuv volt on tuntud germinatiivsirbi
nime
all. // Hüpoblasti rakud koonduvad germinaalsirbi piirkonda, area
pellucida anterioorsesse ossa, sisaldab ka ürgseid sugurakke
(idurakke).
Ekstra-embrüonaalsed
koed: amnion e vesikest ; koorion; rebukott ; allantois e kusekott .
Kirjelda imetaja gastrulatsiooni
( hiir ja inimene) ALATES SLAID 27
- Inimene: epiblasti/hüpoblasti formeerumine. Tsütotrofoblast, süntsiotrofoblast. Platsenta teke, ehitus ja funktsioon. Ekstra-embrüonaalsed koed ( lootekestad ) - koorion, amnion, rebukott, allantois. Ürgjutt, ürgsõlm, endo - ja mesodermi ingressioon läbi ürgjuti, epiteliaal-mesenhümaalne üleminek (EMT). Anterioorse (eesmise) – posterioorse (tagumise) kehatelje kujunemine ( hiire näitel): DVE/AVE vs. ürgjutt. Vasak-parem kehatelje kujunemine ja sõlme roll selles. ---
Platsenta
moodustub 6-7 päeval pärast viljastamist; kõldkest ja emaka
limaskest kasvavad kokku. Seda läbivad veresooned , lisaks eritab platsenta hormoone, östrogeeni ja progesterooni, mis takistavad uute
munarakkude küpsemist ja tagavad raseduse normaalse arengu.
7.
Endoderm (täisk organismis on ülesandeks O2 omistamine ja
toitainete seedimine, imend)
Primitiivne endoderm – ekstraembrüonaalne kude, mis ümbritseb epiblasti.
Parietaalne endoderm – interakteerub trofoblasti gigant rakkudega (peale implanteerumist prim endodermist)
-migreeruvad
mööda trofektodermi sisemist pinda ning mood koos trofoblasti
gigant rakkudega parietaalse rebukoti.
Vistseraalne endoderm (k.a DVE ja AVE roll embrüo arengus) – moodustab blastotsööli sisevooderduse ning epiteliaalse koe epiblasti ja ekstraembrüonaalse ektodermi pinnale. (mood peale implanteerumist prim endodermist)
Definitiivne endoderm – embrüo kõige sisemine looteleht; pärineb epiblastist
Parietaalne
ja vistseraalne endoderm vahendavad toitainete ja jääkide vahetust
arenevas embrüos.
Vists
endoderm on oluline signalisatsioonikeskus embrüo edasises arengus.
-jaguneb
: embrüo distaalses osas distaalseks
vistseraalseks endodermis
(DVE)
millele
järgneb DVE rakkude ümberpaiknemine anterioorsesse poolde ja
formeerumine
anterioorseks
vistseraalseks endodermis
(AVE)
AVE
on embrüo signaliseerimise keskus, mis on vajalik
anterioorse-posterioorse telje moodustamiseks ja embrüo
anterioorsete struktuuride kujunemiseks; ekspresseerib mitmeid
antagoniste.
Endoderm
on vajalik ürgsoole tekkeks, millest areneb 2 torujat organsüsteemi: seede - ja hingamiselundkond .
- Ürgsool (ees-, kesk- ja tagasool; oraalplaat, Rathke tasku )
Inimese
näitel: esmalt tekib ürgsool, mis seejärel spetsialiseerub ees-,
kesk- ja tagasoole piirkonnaks . Kesksool on ühenduses rebukotiga.
Eessoole eesmine osa on blokeeritud ektodermaalse plaadiga –
oraalplaadiga
ehk stomodeumiga (suuava tekkeks see lõheneb)
Oraalplaadi
ektoderm on otseses kontaktis neutraalse ektodermiga (aju ektoderm)
ning need kaks ektodermi interakteeruvad, moodustades Rathe
tasku
– areneb hüpofüüsi näärmeline osa (selle neutraalne osa areneb
vaheajust)
- Neel (neelutaskud ja mis neist areneb)
Neel
järgneb suuõõnele. See kujuneb ekto-, meso - ja endodermist,
moodustades neelukaared ja – taskud .
Neelutaskud
– neelu endodermaalne osa,
imetajatel on 4 paari neelutaskuid. Neist areneb:
paar – keskkõrva kuulmekanalid/ õõnsused, kuulmetõri
paar – mandlite areng; paari ühinemisel mood allapoole kilpnääre
paar – tüümus ja 1.paar kõrvalkilpnäärmeid
paar – 2.paar kõralkilpnäärmeid
- Seedekulgla kujuneb endo- ja mesodermi vastastikusele signalisatsioonile. Soole endoderm ekspresseerib esmalt Shh-d, mis spetsialiseerib ümbritsevat mesodermi ekspresseerima Hox-e. Mesodermist tulenevad signaalid omakorda määratlevad erin soole osade piirid. Piiride määratlemisele järgneb soole ja tema derivaatide diferentseerumine .
Endoderm
mood seedekulgla sisevoodri.
Splanhilisest
mesodermist pärinevad soolte ümber paiknevad lihased.
Sool
lõpeb esialgu endodermaalses kloaagis, mis jaguneb hiljem põieks ja
rektaalseks piirkonnaks. Sool ühineb rektaalse osaga, tekib
sopistis, mis puutub kokku pinnaektodermiga, mida eraldab membraan
(kui see purunub, siis mood anus).
- Maksa ja sapipõie ning pankrease areng
*
Eessoole piirkond spetsialiseerub hepaatiliseks
endodermis,
millest omakorda pärinevad hepatoblastid
(eellasrakud, bi-potentsiaalsed) mood väljakasvu – maksapunga.
Maksapunga epiteel hakkab harunema vastusena mesenhüümist
tulenevatele signaalidele.
Eessoolest
kaugemal asuvad rakud kujunevad hepatotsüütideks (maksarakud)
*Eessoolelähedased
hepatoblastid kujunevad sapijuha / põie epiteeliks ja nendest areneb
sapisüsteem.
*Mesenhüüm
indutseerib kahe pankrease punga kujunemist ( dors ja ventr pung ).
Ventraalse punga arengut mõjutavad südamest pärit signaalid ja
dorsaalset nt seljakeelikust ja dorsaalaordist tulevad signaalid ning
lisaks vajab see retinoolhappe olemasolu. Lõpuks kasvavad dorsaalne ja ventraalne osa kokku. Inimesel säilib vaid ventraalne pankrease juha , mis viib seedeensüüme soolestikku.
- Kopsude areng – hingamiselundkond areneb samuti ürgsoolest vastusena südamest tulevale FGF signaalile. Esmalt tekib larüngotrahheaalne vagu , mis pikeneb ventraalselt.
Vagu
jaguneb kaheks haruks, millest arenevad kopsud
ja bronhid.
Oluline
on endodermaalse epiteeli ja mesenhüümi vastastikune
signalisatsioon.
(Endoderm
vooderdab hingetoru , bronhe ja kopsu alveoole.)
Primitiivsest
endodermist pärineb parietaalne ja vistseraalne endoderm ning neist
mood hiljem rebukott.
Vistseraalne
endoderm on oluline signalisatsiooni keskus arenevas embrüos.
Endoderm
on vajalik mitmete mesodermaalsete organite arengu induktsiooniks
ning ka ürgsoole tekkeks.
Ürgsoolest
areneb seede- ja hingamiselundkond
Endoderm
mood ninde elundkondade kõige sisemise epiteliaalse kihi.
8.
Jäseme areng
- Jäsemeväli kui morfogeenne väli
Jäseme
areng algab jäsemavälja kujunemisega- see on morfogeenne piirkond,
kuhu on koondunud rakud, mis on võimelised tervet jäset moodustama.
Kui see embrüost välja lõigata ja mujale asetada, siis peaks seal
arenema jäse. Ühest jäsemeväljast võib välja kasvada mitu
jäset. Hetkel, mil jäsemepung alles moodustub, on jäsemevälja
rakkudel võime moodustada ükskõik milline jäseme osa. Peale
jäsemevälja mood jäsemepung.
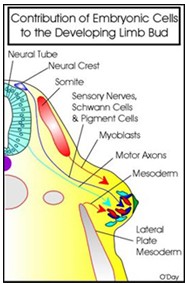
- Millised rakud on olulised jäsemepunga tekkeks?
Külgplaadi
mesodermi rakud (nendest tuleb signaal), Schwanni rakud, aksonid , somiidid (lihasrakud), pigmendirakud…
- Kuidas Hox perekonna geenid mõjutavad jäseme arengut?
Hox
geenid määravad anterio-posterioorsel teljel koha, kuhu jäse tekib
(Hox5, Hox6-esijäse, Hox10-tagujäse) ning määravad sõrmede asetuse ja lahknemise.
(Spetsifitseerib
mesenhüümi rakud kas stülopoodiks, seugopoodiks või autopoodiks.)
- Kuidas mõjutavad jäsemepunga teket ning jäseme arengut FGF8 ja FGF10?
FGF
valgud osalevad jäsemepunga initsiatsioonis.
FGF10
avaldub jäsemepunga mesenhüümis ja indutseerib ektodermis AERi
tekke.
FGF8
avaldub AERis ja hoiab AER-ga külgneva mesenhüümi jagunemas.
- Apikaalse-ektodermaalse valli (AER) ning selle all paikneva mesenhüümi roll
-areneva
jäseme peamine signaalikeskus JUHIB JÄSEME EDASIST ARENGUT ja
vastutab apoptoosi eest (vajalik sõrmede ja varvaste eraldumiseks)
-AER
tagab proksimaalse- distaalse (õlg-sõrmed) jäseme kasvu, hoides mesenhüümi rakke mitootilises prolifereerumisjärgus. Samal ajal
AER säilitab anterioorse-posterioorse (pöial-väike sõrm) telje
tekkeks vajalike faktorite ekspressiooni ja määratleb D-V
(käeselg-peopesa) teljed.
Retinoolhappe
ja FGF/Wnt
vastandlikud gradiendid:
Retinoolhapet
eritub proksimaalsel poolel, FGF ja Wnt distaalsel poolel. Kui
mesenhüümi töödelda retinoolhappega, siis sellisest jäsemepungast
arenevad proksimaalsed osad rohkem ja samuti FGF ja Wnt puhul
(arenevad distaalsed osad).
*Proksimaal-distaalse
telje kujunemine ja jäseme diferentseerumine sõltub
interaktsioonidest AERi ja selle all oleva jäsemepunga mesenhüümi
vahel. Selle distaalse mesenhüümi ehk mittediferentseerunud tsooni
ehk progresseeruva tsooni (PZ) prolifereeruv aktiivsus tagab
jäsemepunga proksimaal-distaalse pikenemise ning hoiab ära
kõhrestumise.
- Anterio-posterioorse (pöial-väike sõrm) telje areng ja peamised faktorid
Määravaks
faktoriks sonic
hedgehog
(Shh)- kutsub esile peegelpildi normaalsest olukorrast; vastutav A-P
telje kujunemise eest. (Seda piirkonda kutsutakse ZPA (polariseeriva
aktiivsusega tsoon))
Shh ekspressioon väljaspool ZPA’d põhjustab lisavarvaste teket.
Sõrmede
täpsustumine sõltub ajast, mil Shh ekspresseerub- selle
ekspressioon on autokriinne- see ekspresseeritakse rakku sisse. Kõige
rohkem ekspressiooni saab väike sõrm (see on aposterioorne), keskm
sõrmes on oluline Shh konts kui ka aeg, nimetissõrme identiteet pannakse paika ainult konts-ga, pöial on Shh-st täielikult
sõltumatu (kui shh ekspress puudub, siis mood ainult pöial)
- Dorso-ventraalse (käeselg-peopesa) telje areng ja peamised faktorid
Dorso-
ventraalse telje areng – määratakse ektodermi poolt. Dorso-ventraalse telje määramisel on eriti oluline Wnt7a
geen, mida ekspresseeritakse dorsaalses jäsemepunga ektodermis.
Dorsaalses mesenhüümis aktiveerib ta Lim1 geeni, mis
spetsifitseerib dorsaalsete rakkude diferentseerumise.
Kui
Wnt7a eemaldada, siis see põhjustab jäsemes ventraalsete
struktuuride tekke dorsaalsele poolele ehk dorsaalne pool saavutab
ventraalse identiteedi.
Lim1
funktsiooni vähenemine/kadumine inimesel põhjustab küüs-patella
sündroomi (küüs pole nii arenenud ja patella on väiksem).
- Apoptoosi roll autopoodi kujunemisel
Selleks,
et autopood välja kujuneks , on oluline apoptoos - õigel ajal peavad
teatud rakud tulevaste sõrmede vahelt kaduma/ surema, et tekiksid
normaalsed sõrmed.
Signaal
autopoodis toimuva apoptoosi jaoks tuleb BMP
valkudelt.
BMPdel on kaks rolli: nad algatavad mesenhüümi rakkude apoptoosi
või kondrotsüütide diferentseerumist.
BMP
on oluline sõrmevahelises ruumis, et sõrmed saavutaksid mingi
identiteedi; kuna alguses on sõrmede vahel lestmed.
Shh
põhjustab Gli3 ekspressiooni, mis on pidurdavaks faktoriks valgule
Gremlin, mis omakorda pidurdab BMPd . Kui Gli3 on puudu, siis Shh-l ei
ole rolli, kuid Gli3 ei pidurda siis Gremlini ja BMP tase on kõrge –
seetõttu mood palju ühesuguseid sõrmi.
Kui
BMPd on palju , siis see soodustab apoptoosi. Gremlin pidurdab
apoptoosi- lestad jäävad alles.
9.
Mesoderm
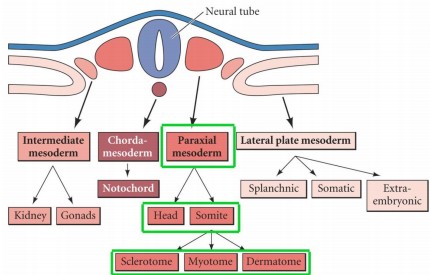
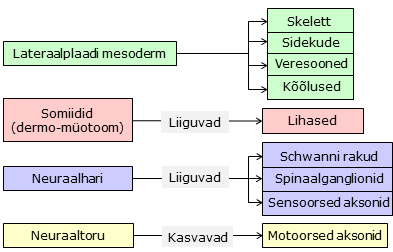
- Nimetage mesodermi liigid ning põhilised koe- ja organialged, mis nendest arenevad
Sklerotoom – selgroolülid, lülidevahelised diskid ja roided
Müotoom
– selja, roietevahelised ja keha eesmised lihased
Dermamüotoom
– skeletilihaste eellased ja selja dermise rakud
- Kordamesoderm ja tema roll selgroogsete arengus
Kordamesoderm
mood korda ehk seljakeeliku, mis paikneb alati neuraaltoru all ja
pikeneb kõhtmiselt.
Kõrgematel
selgroogsetel esineb see embrüonaalses staadiumis ja on embrüos
tugiorganiks.
Kui embrüol hakkab mood. selgroog, jääb seljakeelik selgroo vahediskide koosseisu.
Süstikkaladel
esineb seljakeelik kogu elu.
- Mis on paraksiaalne mesoderm?
..on
kahel
pool neuraaltoru paiknev mesoderm, mis on segmenteerunud (somiidid) /segm-mata
(posterioorne osa). Segmenteerumata mesoderm on mesenhümaalne. Kui
on moodustunud lõhe, siis välimised mesenhümaalsed rakud
epitelialiseeruvad ning moodustub epiteliaalne somiit – toimub
mesenhüüm epiteel transitsioon (EMT)
-jaguneb
pea mesodermiks ja somiitideks- nendest omakorda palju massi.
-somiitide
areng toimub eest tahapoole
-segmenteerumine
kulgeb anterior -posterior teljel
*Notch
signaalirajas olev hairy1 osaleb uue somiidi moodustumises, mis
ekspress lainetena.
-Somiidi
epitelialiseerumine: Uue somiidi eraldumisele eelneb lõhe
moodustumine. Lõhe asukoha määravad ephriin retseptor EphA4
(somiidi anterioorses osas) ja ligand – ephriin B2 (aposterioorses
osas). Nende valkude toimel toimub rakk-rakk eemaletõukumine.
-Somiitide
tekkel on oluline, et mesenhüüm muutuks epiteeliks.
-Somiidi
spetsialiseerumine: Somiitide saatus on tugevalt determineeritud
juba enne, kui nad on tõelisteks somiitideks arenenud. Anterioorsed
somiidid on määratud arenema nt kaelalülideks ja posterioorsed
nimmelülideks. Sellise spetsialiseerumise eest vastutavad Hox
geenid.
-Somiidi
diferentseerumine:
sklerotoom-
skeletielemendid; müotoom-
lihased; dermatoom
-
nahk, osad lihased
Selgroolüli
mood kahe kõrvutioleva somiidi
poolsegmendi ühinemisel.
Eesmise
somiidi tagumine osa+tagumise somiidi eesmine osa=selgroolüli. Seda
nim resegmenteerumiseks – vanad segmendid kaovad ja tekib uus
struktuur. Arvatakse et kõhreline kude diskide vahel on jäänuk
seljakeelikust.
- Skeletiluude erinevad eellased
-somiidid,
millest saadakse selgroog
-lateraalplaat,
millest saadakse jäsemete skelett
-kraniaalne neuraalhari
– näokolju luud ja kõhred
- Kehalihaste päritolu ja üldine areng
Kõik skeletilihased pärinevad ( derma )müotoomist
va pea lihased.
► Dermamüotoomi
alla moodustub müotoom, millest toimub müoblastide vabanemine .
Müoblastid jagunevad kiiresti, millest enamik migreerub keha
osadesse laiali.
**
Müoblastid koonduvad – Müoblastid joonduvad – Rakud fuseeruvad
– Moodustub müotuubul ja toimub diferentseerumine – Müotuubul
moodustab ühtse lihaskiu – Lihaskiud koonduvad lihaskiudude
kimpudeks.
► Lihased
kujunevad kahest müotoomi populatsioonist:epaksiaalsest ja
hüpaksiaalsest müotoomist
- Urogenitaalsüsteemi päritolu ja neerutüüpide vaheldumine embrüogeneesis
Arenevad
vahelmisest
mesodermist
(paraksiaalsest).
Pronefros
on funktsionaalne neer kaladel ja amfiibidel, kõrgematel loomadel
degenereerub pronefors varsti peale moodustumist. Neeru juha
anterioorne osa ning pronefrilised torukesed taandarenevad, juha
kaudaalne osa säilub ning annab aluse Wolffi ehk nefrilisele juhale.
Wolffi
juha indutseerib pronefrosest kaudaalselt uute torukeste moodustumise
juha kõrval paiknevas mesenhüümis. Need torukesed diferentseeruvad
nefroniteks ja nõnda tekib mesonefros
ehk keskneer. Uute torukeste moodustumisega kaudaalses osas toimub
samal ajal anterioorses osas nende lagundamine. Osad mesonefrilised
tuubulid säiluvad
ning
osalevad hiljem isaste suguteede arengus.
Nefrilise
juha jõudmisel kaudaalsesse piirkonda – metanefrilisse mesenhüümi
–indutseeritakse kusejuha punga teke, millega algab metanefrose
areng. Metanefros on amniootide pärisneer.
Nefriline
juha suubub kloaaki (imetajatel embrüonaalne struktuur)
- Mis on kardiogeenne mesoderm?
Tekib tavalisest mesodermist; diferentseerumise tagavad südamesirbi
piirkonna endodermist lähtuvad signaalid (kude kui induktor ).
(Sellest tekib torujas organ süda)
Kui
endoderm eraldada, siis dif ei toimu.
- Missugused rakuliigid diferentseeruvad kardiogeensetest eellasrakkudest?
Endokardi
endoteelist endokardiaalse padjandi rakud, kodade müotsüüdid ja
vatsakeste müotsüüdid ning Purkinje rakud.
- Südame neuraalhari ja tema derivaadid
Paikneb
kõrvaplakoodi ja kolmanda somiidi vahelisel alal.
Sellest
mood veresooned, arterite silelihaskestad; harkelundi,
kõrvalkilpnäärme ja kilpnäärme sidekude ning vatsakeste ja
suurte veresoonte vaheseinad . Osalevad poolkuuklappide moodustumisel
ja tagavad erutusjuhtesüsteemi arengu.
Poolkuuklapid;
kojavatsakeseklapid; kambritevahelised vaheseinad.
- Nimeta südame morfogeneesi põhi etapid
1.
Kardiogeense mesodermi välja kujunemine
2.
Külgplaatide kardiogeense mesodermi lõhenemine (M-E)
3.
Endokardiaalse alge tekkimine
4.
Vasaku ja parema südamepoole liitumine toruks
5.
Südametoru lingustumine
6.
Endokardiaalpadjandi tekkimine (E-M) neuraalharja ja endokardi
rakkudest
7. Klappide ja vaheseinte teke ja areng
8.
Erutusjuhtesüsteemi areng
9.
Neljakambrilise südame areng
Tekib
siis, kui vasak ja parem südamepoole liituvad/sulanduvad AIP
piirkonnas toruks.
Kui liitumist takistada, tekib 2 funktsioneerivat südant (cardia bifida)
Süda
töötab ja on tekkinud veresoonestik. On olemas rebukott, aga see ei
toida ära.
Embrüo
saab ema käest O2 kätte, sest embrüo hemoglobiinil on suurem
affiinsus O2 suhtes.
Rebukoti
veresooned on vajalikud toitainete ja gaasi vahetuseks.
- Missuguseks muutub südameosade järjekord lingustumisel piki välimist ja sisemist südamekõverust?
Sisemisel
kõverusel toimuvad aga kvalitatiivsed ümberkorraldused: OFT – VK
ja PK—VV
Piki
välimist kõverust säilib esialgne kambrite järjestus: OFT
-PV-VV-VK (25)
- Missugused südame arengudefektid on iseloomulikud Fallot ’ tetraloogiale?
Falloti
tetraloogia- 4 defekti on koos: kopsutüve kitsenemine; aorti pääseb venoosne veri ( vahesein on defektne); parema vatsakese hüpertroofia
(suurem kui vasak)
- Millised suured veresooned tekivad lõpusekaare arteritest?
Südamest
väljuvad suured veresooned. Erinevad
unearterid; alanev ja ülenev aort, rangluuarterid
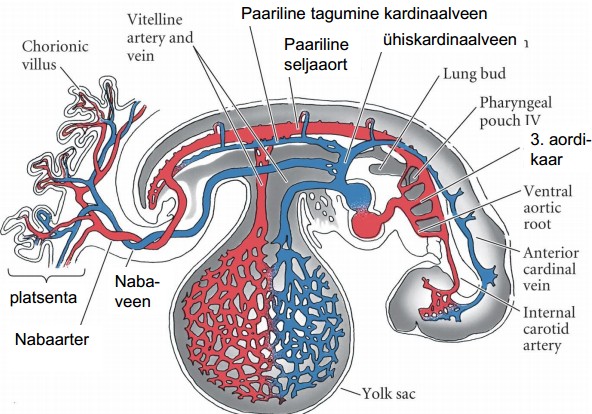
- Vaskulogeneesi üldiseloomustus
(veresoonkonna
areng) Veresoonte moodustumine de novo külgplaadi mesodermist.
*Protsess,
mille käigus moodustuvad esmased veresooned. Kõigepealt
kondenseeruvad hemangioblastid veresaarekesteks, mille sisemised
rakud kujunevad vererakkude eellasteks. Välimised rakud kujunevad
angioblastideks ehk veresoonte eellasteks, mis omakorda dif
endoteelirakkudeks. Need endoteelirakud mood aga torusid , mis
ühinevad kapillaarvõrgustikuks.
- Angiogeneesi üldiseloomustus
On
protsess, mille käigus kujundatakse ümber vaskulogeneesis
moodustunud veresooned ning kujuneb uus veresoonte võrgustik
organite ümber.Tekivad peened kapillaarid ning eristuvad arterid ja veenid . Uued veresooned mood olemasolevatest harude väljakasvamisel
või nende lõhenemisel.
Vererakkude
ja endoteeli rakkude ühine eellane.
- Hematopoeesi toimumise kohad
Hematopoeesi
eelduseks on pluripotentsete tüvirakkude olemasolu.
Imetajate
embrüonaalne vereloome algab rebukoti
veresaarekestes
(looteväline vereloome) või aordi -gonaadi-mesonefrose
piirkonnas
(lootesisene vereloome).
Hiljem
liiguvad tüvirakud mõlemast piirkonnast loote
maksa
ning seejärel luuüdisse,
lümfisõlmedesse,
põrna
ja harknäärmesse
(e. tüümusesse), kus nad jäävad vererakkude allikaks organismi
edasise elu jooksul.
Täiskasvanud
organismis toimub vereloome luuüdis
ja põrnas,
harvem ka maksas ,
neerudes,
neerupealistes
ja rasvkoes.
- Iseloomustage hematopoeetilist tüvirakku
See
on pluripotentne ; võimeline diferentseeruma nii vere- kui ka
lümfirakuks.
Tüviraku
jagunemisel tekib üks diferentseeruma spetsialiseerunud rakk ning
üks tüvirakk, mis on võimeline ennast taastootma.
Hematop
tüvirakud on eellasteks punaverelibledele (erütrotsüütidele),
valgetele verelibledele (granulotsüüdid, neutrofiilid,
vereliistakud) ja lümfotsüütidele.
10.
Ektoderm
- Pinnaektoderm (epidermis e nahk, juuksed, küüned, suuepiteel,
lääts),
- neuraalhari ( perifeerne NS, melanotsüüdid, hamba dentiin)
- neuraaltoru ( peaaju ja seljaaju ehk kesknärvisüsteem, motoorsed neuronid )
- Kesknärvisüsteemi varajane areng ( induktsioon , neuraaltoru teke ja selle anterio-posterioorne ja dorso -ventraalne polariseerumine )
Gastrulatsiooni
tulemusena tekivad 3 lootelehte; rakud pole enam totipotentsed
–ainult teatud rakud ja piirkond on võimeline arenema KNS-ks:
(induktsioon)
blastopoor hakkab ülespoole liikuma ja ’’ütleb’’
dorsaalsele ektodermile, et see peab arenema KNS-ks; seejärel mood
neuraalplaat - dorsaalne ektoderm pakseneb- ja sellest omakorda
neuraaltoru.
Neuraaltoru
teke:
-
dorsaalne
ektoderm pakseneb (mood neuraalplaat).
-
neuraalplaat hakkab sisse voltuma, mood neuraalvagu. (+neuraalvallid)
-
neuraalvallid
saavad kokku ja mood epidermise ning neuraaltoru mood täpselt selle
alla. Tulevase epidermise ja neuraaltoru vahele jääb neuraalhari.
(neuraalvagu sulgub neuraaltoruks)
Notochord-
sellest areneb seljakeelik;
annab signaale, embrüo arenguks vajalik.
ANTERIO-POSTERIOORSE
TELJE KUJUNEMINE:
*Hox-
geenid
– on erin geenisegmentides
*Kesk- tagaaju piirkond on tugev signalisatsiooni tsenter .
*Eesaju
mood otsaju ja vaheaju (sellest areneb silma võrkkest)
*vaheajust
(Diencephalon)
punguvad välja silma vesiikulid.
DORSO-VENTRAALSE
TELJE KUJUNEMINE:
Seljaaju
selgmise identiteedi määrab ära epidermis
ja kõhtmise notochord e seljakeelik.
Seljakeelik
ütleb, et arenema peavad motoneuronid.
- Kust pärinevad kesknärvisüsteemi rakud (kas tekivad kindlates piirkondades, kus? kuidas jõuavad kesknärvisüsteemi rakud oma lõppasukohta?)
Kõik
KNSi rakud tulevad neuraaltorust.
Kui paljunemine lõpeb, siis nad migreeruvad vastavasse paika.
Ventrikulaartsooni
piirkonnas on neuronid, neuroepiteel.
Marginaaltsoonis
on neuronite jätked.
*
Lõppasukohta
migreeruvad nt mööda teisi rakke. (rakud migreeruvad neuraaltorust
väljapoole)
- Neuronite arvu regulatsioonitasemed
-
lateraalne inhibitsioon neuroepiteelis - kõrvutiolevad rakud ei
arene reeglina sama tüüpi rakuks. Kui üks rakk areneb neuroni
suunas, siis tema kõrval olev rakk areneb tugirakuks
-
rakutsükli pikkus – mida noorem on embrüo, seda kiiremini rakud
jagunevad. Vanema embrüo puhul on G1 ja S faas pikemad .
-
sihtmärkrakult tulevad signaalid, parakriinsed faktorid gliia ja
neuronite kehadest, teistelt neuronitelt tulevad neuraalsed signaalid
- Ektodermaalsed plakoodid – teatud piirkonnad, mis on võimelised alust andma neuronitele. Lens - läätseplakood - sellest areneb välja silmalääts, ei anna neuroneid .
Osalevad
meeleelundite arengus (silm, sisekõrv)
Neuraalhari
ei teki neuraaltorust ega ka epidermisest, vaid sealsest piirpinnast.
Inimeses hakkavad neuraalharja rakud migreeruma peale toru sulgumist,
hiires enne sulgumist.
Neuraalharja
rakkudest saavad neuronid, Schwanni rakud, pigmendirakud e
melanotsüüdid ja kromatiini rakud (toodavad adrenaliini)
- Mida kujutab endast perifeerne närvisüsteem ja millised arengulised struktuurid annavad sellele aluse
… ehk
piirdenärvisüsteem koosneb ülejäänud närvidest ja närvirakkude
kehade kogumikest (ganglionitest) väljaspool kesknärvisüsteemi.
Sellele annavad aluse:
- Perifeersed aksonid (neuroni kehad on KNS-s)
- Neuraalhari
- Ektodermaalsed plakoodid
- Mis struktuur annab aluse sisekõrvale ja selle ganglionile? (sh ganglioni mõiste)
Ganglion -
närvirakkude sõlmjas kogum. Nad koosnevad närvirakkude kehadest ja
on ümbritsetud sidekoelise kihnuga.
Sisekõrv
on ektodermaalset päritolu. Aluse annavad poolringkanalid, tigu (?)
- Silma areng (induktsioonid, võrkkesta ja läätse päritolu)
Vaheajust
pungub välja silmavesiikul, mis indutseerib pinnaektodermist läätse
plakoodi tekke. Seejärel silmavesiikul sopistub uuesti sisse.
Lääts
tekib pinnaektodermist
Võrkkest
neuraaltorust.
Silma
arenguks on vaja transkr faktorit Pax6. Kui nt Shh inhibeerib seda,
siis silma ei teki. Samuti ei teki silma siis, kui Shh-d on liiga
palju ning kui seda on vähe, siis võib tekkida 1 silm. Järelikult
peab Shh hulk olema täpselt õige.
11. Metamorfoos , regeneratsioon , vananemine
- Metamorfoos (mis on, milleks vaja, kellel vaja)?
Metamorfoos-
arengu
hormonaalne taasaktiveerimine (reguleeritakse
kilpnäärme poolt)
Elustiili
muutuseks on vajalik ka morfoloogiline muutus: see lubab nt
amfiibidel minna üle veeliselt eluviisilt maismaalisele
-Vastse
vormil on teine ülesanne kui valmikul (liblika rööviku ül on
toitumine ja kasvamine, valmiku ül aga paljunemine)
Need
on kilpnäärmehormoonid (türoksiin T4 ja tri-iodotüroniin T3);
annavad 4 võimalust:
organi
kasv, surm, remodelleerimine või respetsifikatsioon.
Metamorfoosil
on oluline rakkude surm – apoptoos,
mida indutseerib T3 hormoon – lõpuste ja saba rakkude lagunemine .
Remodelleerimise
käigus saadakse kullese pikast sooltorust lühem – kudede
ümberkorraldamisel ei tekitata uusi rakke juurde, vaid olemasolevaid
paigutatakse ümber. Keele arenguks kasvatatakse uus lihas, mida
kullese staadiumis pole. Kullese kolju on kõhreline ja neuraalharja
päritolu. Täisk kolju on neuraalharja päritolu luukude.
Respetsifikatsioon:
Metamorfoosil maks sünteesib ensüüme, mis on vajalikud uurea
tootmiseks (vee kokkuhoid ja maismaaline eluviis). T3 aktiveerib
täisk maksa geenid ja represseerib samal ajal kullese maksa geenid.
Lisaks põhjustabT3 kullese hemoglobiiniga rakkude surma ja
proliferatsiooni rakkudes, kus on täisk hemoglobiin .
Kui
ei toodeta thyrotropin’i ja T4 sünteesi ei aktiveerita, siis
säilib kullese keha, kuid organism on suguküps (neoteenia).
- Amfiibide metamorfoosi muutused, näited ja põhjused
kulles- veeline eluviis, sabauim (lagund osaleb T3) täiskasvanu-
4 jalga
hemoglobiin
teistsugune -’’-
herbivoorne
toitumine, pikk sooltoru karnivoorne toitum, lühike sooltoru
eritavad
ammoniaaki eritavad uureat
limanäärmed
puuduvad limanäärmed
silmad
külgedel - monokulaarne nägemine silmad otse – binokulaarne
nägemine (ühest silmast liiguvad aksonid vastasajupoolkerale) (mõlemast silmast väljuvad aksonid)
kolju
neuraalharja päritolu kõhr kolju neur.harja päritolu luukude
- Putukate metamorfoosi tüübid, näited
Vastse
kudede kaotamine ja nendest tehakse täiesti uued rakupopulatsioonid.
*
Ametaboolne
metamorfoos
– kasv toimub ainult läbi kestumiste, munast koorub peaaegu täisk
isendi sarnane isend . (hooghännalised)
*
Hemimetaboolne
metamorfoos
– munast koorub pronümf, kellest mõne kestumise tagajärjel saab
nümf, kel kõik kehaosad olemas, kuid mitte täielikult arenenud
( rohutirtsud )
*
Holometaboolne
metamorfoos
–täielik metamorfoos, munast koorub vastne, kes ei näe üldse
valmiku moodi välja. Vastne – nukk (ei toitu, valmistatakse
valmiku keha struktuurid)– valmik ( liblikad )
- Imaginaaldiskid- põhiliseks elemendiks, mis on olemas juba vastsel. Uue valmiku organid arenevad nendest diskidest. Enamus vastse keha lagundatakse apoptoosis.
Uued
valmiku organid arenevad mittediferentseerunud imaginaalrakkudest,
mis paiknevad vastsetes kobaratena ja ootavad diferentseerumiseks
signaali.
Erinevad
struktuurid omavad erin imaginaaldiske. Neid on 3 tüüpi:
imaginaaldiski rakud, mis mood vamiku kutikulaarse struktuuri (tiivad, jäsemed, pea, tundlad, rinna, suguelundid )
Histoblastid – valmistavad putuka tagakeha; imaginaalrakkude kobarad
Imaginaalrakkude kogumikud – mood valmiku organid pärast vastse osade lagunemist
- Putukate metamorfoosi hormoonid ja kuidas toimivad ?
Steroid 20-hydroxyecdysone (20E) – koordineerib kestumist . Esimene 20E laine põhjustab nukkumist, peatab rakkude jagunemise diskis ja muudab rakkude kuju. Stimuleerib epidermaalseid rakke, et need sünteesivad omakorda ensüüme, mis lagundavad vana kutiikuli ja sünteesivad uue.
Kui
20E puudub, siis metamorfoosi üldse ei minda.
Lipiid juveniil-hormoon (JH) - tagab pärast kestumist vastse fenotüübi, mitte nuku või valmiku tekkimist (kui on vaja mitu kestumist). (on oluline vastse staadiumis, takistab seda, et loom läheks metamorfoosi)
Enne
nukkumist JH tase langeb ja 20E laine põhjustab nuku tekkimise.
Regeneratsioon
– arengumehhanismide käivitamine post-embrüonaalses eas, et
taastada puuduv või kahjustunud kude.
Tüviraku vahendatud regeneratsioon – tüvirakke kasut kaotatud koe taastamiseks (vererakkude asendamine, karvakasv, nahk- tüvirakud jagunevad ja uuenevad pidevalt, lameussid)
Kui lameuss keskelt pooleks lõigata, siis sabaosa taastab peaosa ja
vastupidi ja kui nii saba- kui ka peaosa puuduvad, siis
regenereeritakse mõlemad õigetesse kohtadesse . Klonogeensed
neoblastid
– migreerumisvõimelised ja pluripotentsed.
Epimorfoos – olemasoleva koe rakud dediferentseeritakse ( kaotavad oma ülesande organismis)- muutuvad tüvirakulaadseks (mittediferentseerunud massiks) ja pärast signaalide najal kasvatavad seda, mida vaja ehk spetsialiseeruvad uuesti ( salamander )
Jäseme eemaldamisel kasvatab tagasi puuduoleva osa. Kõigepealt tekib
plasmataoline hüüve. Haava otsa teib apikaalne ektodermaalne ots,
edasi mittediferentseerunud rakkude mass – regeneratsiooni
blasteem
– need paljunevad ja redif uuteks struktuurideks. Rakud mäletavad,
mis tüüpi nad enne olid.
Morfallaksis – olemasolevate kudede ümber paigutamine (hüdra- tükeldades saadakse mitu hüdrat)
Ehk
rakud remodelleerivad ennast. Rakud on pidevalt mitoosis ning
tekkivad rakud paigutuvad distaalselt. Igal rakul on mitu rolli,
sõltuvalt vanusest . Hüdra tükeldamisel kasvatab iga tükk
apikaalsesse otsa pea ja basaalsesse otsa saba- tekib väiksem hüdra.
Pea aktivatsiooni gradient on kõrgeim tipus ja jala aktivatsiooni
gradient basaalses osas.
*Hüpostoom
– põhiorganisaator peaosas ’tundlate’ vahel, inhibitoorne
gradient väheneb basaalse osa suunas; et ei tekiks uut
organiseerivat organit. Kuidas hüdra teab, millal punguda?- pungumine saab toimuda seal, kus signaalide gradiendid on võimalikult
väikesed ehk siis kui hüdra on saavutanud suurema kasvu.
4)
Kompenseeriv
regeneratsioon
– diferentseerunud rakud jagunevad, säilitavad oma fn’i. Uued
rakud ei tulene tüvirakkudest ega läbi dediferentseerumise.
(imetaja maks)
Dif-nud
rakud paljunevad, et taastada vigastatud organi struktuur ja fn. (nt
maks kompenseerib puuduoleva sagara- uut sagarat ei kasvatata, vaid
suurendatakse teisi sagaraid) kõik rakutüübid maksas suurendavad
oma hulka. HGF – hepatotsüütide kasvu suurendamine.
- Vananemine ja mis seda põhjustab?
-
elamiseks ja paljunemiseks vajalike füsioloogiliste fn-de ajast
sõltuv mandumine/ allakäik. Põhjustavad:
mutatsioonid
(kiirgus, hapniku radikaalid), keskkonna
faktorid,
juhuslikud
epigeneetilised muutused (sobimatu
metülatsioon)
Vead
DNA reparatsioonil; kulumine (punktmutatsioonide arv kasvab vanusega,
sellega väheneb ensüümide võimsus); mõne looma puhul mõjutab
eluiga insuliini tase (koeratõud, kel see madalam on, elavad kauem;
inimese kohta pole eriti täheldatud, sest selle puudus põhjustab
erin surmavaid defekte).
Kui
kromatiini ei remodelleerita e ei parandata selle katkestusi, siis
see kiirendab vananemist .
Vananemise
takistamisel osalevad geenid: DNA reparatsiooni ensüümid, insuliini
signaaliraja valgud, kromatiini remodelleerimise ensüümid.


































Kõik kommentaarid