GENEETIKA AJALUGU
19. sajandil tegutses
Brno kloostris
munk Gregor Mendel, kes viis läbi katseid aedhernega. 1865. aastal
sõnastas ta pärilikkuse üldprintsiibid. Sellega sai alguse ka teadusliku geneetika periood.
Mendeli I seadus ehk esimese hübriidse põlvkonna ühtlikkuse seadus: homosügootsete vanemate
ristamisel saadakse esimeses järglaspõlvkonnas genotüübiliselt identsed ja fenotüübiliselt ühtlikud
järglased.
Mendeli II seadus ehk alleelide lahknemise seadus: monohübriidse ristamise teises
hübriidpõlvkonnas saadakse genotüübiline lahknemissuhe 1:2:1 ja fenotüübiline lahknemissuhe 3:1 või
1:2:1.
Mendeli III seadus ehk sõltumatu lahknemise seadus: erinevad alleelipaarid segregeeruvad ja
kombineruvad üksteisest sõltumatult. Polühübriid moodustab võrdse sagedusega 2n haplotüübiga
gameeti, kus n on heterosügootsete geenipaarde arv. Ganeetide ühinemisel võib tekkida 3n erineva
genotüübiga sügooti kindlate sagedustega.
1900. aastal Mendeli seaduste taasavastamine kolme sõltumatu teadlase poolt- Hugo de Vries, Carl
Correns ja
Eric von Tscermak-Seysenegg.
1910 -1925- pärilikkuse kromosoomiteooria, T.H.
Morgan .
1941. aastal avastati, et geen kodeerib valku
1944. aastal tõestas O. Avery, et pärilikkust
kandev materjal on desoksüribonukleiinhape.
1953. aastal kirjeldasid J. D. Watson ja F. H. C. Crick DNA kaksikheeliksi struktuuri.
1961. aastal avastati, et geneetiline kood on kirjutatud kolmest nukleotiidist
koosnevate triplettidena
1961. aastal avastati mRNA ning F. Jacob ning J. Monod avaldasid geeni regulatsiooni operonimudeli
1970. aastal
avastasid H. Temin ja D.
Baltimore retroviirustest ensüümi pöördtranskriptaas, mille abil
toimub DNA süntees RNA-lt.
1977. aastal töötati välja meetodid, millega oli võimalik DNA-d suurtes hulkades lugeda. PÄRANDUMISE ALUSED DNA (desoksüribonukleiinhape)
Ehitus. DNA on
madalmolekulaarne aine, mis koosneb viiesüsinikulisest
suhkrust desoksüriboos,
fosfaatrühmast ja lämmastikalusest. Fosfaatgrupp ja lämmastikgrupp moodustavad lämmastikaluse.
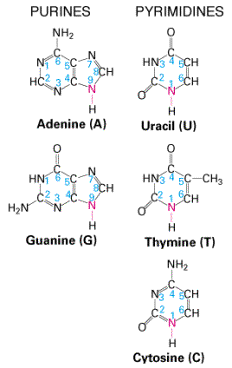
Lämmastikaluseid nimetatakse vastavalt lämmastikgrupile järgnevalt-
adenosiinfosfaat , tsütidiinfosfaat,
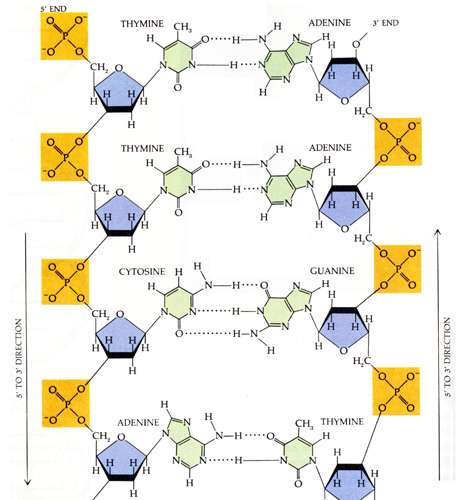
guanosiinfosfaat ja tümidiinfosfaat. DNA primaarseks struktuuriks on aluspaariline järjestus.
Sekundaarseks struktuuriks on aga
biheeliks ehk kaksikahel. DNA primaarne struktuur moodustub
lämmastikaluste üksteisele vastavuse ehk komplementaarsuse alusel. Lämmastikalusteks DNA puhul
on
adeniin , tsütosiin, tümiin ja
guaniin . Üksteisele vatavus on järgnev- adeniin moodustab kahe
vesiniksideme abil sideme tümiiniga ja guaniin kolme vesiniksideme abil tsütosiiniga. DNA on
pakitud histoonide abil. Tähtis on teada ka, et DNA
ahelad on antiparalleelsed, ehk siis
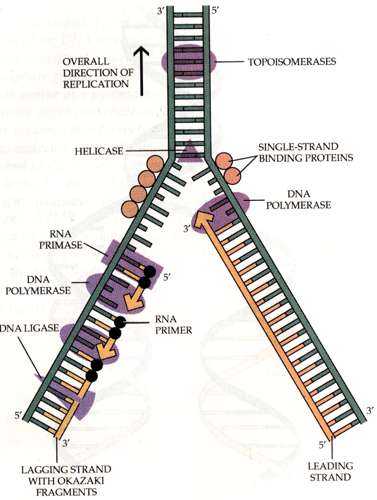
kokkuleppeliselt on üks ahel 5´-3´ ja teine ahel 3´-5´. DNA kahekordistumisprotsessi nimetatakse replikatsiooniks. Protsessi viib läbi ensüüm DNA
polümeraas. DNA kahekordistumine toimub semikonservatiivse mudeli järgi. See tähendab, et
mõlemale vanale
ahelale sünteesitakse kõrvale uus.
Replikatsioon toimub 5´-3´ suunas. Protsessis on
üks ahelatest juhtiv (
leading strand ) ja teine mahajääv ahel (lagging strand). Juhtivalt ahelalt toimub
süntees pidevalt, kuid mahajääval
ahelal toimub süntees 100-1000 nukleotiidiste blokkidena ehk
Okazaki fragmentidena. Nende fragmentide sünteesiks kasutatakse RNA praimereid. Hiljem
liidetakse tekkinud
fragmendid esüümi ligaas abil. Kuna DNA repikatsiooni puhul tekib mõlema DNA
ahelale kõrvale uus DNA ahel, siis nimetatakse vastavat protsessi semikonservatiivseks.
Helikaas - ensüüm, mis lahutab DNA ahelad üksteisest. Enamasti funktsioneeri heksameerina.
SSBP (
single -strand binding
protein )-
seondub üheahelalisele DNA-le,
hoides ära selle kaheahelaliseks
muutumist.
DNA replikatsiooni näol on tegemist väga täpse mehhanismiga. Nimelt, uurimuste kohaselt teeb
tehakse miljon aluspaari
liitmise kohta ainult üks viga, kuigi reaalselt võiks see arv olla palju suurem.
Nimelt on DNA polümeraasil peale sünteesimisaktiivsuse ka ingliskeelse terminiga väljendatud
aktiivsus ehk proofreading. Vigade parandamiseks on DNA polümeraasil 3´-5´ eksonukleaasne
aktiivsus.
Bakterirakus on kolm DNA polümeraasi- DNA polümeraas I, II ja III. Polümeraasid I ja II. DNA polümeraas III on põhiline DNA replikatsooni ensüüm. Imetajarakkudes on vähemalt 5 erinevat
polümeraasi- , , , ja , ja töötavad koos ning on seotud tuuma DNA replikatsiooniga. polümeraasi ülesandeks on
mitokondriaalse DNA replikatsioon. ja osalevad aga tuuma DNA
reparatsioonil.
http://oak.cats.ohiou.edu/~ballardh/pbio475/Heredity/DNA-replication.JPGFunktsioon. DNA peamiseks ülesandekson geneetilise info säilitamine ja selle
korrektne ülekandmine
tütarrakkudesse. Selle juures on vajalik DNA kahekordistumine ehk replikatsioon. RNA (
ribonukleiinhape )
Ehitus. RNA ehituslikeks komponentideks on viiesüsinikuline suhkur riboos, fosfaatgrupp ja
lämmastikgrupp.RNA primarseks struktuuriks on lämmastikaluste järjestus, kuid sekundaarseks
struktuuriks on osaline
heeliks .
Lämmastikalus tümiini asendab
uratsiil . Ehk siis komplementaarsus on A-U ja G-C.
RNA sünteesimine toimub
kolmes järgus:
initsiatsioon , elongatsioon ja terminatsioon. RNA
polümeraasid, mis katalüüsivad transkriptsiooni on komplekssed multimeersed
valgud .
Erinevad RNA liigid:
tRNA - transpordib aminohappeid ribosoomi
mRNA- info valgu sünteesiks
rRNA- ribosoomide
koostisosaFunktsioon: RNA abil toimub geneetilise info
realiseerimine .
Eukarüootide RNA polümeraasid
RNA polymerase I
tuumake ribosomaalne RNA, välja arvatud 5S rRNA
RNA polymerase II tuumake tuuma pre-mRNA-d
RNA polymerase III tuumake tRNA-d, 5 S rRNA ja teised väikesed tuuma RNA-d
Transkriptsiooniks on vajalikud promooterid. Erinevate polümeraasside puhul on ka vastavad
järjestused erinevad. RNA polümeraas II promootor sisaldab lühikesi konserveerunud järjestusi.
Transkriptsiooni alguspunktile lähim konserveerunud element kannab nimetust TATA box, järjestus
TATAAAA. Teised konserveerunud elemendid on CAAT box ja GC box.
Transkriptsioonifaktorid on valgud, mis inistsieerivad või reguleerivad eukarüootsetes rakkudes
trankriptsiooni. Transkriptsioonifaktorid sisaldavad DNA-ga seonduvat järjestust, mis seondub 8-15
nukleotiidi pikkuste
spetsiifiliste DNA aladega ning transaktiveerivat järjestust, mis enamasti
võimaldab sondumist RNA polümeraasi siduvate valkudega. Lisaks nendele aladele on
transkriptsioonifaktoritel mitmeid ühiseid ehituslikke elmente ja selle alusel jagatakse :need järgnevalt:
· heeliks-pööre-heeliks (
helix -turn-helix) motiiviga valgud · homeodomääni valgud
· heeliks-
ling -heeliks (helix-
loop -helix)
· "Tsinksõrme" (Zn-
finger ) sisaldavad valgud
· aluselised leutsiini tõmlukku (leuzine zipper) sisaldavad valgud
· POU-domääni valgud
· steroidhormoonide
retseptoridInimese
mitokondriaalne genoom Mitokondriaalne genoom koosneb tsirkulaarsest kaksikahelalisest DNA-st. Antud genoomi suuruseks
on 16 569 bp. Jaotatakse raskeks ja kergeks ahelaks.
Mitokondrid sisaldavad tavaliselt tuhandeid DNA
molekule. Mitokondriaalne DNA päritakse ema liini pidi. Inimese mitokondri DNA-s sisaldub
praeguste andmete kohaselt 37 geeni.
Tuuma genoom Mitokondriaalne genoom
· suurus 3300 Mb 16.6 kb
· erinevate DNA 23 XX või 24 XY rakkudes, üks tsirkulaarne DNA molekulide arv kõik on
lineaarsed molekul· seonduvad valgud mitmed
histoonid ja teised valgud suures osa valkudest vabad
· geenide arv 30 000-40 000 37
· geeni tihedus 1/90 kb 1/0.45 kb
· korduselemendid suur hulk väga vähe
· transkriptsioon enamik geenidest transkribeeritakse mitmete geenide transk- individuaalselt unikaalsetest promootoritelt riptsioon samalt promootorilt ·
intronid leiduvad
enamikes geendes puuduvad
· kodeeriv DNA 2% 93% VALGUD
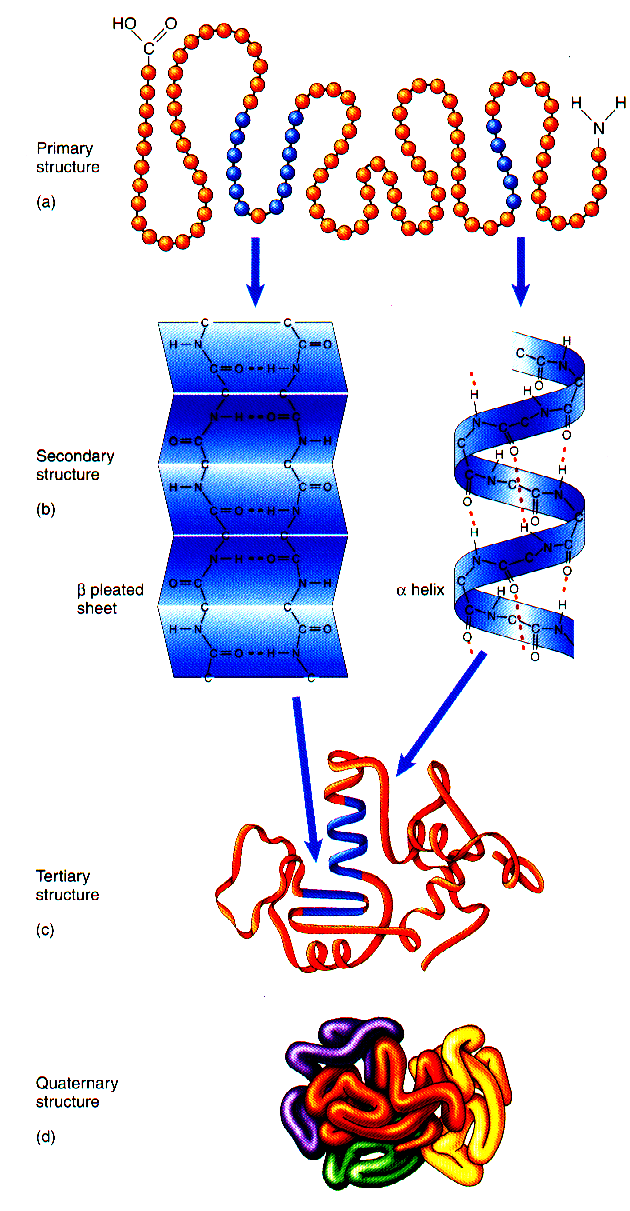
Ehitus. valgu näol on tegemist kõrgmoleklaarse ainega, mille koostisosadeks on
aminohapped .
Primaarseks ehituslikuks tasandiks on aminohappeline järjestus, järgnev tasand on -struktuur või -
heeliks. Tertsiaarselt moodustuvad kas gloobulid või fibrillaarsed
struktuurid . Kvaternaarse tasandi puhul liituvad omavahel kas gloobulid või fibriinid, võimalik ka nende omavaheline kombinatsioon.
Valkude süntees kannab nimetust
translatsioon . Protsess toimub ribosoomides.
Ribosoomid on
rakuorganellid, mis koosneb rRNA moleulidest. Eukarüoodi ribosoom (80 S) koosneb järgnevatest
komponentidest:
1. väike alaüksus ehk 40S, sisaldab 18S rRNA-d ja 33 erinevat valku.
2. suur alaüksus ehk 60S, mis sisaldab 5S rRNA-d, 5.8S rRNA-d, 28S rRNA-d ja 49 erinevat valku.
Neid kromosoomipiirkondi, kus toimub RNA geenide transkriptsioon, nimetatakse tuumakese
piirkondadeks. Ribosoomide
geenid asetsevad
genoomis tandeemsete duplikaatidena, mis on omavahel
eraldatud intergeensete mittetranskribeerivate speisseraladega. rRNA geenide transkribeerimise
tulemusena saadakse prekursorRNA, mis allub posttranskriptsioonilisele protsessingule
Funktsioon. valgud osalevad samuti geneetilise info realiseerimises. See toimub geneetilise koodi abil.
Geneetilises koodis määravad aminohappe ära tripletid.
Kromosoomid Organismide genoom on suuremas osas jaotunud kromosoomidesse. T.
Brown defineeib kromosoomi
kui isereplitseeruvat nukleiinhappe molekuli, mis sisaldab geene. Prokarüootide
kromosoomiks on
rõngas-DNA molekul. Kogu prokarüoodi genoom on pakitud ühte kromosoomi. DNA on kokku
pakitud mitmekümneks linguks, mida hoiab koos RNA. Valkude abil kinnituvad DNA lingud basaalse
alaga bakteriraku sisemembraanile, moodustades nukleoidi ala. Eukarüootidel on nukleoomse
struktuuriga kromosoomid, mis asuvad rakutuumas ja on tsütoplasmast eraldatud tuumamembraaniga.
Lisaks tuumagenoomile on veel tuumaväline mitokondrigenoom, millele taimede ja mõnede vetikate
puhul lisandub kloroplastigenoom. Kromosoomide koostisse kuuluvad DNA, RNA ja valgud.
Kromosoomistikku iseloomustab kromosoomide kuju, mille määrab primaarsoonise ehk tsentromeeri
asukoht. Selle alusel eristatakse telo-, akro-, submeta- ja metatsentrilisi kromosoome. Metatsentriku
tsentomeer
jagab kromosoomi kaheks enam-vähem võrdseks õlaks. Telotsentriku
tsentromeer paikneb
aga telomeeri piirkonnas ja
kromosoomil on visuaalselt
eristatav vaid üks õlg. Inimesel esineb
karüotüübis nii
meta -, submeta- kui ka akrotsentrilised kromosoomid. Kõrgemalt arenenud liikidel on
rohkem asümmeetrilisi kromosoome.
Tuuma DNA hulk on määratud liigi kromosoomide arvu ja suurusega. Igal liigil on genoomis kindel
hulk DNA-d ja seda suurust nimetatakse C-väärtuseks. Antud suurus võib olla väljendatud nii pikogrammides kui ka aluspaarides. On täheldatud, et DNA hulk
varieerub lähedastel liikidel väga
suures skaalas ja samas suur sarnasus morfoloogiliselt väga erinevate ja süstemaatiliselt kaugete liikide
vahel. Seda nähtust nimetatakse C-väärtuse paradoksiks.
Eukarüoodi DNA-seoselised valgud jagunevad kaheks- histoonid ja mittehistoonsed valgud.
Mõlemat tüüpi valkude kompleksid tuuma DNA-ga moodustavad
kromatiini . Histoone nimetatakse ka
kromosoomi struktuurvalkudeks, ülejäänud on regulatoorsed valgud. Histoonid sisaldavad palju
positiivselt laetud aminohappeid arginiini ja lüsiini. Sellest tulenevalt on histoonid aluselised.
Positiivne laeng aitab histoonidel DNA-ga seonduda. Eristatavad on 5 põhilist histoonide klassi: H2A,
H2B, H3 ja H4 kuuluvad nukleosoomi koostisesse; histoon H1 asub
linker -DNA
alas . Protamiinid on
tugevalt aluselised valgud, mis on ehituselt histoonidest lihtsamad ning ei sisalda türosiini. Protamiinid
asendavad histoone spermides, võimaldades DNA väga tugeva kokkupakkimise. Histoone määravad
geenid asuvad eükarüootide genoomis tandeemsete kordustena.
Mittehistoonsete kromosoomivalkude hulka kuuluvad DNA topoisomeraasid, HMG valgud, teatud
DNA-ga järjestsspetsiifiliselt
seonduvaid valke, RNA polümeraasid, DNA süntetaasid ja
proteiinkinaasid. Ensüüm
topoisomeraas I ülesandeks on keerata lahti DNA
keerd , tehes enne lõike
ühte ahelasse. Topoisomeraas II lõhub kaheahelalise DNA, võimaldades DNA lahtikeerrumise
parandab lõikekoha ehk siis katalüüsib DNA
heeliksi pöörduvaid katkemisi.
Eukarüootide kromosoomides on DNA umbes 1200-kordselt kokku pakitud. Kromatiini pakkimise
ühikuks on
nukleosoom . Nukleosoom koosneb umbes 200 bp
pikast DNA lõigust ja histoonsest
oktameerist. Viimase moodustavad histoonid H2A, H2B, H3 ja H4,
kusjuures igaühte on genoomis 2
molekuli. Umbes 140 bp DNA-st keerdub kaks korda ümber histoonse südamiku, ülejäänud DNA on
linker-alas. 2 nm diameetriga DNA kaksikahela keerdumisel ümber histoonse oktameeri moodustub 10
nm kromatiinniit. Kromatiinniit moodustab H1 histooni osalusel 30 nm kromatiinkiu. Järgnevalt,
nukleosoomne kiud moodustab spiraalse konformatsiooni-
solenoidi . On ka mudeleid, mille kohaselt
on solenoidi asemel hoopiski dinukleosoomne
spiraalne lint. 1989. aasta Pienta mudeli järgi
organiseeruvad solenoidid lingudeks. Ling on tõenäolisemalt DNA kõrgemat järku struktuuri põhiühik,
eksisteerides kogu rakutsükli vältel nii spermides kui ka diploidsetes tuumades.
Kromatiini kokkupakkimisel ja
geeniekspressiooni regulatsioonil on olulised nii histoonide
atsetüleerimine kui ka fosforüleerimine. Histoonide fosforüleerimise eest vastutab proteiin
kinaas .
Komosoomide
kondenseerumine on seotud selliste valkudega nagu XCAP-C ja XCAP-E.
Lisa-ehk B-kromosoomid
Igat liiki iseloomustab kindel kromosoomistik, niinimetatud A-kromosoomid. A-kromosoomide
mutatsioonid viivad geneetilise tasakaalu rikkumisele ja on reeglina kahjulikud. Paljudel taime-ja
loomaliikidel (mitte inimesel) võivad karüotüübis esineda ka B-ehk lisakromosoomid, mis on suures
osas heterokromatiinsed ja mille vähest eukromatiini ei transkribeerita. B-kromosoomid on tavaliselt
väiksemad kui A-kromosoomid ning nende arv ja kuju varieerub ühe liigi isenditel ja ka ühe isendi
erinevates rakkudes. B-kromosoomide olemasolu karüotüübis võib paljudel liikidel viia kiasmide
sageduse tõusule A-kromosoomides, suurendades geneetilist rekombineerumist ja ka varieeruvust
järglaste hulgas. B-kromosoomid replitseeruvad S-faasi lõpus, kusjuures rakutsükkel pikeneb vastavalt
lisakromosoomide arvu suurenemisele.
Kromosoomide spetsialiseerunud alad-
· soonised ehk heledamini värvunud alad- primaarsoonis ehk tsentromeer jagab kromosoomi kaheks õlaks. Sekundaarsoonis on soonis, mis pole seotud tsentromeeriga ja avaldub kui heledalt värvunud ala kromosoomis.
· satelliidid- sekundaarsoonistest distaalselt asuvad alad.
·
telomeerid - kromosoomiotsad ehk terminaalsed alad.
· kromomeerid-
tumedad vöödid, mis tekivad kromatiinniidi keerdumise tagajärjel. Neid võib näha meioosi
profaasis tumedate vöötidena.
· kromosoomivöödid- heledamini ja tumedamini värvunud alade
vaheldumine piki kromosoomi. Kromosoomivöödistud saadakse
diferentsiaal - ja selektiivvärvimise tulemusena või taimede puhul näiteks ka temperatuuriga mõjutades. Kromosoomide duplitseerumiseks ja repltseerumiseks on vajalik element replikatsiooni lähtepunkt ehk
origin . Origin on defineeritud kui DNA lõik, mis on vajalik ja piisav DNA replikatsiooni tagamiseks. Replikatsiooni origin peab sisaldama autonoomselt replitseeruvaid järjestusi ehk ARS- e. Kui prokarüootide genoomis on tavaliselt üks origin, siis eukarüootides algab replikatsioon paljudest kohtadest korraga. Tsentromeer ehk cen on kromosoomipiirkond, mis koosneb erinevatest DNA kordusjärjestustest, millega seonduvad tsentromeerivalgud. Tsentromeer
vahendab kromosoomide kinnitumist mitootilise või meiootilise käävi külge, osaleb kromosoomide liikumisel
poolustele ja hoiab koos tütarkromatiide. Tsentromeeri DNA koosneb lühikestest, enamasti alla mõnesaja aluspaari pikkustest lõikudest, mis on
miljoneid kordi tandeemselt korratud. Osas tsentromeerides esineb lihtne järjestus (GGAAT)n, mida peetakse tsentromeerseks konserveerunud järjestuseks. Inimesel paiknevad kõigi kromosoomide tsentromeerses alas alfa.
satelliit -DNA järjestused, mis on 171 bp pikad ja korratud umbes 5000 korda. Kõige paremini iseloomustatud kromosoomivalkudeks on CENP-A, CENP-B ja CENP-E. Kinetohoor on mitmekihiline valguline paaris-struktuur, mis moodustub jagunevas rakus tsentromeeri külge, ehk siisala, millele kinnituvad käävi mikrotuubulid. Kinetohoorid sisaldavad mikrotuubuli mootorvalke ja rakutsükli regulaatorvalke. Kinetohooride vahendusel seonduvad replitseeruvad kromosoomid vastaspoolustele, kinetohoorid aitavad kromosoomidel käävis õigesti paigutuda ja hoiavad ära kromosoomide lahknemise enne, kui kõik kromosoomid on kinnitunud ja õigesti asetsenud. Kinetohoor on kolmekihiline struktuur, mis paikneb kahe kromosoomiõla struktuurse heterokromatiini vahel. Tsentromeerialade analüüs Struktuurse heterokrmatiini alasid saab kromosoomides esile tuua C-vöötide meetodil, töötlemisel restriktaaside või antikehadega ning fluorestsentsanalüüsil.
Barri kehake: selle moodustab üks kahest X-kromosoomist. Barri kehakest võib leida naise teatud
kudedes interfaasi rakkude tuumades, näiteks neeru, maksa ja limaskesta epiteeli diploidsetest
rakkudest.
Telomeer on kromosoomide
otstes asuv järjestus. See hoiab ära DNA degradeerumise ehk lagundamise
nukleaaside ja kromosoomide
omavahelise ots-otsaga
liitumise . Kromosoomiotsad sisadavad kaht tüüpi
järjestusi: lihtsad telomeersed järjestused ja telomeeriga assotsieerunud polümorfsed järjestused.
Telomeerne DNA järjestus on 6-8 bp pikk ja tadeemselt korratud mitusada kuni tuhandeid kordi.
Inimese telomeeris kordub tandeemselt järjestus TTAGGG. See järjestus on arvatavasti olemas kõigil
selgroogsetel kromooomidel. Kuna telomeer lüheneb DNA sünteesil DNA primaasi mõjul, siis
kromosoomi pikkuse säilitamiseks on vajalik pidev telomeersete järjestuste lisamine.
Pikenemine toimub RNA-d sisaldava telomeraasi ehk telomeeri terminaalse transferaasi abil, mis lisab kordusi 3
´otsale. Telomeraasi RNA
molekulil on telomeerse DNA-ga lühike komplementaarne ala, mis täitab
matriitsi rolli telomeersete järjestuste sünteesil. Kõrgematel organismidel hakkab telomeraas
represseeruma kohe peale sündi. Nii hakkavad telomeerid lühenema ja see protsess kujutab endast
mitootilist kella. Telomeraasi reaktiveerumine võib aga viia vähi tekkele.
Telomeeri valke on kirjeldatud kümmekond. Neist osa seaondub vaid telomeerse DNA tipmisele osale, teised aga kogu telomeersele DNA-le. Telomeersed valgud moodustavad telomeerile nii-öelda kaitsva
mütsi. Telomeeri valgud reguleerivad telomeeri pikkust, aitavad kaasa antud järjestuse pakkimisele ja
vahendavad TTAGGG korduste kinnitumist tuuma maatriksile. Telomeeri kromatiini struktuur mõjutab
ka telomeeriga külgnevae geenide ekspressiooni, mistõttu telomeeri vahetus läheduses on geenid sageli
represseeritud. Tuumakese organisaatori piirkond ehk NOR
Tuumakese organisaatori piirkond on kromosoomiala, mis moodustab tuumakese. NOR-is paiknevad
rRNA tandeemselt korratud geenid. Iga
rDNA transkriptsiooniühik sisaldab rRNA geene kodeerivaid
järjestusi ja mittetranskribeeritavat intergeenset speisserit. Inimese 5S RNA geenid, mis on 120 bp
pikad ja mitusada korda tandeemelt korratud, paiknevad 1. kromosoomi pikal õlal ja ei ole seotud
tuumakesega. 18S, 5.8S ja 28S rRNA geenid on 44 bp pikad ja paiknevad akrotsentriliste
kromosoomide 13-15 ja 21-22 NOR-alades.
DNA kordusjärjestused
Kordusala Korduse Korduse arv Kordusühik Lookuste arv Lokalisatsioon tüüp
Satelliit DNA
tandem 10 3-107 1-103 bp Väike (1-2 struktuurses kroosoomi heterokromatiin kohta) is
Minisatelliit tandem 10-103 15-100 bp mõni
tuhat geenide
(VNTR) genoomis vaelistes alades, telomeerides geenide vahelistes alades (ka geenides)
Mikrosatelliit tandem 1-6 bp üle genoomi Geenide
(STR) vahelistes alades
Hajuskordus üksik
(IRS) SINES 105-106 150-300 bp üle genoomi R-vöötides LINES 104-105 1.5-6 kb üle genoomi G-vöötides DNA lateraalsed kordused
Polüteniseerumine on endoreduplikatsiooni
modifikatsioon , misläbi tekivad polüteensed
kromosoomid. Polüteenseid kromosoome on kirjeldatud nii ainuraksetel, taimedel kui ka loomadel.
Neid esineb süljenäärmete rakkudes, Malpighi rakkudes, ovariaalsetes toiterakkudes, naha-ja
sooleepiteelis, trofoblasti rakkudes ja taime endospermi rakkudes. Polüteensed kromosoomid on üle
saja korra suuremad kui
metafaasi kromosoomid. Polüteensed kromosoomid tekivad kromatiidide
korduva replikatsiooni tagajärjel rakutsükli S-faasis. Kromatiidid jäävad kokku ja paarduvad. Tuuma ja
raku jagunemist ei järgne. Tihedalt kokku pakitud kromomeersete alade reastumisel üksteise kõrvale
moodustuvad tumedad vöödid.
Kromoosomikaardid
Geneetilised kromosoomikaardid: antud kaart kujutab endast aheldatud geenide vastastikust
paiknemist piki kromosoomi. Geenide suhtelist kaugust üksteisest arvutatakse rekombinatsioonide
sageduse kaudu ja väljendatakse sentimorganites ehk morganiidides. Mida kaugemal asuvad geenid
üksteisest, seda suurem on nendevahelise ristsiirde tõenäosus. 1 cM (sentimorgan) on suhtelin kaugus
kahe geeni vahel, mille rekombinatsioonisagedus ristsiirdel on 1 %. Rekombinatsioonisagedus ei ole
stabiilne suurus, see varieerub ka sama organismi erinevate kromosoomide puhul.
Füüsilised kromosoomikaardid: jagunevad molekulaargeneetilisteks ja tsütogeneetilisteks.
Tsütogeneetiline kaart näitab tsentomeeri, sekundaarsooniste, kromomeeride, eu-ja
heterokromatiinivöödi asukohti. Molekulaargeneetilised kaardid on kromosoomi füüsikalis-keemilise
analüüsiga määratudjärjestustüübid, ensüümide toimepunktid, geenid ja geenivahemikud. Pikkusi ja
suurusi mõõdetakse nukleotiidipaaride arvuga.
Kromosoomianalüüs Kromosoomiuuringu meetodid
In
vivo mittejagunevad
rakud . Antud olukorras saab kromosoome uurida interfaasi tuumades.
Klassikaline materjal selleks on kahetiivaliste vastsete süljenäärmete polüteensed kromosoomid. Interfaasi kromosoomid on reeglina dekondenseerunud,
pikemad kui metafaasi kromosoomid ja sassis.
Seetõttu ei saa neid
tavaliste meetoditega uurida. Antud kromosoome on võimalik uurida,
kombineerides uusi
tehnoloogiaid ja FISH-tehnoloogiat.
In vivo jagunevad rakud. Tavaliselt analüüsitakse kromosoome mitoosi metafaasi staadium. Kuid
seda uuringut saab teostada somaatilistes kudedes, kus on jagunevaid rakke, meioosi kromosoome aga
generatiivses koes. Inimese kromosoome saab kultiveerimata uurida luuüdi rakkudest, koorioni
hatulisest trofoblastist, kasvajakoest, testikulaarsest ning ovariaalsest koest.
In
vitro jagunevad rakud. Mitmete kromosoomianalüüsiks
sobivate kudede rakud on lõpuni
diferentseerunud ja jagunemise lõpetanud. Sel juhul viiakse kromosoomianalüüs läbi in vitro
kultiveeritud ja jagunema
stimuleeritud rakkudes. Imetajatel uuritakse kromosoome peamiselt
perifeerse vere lümfotsüütides. Kasutatakse kas täisverd või isoleeritud lümfotsüüte. Rakke
stimuleeritakse jagunema
taimsete lektiinidega. Enamkasutatavaks T-rakkude spetsiifiliseks lektiiniks
on türgioa seemnetest eraldatud fütohemaglutiniin (PHA). Esimesed mitoosid
ilmuvad 38-40 tundi
peale stimuleerimist, maksimaalset proliferatsiooni võib täheldada 72-96 tunni möödudes. Tavaliselt
kasutatakse 68-72 tunni kultuuri ja analüüsitakse teist või kolmandat mitoosi. Sünnieelses diagnostikas
kasutatakse kultiveeritud
amnioni - ja koorionirakke. Nii täiskasvanu kui embrüo rakud kasvavad
kultuuris vaid piiratud aja. Selleks, et rakke kauem kasvatada, tuleb need immortaliseerida.
Püsirakuliine saadakse kas spontaanset immortaliseerunud kasvajarakkudest või keemiliste ainete,
kiirguse ja viiruste tomel transformeerunud rakkudest. Tuleb aga arvestada, et pikaajalisel
kultiveerimisel mutub rakupopulatsioon aina heterogeensemaks ja rakud polüploidiseeruvad.
Kromosoomipreparaadi saamine
Kromosoomipreparaadi saamiseks töödeldakse rakke järgnevalt:
· ihibeeritakse mitoosi kääv
· töödeldakse hüpotoonilise lahusega
· fikseeritakse
· tehakse kromosoomipreparaat
· värvitakse Käävi inhibeerimine. Rakkude jagunemise peatamiseks
metafaasis kasutatakse enamasti sügislillest (Colchicum autumnale) saadud alkaloidi kolhitsiin ja selle sünteetilist derivaati koltsemiidi. Mitoosi käävi inhibeerimiseks kasutatakse veel taimest Vinca saadud vähivastaseid ravimeid vinblastiin ja vinkristiin. Hüpotooniline töötlemine. Hüpotoonilise lausena kasutatakse 0.56%
kaaliumkloriidi või 0.8% naatriumtsitraadi lahust. Kui rakud viia hüpotoonilisse lahusesse, siis võtavad need soolade kontsentratsiooni ühtlustamiseks väliskeskkonnast vett ja
paisuvad . Kuna profaasis on
tuumamembraan lagunenud ja mitoosi kääv tänu kolhitsiinile depolümeriseerunud, siis saavad kromosoomid paisuvas rakus vabalt
liikuda . Fikseerimine. Rakke fikseeritakse metanooli ja jää-äädikhappe segus vahekorras 3:1. Metanool denatureerib ja sadestab valgu, äädikhape aga koaguleerib nukleoproteiini. Kromosoomipreparaadi tegemine. Preparaadi valmistamisel tilgutatakse fikseeritud rakususpensioon
puhastatud märjale alusklaasile ning kuivatatakse kas õhus või leegil. Niiskes keskkonnas on kromosoomid
pehmed , kuid muutuvad kõvaks kohe peale kuivamist. Pehmed kromosoomid on väga elastsed, eriti kui neid on väga lühikest aega fikseeritud. Elastseid kromosoome on võimalik faaskontrastmikroskoobi all mitu korda pikemaks venitada nagu kummipaela. Niiskes keskkonnas läbi
viidud kromosoomide pikendamist nimetatakse kromosoomide venitamiseks ehk sirutamiseks (chromosome
stretching ).
Kõige lihtsam on kromosoome värvida tavavärvimise meetodil Giemsa värviga. Töötluse tulemusena
kromosoomid ei vöödistu, vaid värvuvad ühtlaselt. Kõige rohkem kasutatakse Giemsa värvi, mille
koostisesse kuuluvad happeline
eosiin Y ning aluseline metüülsinine ja thaziinvärv azuur B. Värv
muudab kromatiini lillakaspunaseks. Giemsa-kompleksil on üks negatiivne ja kaks
positiivset langut.
Positiivselt laetud molekuli osa inerakteerub negatiivselt laetud DNA-ga, samas kui negatiivselt laetud
osa seostub histoonidega.
Kromosoomivöödid
Kromosoomivööt ehk bänd on kromosoomiala, mis naaberaladest eristub, olles heledam või
tumedam . Kromosoomivöödistus tekib lineaarselt külgnevate heledate ja tumedate vöötide
vaheldumise tõttu. Lisaks vöötidele eristatakse kromosoomimustris veel rajamärke, mille all
mõeldakse selgelt eristatavaid ja kromosoomi identifitseerimisel olulisi morfoloogilisi alasid.
Rajamärkide hulka kuuluvad telomeerid, tsentromeer ja suured iseloomulikud vöödid. Regiooniks
nimetatakse kromosoomipiirkonda kahe rajamärgi vahel.
Vöödistude
tasemed . Kui rakendada kromosoomidel Q-, G- või R-vöödistust, siis saab inimese
kromosoomistikus umbes 300-400 vöödi lahutuse. Pro- ja prometafaasi kromosoomides diferentsiaalvärvimise tulmusena saadud vöödistusi nimetatakse kõrglahutusvöödistuseks, HRB ehk
kõrglahutus-tsütogeneetikaks, HRC. Maksimaalne lahutus on 1000-1250 vööti genoomi kohta. Üks
vööt sisaldab tavaliselt 2-5 Mb DNA-d.
Eukromatiini diferentsiaalvärvimine
Eukromatiini värvimiseks on Q-, G- ja R-vöötide meetodid.
Q-vöötide meetod töötati välja 1968. aastal. Selle meetodi puhul värvitakse kromosoome lühiajaliselt
fluorokroomiga ja analüüsitakse seejärel fluorestsentsmikroskoobis. Töötluse tulemusena vahelduvad
alad tuhmi hiilgusega aladega.
G-vöötide meetod ehk GTG-meetod töötati välja
1971 .aastal Seabright´i poolt. Meetod hõlmab endas
kromosoomide lühiajalist mõjutamist trüpsiiniga ja värvimist Giemsa värviga. Piki kromosoomi
saadakse tumedate ja heledate vöötide muster, mis langeb kokku Q-vöötide hiilgavate ja tuhmide alade
vaheldumisega. Arvatakse, et vöödistus tuleneb kromosoomide
erinevast kokkupakkimisastmest.
R-vöötide meetod ehk RFA- või RHG-meetod töötati välja 1971. aastal Dutrillaux ja Lejeune poolt.
R-vöödid on vastupidised Q- ja G-vöötidele. R-vöötide saamiseks on kaks erinevat võimalust:
1. RFA-meetod: preparaate inkubeeritakse 85ºC juures fosfaatpuhvris, värvitakse akridiinoranziga ja analüüsitakse fluorestsentsmikroskoobis. Punased kromosoomivöödid vahelduvad rohelistega. Akridiinoranz värvib üheahelalise DNA punaseks ja kaheahelalise roheliseks.
2. RHG-meetod: preparaate inkubeeritakse lühiajaliselt fosfaatpuhvris 85ºC juures ja värvitakse Giemsa värviga. Analüüsitakse tavalises mikroskoobis. Töötluse tulemusena vahelduvad heledad alad tumedatega.
Heterokromatiini
esiletoomine kromosoomides
Kõrgematel organismidel moodustab struktuurne heterokromatiin umbes 10% genoomist ja jaotub
karüotüübis kindla mustri järgi. Neid alasid saa kromosoomides esile tuua C-vöötide meetodil,
töötlemisel restriktaasidevõi atikehadega või fluorestsentsanalüüsil. Kuna kõik need meetodid toovad
sageli välja tsentromeeri piirkondades asuva heterokromatiini, siis nimetatakse vastavaid alasid ka C-
vöötideks.
C-vöötide meetod ehk CBG-meetod (C-vöödid baariumhüdroksiidi ja Giemsa värviga). Meetod hõlmab endas denatureerimist leelisega, järgnevat inkubeerimist 65ºC juures ja värvimist Giemsa
lahusega. C-vöödistusega värvuvad valikulkiselt struktuurse heterokromatiini alad nii metafaasis kui ka
interfaasi tuumas. C-vöödid on päranduvad kromosoomielemendid.
Restriktsiooni endonukleaas/ Giemsa meetod sisaldab endas kromosoomide fikseerimist metafaasis
metanool/jää-äädikhappega ja töötlust Dnaasidega. Töötluse tagajärjel tekib C-vöödi sarnane muster.
DA/
DAPI fluorestsentsanalüüs. Kui inimese kromosoome värvida ainult DAPI-ga (4´-6-diamidino-2-
fenüülindool), siis näeb lisaks Q-vöödistusele sarnase mustri ka suuri briljantselt hiilgava
heterokromatiini blokke, kuid seda vaid 1. ja 16. kromosoomi puhul. Kui kromosoome värvida
disramütsiin A-ga ja seejärel DAPI-ga, siis on briljantselt hiilgavad alad näha paljudes kromosoomides.
Nukleosiid-
vastaste antikehadega töötlus. Meetod seisneb DNA denatureerimises UV-ga,
töötlemises 5-MeC AK-ga ja FITC-iga konjugeeritud sekundaarse AK-ga töötlemises. Analüüs toimub
fluorestsentsmikroskoobi abil.
Kromosoomide diferentsiaalvärvimine
Meetod uurimisobjekt I kromosoomide tavavärvimine kromosoomid II kromosoomivöödistused 1. diferentsiaalvärvimise meetodid eukromatiin
· Q-vöödistus
· G-vöödistus
· R-vöödistus 2. Selektiivse värvimise meetodid
· C-vöödistus heterokromatiin
· NOR-vöödistus tuumakese organisaatori piirkond 3. Fluorokroomidega värvimine heterokromatiin 4. Antkehade kasutamine kromosoomide värvimisel heterokromatiin 5. Restriktsiooni endonukleaas/Giemsa vöödistused heterokromatiin III värvimise erimeetodid 1. kõrglahutusvöödistus prometafaasi kromosoomide 2. fragiilsaitide esiletoomine fragiilsaidid 3. õdekromatiidide eristamine kromosoomide
reproduktsioon 4. repliokatsioonivöödistused kromosoomide reproduktsioon IV In situ hübridiseerimise meetodid kromosoomid
Rakutsükkel
Perioodi raku elust alates mitoosist läbi interfaasi teise mitoosini nimetatakse rakutsükliks. Selleks
protsessiks kuluvat aega nimetatakse generatsiooniajaks.
Interfaas hõlmab sellest ajast 90 %. Interfaasi
alla käivad G1, M ning G2 faas. G1 perioodil toimub ettevalmistus DNA sünteesiks, rakk kasvab ja
toodab vajalikke aineid. S-perioodil toimub DNA replikatsioon ja kromosoomivalkude süntees. G2-
perioodistoodetakse peamiselt käävimaterjali ja mitoosiks vajalikku ATP-d.
Selleks, et jaguneda, peab eukarüootne rakk oma massi kahekordistama ning siis
jagama komponendid
võrdselt kahe tütarraku vahel. Massi
suurendamine on pidev protsess, mis tuleneb konkreetse raku
fenotüüpi määravate geenide transkriptsioonist ja translatsioonist.
Rakutsükli läbimine on
rangelt kontrollitud. Rakutsüklis on olemas kaks kontrollpunkti, millest edasi
kulgeb rakutsükkel ainult juhul, kui
eelmine faas on täiesti lõpetatud. Esimeseks kontrollpunktiks on
G1 faasis nn R-punkt ja G2 faasi lõpuosas.
Mõned rakkudest ei jagune üldse. Sel juhul on rakk faasis, mis on sarnane G1 faasile, kuid erinevalt
sellest pole võimeline
sisenema S faasi. Seda nimetatakse G0 faasiks.
Eukarüootset rakutsüklit käivitav
mehhanism koosneb valgukompleksidest, mida kindlas järjekorras
aktiveeritakse.
Rakutsükli
aparaati ja rakuväliseid signaale ühendavad signaalide ülekanderajad. Mitogeensed
kasvufaktorid seonduvad raku pinnal oma spetsiifiliste retseptoritega ja initsieerivad sündmuste
kaskaadi, milles on
kesksel kohal erinevate kinaassete ansüümikomplekside ekspressioon ja
aktiveerumine. Need
spetsiifilised kompleksid koosnevad kahest subühikust: regulaatorsubühikust
tsükiinist ja katalüütilisest subühikust, mida nimetatakse tsükliinsõltuvaks kinaasiks (CDK). Loomaraku mitoosi reguleerivad CDK1 (CDC2) ja tsükliinid A ja B.
Mitoos Selle protsessi abil toimub keha- ehk
somaatiliste rakkude paljunemine. Mitoos koosneb profaasist,
metafaasist, anafaasist ja telofaasist.
Profaas - algab kromosoomide kondenseerumine, kaob tuumake, toimub tsütoskeleti osaline
lagundamine. Moodustub käävisüsteem. Protsessi juhivad kaks tsentrosoomi, mis liiguvad raku
poolustele.Tsentrosoomide baasilt piknevad
mikrotorukesed ehk kääviniidid kromatiidide
tsentromeeridele. Mikrotorukeste kinnitumiseks on tsentromeeride läheduses
spetsiaalsed alad-
kinetohoorid.
Metafaas - toimub kromosoomide lühenemine. Mikrotorukesed ja kromosoomid asetuvad raku
kesktasandile. Moodustub metafaasiplaat.
Anafaas - paarilised tsentromeerid eralduvad üksteisest, kääviniidid hakkavad lühenema ning
tütarkromatiidid liiguvad seejuures raku eri poolustele.
Telofaas- moodustuvad uued tuumakatted, tuumakesed. Kromosoomid dekondenseeruvad.
Telofaasile järgneb tsütokinees ehk tsütoplasma jagunemine ja tütarrakkude teke.
Meioos Tegemist on
rakujagunemise viisiga, mille käigus tekivad sugurakud, kusjuures ühest
eellasest ekib
neli uut rakku. Meioos koosneb reduktsioonijagunemisest ja ekvatsioonijagunemisest, mis koosnevad
faasidest, mis esinevad ka mitoosis. Oluline on ka see, et kahe jagunemise
vahepeal ei toimu DNA
replikatsiooni. Ehk siis tähtis erinevus mitoosi ja mitoosi vahel on see, et erinevalt
esimesest tekivad
meioosi tulemusena haploidse genoomiga rakud.
Meioosi peamised erinevuse võrreldes mitoosiga on järgnevad:
· meioosi profaas kestab kaua
· tuum jaguneb kaks korda järjest ning kahe jagunmise vahepeal puudub S-faas, see tähendab, et kromosoomid replitseeruvad ühel korral.
·
homoloogsed kromosoomid paarduvad ja orienteeruvad vastaspoolustele.
· meioosi-spetsiifiline mehhanism tagab ainult ühe funktsionaalse kinetohoori
moodustumise duplitseerunud tsentromeeri piirkonda.
· homoloogide vahel esineb sageli ritsiire.
· homoloogsed kromosoomid jagunevad tütartuumadesse.
· õdekromatiidid jäävad kokku 2. jagunemise metafaasini.
· kromosoomiarv väheneb kahekordselt.
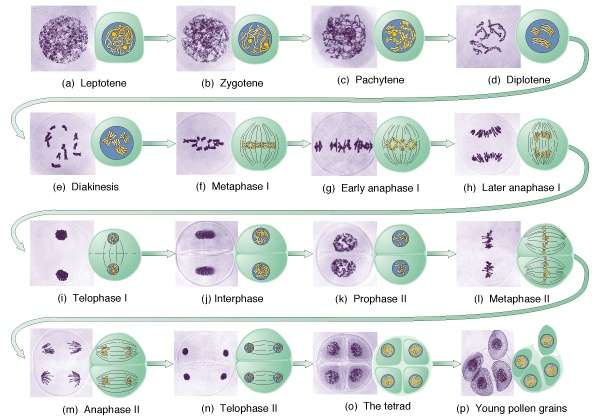
1.
Reduktsioon - ehk taandjagunemine Meioosi I profaas on kõige pikem meioosi staadium ning selles eristatakse 5 staadiumi:
· leptoteen- selles
etapis kromosoomid kondenseeruvad ja muutuvad nähtavaks. Iga
kromosoom kinnitub tsentromeersete aladega
tuumamembraani külge erilise kinnitusdiski abil.
· sügoteen- sünaptonemaalse kompleksi moodustumine algas leptoteeni lõpus ning jätkub sügoteeni vältel.
Esmalt liituvad mõlema homoloogi külge sünaptonemaalse kompleksi lateraalsed elemendid, mis koosnevad valgust ja RNA-st. Lateraalsete elementide vahele tekib
tsentraalne element. Konjugeeritud homoloogseid
paare nimetatakse bivalentideks ja kuna kumbki homoloog koosneb kahest õdekromatiidist, siis nimetatakse taolist struktuuri ka tetraadiks.
· pahhüteen- selles etapis on sünaps täielik. Kromomeeride muster vastab mitoosi kromosoomide G- vöödistuse mustrile. Kui homoloogsed kromosoomid on kogu pikkuses aardunud, siis moodustuvad sünaptonemaalse kompleksi rekombinatiivsed sõlmed, mis kujutavad endast multiensüümkomplekse. Nendes alades toimub pahhüteenis krossingover (
ristsiire ) ehk siis homoloogsete segmentide vahetus kahe mitte õdekromatiidi vahel.
· diploteen- jätkub kromosoomide kondenseerumine.
Homoloogid hakkavad eralduma, jäädes kokku vaid kohtades, kus on toimunud ristsiire. Diploteeni
staadiumis võib meioos
peatuda . Inimese oogeneesis on loote arengu 9. kuuks kõik ootsüüdid jõudnud diploteeni staadiumisse ja seal meioos peatub (staadiumi nimetatakse siis ka diktioteeniks). Edasine meiootiline jagunemine toimub ootsüüdi küpsemse käigus tsükliliselt suguhormoonide toimel. Nii kestab diktioteen ootsüütides 12- 50 aastat. Diktioteeni kromosoomid on lambiharikromosoomid, kus toimub aktiivne RNA ja valkude süntees.
· diakinees- selles staadiumis lõppeb RNA süntees, kaob kromosoomide lambihari välimus ja jätkub nende kondenseerumine. Kromosoomid eralduvad tuumamembraani küljest ning valgusmikroskoobis saab bivalendis eristada 4 kromatiidi. metafaas I- lühikeses prometafaasis kaob tuumamembraan. Järgnevalt kogunevad bivalendid metafaasiplaadile. Kahe mitoositsentri vahele moodustub mitoosikääv. Homoloogsed kromosoomid, mida kuni anafaasini hoiavad koos kiasmid, orienteeruvad juhuslikult pooluste suunas. Anafaas I- homoloogsed kromosoomid segregeeruvad. Tänu ema- ja isapoolsete kromosoomide sõltumatule lahknemisele kombineeruvad mittealleelsed geenid. Võimalike kombinatsioonide arv sõltumatu lahknemise korral on 2n.
· telofaas I- eristub kaks võrdset gruppi kahekromatiidilisi kromosoome, kus igat homoloogi on üks.
· Interkinees- kahe jagunemise vahel toimub kinetohooride ümberorientatsioon vasatavalt teise jagunemise tasapinnale. Kromosoomid ei reprodutseeru.
http://www.bio.miami.edu/dana/250/meiosis.jpg· 2.
Ekvatsioon - ehk võrdjagunemine. Ekvatsioonjagunemine on võrreldes reduktsioonijagunemisega kiirem. Protsess on sarnane mitoosile, kuid kromosoomiarv on haploidne ja kromosoomid on lühemad kui mitoosis.
Krossingover hõlmab sündmusi, mis viivad geneetilisele rekombineerumisele aheldunud markerite vahel. Formaalselt on ristsiire homoloogsete kromosoomide vastavate alade retsiprookne vahetus
sümmeetrilise murru ja ristpidise taasühinemise tagajärjel. Ristsiirdel osalevad kaks kromatiidi
bivalendi neljast kromatiidist. Eukarüootidel esineb nii mitootiline kui ka meiootilne krossingover.
Ristsiire toimub komplementaarsete DNA
ahelate murru ja taasühinemise läbi sünaptonemaalse
kompleksi moodustumise käigus. Tüüpilises bivalendis on vähemalt üks
kiasm . Arvatakse, et
esimene kiasm tekib juhuslikus kohas, järgmine ei saa aga tekkida väga lähedal, sest kahe
kiasmi vahel peab olema teatud
vahemaa . Seda fenoeni nimetatakse kiasmi interferentsiks.








































Kõik kommentaarid