Valgu struktuur (2)
VALKUDE STRUKTUUR
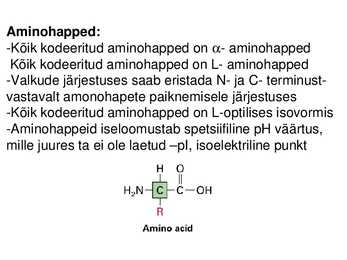
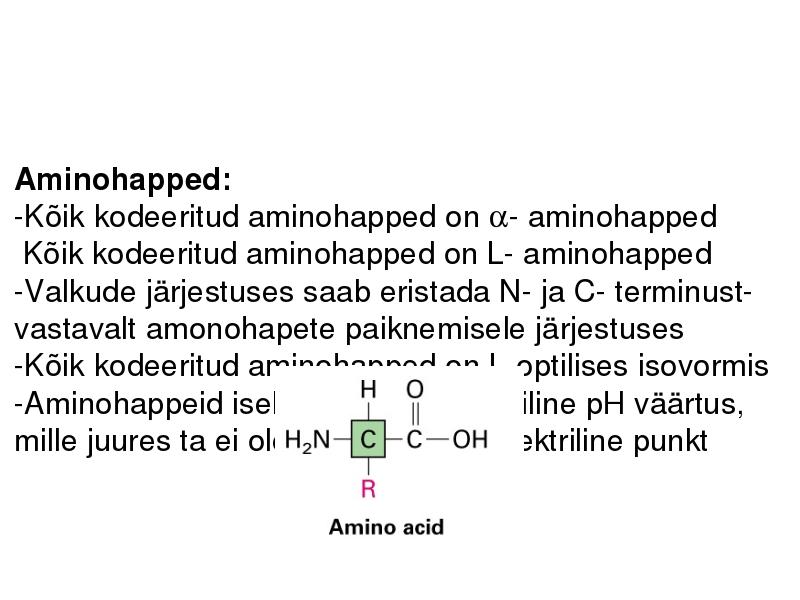
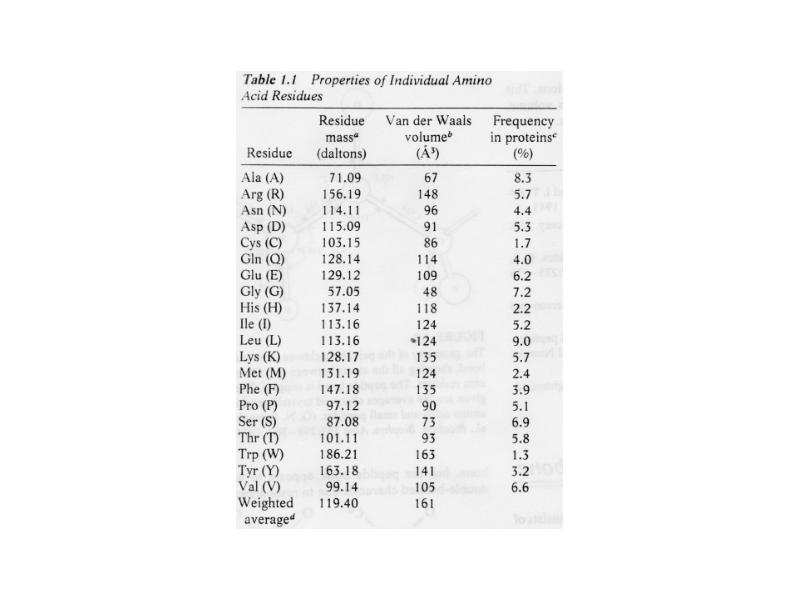
Aminohapped:
-Kõik kodeeritud aminohapped on - aminohapped
Kõik kodeeritud aminohapped on L- aminohapped
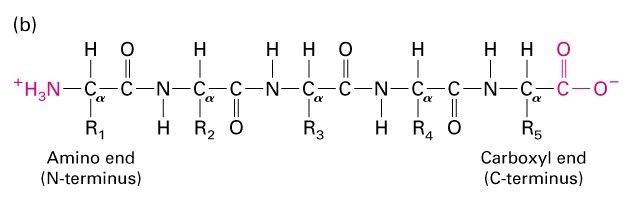
-Valkude järjestuses saab eristada N- ja C- terminust-
vastavalt amonohapete paiknemisele järjestuses
-Kõik kodeeritud aminohapped on L-optilises isovormis
-Aminohappeid iseloomustab spetsiifiline pH väärtus,
mille juures ta ei ole laetud pI, isoelektriline punkt
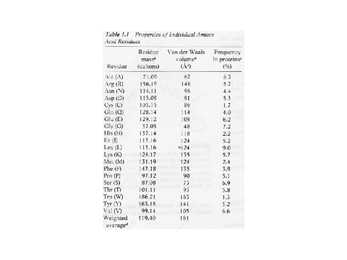
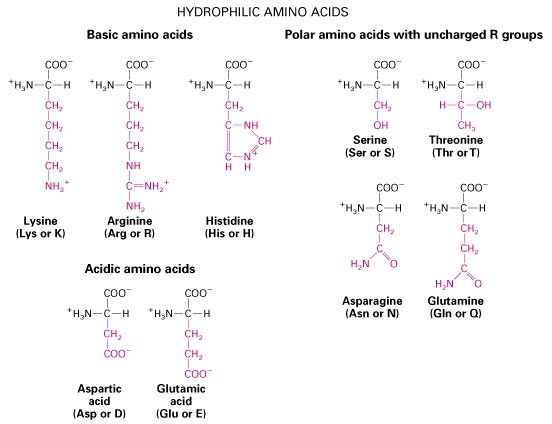
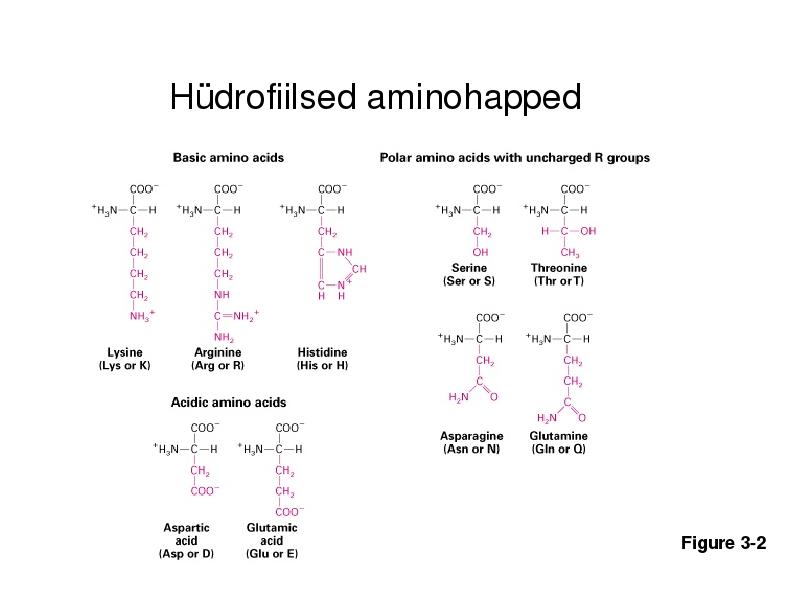
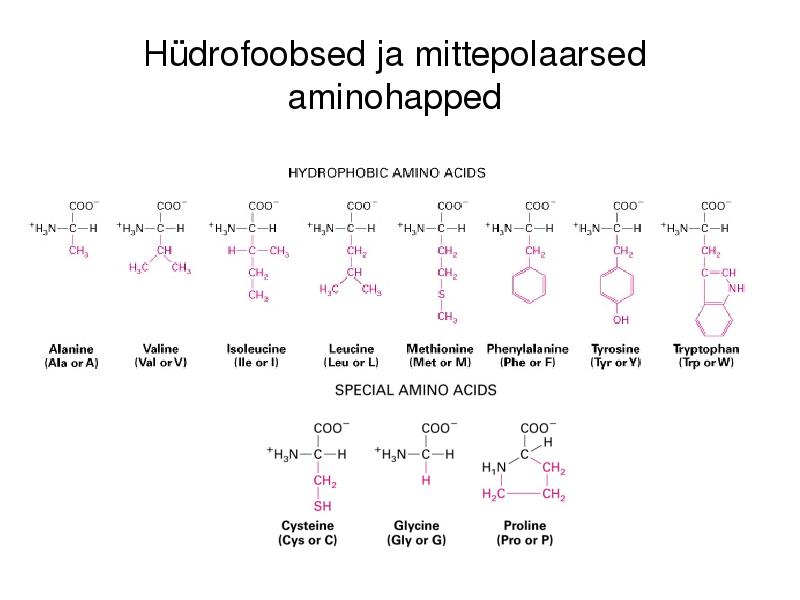
Hüdrofiilsed aminohapped
Figure 3-2
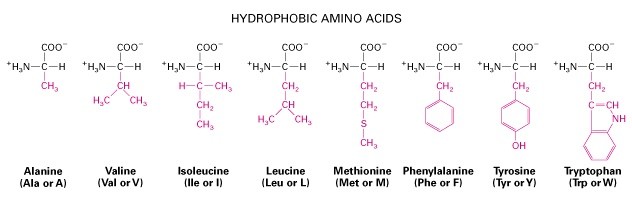
Hüdrofoobsed ja mittepolaarsed
aminohapped
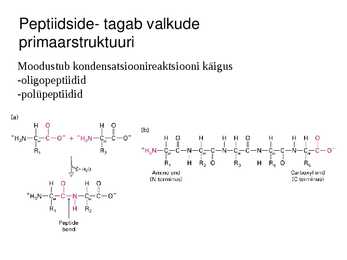
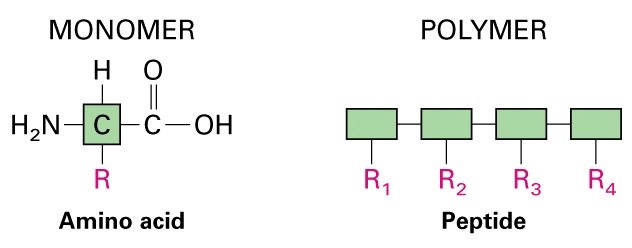
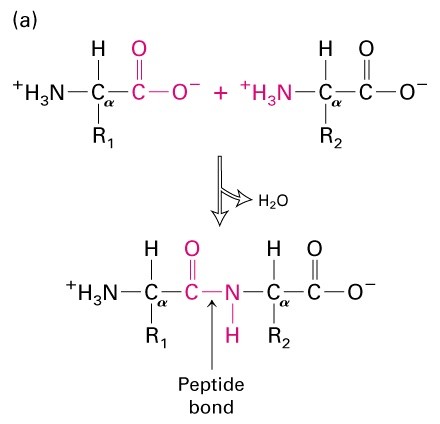
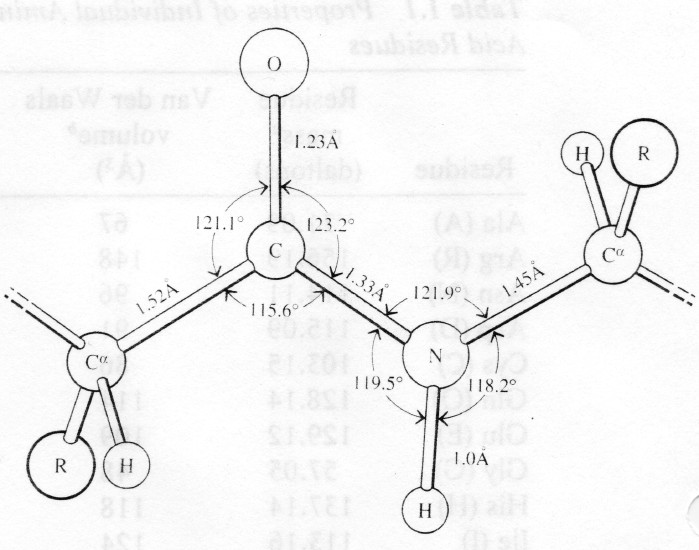
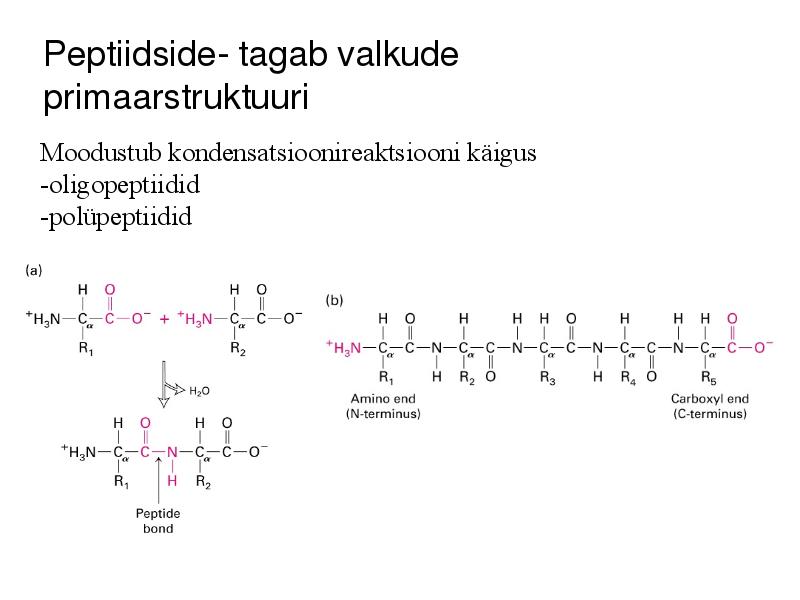
Peptiidside- tagab valkude
primaarstruktuuri
Moodustub kondensatsioonireaktsiooni käigus
-oligopeptiidid
-polüpeptiidid
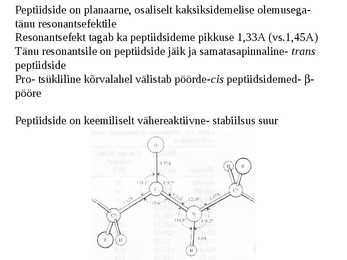
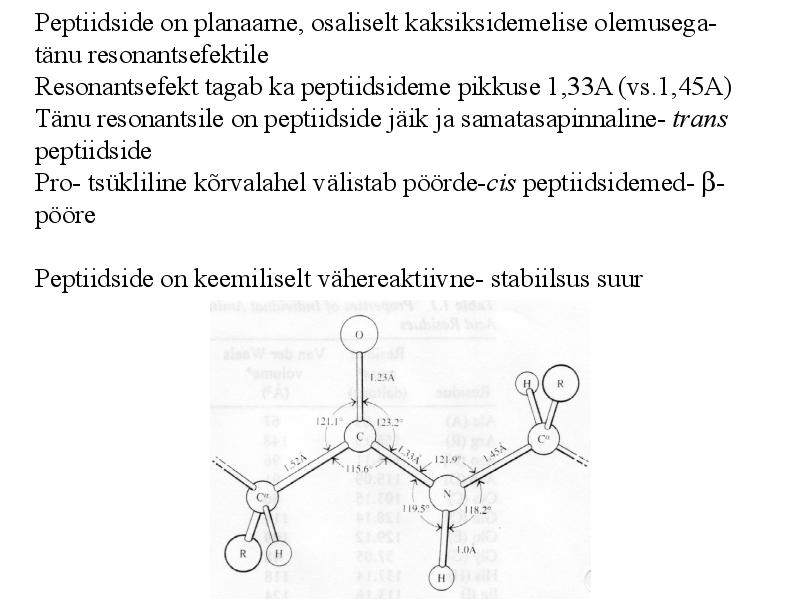
Peptiidside on planaarne, osaliselt kaksiksidemelise olemusega-
tänu resonantsefektile
Resonantsefekt tagab ka peptiidsideme pikkuse 1,33A (vs.1,45A)
Tänu resonantsile on peptiidside jäik ja samatasapinnaline- trans
peptiidside
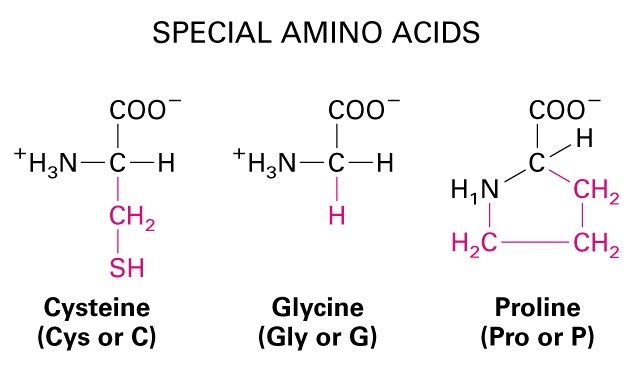
Pro- tsükliline kõrvalahel välistab pöörde-cis peptiidsidemed- -
pööre
Peptiidside on keemiliselt vähereaktiivne- stabiilsus suur
Polüpeptiid aminohapete järjestus
Peptiid aminohapete järjestus, millel puudub selgelt
defineeritud kolmemõõtmeline struktuur (praktikas
on sellised järjestused lühemad kui 100
aminohapet).
Valkdefineeritud 3 mõõtmelise struktuuriga
polüpeptiidahelad, sageli olemas ka
kvaternaarstruktuur
VALKUDE STRUKTUUR JA FUNKTSIOON
Valgud on hargnemata polümeerid, mis moodustuvad peptiidsideme
vahendusel aminohapetest
Valkude funktsionaalsus on tagatud nende kolmemõõtmelise
struktuuriga, mis omakorda tuleneb valkude aminohappelisest
järjestusest.
Aminohapete kõrvalahelate erinevad omadused tingivad nende
erinevad konformatsioonid vesilahustes
Genoom kodeerib 20 aminohapet
Valkude 3M struktuur-konformatsioon-on determineeritud tema
aminohappelise järjestusega
Valgu struktuur määrab tema bioloogilise aktiivsuse (funktsiooni)
Liht ja liitvalgud:
· Lihtvalk on ehitatud ainult aminohapetest.
· Liitvalgus esineb veel täiendav rühm,
milleks võib olla sahhariidi, rasvataolise või
mõne muu lihtsama aine molekul, millega
omakorda võivad olla seotud metalliioonid.
Sekundaarsruktuur
Valgu sekundaarstruktuur kirjeldab, kuidas
polüpeptiidahel ennast ruumiliselt paigutab.
· Enamlevinud struktuurid on:
heeliks, kus valk on keerdunud spiraalina;
voldik, kus ahela osad paiknevad kõrvuti.
· Sekundaarstruktuuri hoiavad koos erinevate
aminohappejääkide vahelised vesiniksidemed.
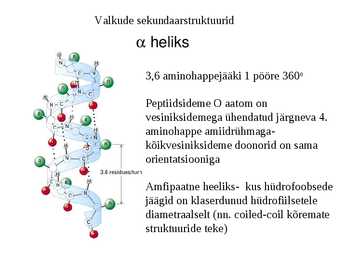
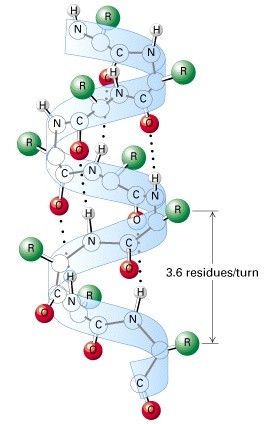
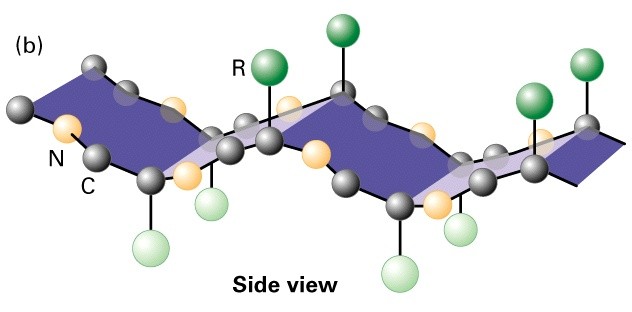
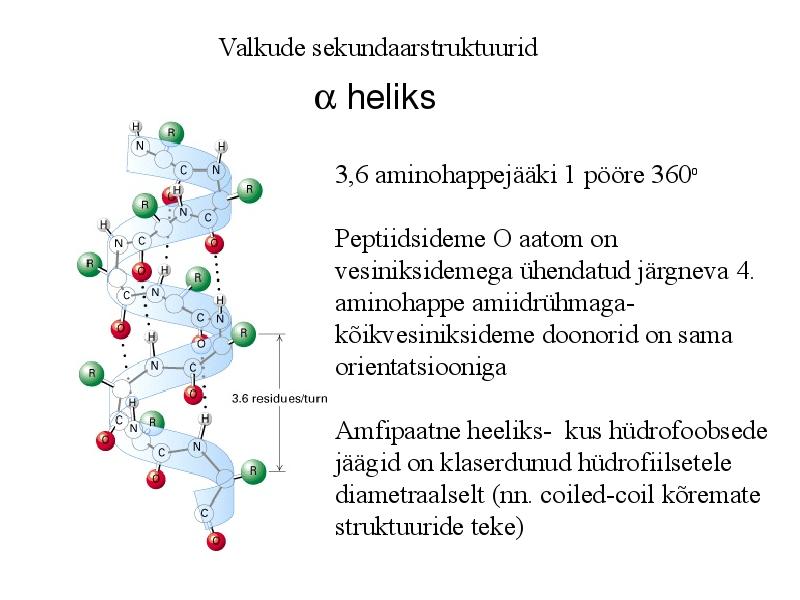
Valkude sekundaarstruktuurid
heliks
3,6 aminohappejääki 1 pööre 360o
Peptiidsideme O aatom on
vesiniksidemega ühendatud järgneva 4.
aminohappe amiidrühmaga-
kõikvesiniksideme doonorid on sama
orientatsiooniga
Amfipaatne heeliks- kus hüdrofoobsede
jäägid on klaserdunud hüdrofiilsetele
diametraalselt (nn. coiled-coil kõremate
struktuuride teke)
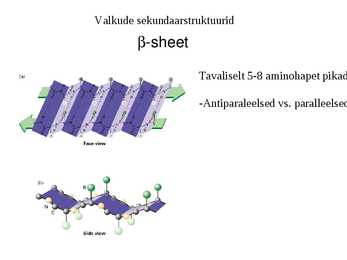
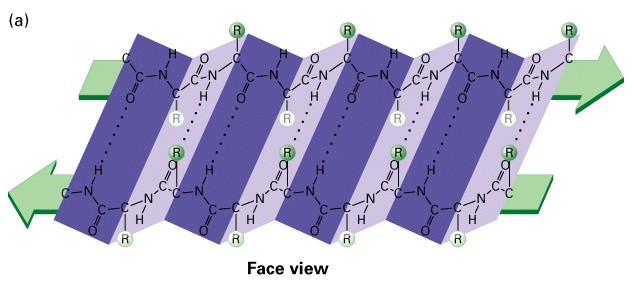
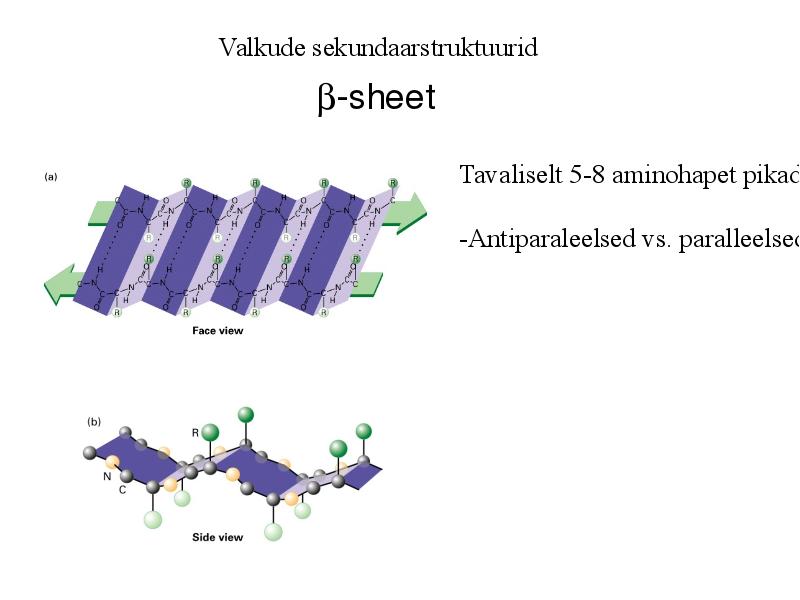
Valkude sekundaarstruktuurid
-sheet
Tavaliselt 5-8 aminohapet pikad
-Antiparaleelsed vs. paralleelsed
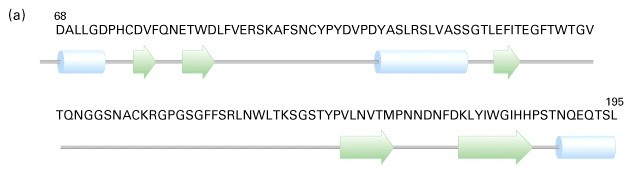
Hemaglutiniini primaar ja sekundaarstruktuur
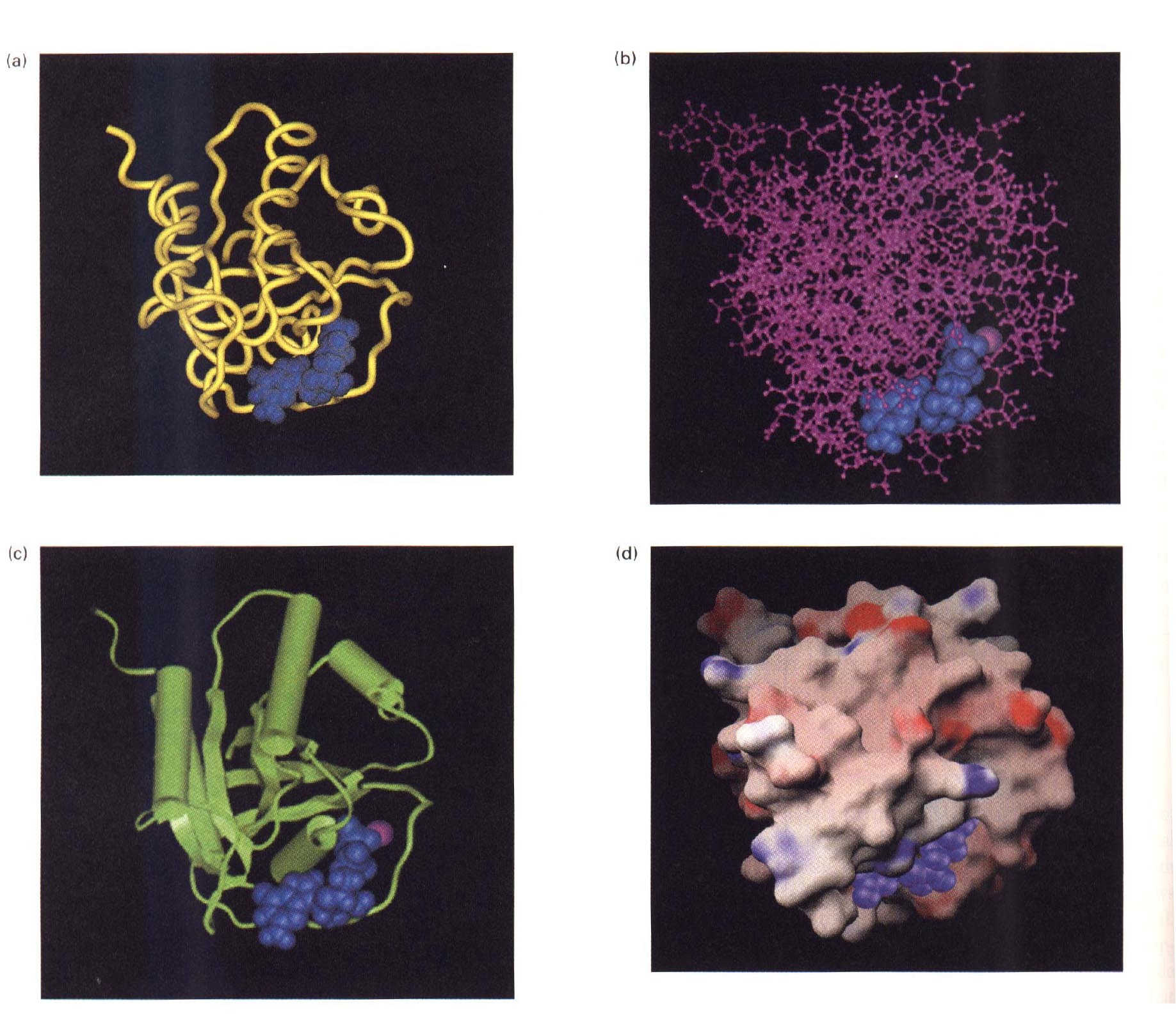
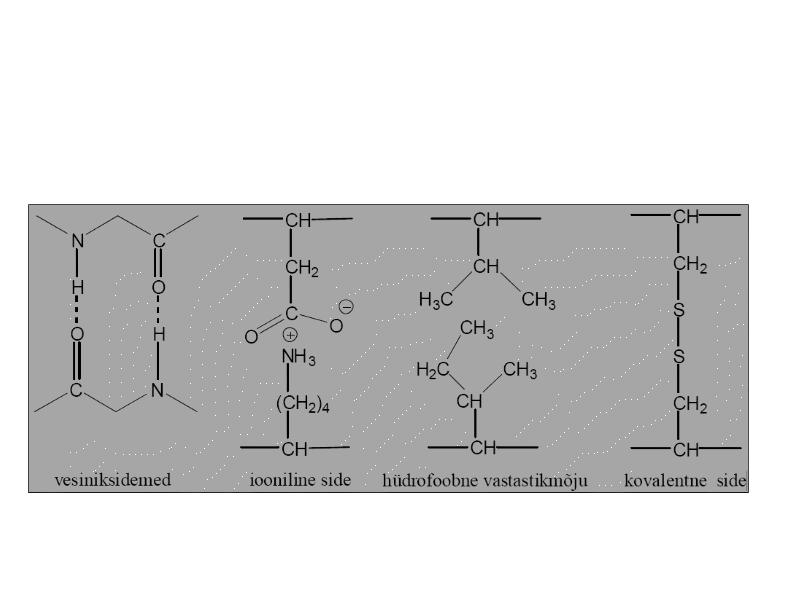
Terstiaarstruktuur
Valgu tertsiaarstruktuur kirjeldab, kuidas
paiknevad ruumiliselt polümeeri erinevad
osad (heeliksid ja voldikud).
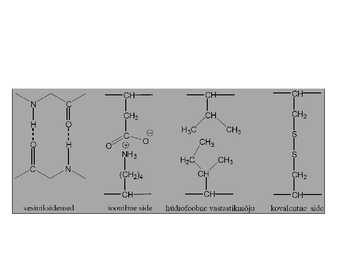
Interaktsioonid, mis stabiliseerivad valkude
kõrgemad struktuurid
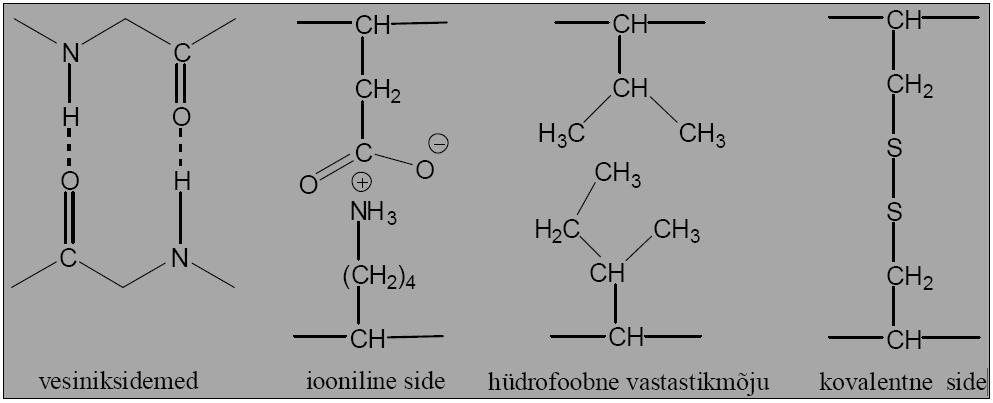
Vesiniksidemed polaarne interaktsioon, kus
elektropositiivne H on jagatud kahe elektronegatiivse
aatomi vahel (0,30nm)
Ioonsidemed elektrostaatilised interaktsioon
erilaenguliste aatomite vahel (0,25nm)
Van der Waals interaktsioonid kahe kõrvutipaikneva
aatomi elektronpilve fluktuatsioonidest tulenev jõud
(0,35nm)
Hüdrofoobne interaktsioon kahe hüdrofoobse
kõrvalahela vahel
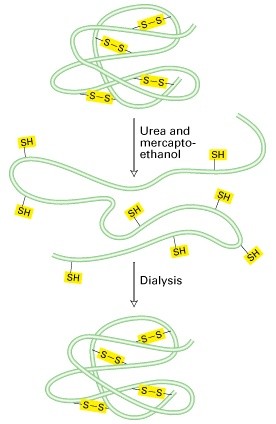
Disulfiidsidemed kovalentsed sidemed Cys vahel
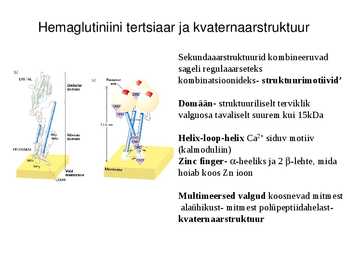
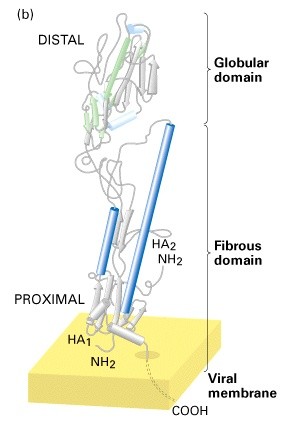
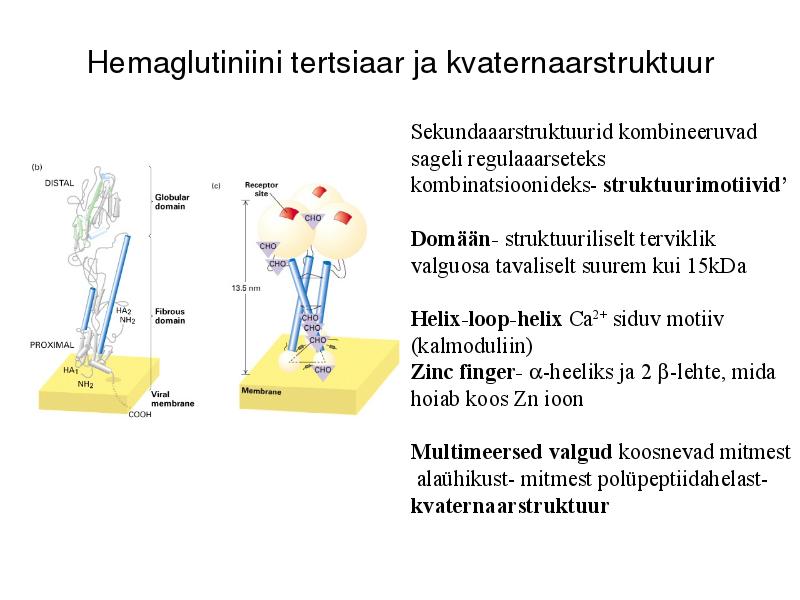
Hemaglutiniini tertsiaar ja kvaternaarstruktuur
Sekundaaarstruktuurid kombineeruvad
sageli regulaaarseteks
kombinatsioonideks- struktuurimotiivid'
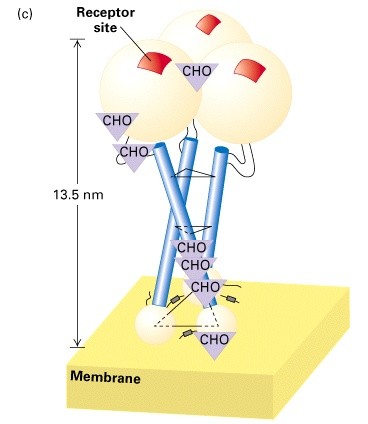
Domään- struktuuriliselt terviklik
valguosa tavaliselt suurem kui 15kDa
Helix-loop-helix Ca2+ siduv motiiv
(kalmoduliin)
Zinc finger- -heeliks ja 2 -lehte, mida
hoiab koos Zn ioon
Multimeersed valgud koosnevad mitmest
alaühikust- mitmest polüpeptiidahelast-
kvaternaarstruktuur
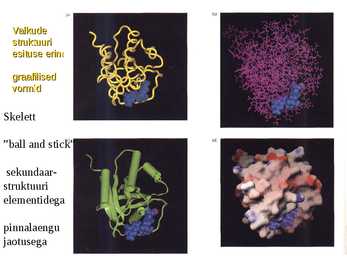
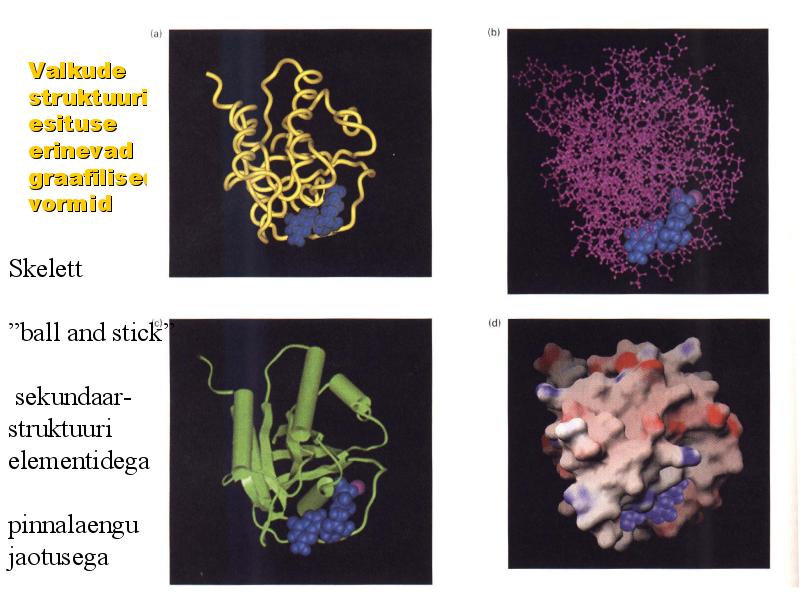
Valkude
struktuuri
esituse erinevad
graafilised
vormid
Skelett
"ball and stick"
sekundaar-
struktuuri
elementidega
pinnalaengu
jaotusega
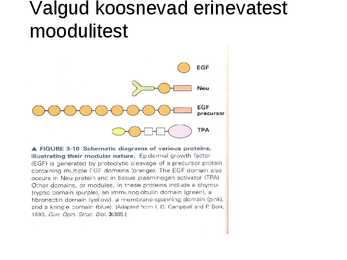
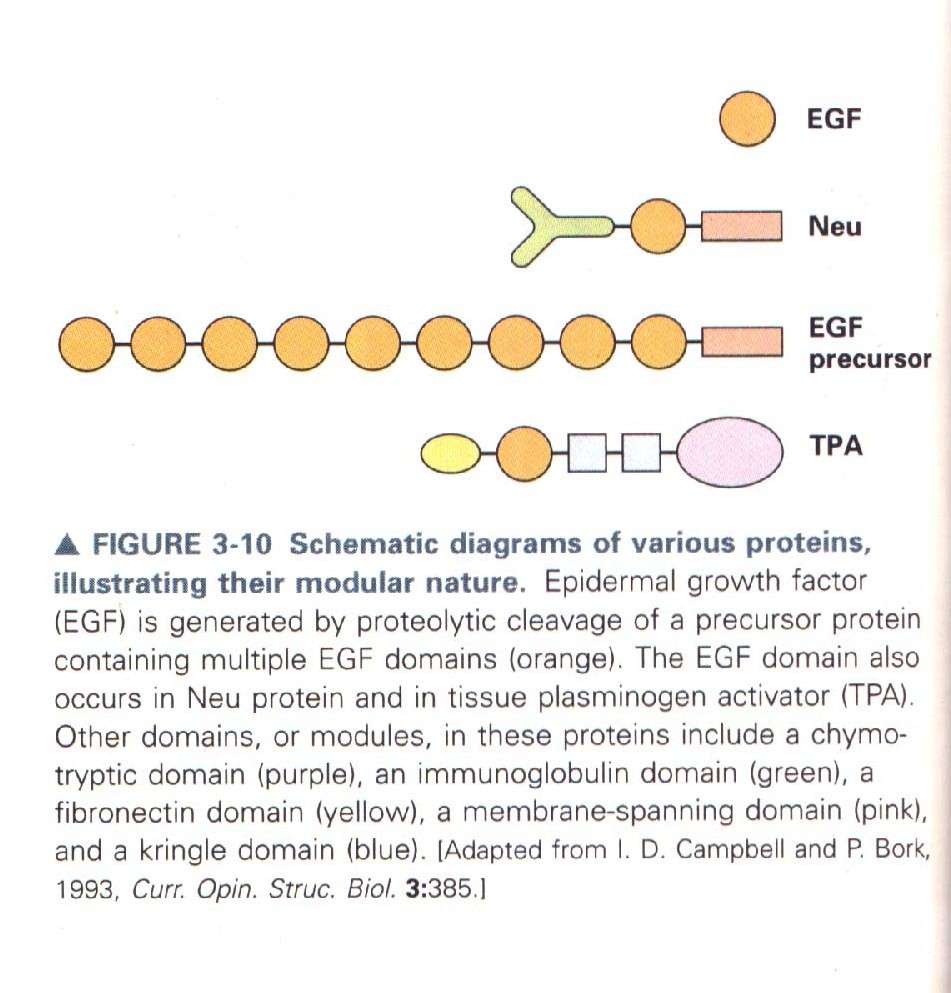
Valgud koosnevad erinevatest
moodulitest
Valkude homoloogia
Sarnane struktuur-sarnane järjestus (Max Peruz 1960)
Homoloogilised struktuurid- evolutsiooniliselt samast eellasest
pärinevad, ei pea täitma sarnaseid funktsioone (homoloogid vs.
analoogid)
Valgud jagatud perekondadesse vastavalt struktuurihomoloogiale
-liigisisene
-liikidevaheline
Järjestuste homoloogia võimaldab seondada erinevad valgud
evolutsiooniliselt
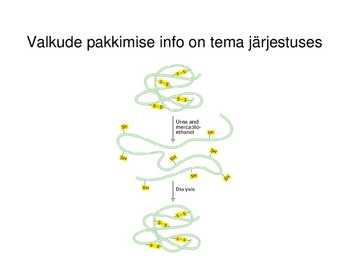
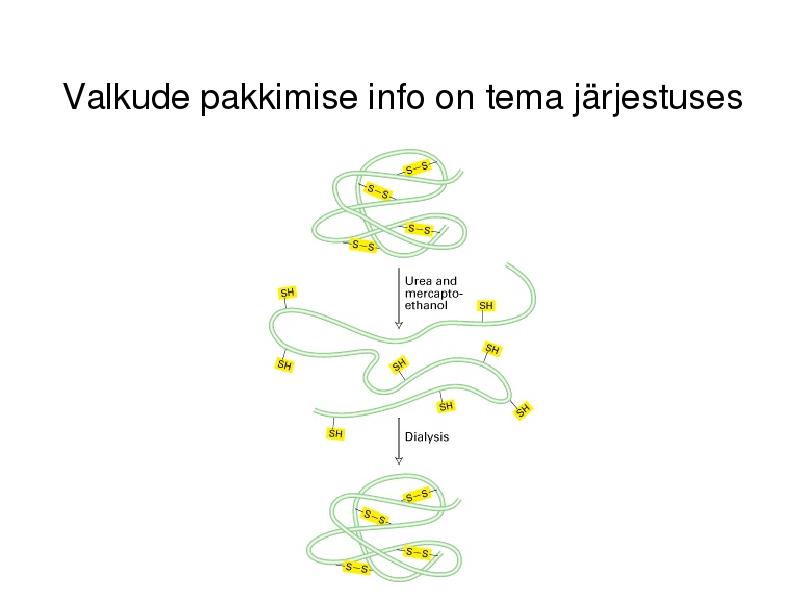
Valkude pakkimise info on tema järjestuses
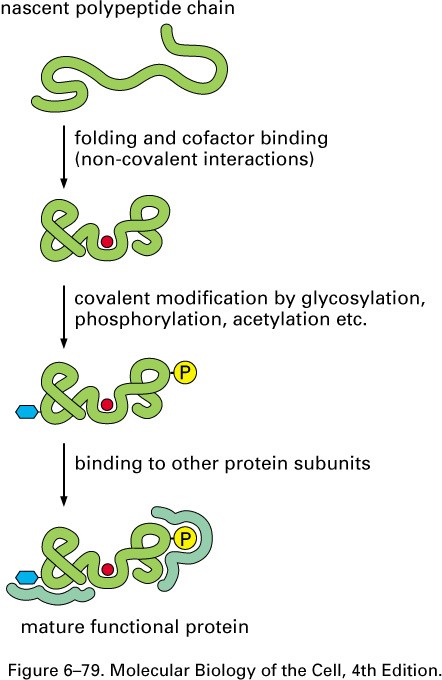
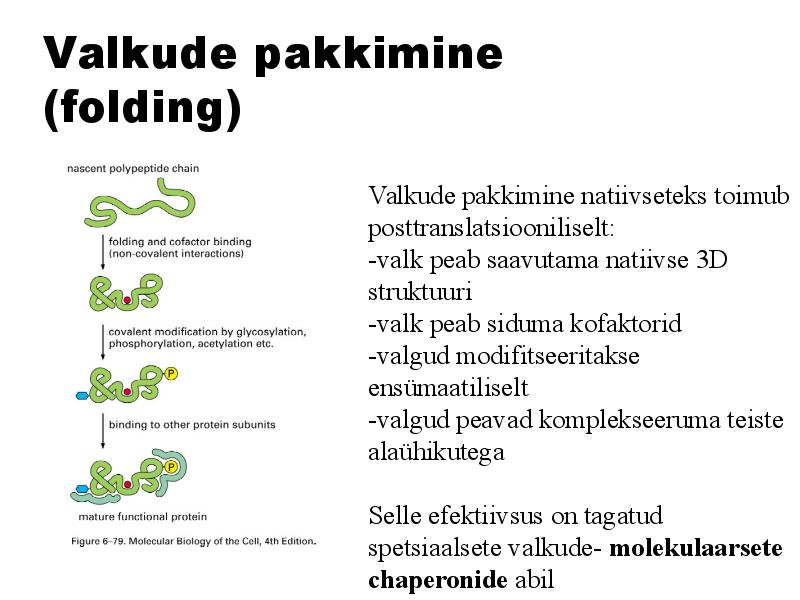
Valkude pakkimine (folding)
Valkude pakkimine natiivseteks
toimub
posttranslatsiooniliselt:
-valk peab saavutama natiivse 3D
struktuuri
-valk peab siduma kofaktorid
-valgud modifitseeritakse
ensümaatiliselt
-valgud peavad komplekseeruma teiste
alaühikutega
Selle efektiivsus on tagatud
spetsiaalsete valkude- molekulaarsete
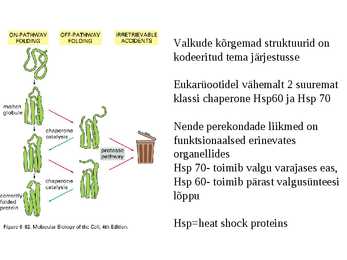
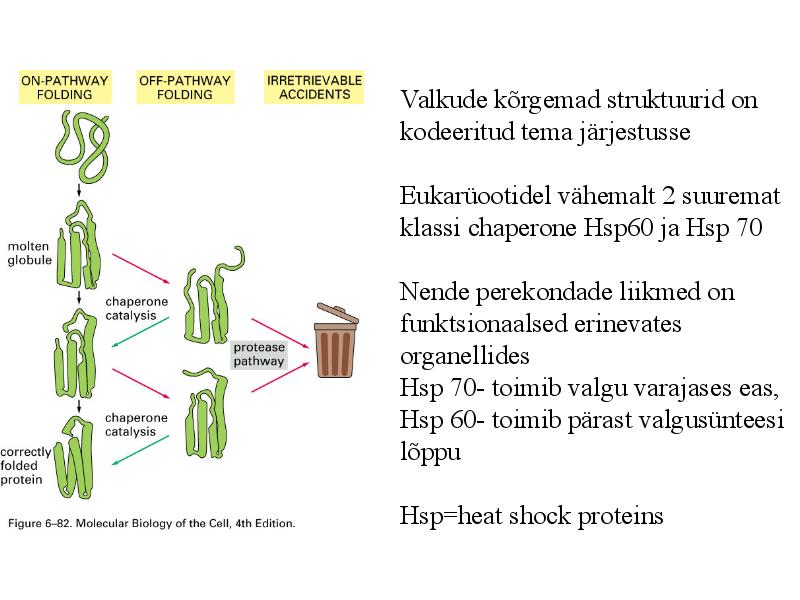
Valkude kõrgemad struktuurid on
kodeeritud tema järjestusse
Eukarüootidel vähemalt 2 suuremat
klassi chaperone Hsp60 ja Hsp 70
Nende perekondade liikmed on
funktsionaalsed erinevates
organellides
Hsp 70- toimib valgu varajases eas,
Hsp 60- toimib pärast valgusünteesi
lõppu
Hsp=heat shock proteins
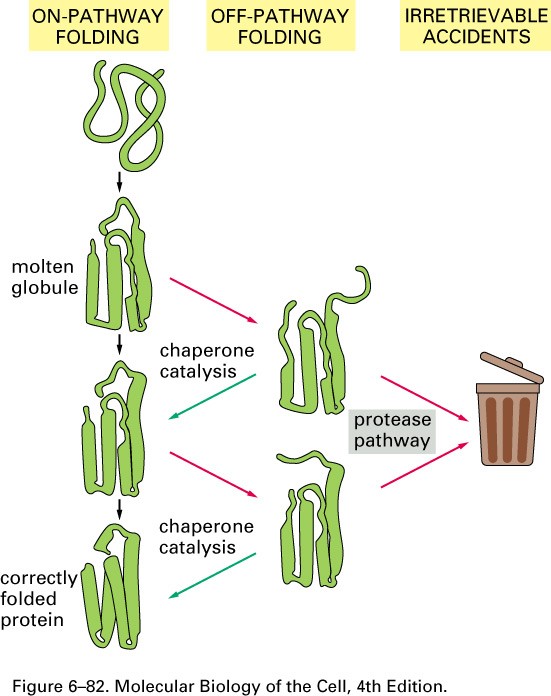
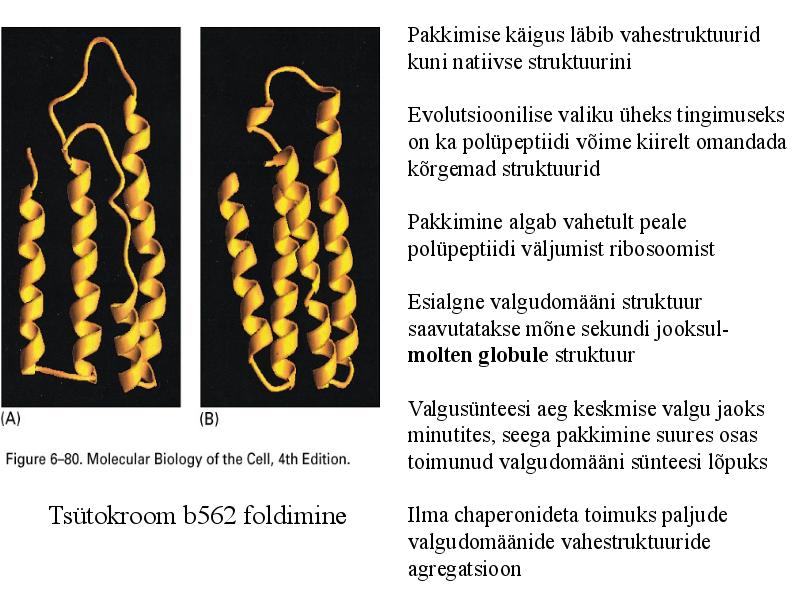
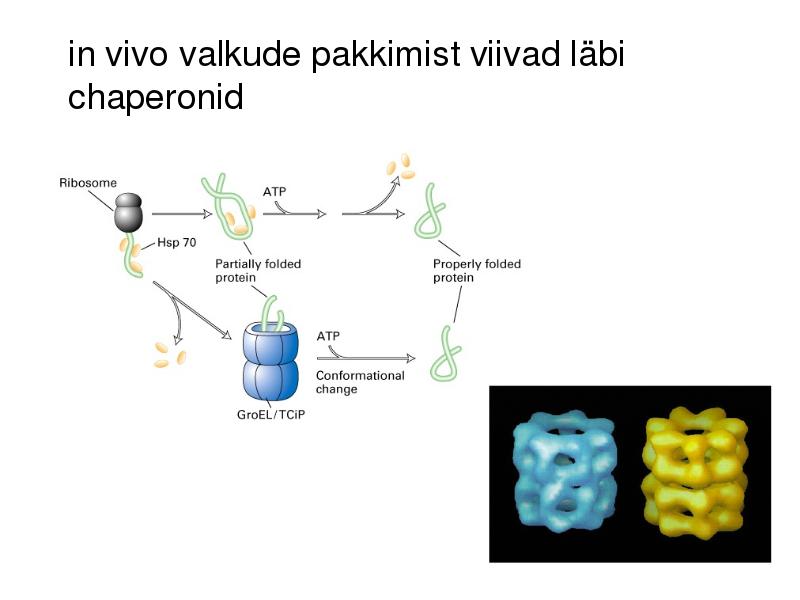
Pakkimise käigus läbib vahestruktuurid
kuni natiivse struktuurini
Evolutsioonilise valiku üheks tingimuseks
on ka polüpeptiidi võime kiirelt omandada
kõrgemad struktuurid
Pakkimine algab vahetult peale
polüpeptiidi väljumist ribosoomist
Esialgne valgudomääni struktuur
saavutatakse mõne sekundi jooksul-
molten globule struktuur
Valgusünteesi aeg keskmise valgu jaoks
minutites, seega pakkimine suures osas
toimunud valgudomääni sünteesi lõpuks
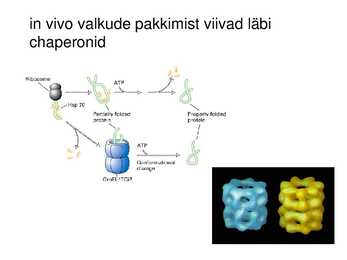
Tsütokroom b562 foldimine Ilma chaperonideta toimuks paljude
valgudomäänide vahestruktuuride
agregatsioon
in vivo valkude pakkimist viivad läbi
chaperonid
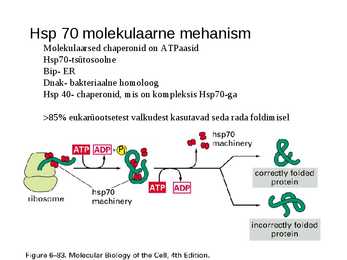
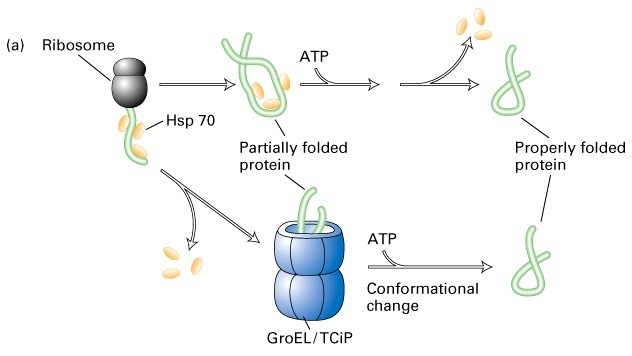
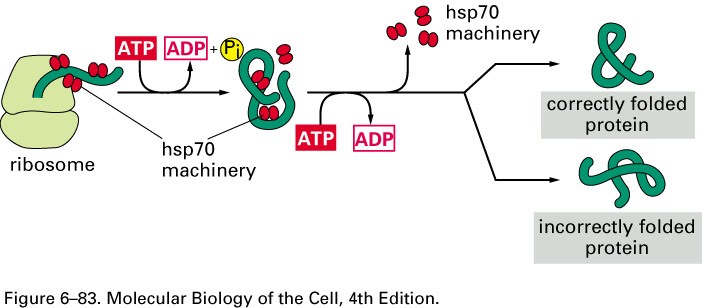
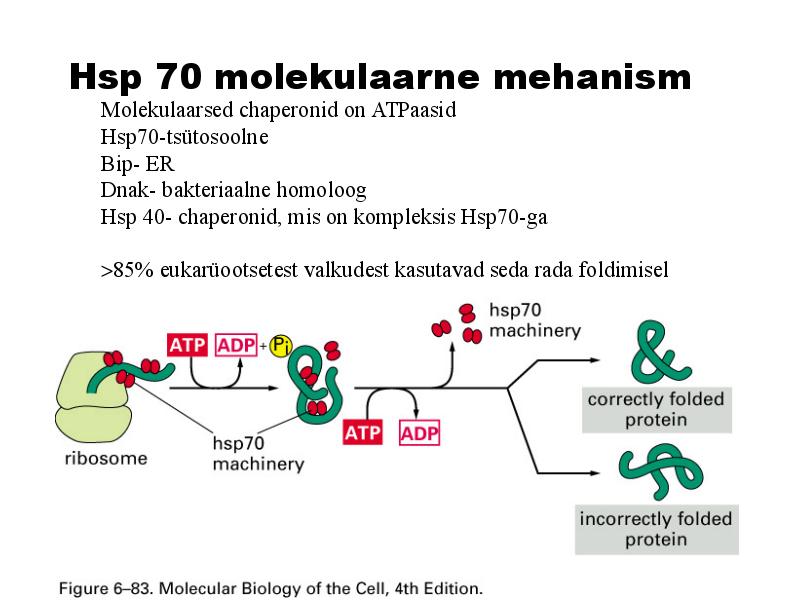
Hsp 70 molekulaarne mehanism
Molekulaarsed chaperonid on ATPaasid
Hsp70-tsütosoolne
Bip- ER
Dnak- bakteriaalne homoloog
Hsp 40- chaperonid, mis on kompleksis Hsp70-ga
>85% eukarüootsetest valkudest kasutavad seda rada foldimisel
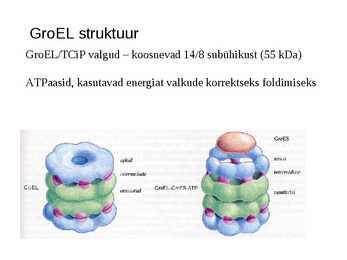
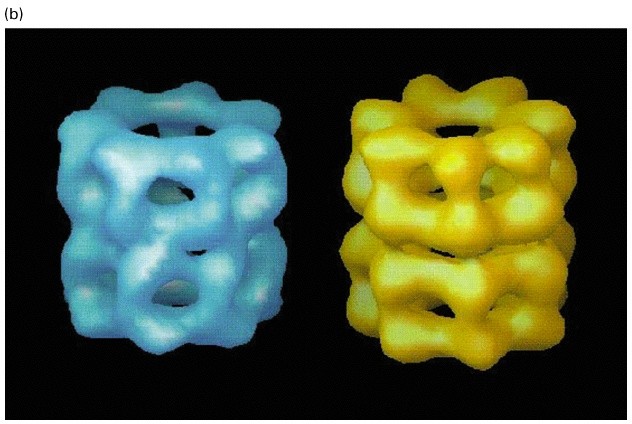
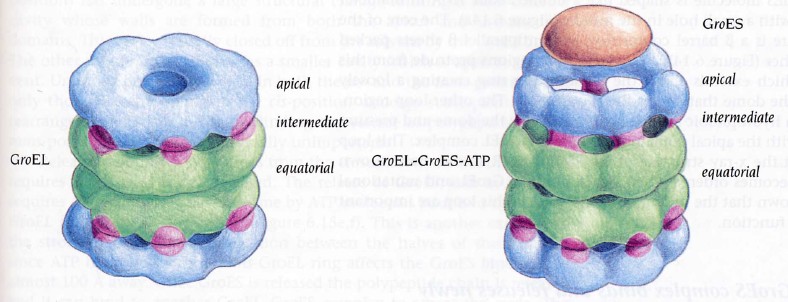
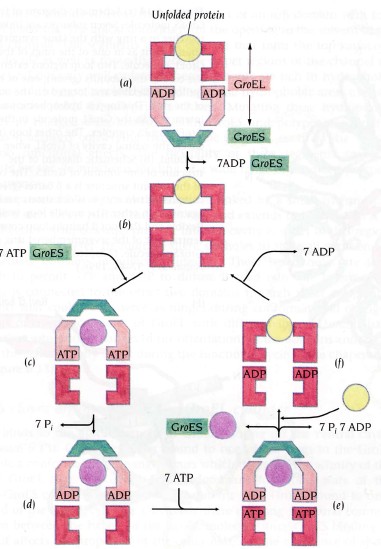
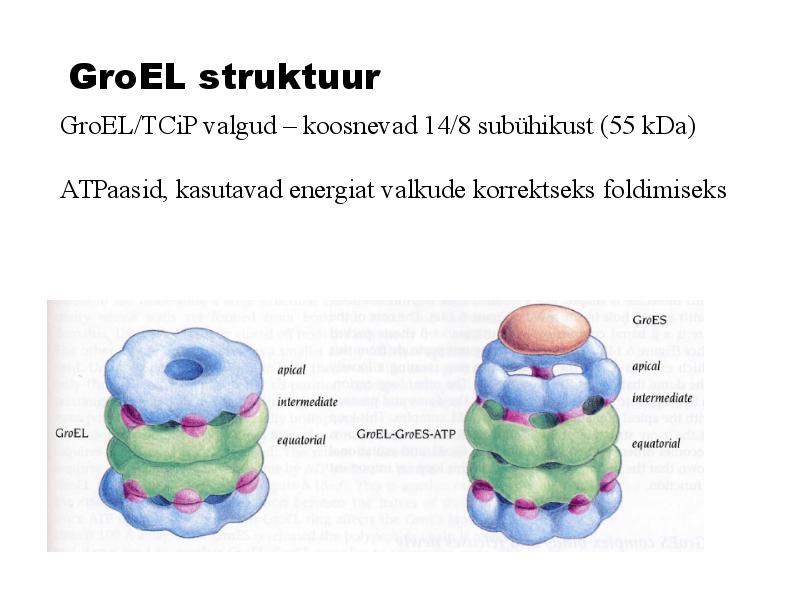
GroEL struktuur
GroEL/TCiP valgud koosnevad 14/8 subühikust (55 kDa)
ATPaasid, kasutavad energiat valkude korrektseks foldimiseks
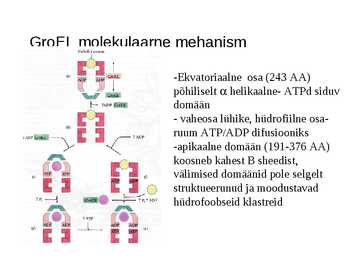
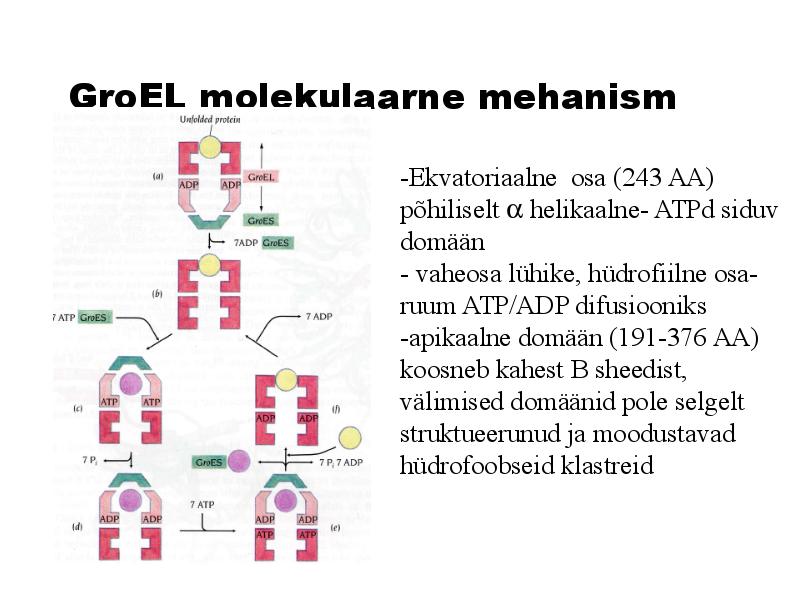
GroEL molekulaarne mehanism
-Ekvatoriaalne osa (243 AA)
põhiliselt helikaalne- ATPd siduv
domään
- vaheosa lühike, hüdrofiilne osa-
ruum ATP/ADP difusiooniks
-apikaalne domään (191-376 AA)
koosneb kahest B sheedist,
välimised domäänid pole selgelt
struktueerunud ja moodustavad
hüdrofoobseid klastreid
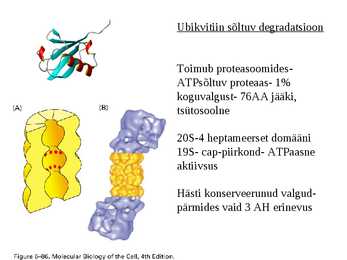
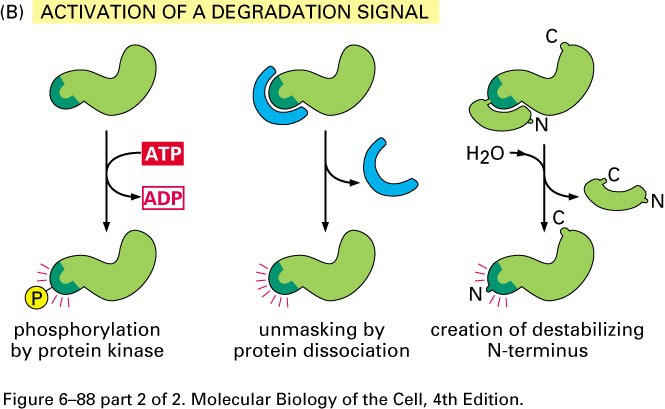
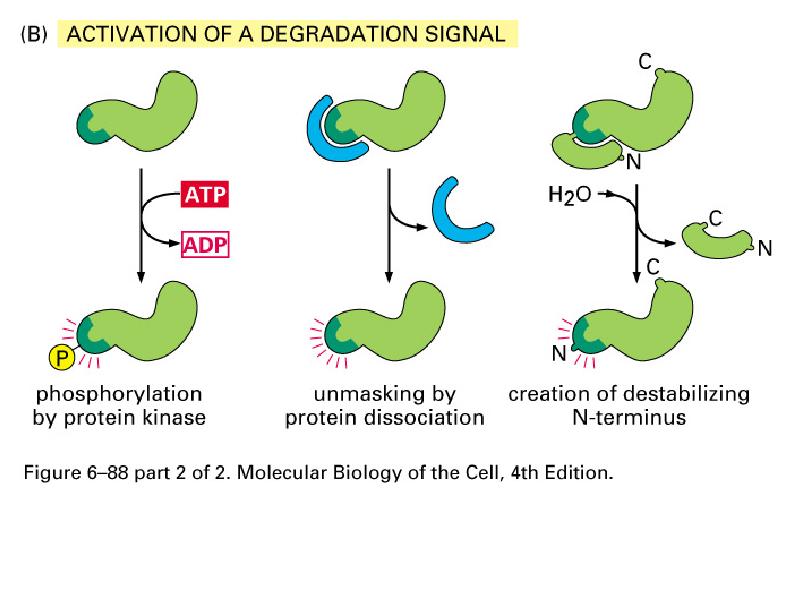
Valkude degradeerimine
Ekstratsellulaarselt toimub valkude lagundamine proteaaside
abil: endoproteaasid, eksoproteaasid
Valkude vananemine:
-keemiline vananemine Gln Asn deaminatsioon
-Met Cys oksüdeerimine
-Ratsemisatsioon
Valkude eluiga determineeritud tema järjestusega
Intratsellulaarne valkude degradatsioon:
-Lüsosomaalne
-Ubikvitiin-sõltuv
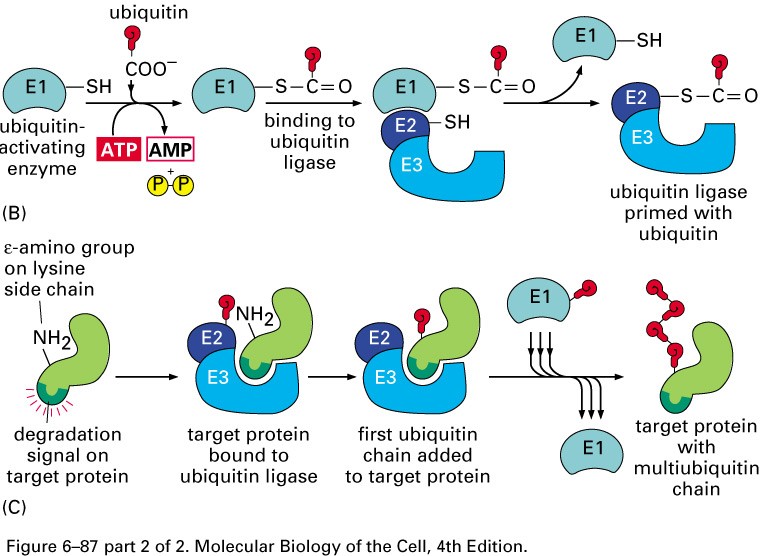
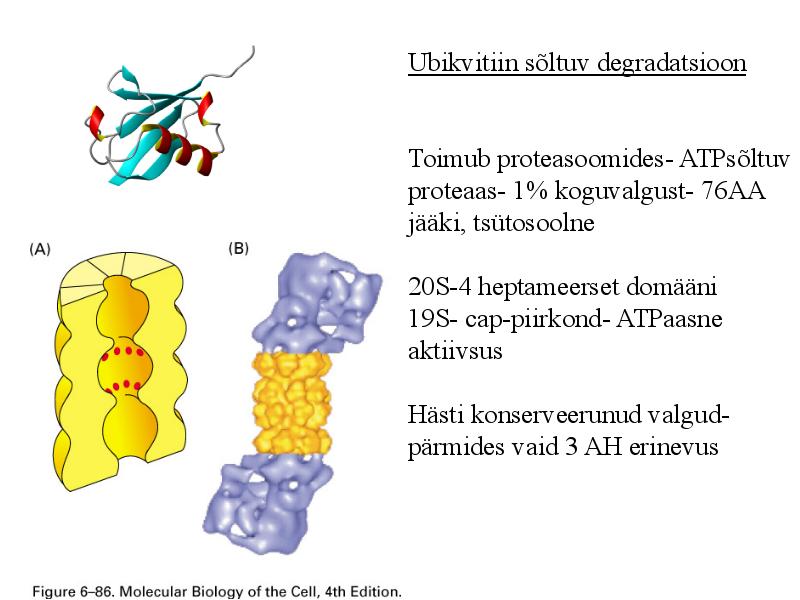
Ubikvitiin sõltuv degradatsioon
Toimub proteasoomides-
ATPsõltuv proteaas- 1%
koguvalgust- 76AA jääki,
tsütosoolne
20S-4 heptameerset domääni
19S- cap-piirkond- ATPaasne
aktiivsus
Hästi konserveerunud valgud-
pärmides vaid 3 AH erinevus
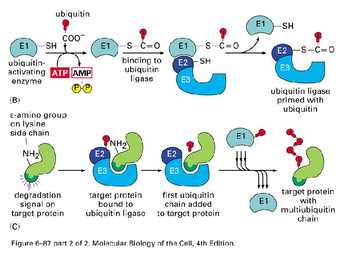
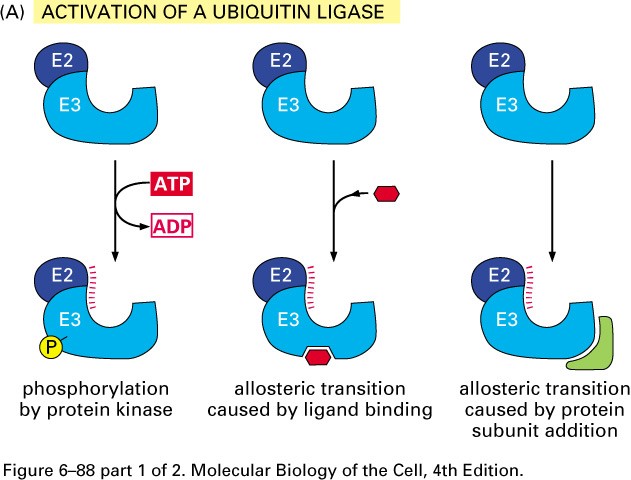
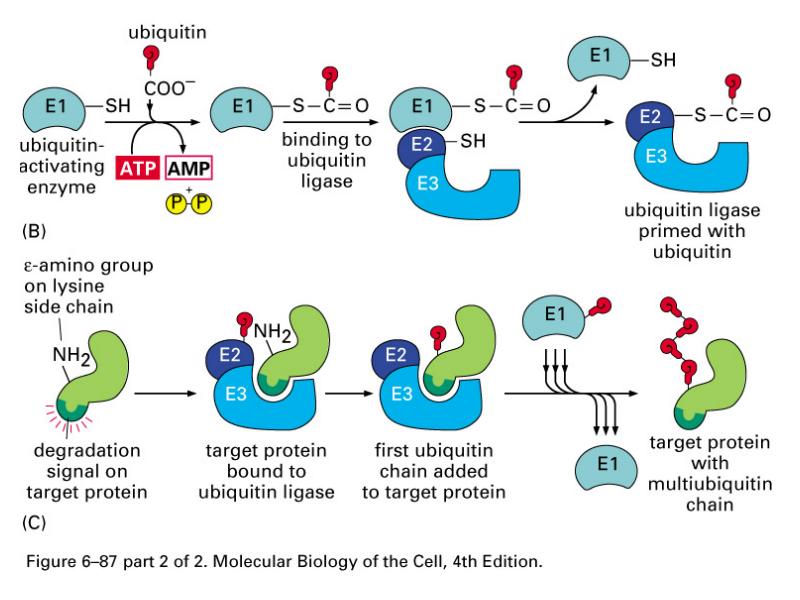
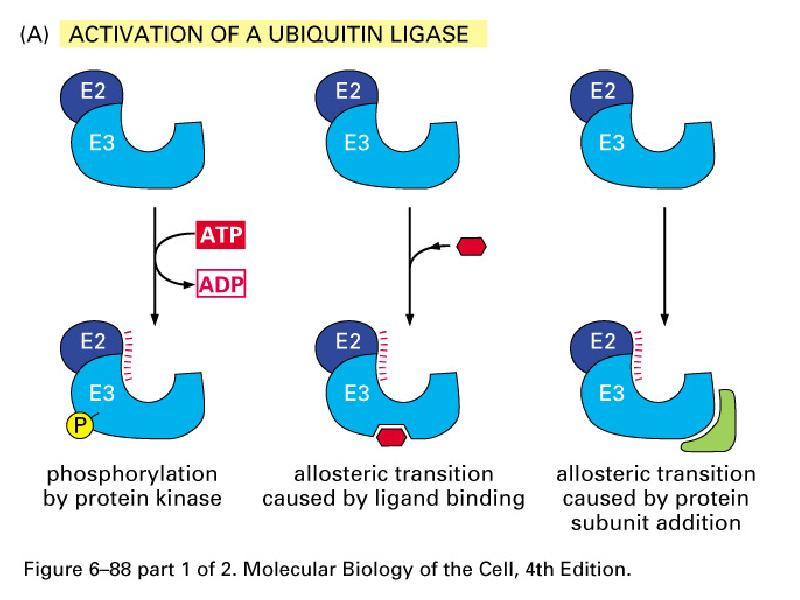
Ubikvitiin-konjugeerivad valgud
E1- ubikvitiini aktiveerivad valgud
E2- ubikvitiin konjugeerivad valgud
E3- aksessuaarne valk, moodustab E2-E3 kompleksi-ubikvitiini
ligaasid
E2-E3- kompleks seondub E3 abil märklaudvalgu Lys ja moodustab E2
abil polüubikvitiinahela- ligandiks proteasoomile
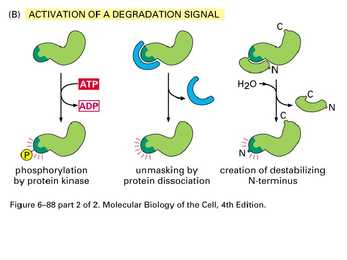
Valkude aktiivsus sõltub tema N terminuse struktuurist
-amidatsioon
-Met Ile Val Gly Pro Thr
-tsükliinidel konsensus: X-X-Leu-GlyX-Ile-Gly-Asn
-PEST rikkad alad
3 tüüpi E3 valke, mis tunnevad erinevaid N terminusi:
1.Aluselised AHd Cys, His
2.Hüdrof. Ahd Phe Tyr Trp Leu
Valkude topoloogia
Valkude bioloogilise aktiivsuse avaldumine toimub kindlates
kohtades, selle realiseerumiseks kasutatakse erinevaid
signaaljärjestusi:
2 tüüpi:
-võivad olla järjestikused
- moodustuda valgu eri osadest selle pakkimise tulemusena
20% juhuslikest järjestustest moodustavad erinevaid topoloogilisi
signaale
Valkude posttranslatoorne modifitseerimine
Keemiline modifitseerimine:
AtsetüleerimineNatsetüültransferaas
Nterminaalne metülatsioon
Nterminaalne müristoüleerimine
Lipiidide lisamine
Cterminaalne amidatsioon
Glükosüleerimine
Fosforüleerimine
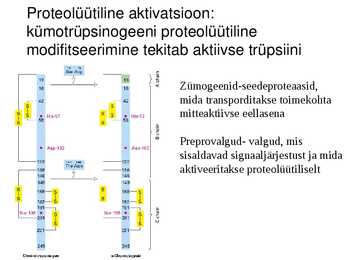
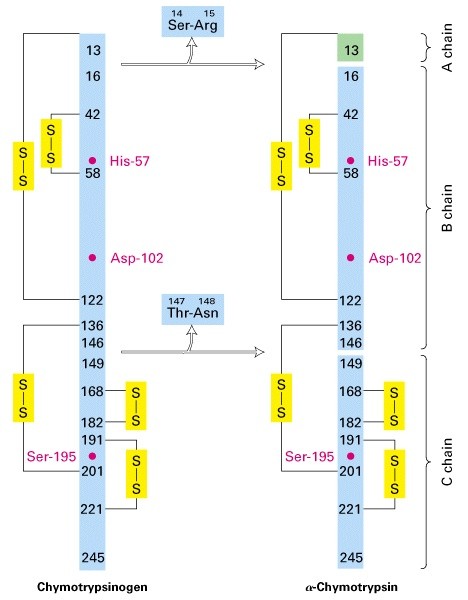
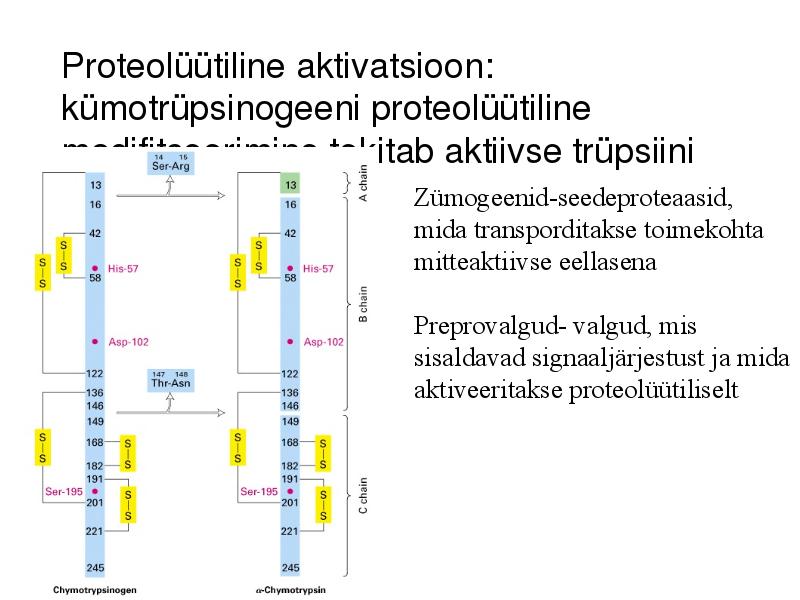
Proteolüütiline aktivatsioon:
kümotrüpsinogeeni proteolüütiline
modifitseerimine tekitab aktiivse trüpsiini
Zümogeenid-seedeproteaasid,
mida transporditakse toimekohta
mitteaktiivse eellasena
Preprovalgud- valgud, mis
sisaldavad signaaljärjestust ja mida
aktiveeritakse proteolüütiliselt
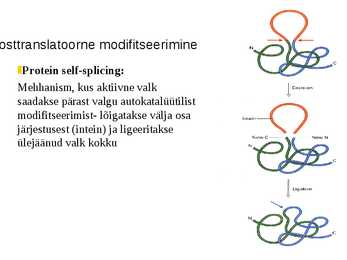
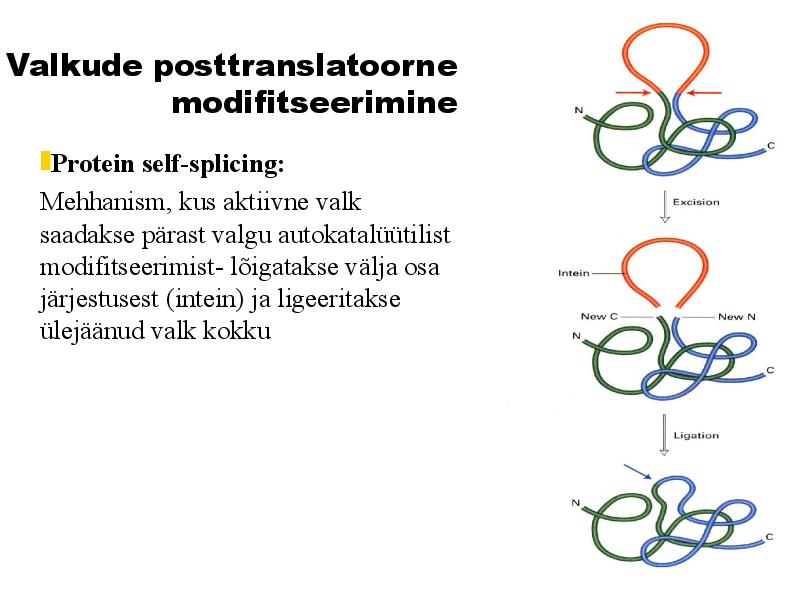
osttranslatoorne modifitseerimine
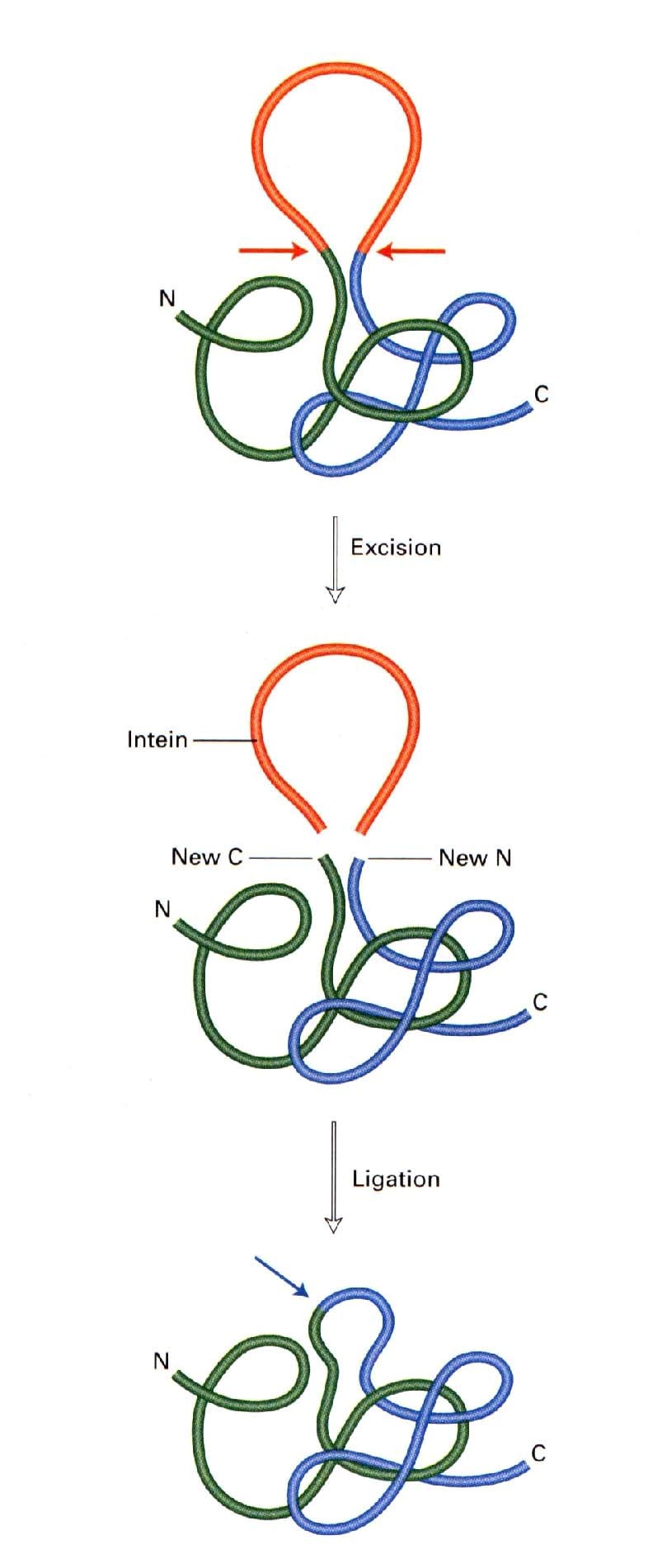
Protein self-splicing:
Mehhanism, kus aktiivne valk
saadakse pärast valgu autokatalüütilist
modifitseerimist- lõigatakse välja osa
järjestusest (intein) ja ligeeritakse
ülejäänud valk kokku
Valkude struktuurid ja funktsioon
Valkude põhiomadus siduda teisi valkemolekulaarne
komplementaarsus
Ligand mingi valguga spetsiifiliselt interakteeruv molekul
Polüpeptiidid
Mittevalgulised
Ligandi seondumine kutsub esile märklaudvalgu (retseptori või
ensüümi) konformatsiooni muutuse
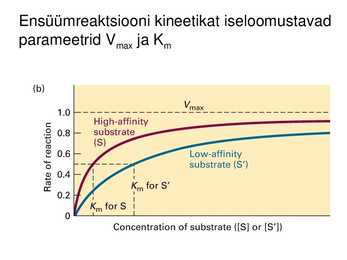
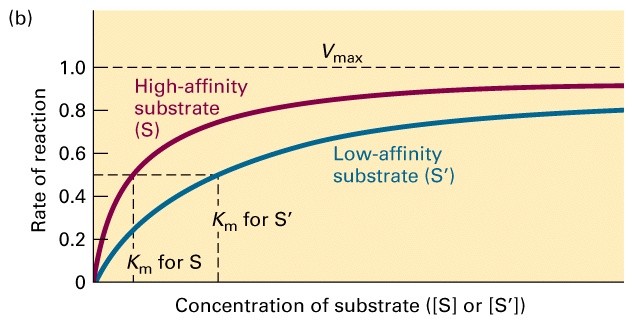
Ligandi (k.a ensüümi) spetsiifilisuse mõõt on afiinsus
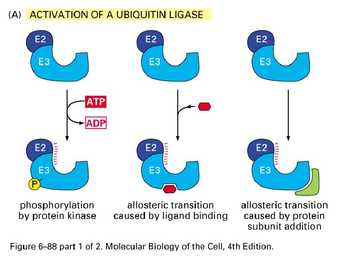
Mehhanismid, mis reguleerivad valkude
funktsionaalsust
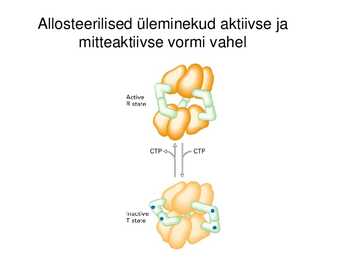
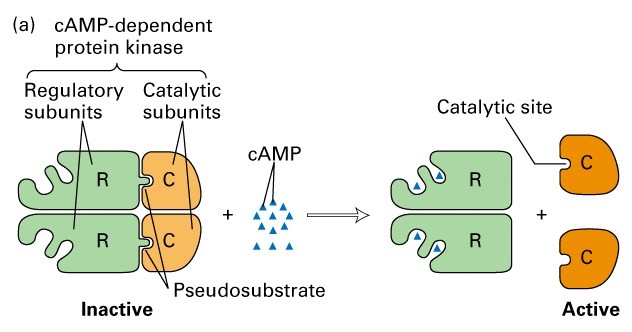
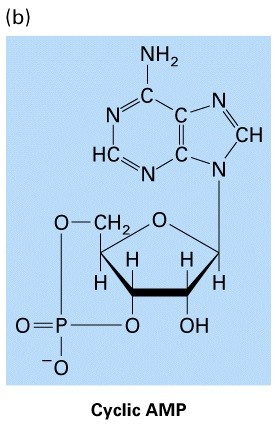
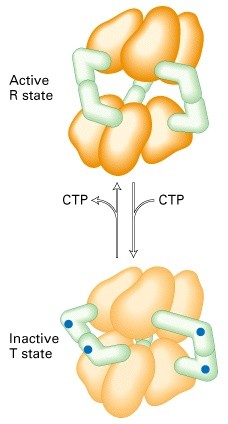
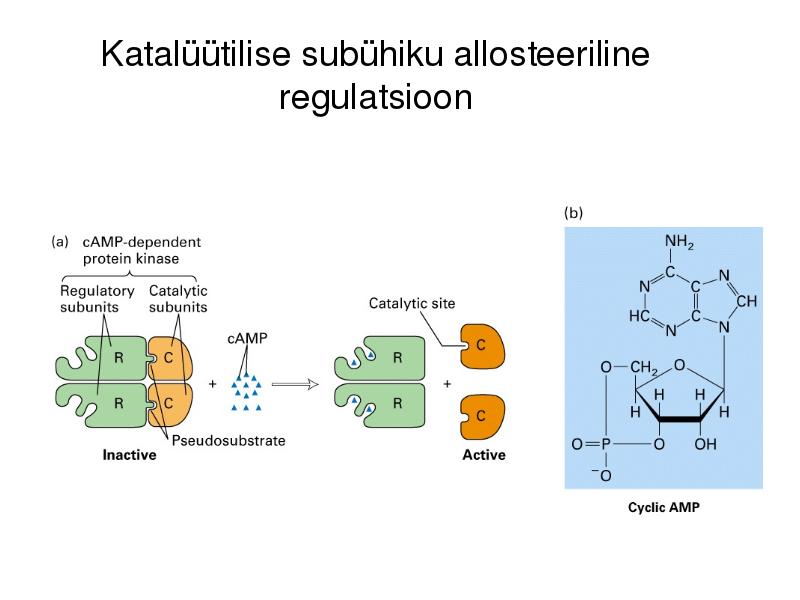
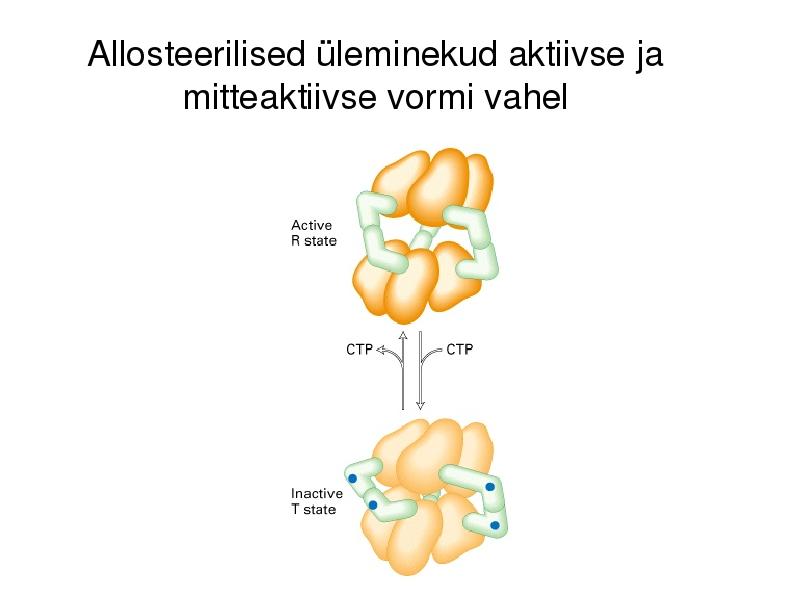
Allosteerilised üleminekud (allosteeriline kontroll)
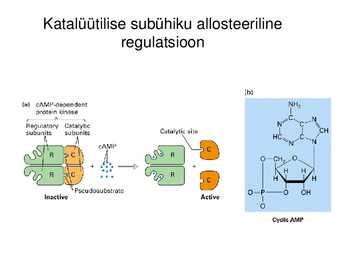
Katalüütiliste subühikute konformatsiooni muutus allosteerilise
regulaatori seondumise järgselt, üleminek aktiivse ja mitteaktiivse
vormi vahel (active and inactive state)
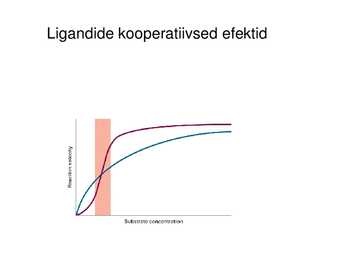
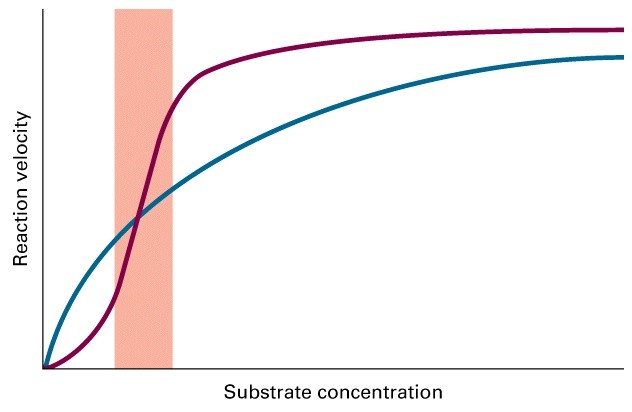
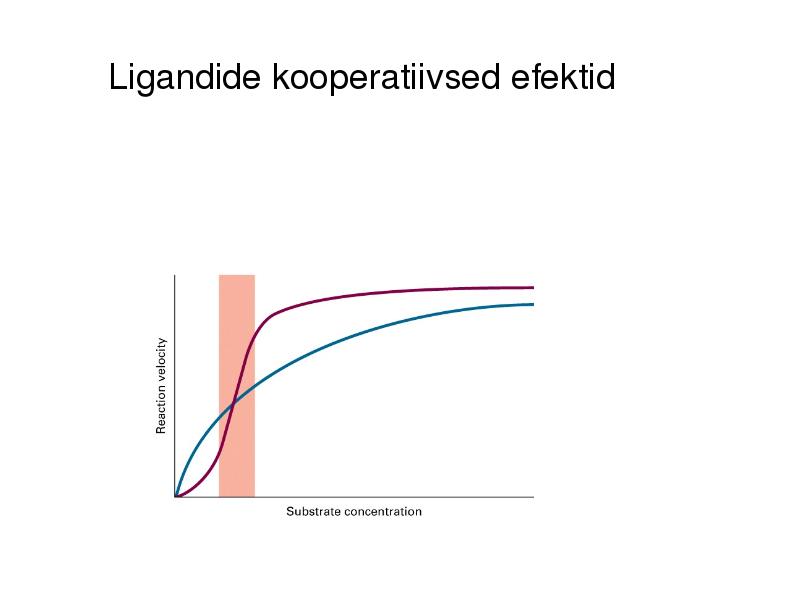
Ligandide kooperatiivsed efektid (ühe ligandi seondumine kas
inhibeerib või soodustab järgmise seondumist, Hilli koefitsioent)
Fosforüleerimine - defosforüleerimine
Proteolüütiline modifitseerimine (aktivatsioon või
innakstivatsioon, nt trüpsinogeen ja trüpsiin-
seedeensüümid)
Valkude kompartmentalisatsioon- ensüüm kas pääseb või ei
pääse substraadile ligi
Ligandide kooperatiivsed efektid
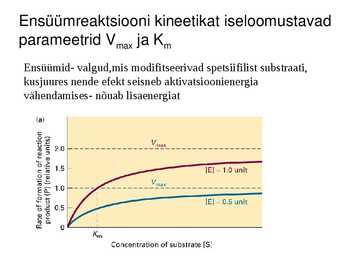
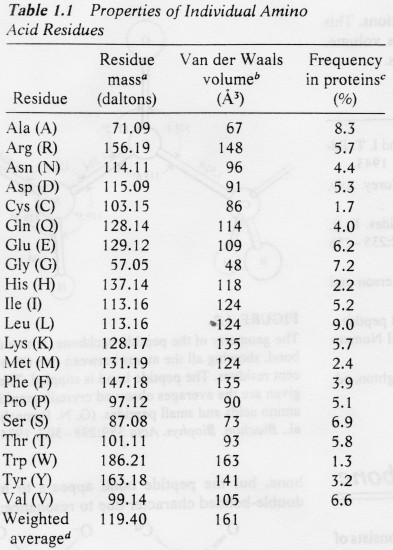
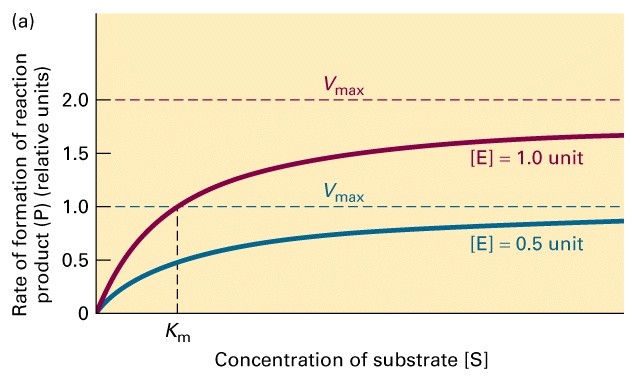
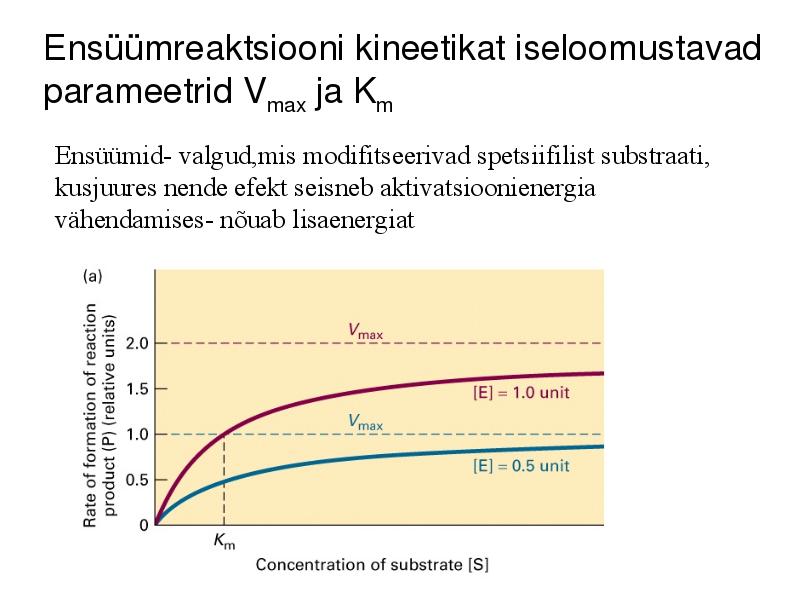
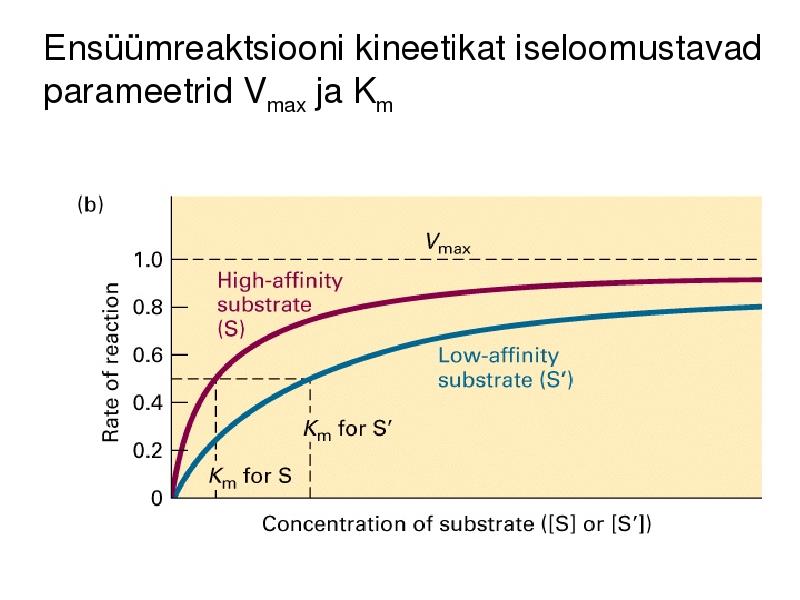
Ensüümreaktsiooni kineetikat iseloomustavad
parameetrid Vmax ja Km
Ensüümid- valgud,mis modifitseerivad spetsiifilist substraati,
kusjuures nende efekt seisneb aktivatsioonienergia
vähendamises- nõuab lisaenergiat
Ensüümreaktsiooni kineetikat iseloomustavad
parameetrid Vmax ja Km
Katalüütilise subühiku allosteeriline
regulatsioon
Allosteerilised üleminekud aktiivse ja
mitteaktiivse vormi vahel
Info valgustruktuuridest
Sarnased õppematerjalid
![Molekulaar - ja rakubioloogia I kontrolltöö kordamisküsimused]()
15
docx
Molekulaar - ja rakubioloogia I kontrolltöö kordamisküsimused
Osaliselt laetud: amiidrühma N on osaliselt
positiivne, karbonüülrühma O osaliselt negatiivne. Keemiliselt väheaktiivne –
stabllisus suur. Pro- tsükliline kõrvalahel välistab pöörde-cis peptiidsidemed- b-
pööre
Valkude järjestuses eristatakse N- ja C- terminust (vaba aminorühm ja
karboksüülrühm). Valk kasvab alati N-terminusest C-terminusse.
4. Kuidas on defineeritud peptiidid ja valgud?
Peptiid – aminohappejärjestus, millel puudub selgelt defineeritud
kolmemõõtmeline struktuur, enamasti lühikesed ahelad (<100 aminohappe)
Valk – polüpeptiidahel, millel on kindel kõrgemat järku struktuur (ja bioloogiline
funktsioon), sageli kvaternaarstruktuur
5 Valgu struktuuritasemed, interaktsioonid, mis stabiliseerivad
struktuure
Valkude funktsionaalsus on tagatud nende kolmemõõtmelise struktuuriga-
konformatsiooniga-, mis omakorda tuleneb valkude aminohappelisest
järjestusest. Aminohapete kõrvalahelate erinevad omadused tingivad nende
Molekulaar - ja rakubioloogia loengud
![Molekulaar- ja rakubioloogia KT I kordamisküsimused]()
8
docx
Molekulaar- ja rakubioloogia KT I kordamisküsimused
MOLEKULAAR- JA RAKUBIOLOOGIA |
YTM0011
I KONTROLLTÖÖ KORDAMISKÜSIMUSED |
20/09/10
I VALKUDE STRUKTUUR
1. Aminohapped, aluselised ja happelised, hüdrofiilsed ja hüdrofoobsed, polaarsed vs.
mittepolaarsed, kõrvalahelate tüübid.
Aluselised: Lüsiin, Arginiin, Histidiin. Happelised: Aspartaat, Glutamaat.
Hüdrofoobsed: Alaniin, Valiin, Leutsiin,
Metioniin, Isoleoutsiin, Fenüülalaniin, Trüptofaan,
Tyrosiin. Hüdrofiilsed: Arginiin, Lüsiin, Aspargiin,
Glutamaat, Proliin, Aspartaat.
Molekulaar - ja rakubioloogia loengud
![Valkude ruumiline struktuur]()
37
ppt
Valkude ruumiline struktuur
Valkude ruumiline struktuur
�1. Valkude struktuuri määravad
faktorid
2. Valkude sekundaarstruktuur
3. Valkude tertsiaarstruktuur
4. Valkude kvaternaarstruktuur
5. Valkude struktuuri näiteid:
RibonukleaasA, Müoglobiin,
Hemoglobiin, Insuliin
� Valkude struktuur?
· Bioloogiliste makromolekulide
struktuur kirjeldatakse erinevatel
tasanditel PRIMAARSTRUKTUUR. Aminohappe jääkide lineaarne järjestus
· primaarstruktuur
· sekundaarstruktuur
· tertsiaarstruktuur
· kvaternaarstruktuur
heeliks leht
SEKUNDAARSTRUKTUURID DOMEENID ehk SUPERSEKUNDAARSTR
Millised faktorid määravad valkude
![Biokeemia I test]()
48
pdf
Biokeemia I test
Mõõtmed 20nm ...
1µm; geenide arv 3...200; nukleotiidide arv 3 tuhat kuni 300 tuhat.
Kuna puudub iseseisev ainevahetus ja reprodutseerumisvõime, siis viirusosakesed pole
elusorganismid.
� LIISI KINK 6
BIOKEEMIA test I
II. VESI JA VESILAHUSED. TERMODÜNAAMIKA ALUSED.
1. Vesi omadused, struktuur, H-sidemed vees ja jääs.
Omadused
Kõrge sulamis- ja keemistemperatuur (0°C ja 100°C)
Suur aurumissoojus (540 kcal/kg = 2260 kJ/kg)
Suur soojusmahtuvus (1 kcal/kg·deg)
Kõrge pindpinevus
Kõrge dielektriline konstant
Maksimaalse tihedus vedelas olekus
Struktuur
Jäik struktuur teeb vee molekuli polaarseks. Võime moodustada neli H-sidet molekuli kohta.
Hapnikuaatomi kaks paardumata elektroni moodustavad kaks negatiivse laengu piirkonda.
![Molekulaarbioloogia konspekt]()
38
pdf
Molekulaarbioloogia konspekt
RNA polümeraase on väga
palju erinevaid tüüpe. Eukarüootides on kolm erinevat RNA polümeraasi, mis
sünteesivad erinevaid RNA molekule. RNA sünteesil on substraatideks (ribo-)
nukleosiid 5'-trifosfaatidest. Sünteesitud RNA ahel vastab üks-üheselt temaga
antiparalleelsele DNA matriitsahelale komplementaarsusprintsiibi alusel. RNA
järjestusega identset DNA ahelat nimetatakse kodeerivaks ahelaks. RNA sünteesi
käigus toimub DNA ahelate lahtiharutamine. Algne DNA struktuur taastub peale
transkriptsiooni lõppu. Transkriptsiooniga on seotud RNA protsessing ja
modifitseerimine.
3. Translatsioon - valgu biosüntees. Translatsioon tähendab tõlkimist.
Molekulaarbioloogias tähendab translatsioon RNA (seega ka DNA) nukleotiidse
järjestuse tõlkimist valkude aminohappeliseks järjestuseks. Valkude sünteesiks
vajalikku geneetilist informatsiooni kannab mRNA (matriits- ehk informatsiooniline-
RNA)
![Biokeemia mõisteid ja termineid]()
7
doc
Biokeemia mõisteid ja termineid
Peptiidahela Φ ja Ψ nurgad – võimalikud nurgad, mille all võivad planaarse
peptiidsidemega (-N(H)-C(=O)-) külgnevad aminohappejäägid paikneda (planaarse
peptiidsideme suhtes).
Peptiidside – side, mis ühendab valkude primaastruktuuris esinevaid aminohappejääke.
Peptiidside on planaarne, sest seal esineb resonantsstruktuur.
N ja C terminus – peptiidahela (valkude primaarstruktuuri) otsas; N-ots tähistab vaba
aminorühmaga otsa ning C-ots vaba karboksüülrühmaga otsa. Valgu süntees toimub N-
terminusest C-terminuse poole, st. uusi aminohappeid liidetakse eelmise aminohappe
karboksüülrühma külge. Üldiselt aminohappe aminorühmapoolne ots ja
karboksüülrühmapoolne ots.
Polüpeptiid ja oligopeptiid – polüpeptiid tähistab (pikka) polümeeri, mille monomeerideks
on aminohapped; oligopeptiid on lühike aminohapete polümeer (5-7? aminohapet).
Natiivne ja denatureeritud struktuur – natiivne struktuur on valgu selline konformatsioon,
![MOLEKULAARBIOLOOGIA-ja RAKUBIOLOOGIA]()
54
pdf
MOLEKULAARBIOLOOGIA ja RAKUBIOLOOGIA
TBP Nterminaalne domään on erinevatelt organismidelt pärit
TBP valkudel nii pikkuselt kui järjestuselt väga varieeruv. Sellel on oluline roll RNAPolII-katalüüsitud
snRNAde geenide transkriptsioonis. TBP monomeer pakkub moodustades sadul-struktuuri, kus molekuli 2 poolt
omavad diaadset (polaarset, kaheli) sümmeetriat, ent pole identsed. Nagu HMGI ja teised DNAd-painutavad
valgud, interakteerub TBP DNA väikese vaoga, painutades tugevasti kaksikheeliksit. DNAd-siduv valgu pind
on TBP puhul konserveerunud kõigis eukarüootides, mis selgitab ka TATAbox promootorelemendi kõrget
konserveerumist läbi evolutsiooni.
11. Kas transkriptsioonifaktorid (aktivaatorid) interakteeruvad otse(vahetult) TBPga?
Jah, Kui TBP istub TATAboxil, siis saab TFIIB seonduda. TFIIB on monomeerne valk, mis on natuke väiksem
TBPst. TFIIB C-terminaalne ots interakteerub kahel pool TATAboxi TBP ja DNAga, samas kui tema N-
Molekulaar - ja rakubioloogia loengud
![Molekulaarbioloogia]()
194
docx
Molekulaarbioloogia
funktsioneerimise. Uurib füüsikalis-keemiliste struktuuride ja biokeemilis-füsioloogiliste
funktsioonide vastavust. Teadussuund hakkas arenema pärast makromolekulide
ruumilise struktuuri kindlakstegemist (DNA 3-ruumiline struktuur).
Molekulaarbioloogia dimensioon – 1 A – 300 A (üle 500 – rakubioloogia, alla 1 -
biofüüsika)
1 A (ongström) = 10 -10 m
1nm = 10 A
2-ahelalise DNA läbimõõt – 20 A
kovalentne side – 1,5 A
globulaarse valgu d – 50 A
dsDNA (double stranded) d – 50 A
ribosoomide, valgumolekulide d – 200-300 A
DNA aluspaaride vahe – 3,4 A
vesiniksideme pikkus – 3 A
nukleosoom – 60x110x110 A
bakteri ribosoom – 200x200x230 A
tuumapoorid – 120x120x75 A
bakteriaalne RNA polümeraas – 90x90x60 A
Molekulaarbioloogia põhidogma
DNA↔ RNA →valk
DNA sünteesitakse nii DNA kui RNA alusel!
RNA-sõltuv DNA polümeraas – pöördtranskriptaas – revertaas – katalüüsib DNA
Meedia
Kommentaarid (2)




















































Kõik kommentaarid