unikaalne tunnus. See on kodeeritud vastava DNA lõigu ehk geeni nukleotiidide järjestusega. Valgu AH järjestuse kaudu realiseerub geneetiline informatsioon. Järjestust loetakse alati amino-ehk N-terminusest karboksüül-ehk C-terminus suunas. Monomeerne valk – üks polüpeptiidahel. Multimeerne valk – mitu polüpeptiidahelat. Lihtvalk on ehitatud ainult aminohapetest. Liitvalgus esineb veel täiendav rühm, milleks võib olla sahhariidi, rasvataolise või mõne muu lihtsama aine molekul, millega omakorda võivad olla seotud metalliioonid. Kui mitte-AH osa on funktsionaalselt oluline, siis nim seda prosteetiliseks rühmaks. Sõltuvalt mitte-AH osast jaotatakse valgud glüko-, lipo-, nukleo-, fosfo-, metallo-, hemo- ja flavoproteiinideks. Valkude arhidektuuris eristatakse nelja struktuuritasandit: - Primaarstruktuur – aminohappe järjestus polüpeptiidis – kovalentsed (peptiid-ja disulfiid-) sidemed
5 molekuli. Lehekülg RNA transkriptsioon on matriitssüntees, mille käigus saadakse DNA molekuli ühe ahela nukleotiidse järjestusega komplementaarne mRNA molekul Nukleotiid 2. Nukleiinhapete komponendid. Pürimidiinid: Tsütosiin (DNA, RNA); Uratsiil (RNA); Tümiin (DNA) Puriinid: Adeniin; Guaniin. Suhkrud: Riboos/desoksüriboos. Fosfaat. Komplementaarsus on kindlate
Tsüklilised nukleosiidmonofosfaadid. Nukeosiiddi- ja trifosfaadid: roll, ehitus, fosfoanhüdriidsidemed. Nukleosiid = N-alus + suhkur Nukleotiid ehk nukleosiidfosfaat(NTP) = N-alus + suhkur + 1-3 fosforüülrühma Enamik vabu nukleotiide on ribonukleotiidid, millel fosforüülrühm on riboosis 5'-asendis. Nukleotiidid on mitme prootiniga happed. NTPd on koensüümide komponendid; energiakandjad ATP on keskne molekul energia metabolismis; GTP on peamine energiaallikas valgu sünteesis; CTP on oluline metaboliit fosfolipiidide sünteesis; UTP aktiveerib suhkrud polüsahhariidide sünteesis. Nukleotiideide kooseisu kuuluvad N-alused toimivad äratundmisühikutena, ise nad metabolismis ei osale. 4. DNA primaarstruktuur- polünukleotiidahela ehitus, 3' ja 5' otsa tähendus, fosfodiesterside. Ahela lihtsustatud (skemaatiline) esitusviis. Miks on DNA hape?
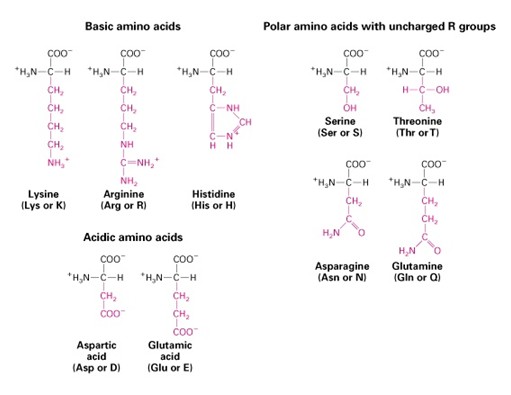
järjestusest. Aminohapete kõrvalahelate erinevad omadused tingivad nende erinevad konformatsioonid vesilahustes Genoom kodeerib 20 aminohapet Valkude 3M struktuur-konformatsioon-on determineeritud tema aminohappelise järjestusega Valgu struktuur määrab tema bioloogilise aktiivsuse (funktsiooni) Liht ja liitvalgud: · Lihtvalk on ehitatud ainult aminohapetest. · Liitvalgus esineb veel täiendav rühm, milleks võib olla sahhariidi, rasvataolise või mõne muu lihtsama aine molekul, millega omakorda võivad olla seotud metalliioonid. Sekundaarsruktuur Valgu sekundaarstruktuur kirjeldab, kuidas polüpeptiidahel ennast ruumiliselt paigutab. · Enamlevinud struktuurid on: heeliks, kus valk on keerdunud spiraalina; voldik, kus ahela osad paiknevad kõrvuti. · Sekundaarstruktuuri hoiavad koos erinevate aminohappejääkide vahelised vesiniksidemed. Valkude sekundaarstruktuurid heliks 3,6 aminohappejääki 1 pööre 360o
esinemisest tingitud omadused. Peptiid- ehk amiidsideme omadused: · Planaarne ehk tasandiline · Sarnane kaksiksidemele (elektronide resonants) · Pöörlemine ümber sideme on takistatud · Pikkus 1,32 Å, mis on üksik- ja kaksiksideme vahel · Puudub laeng. III. VALGU STRUKTUUR, ISELOOMUSTUS JA BIOLOOGILINE ROLL. (Õpik lk 45-58) 1. Üldine iseloomustus molekulide suurus, makrostruktuur, mono- ja oligomeerse valgu mõiste. Peptiid on molekul, mis koosneb kahest või enamast aminohappejäägist, mis on omavahel ühendatud peptiidsidemetega. Sõltuvalt jääkide arvust nimetatakse peptiide: 2 jääki dipeptiid 3 jääki tripeptiid 12-20 jääki oligopeptiid 20 -100 jääki polüpeptiid >100 jääki Mr>10 000, valk Valgud koosnevad ühest või mitmest polüpeptiidahelast: Monomeerne valk üks polüpeptiidahel; Multimeernevalk mitu polüpeptiidahelat 2. Lihtvalgud ja konjugeeritud e
Glükolüüs Glükolüüs on ensümaatiliste reaktsioonide ahel, mille käigus glükoos muudetakse püruvaadiks. Glükolüüs on raku tsütoplasmas kulgev universaalne ainvahetusrada, anaeroobsetes rakkudes ainus ATP tootev rada. Aeroobsetes rakkudes esimeseks etapiks süsivesikute oksüdatsioonil. Glükoos + 2 ADP + NAD + 2 P 2 Püruvaat + 2 ATP + 2 NADH + 2 H2O Glükolüüsi lähtesubstraadiks on glükoos, mis siseneb rakku transportvalgu abil. Glükolüüsi I faasis glükoosi molekul muundatakse viie keemilise reaktsiooni abil kaheks glütseeraldehüüd- 3-fosfaadiks. I faasis tarbitakse 2 ATP molekuli. Glükolüüsi II faasis muundatakse glütseeraldehüüd-3-fosfaat püruvaadiks, kasutakase 1,3-BPG ja PEP. Lisaks püruvaadile tekib II faasis ka ATP ja NADH. 1 molekuli glükoosi kohta tekib 4 molekuli ATP ja 2 molekuli NADH. Enamik reaktsioone glükolüüsis omavad Gibbsi vabaenergiat, mis on ligilähedane nullile.
Kontrolli, kas tead järgmiseid mõisteid ja termineid I 1. Kas oskad nimetada kõiki loengus loetletuid funktsionaalrühmi 2. Kas oskad nimetada eesliiteid, mida kasutatakse sageli biokeemiliste suuruste iseloomustamisel 3. Kas oskad ära tunda D ja L isomeere, R ja S isomeere Hüdrofiilne – suures osas polaarsete või iooniliste rühmadega ühend, moodustab veega sidemeid. Hüdrofoobne – suures osas mittepolaarsete rühmadega molekul, ei ole veega olulist vastastikmõju. Amfipaatne – molekul, millel on eristatavad hüdrofiilsed ja hüdrofoobsed osad. Elektronegatiivsus – aatomi võimekus endaga elektrone liita (madala elektronegatiivsusega aatom loovutab kergelt oma elektronid). Vesinikside – keemiline side, mis moodustub liigsete elektronidega (- laeng või osalaeng) elektronegatiivse aatomi ning vaba orbitaaliga (kasvõi osaliselt, st + osalaeng) vesiniku
Milles seisneb geenide positsiooniefekt? Geenide kohasõltuv avaldumine. Kui heterokromatiini piirkonnas olev DNA tuumas oma asukohta vahetab ja liigub eukromatiini alasse, võib see algatada selle DNA transkribeerimise. Sama efekt esineb ka eukromatiini puhul – kui ekspresseeruv geen viia eukromatiini piirkonnast üle heterokromatiini piirkonda, geen vaigistatakse. Millised on kolm kromosoomi säilitamiseks vajalikku struktuurielementi? Et DNA molekul oleks funktsionaalne ehk võimeline edukalt tütarrakkudesse kanduma, vajab ta kolme strukturaalset ühikut – telomeeri, tsentromeeri ja replikatsiooni alguspunkte Replikatsiooni alguspunkt – milleks vajalik, erinevused pro- ja eukarüootidel. Struktuur, mis oma järjestuselt pole küll korduselement, aga on vajalik kromosoomi kui terviku toimimiseks. Prokarüootidel esinevad kindlad DNA järjestused, kuhu DNA polümeraas seostub. Eukarüootidel

























Kõik kommentaarid