32
“Valgu biosüntees 2012” loengute
põhipunktidValgusünteesi uurimise meetodidRakuvaba valgusüntees
- Milliseid komponente peab sisaldama rakuvaba valgusünteesi ekstrakt (mis peab ekstraktis olema ja mida tuleb lisada) On olemas ribosoomid ja translatsiooni faktorid . Rakud lüüsitakse DNaasi juuresolekul, tsentrifuugitakse ja dialüüsitakse. Lisatakse: mRNA või DNA + RNAP (NTP), Aminohapped , ATP, GTP ja energia regeneratsiooni süsteem (PEP/PK, AcP/AK, KrP/KP) S12 puhul ka glükoos vms energia allikas, SH reagent (DTT, 2-ME), Mg2+, K+, Na või NH4+
- Mille poolest erinevad bakterite ja eukarüootide rakuvaba valgusünteesi ekstraktid
Bakterite lüsaadid saavad
produtseerida vaid teatud komplekssusega valke, kuna puuduvad
võimalused teatud protsessimisteks ja voltumisteks. Samas kõige
kiiremini kasvatatav ning kõige kuluefektiivsem.
- Küüliku retikulotsüüdi ekstrakt Küüliku retikulotsüütide lüsaat valmistatakse aneemiliste küülikute retikulotsüütidest ja dialüüsitakse. Lisatakse AH, ATP, GTP ja energia regeneratsiooni süsteem.Sünteesitavaid valke analüüsitakse geelelektroforeesiga.
- Kuidas teha rakuvaba valgusünteesi süsteem sõltuvaks lisatavast mRNAst
Tehes lüsaadile nagu
restardi, paned lahusesse Ca-sõltuva nukleaasi (RNaas?), sellega
vabaned üksnes endogeensest mRNA-st (mitte aga rRNA-st), siis lisad
EGTA, mis aktiveerib lüsaadi.
- Millised on fraktsioneeritud valgusünteesi süsteemid ja mida need sisaldavad. Bakteriaalsed ribosoomid, mRNA, tRNA, S-100 valgufraktsioon, Foolhape (tetrahüdrofolaat), aminohapped, ATP, GTP, energia regeneratsioon SH reagent, Mg2+, K+ või NH4+
- DNA sõltuvad valgusünteesi süsteemid DNA suunatud valgusüntees. Seda süsteemi saab teha nii S12, S30 kui fraktsioneeritud süsteemi baasil. S30 puhul võib kasutada endogeenset RNA polümeraasi. Süsteem sisaldab lisaks eelnevale ka UTP, CTP, DNA matriitsi ja vajadusel sobivat RNA polümeraasi (T7, T5, SP6, E. coli RNAP)
- Geneetilise koodi laiendamine in vitro ja in vivo . In vitro. kasutatakse mingit UAG supressor tRNA-d, mille viimased 2 nukleotiidi puudu on (xxxcca-->xxxc). Sellele lisatakse mingi kunstlik CA(3’O) ( aminohape ?) ja tulemuseks ongi kunstlik aa-tRNA ja siis tuleb see lisada lihtsalt valkudesse.
In vivo: geneetilise koodi muutumine toimub
väikeste genoomide puhul, kui kõiki koodoneid ei kasutata.
Kasutamata koodonitele saab määrata uue funktsiooni ilma et see
valgusünteesi häiriks)
- Rakuvaba valgusünteesi rakendused . Valgud , mida on raske ekspresseerida in vivo (tsütotoksilised, ebastabiilsed). Valkude märkimine stabiilsete või radioaktiivsete isotoopidega, spin märke või fotoaktiveeritava rühmaga. Geneetilise koodi laiendamine. In vitro evolutsioonilised lähenemised .
Valgusünteesi uurimine rakkudes - Ribosoomide profileerimise meetod, millel põhineb ja kuidas toimib, mida selle meetodiga on võimalik teada saada. Süvasekventsiga määratakse ribosoomidega seotud mRNA osad, mis näitab translatsioooni dünaamikat mRNA-l. Kaitstud fragmentide sagedus peegeldab konkreetse piirkonna translatsiooni kiirust. Rakkudest eraldatakse polüsoomid. Ribosoome töödeldakse ribonukleaasiga. Tekivad 36-40 nt pikkused fragmendid , mis suunatakse sekveneerimisse. Kõik sobivate pikkustega fragmentide järjestused saadakse teada ja paigutakse genoomidesse. Vaadates palju sarnaseid fragmente on, saame määrata nende aktiivsuse tase. Mida rohkem ühte lõiku, seda aeglasem on sealkohal translatsioon . Saame ribosoomide profiili mRNA-de peal. Töödeldes polüsoome nukleaasiga, muutuvad nad kõik 70S-ks. Kui mRNA sisaldab Shine-Dalgamo järjestsutega sarnased, siis jääb sel kohal translatsioon seisma, katkeb.
Füüsikaline meetod, mis
võimaldab väga täpselt määrata mingisuguste osakeste massi ja
laengust (selle suhet). Nt peptiidide hulka segus. Põhineb
stabiilsete isotoopidega märkimisel. Kasutatakse kahte valkude segu
(standart ja uuritav). Märgitakse nt metaboolselt. Raskete ja
kergete
ahelate suhe, leiame keskmise. Leiame valgu suhte nendes
kahes proovis. Sarnast märkimist võib teha kasutades AH keemilisi
omadusi (
aminorühm , millel eriline
reaktsioon mingi kindla ainega,
mida teistel AH pole). Ensümaatiline märkimine (karboksüterminaalse
AH märkimine mingi kemikaaliga).
- SILAC märkimine, pSILAC valgusünteesi dünaamikma uurimisel. SILAC Valgud märgitakse stabiilsete isotoobidega. Rakukultuuris märgitakse valgud lisades söötmele raskeid aminohappeid . Harilikult lüsiin, arginiin või leutsiin, mis sisaldavad 2H, 13C või 15N aatomeid. Valgud lõigatakse peptiidideks ja peptiidid eraldatakse kromatograafiliselt. Kindla väärtusega m/z ioonide kvantitatiivne suhe näitab peptiidide ja seega ka valkude suhet. Trüpsiin lõikab valguahelat positiivselt laetud AH (R, K) C-terminaalselt. LysC lõikab vaid lüsiini tagant. Kümotrüpsiin lõikab aromaatsete AH (F, Y, W) tagant. Kasutades massi märkega ühte neist AH-st, saab kindla massnihkega peptiide.pSILAC. Pulss märkimisega SILAC. Rakkudele lisatakse erineva märkega aminohappeid (söötmevahetus). Jälgitakse kahe või enama märkega peptiidide suhet LQC/MS abil. Võimaldab analüüsida paljude valkude sünteesi kineetikat samades rakkudes. Näide: hiire fibroplastide kultuur – globaalne geeniekspressiooni regulatsioon . mRNA ja valkude eluead ning koopiaarvud.
Aminoatsüül-tRNA süntetaasid - Aminoatsüül-tRNA süntetaaside jaotus kahte klassi. aaRS-id jagatakse kahte klassi (I ja II) ja kummassegi klassi kuulub 10 liiget. Klassidesse jagamise aluseks on konseveerunud ah järjestused ja iseloomulikud struktuursed motiivid. Erinevused järjestustes annavad tulemuseks aktiivsaidi erineva 3D struktuuri ja erineva tRNA ja aaRS-i molekulide stereomeetrilise seondamise. Klassid jaotakse omakorda kolme alamklassi nende homoloogia ja domäänide struktuuri alusel. Klass I süntetaaside katalüütiline sait sisaldab Rossmanni volti (beeta-paralleelne kiht). Seal leidub kaks konserveerunud järjestusmotiivi- HIGH(esimene alfaheeliks) ja KMSKS(kujutab endast lingu , mille konformatsiooni muutus substraadi seondamisel aitab aktiveerida ah-t. Klass II aaRS aktiivtsenter on suurem, koosnev antiparalleelsest beeta-kihtidest, mis on ümbritsetus alfa-heeliksitega. Aktiivtsentris eristatakse kol motiivi: ATP sidumiseks, ah sidumiseks ja dimerisatsiooniks. Klass I : on mono - või dimeerid, ATP konformatsioon on venitatud, aminoatsüleerimissait: tRNA terminaalse adenosiin 2´-OH. Klass II on di- või tetrameerid, ATP konformatsioon painutatud, aasait on tRNA terminaalse adenosiin 3´-OH
- Aminoatsüleerimisreaktsioon ja selle erandjuhud. Aminoatsüleerimiseks nimetatakse valgusünteesi etappi , mille käigus liidetakse ah vastava tRNA terminaalse adenosiini 3´-OH või 2´-OH külge. Üldjuhul toimub ah liitmine tRNA molekulile kaheastmelise reaktsioonina. (1) aaRS + ATP:Mg2+ + aa = aaRS:aa:AMP + PPi:Mg2+. Seda esimeset etappi nimetatakse ka ah aktiveerimiseks. Süntetaas omab kolme seostumissaiti – tRNA, ah, ja energiaallika ATP jaoks. Enamasti senduvad vesiniksideme abil ensüümiga esmalt ah ja ATP. Mõned klass I süntetaasid vajavad ah aktivatsiooniks tRNA juuresolekut, mis muudab ensüümi konformatsiooni nii, et suureneb süntetaasi afiinsus ah suhtes. Ah karboksüülrühma hapnik reageerib ATP alfa-fosfaadiga. Mg on aaRS-dele kofaktoriks, aidates muuhulgas stabiliseerida ATP konformatsiooni. Süntetaasi pinnal moodustub aminoatsüüladenülaat, mis kujutab endast aminohappest ja ATP-st koosnevat aktiveeritud intermediaati. Reaktsiooni käigus vabaneb pürofosfaat. (2) aaRS:aa:AMP + tRNA = aaRS + aa-tRNA + AMP. tRNA seondamine toimub tänu spetsiifiliste interaktsioonide süntetaasi ah ja tRNA nukleotiidide vahel. Teises etapis toimub tRNA 3´-terminaalse adenosiini 2´või 3´ süsiniku küljes oleva karboksüülrühma nukleofiilne atakk aminoatsüül-adenülaadi karboksüülrühmale. Selle tulemusna moodustub ah karboksüülrühma ja tRNA A76 vahel esterside . Nii modustub aa-tRNA ja vabaneb AMP. Nüüd seosub aminoatsüül-tRNA-ga EF-Tu, mis transpordib aminoatsüül-tRNA ribosoomi A-saiti. Kus ta käitub peptiidi aktseptorina. Erandjuhud. ND-GluRS- mitte-dikrimineeriv glutamüül-tRNA süntetaas(kui Gln süntetaas puudub, siis tRNA Gln saaab sünteesida ND Glu abil Kaks etappi. Esimeses sünteesitakse ND-GluRs vahendusel Glu-tRNA glu-tRNAGln, järgnevalt konventeeritakse Glu-tRNAgln ensüümi amidotransferaasi abil Gln-trnagln-ks reaktsiooni vajatakse ATP-d. Sünteesib nii glu kui gln. ND-AspRS-sünteesib nii Asp+tRNAasp kui ka Asp-tRNAAsn. SepRS-Fosfoserüül-tRNA süntetaas osaleb Cys-tRNAcys sünteesil( liidab esialgul fosfoseriini, seejärel transformeeritakse see tsüsteiiniks. Selle reaktsiooni viib läbi SepCysS, kofaktoriks on püridoksaal fosfaat PLP. PylRS- pürrolüsüül-tRna süntetaas.
- Aminoatsüül-tRNA süntetaaside editeerimismehhanismid. See on aa-RS hüdrolüütiline aktiivsus, mille abil välditakse vale ah laetud tRNA vabanemist. Võib jagada kaheks. Ülekande- eelne ja ülekande järgne. Ülekande-eelne editeerimine toimub aminoatsüül-adenülaadi tasemel. Kontrollitakse aminoatsüleerimisreaktsiooni esimest etappi. Kui süntetaasi pinnal moodustub vale aminoatsüüladenülaat, siis see hüdrolüüsitakse (või hüdrolüüsub ebastabiilsuse tõttu ise) enne estersideme tekkimist aminohappe ja tRNA vahel. Reaktsiooni toimumiskohaks on enamasti ensüümi aktiivsait või eraldi editeerimissait. Reaktsiooni tulemusena vabanevad aminohape ja AMP. Ülekande-järgsest editeerimisest räägitakse siis, kui esterside vale aminoatsüül-adenülaadi ja tRNA vahele on juba moodustunud.
Ülekande-järgne editeerimine liigitatakse
omakorda:
- editeerimine in cis – toimub enne valesti laetud aa-tRNA vabanemist
- editeerimine in trans – toimub pärast aa-tRNA vabanemist
In cis
editeerimise puhul toimub estersideme
hüdrolüüs enamasti
süntetaasi aminoatsüleerimissaidist eemal spetsiaalses
editeerimissaidis. Klass Ia aaRS-del on selleks CP1 domään.
Klass II ensüümide editeerimisdomäänid pole üldiselt
konserveerunud ja ei sarnane CP1-ga.
Ülekande-järgset editeerimist
in
trans viivad läbi kas
spetsiifilised hüdrolaasid või süntetaasid ise. Ka EF-Tu võib selles protsessis
osaleda. Enamasti translokeerub Tyr-tRNAPhe
3’ ots koheselt peale moodustumist aaRS editeerimissaiti (
in
cis editeerimine). Kui aga Tyr-tRNAPhe
vabaneb, konkureerivad tema pärast aaRS ise ja EF-Tu. Kuna EF-Tu
afiinsus valesti laetud tRNA suhtes on väike,
seondub Tyr-tRNAPhe
uuesti PheRS-ga nii et Tyr satub otse editeerimissaiti (
in
trans editeerimine). Selline
mehhanism on üks võimalus, kuidas valesti aminoatsüleeritud tRNA
translokaliseerub aminoatsüleerimissaidist editeerimissaiti.
In trans
editeerimist võivad läbi viia erilised valgud, mis omavad
hüdrolüütilist aktiivsust. Siin on näiteks valk AlaX, mis
deatsüleerib Ser-tRNAAla
ja Gly-tRNAAla.
AlaX on leitud kõikidest eluslooduse riikidest. Asjaolu, et AlaRS
võib ekslikult sünteesida ka Ser-tRNAAla,
on üllatav, kuna seriin on alaniinist suurem ja teistele aaRS-dele
pole suuremate aminohapete
vältimine probleem. Ser-tRNAAla
süntees on tingitud seriini biokeemilisest eripärast, mistõttu ei
saa toimuda evolutsiooniliselt konserveerunud aaRS:aminohappe
interaktsioon . Seetõttu arvatakse, et AlaX, mis sarnaneb AlaRS
editeerimisdomääniga, on tekkinud samal ajal koos AlaRS-ga.
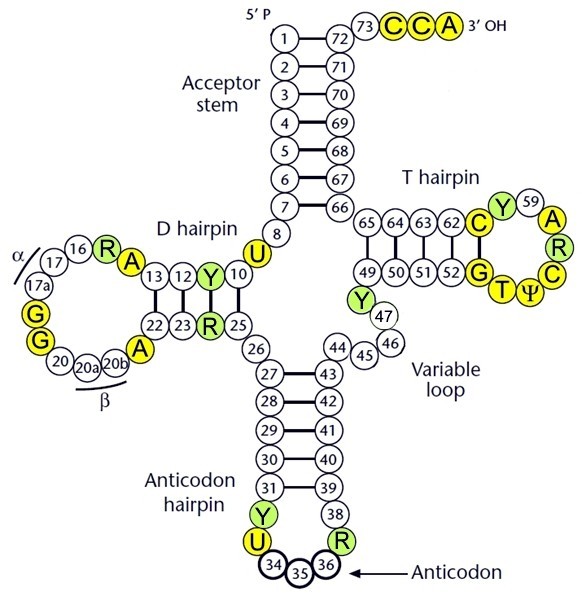
tRNA struktuur - tRNA sekundaarstruktuur. Kõikide tRNA molekulide sekundaar - ja tertsiaarstruktuur peab olema sarnane, et nad oleksid ära tuntavad elongatsioonifaktorite poolt ning et nad saaksid seonduda ribosoomiga. tRNA molekulidele on omane ristikheinalehe-kujuline sekundaarstruktuur. tRNA sekundaarstruktuuri moodustavad 4 kaksikahelalist osa, õlga (stem) ja 4 üksikahelalist piirkonda. Üksikahelalised regioonid - 3 lingu ( loop ) ja 4 paardumata nukleotiidi molekuli 3’ otsas - paiknevad vastavate õlgade tippudes. Kaksikahelalised osad on stabiliseeritud vesiniksidemete poolt. Heeliksite ja lingude pikkused on suurel määral konserveerunud.
tRNA molekuli
otsad asuvad lähestikku, nende
paardumisel tekkiv kaksikahelaline osa kannab nime aktseptoorne
õlg (acceptor arm).
Aktseptoorse õla 3’ otsas paikneb üheahelaline CCA76
järjestus, millele liidetakse aminohape. CCA järjestuse olemasolu
on tRNA’de universaalne omadus, sest kõikide tRNA’de 3’ ots
peab sobituma ribosoomi dekodeerivasse tsentrisse. Nukleotiidi
positsioonis 73 nimetatakse diskriminaatoriks, kuna see on oluline
tRNA äratundmisel aa-tRNA süntetaasi (aaRS) poolt. tRNA 5’
terminaalne
nukleotiid omab 5’-fosfaatgruppi. Aktseptorõla pikkus
on 7 bp (
erandiks on tRNAHis).
Aktseptorõla ja D-õla vahel on alati 2 nukleotiidi.
D-õla nimetus tuleneb sellest, et pea-aegu alati
esineb selles tRNA osas üks või mitu dihüdrouridiini (uridiini
pürimidiinring on küllastunud). Tegemist on
post-transkriptsioonilise modifikatsiooniga. D-õla pikkus on
enamasti 4 bp ning D-lingu suurus
varieerub 8-11 aluseni. D-õla ja
antikoodon -õla vahele jääb alati üks nukleotiid.
Antikoodon-õlg on alati 5
aluspaari pikk ja
sisaldab antikoodonit. Antikoodon lingus on alati 7 nukleotiidi.
Antikoodoni nukleotiidid 34, 35 ja 36 paarduvad vastavalt mRNA-l
paikneva koodoni 3. 2. ja 1. positsiooniga. Paljud tRNA molekulid
sisaldavad lisaõlga ja/või lisalingu, mille pikkus võib varieeruda
laias vahemikus (4-21 alust). Vastavalt lisalingu pikkusele
eristatakse klass I ja klass II tRNA-sid. Klass I tRNA-del on
lisalingus 4-5 nukleotiidi. Klass II tRNA-del paikneb samas regioonis
10-21 nukleotiidi, mistõttu sellistel tRNA-del moodustub viies
heeliks . Klass II tRNA-sid on vähe – nende hulka kuuluvad vaid
tRNALeu,
tRNASer
ja eubakterite ning organellide tRNATyr.
T-õla (tähistatakse ka TΨC õlg)
pikkuseks on 5
bp. 7 aluse
pikkune T-
ling sisaldab sageli
modifitseeritud lämmastikalust ribosüültümidiin (
tümiin paikneb
riboosi küljes;
nimetatakse ka 5-metüüluridiiniks, m5U).
Teiseks modifitseeritud nukleotiidiks selles piirkonnas on
pseudouridiin Ψ (uratsiil on liidetud riboosi külge C5 kaudu, mitte
tavapärase N1 kaudu).
tRNA molekuli pikkus varieerub tavaliselt 75-85
nukleotiidini, kuigi leidub ka
erandeid . tRNA nukleotiidid on
nummerdatud ühtse nomenklatuuri alusel: esimene nukleotiid on 5’
otsas, antikoodoni moodustavad nukleotiidid 34, 35, 36 ja 3’ otsa
konserveerunud järjestus CCA kannab numbreid 74...76 olenemata
sellest, mitu nukleotiidi konkreetses tRNA molekulis on. Näiteks
võivad lisalingus paikneda nukleotiidid 47, 47:1, 47:2 jne (või
47A, 47B, 47C jne).
- tRNA tertsiaarstruktuuri moodustumine ja mis seda stabiliseerib. Üheks tRNA molekuli ruumilist struktuuri stabiliseerivaks elemendiks on lämmastikaluste vaheline stäking. Teiseks tähtsaks teguriks on mitmesugused tertsiaarsed vesiniksidemed , mis moodustuvad tRNA D-õla ja molekuli üheahelaliste regioonide vahel. Voltumise tulemusena saavad vesiniksidemed tekkida ka tRNA erinevate üheahelaliste piirkondade vahel. Lämmastikaluste vahel leiavad aset hüdrofoobsed stäking-interaktsioonid. Selle tulemusel paigutuvad lämmastikalused teineteise suhtes nii, et nende kokkupuude veega oleks minimaalne. Teineteise lähedale sattunud tRNA heeliksite vahel toimub samuti stäking-interaktsioon (coaxial stacking) ja tulemuseks on heeliksite liitumine. Aktseptorõlg ja T-õlg liituvad ning moodustavad ühise heeliksi . Omavahel liituvad ka D-õlg ja antikoodon-õlg ja nii tekib teine heeliks. Antikoodon- ja aktseptorõlg jäävad molekuli vastaspooltele. Erinevate tRNA molekulide ruumiline struktuur peab olema sarnane, kuna nad kõik peavad seonduma ribosoomil samasse piirkonda.
tRNA sekundaarstruktuuris leiduvad üheahelalised
regioonid saavad paarduda D-õla aluspaaridega, tekitades seeläbi
kolmikahelalisi fragmente. Lisaks heeliksite paardumisele
interakteeruvad tRNA üheahelalised regioonid ruumilises struktuuris
ka omavahel.
D- ja T-ling satuvad voltumise tulemusena
lähestikku, mistõttu saavad tekkida interaktsioonid selle nn
DT-regiooni üheahelaliste piirkondade vahel.
Pole lõplikult selge, kuidas toimub tRNA
voltumine tertsiaarsesse L-kujulisse struktuuri, kuid oluliseks
abikomponendiks on
magneesium . D-õlas ja antikoodon-lingus paikneb 4
magneesiumiooni seondumissaiti. Mg2+
seondub vesiniksidemete abil mitte-helikaalsete regioonidega ning
stabiliseerib tRNA tertsiaarstruktuuri linge ning kurve. Mg2+
puudumisel võib tRNA voltuda ebakorrektsesse struktuuri.
Lisafaktoriks tRNA 3D struktuuri stabiliseerimisel on mõningad
nukleotiidide modifikatsioonid. On arvatud, et modifikatsioone läbi
viivad
ensüümid võivad toimida omalaadsete chaperonidena, aidates
tRNA voltumisele kaasa.
- initsiaator -tRNA ja elongaator-tRNA erinevused. Initsiaator-tRNA (tRNAfMet) ehitus on võrreldes elongaator-tRNA eriline. Bakteriaalsel initsiaator tRNA-l (v.a. termofiilid ja halofiilid) puudub aluspaardumine nukleotiidide 1 ja 72 vahel. See võimaldab metioniini formüleerimist. D-õlas paikneb Y11:R24 asemel R11:Y24 aluspaar . Antikoodon-heeliksis paikneb kolm G:C aluspaari, mille abil suunatakse initsiaator tRNA otse ribosoomi P-saiti. Eukarüootsel initsiaator-tRNA-l on N1:N72 aluspaardumine säilunud. Kuid T-lingus puudub TΨ-järjestus. Antikoodon-heeliksis paikneb kolm iseloomulikku G:C aluspaardumist.
- precursor-tRNA protsessimise etapid. tRNA geenide esmased transkriptid omavad 5'- ja 3'-otstes lisajärjestusi ja funktsionaalsete tRNA molekulide moodustumiseks peavad nad läbima protsessingu. tRNA transkripti 5’ otsas paikneva liigse järjestuse eemaldab endoribonukleaas RNaasP, mille variant on olemas kõikides organismides. RNaasP omab nii RNA komponenti kui ka valgulisi subühikuid. Üldiselt leiab 5’ otsa protsessing aset enne 3’ otsa protsessingut. tRNA geeni transkripti 3’ poolne protsessing on komplitseeritum. Erinevaid protsessingu radasid võidakse kasutada isegi ühe ja sama organismi piires. Esmalt lõikab prekursorit 3’-otsast endoribonukleaas RNaasE. Seejärel treivad eksoribonukleaasid RNaas PH ja RNaas T 3'-otsa parajaks. Kui tRNA on transkribeeritud pikemate prekursoritena, siis lõikavad eksonukleaasid PNPaas ja RNaas II esmalt transkripti lühemaks ja nüüd asuvad tööle 5' otsas RNaas P ja 3' otsas RNaas PH ja RNaas T. Kui tRNA geen ei kodeeri CCA-otsa, osaleb 3’ otste protsessimisel endonukleaas RNaas Z. RNaas Z sooritab lõike prekursori 3' otsas kohe pärast diskriminaatornukleotiidi (N73). Lisaks RNaas Z-le võib 3’ otsa protsessingul osaleda ka eksonukleaase, näiteks pärmi ensüüm Rex1p. Eukarüootidel leiavad nii tRNA protsessing kui ka 3'-terminaalse CCA järjestuse lisamine aset tuumas. organismides lisatakse CCA järjestus pärast 3' otsa protsessimist posttranskriptsiooniliselt ensüümi ATP(CTP):tRNA nukleotidüültransferaasi poolt.
- peamised modifikatsioonid tRNAs ja nende tähtsus. RNA molekulidest on siiani leitud 109 modifikatsiooni tüüpi, neist tRNA-del on avastatud 85. Seega on tRNA kõige modifitseeritum nukleiinhape rakus. Reeglina on lihtsate organismide ning organellide tRNA'd vähem modifitseeritud kui kõrgemate eukarüootide tsütoplasmaatilised tRNA'd. Kõige sagedasemad on lihtsad nukleosiidide modifikatsioonid: riboosi või aluse metülatsioonid, aluse isomerisatsioonid, aluste redutseerimised, aluste tiolatsioonid ja aluste deamineerimised. Keerulisematel juhtudel liidetakse nukleosiididele lisagruppe ja esineb ka aluste ja/või riboosi mitmekordseid modifikatsioone.
Modifitseerimine viiakse läbi
post-transkriptsiooniliselt spetsiifiliste ensüümide poolt.
Prokarüootidel ja arhedel toimub modifitseerimine
tsütoplasmas. Eukarüootidel võib see toimuda tuumas, tsütoplasmas
või organellides. Kuid modifitseerimine muudab pisikesi tRNA
molekule üksteisest erinevamaks, tõstab valgusünteesi täpsust ja
efektiivsust.
Loetelu mõnedest modifitseerimise funktsioonidest:
Antikoodoni struktuuri ja
koodon -antikoodon
interaktsiooni stabiliseerimine ning lugemisraami nihke vältimine, tRNA üldise
struktuuri stabiliseerimine, D-lingu painduvuse
suurendamine :
dihüdrouridiin, tRNA ebaõige voltumise vältimine, Kaitse
endonukleaaside eest, Fotoprotektsioon, Identsuselemendid aaRS-dele,
Antideterminandid aaRS-dele või teistele valkudele, Aluspaardumise
spetsiifika laiendamine
wobble-positsioonis
N34, Termostabiilsuse tõstmine.
Ribosoomi struktuur - Ribosoomi subühikud PROKARÜOODID :- 2 subühikut :väike e. 30S (16S rRNA-st sisaldab 4 domeeni; 5’- domeen , tsentraalne, 3’-major ja 3’- minoor ; 3’ots sialdab anti-SD järjestus, mis seondub start AUG-ga)ja suur e. 50S (23S rRNA sisaldab 6 domeeni; II IV – subühikutevahelised kontaktid, V peptidüültransferaasne aktiivsus, VI-faktorite sidumine+ aktivatsioon ; ja 5S rRNA on põhimiseks funktsiooniks siduda omavahel 23S rRNA domeen 2 ja 5 ). Funktsionaalne ribosoom : 70S (2.30 MDa). 3 rRNA molekuli : 5S & 23S rRNA (suures), 16S rRNA (väikeses). Suur subühik: ~ 34 valku. Väike subühik: ~ 21 valku. EUKARÜOODID :2 subühikut : 40S (väike), 60S (suur). Funktsionaalne ribosoom : 80S (~ 4 MDa). 4 rRNA molekuli : 5S, 5.8S (on suure subühiku mittekodeeriv RNA komponent ) & 28S rRNA (suur), 18S rRNA (väike). Suur subühik : ~ 50 valku. Väikse subühik : ~ 33 valku
- mida tähendab S nimedes 30S, 50S jne. Näitab, kui kergesti mingi osake tsentrifuugimisel põhja sadestub
- Valkude ja RNA struktuuri erinevad tasemed
- primaar- (1’) (aminohappe lineaarne järjestus), sekundaar- (2’) (AH järjestus on seotud vesenik sideme abil ) ja tertsiaarstruktuur (3’) (ühe polüpeptiid ahel ühe v mtme valgu sekundaarse struktuuriga )
- valkude sekundaarstruktuuri elemendid (α-heeliks- iga aminogrupp on seotud AH karbonüül rühmaga, mis asub 4 AH varem, β-leht- koosneb beeta ahelast , mis on külgsuunas seotud kahe v kolme vesiniksidemega)
- RNA sekundaarstruktuuri elemedid (stem-loop, pseudosõlm, A-minoor- ahelate seondumine, kui vähemalt 2 ahelat on omavahel juba alfa-heeliksis ja siis midagi veel selle külge kleepuda üritab.)
- Domeeni mõiste valkudes ja RNA-s Valgu või nukleiinhappe osa, mis on võimeline voltuma (ingl. fold), funktsioneerima, evolutsioneeruma sõltumatult molekuli ülejäänud osadest. Võtab iseseisvalt struktuuri. Üldiselt üks evolutsioneeruv osa.
- rRNA ruumilise struktuuri kujunemise üldpõhimõtted
- kaksikhelikaalsete piirkondade osakaal rRNA-s
- coaxial stacking ja selle tähtsus rRNA 3D struktuuri jaoks
- A-minoorne interakstioon-
- hoiab rRNA ruumilist struktuuri paigas. RNA kaksikheeliksi väikese vao kaudu toimub interaktsioon A-nukleotiidide (A nukleotiidid pärinevad üksikahelast; adeniin ) ja rRNA vahel. Moodustatakse põhiliselt vesiniksidemeid kaheahelalises osas osalevate 2’ hapnikega (riboosi hapnikud). Eristatakse 4 erinevat tüüpi interaktsiooni, olenevalt sellest, milliste kaksikahelaliste piirkondadega sidemet tekivad.
- 16S rRNA sekundaarstruktuur: Sisaldab 4 domeeni : 5’-domeen : KEHA, tsentraalne : PLATVORM, 3’-Major : PEA, 3’- minor : PENULTIMATE STEM. Domeenid 30S tasemel suhteliselt iseseisvad. Domeenide liikumine teineteise suhtes valgusünteesi käigus.
- 23S rRNA sekundaarstruktuur 23S rRNA : Sisaldab 6 domeeni, Domeenid tihedalt läbipõimunud - moodustavad 50S tasemel monoliitse ja väheliikuva struktuuri, Erandiks L1 ja L11 stalk – liikuvad ja ülejäänud 50S-ist suhteliselt omaette . Domeenidel 50S tasandil erinevad funktsioonid:Dom II : subühikutevahelised kontaktid. Dom IV : subühikutevahelised kontaktid, dekodeerimine. Dom V : peptiidsideme teke, faktorite sidumine + aktivatsioon. Dom VI : faktorite sidumine + aktivatsioon.
- 23S rRNA domeenid on üksteisega tihedalt läbipõimuinud, sestap on 50S tunduvalt monoliitsem kui 30S. Väikese subühiku struktuur on tunduvalt paindlikum/liikuvam kui suure subühiku oma. Omab keskset kühmukest. 30S subühiku pea pöörleb võrreldes kehaga ja kogu subühik põõrleb veel lisaks ka suure subühikuga võrreldes
- Ribosoomi valgud (r-valgud). Valdavalt lühikesed (25 – 300 aminohapet). Valdavalt aluselised (pI = 10.1) (erandiks happeline L7/L12; pI = 4.9). Interakteeruvad rRNA-ga (erandiks L7/L12). Struktuur: globulaarne, globulaarne + struktuuritu, struktuuritu. Globulaarsed osad ribosoomipartikli pinnal. Struktuuritud osad ulatuvad partikli sügavusse. Puuduvad subühikute kokkupuutepinnalt (interface). 16S ja 23S rRNA struktuuri kujunemine & säilitamine. tRNA positsioneerimine ribosoomis. Dekodeerimine. mRNA mobilisatsioon (ingl. recruitment) (S1). mRNA sekundaarstruktuuride lammutamine (ribosoomi helikaasne aktiivsus). Elongatsioonifaktorite – IF2, EF-Tu, EF-G - mobilisatsioon . Interaktsioon kasvava peptiidiga.
- r-valkude jaotus suures ja väikeses, subühikus 50S sisemuses paiknevad valguosad positiivselt laetud. 50S pinnal paiknevad valguosad negatiivselt laetud
- kust on pärit r-valkude nimed (a la S5, L4) S5 väikses subühikus: vähendab translatsiooni täpsust ja tõstab spectinomyciine AB resistentsus ; L4 reguleerib S10 operooni transkriptsiooni.
- Subühikutevahelised sillad . Kontakteeruvad piirkonnad 30S & 50S subühikus. Hoiavad subühikuid koos 70S ribosoomis. Jaotunud üle terve subühikute kokkupuutepinna (ingl. interface). Kokku ≈ 12 silda : B1 – B8. RNA-RNA sillad interface´i keskosas: staatilised. RNA-valk, valk-valk sillad perifeerias: dünaamilised
- kus paiknevad. 50S dom II ja IV
- millest koosnevad
- RNA-RNA sillad versus RNA-valk/valk-valk sillad (kus asuvad, kuidas muutuvad/ei muutu elongatsiooni käigus)
- sildade funktsioonid seovad translatsioonis seovad subühikud omavahel valkude sünteesil
- Kahevalentsed metallid ja rRNA stabiilsus ning subühikute koospüsimine
- Eukarüootse ribosoomi struktuur võrreldes prokarüoodi omaga
- 40 % mahukam rRNA
- 32 täiendavat r-valku
- 4 täiendavat subuühikutevahelist silda
- 2 x suurem kontaktipind subühikute vahel
- rohkem üheahelalist RNA-d
Ekstensioonisegmendid on pärmidel olemas, kuid
puuduvad eubakterites. Bakteriribosoomile vahele lisatud
sekundaarstruktuuri elemendid. Enamasti
heeliksite otsas
pikemad jätked (nt lisalingud). 5S on eukarüootidel
ja bakteritel väga sarnased. Väiksema subühiku rRNA on väikeste
lisadega, suur subühik on suuremate lisadega, millest üks on eriti
pikk ja massiivne. Paiknevad ribosoomi välimisel küljel.
Ribosoomi subühikutevahelised sillad
eukarüootides on samad mis bakteritel, lisaks 3-4tk.
Ribosoomivalgud – riigispetsiifilised ja siis
üldised.
- Ribosoomi ” funktsionaalsed tsentrid” ja nende roll valgusünteesil: 30S Dekodeeriv tsenter - koodon : antikoodon (paardumise kontroll). P/E-saidi “värav”- tRNA liikumine P-saidist E-saiti translokatsiooni ajal. 16S rRNA 3’-ots- mRNA kinnitumine ribosoomile SD-anti-SD interaktsiooni abil. 16S rRNA heeliks 44- subühikutevahelised kontaktid.
50S:
Peptidüültransferaasne
tsenter-peptiidsideme moodustumine.
L11 stalk-Translatsioonifaktorite
“kinnipüüdmine” ; translatsiooniliste
GTPaaside aktivatsioon.
Sartsiin-ritsiin ling -
Translatsiooniliste GTPaaside
aktivatsioon.
L1
stalk-tRNA
väljutamine E-saidi kaudu.
A-sait-aminoatsüül-tRNA sidumine.
P-sait peptidüül-tRNA
sidumine.
E-sait-
deatsüleeritud-tRNA sidumine & väljutamine.
Valgusünteesi
elongatsioonifaas
Enne kui algab
translatsiooni elongatsioonifaas, peab olema toimunud
initsiatsioonifaas (initsiatsioonist tuleb kunagi eraldi loeng).
Initsiatsiooni põhiline
keerukus valgusünteesis seisneb tõsiasjas,
et kui RNA süntees algab 1 nukleotiid enne või pärast, siis suurt
funktsionaalset kaotust pole.
Tuleb leida üles õige
lugemisraam ja alustada valgusünteesi sealt kohast, kus ta on
ettenähtud
algama – initsiatsiooni kõige tähtsam ülesanne.
Translatsiooni initsiatsioonil osalevad mitmed
initsiatsioonifaktorid: IF1, IF2, IF3. Initsiaatorkoodoni üles
leidmisel osaleb Shine-Dalgarno (SD) järjestus, mis asub mRNA
initsiaatorkoodoni ees. rRNA 3’ ots seondub SD järjestusega.
Initsiaatorkoodoniks on AUG. Kui 30S initsiatsioonikompleks on paika
saanud, siis
lahkub sealt IF3 faktor ning järelejäänud kompleksiga
ühineb suurem subühik, misjärel võib
elongatsioon peale alata.
AUG start-koodon on komplementaarne
initsiaator-tRNA-ga (ehk
fMet -tRNA, antikoodoniks 3’ – UAC –
5’, mis võimeline seega seondama AUG start-koodoniga).
Initsiatsioonisaidi valik sõltub suuresti 30S subühiku
omavahelisest interaktsioonist mRNA järjestusega.
Elongatsioon on tsükliline
protsess, mille ühe tsükli käigus pikeneb peptiidahel ühe
aminohappe võrra. Valgud sünteesitakse N-terminusest C-terminuse
poole. Igal aminohappel on alfa-aminogrupp ja alfa-karbosküülgrupp,
järgmine aminohape lisatakse karboksüülhappele. Uue aminohappe
aminorühm atakeerib alfa-karboksüülrühma süsinikku. Ribosoom
loeb mRNA-d 5’ 3’ suunas (RNA sünteesi suund sama).
Ribosoomi sees tRNA-d liiguvad alguses A, siis P ja lõpuks E saiti.
Ribosoomi subühikud liiguvad teineteise suhtes iga
elongatsioonitsükli käigus. Elongatsioonil osalevad translatsiooni
elongatsiooni faktorid, mille hulka kuuluvad 3 universaalset
faktorit : EF-Tu (+ GTP) – vahendab aminoatsüül-tRNA sisenemist
ribosoomi vabasse A-saiti; EF-Ts – EF-Tu guaniini
nukleotiidivahetusfaktor, katalüüsides GDP vabanemist EF-Tu-lt;
EF-G (+ GTP) – katalüüsib tRNA ja mRNA translokatsiooni mööda
ribosoomi pärast iga polüpeptiidi elongatsioonitsükli lõppu.
Terminatsiooni käigus peab
vabanema valmissünteesitud peptiidahel ja ribosoomid tuleb vabastada
kõigist tema ligandidest.
Terminatsiooni- (ehk
nonsense ) ehk STOP-koodoneid on 3:
• UAG -
amber • UAA - ochre
• UGA - opal
Kui ribosoomi A-saiti satub
üks
nendest koodonitest ja P-saidis on peptidüül-tRNA, siis algab
terminatsioon. Peptiidi vabanemisfaktor ehk klass I polüpeptiidi
vabanemisfaktor (RF ehk
release factor ) peab seonduma, tundes ära
A-saidis oleva STOP-koodoni. Katalüüsib ka peptidüül-tRNA
hüdrolüüsi.
Ribosoomide retsükleerimine
tähendab ribosoomine viimist sellisesse seisu, et nad võiksid jälle
alustada uue peptiidahela sünteesi. Ribosoomi
retsükleerimine – ribosoomid vabastatakse, polüpeptiid eraldub,
mRNA eemaldatakse ribosoomide küljest, subühikud dissotsieeruvad ja
saavad uuesti alustada järgmist ringi translatsiooni
initsiatsiooniga.
- Ülevaade elongatsioonifaasi põhisündmustest. Polüpeptiidahel pikeneb 1 ah kaupa. Ahela pikenemise suund: N- term → C-term. Ribosoomi liikumine mRNA-l: 5’ → 3’. tRNA-de liikumine saitides: A →P → E. 30S ja 50S liiguvad üksteise suhtes. Osalevad elongatsioonifaktorid :EF-Tu (+ GTP), EF-Ts, EF-G (+ GTP).
- Valgusünteesi üldine mehhanism : erinevus DNA/RNA biosünteesist (a la väike molekul suure otsa versus suur molekul väikese otsa). NB! Vaadake loengu ”Elongatsioon I” slaidi 8.
- Ühe elongatsioonietapi ligikaudne kiirus prokarüootides pikenes valk mingi (15-)20 ah/s.
- Ribosoomi saidid (A kuni E) ja milliseid tRNA-sid seovad:
- aminoatsüül-tRNA
- peptidüül-tRNA
- deatsüleeritud tRNA
- Elongatsioonifaktorid pro- ja eukarüootides ja nende roll. EF-Tu (EF1a)- aa-tRNA transport ribosoomi A-saiti. Koodon-antikoodon interaktsiooni kontroll. EF-Ts (EF-1b)- GDP asendamine GTP-ga EF-TU-l. EF-G (EF-2) – katalüüsib ribosoomi translokatsiooni mRNA-l.
- GTPaasid ja nende rollid rakus. Osalevad: rakulises signaaliülekandes, valkude translokatsioonil, rakujagunemise kontrollis , valgusünteesil. Mehhanism: GTP sidumine ja hüdrolüüs, pendeldavad 2 oleku vahel – GTP- seoseline AKTIIVNE; GDP-seoseline INAKTIIVNE.
- GTPaasi tsükkel , G-nukleotiidi vahetusfaktor GEF, GTPaasi aktiveeriv valk GAP
- GTPaasid ja G-domeen: Konserveerunud piirkonnad: switch I, switch II, P-loop
P-loop seob GTP
fosfaatrühmi
Switch-id
olulised GTP hüdrolüüsiks
NB! Vaadake slaide 12 - 13
- EF-Tu Tu” : T –” transfer”, u - “unstable”. Monomeerne GTPaas, 3 domeeni
GTPaasne aktiivsus.
GTPaasi stimuleerib ribosoom.
Kolmikkompleks :
EF•Tu-GTP•aa-tRNA. EF-Tu
seondub ribosoomiga kolmikkompleksis. EF-Tu konformatsioon muutub GTP
hüdrolüüsi järel. EF-Tu•GTP
kompleks seob aa-tRNAd
tugevalt EF-Tu nn. “suletud”
vormis GTP hüdrolüüs EF-Tu-l. EF-Tu omandab “avatud”
konformatsiooni EF-Tu•GDP
dissotsieerub aa-tRNA küljest.
His84 aktiveerib H2O
molekuli, see atakeerib γ-fosfaatrühma
GTPs. Val20/Ile60 “värav” kaitseb GTP-d His84 toime eest.
Ribosoom stimuleerib “värava” avanemist.
“Avatud” värava korral toimub GTP
hüdrolüüs .
- EF-Ts. Ts: T – “temperature”, s- “stable”. Monomeerne valk. G-nukleotiidi vahetusfaktor (GEF) EF-Tu jaoks:
EF-Tu•GDP + GTP
EF-Tu•GTP + GDP
- EF-G. Monomeerne valk. 5 domeeni. GTPaasne aktiivsus (ribosoom-stimuleeritud). Katalüüsib mRNA-tRNA kompleksi translokatsiooni. Sarnaneb struktuurilt aa-tRNA-EF-Tu kolmikkompleksiga - molecular mimicry. Domeen IV liigub A-saiti sarnaselt tRNA antikoodon õlale. mRNA-tRNA kompleksi liikumine ühe koodoni võrra mRNA 3’ suunas .
PRE-olek :
deatsüleeritud tRNA P-saidis, peptidüül-tRNA A-saidis
POST-olek :
deatsüleeritud tRNA E-saidis, peptidüül-tRNA P-saidis
Translokatsioon mRNA-tRNA kompleksi liikumine ühe
koodoni võrra mRNA 3’ suunas, EF-G katalüüsib seda.
- Hübriidsaidid (A/P, P/E) Translokatsiooni eel ja järel tRNA-d A/A ja P/P olekus . Translokatsiooni käigus tRNA-d A/P ja P/E HÜBRIIDSAITIDES. Hübriidsaidid “vahetult” näha ribosoomide krüo-EM-is. Hübriidsaitide ajas moodustumise järjekord : P/E → A/P. Hübriidsaitide tekkeks vajalik P-saidi tRNA deatsüleeritud olek !!! tRNA-de minek hübriidsaitidesse sõltumatu EF-G-st !!!
- Ribosoomi konformatsioonilised muutused elongatsioonil. Väike subühik pöördub suure subühiku suhtes. ~ 60 → ratchet liikumine. Samaaegselt hübriidsaitide moodustumine. Ratcheti vallandab P-saidi tRNA deatsüleerimine (nn. ribosoomi unlocking). Ratchet toimub ka EF-G puudumisel. EF-G stabiliseerib ribosoomi “ratchet” olekut. Ümberkorraldused B1a ja B1b sildade. L1 stalk liigub ratcheti ajal E-saiti sissesubühikute ”ratchet” :
- Translokatsioon ja antibiootikumid : spektinomütsiini juhtum
Spectinomycin interacts
specifically with the residues G1064 and C1192 in 16S rRNA, thereby
potentially freezing this molecule into an inactive conformation.
Takistab kuidagi EF-G seondumise, mis peatab ka A-->P sait
liikumise
Spektinomütsiin võib
blokeerida olulise liikumise ribosoomis. Seondub ribosoomi kaela
piirkonda (väike subühik), translatsioon enam toimuda ei saa (jääb
kinni).
Valgu biosünteesi täpsus ja
koodon-antikoodon interaktsioonid
Valgusünteesi täpsus on mõjutatud kahe protsessi poolt. Esiteks
peavad tRNA-d ribosoomis korrektselt sobituma neile vastavate mRNA
koodonitega ja teiseks teguriks on tRNA
korrektne aminoatsüleerimine.
Korrektne aminoatsüleerimine sõltub otseselt aaRS suutlikusest ära
tunda ja aminoatsüleerida õige tRNA.
Vigade sagedus. DNA süntees:
1:10e8-10e10. RNA süntees: 1:5000-10000. Valgusüntees 1:1000. DNA
replikatsioonil on reparatsiooni süsteem. Pärast DNA sünteesi
moodustub metüleeritud vorm ( vale nukleotiidi sisse lülitamisel
lagundatakse mitte metüleeritud DNA.) See tõstab täpsust.
Valgusünteesil vigade sagedus tõstab mRNA lugemisraami nihke, vale
tRNA
sattumine ribosoomi ja tRNA aminoatsüleerimine vale ah-ga.
Geneetiline kood. Koodon on
ühele aminohappele vastav mRNA molekuli
nukleotiidikolmik (triplett) geneetilises
koodis.
Antikoodon on tRNA molekulis olev kolmest nukleotiidist koosnev
spetsiifiline järjestus, mis valgu biosünteesis võib mRNA
komplementaarse koodoniga moodustada aluspaare. Kõdumine on
1aminohappele vastab rohkem kui 1 koodon. Koodoni 1.&
2. Positsioonil onWatson-Crick
paardumine : A – U, G – C. Koodoni
3. Positsioon
Wobble paardumine A/C/U – I(nosiin), A/G –
U, C - G
- Mis on sense - ja mis nonsense koodon. Sense koodon, mis kodeerib ah, nonsense+ terminaator koodon.
- Mõisted ”cognate”, ” near -cognate” ja ”non-cognate” koodon-antikoodon paardumise korral. Cognate- antikoodon paardub koodoniga, near-cognate- esimeses positsioonis ei paardu, non- cognate 1. Ja 2. Ei paardu.
- Valgusünteesi täpsuse tagamine
- Kas erinevus koodon-antikoodon paari stabiilsuses cognate ja near-cognate tRNA korral on piisav, et ühes selektsioonietapis eristada cognate tRNA near-cognate tRNA-st (Francis Cricki hüpotees). Valepaardumine = vähem H- sidemeid . Koodon-antikoodon paardumine VÄHEM STABIILNE. Vale tRNA dissotsieerub ribosoomilt enne, kui peptiidside moodustub. Near-cognate tRNA korral suurem GTP kulu
- Milline parameter iseloomustab koodon-antikoodon paardumise stabiilsust (tuletage meelde mõisted “vabaenergia”, “tasakaal”, “tasakaalukonstant”) vabaenergia, ning tasakaalukonstant
- Kas on tõendeid, et ribosoom osaleb koodon-antikoodon paardumise õigsuse kontrollis : Streptomütsiin: seondub 30S subühikuga, valede aminohapete lülitumine valkudesse, stop-koodoni läbilugemine. SmR-mutandid : mutatsioonid valgus S12, resistentsus streptomütsiini suhtes, “hüpertäpsed” ribosoomid. Ram-mutandid : mutatsioonid valkudes S4 & S5: jäljendavad streptomütsiini toimet,- SmR- & Ram-mutatsioonide antagonism.Hüpotees dekodeerivast tsentrist 30S subühikul
- tRNA kaheetapiline selektsioon : Near-cognate tRNA dissotsiatsioon kiireneb
- initial selection (IS)
ajal
- proofreading (F) ajal
• Non-cognate tRNA IS-ist
edasi ei jõuagi
• GTP hüdrolüüsi kiirus
tRNA-st ei sõltu
• EF-Tu kui “
internal clock ”
• Near-cognate tRNA korral
suurem GTP kulu
NB! Vaadake ”Translatsiooni
täpsus” slaidi nr. 34
- Koodon-antikoodon dupleksi struktuur ja interaktsioon dekodeeriva tsentriga
- DNA/RNA kaksikheeliksi suur ja väike vagu
- A-minor interaktsioon
Dekodeeriv tsenter :
dekodeerivas tsentris on kaks lapatsina liikuvat RNA nukleotiidi
A1492 ja A1493 käib vaid koodonites 1 ja 2 (väikese vao poolt)
kolmandas ei ole, seal saab toimuda too wobble paardumine. A minor
paardumine aitab energeetiliselt paardumisele kaasa, ilma selleta
tuleb lisakulutused
- asukoht ribosoomil
- olulisemad komponendid
- A1492 ja A1493 ”flipping-out” ja A-minor interaktsioon koodon-antikoodon miniheeliksiga
Interaktsioonid
DC-s.
1. Koodoni positsioonis.
A-minoor I. A1493 täidab väikese vao kogu ulatuses. A1493
kontakteerub koodoni 1 nt, antikoodoni 3. Nt. See on A-minoor I
interaktsioon. A minor I on tundlik k-ak paari geomeetria suhtes. G-U
mismatch kaotab H-sideme A1493 ja U1 2’-OH vahel. Mismatchi
energeetiline hind 16-40 kJ/mol.
2. Koodoni
positsioonis.A1492 täidab poole väikesest vaost. A1492
kontakteerub koodoni teise nt ja antikoodoniga läbi G530-i. See on A
minor II interaktsioon. Oluline on ka valgu S12 Ser50. G-u mismatch
kaotab A1492 kontakti koodon-antikoodon paariga, selle hinnaks on
15-20 kJ/mol.
3. Koodoni positsioonis.
Wobble. 16S rRNA ei interakteeru koodon-antikoodon paariga.
Dekodeeriv tsentr k-ak paari kuju suhtes „tundetu“. Koodoni
kolmandas positsioonis on võimalikud ebastandartsed aluspaarid –
Wobble (G-U, I-A, I-C).
30S „
sulgumine “. Õige
antikoodon A-saidis → dekodeeriva tsentsi konformatsioon muutub,
see levib üle 30S-i. 30S „suletud“ vormi („pea“ ja „õlg“
liiguvad A-saidi poole). 30S konformatsiooni muutus viib EF-Tu
GTPaasi aktivatsioonile. aa-tRNA 3’-CCA ots liigub PT
tsentrisse.kuidas
- seletavad struktuuriandmed “wobble” nähtust
- A1492/ 1493 “flipping-out” ja 30S konformatsioonilised muutused
kui on Õige antikoodon
A-saidis :
dekodeeriva tsentri konf.
muutub
• Konf. muutus levib üle
30S-i
• 30S “suletud” vormi
:
- “pea” & “õlg”
liiguvad A-saidi poole
• 30S konf. muutus viib
EF-Tu GTPaasi aktivatsioonile
• aa-tRNA 3’-CCA ots
liigub PT tsentrisse
- Kuidas viib korrektne (cognate) koodon-antikoodon paardumine GTP hüdrolüüsile EF-Tu-s
Translatsiooni terminatsioon ja
retsükleerimine - Mis on terminatsioon; peptiidi vabanemine ; subühikute retsükleerimine
Kui polüpeptiid on valmis, siis A-saidis on üks
kolmest STOP ehk nonsense-koodonist (UAG – „amber“, UAA –
„ochre“, UGA – „opal“). P-saidis on peptidüül-tRNA. Klass
I polüpeptiidi vabanemisfaktor tunneb ära stop-koodoni A-saidis
ning katalüüsib peptidüül-tRNA hüdrolüüsi. Järgnevalt
eemaldab klass II vabanemisfaktor ribosoomilt klass I faktori ja
lahkub seejärel ise ribosoomilt. Klass II vabanemisfaktor on GTPaas
(sarnaneb EF-Tu-le ja EF-G-le). RRF (ribosome
recycling factor) ja
EF-G lammutavad ribosoomi subühikuteks, mis on valmis alustama uut
valgusünteesiraundi. Eristatavaks on 3 etappi:
● Peptiidi vabanemine (katalüüsib klass I
faktor).
● Klass I faktori vabanemine (katalüüsib klass
II faktor, protsess nõuab GTP energiat).
● Ribosoomi
lagunemine subühikuteks
(katalüüsivad RRF ja EF-G, protsess nõuab GTP energiat)
- mis on signaaliks, mis käivitab terminatsiooni
Klass I polüpeptiidi vabanemisfaktor tunneb ära
stop-koodoni A-saidis ning katalüüsib peptidüül-tRNA hüdrolüüsi
UAG ("amber") "U Are Gone”
UAA ("ochre") "U Are
Away "
UGA ("opal") "U Go Away"
- Klass I ja klass II polüpeptiidi vabanemisfaktor : mida teeb klass I faktor, mida klass II faktor
Klass I polüpeptiidi vabanemisfaktor (ingl.
release factor)-tunneb ära stop-koodoni A-saidis, katalüüsib
peptidüül-tRNA hüdrolüüsi. Järgnevalt eemaldab klass II
vabanemisfaktor ribosoomilt klass I faktori ja lahkub seejärel ise
ribosoomilt.
- Klass I polüpeptiidi vabanemisfaktorid prokarüootides
Prokarüootides: RF1 : tunneb ära UAG ja UAA
koodoni. RF2 : tunneb ära UGA ja UAA koodoni. Ribosoomiga
seondumisel konf. muutus RF1/2-s. Domeen III liigub eemale dom.
II-st. Domeen III GGQ
motiiv satub peptidüültransferaassesse tsentrisse. GGQ-motiiv : sama nii RF1-s kui RF2-s, domeenis III,
vajalik pept-tRNA hüdrolüüsiks. “Tripeptiidne antikoodon” - :
domeenis II, P(A/V)T – RF1, SPF – RF2, vajalik STOP-koodoni
äratundmiseks. “antikoodoni” vahetus RF1 ja RF2 vahel paneb RF1
ära tundma RF2
koodonit (UGA) ja
vice versa (Ito et al. 2000).
RF1/RF2. Pos 1. Ainult pürimidiin (C,U)
mahub “seondumise
taskusse ” Aga C ei annaks vajalikke H-sidemeid
RF1 jääkidega T186 ja G116↓ Seega sobib ainult U. RF1. Pos2.
Pürimidiinid (C,U) ei anna hüdrofoobseid interaktsioone U1 ja
Glu119/Pro184-ga Jäävad
puriinid A ja G. Aga G ei annaks vajaliku
H-sidet RF1 jäägiga T186. Seega sobib ainult A. RF2. Pürimidiinid
ei annaa vesinuk sidemeid. Seega Avõi U. RF1pos3 Avõi U, RF2- C ja
U ei anna hüdroboobseid sidemed. Peptidüül-tRNA estersideme
hüdrolüüsiks vajalik GGQ (glütsiin, glütsiin, glutamiin) motiiv
RF1/2 domeenis 3. Hüdrolüüs :
ester + H2O. Seondumistasku H2O
molekulile
▫ Q240 (RF)
▫ C2063 (23S rRNA)
▫ A2451 (23S rRNA)
▫ A76 2’-OH (pept-tRNA)
GGQ stabiliseerib vaheoleku.
- Mis on ”tripeptiidne antikoodon”
RF1 on neli domeeni: Domäänis 2
on
tripeptiidne antikoodon, mis tunneb ära terminatsioonikoodoni
A-saidis
NB! Vaadake loengu “Terminatsioon ja
retsükleerumine” slaidi 23 - Klass II vabanemisfaktor RF3
- Prokarüootides : RF3. Eukarüootides : eRF3. GTPaas : sarnane IF2, EF-Tu ja EF-G-ga. Kolm domeeni : G(incl. G’), II ja III. Domeen III : unikaalne “fold”. RF3-e substraat : Ribosoom•deac-tRNAPsite •RF1/2. RF3 eemaldab RF1/2 ribosoomilt ja siis hüdrolüüsib GTP. Peale GTP hüdrolüüsi RF3 dissotsieerub ribosoomilt. RF3 seondub ribosoomile GDP-vormis. Ribosoom põhjustab GDP→GTP vahetuse . RF3 GTP-vormis kutsub esile subühikute omavahelise liikumise: ratchet. See liikumine on sarnane translokatsioonil esineva ratchetiga. Subühikute omavaheline ~ 10 kraadi nihkumine katkestab kontaktid RF1/2 ja ribosoomi vahel:
▫ RF1/2 – dekodeeriv tsenter
▫ RF1/2 – L11 stalk
RF1/2 dissotsieeruvad ribosoomilt.
RRF ehk ribosoomi retsükleerimisfaktor (ingl.
ribosome recycling factor). 2 domeeni: domeen I (“saba”) ja
domeen II (“pea”). Seondub 70S ribosoomiga : domeen I : 50S ja
domeen II : 30S. Koos EF-G-ga põhjustab 70S lagunemise subühikuteks.
RRF-i kaks seondumispositsiooni : Pos 1 – 70S ribosoomil ja Pos 2 –
50 subühikul. RRF seondub RS-ga enne EF-G-d. RRF ja EF-G
seondumiskohad kattuvad kui RRF positsioonis 1. Järeldus : EF-G
seondumisel võtab RRF sisse positsiooni 2 ( GTP hüdrolüüsi järel
liigud EF-G dom Iv dom III poole ja selle väänad välja RF pea).
RRF minek positsiooni 2 lõhub subühikutevahelised sillad B2a ja
B3.B2a ja B3 on ühed kõige tugevamad sillad. Järeldus : subühikud
dissotsieeruvad. Aminoglükosiidid takistavad H69 liigutamist RRF-i
poolt. See hoiab B2a silla koos. Ja järelikult pärsib subühikute
dissotsiatsiooni.
Translatsiooni terminatsioon ja
retsükleerimine - Mis on terminatsioon- valgu sünteesi lõpetamine , polüpeptiidi vabanemine.
Kui polüpeptiid on valmis,
siis A-saidis on üks kolmest STOP ehk nonsense-koodonist (UAG –
„amber“, UAA – „ochre“, UGA – „opal“). P-saidis on
peptidüül-tRNA. Klass I polüpeptiidi vabanemisfaktor tunneb ära
stop-koodoni A-saidis ning katalüüsib peptidüül-tRNA hüdrolüüsi.
Järgnevalt eemaldab klass II vabanemisfaktor ribosoomilt klass I
faktori ja lahkub seejärel ise ribosoomilt. Klass II vabanemisfaktor
on GTPaas (sarnaneb EF-Tu-le ja EF-G-le). RRF (ribosome recycling
factor) ja EF-G lammutavad ribosoomi subühikuteks, mis on valmis
alustama uut valgusünteesiraundi. Eristatavaks on 3 etappi:
- Peptiidi vabanemine (katalüüsib klass I faktor).
- Klass I faktori vabanemine (katalüüsib klass II faktor, protsess nõuab GTP energiat).
- Ribosoomi lagunemine subühikuteks (katalüüsivad RRF ja EF-G, protsess nõuab GTP energiat) mis on signaaliks, mis käivitab terminatsiooni: A saidis on stop koodon
- STOP-koodonid: UAG – “amber”, UAA – “ochre”, UGA – “opal”
- Klass I ja klass II polüpeptiidi vabanemisfaktor : mida teeb klass I faktor, mida klass II faktor. Klass I polüpeptiidi vabanemisfaktor (ingl. release factor)-tunneb ära stop-koodoni A-saidis, katalüüsib peptidüül-tRNA hüdrolüüsi. Järgnevalt eemaldab klass II vabanemisfaktor ribosoomilt klass I faktori ja lahkub seejärel ise ribosoomilt.
- Klass I polüpeptiidi vabanemisfaktorid prokarüootides. Prokarüootides: RF1 : tunneb ära UAG ja UAA koodoni. RF2 : tunneb ära UGA ja UAA koodoni. Ribosoomiga seondumisel konf. muutus RF1/2-s. Domeen III liigub eemale dom. II-st. Domeen III GGQ motiiv satub peptidüültransferaassesse tsentrisse. GGQ-motiiv : sama nii RF1-s kui RF2-s, domeenis III, vajalik pept-tRNA hüdrolüüsiks. “Tripeptiidne antikoodon” - : domeenis II, P(A/V)T – RF1, SPF – RF2, vajalik STOP-koodoni äratundmiseks. “antikoodoni” vahetus RF1 ja RF2 vahel paneb RF1 ära tundma RF2 koodonit (UGA) ja vice versa (Ito et al. 2000). RF1/RF2. Pos 1. Ainult pürimidiin (C,U) mahub “seondumise taskusse” Aga C ei annaks vajalikke H-sidemeid RF1 jääkidega T186 ja G116 ↓ Seega sobib ainult U. RF1. Pos2. Pürimidiinid (C,U) ei anna hüdrofoobseid interaktsioone U1 ja Glu119/Pro184-ga Jäävad puriinid A ja G. Aga G ei annaks vajaliku H-sidet RF1 jäägiga T186. Seega sobib ainult A. RF2. Pürimidiinid ei annaa vesinuk sidemeid. Seega Avõi U. RF1pos3 Avõi U, RF2- C ja Uei anna hüdroboobseid sidemed. Peptidüül-tRNA estersideme hüdrolüüsiks vajalik GGQ motiiv RF1/2 domeenis 3. Hüdrolüüs : ester + H2O. Seondumistasku H2O molekulile
▫ Q240 (RF)
▫ C2063 (23S rRNA)
▫ A2451 (23S rRNA)
▫ A76 2’-OH (pept-tRNA)
GGQ stabiliseerib vaheoleku.
- Klass II vabanemisfaktor RF3. Prokarüootides : RF3. Eukarüootides : eRF3. GTPaas : sarnane IF2, EF-Tu ja EF-G-ga. Kolm domeeni : G(incl. G’), II ja III. Domeen III : unikaalne “fold”. RF3-e substraat : Ribosoom•deac-tRNAPsite •RF1/2. RF3 eemaldab RF1/2 ribosoomilt ja siis hüdrolüüsib GTP. Peale GTP hüdrolüüsi RF3 dissotsieerub ribosoomilt. RF3 seondub ribosoomile GDP-vormis. Ribosoom põhjustab GDP→GTP vahetuse. RF3 GTP-vormis kutsub esile subühikute omavahelise liikumise: ratchet. See liikumine on sarnane translokatsioonil esineva ratchetiga. Subühikute omavaheline ~ 10 kraadi nihkumine katkestab kontaktid RF1/2 ja ribosoomi vahel :
▫ RF1/2 – dekodeeriv tsenter
▫ RF1/2 – L11 stalk
RF1/2 dissotsieeruvad ribosoomilt.
- RRF ehk ribosoomi retsükleerimisfaktor (ingl. ribosome recycling factor). 2 domeeni: domeen I (“saba”) ja domeen II (“pea”). Seondub 70S ribosoomiga : domeen I : 50S ja domeen II : 30S. Koos EF-G-ga põhjustab 70S lagunemise subühikuteks. RRF-i kaks seondumispositsiooni : Pos 1 – 70S ribosoomil ja Pos 2 – 50 subühikul. RRF seondub RS-ga enne EF-G-d. RRF ja EF-G seondumiskohad kattuvad kui RRF positsioonis 1. Järeldus : EF-G seondumisel võtab RRF sisse positsiooni 2 ( GTP hüdrolüüsi järel liigud EF-G dom Iv dom III poole ja selle väänad välja RF pea). RRF minek positsiooni 2 lõhub subühikutevahelised sillad B2a ja B3.B2a ja B3 on ühed kõige tugevamad sillad. Järeldus : subühikud dissotsieeruvad. Aminoglükosiidid takistavad H69 liigutamist RRF-i poolt. See hoiab B2a silla koos. Ja järelikult pärsib subühikute dissotsiatsiooni.
Eukarüootse translatsiooni initsiatsioon ja
regulatsioon - Eukarüootse ja prokarüootse translatsiooni võrdlus. Mis on sarnane, mis erinev ja mis võiksid olla erinevuste laiemad põhjused
- Eukarüootse translatsiooni initsiatsioon
- Põhilised eIF-id : eIF2- moodustab kompleksu Met-tRnai-ga ja GTP-ga, mis annab algust inisatsionile, siis seondab vaba 40S subühikuga. Toob inisatsiooni tRNA. eIF3seondab 40S ja eIF5 on GAP eIF2jaoks( gtp aktivaator). eIF4A- helikaas, mis aitab lahti keerata mRNA. eIF4E - tunneb ära cap struktuuri. eIFA- sarnane bakterite IF-ga. eIF4A, 4B ja 4fF, seondavad mRNA ribosoomi 40S subühikuga.
- Põhilised regulatsioonipunktid
- Polü(A) mõju translatsioonile
- Initsiaator-tRNA leidmine. eIF2 (seotud GTP-ga) seondab Met-tRNAi.
- 43S kompleksi assambleerimine. eIF2 toob initsiaator-tRNA 40S kompleksi ning mingil hetkel seondub eIF5, mida on vaja selleks, et eIF2 saaks vabaneda (stimuleerib GTP hüdrolüüsi).
- 43S kompleksi seondumine mRNA-ga. Ribosoomi väike subühik saab mRNA kompleksiga kokku. mRNA docking on the 43S - mRNA on ümber ribosoomi väikese subühiku kaela keeratud.
- mRNA skanneerimine AUG koodoni leidmiseks. Kogu kompleks hakkab liikuma ja otsima start-koodonit. eIF4E on siiani seotud CAP-struktuuriga, GTP hüdrolüüsi indutseerimisel lahkub eIF2.
- 80S kompleksi assambleerimine. eIF5B (GTPaas) toob ribosoomi suure subühiku ning sel ajal lahkuvad osad eIF-id (PILDILT). eIF5B ühendab subühikud, seejärel vabanevad initsiatsioonifaktorid. Tõenäoliselt jääb eIF4E siiski CAP-struktuuri külge ning samuti mRNA rõngas ei tohiks laguneda.
- cap-sõltuva ja sõltumatu translatsiooni initsiatsiooni võrdlus. Miks kasutada IRES -t ? CAP- dependent initiation. Initiation of translation usually involves the interaction of certain key proteins with a special tag bound to the 5'-end of an mRNA molecule, the 5' cap. The protein factors bind the small ribosomal subunit (40S), and these initiation factors hold the mRNA in place . The eukaryotic Initiation Factor 3 (eIF3) is associated with the small ribosomal subunit, and plays a role in keeping the large ribosomal subunit from prematurely binding. CAP-sõltumatu initsiatsioon. The best studied example of the cap-independent mode of translation initiation in eukaryotes is the Internal Ribosome Entry Site (IRES) approach. What differentiates cap-independent translation from cap-dependent translation is that cap-independent translation does not require the ribosome to start scanning from the 5' end of the mRNA cap until the start codon.
- Upstream ORF-ide mõju eukarüootsele translatsioonile. GCN4 süsteem.
Снижение
эффективности трансляции из-за наличия
uORF в 5’-UTR является хорошо изученным
эффектом; одним из примеров, иллюстрирующих
его, может служить ген поли(А)-полимеразы
α[en] (англ.
poly (A)-polymerase α, PAPOLA), чья мРНК
содержит две высококонсервативные uORF
в 5’-UTR. Мутация проксимального uAUG
вызывает повышение эффективности
трансляции этой мРНК, следовательно,
uORF значительно снижает экспрессию этого
гена. Другим примером является рецептор
тиреоидного гормона, оказывающий
активирующее или репрессирующее действие
на транскрипцию ряда генов-мишеней:
сильная репрессия его трансляции
осуществляется uORF длиной 15 нуклеотидов
внутри 5’-UTR его мРНК.
GCN4 on eukarüootne transkriptsionaalne aktivaator valk, mis seondub
sihtmärk DNA-ga dimeerselt.
- Translatsiooni kontroll üle eIF2.
eIF2 kineerimine peatab translatsiooni. eIF2
seondub fosforüleeritult eIF2B-le (mida on rakus ca 10x vähem).
Piisab kui fosforüleerida 1/10 eIF2, see seondub sel juhul eIF2B-le
ning enam ei toimu GTP/GDP vahetust → translatsiooni blokk.
Translatsiooni kontroll üle
eIF4E. cap ja polü(A) saba. Klassikaline initsiatsioonirada ja
faktorid (eIF1, eIF1A, eIF2, eIF3, eIF4A, eIF4B, eIF4E, eIF4G, eIF5,
eIF5B, PABP). Sisemine initsiatsioon, IRES. Valgusünteesi
regulatsioon initsiatsioonifaktorite kineerimise kaudu (näitena
eIF2, eIF4E-BP kineerimine).Ferritiini ja transferriini retseptori
süneesi regulatsioon.
iIF4-BP seondub rakus
eIF4E-le ning sel juhul pole rakus piisavalt eIF4E-d, et
translatsioon saaks alata. Fosforüleerides eIF4-BP-d, vabaneb selle
küljest eIF4E ning translatsiooni initsiatsioon saab jätkuda. Seega
eIF4E-eIF4-BP kompleksi kineerimine aktiveerib translatsiooni.
Translatsiooni initsiatsioon bakterites - - bakteriaalsed initsiatsioonifaktorid ja intsiaator tRNA. IF1(blokeerib Asaidi, ja suunab itRNA kohe P-saitistimuleerib IF2 ja IF3 ), IF2(seondab suunab itRNA P-saiti, GTPaas, ja aitab 70S vormeerimist.), IF3(ei anna võimalust ribosoomi subühikute dissotsieerumiseks, suunab mRNA startkoodoni Psaiti, aitab koodon-antikoodon interaktsioonile). itRNA- 5´esimene nukleotiid ei paardu 73 nukleotiidiga, antikoodon sõlmus on kolm G:C järjestust, Met on N-formüülMet.
Initsiatsioonil osalevad
komponendid:
- Initsiaator-tRNA, milleks on f-Met
- Aminohape Met on formüleeritud (D-loopis asetseva paari muutus on oluline).
- Siseneb ribosoomi P-saiti otse (mitte A-saiti);
- f-Met-tRNA on resistentne peptidüül-tRNA hüdrolaasile (ensüüm, mis juhul kui valgusünteesil juhtub midagi ja peptidüül-tRNA kukub ribosoomist maha sünteesitakse poolik valk vabastab peptidüül-RNA tRNA otsast). Esimese ja 72. aluspaari vahel puudub paardumine, mis tagab resistentsuse sellele ensüümile.
- - põhilised protsessid klassikalisel (SD-sõltuval) initsiatsioonil ja neile vastavad kompleksid. SD paikneb umbes 5-7 nukleotiidi kaugel start koodonist. SD- on AGGAGG, mis paardub vastava anti-SD järjestusega 16S rRnal.
- - initsiatsioonitüübid (SD- juhitud , SD-ta, liiderjärjestuseta, reinitsiatsioon
liiderregioonita mRNA seondub otse 70S
ribosoomile. Vajalik inits.tRNA olemasolu ning IF2
stimuleeribpaardumist. SD-sõltuv initsiatsioon – 30S subühikule
seondub IF3, siis teised, moodustub 30S PIC, pannakse mRNA paika,
toimub koodonantikoodon paardumine, on moodustunud 30S IC, seondub
50S ja eemaldub IF3, siis teised, on moodustunud 70S IC. IF2
hüdrolüüsi (GTP hüdrolüüs) tagajärjel läheb ribosoom üle
elongatsiooniliseks ribosoomiks. Järjekorrad pole 100% kindlad.
Reinitsiatsioon – algab terminatsioon, ribosoom
laguneb, aga enne kui mRNA ära läheb, tuleb uus IF3 ja võib uuesti
alustada. Kasutatakse siis kui mitu ORFi on järjest, võib-olla on
kahe ORFi vahel järjestused, mis seda soodustavad.
Translatsiooniline kontroll bakterites
==
- poomisvastus (stringent response) Aminohapete
nälja tingimustes pole võimalik tRNA-sid aminoatsüleerida. Kui
ribosoomi siseneb aminoatsüleerimata tRNA, aktiveerub ribosoomiga
seondunud valk RelA. RelA katalüüsib guanosiin tetrafosfaadi
(ppGpp) ja pentafosfaadi (pppGpp) sünteesi. (p)ppGpp omakorda
seondub RNA polümeraasiga, mis seepeale pole enam võimeline
transkriptsiooni initsieerima. Nii peatab
rakk energeetiliselt kuluka transkriptsiooni-translatsiooni
protsessi. - ribosoomi valkude
regulatsioon. Iseregileerivad. Mõned valgus seondavad operooniga,
kus paikneb nendee geen ja blokeerivad translatsiooni.
- - translatsiooni reguleerivad mRNA primaarjärjestuse elemendid : SD järjestuse ära tundmine ja seondamine.
- - translatsiooni ja transkriptsiooni attenuatsioon
- - ribolülitid. Üks võimalus translatsiooni reguleerimiseks mRNA sekundaarstruktuuri kaudu on ribolülitid (riboswitch). Ribolülitid koosnevad kahest struktuursest osast: aptameerist ja ekspressiooniplatvormist. Ribolüliti aptameerne domeen on mRNA struktuur, mis on võimeline seondama spetsiifilist metaboliiti. Ekspressiooniplatvorm on mRNA piirkond, mille kaudu metaboliidi seondamisest või mitteseondamisest tingitud muutused mõjutavad geeniekspressiooni. Enamasti osalevad ribolülitid negatiivses regulatsioonis madalmolekulaarse metaboliidi seondumisel aptameeriga blokeeritakse tema sünteesiks vajalike geenide transkriptsioon või translatsioon. Translatsiooni tasemel toimib geeniekspressiooni regulaatorina näiteks tiamiini ribolüliti, mis moodustab tiamiini või tiamiin-pürofosfaadi seondamisel sekundaarstruktuuri, mis varjestab SD ning takistab seega translatsiooni initsiatsiooni.
- - sRNA-d (valku siduvad , RNA-d siduvad). Kui sidub valku, siis muutub tema funktsiooni, kui mRNA, siis reguleerib geeni ekspressiooni. Võib olla cis ja trans.
- - toksiin -antitoksiinid ja trans-translatsioon. T-Anti I tüüp. mRNA translatsiooni kus paikneb toksiini geen on inhibeeritud väikese mittekodeeriva RNA antotoksiiniga seondumisega. Toxin-antitoxin systems and trans-translation. Koosnevad alati valgulisest toksiinist ja selle valgulise toksiini omadusi/võimeid neutraliseerivast antitoksiinist (valk või RNA). Toksiin on enamasti stabiilne, antitoksiin on ebastabiilne ning seda peab koguaeg juurde sünteesima.
- - 100S ribosoomid dimerisatsioon
Translatsioonilised GTPaasid - Mis on ortoloogid ja mis on paraloogid. Ortoloog - eri liikidest pärit järjestused, millel on üks ühine eellane. Paraloog - samast liigist pärit järjestused, millel on üks ühine eellane. geenid mis on tekkinud geeniduplikatsiooni tulemusena sama liigi piires
- Kas
- EF-Tu/eEF1A/aEF1A on ortoloogid/paraloogid
- IF2/eIF5B/aIF5B on ortoloogid/paraloogid
- EF-G/eEF2/aEF2 on ortoloogid/paraloogid
- RF3/eRF3 on ortoloogid/paraloogid
- Järgmiste valkude funktsioon
- EF-Tu/eEF1A/aEF1A-Toimetab aa-tRNAsid ribosoomi. aa-tRNA*EF-Tu*GTP on tuntud kui kolmikkompleks
- EF-G/eEF2/aEF2- EF-G'd kutsutakse ka "translokaasiks". Katalüüsib pep-tRNA liikumist A sidist P saiti. Samas liiguvad ka P saidis olev deats-tRNA E saiti ja seal olnud deats-tRNA lahkub ribosoomist. A sait vabaneb, valmis uueks koodoni äratundmiseks.
- SelB/eSelB/aSelB- Transpordib selenocysteinyl-tRNA ribosoomi.
- eIF2γ/aIF2γ -Arhetes ja eukarüootides viib fMet-tRNAfMet seondumist läbi
- RF3/eRF3- RF3 katalüüsib class I relese factors ite RF1 ja RF2 vabanemist peale terminatsiooni.
- Hbs1- Kui ribosoom "jääb kinni" - hangub
- Ski7- Peale stopp coodoni läbilugemist.
- Tet- Nad kaitsevad ribosoomi antibiootikum tetratsükliin eest.
- Translatsioon mitokondrites ja kloroplastides
Antibiootikumid - Antibiootikumide märklauad bakterirakus
- Translatsioon. Aminoglükosiidid (kanamütsiin, gentamütsiin, neomütsiin, streptomütsiin), Erütromütsiin, Spektinomütsiin, Kloramfenikool
- Transkriptsioon Rifampitsiin
- Foolhappe süntees. Sulfonüülamiid-inhibeerib foolhappe sünteesi
- DNA topoisomeraaside inhibitsioon Norfloksatsiin
- Rakukesta sünteesi inhibitsioon Penitsiliin, Vankomütsiin
- Antibiootikumide väljatöötamise ajalugu
- Sulfonüülamiidid. Esimest korda oli sünteesitud aastal 1908. 1932 – protnosiil. Gerhard Domagk.
- Penitsilliin - 1929, FLEMING
- Streptomütsiin- 1944, Albert Schatz, Selman Waksman
- Tetratsükliini mõju kolmikkompleksi seondumisele ribosoomiga. Tetratsükliin seondab Asaidi lähedusse ja pärsib tRNA seonduist ribosoomiga.
- Aminoglükosiidide mõju translatsiooni täpsusele
Aminoglükosiidid seonduvad
A-saiti väiksele subühikule. Vastupidiselt tetratsükliinile
seondavad aminoglükosiidid tRNA-d palju tugevamini ning seega ei
toimu translokatsiooni. Lisaks ei saa vale tRNA lahkuda A-saidist
ning tänu sellele suureneb valgusünteesi vigade sagedus.
Paromomütsiin tõstab
vigade sagedust peamiselt soodustades induced fit-i. Täpselt
interaktsioone ega detaile ei teata, aga tõhustatakse tRNA
seondumist.
- Makroliidide mõju kasvava peptiidi liikumisele väljumistunnelis
Makroliidid - laktoonrõngas,
mille küljes on erinevad suhkrujäägid.
Ribosoomi suure subühiku
PTC asub peptiidi kanali juures. Makroliidid seonduvad kanalisse PTC
ette ning sünteesitud valk ei suuda väljuda. tRNA ei suuda seonduda
korralikult ribosoomile ning kukub välja. Translatsioon peatub.
- Kloramfenikool ja peptidüültransferaasse reaktsiooni inhibeerimine
Chloramphenicol prevents
protein chain elongation by inhibiting the peptidyl transferase
activity of the bacterial ribosome. It specifically binds to A2451
and A2452 residues in the 23S rRNA of the 50S ribosomal subunit,
preventing peptide bond formation.
While chloramphenicol and the
macrolide class of antibiotics
both interact with ribosomes,
chloramphenicol is not a macrolide. It directly interferes with
substrate binding, whereas macrolides sterically block the
progression of the growing peptide.
- Pruomütsiin kui A-saidi substraadi analoog
Puromütsiin seondub
ribosoomi aktiivsaiti (ensüümi inhibiitor). A nukleotiidi analoog,
seondumine toimub
suurele subühikule.
- Antibiootikumiresistentsuse kolm mehhanismi
Kui meil mingi AB märklaud (ribosoom, topoisomeraas jne), kuid selle märklaua geenis toimub mingi muutus AB ei saa enam oma märklaulale seonduda. Tekib geeniresistentne variant.
Bakter omandab pumba, millega AB bakterirakust välja pumpab ja vähendab AB-de kontsentratsiooni rakus.
AB lihtsalt inaktiveeritakse. Nt penitsiliini puhul lõigatakse AB hüdrolüütilise ensüümi poolt katki. Mõnede teiste puhul ka nt fosforüleerimine, metüleerimine jne.
Geneetiline kood.
- Koodoni wobble positsioon (inosiin ja tRNA modifitseerimine).
Kolmandas positsioonis on koodoni kasutus lõdvem,
tänu millele võib tekkida seal rohkem vigu. Koodoni 1 ja 2 täht
olulisemad (seos ribosoomi dekodeeriva tsentriga). Kolmandas
positsioonis on puriin või pürimidiin. Koodon-antikoodon
äratundmisel toimub koodoni 1. ja 2. positsioonis WC tüüpi
aluspaardumine. Koodoni 3ndas positsioonis võib toimuda ka G-U
paardumine (nn „wobble“), mis lubab U-ga algava antikoodoniga
tRNA-del transleerida koodoni 3ndas positsioonis nii A kui G ja G
algavail antikoodonitel transleerida nii C kui U koodoneid.
Initsiaator tRNA puhul esineb bakteritel „wobble“ paardumine
koodoni esimeses positsioonis (GUG ja UUG on sagedased 10%).
tRNAde väiksema arvu jaoks kasutatakse
mitokondrites kolmandas positsioonis tihti inosiini, mis on guaniini
sarnane. Lisaks on antikoodoni kõrval metülatsioon ja
pseudouridiin.
Wobble paardumine. Inosiin
esineb tRNA-s ja on vajalik wobble aluspaaride õigeks
translatsiooniks. Inosine (I) at position 34 (wobble position) of
tRNA is formed by the hydrolytic deamination of a genomically encoded
adenosine (A) (reaktsiooni katalüüsib tRNA A:34 deaminaas). Adeniin
muudetakse kas adenosiin-/inosiinmonofosfaadiks, mis järgnevalt
muudetakseinosooniks.
- Alternatiivsed geneetilised koodid (mitokondrite erinevad koodid, alternatiivsed koodid bakteritel, pärmidel)
- Mitokondris:
- AGA - Ter* (Arg)
- AGG - Ter* (Arg)
- AUA - Met (Ile)
- UGA - Trp (Ter*)
Muutused koodis ehk koodoni tähenduse muutused, saavad toimuda vaid
siis kui koodon esineb vaid vähestes geenides. Alternatiivsed
koodid on peamiselt mitokondrites ja plastiidides, st väga väikestes
genoomides. Pärmi Candida
mõnedel liikidel (nt albicans)
on CUG koodon kas kahetähenduslik või vastab seriinile (Leu
asemel). Mycoplasma capricolum
kasutab ainult 56 koodonit, st 5 koodonit valgu geenides ei esine.
- Selenotsüsteiin ja pürrolüsiin (esinemine, koodonid ja valku lülitumise teed)
Koodonikasutamise indeksid (CAI - codon adaptation index)
Programmeeritud raaminihe (RF2, retroviirused)
Cai – igale koodonile antakse arvuline väärtus
vastavalt tema esinemisele sagedastes valkudes. Kõige kasutatavam
koodon saab väärtuse 1, teiste kasutatavus arvutatakse selle järgi.
kõigi alternatiivsete koodonite kasutuse hulgast võetakse
geomeetriline keskmine, mis ongi CAI. Terminatsioonifaktor II (RF II)
tunneb ära UGA koodoni. Kui RF II-te on rakus vähe, siis toimub +1
raaminihe ja sünteesitakse RF II-te juurde. Enne seda on SD
järjestus, et ribosoom oleks veits aeglasem.
HIVis kapsiidi ja polümeraasi valgud kattuvad,
aga kapsiidi on vaja umbes 100 korda rohkem. Transkribeeritakse üks
mRNA. Enamus ribosoome transleerib kapsiidivalgu. Aga mõni ribosoom
nihkub ühe nukleotiidi võrra tagasi (-1 raaminihe) ja
terminatsioonikoodonit enam pole ja sünteesitakse liitvalk, mis
hiljem üksteisest lahti lõigatakse. Raaminihke järel on
pseudosõlme struktuur, kus ribosoom ei suuda läbi sõita, ja tänu
sellele toimubki nihkumine. Pseudosõlm on see, kui juuksenõela
paardumata osa paardub mingi edasise ahela osaga.
Valkude väljumine ribosoomist.
- Kasvava peptiidi tunnel
Geeniekspressiooni regulatsioon tunnelis (SecM, TnaC)
Valgu modifitseerimine ribosoomil (metioniini eemaldamine, atsetüleerimine)
Molekulaarsed chaperonid
Valkude transport endoplasmaatilisse retiikulumi
Seinas ei ole laetud AH, hüdrofoobseid AH.
Laenguta hüdrofiilsed ühendid, et midagi kinni ei jäeks. Me ei tea
mis konformatsioonis kasvav peptiid tunnelis on, kuid teame, et see
on piisavalt peenike, et enne tunneli lõppu ei saa toimuda migit
voltumist (võib ehk olla sunnitud alpha-heeliksina). Tunnelisse
mahub 35-45 AH. Enamus peptiide mahuvad tunnelist läbi, saab
kasutada regulatsioonis (geeniekspressiooni omas nt). Bakterites on
valgu ekspordisüsteem. SecYEG (sekretsioon), SecA (ATPaas, tunneb
ära kasvava peptiidi N-terminuses oleva ekspordisignaalijärjestuse,
hakkab kotranslatsiooniliselt valku rakust välja pressima (tõmbab
ribosoomist ka välja, lükkab SecYEG poori)). Ribosoomist väljuvat
peptiidi modifitseeritakse kotranslatsiooniliselt.
A. Nii kui peptiid pistab nina välja, võetakse
formüülrühm ära, samuti võetakse Met ära N-terminuses.
B. Met võidakse ära võtta, aga ei pea.
Atsetüülrühma viimine küpse valgu N-terminuses (enamustes
valkudes) – funktsiooni ei teata, aga nii juhtub.
N-atsetüültrasnferaas teostab oma tööd juba ribosoomi peal. PDF
(peptiidi-de-formülaas) seondub L22 külge. Modifikatsiooni
teostavad ensüümid, mis on otseselt ribosoomiga seotud. Chaperon –
valk mis hoiab ära väljuva/kasvava peptiidi ebasoovitavaid
interaktsioone. Erinevaid chaperone, mõned on fülogeneetilise rühma
spetsiifilised, osad mitte; osad spetsiifilised kindlatele
peptiididele, osa universaalsed .
ÜLDISTE CHAPERONIDE PEREKONNAD:
● DNA K HSP 70 On kõige
paremini kirjeldatud väike (70 kDa) chaperon. Vajab Hsp40 abi, mis
suurendab ATP kasutatavust ja Hsp70 aktiivsust. On leitud, et Hsp70 suurendatud ekspressioon rakkudes vähendab apoptoosivõimalust.
● DNA J HSP 40
● GRO EL HSP 60 Kõige paremini
uuritud suur ( 1 MDa) chaperonide kompleks. On molekulaarseks
chaperoniks ka mitorkondris.
● GRO ES HSP 10
● HTP G HSP 90 Saadakse sellest
kõige vähem aru. 90 kDa, vajalik eukarüootides normaalseks
eluvõimeks (arvatavasti ka prokarüootides). Igal on ATP-siduv
domeen, keskmine domeen ja dimerisatsiooni domeen.
Chaperonid vaatavad kasvava peptiidi hüdrofoobseid
regioone, mille järgi saavad aru, et peavad tööle asuma.
ER-i läheb kotranslatsioonilises. Signaalpeptiid
on N-terminuses. Peptiid tuleb tunnelist välja, selle tunneb ära
(RNA-st ja valkudest koosnev kompleks) – SRP (signal recognising
particle), paneb translatsiooni seisma kui signaali ära tunneb, viib
ribosoomi kasvava peptiidiga ER-i juurde, kompleks eemaldub ning
ribosoom laseb kasvava peptiidi otse ER-i.
RNAi
- RNAi üldiseloomustus : komponendid, mehhanism, tagajärjed
RNA interferents on süsteem elavates rakkudes,
mis osaleb geeniaktiivsuste määramisel. RNA interferentsiks on
olulised kaks tüüpi väikeseid RNA molekule: mikroRNA ( miRNA ) ja
väike interfereeriv RNA ( siRNA ). Väikesed RNA-d võivad seonduda
teistele RNA-dele, tõstes või alandades nende aktiivsust (nt
seondudes mRNA-le, takistavad nad sellest toimuva translatsiooni. RNA
interferentsil on oluline roll rakkude kaitses parasiitsete geenide
vastu, organismi arengu suunamises ja geeniekspressioonis üldiselt.
- Dicer ja Argonaute : nende valkude funktsioon, olulisemad domeenid ja toimemehhanism
- Dicer on RNaas III perekonna endoribonukleaas, mis lõikab dsRNA-d and pre-mikroRNA-d lühikesteks 20-25 bp pikkusteks dsRNA- deks (2 bp jääb 3’-otsast üle ulatuma). Dicer koosneb 3 domeenist: 2 RNaas III domeeni ning üks PAZ domeen. Interageerub abivalkudega.
- Argonaut on RISC kompleksi katalüütiline komponent. Seondab endaga erinevate klasside väikseid mitte-kodeerivaid RNA-sid (miRNA, siRNA, piRNA). Väikesed RNA-d juhivad agronaudi valgud komplementaarsete mRNA-de juurde. Osadel agronautidel on endonukleaasne aktiivsus, mis lõikab mRNA-d, lisaks osaleb siRNA passenger-ahela degradatsioonis.
- Kolm rühma: argonaudi-sarnased AGO1; piwi-like proteins, group 3
- mikroRNA ja siRNA biogeneesi võrdlus
miRNA
siRNA
Toimumine
Looduslikult esineb taimedes ja loomades .
Looduslikult esineb taimedes ja osades loomades.
Esinemine
Üheahelaline.
Kaheahelaline.
Pikkus
19-25 nt.
21-22 nt.
Sihtmärk-mRNA komplementaarsus
Mitte väga suur, tänu millele võib üks miRNA seonduda sadade mRNA-dega.
100%, igal siRNA-l on kindel sihtmärk (mõni erand võib leiduda).
Biogenees
Ekspresseeritakse miRNA geenidelt, reguleerivad teiste geenide mRNA-sid.
Reguleerivad enda ekspressioonigeene.
Tegevus
mRNA translatsiooni inhibeerimine.
Lõikavad mRNA-d.
Funktsioon
Geenide regulatsioon (inhibeerimine).
Nn geenivaigistamise valvurid.
Kasutatavus
Possible therapeutic uses either as drug targets or as drug agents themselves. Expression levels of miRNAs can be used as potential diagnostic and biomarker tools .
siRNAs are valuable laboratory tools used in nearly every molecular biology laboratory to knock down genes. Several siRNAs are in clinical trials as possible therapeutic agents.
- siRNA rakku viimise meetodid, nende eelised ja puudused
- lipiidimitsellis
- rakulises vesiikulis
- viiruste sisse pakitult
- lühemad kui 14 lähevad iseseisvalt läbi raku
- bakteritega
- RNAi kasutusvõimalused meditsiinis ning sellega seotud probleemid
- Geenide funktsionaalsed uuringud - siRNA abil surutakse alla uuritava geeni või tema partneri ekspressioon. Lülitades välja geen on võimalik jälgida muutusi rakkudes. Võimalik on vaadata näit. mõju geeniekspressioonile, valkude modifikatsioonidele, mingitele kindlatele protsessidele rakus – näiteks apoptoos, transport jne.
- Bakterite raamatukogu, kus erinevad kloonid toodavad erinevaid pikki kaheahelalisi RNA-sid, lisatakse toidule.
- Drosophila rakud võtavad RNA söötmest ise sisse. Organismi tasemel süstimine embrüodesse.
- Transfektsioon või nakatamine retroviirusvektorite raamatukoguga, mis sisaldavad erinevaid shRNA-sid.
- Funktsionaalsete partnerite ja märklaudgeenide otsing - geneetika ja funktsionaalne genoomika.
- Kasulikud transgeenid. Viirusvastaseid siRNA vektoreid kandvad haigusresistentsed taimed. Putuk-kahjurite vastast RNAi vektorit kandvad transgeensed taimed. Seemneteta tomatid.
- Geeniteraapia, haiguste diagnostika. RNAi induktsioon in vivo imetajatel. Peamine probleem praegu: kuidas viia siRNA-d õiges koes või vähis märklaud-molekulini kõrvalmõjusid tekitamata?
- Bakteriaalsed minirakud.
- Kolesterooliga pakitud nanopartiklid (+ligand).
- Raku membraani ise läbivate peptiididega konjugeeritud siRNA-d.
- siRNA-d või miRNA inhibiitorid , mis on lühemad kui 14 nt lähevad ise imetaja rakkudesse sisse.

















































Kõik kommentaarid