Ensümoloogia alused.
KordamisküsimusedEnsüüm kui
valk: valgu struktuur,
aminohapped ,
mittekovalentsed
interaktsioonid ,
vesilahused ja
unikaalsed vee
omadused.
Valgu
funktsioneerimise tagab tema
struktuur.
Ensüüm kui
katalüsaator: keemiline
reaktsioon ,
termodünaamika,
kineetika , katalüüs,
mehhanism , ensüümide
kasutamine tööstuses.
Ensüüm kui
bioloogiline katalüsaator:
sidustatud
reaktsioonid, bioenergeetika,
metabolism ,
regulatsioon ,
klassifikatsioon ja nomenklatuur.
Ensüümid on organismide tööhobused.
1) Ensüümkatalüüsi põhimõisted ja printsiibid +
Ensüümkatalüüsi peamised tunnus-jooned. - Ensüümkatalüüs põhineb rangelt füüsikalistel ja keemilistel vastasmõjudel.
- Kõik ensüümid on evolutsioonilise arengu produktid ja kujunenud selliseks , nagu me neid täna näeme, evolutsiooni ja loodusliku valiku tulemusel.
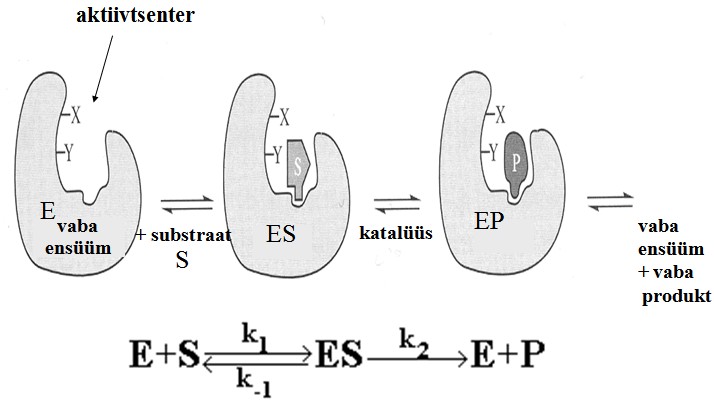
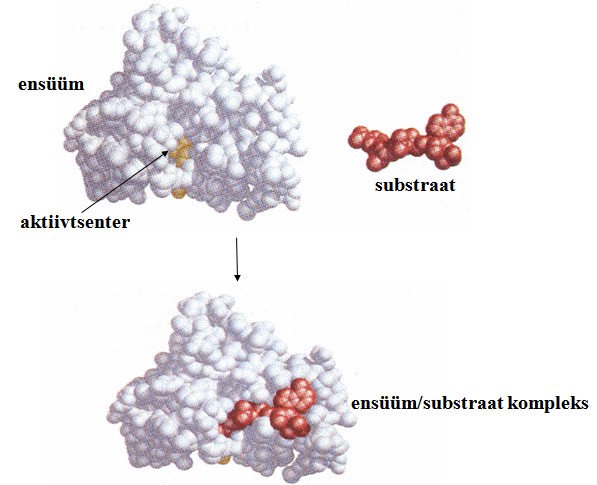
Substraat seostub ensüümi aktiivtsentrisse, mis
võtab enda alla tavaliselt vähem kui 5%
ensüümi pinnast.
Ensümaatilise reaktsiooni kiirust võib määrata
substraadi kontsentratsiooni vähenemise või
produkti kontsentratsiooni suurenemise
kaudu.
- Ensüümid on väga efektiivsed katalüsaatorid, mis kiirendavad reaktsioone kuni 1017 korda
- Reaktsioonid toimuvad pehmetes tingimustes (madal temperatuur ja rõhk, neutraalne pH)
- Reaktsioonidel on väga suur spetsiifilisus – iga ensüüm katalüüsib vaid teatud kindlaid reaktsioone
- Reaktsioonid alluvad regulatsioonile ( aktivaatorid , inhibiitorid )
- Ensüümid võivad muuta energia ühte vormi teiseks.
Substraat (S) on aine, mille muundumist ensüüm (E)
katalüüsib.
Produkt (P) on
aine, mis tekib substraadist ensüümi toimel .
Ühes reaktsioonis võib osaleb
mitu erinevat substraati ja tekib mitu
erinevat produkti
(substraadid A, B,C jne., produktid P, Q, R jne.)
Inhibiitor on
aine
või faktor, mis vähendab keemilise
reaktsiooni kiirust
või takistab reaktsiooni.
Tehislikud inhibiitorid on kasutusel kui
ravimid - näiteks proteaaside inhibiitorid HIV infektsiooni vastu.
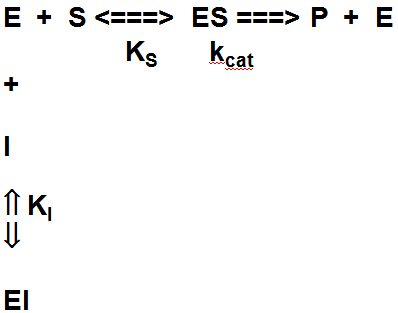
Konkurentsed inhibiitorid: seostuvad ensüümi
aktiivtsentrisse ja välistavad substraadi seostumise kas blokeerides
substraadi sidumistsentri või kutsudes esile valgu
konformatsioonimuutuse, mis välistab substraadi
sidumise .
Mittekonkurentsed
inhibiitorid: seostudes ensüümile
võimaldavad substraadi sidumist (kolmikkompleks moodustub) kuid ei
võimalda reaktsiooni
toimumist (
i.e.
EIS
ei ole reaktsioonivõimeline).
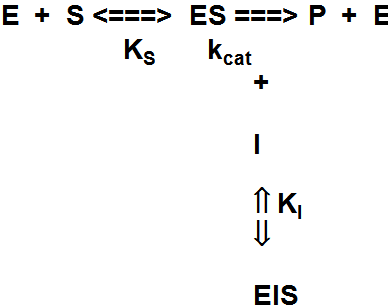
Ebakonkurentsed (un-competitive)
inhibiitorid:
seostuvad ainult
ensüüm-substraat kompleksiga, aga mitte vaba ensüümiga.
Suhteliselt harvaesinev inhibeerimise tüüp - esineb põhiliselt
multisubstraatsete reaktsioonide korral teatud tingimustes.
Substraat seostub ensüümi
aktiivtsentrisse
(aktiivalasse ehk aktiivsaiti).
Seejärel toimub katalüütiline
reaktsioon , mille käigus S muundub
P-ks.
Katalüsaatorid:
- Alandavad aktivatsioonienergiat .
- Ei mõjuta tasakaaluolekut.
- Osalevad reaktsioonis kuid ei muundu selle tulemusel (taastuvad reaktsiooni käigus)
- üks katalüüsitsenter või katalüsaatori molekul osaleb reaktsioonis korduvalt
- mõjutavad reaktsiooni kiirust juba väga väikses koguses
Biloogilised katalüsaatorid - ensüümid:
- Suurendavad reaktsiooni kiirust alandades aktivatsioonibarjääri EA kõrgust.
- EI MUUDA reaktsiooni SUUNDA
- On võimelised reaktsioone energeetiliselt omavahel sidustama - st teostama endergoonilisi protsesse eksergooniliste protsesside käigus vabaneva energia arvel
- Toimivad väga pehmetes tingimustes
- On väga selektiivsed ja äärmiselt kõrge spetsiifilisusega.
- Alluvad regulatsioonile
Ensüümkatalüüsi iseloomustab:
- Substraatspetsiifilisus (mis läheb sisse)
Erinevalt keemilistest katalüsaatoritest katalüüsivad ensüümid
suhteliselt kitsa substraatide valiku reaktsioone.
Näiteks: restriktaasid või
proteiinkinaasid.
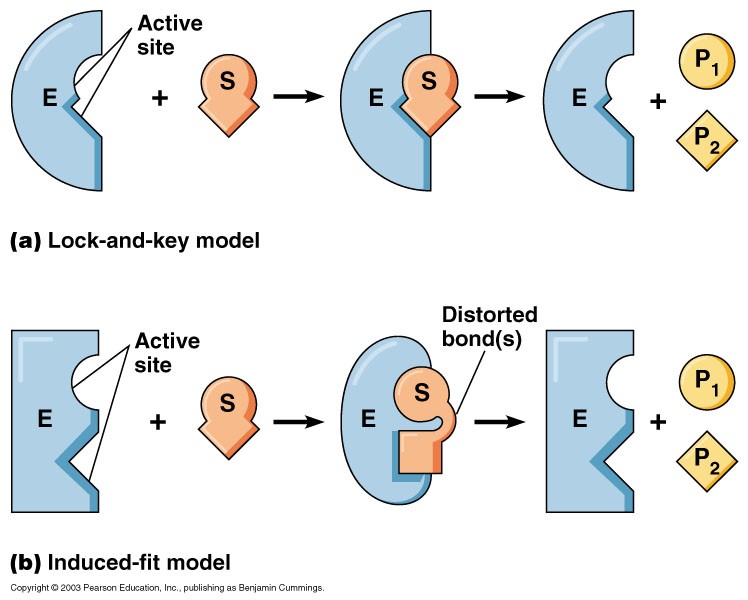
Substraatspetsiifilisus põhineb
eelpool loetletud interaktsioonidel
ja substraadi ning ensüümi ruumilisel sobivusel ning seda
kirjeldavad põhiliselt kaks hüpoteesi:
Luku ja võtme hüpotees (Fisher, 1894)
Indutseeritud sobivuse hüpotees
(Koshland, 1958)
- Stereospetsiifilisus (mis tuleb välja)
- Katalüütiline efekt (kui kiiresti kõik toimub
Ensüümkatalüüs: Kuidas ensüüm vähendab aktiveeritud ja
lähteoleku energiate vahet (alandab aktiveeritud oleku energiat):
- Viib reageerivad rühmad kokku sobivas orientatsioonis (Proximity)
- Solvateerib aktiveeritud olekut paremini kui lähteolekut
Lisaks võib ensüüm reaktsioonis “aktiivselt” osaleda
Igas metaboolses rajas on regulatoorne ensüüm ehk
võtmeensüüm,
mille
poolt katalüüsitava reaktsiooni kiirus määrab kogu metaboolse
raja kiiruse.
Võtmeensüümi poolt katalüüsitav reaktsioon toimub kõige
“aeglasemalt”, seetõttu limiteerib selle kiirus kogu raja
toimimist.
Metaboolse raja võtmeensüümi hulga ja aktiivsuse kaudu
reguleeritakse
kogu metaboolse raja kiirus. Selliste võtmeensüümide poolt
katalüüsitavate protsesside
statsionaarne olek on kaugel nende
tasakaaluolekust.
Ensüümide
aktiivsuse regulatsioon:
- Kovalentne modifitseerimine
- Pöörduv (fosforüülimine)
- Mittepöörduv (limiteeritud proteolüüs, metüleerimine)
- Mittekovalentsed regulatsioonimehhanismid
- Spetsiifilised aktovaatorid-inhbiitorid
- Produktpidurdus
- Keskonnatundlikud konformatsioonimuutused
- Kooperatiivsus ja allosteerika
- Kompartmelisatsioon
- Iga ensüüm paikneb kindlas raku piirkonnas või struktuuris
- See tagab ensüümreaktsioonide eraldatuse ja kindla järjestuse
- Substraadi ja kofaktorite kontsentratsioon
Ensüümid - katalüütiliste
omadustega
valgud (lihtensüümid) või valkude
kompleksid (liitensüümid). Ensüümid osalevad peaaegu kõikides
biokeemilistes reaktsioonides elusorganismides. Nii kontrollivad
ensüümid metaboolsete protsesside toimumise
kiirust;
ribosüümid - katalüüsivad ribonukleiinhapete
hüdrolüüsi ja ümberesterifikatsiooni reaktsioone (RNA
protsessing), mille käigus muudetakse primaarne RNA
transkript mRNA-ks. Ribosoomi RNA katalüüsib peptiidsidemete moodustumist;
absüümid - ehk katalüütilised
antikehad saadakse organismi
immuniseerimisel substraadi aktiveeritud oleku analoogiga. Erinevalt
ensüümidest ja ribosüümidest pole absüümidel tõelist
bioloogilist funktsiooni organismis. Võrreldes ensüümidega on
absüümide katalüülitiline aktiivsus palju väiksem.
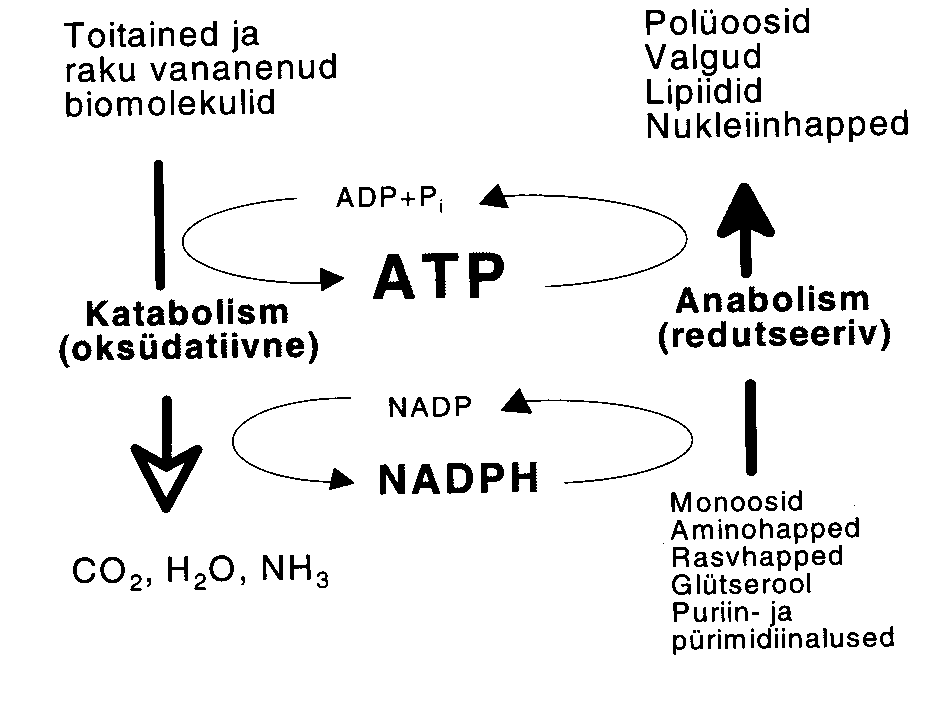
Ensüümkatalüüsi termodünaamika.
Energia on võime teha tööd.
- Energia liigid
- Potentsiaalne energia
- Kineetiline energia
- Energia muundumine
- Vabaneb soojus
- Tööd tehakse keha viimiseks suurema potentsiaalse energiaga olekusse. Tööd saab teha ainult süsteemi vabaenergia arvel.
Energia allikad:
- Päikesevalgus
- Keemiliste sidemete energia
Keemilistes
sidemetes salvestatud potentsiaalne energia võib:
- kanduda produktide keemilistesse sidemetesse (keemiline töö)
- Kasvamine, valgu süntees jne.
- realiseeruda mehhaanilise tööna - liikumine, transport raku sees
- realiseeruda elektrokeemilise tööna - ioonide transport läbi membraanide
- eralduda soojusena
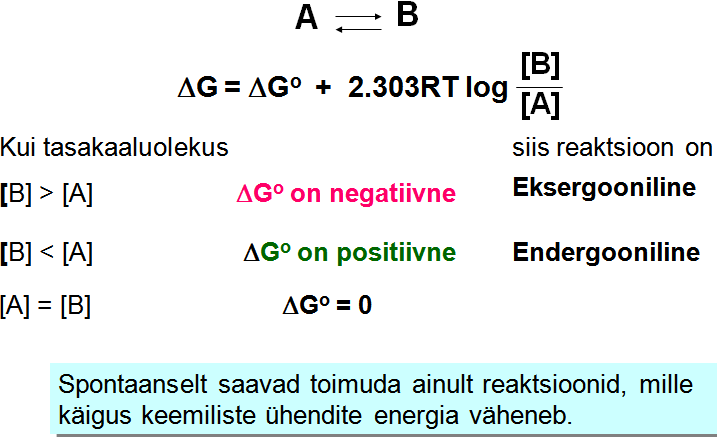
Vabaenergia väärtus näitab energia hulka, mis vabaneb reaktsiooni
käigus või vajatakse reaktsiooni teostamiseks. Märk näitab
reaktsiooni võimaliku kulgemise suunda.
Kui reaktsiooni Go
on positiivne, võib
G
väärtus sõltuvalt substraatide ja produktide
kontsentratsioonist
olla siiski negatiivne.Mitmed reaktsioonid, mille Go
on positiivne, kulgevad spontaanselt, kuna teised reaktsioonid
lagundavad produkte või tagavad lähteainete (substraatide) kõrge
kontsentratsiooni.
Aktiivne transport:
ATP hüdrolüüsi energia arvel (DG0).
ATP süntees:
Spontaanse H+
voo (DG0)
-
mitokonder .
Vabaenergia muutus (G)
määrab ära reaktsiooni toimumise võimaliku suuna, aga ei
ütle midagi reaktsiooni tegeliku toimumise kohta.
* Miks logaritmitakse konstantide väärtusi?
Tasakaalukonstandi
logaritm on
lineaarses sõltuvuses
interaktsiooni energiast. Energiad on aditiivsed ning me saame arvutada erinevate
ensüümi või substraadi üksikute osade
energeetilised panused ning teha neist järeldusi protsessi olemuse kohta.
Aktiveeritud vaheoleku teooria.
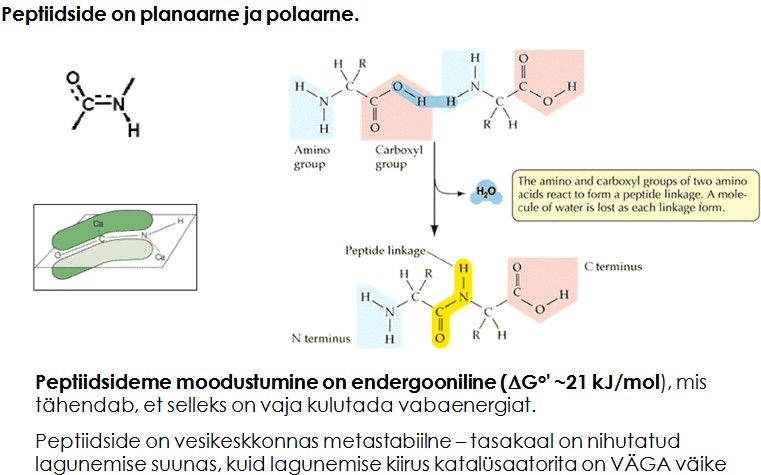
Ensüümide struktuur.
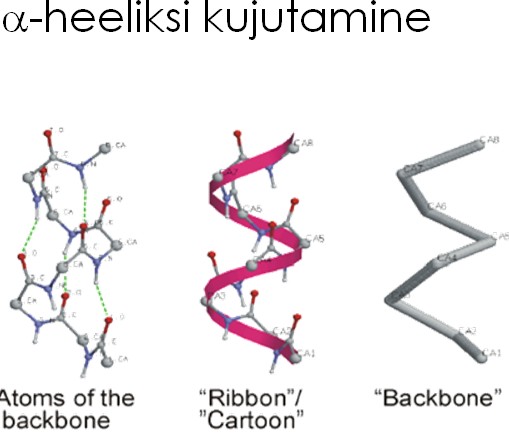
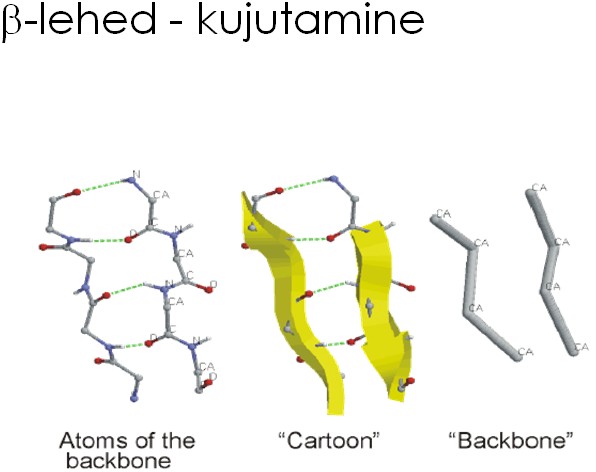
- Primaarstruktuur – valgu järjestus
- Sekundaarstruktuur - peptiidahela konformatsioon või organiseeritus (kõrvalahelate paiknemist arvestamata). (peptiidahela lokaalne organiseeritus (aminohapete kõrvalahelate – R-rühmade - konformatsioone ignoreeritakse).)
- Tertsiaarstruktuur - kogu peptiidahela (k.a. kõrvalahelad) ruumiline paigutus .
- Kvaternaarstruktuur - valgu subühikute (ja domäänide) omavaheline ruumiline paigutus.
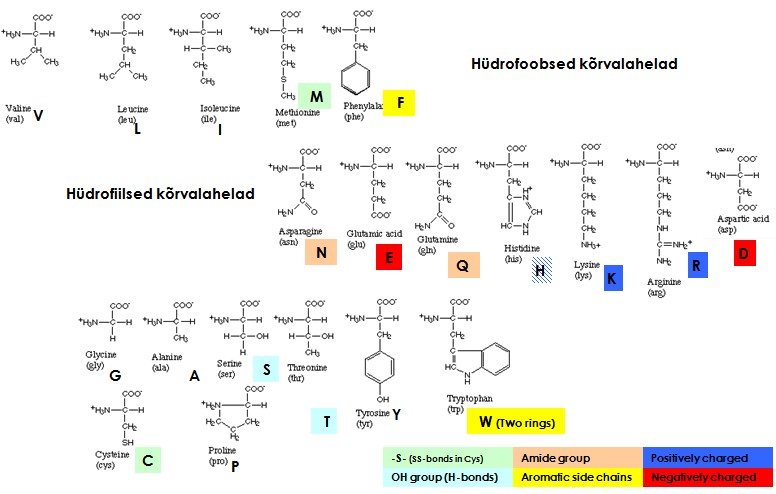
- Valkudes esineb 20 aminohapet (lisaks post-translatoorsed modifikatsioonid) , mille omadused tagavad valkude struktuuri ja toimimise:
Aminohapete omadused:
- suhteline hüdrofoobsus ja polaarsus
- ionisatsioon
- muud keemilised omadused
Gly increases the flexibility of the polypeptide backbone since there is no steric
hindrance from the side chain - this permits sharp turns or bends and
the ability to fit in tight spots. Gly residues are often highly conserved, and often found at turns.
Ala
with its small side-chain is found equally in the interior and on the
surface and is a very abundant amino acid in proteins.
Val, Leu, Ile
with their bulky hydrophobic side-chains are stiffer due to steric
hindrance, their bulk also tends to make them helix breakers.
Hydrophobic side chains are most commonly found in the interior, i.
e. buried , of proteins.The aromatic amino acids have a b-methylene to provide some flexibility; they are non- polar , although Tyr
has the polar phenol group. Aromatic side-chains tend to pack
together at 90°C , rather than side-to-side. Trp
has the largest side-chain.
Pro is
non-polar and has a very rigid side-chain, causing a "kink"
in the peptide backbone. Usually Pro breaks a helix, and many turns contain Pro. It usually is found in the more stable trans
configuration, but occasionally in the cis configuration.
Polar side chains (Ser,
Thr,
Cys,
His,
Asn,
Gln)
have the potential to form H-bonds with the solvent , each other and
the peptide bond. This is important in what is called helix-capping,
in which the unfulfilled H-bonds of the backbone are satisfied. When H-bonded and buried they are effectively no longer polar. The pK of
the imidazole of His (6.7) means that at physiological pH it can function either as an acid or a base . Many enzymes use His as a
general acid/base catalyst.
Cys can
make disulfide bonds, which act
as covalent cross- links .
Charged side chains are usually found on the
surface. Buried ones tend to have several H-bonds to "diffuse"
the charge . Salt bridges are ion pairs between an acid and basic amino acid. The Asp
side chain is more rigid than that of Glu
due to the shorter chain in Asp, thus Asp is more frequently found in
the active sites of enzymes.
Kvaternaarstruktuur on eriti oluline mitmest subühikust
koosneva valgu funktsioneerimiseks (ensüümide korral allosteeriline
regulatsioon)
Ensüümkatalüüsi iseloomustavad kineetilised parameetrid.
Homogeense keemilise reaktsiooni kiirus on reagentide
kontsentratsiooni muutus ajas ja selle ühikuks on M s-1
(ensüümide korral sageli μM/min)
Kiirust võib väljendada nii produkti tekkimise kui lähteainete
kadumise kaudu
Reaktsiooni A
® B
kiirus= v
= -D[A]/ D t = D [B]/ D t üldisemalt,
v
= -d[A]/dt = d[B]/dt Kiiruskonstant on tegur, mis muudab valemi
täpseks võrrandiks kiirus = k[A].
Kiirus-konstandi ühikud peavad olema sellised, et kiirus
oleks väljendatud sobivates ühikutes - antud juhul on konstandi
dimensiooniks aeg-1.
Rekatsiooni järk: A+B® C+D.
Reaktsiooni järk on number, mis näitab
kontsentratsiooni väljendavate liikmete hulka kineetilises võrrandis
Esimest järku reaktsioon:
rate = k[A]
Kiiruskonstandi dimensiooniks on aeg-1
- ühikuks on s-1
Teist järku reaktsioon: rate = k[A][B]
rate = k[A]2
Kiiruskonstandi dimensiooniks on kontsentratsioon-1•
aeg-1
- M-1
s-1
Null-järku reaktsioon: rate = k
Kiirus on sõltumatu
reagentide kontsentratsioonist ja kiiruskonstandi dimensioon
langeb kokku kiiruse dimensiooniga
kontsentratsioon•aeg-1
- Ms-1
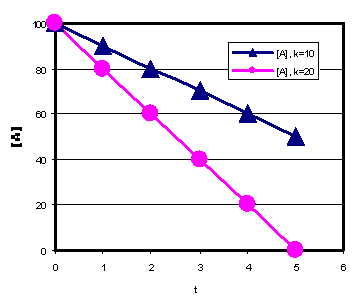
- Kiiruskonstant sõltub reageerivate ainete iseloomust ja väljendab reaktsiooni kiirust kindlates tingimustes (pH, temperatuur, puhverlahus , rõhk jne.)
- Mida suurem on konstandi väärtus, seda kiirem on teiste parameetrite (järk ja kontsentratsioonid) võrdsuse korral reaktsioon
- Ainult ühte ja sama järku reaktsioonide kiiruskonstantide võrdlus on mõtestatud.
0-järku:
- Reagendi kontsentratsioon väheneb ajas lineaarselt, sirge tõus=k
- Mida suurem on k väärtus, seda kiirem on A kontsentratsiooni vähenemine
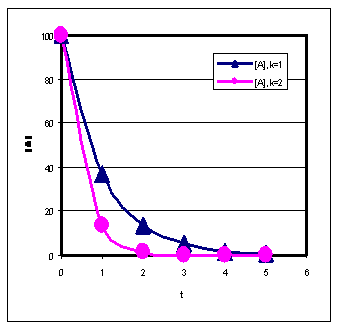
1-järku:
- Kiiruskonstandi määramiseks on soovitatav kasutada eksponentsiaalset kuju. Kui me mõõtsime produkti, siis on lineaarne võrrand oluliselt ebatäpsem.
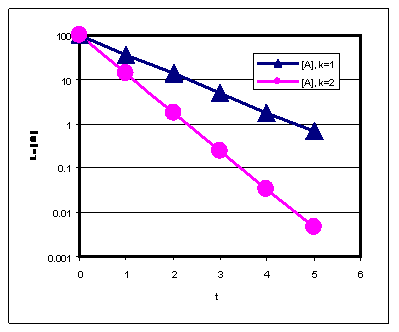
Poolestusaeg :
- Defineerime t1/2 = aeg, mille jooksul pool lähteainest on muundunud produktiks.
- I järku reaktsiooni poolestusaeg ei sõltu kontsentratsioonist.
- Pseudo esimest järku reaktsioon on reaktsioon, mis näib olevat esimest järku (st produkti teke toimub vastavalt esimest järku kineetilisele võrrandile), aga tegelikult on reaktsioon teist järku (tema kiirus sõltub mõlema reagendi kontsentratsioonist).
- Tavaliselt on pseudo esimest järku tingimustes ühe reageeriva ühendi kontsentratsioon konstantne , kuna
- see on teisest oluliselt suurem (näiteks reaktsioon solvendiga)
- tegemist on katalüsaatoriga
- selle kontsentratsioon on puhverdatud
Ensüümi juuresolekul muutub sõltuvus reaktsioonikiiruse ja
substraadi kontsentratsiooni vahel hüperboolseks.
Ensüümi poolt katalüüsitaval reaktsioonil:
Madalatel [S] väärtustel,
vcat = f([S])
Kõrgetel [S] väärtustel,
vcat -->
constant .
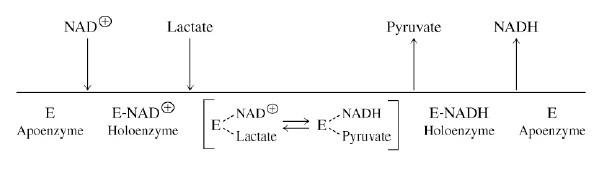
Kofaktorid ja koensüümid.
Koensüümid on orgaanilised või metallo-orgaanised, mis on
ensüümiga tihedalt seotud ja vajalikud katalüüsi toimumiseks.
Ensüümiga kovalentselt seotud kofaktorit
nimetatakse
prosteetiliseks
rühmaks
Kofaktorid osalevad sageli otseselt katalüütilises protsessis
kandes üle elektrone või funktsionaalseid rühmi substraadilt
produktile.
Ensüümkatalüüsi spetsiifilisus.Homogeenses katalüüsis on reageerivad ained ja katalüsaator
samas faasis.
Heterogeenses katalüüsis
moodustab katalüsaator eri faasi, katalüüs
toimub katalüsaatori pinnal (katalüüs metallide ja mitmete
oksiidide pinnal).
Ensüümkatalüüsil
on mõlema tüübile
iseloomulikke jooni:
1) ensüüm on tavaliselt reageerivate
ainetega samas faasis (vedelas); 2) ensüümide suuruse tõttu on
nende pind, kus toimub katalüütiline reaktsioon, teistsuguste
omadustega kui ümbritsev lahus.
2.Ensüümide nomenklatuur, klassifikatsioon ja EC kood:Ensüümi nimetus tuleneb tema poolt lõhustatava substraadi
nimetusest (
laktoos –
laktaas ). Ensüümile
viitab substraadi nimetuse lõpp “aas” (
lipaas
katalüüsib
lipiid triglütseriidi hüdrolüüsi ja
sahharaas katalüüsib
sahharoosi hüdrolüüsi glükoosiks ja
fruktoosiks). Peale substraadi nimetuse märgitakse
ensüümi nimetuses tavaliselt ära
katalüüsitava reaktsiooni nimetus/tüüp (laktaadi
dehüdrogenaas tähistab seda, et
substraadiks
on
laktaat
ja toimub selle
dehüdrogeenimine).
Multiensüümkomplekside puhul kasutatakse lisandit “
kompleks ”
(Püruvaadi dehüdrogenaasne kompleks). Tihti kasutatakse ka
ajaloolisi nimetusi, näiteks Trüpsiin ja pepsin.
Vastavalt katalüüsitavatele
reaktsioonidele jaotatakse ensüümid
kuude klassi. Iga klass jaotub alaklassideks (subclass) ja need
omakorda alaalaklassideks (subsubclass).
Ensüümi
klassid :
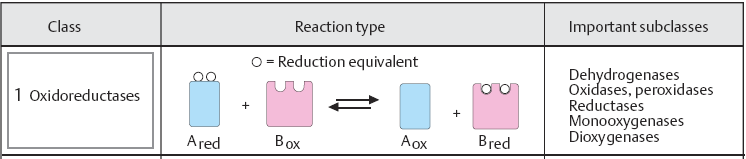
Oksüdoreduktaasid (Oxidoreductases)
Transferaasid (Transferases)
Hüdrolaasid (Hydrolases)
Lüaasid (Lyases)
Isomeraasid (Isomerases)
Ligaasid (Ligases)
Ensüümi alaklassid (igas klassis on alaklassi tähendus erinev):
oksüdoreduktaasid – redoksreaktsioonis osaleva rühma keemilist loomust
transferaasid – ülekantava rühma nime
hüdrolaasid – hüdrolüüsitava sideme tüüpi
lüaasid – lõhustatava sideme tüüpi
isomeraasid – isomerisatsiooni tüüpi
ligaasid – moodustuva sideme tüüpi
Ensüümi alaalaklassid:
milline elektronide/vesinike aktseptor osaleb reaktsioonis
täpsustab ülekantava rühma struktuuri
millisesse ühendiklassi kuulub hüdrolüüsitav substraat
millisesse ühendiklassi kuulub substraat, mis lagundatakse
substraadi ühendiklassi
reaktsioonil tekkiva produkti ühendiklassi
Iga ensüümi jaoks on neljanumbriline kood. Koodile eelneb lühend
EC ( Enzyme Comission). Esimene number näitab ensüümi klassi (1
kuni 6) ehk millist tüüpi reaktsiooni ensüüm katalüüsib, teine
number näitab alaklassi, igas klassis on alaklassi tähendus erinev,
kolmas number tähistab alaalaklassi ja neljas number on
järjekorranumber rühma piires.
Täpsem info ala ning alaalaklasside kohta: http://www.chem.qmul.ac.uk/iubmb/enzyme/ .
EC 1. Oksüdoreduktaasid ja nende süstemaatilised nimetused:
Orgaanilise aine oksüdeerumisel loovutatakse koos elektronidega
prootoneid või asendatakse vesinikuaatomid mõne elektronegatiivsema
aatomiga (näit. hapnikuga). Oksüdeeritust näitab süsinikega
seotud vesinike arvu ja süsinike üldarvu suhe. Mida väiksem on see
suhe, seda oksüdeeritum on ühend.
Oksüdoreduktaasid katalüüsivad redoksreaktsioone, ehk nad on
ensüümid, mis kannavad elektrone ühelt molekulilt teisele
oksüdatsioonireaktsiooni käigus.
A + D-
→ A-
+ D
Redoksreaktsioonides toimub elektronide ülekanne doonorilt
aktseptorile.
Kõik biokeemilised redoksreaktsioonid toimuvad koensüümide
osavõtul.
Redoksreaktsioonis doonor oksüdeerub ja aktseptor redutseerub.
Doonoriks on metaboliit (metabolismi vaheprodukt). Aktseptoriks
võivad olla koensüümid
(NAD+
, FAD, jt.), mis on seotud
oksüdoreduktaaside aktiivtsentrisse või hapnik , metalliioonid, lipohape , disulfiidid.
Oksüdoreduktaaside süstemaatilised nimetused:
Klassikalise nimena kasutatakse doonor:aktseptor
oksüdoreduktaasi,
ka nimi dehüdrogenaas
on kasutatav. Võib kasutada ka nime
aktseptor reduktaas,
kuid need klassifikatsioonid võivad olla keerulised juhtudel,
kui mitmeid erinevaid retseptoreid kasutatakse sama ensüümi poolt.
Näiteks: alkohol :NAD+ oksüdoreduktaas
(alkoholdehüdrogenaas), kus alkohol on
doonor ning NAD+ on akseptor .
EC 2. Transferaasid ja nende süstemaatilised nimetused:
Transferaasid katalüüsivad rühmade ülekannet ühelt molekulilt
teisele:
X-Y + Z → X + Z-Y ,
kus Y on ülekantav rühm.
Need ensüümid kannavad C, N, P või
S sisaldavaid gruppe (alküül,
atsüül, aldehüüd, amino, fosfaat või glükosüül)
ühelt substraadilt teisele.
Metüültransferaasid – metüülrühmade ülekanne
Atsüültransferaasid – atsüülrühmade ülekanne
Aminotransferaasid – aminorühmade ülekanne
Glükosüültransferaasid – glükosüüljäägi ülekanne
Ketotransferaasid – k etorühmade ülekanne
Transferaaside süstemaatilised nimetused:
Näiteks: ATP : heksoos 6 – fosfo transferaas
(heksoosi kinaas ), kus ATP in doonor, heksoos on
akseptor ning 6 – fosfo on ülekantav rühm.
EC 3. Hüdrolaasid ja nende süstemaatilised
nimetused:
Katalüüsivad hüdrolüüsireaktsioone (reaktsioone veega) ning
nende reaktsioonide käigus katkevad sidemed C-O, C-N, C-P, C-C, C-S.
Hüdrolaaside süstemaatilised nimetused:
Näiteks: atsetüül-CoA hüdrolaas, kus atsetüül-CoA
on substraadi nimi.
EC 4.0 Lüaasid ja nende süstemaatilised nimetused:
Lüaasid on ensüümid, mis katalüüsivad lagunemis- ehk lüüsumisreaktsioone. Sidemete lõhustumises ei osale veemolekulid ja oksüdeerijad. Lüaaside reaktsioonide käigus tekib reeglina
kaksikside või ringstruktuur. Vastureaktsioonis katalüüsitakse
kahe substraadi ühinemist, seetõttu kutsutakse lüaase ka
süntaasideks
Lüaaside süstemaatilised nimetused:
Näiteks: tsitraadi hüdro-lüaas, kus tsitraat
on substraat ning hüdro (H2O) on eralduv rühm.
Lüaas
Reaktsioon
dehüdrataas
vee elimineerimine
hüdrataasid
vee liitmine
dekarboksülaas
CO2 elimineerimine
süntaas
kondensatsioonireaktsioonid, mis
ei vaja ATP, GTP jt energiat
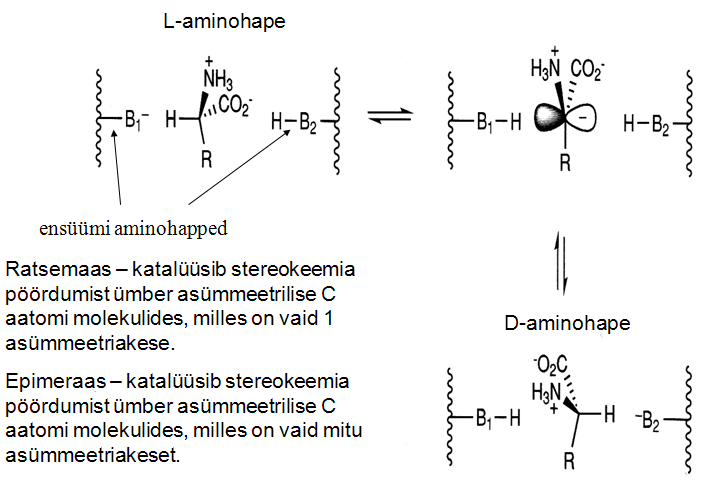
EC 5. Isomeraasid ja nende süstemaatilised nimetused:
Isomeraasid katalüüsivad muutusi ühe molekuli piirides ehk
isomerisatsiooni. Isomeraaside reaktsioonides on üks substraat ja
üks produkt. Isomeraasid jagunevad: ratsemaasideks, epimeraasideks,
mutaasideks ning cis-trans-isomeraasideks.
Isomeraaside süstemaatilised nimetused:
Aluseks on substraadi nimetus ja reaktsioonitüüp.
Näiteks: Laktaadi ratsemaas, see ensüüm katalüseerib
reaktsiooni (S)-laktaat ↔ (R)-laktaat.
EC 6. Ligaasid ehk süntetaasid ja nende süstemaatilised
nimetused:
Katalüüsivad ühinemisreaktsioone (ligation – ing.k
kokkuliitmine). Reaktsioonid toimumiseks on tarvis täiendavat
energiat, see saadakse ATP või mõne muu ühendi makroergiliste
sideme(te) lagundamisest. Ligaasid katalüüsivad C-C,
C-O, C-N, C-S ja C-halogeen sidemete
moodustumist ilma hüdrolüüsi või oksüdatsioonita.
Ligaaside süstemaatilised nimetused:
Näiteks: L- aspartaat : ammoniaak ligaas (EC 6.3.1.1), kus
L-aspartaat ja ammoniaak on substraadid. Reaktsioon,
mille toimumist see ensüüm katalüüsib on L-aspartaat + NH3
+ ATP → L-asparagiin + AMP + difosfaat .
Isoensüümid:
Isoensüümid on ensüümid, mis katalüüsivad ühesuguseid
reaktsioone, kuid erinevad üksteisest aminohappelise
koostise/järjestuse poolest (neid kodeerivad erinevad geenid ).
Näiteks südamelihases esineb kaks malaadi dehüdrogenaasi – üks
tsütoplasmas, teine mitokondris. Ensüüme, mis erinevad ainult
posttranslatsiooniliste modifikatsioonide poolest (fosforüülimine, lipideerimine , glükosüleerimine), isoensüümide hulka ei loeta.
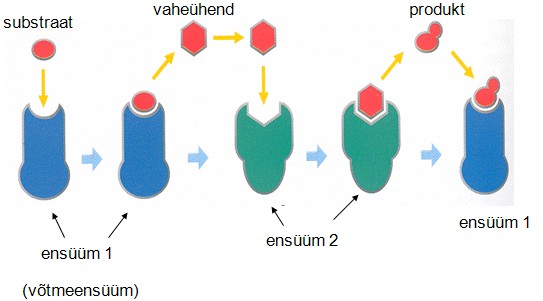
Multiensüümsed süsteemid:
Multiensüümsed süsteemid koosnevad mitmest erineva katalüütilise
aktiivsusega ensüümist. Multiensüümsed süsteemid viivad reeglina
läbi üksteisele järgnevate reaktsioonide toimumise. Ühe
reaktsiooni produkt on teise reaktsiooni lähteaineks. Multiensüümide
jaoks pole EC koodi, nende koostises olevatel ensüümidel on oma EC
koodid.
Multiensüümsed süsteemid jagunevad multiensüümseteks
kompleksideks (koosnevad mitmest erineva katalüütilise
aktiivsusega polüpeptiidahelast) ja multiensüümseteks
polüpeptiidideks (Koosnevad ühest polüpeptiidahelast. See
polüpeptiid moodustab mitu erineva katalüütilise aktiivsusega
tsentrit).
Multiensüümsete komplekside esindajad: püruvaadi dehüdrogenaasne
kompleks (3 erinevat aktiivtsentrit), trüptofaani süntaas (2
aktiivtsentrit) ja DNA polümeraas (3 aktiivtsentrit). Multiensüümse
polüpeptiidi esindajad: rasvhappe süntaas (7 erinevat
aktiivtsentrit).
Koensüümid:
Paljudes ensüüm-katalüüsitavates reaktsioonides kantakse ühelt
substraadilt teisele elektrone või aatomite gruppe. Sellistes
reaktsioonides osalevad alati lisamolekulid, mis ajutiselt võtavad
endale transporditava grupi. Neid “aitajaid” nim. koensüümideks.
Mittevalgulised orgaanilised ühendid, mis tavaliselt sisaldavad
vitamiini või mineraali. Koensüümid ei ole ise katalüütiliselt
aktiivsed. Koensüümid pole substraadi-spetsiifilised, vaid toimivad
koos erineva spetsiifikaga ensüümidega.
Näiteid erinevatesse klassidesse kuuluvate ensüümide
toimemehhanismidest:
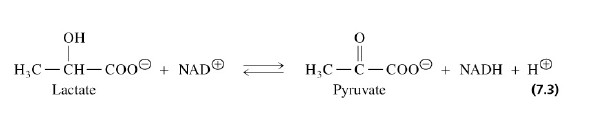
Laktaadi dehüdrogenaas:
Laktaadi dehüdrogenaas tähistab seda, et
substraadiks on
laktaat ja toimub selle
dehüdrogeenimine (näiteks: CH3CH2OH (alcohol) ↔ CH3CHO (aldehüüd)).
Proteiinkinaas :
Lipaas :
Proteaasid :
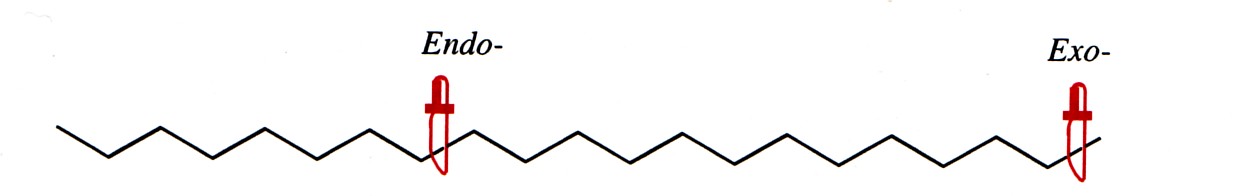
Endoproteaasid katalüüsivad sidemete hüdrolüüsi peptiidahela
keskel, eksoproteaasid ahela lõpus.
Karbonaadi hüdro-lüaas:
Aitab CO2 eemaldamisel kudedest ja transporti punastest
vererakkudest kopsudesse, kofaktoriks on Zn2+ .
CO2 + H2O ↔ HCO3 + H+
Ratsemaasid:
3.
Eksperimentaalsed meetodid ensümoloogias:
Eksperimendi eesmärgid:
ensüümi kontsentratsiooni ja aktiivsuse määramine
substraadi hulga/kontsentratsiooni määramine
- metaboolsete protsesside jälgimine
- meditsiiniline diagnostika
- keskkonna seisundi jälgimine
ensüümkatalüüsi mehhanismide uurimine
- ensüümi struktuuri kindlakstegemine
- ensüüm-substraat või ensüüm-inhibiitor kompleksi struktuuri kindlakstegemine ensüümreaktsiooni iseloomustavate tasakaalu- ja kiiruskonstantide määramine produktide ja vaheühendite kindlakstegemine
ensüümide puhastamine
- laboratoorseks kasutamiseks
- tööstuslikuks kasutamiseks
ensüümide omaduste muutmine ja uute ensüümide disain
ensüümide tehnoloogilised rakendused
- orgaaniline peensüntees
- toiduainetetööstus
- ravimitööstus
- tekstiilitööstus
Ensüümi aktiivsus ja selle määramine:
Aktiivsuse määramiseks on tarvis määrata substraadi või
produkti kontsentratsiooni muutumine ajas ehk reaktsiooni toimumise
kiirus. Kuivõrd produkti kontsentratsiooni suhteline muutus on
suurem kui substraadil, on eelistatum määrata reaktsiooni kiirus
produkti tekkekiiruse järgi. Reaktsiooni
kiiruse määramiseks tuleb kasutada meetodit, mis võimaldab
eristada produkti kontsentratsiooni muutumise substraadi omast.
Aktiivsuse ühikuks on kogus ensüümi, mis muundab kindla koguse
substraati produktiks teatud kindla ajavahemiku jooksul. Kõige enam
on aktiivsuse ühikuna kasutusel Unit , lühendina U või IU
(International Unit). See vastab ensüümihulgale, mis muundab 1
μmooli substraati produktiks 1 minuti jooksul. Vähem kasutatakse
aktiivsuse ühikuna SI süsteemi ühikut katal (kat). 1 katal on
ensüümihulk, mis muundab ühe 1 mooli substraati produktiks 1
sekundi jooksul.
1 kat = 6 x 107 Unit | 1 Unit = 16.67 nkat
Ensüümi aktiivsuse määramisel tuleb kontrollida järgmisi
parameetreid:
- temperatuur
- pH
- ioonne jõud
- aktivaatorite või stabilisaatorite juuresolek
- redokspotentsiaal
- (raskemetallid, lisandid)
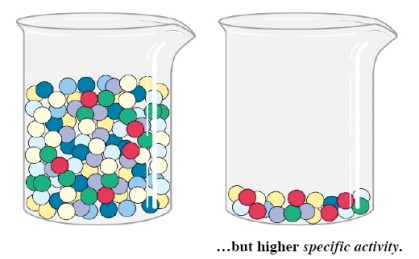
Eriaktiivsus :
Eriaktiivsus on ensüümi ühikute arv proovi kuivmassi või proovi
valgumassi kohta. Tavaliselt väljendatakse eriaktiivsus 1 mg valgu
kohta.
Eriaktiivsuse arvutamine:
- määratakse ühikute arv proovi mingis koguses (näiteks ml) ehk ühikute kontsentratsioon
- määratakse valgu kontsentratsioon proovis
Eriaktiivsuse saame, kui jagame ühikute kontsentratsiooni valgu kontsentratsiooniga . Eriaktiivsuse ühikuks on seega U/mg , mU/g,
kat/kg jne.
Eriaktiivsuse väärtus sõltub:
- ensüümi katalüütilist efektiivsusest
- aktiivse ensüümi sisaldusest proovis
Eriaktiivsust kasutatakse peamiselt ensüümipreparaadi kvaliteedi ja puhtuse hindamiseks. Kui puhta ensüümi
eriaktiivsus on teada, iseloomustab uuritava preparaadi eriaktiivsuse
ja tõelise eriaktiivsuse väärtuse suhe seda osa üldvalgust, mis
kuulub aktiivsele ensüümile.
Aktiivsete ensüümimolekulide (punased) arv ehk ühikute arv on
sama, kuid eriaktiivsus on kõrgem keeduklaasis 2:
Ensüümi aktiivsuse faasid :
Prestatsionaarne faas - eelstatsionaarses faasis ES tekkimis - ja lagunemis- kiirused pole võrdsed. Sõltuvalt mehhanismist esineb „lag”( viivitus ) või „burst” (purse). See faas kestab tavaliselt milli- või mikrosekundeid.
Statsionaarne faas - on reaktsioonikiirus konstantne, ES tekkimis- ja lagunemiskiirused on võrdsed.
Lõpp faas - substraadi kontsentratsioon on vähenenud sedavõrd, et ES kontsentratsioon hakkab märgatavalt langema ning reaktsiooni kiirus langeb. Reaktsioon lõpeb, kui kogu substraat on reageerinud või süsteem on jõudnud tasakaalu.
Aktiivsuse määramisviisid:
Otsene-pidev ensüümi aktiivsuse määramine: Reaktsioonisegus on võimalik pidevalt jälgida produkti/substraadi kontsentratsiooni muutust ajas. Mõõdetakse pidevalt mõne füüsikalise parameetri väärtust (optiline tihedus, pH, juhtivus jne.), mis on võrdeliselt seotud produkti või substraadi kontsentratsiooniga. Kõige mugavam ja väikseima töömahuga meetod
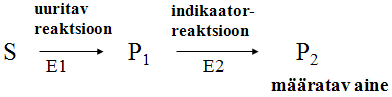
Kaudne-pidev ensüümi aktiivsuse määramine: Kasutatakse juhul, kui pidevate jälgimismeetodite abil pole produkti ja substraadi eristamine võimalik. Uuritava ensüümi (E1) toimel tekkinud produkt P1 muudetakse reaktsioonisegus teise reaktsiooni ehk indikaatorreaktsiooni abil ühendiks P2, mille hulka/kontsentratsiooni saab teatud meetodi abil pidevalt mõõta.
Uuritava
ensüümi kvantitatiivseks määramiseks ei tohi indikaatorreaktsioon
olla kiirust
limiteerivaks
staadiumiks
Aktiivsuse määramine proovide võtmisega: Teatud ajavahemike (t1, t2, t3, t4 jne) järel võetakse reaktsioonisegust proov , milles reaktsioon peatatakse. Kõikides proovides lahutatakse produkt substraadist ja määratakse nende mõlemavõi ühe kontsentratsioon.
Produkti
kontsentratsioon on määratud iga minuti järel ning kantakse
graafikule, kus x- teljel on aeg ning y-teljel produkt.
Instrumentaalsed meetodid aktiivsuse määramiseks:
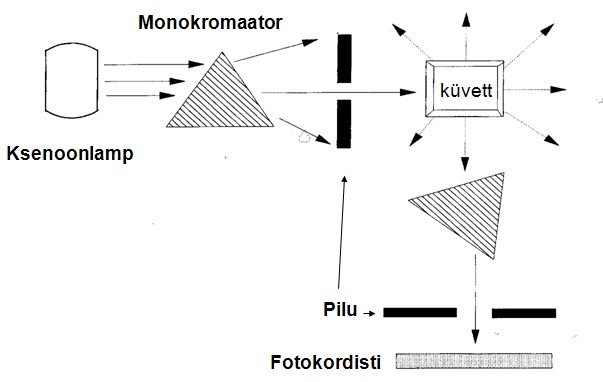
Spektrofotomeetria :
Spektrofotomeetriat saab kasutada, kui substraat ja produkt neelavad elektromagnetkiirgust erinevalt. Registreeritakse optilise tiheduse
muutusi, mis toimuvad spektri nähtava valguse või
ultraviolettkiirguse (UV-kiirgus) piirkonnas, lainepikkuste 200-700
nm vahel. Tavaliselt fikseeritakse optilise tiheduse muutus ühel
kindlal lainepikkusel. Nähtava valguse ja ultraviolettkiirguse
piirkonnas eelavad ained, millede koostises on kaksiksidemeid , eriti
hästi aga aromaatseid tsükleid sisaldavaid rühmi. Selliseid rühmi
nimetatakse kromofoorideks.
Lahuse optiline tihedus A (absorbance) avaldub:
kus Io –
valgusallikast tuleva valguse intensiivsus ning I -
lahuse läbinud valguse intensiivsus.
Valguse neeldumist kirjeldab Lambert -Beeri seadus:
Fluorestsents- spektroskoopia :
Fluorestsentsi võib kirjeldada järgmise skeemi abil:
Fluorestsents-spektroskoopia on umbes 100 korda tundlikum kui
spektrofotomeetria. Looduses on väga vähe piisavalt tugeva
fluorestsentsiga ühendeid. Seetõttu kasutatakse sünteetilisi
substraate, mille koostisesse on viidud fluorogeenseid rühmi.
F – fluorestsentsi intensiivsus
Io - ergastava kiirguse intensiivsus
ε - ekstinktsioonikoefitsent
c - molaarne kontsentratsioon
l - neelava kihi paksus
Q - kvantsaagis
Kui εcl on väike, lihtsustub valemiks:
kus F on proportsionaalselt sõltuv kontsentratsioonist.
Mida madalam on lahuse optiline tihedus, seda korrektsem on
spektrofluorimeetriline mõõtmine!
Turbidimeetria:
Mitmete ensüümreaktsioonide substraadid või produktid ei lahustu
vees. Lahustumatud osakesed ehk agregaadid hajutavad oluliselt
kiirgust ja nende lahused ei allu samadele seaduspärasustele kui
homogeensed lahused. Turbidimeetria on meetod hajunud valguse
intensiivsuse määramiseks. Mida rohkem ja mida suuremad on
agregaadid, seda enam kiirgust hajutatakse.
Turbidimeetria rakendused:
- lüsotsüüm (bakterite seinu lõhustav ensüüm)
- lipaasid
- vere hüübimises ja fibrinolüüsis osalevad proteaasid
Polarimeetria:
Polarimeeter on seade, mis võimaldab mõõta valguse
polarisatsioonitasandi pöördenurga muutumist. Polarimeetrit saab
kasutada selliste ensüümreaktsioonide jälgimiseks, milles osalevad
optiliselt aktiivsed ühendid.
Polarisatsioonitasandi pöördenurk on määratud valemiga:
c – aine kontsentratsioon
l – valguse poolt läbitava lahusekihi paksus
[α] - eripöörang,
iseloomustab ainet
α - polarisatsioonitasandi pöördenurk (o)
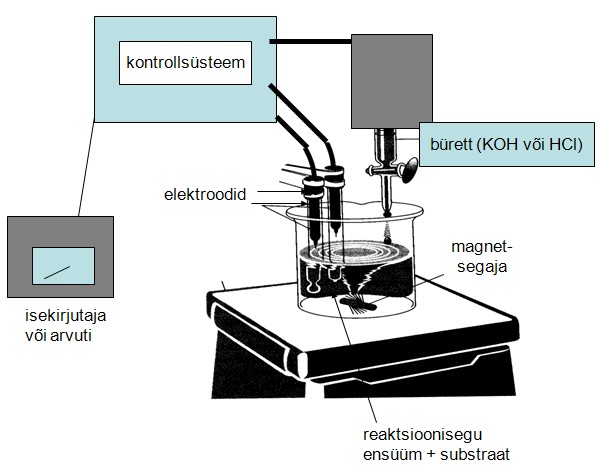
Elektrokeemilised meetodid (pH staat, hapnikuelektrood):
- pH-staat: Kasutatakse reaktsioonide puhul, millede käigus tekib H+ või OH-. pH-staat hoiab lahuse pH konstantsena, lisades sõltuvalt reaktsioonist kas leelist või hapet. Lisatud leelis või hape neutraliseerib reaktsioonil moodustuva happe või leelise. Kuivõrd lahuse pH hoitakse konstantsena, siis on reaktsioonikiirus võrdne leelise (happe) lisamiskiirusega. pH-staati kasutatakse peamiselt hüdrolaaside aktiivsuse määramiseks (tundlikkus kuni 50 nmooli).
- Hapnikuelektrood: Hapnikuelektrood võimaldab mõõta hapniku kontsentratsiooni lahuses. Seega saab hapnikuelektroodi abil saab mõõta selliste ensüümreaktsioonide kiirust, millede käigus tekib või reageerib O2. Hapnikuelektrood kuulub amperomeetriliste mõõteriistate hulka. Mõõdetakse hapniku redutseerimise tulemusel tekkiva redokselektronide voo tugevus ehk voolutugevust . See voolutugevus sõltub sellest, kui kiiresti difundeerub hapnik lahusest elektroodile. Difusioonikiirus on võrdeliselt seotud hapniku kontsentratsiooniga lahuses. Tsentraalne plaatina katood, mida ümbritseb Ag/AgCl anood. Nende vahele rakendatakse konstantne pinge (0.6V). Elektroodid on eraldatud reaktsiooni toimumiskohast õhukese (teflonist) membraani abil. See membraan on läbitav ainult hapnikule. Mõõdetakse voolutugevust, mis on määratud O2 difusiooniga katoodile. Difusiooni kiirus on võrdeliselt sõltuv hapniku kontsentratsioonist.
Kromatograafia ja ekstraktsioon:
Produkti ja substraadi lahutamismeetodid:
- Sadestamist kasutatakse reaktsioonides, millede käigus toimub makromolekulide (DNA, RNA, polüpeptiidide, polüsahhariidide) süntees madalmolekulaarsetest radioaktiivmärgitud substraatidest. Polümeerid on reeglina väiksema lahustuvusega kui madalmolekulaarsed ained. Näiteks saab polüpeptiide eraldada aminohapetest etanooli, triklooräädikhappe või ammooniumsulfaadiga sadestamise teel.
- Ekstraktsiooni kasutatakse juhtudel, kui substraadi ja produkti jaotus kahe omavahel mitteseguneva solvendi vahel on väga erinev.
- Produkti adsorbeerimine tahkele kandjale nagu paber, klaas, ioonvahetuskandjad.
- Kromatograafilised lahutusmeetodid :
- lahutus võib põhineda substraadi ja produkti erineval ,
- molekulmassil (geelfiltratsioon),
- hüdrofoobsusel (pööratud faasi kromatograafia),
- ioniseeritusel (ioonvahetuskromatograafia) ,
- adsorptsioonil (adsorptsioon-kromatograafia).
HPLC ehk kõrgsurve vedelikkromatograafia, paberkromatograafia,
kromatograafia õhukesel kihil.
Radioaktiivsuse kasutamine ensüümreaktsioonide jälgimiseks:
Aktiivsuse määramise radioaktiivselt märgistatud substraadiga:
Ensümaatiline reaktsioon:
Reaktsiooni peatamine ning P* ja S* lahutamine (kromatograafia, sadestamine, ekstraktsioon jt.)
Radioaktiivsuse mõõtmine ja produkti hulga arvutamine
Eelised:
- Kõige tundlikum meetod, võimalik on määrata produkti koguses10-18-10-17 mooli (umbes 6-7 suurusjärku tundlikum kui spektrofotomeetria). Seetõttu kasutatakse laialdaselt kliinilistes analüüsides, farmakoloogias ja rakubioloogias .
- Aktiivsuse määramiseks pole oluline ensüümi eelnev puhastamine.
- Universaalne, rakendatav kõikide ensüümide jaoks.
Ensüümiaktiivsuse määramisel kasutatakse peamiselt
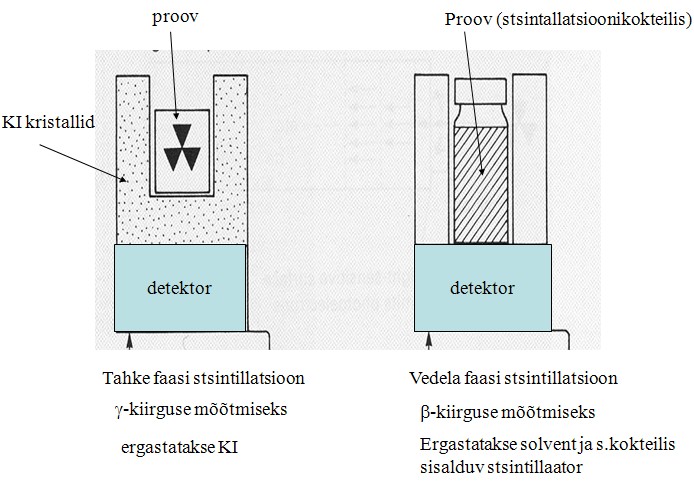
stsintillatsioonimeetodit. Stsintillatsioonimeetodi põhimõte:
kasutatakse radioaktiivset substraati, substraat ja produkt on
teineteisest kergesti eraldatavad (näiteks ekstraktsioon),
Radioaktiivsel lagunemisel tekkivad osakesed ( γ või β
–osakesed) põrkuvad läheduses asuvate molekulidega ja
ergastavad neid, ergastatud molekulid kiirgavad valguskvandi (fluorestsents), mis registreeritakse fotokordisti vooluimpulssidena,
stsintillatsiooniloenduritega mõõdetakse radioaktiivsust cpm
ühikutes (counts per minute ), reaktsioonis tekkiva produkti hulga
arvutamiseks tuleb eelnevalt määrata substraadi eriradioaktiivsus
(cpm/ mool ).
4.
Ensüümkatalüüsi regulatsioon:
Ensümaatilise reaktsiooni kiirus sõltub ensüümi
kontsentratsioonist ja ensüümi katalüütilisest aktiivsusest:
Igas metaboolses rajas on regulatoorne ensüüm ehk võtmeensüüm,
mille poolt katalüüsitava reaktsiooni kiirus määrab kogu
metaboolse raja kiiruse.
Võtmeensüümi poolt katalüüsitav reaktsioon toimub kõige
“aeglasemalt”, seetõttu limiteerib selle kiirus kogu raja
toimimist. Metaboolse raja võtmeensüümi hulga ja aktiivsuse kaudu
reguleeritakse kogu metaboolse raja kiirus. Selliste võtmeensüümide
poolt katalüüsitavate protsesside statsionaarne olek on kaugel
nende tasakaaluolekust.
Ensüümivalgu hulka mõjutavad protsessid:
Biosüntees:
Degradatsioon :
Pikema elueaga ensüümid lagundatakse reeglina lüsosoomis. lühema
elueaga ensüümid, mis on sageli regulatoorse funktsiooniga,
lagundatakse proteasoomis.
Ensüümide aktiivsuse regulatsioon:
- Kovalentne modifitseerimine
- Pöörduv (fosforüülimine)
- Mittepöörduv (limiteeritud proteolüüs, metüleerimine)
- Mittekovalentsed regulatsioonimehhanismid
- Spetsiifilised aktovaatorid-inhbiitorid
- Produktpidurdus
- Keskonnatundlikud konformatsioonimuutused
- Kooperatiivsus ja allosteerika
- Kompartmelisatsioon
- Iga ensüüm paikneb kindlas raku piirkonnas või struktuuris
- See tagab ensüümreaktsioonide eraldatuse ja kindla järjestuse
- Substraadi ja kofaktorite kontsentratsioon
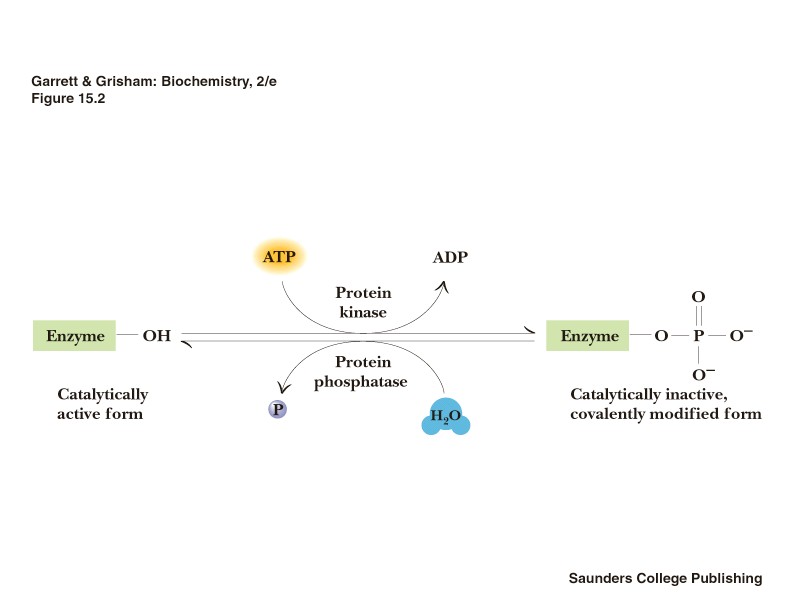
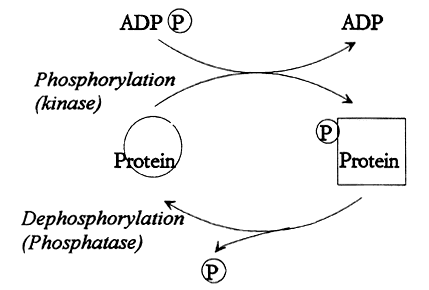
Kovalentne modifitseerimine:
Fosforüleerimine(pöörduv):
- Fosfaatrühma liitumist ensüümi türosiini-, seriini - või treoniinijäägiga katalüüsivad proteiinkinaasid
- Fosfaatrühma allikaks on ATP, cAMP , GTP
- Defosforüülimisreaktsiooni katalüüsivad fosfataasid
- Inimese genoomis on teada 575 proteiinkinaasi kodeerivat geeni
- Fosforüülimisreaktsioonid toimuvad kõikides elusorganismides
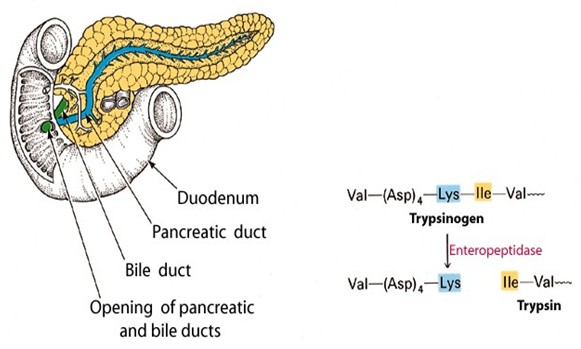
Limiteeritud proteolüüs:
Ensüümi polüpeptiidahela teatud peptiidsideme(te) hüdrolüüs
spetsiifilise proteaasi toimel aktiveerib ensüümi. Näit. Pankrease proteaaside aktivatsioon kaksteistsõrmiksooles (duodenum). Pankrease
ensüümid on pankrease nõres inaktiivsete proensüümide ehk
sümogeenidena. Ensüümide aktivatsioon toimub alles
kaksteistsõrmiksooles, see algab trüpsinogeeni (sümogeeni)
muundamisega aktiivseks trüpsiiniks enteropeptidaasi toimel. Selles
reaktsioonis ‘lõigatakse’ trüpsinogeeni N-terminaalsest osast
ära heksapeptiid. Tekkinud trüpsiin aktiveerib teised pankrease
ensüümid.
Glükosüleerimine (sahhariidide liitmine):
Peaaegu kõik ekstratsellulaarsed ensüümid on glükosüleeritud.
Oligosahhariidiahelad liidetakse ensüümi asparagiinile, treoniinile
või seriinile. Oligosahhariidiahelad liidetakse ensüümi
asparagiinile, treoniinile või seriinile. Oligosahhariidiahelad ei
osale otseselt katalüütilistes reaktsioonides, kuid stabiliseerivad
ensüüme ning on olulised nende kokkupakkimisel ja transpordis .
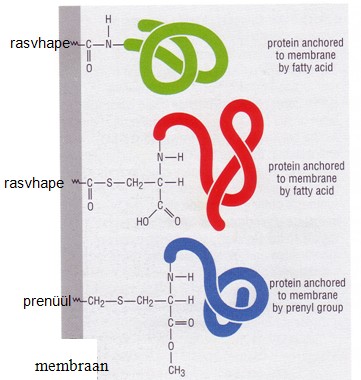
Lipideerimine:
Lipideerimine on rasvhapete ja teiste lipiidide liitmine
valgumolekuliga. lipideeritud ensüümid paiknevad reeglina
membraanides.
Mittekovalentsed regulatsioonimehhanismid:
Aktivaatorid ja inhibiitorid:
Aktivaatorid:
Inhibiitorid:
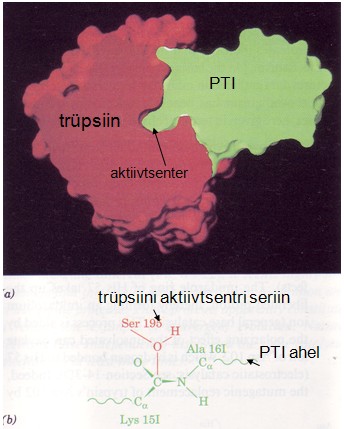
Paljude proteaaside aktiivsust inhibeerivad valgulised inhibiitorid.
Trüpsiini aktiivsust alandab pankrease trüpsiini inhibiitor
(PTI), mis kuulub pankrease nõre koostisesse. Selle valgu olemasolu
väldib trüpsiini aktiveerumise pankreases. Antitrüpsiin moodustab
trüpsiiniga väga tugeva kompleksi (Kd = 10-13M).
Trüpsiini aktiivtsentrisse seostub PTI vaheolek, kus Lys15.
Sarnaselt substraadiga moodustub tetraeedriline trüpsiini
aktiivtsentri Ser195 on seotud karbonüülse süsinikuga. Kuid see
kompleks ei lagune produktideks ,sest trüpsiin ja PTI on väga
jäigalt seotud ning veemolekulid ei pääse reaktsioonitsentrisse.
Produktinhibeerimine:
Reaktsioonil tekkiv produkt on sama reaktsiooni inhibiitoriks, see on väga levinud regulatsioonimehhanism.
Metaboolse raja vahe- või lõpp-produkt inhibeerib võtmeensüümi aktiivsuse.
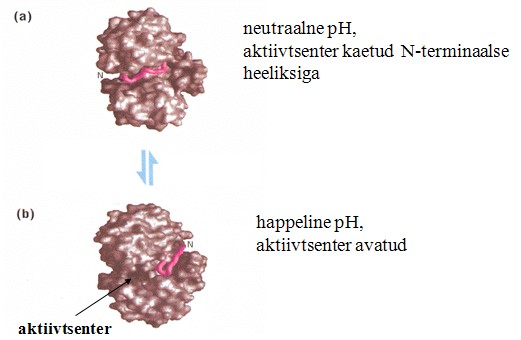
Keskkonnast sõltuvad konformatsioonilised muutused:
Näit. Katepsiin ( proteaas ) on aktiivne endosoomis, kus pH happeline
ja inaktiivne tsütoplasmas, kus pH on neutraalne.
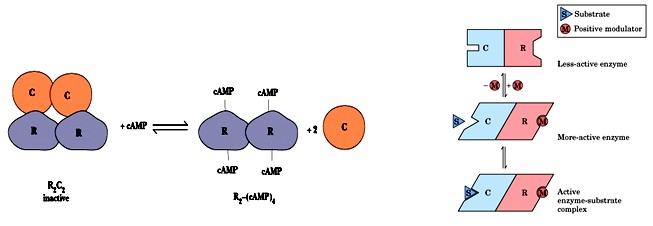
Modulaatorvalgud on regulatiivsed subühikud, mis võivad ensüümi
aktiveerida või inhibeerida tingimustest sõltuvalt. cAMP
proteiinkinaas on tetrameerina inaktiivne.
cAMP seostumisel regulatoorsed subühikud dissotsieeruvad ja
ensüüm muutub aktiivseks. Paremal on kujutatud reaktsiooni
hüpoteetilise aktiveeriva modulaatoriga.
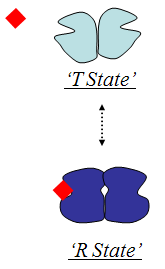
Kooperatiivsus:
Kooperatiivne substraatide sidumine:
Allosteerika ja regulatsioon:
Allos – teine; steric – kuju.
- Raku elutegevuse käigus tuleb kõigi ensüümide aktiivsused hoida pideva kontrolli all. Seda mitte ainult aktiivsete protsesside teostamiseks vaid ka homeostaasi säilitamiseks.
- Tagasiside - paljudest ensümaatilistest protsessidest koosneva metaboolse raja üks ensüüm (regulatoorne) on tundlik inhibeerimisele raja lõpp-produkti poolt.
- Allosteerika. Allosteerilised ensüümid:
- Muudavad oma konformatsiooni modulaatori sidumisel
- Lisaks katalüütilistele subühikutele võivad sisaldada ka spetsiaalseid regulatoorseid subühikuid
- On tavaliselt suuremad ja keerukamad kui MM kineetikale vastavalt käituvad ensüümid
Allosteerilised ensüümid:
- Kui aktiivsuse modulaatoriks on substraat, siis on regulatsioon homotroopne, kui mõni muu ühend, siis heterotroopne.
- Jagunevad K ja V tüüpi ensüümideks (paljud on segatüüpi).
- K tüüpi allosteeriliste ensüümide kineetika ei allu MM võrrandile - v sõltuvust [S]st ei iseloomusta hüperboolne vaid S-kujuline (sigmoidne) sõltuvus. Substraadi kontsentratsiooni, kus kiirus on pool maksimaalsest nimetatakse K0.5 või KS, aga mitte Km.
- Sigmoidse substraatsõltuvuse põhjustab kooperatiivsus subühikute vahel:
- Modulaatori seostumine ühele subühikule põhjustab konformatsioonimuutusi teistes subühikutes
- Konformatsiooni muutus ühes subühikus põhjustab muutusi teistes subühikutes
Näited: hemoglobiin (mis ei ole küll ensüüm) ja aspartaadi transkarbamülaas (ATCase).
Rõhuv enamus allosteerilisi ensüüme ilmutavad substraadi sidumisel
positiivset kooperatiivsust. Võrreldes sarnase mitte-allosteerilise
ensüümidega on nende aktiivsus madalatel substraadi
kontsentratsioonidel madalam. S-kujuline substraatsõltuvus on
erinevate ensüümide korral erinev - võib toimuda järsk tõus väga kitsas kontsentratsioonivahemikus.
5) Ensümaatiliste reaktsioonide kineetika:
Michaelis- Menteni võrrand ja kineetilised parameetrid.
MM pakkusid välja reaktsiooni skeemi:
k1 kcat
E + S ES ===> P + E
k-1
, mille tähtsaimaks eripäraks on mittekovalentse
ensüüm-substraat kompleksi, (ES),
teke.
k1
on ESi moodustumise kiiruskonstant (v = k1[E][S])
k-1
on ESi dissotsiatsiooni kiiruskonstant (v = k-1[ES])
Reaktsiooni kiirus on:
2 peamist lihtsustust:
Esiteks eeldame, et k-1
on suurem kui kcat nii et ensüüm, substraat ja ensüüm-substraat kompleks on
omavahel tasakaalus.
Tasakaaluolekus on ES-kompleksi
moodustumise ja lagunemise kiirused võrdsed, järelikult:
k1[E][S] = k-1[ES]
Ehk :
kus KS
on defineeritud kui dissotsiatsiooni tasakaalukonstant .
Teiseks eeldatakse, et substraadi algkontsentratsioon,
[S]o,
on palju suurem kui lisatud ensüümi
kontsentratsioon, [E]tot,
mistõttu reaktsiooni algfaasis, kui substraati on produktiks
muundunud veel vähe, on substraadi kontsentratsioon lahuses võrdne
substraadi algkontsentratsiooniga (kuna ES
kompleksi läheb isegi tingimustel, kus ensüüm on substraadiga
küllastunud, täiesti ebaoluline kogus substraati):
i.e. [S]o
>> [E]tot ja [S] ≈ [S]o
Lisaks piirdume algkiiruse
arvutamisega, et teine eeldus paika peaks.
Mis juhtub, kui esimene eeldus (k-1
>> kcat
) ei kehti?
Siis ei ole ensüüm, substraat ja ES-kompleks tasakaalus ja MM
võrrandit ei saa selle originaalkujul kasutada. Tavaliselt
mõeldaksegi MM võrrandi kasutamise all hoopis selle võrrandi
kasutamist, mille tuletasid Briggs ja Haldane, kes eeldasid, et
enamuse reaktsiooni kulgemise ajast on ES statsionaarses
( steady -state) olekus, kus [ES] on praktiliselt konstantne.
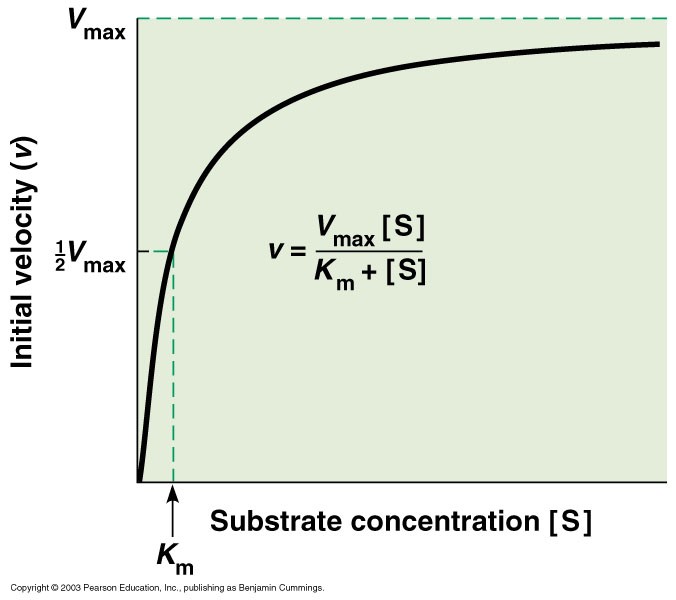
Michaelis-Menten’I võrrandi dualistlik
olemus:
- Kui [S] on madal (> Km), siis on kiirus [S] suhtes 0-järku
Vmax tähendus:
Maksimaalne teoreetiliselt
võimalik kiirus
- Vmax on konstant, mis sõltub lineaarselt ensüümi kogusest.
- Vmax on teoreetiline kiiruse ülempiir – reaalselt EI SAAVUTATA seda kunagi
- Vmax saavutamiseks peavad KÕIK ensüümi molekulid olema seotud substraadiga
- Vmax on piir millele kiirus läheneb asümptootiliselt substraadi kontsentratsiooni suurenedes
Km tähendus:
- Km on kiiruskonstantide kombinatsioon:
= (k-1+k2)/k1
- Km on ES kompleksi dissotsiatsioonikonstandi võimalikuks kõige väiksemaks väärtuseks (lihtsa kahestaadiumilise ensüümreaktsiooni korral), aga ta ei ole tasakaalukonstant, mis alati iseloomustaks sidumist.
kcat/Km
tähendus:
- kcat/Km on teist järku kiiruskonstant :
substraadi kadumine madalal substraadi kontsentratsioonil on
pseudo-esimest-järku protsess, mille kiiruskonstant sõltub ensüümi
kontsentratsioonist
- kcat/Km on ensüümi katalüütilise efektiivsuse mõõduks, kuna näitab vaba S produktiks konverteerimise kiirust
- kcat/Km logaritmimisel saame arvutada aktivatsiooni vabaenergia reaktsiooni jaoks, mis lähtub vabast substraadist ja ensüümist lahuses
- kcat/Km ülempiir on määratud difusioonikiirusega
pH ja temperatuuri mõju aktiivsusele.
Ensüümreaktsiooni kiirust (ensüümi aktiivsust) mõjutavad faktorid :
- Substraadi kontsentratsioon
- Ensüümi regulatsioon
- Temperatuur
- pH
- Puhvrid , soolad etc.
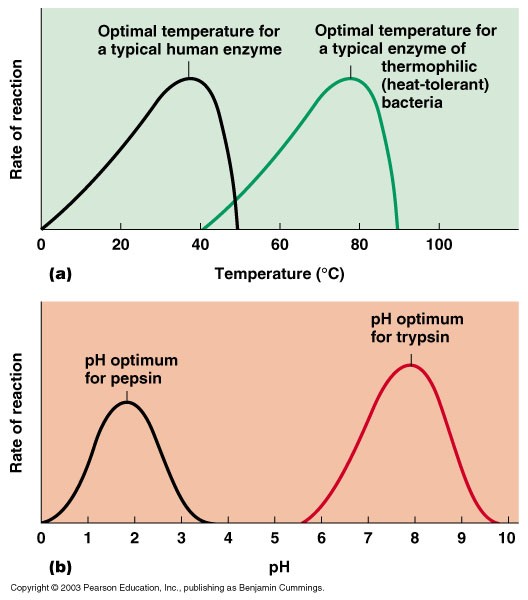
Temperatuuri mõju keemilise reaktsiooni kiirusele:
- Kiirus kasvab temperatuuri tõustes.
- Arrheniuse võrrand
Ensümaatiliste reaktsioonide sõltuvus temperatuurist - kiirus sõltub toimuva keemilise reaktsiooni
aktivatsioonienergiast ja ( teiselt poolt) ensüümi struktuuri koos
hoidvate nõrkade vastasmõjude stabiilsusest. Optimum on tavaliselt
veidi kõrgem kui ensüümi loodusliku keskkonna tavaline
temperatuur.
pH mõjutab reaktsiooni kiirust muutes
aktiivtsentri aminohappejääkide protonisatsiooniastet. Optimum
vastab tavaliselt ensüümi natiivse keskkonna pHle.
pH sõltuvus:
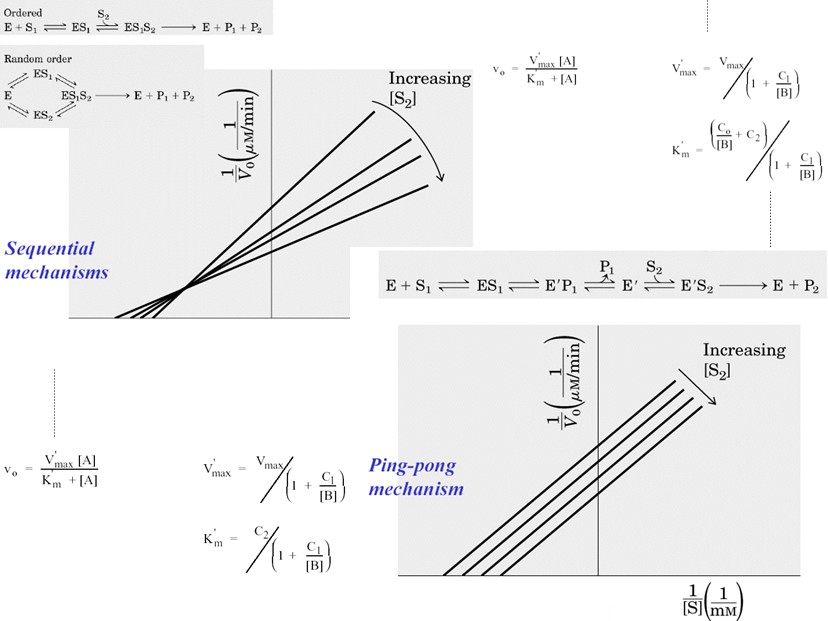
Multisubstraatsed reaktsioonid – korrastatud ja juhuslik ning ping - pong mehhanism.
Ensüümreaktsioonide liigitus substraatide ja produktide arvu järgi:
Järjestikuline rekatsioon kahe substraadiga:
Järjestikuse
mehhanismi järgi toimuva ensümaatilise reaktsiooni korral peavad
mõlemad substraadid seostuma ensüümiga enne reaktsiooni toimumist
- reaktsioon kahe substraadiga võib toimuda korrastatud või juhusliku mehhanismi järgi.
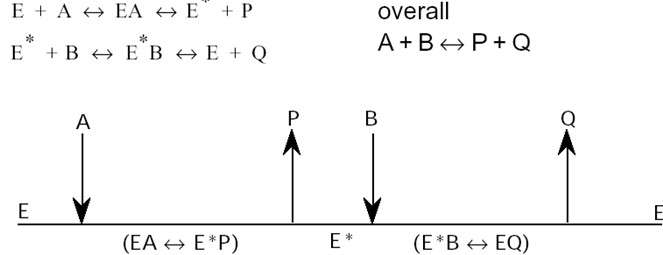
Korrastatud järjestikune mehhanism:
Nii substraatide seostumine kui ka produktide dissotsiatsioon
toimuvad kindlas järjekorras.
Substraatide tähiseks on A, B C jne; produktil vastavalt P, Q, R
jne.
Clelandi meetodil kirjutatakse reaktsioonid mitme substraadiga üles
järgnevalt:
Esimesena seostub ensüümiga NAD+, mis kutsub esile ensüümi
konformatsiooni muutuse ja afiinsuse suurenemise laktaadi suhtes
Juhuslik
bi-bi mehhanism:
Järjestikuse mehhanismi alusel toimuva kahesubstraatse
reaktsiooni kineetika:
Kiiruse üldine avaldis nii korrastatud kui juhusliku mehhanismi
jaoks on võimalik tuletada nii tasakaalu kui statsionaarset olekut
eeldades:
Ping-pong mehhanism:
Puudub kompleks EAB - kõigepealt reageerib ensüümiga esimene
substraat ja peale esimese produkti dissotsiatsiooni reageerib
(modifitseerunud) ensüümiga teine substraat. Ping-pong mehhanismi
eripäraks on kovalentse vaheühendi teke - teise substraadi
puudumisel on võimalik see “kinni püüda” ja määrata näiteks
radioaktiivse märgise abil. (N: tranamidaasid)
Teiseks substraadiks on antud juhul vesi ja reaktsiooni kineetikat
kirjeldab vesilahuses lihtne MM võrrand. “Katseklaasis” on
võimalik vee asemel kasutada alternatiivseid “teisi substraate”,
mis avab võimaluse kasutada seriinhüdrolaase katalüsaatorina
vajalike ühendite sünteesil. Eriti laia rakendust on leidnud bakteriaalsed lipaasid.
Reaktsioonid kahe substraadiga:
- Alluvad MM kineetikale kui varieerida ühe substraadi kontsentratsiooni
- Kui varieerida ühe substraadi kontsentratsiooni ja hoida teist konstantsena, siis saab substraatsõltuvuste kuju järgi eristada järjestikust ja ping-pong mehhanismi
- Enamus sidustatud reaktsioone on kahe substraadi reaktsioonid.
6) Ensüümkatalüüsi uurimismeetodid:
- Modelleerimine –
- Modifitseerimine - Disainitakse ühend, mis sarnaneb substraadiga ja on võimeline reageerima oletatavate aktiivsete rühmadega. Inkubeeritakse ensüümi reagendiga. Määratakse kindlaks, milline aminohappejääk modifitseerus. Substraat peab pidurdama inhibeerimise kiirust!
- Mutatsioonid – punktmutatsioon, on kaasaegne meetod ensüümi aktiivtsentri ja reaktsioonimehhanismi uurimiseks. Eelistada tuleb konservatiivseid asendusi Glu, Asp - Gln, Asn; Ser - Ala jne..
Näide: Asp102 rolli seriinproteaasides eeldati tema läheduse tõttu
His57le. W. Rutter valmistas trüpsiini, kus Asp102 oli asendatud asparagiiniga - mutandi aktiivsus oli 10 000 korda madalam kui
natiivsel ensüümil. Mis juhtub kui muteerida kogu katalüütiline triaad ? “Jääkensüüm” ilmutab ikkagi mõningast
katalüüsivõimet - sidumisvõime ja aktiveeritud olekut
solvateerivad rühmad jäid ju alles.
- keskkonnaefektid,
- substraadi struktuuri mõju,
- inhibiitoranalüüs.
- Ensüümkatalüüsi teooriad.
- Ensüümide spetsiifilisust ja katalüütilist aktiivsust määravad tegurid.
- Ensüümide regulatsioon ja multiensüümsed kompleksid metabolismis: regulatsiooni erinevad tasemed - allosteerika, kofaktorid, post-translatoorne modifitseerimine.
- Ensüümide biomeditsiinilised rakendused – paljud ensüümid on higuste markeriteks nt. kreatiini kinaas.
Ravimid kui ensüümide inhibiitorid:
- Mõned antibiootikumid on pöördumatud inhibiitorid. Näiteks penitsilliin , mis on inhibiitoriks bakteri membraani terviklikkust tagavale transpeptidaasile.
- Seostuvad kovalentselt
- Väga reaktiivsed ja võivad seostuda ka mitte-märklaud rakkudega
- Osad AB on pöörduvad inhibiitorid. Näiteks sulfaniilamiid, mis inhibeerib foolhappe sünteesi. Foolhape on vajalik mõningate mikroobide kasvuks.
- Nad on konkureerivad inhibiitorid ja seostuvad mittekovalentselt
- Vähem kõrvaltoimeid kuid nende toime on lühiajalisem
Ensüüm Patoloogiline seisund
Alaniini aminotransferaas Hepatiit
Aspartaadi aminotransferaas Infarkt ja hepatiit
Alkaalne fosfataas Maksa- ja luuhaigused
Kreatiini kinaas Südame infarkt
Amülaas Kõhunäärmehaigused
Happeline fosfataas Eesnäärmevähk
- Proteaasid - Esinevad naturaalselt kõikides organismides. Katalüüsivad peptiidide ja valkude peptiidsidemete hüdrolüüsi. Seetõttu kutsutakse neid ka proteaasideks. Proteaasidel osalevad toiduvalkude lagundamises, vere koagulatsioonis, mitmete teiste ensüümide aktiivsuse regulatsioonis. Kasutatakse paljudes pesupesemisvahendites, leivatööstuses. Peptidaasid võivad lõhkuda:
- spetsiifilist peptiidsidet (limited proteolysis), mis
sõltub valgu aminohappelisest järjestusest või
- kogu peptiidi aminohapeteks
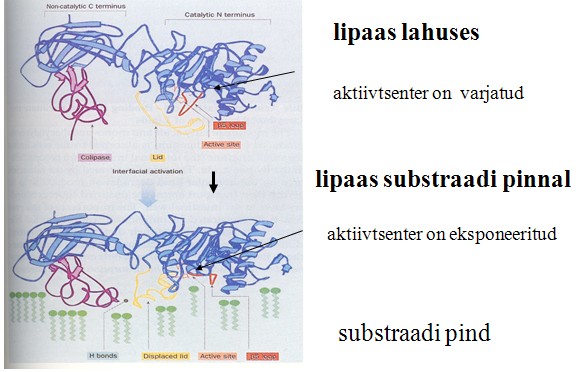
Lipaasid toimivad substraadi-agregaadi (rasvatilga) pinnal























































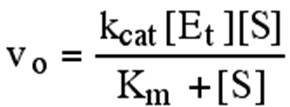
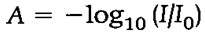

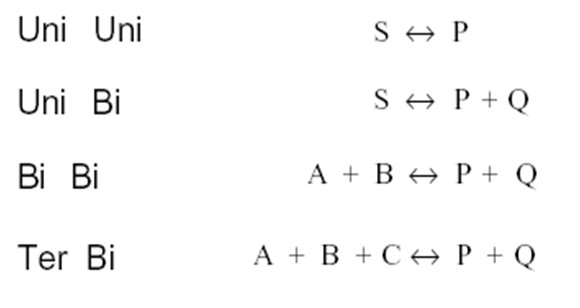



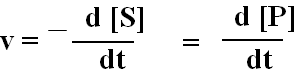


































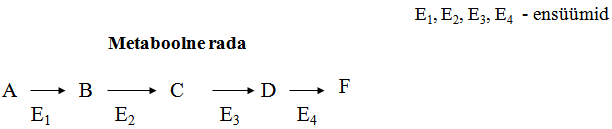







Kõik kommentaarid