1
GENEETIKA I KORDAMISKÜSIMUSED EKSAMIKS
1. Kaasaegse geneetika rakendusalad meditsiinis ja kohtumeditsiinis.
MEDITSIIN
Geneetilised uuringud on alati olnud suures ulatuses seotud meditsiiniga ja nende eesmärgiks on olnud
meditsiiniprobleemide lahendamine. Need uuringud on võimaldanud leida viise võitluses
nakkushaigustega ning kindlaks teha geene, mis on otsustavad pärilike haiguste tekkel. Geneetikute töö
tulemuseks on ka efektiivselt töötavad vaktsiinid.
1.
Molekulaarne diagnostika ehk teha kindlaks geenid, mis on otsustavad pärilike haiguste tekkel.
Molekulaarsete diagnostikameetoditega on võimalik tuvastada haigusi põhjustavaid mutantseid geene. See
aitab leida optimaalseid ravivõimalusi.
Nt alpaktonuuria on perekonniti päranduv, lisaks huntingtoni tõbi, tsüstiline fibroos.
2.
Geeniteraapia rakendamine. Geeni defekt kompenseeritakse uue, funktsionaalse geeni rakku
viimisega. Nt immuunpuudulikkuse ja tsüstilise fibroosi korral. Terve geen viiakse organismi lisaks
defektsele geenile: Sisseviidud geeni ekspressioonitase on kontrollimatu, Sisseviidud geen integreerub
suvalisse kohta genoomis, võivad tekkida soovimatud mutatsioonid
3.
Sünnieelne diagnostika. See on oluline eriti juhul, kui vanemate suguvõsas esineb geneetilisi haigusi.
4.
CRISPR Cas9 - uus lahendus! Peaks asendama defektse geeni terve geeniga selle õiges
asukohas. Katsejärgus. Praegu inimeste puhul veel ei rakendata, va. Hiinas.
Cas9 endonukleaas teeb DNA-sse vajalikku kohta katked tänu selle kohaga paardunud komplementaarsele
RNA-le. DNA järjestuste deletsioonid, uue järjestuse sisestamine rekombinatsiooni abil
Eelmise sajandi lõpul selgitati samuti, et ka komplekssete haiguste puhul on tegevad konkreetsed geenid.
5. Vähialased uuringud. Geenmutatsioonid. Ka vähk on põhiolemuselt geneetiline haigus.
Kuna rakkude jagunemist ja diferentseerumist kontrollivad geenid, siis nende geenide mutatsioonid
võivad organismi eluajal muuta raku kasvu kontrollimatuks. Seepärast on ka vähk arengubioloogiline
haigus. Üldiselt on vähi tekkel tegemist mitmete geenidega, mis vähi väljaarenemiseks peavad kõik
muteeruma. Vähi tekke sagedust vähendab siiski olukord, kus vastavate geenide mutatsioonid ei pruugi
tekkida ühes ja samas rakus ning rakk ei muutu kasvajalikuks. Mitme mutatsiooni esinemise tõenäosus
suureneb vanusega. Teisalt, kui mingi vähi geene organismis pole, siis puudub sel organismil ka
eelsoodumus vastava haiguse tekkeks.
6.
Farmakogeneetika. Personaalmeditsiin. Inimgenoomi puhul on päevakorda tõusnud personaal- ehk
individuaalmeditsiini küsimused. Ravimid pole universaalsed – nende toime sõltub organismist, tema
genotüübist. Seega, kui meil oleksid nn geneetilised ravimid, mis toimivad vaid kindlate geenide ja
genotüüpide puhul, oleks ravi kindlasti palju edukam. Personaalmeditsiini tungimine igapäevameditsiini
on seniste teadsandmete põhjal veel kauge ootus.
KOHTUMEDITSIINIS – inimeste tuvastamine
DNA fingerprinting. Inimese DNAs on tohutult erineva pikkusega kordusjärjestusi. Praegu kasutatakse
STR (short tandem repeat). Toimub STR-de amplifitseerimine PCR meetodil ja Sõltuvalt korduste arvust
saadakse erineva pikkusega DNA fragmendid
-
Isaduse tõestamine
-
kohtumeditsiinis
2.
Kaasaegse geneetika rakendusalad põllumajanduses. Transgeensed organismid.
Organismi kloonimine.
1
Tänapäeval on paljud taimesordid geneetiliselt modifitseeritud, ehk need sordid sisaldavad
lisageene.
GMO – transgeenselt modifitseeritud organismid
Sordiaretus: Üks võõraste geenide ülekande põhimeetodeid seisneb bakteri Agrobacterium
tumefaciens’i Ti-plasmiidi vahendatud geenide ülekandes taimedesse. GMO-de kasutamine on
tänapäeval rahvusvaheliselt ja riigiti küllalti rangelt kontrollitud. Põhjuseks potentsiaalne biooht:
GMOde kontrollimatu levik looduses, mitteteadaolevate allergiliste või toksiliste ainete süntees
või ravimiresidentsusgeenide horisontaalne looduslik ülekanne. Tootmiseks soovitatud taimedel
neid kahjulikke mõjusid siiani veel kindlalt tõestatuna pole avastatud.
Transgeensed taimed:
Suurenenud vastupanuvõime kahjuritele
Viljade säilivusaja pikenemine (tomat).
“kuldne riis” toodab β-karoteeni. On suurendatud riisi toiteväärtust.
Tõuaretus. Koduloomade tõuaretuses saadakse klassikaliselt suurema muna- piima- või
lihatoodanguga tõuge. Tõuaretusse tõi pöörde kunstliku seemendamise massiline rakendamine,
mis võimaldas hakata jälgima tõuaretuse kulgu. Isaslooma sperma kogumisel ja külmutamisel
saab seda kasutada tuhandeteks seemendusteks.
Järgmisele tasemele jõudis tõuaretus aga loodete siirdamise metoodi väljatöötamisega, mis
võimaldab lühikese ajaga kasi välja vahetada või luua kiiresti uus kari.
Järgmine tase võib saabuda transgeensete ja kloonitud loomade kasutamisega. Siiani on nende
meetodite laialdasem kasutus piiratus oluliste probleemidega elujõuliste ning geneetiliselt
täisväärtuslike järglaste saamisel.
Transgeensed loomad:
tõuomaduste parandamine. Oleks võimalik loomade piimaga koos toota ka muud
vajalikke aineid (nt verehüübe faktorit IX lambaste piimast).
teaduslikud uuringud. Transgeensete hiirte kasutamine: Imetajate geeniekspressiooni
uurimine ja Geenide ülekandevektorite ja meetodite katsetamine, et neid hiljem kasutada
inimese puhul.
Transgeensed kanad, kes on resistentsed ALV (avian leukosis virus) vastu. Nende
organism sisaldab defektset ALV-d, mis produtseerib loodusliku viiruse paljunemist
blokeerivat valku.
Ravimunad: raviotstarbelised valgud ja vaktsiinid munavalges.
Transgeensed mikroorganismid:
Inimese kasvuhormooni ja insuliini tööstuslik tootmine
Organismi kloonimine.
Imetajate kloonimine:
Organismi kloonimise tulemusena saadakse kaks või enam geneetiliselt identset isendit
1. Sisuliselt on kloonid ka ühemunakaksikud, sest nad arenevad ühest viljastatud munarakust
2.
Embrüokloonimine: koduloomade puhul rakendatud varajase embrüo (2-8 raku staadium) rakkude
lahutamist ning nendest identsete järglaste saamist.
3. Kloonimine
lähtudes imetajate somaatilistest rakkudest
4. Somatic cell nuclear transfer –
SCNT
Esimesed kloonitud loomad: 1997 lammas Dolly. Ka lammas Dollyl tekkisid erinevad tervisehäired –
pandi magama. Kloonimise tehnoloogiad on alles algstaadiumis, tehakse palju vigu.
Kloonimise rakendusalad:
2
1. Transgeensete loomade paljundamine
2. Väljasurevate liikide paljundamine
3. (kloonitakse ka koduloomi, eriti Lõuna-Koreas)
Kloonimisega kaasnevad probleemid:
1. Geneetilised ja epigeneetilised efektid kloonimisel
o Kromosoomide aberratsioonid
o Muutused geenide avaldumise mustris, mida mõjutab DNA metülatsioon ja histoonide
atsetülatsioon
2. Edukus on alla 3%.
Inimese reproduktiivne kloonimine on seadusega keelatud
Eriti kui pragu on selles metoodikas veel väga palju vigu. Lisaks inimese isiksus on geenide ja keskkonna
koostoime, ehk kloonides ei saaks sama isiksust. Peale lammas Dolly edukat kloonimist reguleeriti
inimeste kloonimine kiiresti seadusevastaseks.
3. Geneetika väärkasutused.
Meie käitumine ja isiksuseomadused on suures ulatuses geneetiliselt määratud. Alkoholismil ja
skisofreenial on nt geneetiline eelsoodumus. Kui aga keskkonnarisk, nt alkohol, puudub, siis
vastavat haigust ehk alkoholismi ei teki. Loodusliku valiku alusel organismi ebasobivaid
tunnuseid asendatakse kasulikumatega. Galton arendas seda ideed edasi: Kui inimese vaimsed ja
füüsilised tunnused on evolutsioneerunud siis on need ka päritavad ja neile rakendub valik.
Inimsoo kiiremaks parandamiseks saaks kasutada kunstlikku valikut – eugeneetikat.
19. sajandil leiti, et tsiviliseeritud ühiskondades on loodusliku valiku toime vähenenud: kõrgvõimekad
isikud sigivad märksa vähem kui madalamate võimetega inimesed. Kuidas inimest „parandada“?
Eugeenika - teadus kõigist mõjudest ja tingimustest, mis määravad populatsiooni vaimseid ja kehalisi
kvaliteete tulevastes põlvkondades; selle eesmärk on kindlustada iga rass, klass või sekt kehaliselt ja
moraalselt parimate ja vaimselt võimekamate esindajatega.
Eugeenikas oli kaks lähenemist:
Positiivne eugeenika – parimate omadustega isikute sigimise riiklik stimuleerimine (nt rahalised toetused
võimekamate motiveerimiseks).
Negatiivne eugeenika – kahjulike omadustega indiviidide sigimise piiramine (mitmetes riikides rakendati
ainult seda). Piirati vaimuhaigete või muude vigadega, aga ka kriminaalide ja nende, kelle seksuaalne
orientatsioon oli muutunud, sigimist. USAs steriliseeriti indiviidid, keda peeti idiootideks või
retsidivistideks.
Kuigi eugeneetika eesmärgid olid pigem humaansed – inimsoo pirandamine, siis olid eugeneetika
rakendused inimsusevastased.
1895 Alfred Ploetz – kirjutas raamatu “Rassenhygiene”, ei ole tegelikult seotud eugeenikaga, põhines
rasside bioloogilisel ebavõrdsusel (aarialaste rassiline ülimus)
Rassihügieeni seadus 1923 – hakati hiljem nimetama eugeenikaseaduseks. Juutide jt. rahvaste hävitamine
natsistliku Saksamaa poolt. Nii sai eugeenika negatiivse varjundi.
Nii muutus võimatuks ka positiivse eugeenika propageerimine (nt. geeniuste spermapangad ja sellega
võimekate inimeste geene levitada). Kogu eugeenika muutus pikaks ajaks tabuteemaks.
1948 keelati N. Liidus geneetikaalased uuringud täielikult. Põlu alla sattusid ka statistikaalased uuringud
ja küberneetika. Toimus teadlaste sattumine isolatsiooni, paljud kunagi kuulsad Vene teadlased hukati või
surid vangis. Näiteks Nikolai Koltsov (avastas 1903 tsütoskeleti, 1927 oletas, et geneetiline info on
salvestatud 2-ahelalisse hiidmolekuli, mille ahelad on komplementaarsed) , Sergei Tšetverikov (üritas
leida geneetilist põhjust looduslikult valikule).
3
4. Võrrelge eukarüootset ja prokarüootset genoomi.
EUKARÜOOTNE GENOOM
PROKARÜOOTNE GENOOM
TUUM JA
TUUMA-
MEMBRAA
N
On nii rakutuum kui tuumamembraan.
Tuum puudub. Geneetiline info on
koondunud tsütoplasmas raku piirkonda,
mida nimetatakse nukleoidiks.
GEENIDE
ARV
Enamasti üle 5000
Keskmiselt ~1000-5000
VALGU
SÜNTEES
Transkriptsioon ja valgusüntees on teineteisest
lahutatud nii ajaliselt kui ruumiliselt.
1.DNA asub rakutuumas ja seal toimub ka
RNA süntees – transkriptsioon.
2.RNA transporditekse läbi tuumapooride
tsütoplasmasse
3.Ribosoomides, mis paiknevad ERil, toimub
valgusüntees mRNA-s sisalduva geneetilise
info põhjal
Nii kui RNA süntees algab, on
samaaegselt võimalik valgusüntees.
REPLIKATS
IOON
Replikatsioon
algab
paljudest
replikatsioonialguspunktidest,
kuni
replikatsioonikahvlid ühinevad.
Replikatsioon algab ühest kohast, kuni
teeb rõngaskromosoomil ringi peale
INTRONID
Esinevad intronid valke kodeerivates geenides.
Introneid ei esine, splaissing ei ole
vajalik.
RNA
PROTSESSI
NG
Esineb splaissing enne valgusünteesi.
Ei esine. Nii kui RNA süntees algab, on
samaaegselt võimalik valgusüntees
5. Võrrelge raku jagunemist mitoosi ja meioosi teel.
1. GENEETILINE SARNASUS. Mitoosi teel jagunevad tütarrakud on emarakuga geneetiliselt identsed.
Meioosi teel jagunevad tütarrakud on emarakust geneetiliselt erinevad, tagades geneetilise varieeruuvuse
indiviidide vahel.
Meioosis erinevad juba peale esimest jagunemist tütarrakud emarakust, mis peale seda saavad veel üks
kord jaguneda. Lisaks toimub meioosi esimeses profaasis kromosoomide ristsiire ning esimeses anafaasis
liiguvad raku vastaspoolustele homoloogilised kromosoomid, mitte tütarkromatiidid.
2. PLOIDSUS. Mitoosi teel jagunedes on nii emarakk kui tütarrakud on diploidsed. Meioosi teel
jagunedes moodustub kokku 4 haploidse genoomiga gameeti.
3. AEG. Sõltuvalt organismist ja rakkude keskkonnast, võib mitoos kesta mõnest tunnist mõne päevani.
Meioosi puhul võib aga juba ainult esimene profaas osadel loomadel kesta aastaid.
4. RAKUJAGUNEMISTE ARV. Meioosis toimub rakujagunemine 1 kord, kus tekib 2 diploidset rakku.
Meioosis toimub kaks rakujagunemist:
1) redutseeriv jagunemine: Homoloogilised kromosoomid paarduvad omavahel ja lahknevad seejärel
juhuslikkuse alusel tütarrakkudesse.
4
2) võrdväärne jagunemine - Tütarkromatiidid jaotuvad tütarrakkudesse nii nagu mitoosis
5. TOIMUMISKOHT. Mitoos toimub somaatilistes rakkudes, meioos aga idutee rakkudes.
6. DNA REPLIKATSIOON. Kuigi meioosis toimub rakujagunemine 2 korda, on nii meioosis kui mitosis
1 S-faas.
6. Meioos geneetilise muutlikkuse suurendajana. Meioosi häiretest tulenevad defektid.
Mida vanemad eas naine lapse sünnitab, seda suurem tõenäosus on saada defektne laps. Downi sündroom
on levinud. Ka teiste kromosoomide lisakoopiaid esineb, kuid need arenguhäired on veelgi suuremad ja
üldiselt rasedus katkeb ise.
Vead toimuvad kromosoomide jaotumisel tütarrakkudesse. Kuni 5% seemnerakkudest sisaldavad samuti
ebanormaalset kromosoomide komplekti.
Meioosi evolutsiooniline tähtsus:
Isalt ja emalt saadud homoloogiliste kromosoomide juhuslik lahknemine sugurakkudesse.
Esimeses meioosis toimub homoloogiliste kromosoomide juhuslik lahknemine tütarrakkudesse.
Kromosoomide võimalike kombinatsioonide arv on 223 (~1000000 kombinatsiooni).
Ristsiirde toimumine meioos I profaasis suurendab veelgi geneetilist muutlikkust. 1 homoloogilistest
kromosoomidest on pärit isalt ja teine emalt.
-
Suurem võimalus positiivsete mutatsioonide tekkeks ja kinnitumiseks. Kiirem kohastumisvõime.
Kuigi toimub munarakkudes 4 jagunemist, saadakse ikkagi korraga 1 munarakk. Tekib 1 munarakk ja 3
polaarkeha.
7. Mendeli avastatud pärilikkuse üldprintsiibid monohübriidsel ja dihübriidsel ristamisel.
1865 – Mendel summeerib pärilikkuse üldprintsiibid. 1900 Mendeli seadused taasavastati 3 sõltumatu
teadlase poolt.
Modohübriidne ristamine: dominantsuse ja lahknemise printsiip.
Monohübriidne ristamine: vaadatakse ühte tunnustepaari (kääbuskasvulised/kõrgekasvulised)
Mendel ristas kõrgekasvulisi hernetaimi kääbuskasvulistega – hübriidid olid kõrgekasvulised.
Mendeli katsetest järeldus, et geenid esinevad paaridena. Retsessiivne alleel avaldub vaid
homosügootsena.
Mendeli I seadus:
DOMINANTSUSE PRINTSIIP: Erinevate homosügootsete isendite ristamisel on
esimese põlvkonna järglased F1 kõik ühetaolised heterosügoodid. Heterosügootides esineb üks alleelidest
varjatud kujul.
Mendeli II seadus:
SEGREGEERUMISE PRINTSIIP: Kaks erinevat alleeli lahknevad heterosügootide
gameetide moodustumisel.
Heterosügootide (hübriidide)
järglaskonnas F2 toimub geneetiline lahknemine, nii et kindlates
sagedussuhetes tekivad nii homosügootsed kui ka heterüsügootsed
järglased.
Dihübriidne ristamine – Mendeli III seadus
Dihübriidne ristamine: vaadeldud on kahe tunnuse pärandumist, mida
määravad geenid paiknevad erinevates kromosoomides. Kõik
vaadeldavad tunnused peavad olema erinevates kromosoomides,
vastasel juhul seaduspära ei kehti.
GEENIDE SÕLTUMATU LAHKNEMISE PRINTSIIP.
Dihübriidsel ristamisel erinevate geenide alleelid lahknevad ja
kombineeruvad üksteisest tõltumatult.
5
8. Geenide alleelne varieeruvus ja mõju fenotüübile: semidominantsus, kodominantsus,
mitmealleelsus. Tooge näide!
Alleel on täielikult dominantne siis, kui ta annab täpselt samasuguse fenotüübilise efekti homo- ja
heterosügootses olekus (fenotüübiliselt on ju AA ja Aa üksteisest eristamatud). Ebatäielikult või osaliselt
dominantne on heterosügootide(Aa) fenotüüp aga siis, kui ta on homosügootidest (AA/aa) erinev.
Erisatakse kahte tüüpi vastasmõju erinevusi täielikust domineerimisest:
1. SEMIDOMINANTSUS = intermediaansus
2. KODOMINANTSUS
SEMIDOMINANTSUS - F1 põlvkonnas avaldub vahepealne tunnus.
Ristatakse kaks homosügooti (punane värvus, pigmendi puudumine). Järglased on roosade õitega.
Dominantne tunnus avaldub osaliselt.
Kui edasi F1 põlvkonda ristata siis vaid ¼ on punane, ¼ valge ja 2 neljast on roosad.
Õite värvus sõltub geenidoosist. Pigmenti toodetakse nii homosügootsete dominantse alleeli esindajates
kui ka heterosügootides (Aa), kuid seal sünteesitakse vähem ensüümi, mis annab punase õite värvuse.
Dominantne alleel avaldub heterosügootides nõrgemini, tulemuseks õite roosa värvus.
(vähem dominantset alleeli = vähem punast värvi andvat ensüümi)
NÄIDE: Punaste ja valgete õitega lõvilõugasortide ristamisel on järglased roosade õitega.
KODOMINANTSUS - Alleelid avalduvad võrdselt, teineteisest sõltumatult.
Mõlema homosügoodi poolt määratud produkt esineb heterosügootides, ehk heterosügootides avalduvad
mõlema homosügoodi geeni alleelid võrdselt (muidu avaldub dominantne).
NÄIDE: Inimese vererakud võivad toota M ja N antigeeni korraga. Inimestel esineb selline olukord
vererakkude M- ja N-antigeenide äratundmisel. Neid inimese immuunsüsteemi poolt toodetavaid
antigeene, määratakse sama geeni kahe erisuguse alleelse variandi poolt. M- ja N-veregruppidega
homosügoote (AA) saab veretüüpide detekteerimisel selgitada vastavalt anti-M ja anti-N seerumitega
(antikehadega), mis annavad aglutinatsioonireaktsiooni (antigeen+antikeha=sade). Heterosügootidel (Aa)
esineb aga reaktsioon mõlema antiseerumiga, see tähendab, et mõlemad alleelid avalduvad teineteisest
tõltumatult. Nt inimestel on veres kaks antigeeni L ja M.
6
NÄIDE: Rodotendronil võivad olla siiru-viirulised õied. Mõlemad alleelid võivad avalduda.
MITMEALLEELSUS=POLÜALLEELSUS - Tunnuse määravad ühe geeni 3 või enam alleeli. Geen
võib esineda rohkem kui kahe erisuguse alleelina.
NÄIDE: Küülikute kasva värvust kontrollivate geenide alleelid.
c – albiino
ch – himaalaja
cch – tšintšilja
c+ - metsiktüüp. Looduslikus populatsioonis on enamik küülikuid c+ alleeli suhtes homosügootsed.
Homosügooses olekus on igal alleelil oma fenotüüp:
cc – üleni valged
chch – valged jänesed, mustad kõrvad/käpad/ninaots
cchcch – valge karvkate, karvade otsad on mustad
c+c+ - tumeda karvaga
fenotüübiline variatsioon
Metsiktüüpi c+ domineerib kõigi teiste üle. c on retsessiivne kõigi ülejäänute suhtes. cch on osaliselt
dominantne, ehk dominantne ainult ch suhtes.
c+ > cch > ch > c
9. Mutatsioonide toime organismile. Testertüved mutatsioonide alleelsuse testimiseks.
Mutatsioon – muutus geneetilises materjalis.
Mutatsioonide jaotus nende toime alusel:
a. Nähtavad mutatsioonid – muudavad fenotüüpi
b. Neutraalsed mutatsioonid – ei muuda fenotüüpi
c. Steriilsed mutatsioonid – ei võimalda mutatsiooni kandvatel
isenditel järglasi saada
d. Letaalsed mutatsioonid – ei arene elujõulist organismi
1. Retsessiivne amorfne mutatsioon, funktsiooni kaotanud alleel – vajalikku valku ei sünteesita,
tugev fenotüübiline efekt homosügootses olekus
2. Retsessiivne hüpomorfne mutatsioon, kus polüpeptiidi funktsioon osaliselt säilib. Mutantne
fenotüüp võib ilmneda aga tunduvalt nõrgemalt.
3. Dominantne neomorfne mutatsioon, on tekkinud uue funktsiooniga alleel – uute omadustega
polüpeptiid, mis põhjustab uue, mutantse fenotüübi
Heterosügootide fenotüüp erinevate mutantsete alleelide puhul
a – retsessiivne amorfne (funktsiooni kaotanud) alleel – metsiktüüp
ah – retsessiivne hüpomorfne (funktsioon osaliselt säilinud) alleel – metsiktüüp
aD – dominantne neomorfne (uue funktsiooniga) alleel – mutantne
a+ – metsiktüüpi alleel
7
RETSESSIIVSETE MUTATSIOONIDE ALLEELSUSE TESTIMINE
Mutatsioonid võivad olla ka erinevates geenides. Kuidas teha kindlaks kas mutatsioonid on samas geenis
(alleelsed) või erinevates geenides (mittealleelsed)?
Seda saab kindlaks teha retsessiivsete mutatsioonide korral, mitte aga dominantsete mutatsioonide korral.
Mutatsioonide alleelsust saab kontrollida, ristates neid
testertüvedega – neis on retsessiivsete
mutatsioonide esinemine kindlates geenides eelnevalt välja selgitatud.
Ristamisse võetav testertüvi on homosügootne teatava geeni retsessiivse alleeli suhtes. Kaks mutatsiooni
on sama geeni alleelid, kui hübriidide fenotüüp on mutantne. Metsiktüüpi fenotüübi puhul on
mutatsioonid erinevates geenides (ei ole alleelsed)
Nt: sabata kassid manni saarelt. Kui mõlemad alleelid on homosügootses olekus siis loomad ei arene välja.
Kui on heterosügootne, siis sünnib sabata kass.
10. Geenide fenotüübilist avaldumist mõjutavad tegurid. Mõisted penetrantsus ja
ekspressiivsus.
Füüsikaline keskkonna mõju. Keskkonna füüsiline mõju mõjutab geenide avaldumist
Temperatuuritundlikud mutatsioonid – äädikakärvse shibire mutantidel. 25 kraadi juures puudub
nähtav efekt – siiski surevad raputamisel. 29 kraadi juures paralüseeruvad kärbsed koheselt ilma
raputamata.
Fenüülketonuuria – retsessiivne aminohapete metabolism efekt. Mutatsiooni osas homosügootsetel
väikelastel koguneb ajju toksiline ühend, mis põhjustab vaimset alaarengut. Toksilise ühendi
kogunemist saab takistada, kui dieedis vähendada aminohappe fenüülalaniini hulka minimaalsele
tasemele.
Bioloogilise keskkonna mõju
Kiilaspäisus. Kiilaspäisust määrav alleel avaldub sugupooltel erinevalt: meestel dominantsel ja naistel
retsessiivselt. Põhjuseks on meessuguhormoon testosteroon, mida on meestel rohkem.
GEENI
PENETRANTSUS – juht, kus indiviidil ei ilmne tunnust isegi siis, kui tal esineb vastav
genotüüp. Geeni penetrantsuse näiteks on inimesel polüdaktüülia – lisasõrmede või lisavasvaste
olemasolu. Seda põhjustab dominantne mutatsioon P, mis avaldub vaid osadel selle alleeli kandjatel.
Penetrantsus – sagedus protsentides, millega mingi genotüüp avaldub selle kandjate fenotüübis (sõltub nii
geneetilisest taustast kui ka elukeskkonnast)
EKSPRESSIIVSUS – mõistega geeni ekspressiivsus märgitakse olukorda, kus samaste genotüüpide
korral on tunnuste avaldumise aste isenditel erinev. Nt Habsburgide dünastia liikmetle iseloomulik
etteulatuv lõug avaldus indiviiditi erinevalt.
8
11. Mõisted epistaas ja pleiotroopsus. Tooge mõni näide.
EPISTAAS – Ühe geeni mahasuruv (tõkestav, varjutav) toime teise geeni avaldumise suhtes. Geenid,
mida alla surutakse on hüpostaatilised.
NÄIDE: epistaas lillherne õievärvuse kujunemisel. Õie värvust määravat pigmenti antotsüaani
sünteesitakse vaid siis, kui ainevahetusahela mõlematel geenidel esinevad dominantsed alleelid. Kuivõrd
geenid C ja P kodeerivad erisuguseid antotsüaniini sünteesi etappe, siis purpurne värvus saab esineda vaid
mõlema geeni funktsionaalsete produktide olemasolul (alleelid C ja P).
PLEIOTROOPSUS – Kui geen mõjutab mitme teise geeni avaldumist, nimetatakse seda geeni
pleiotroopseks.
NÄIDE: fenüülketonuuria puhul on esmasefekti põhjustajaks retsessiivne mutatsioon, mis tingib
ainevahetuse käigus fenüülalaniinist toksilise aine moodustumise mis omakorda mõjutab aju närvirakkude
arengut ning sellega kaasneb vaimne alaareng. Samal retsessiivsel mutatsioonil on teinegi efekt: see
vähendab pigmendi melaniini sünteesi. Seetõttu on fenüülkenotuurikud tavaliselt helepruunide või
blondide juustega.
12. Mis põhjustab kompleksse tunnuse erinevat avaldumismäära populatsioonis?
Komplekssed tunnused – mitme geeni poolt kontrollitud, polügeensed. Komplekssete tunnuste puhul
ilmneb pidev fenotüübiline varieeruvus – nt inimeste nahavärvus. On leitud umbes 20 geeni, mis osalevad
suuremal või vähemal määral naha pigmentatsiooni määramisel.
Kvantitatiivseid tunnuseid määravad paljud geenid
Kvantitatiivsete tunnuste avaldumist mõjutavad paljud keskkonnafaktorid.
Tunnuste pidev muutlikkus on määratud kahe teguri poolt:
1) Kaks ja enam alleelipaari lahknevad ja kombineeruvad järglaskonnas üksteisest sõltumatult
2) Keskkonna mõjutused
9
KESKKONNA MÕJU FENOTÜÜBILISELE VARIEERUVUSELE.
Katse tubakataimedega, mis erinesid õiekroonide pikkuse poolest, õiekroonide pikkust kontrollivad 9
erinevat alleelipaari, ehk 9 geeni.
NII KESKKONNA KUI KA ERINEVATE ALLEELIPAARIDE
SÕLTUMATU LAHKNEMISE JA KOMBINEERUMISE OSA JÄRGLASKONNA PIDEVA FENOTÜÜBILISE
MUUTLIKKUSE MÄÄRAMISEL.
Mida rohkem geene (ja nende alleele) kontrollib antud tunnust, seda sujuvam on fenotüübiline varieeruvus
populatsioonis
10
13. Kvantitatiivsete tunnuste analüüs: keskmine ja modaalklass, valimi varieeruvus ja
standardhälve.
Kvantitatiivseid tunnuseid iseloomustatakse kvantitatiivselt. Uurimise all on juhuslik osa populatsiooni
isenditest ehk
valim ning nende analüüsist saadud tulemustest tehakse üldistused kogu populatsiooni
kohta.
(X) Valimi keskmine: peab andma populatsiooni iseloomustava tüüpilise tunnuse keskmise väärtuse.
Keskmise (X) leidmiseksliidetakse kõigi isengite andmed (Σxi) ja jagatakse need uuritud isendite arvuga
(n).
X= Σxi/n
Modaalklass: väärtusklass, kuhu jaotub analüüsitud valimist kõige enam isendeid. Nii nagu valimi
keskmine iseloomustab ka modaalklass populatsiooni keskmist väärtust. Enamasti on kvantitatiivsete
tunnuste väärtusi iseloomustav kõver keskmise väärtuse suhtes sümmeetriline. Sellist jaotust nimetatakse
normaaljaotuseks, mille korral populatsiooni keskmine ja modaalklass langevad kokku jaotuskõvera
keskele.
Valimi keskmine ja modaalklass ei iseloomusta andmete hajuvust, ehk seda, kui suures ulatuses andmed
keskmisest väärtusest erinevad.
Valimi varieeruvus mõõdab üksikute andmepunktide hajuvust (s2)
keskmisest punktist ja seda arvutatakse:
s2 = Σ (xi – X)2/(n-1)
Standardhälvet (s) kasutatakse hajuvuse iseloomustamiseks ilma ruutkorrutiseta arusaadavuse
suurandamiseks.
s =
√s2
14. Päritavus. Mida näitab see, kui teatava tunnuse päritavuskoefitsient on väärtusega 0,7?
Päritavus – kvantitatiivse tunnuse populatsioonisisese muutlikuse osa, mis on tingitud indiviidide
genotüüpide erinevusest (genotüübilisest muutlikusest)
Selleks, et mõõta päritavuse osa tunnuse muutlikkuses, tuleb määrata tunnuse kogu muutlikkus Vt ning
leida selles geneetilistest erinevustest tingitud muutlikkuse Vg ja keskkonnateguritest tingitud muutlikkuse
Ve osa. Seega:
Vt = Vg + Ve
Päritavuskoefitsient h2 - väljendab geneetilise muutlikkuse suhteosa Vg tunnuse üldisest
populatsioonisisesest muutlikkusest Vt antud keskkonna tingimustes. See näitab, kuivõrd on fenotüübiline
tunnus põhjustatud geneetilisest erinevusest.
h2 = Vg / Vt
Nt kui nisuterade h2=0,7, siis see tähendab, et 70% ulatuses on nisuterade küpsemine põhjustatud isendite
geneetilisest erinevusest.
Väärtus 0 – fenotüübiline muutlikkus ei ole tingitud genotüübilistest erinevustest
Väärtus 1 – kogu fenotüübiline varieeruvus on põhjustatud geneetilistest faktoritest
15. Kunstlik valik, sellega seotud piirangud. Inbriidingu mõju organismi fenotüübile.
Teades kvantitatiivsete tunnuste päritavuskoefitsienti, on võimalik ennustada kunstliku valiku teel
saadud järglaskonna kvantitatiivseid tunnuste omadusi. Kunstlik valik seisneb peamiselt valitud
genotüüpidega (või fenotüüpidega) isendite ristamises ning järglaste valikus geneetiliste omaduste järgi
.
Kunstlikul valikul kasutatakse kaht põhilist valikumeetodit:
1. Individuaalvalik – järglastest valitakse ristamiseks välja konkreetsed isendid, kasutades
fenotüüpide ettearvamise meetodit
11
2. Massvalik – ristamiseks valitakse vanemate hulgast välja kindlate omadustega väike grupp
isendeid.
Inbriiding – lähedalt suguluses olevate isendite ristumine. Järglaskonnas suureneb
homosügootide aste, lähisugulaste lastel on suurem risk geneetilistele haigustele.
Nt on inbriidingu efekt albinism, ehk pigmendi puudumine. Pigmendi puudumise oõhjustav allele
sattus sugulusabielude tõttu homosügootsesse olekusse.
16. Sugukromosoomid erinevatel organismidel. Eksperimentaalsed tõendid selle kohta, et
pärilikkus on seotud kromosoomidega.
Homogameetne sugupool – moodustab sugukromosoomide osas ühesuguseid sugurakke (gameete)
Naised XX, sugurakus lisaks autosoomidele X kromosoom
Heterogameetne sugupool – moodustab sugukromosoomide osas erinevaid sugurakke
Mehed XY, sugurakus lisaks autosoomidele kas X või Y kromosoom
Sugukromosoomide arv võib liigiti erineda:
Osadel putukatel avastati lisakromosoomid. Nt on emastel rohutirtsudel üks X-kromosoom rohkem
kui isastel. Emased XX, isased X0.
Inimestel, enamikel imetajatel ja osadel putukatel on XY-soomääramise süsteem.
Inimese Y kromosoom on X kromosoomist morfoloogiliselt eristatav. Ühist geneetilist materjali on X
ja Y kromosoomil vähe.
Lindudel ja osadel reptiilidel ja putukatel on olukord vastupidine: isased on homogameetsed (ZZ) ja
emased heterogameetsed (ZW). See on ZW-soomääramissüsteem.
Ekspetimentaalsed tõendid sellele, et geenide pärandumine on seotud kromosoomidega:
Geen white on suguliiteline geen, sest paikneb sugukromosoomis. X kromosoomis – X-liiteline geen.
Thomas Morgan näitas, et punast silmavärvust määrav geen white asub äädikakärbsel X kromosoomis. F2
põlvkonnas olid pooled isastest kärbestest valgesilmsed, kuid emased kõik punasesilmsed. Isased on
punast silmavärvid määrava geeni osas geeni osas hemisügootsed – neil on ainult üks geenikoopia.
Valgesilmsus avaldub emastel järglastel ainult siis, kui mõlemad X kromosoomid sisaldavad retsessiivset
mutatsiooni white.
Tulemuseks ootuspärasest erineva fenotüübiga järglased (punasesilmsed isased ja
valgesilmsed emased). Need tulemused kinnitasid pärilikkuse seost kromosoomidega.
Ta näitas, et silmavärvust määrav geen paikneb X-kromosoomis ja et Y-kromosoomis vastav geen puudub.
Silmade valget värvust põhjustas retsessiivne mutatsioon, mis avaldus isaskärbestel.
17. Selgitage Mendeli seadusi lähtudes kromosoomiteooriast.
12
Mendeli seadused tulenevad sellest, milliste seaduspärasuste alusel toimub kromosoomide lahknemine
sugurakkudes ning sattumine järglaskonda.
Mendeli katsetes vaadeldud tunnuseid määravad geenid asuvad erinevates kromosoomides.
Lahknemisseadus (Mendeli II seadus) – heterosügootide järglaskonnas toimub geneetiline lahknemine, nii
et homosügootseid ja heterosügootseid isendeid saadakse kindlates sagedussuhetes.
Anafaasis, pärast esimest meiootilist jaotumist liiguvad heterosügootsete vanemate kromosoomid, mis
sisaldavad erinevaid alleele, raku erinevatele poolustele ja lahknevad tütarrakkudesse.
Dihübriidsel ristamisel ilmnev (Mendeli III) seadus (sõltumatu lahknemise e. vaba kombineerumise
seadus) baseerub samuti homoloogiliste kromosoomide juhuslikul lahknemisel tütar-rakkudesse
Mendeli seadused lähtudes kromosoomiteooriast:
18. Suguliitelised geenid ja nende avaldumine inimesel. Tooge näide.
X-liiteliste geenide pärandumine seondub X-krmosoomiga.
X-liitelised geenid inimesel
X kromosoom moodustab 4% genoomist, sisaldab 1098 geeni.
10% teadaolevatest geneetilistest haigustest, mis päranduvad Mendeli seaduste järgi, on X-liitelised.
13
Nt: Hemofiilia – X-liiteline retsessiivne tunnus, mida avaldus Euroopa riikide kuninglikes perekondades
tänu sugulusabieludele.
X-liiteline hemofiilia vorm A: puudub verehüübefaktor VIII. Hemofiiliat põhjustab mutatsioon geenis
HEMA, mis kodeerib verehüübe faktorit VIII.
Y kromosoomi-spetsiifilised geenid
TDF (testes determining factor), tuntud ka nimetuse all SRY (sex-determining region Y gene) mõjutab Y
kromosoomis asuvate geenide avaldumist, mis kontrollivad testiste arengut.
Pseudoautosomaalsed regioonid Y kromosoomi otstes on homoloogilised X kromosoomiga. Meioos I
profaasis paarduvad Y kromosoomi otsad X kromosoomidega ning nende alade vahel võib toimuda
rekombinatsioon (ristsiire). XX mehed, kellel puudub Y kromosoom, kannavad SRY regiooni X
kromosoomis
NÄIDE:
Looduses esinevad metsiktüüpi äädikakärbsed on punasilmsed.
Punast silmavärvust määrava geeni dominantne alleel – w+
Valgesilmsust määrava geeni retsessiivne alleel – w
Punaste emaste (w+w+) ristamisel valgesilmsete isastega (w): F1 põlvkonnas kõik punasilmsed, F2
põlvkonnas on puna- ja valgesilmsete suhe 3:1, nagu autosoomsel monohübriidsel ristamiselgi kuid
erinevus seisneb selles, et valgesilmsus esineb vaid isastel. Isastel on pooled puna- ja pooled valgesilmsed.
Järelikult: toimus suguliitelisel pärandumisel F2 põlvkonnas tunnuste lahknemine sõltuvalt soost.
19. Soomääramine erinevatel organismidel.
Lahksugulised – liigid, kelle puhul eristuvad selgelt isased ja emased.
Osadel juhtudel on lahksugulisus määratud pigem keskkonnafaktoritega kui kromosoomidega. Näiteks
osadel kilpkonnaliikidel ja krokodillidel määrab soo temperatuus. Munade haudumisel temperatuuril üle
25 kraadi moodustuvad kilpkonnadel emasjärglased, madalamal isasjärglased. Krokodillidel on vastupidi:
madalal temperatuuril moodustuvad emased, kõrgemal isased. Temperatuuril on sugu määravate geenide
aktiveerimise roll.
Inimesel ja teistel imetajatel määrab soo Y kromosoomi olemasolu.
XO indiviidid on naissoost
XXY indiviidid on meessoost
14
Testist määrav faktor TDF on geeni SRY ehk sugu määrava piirkonna produkt. TDF puudumisel (SRY
geeni defektsus) avalduvad naistele iseloomulikud sootunnused. Järelikult on SRY piirkond isasust määrav
segment ja see on selgelt vajalik isasorganismide tekkeks. SRY geen Y kromosoomis on seotud inimese
sootunnuste kujunemisega.
XX mehed on väiksemat kasvu, enamasti väiksemate testistega ja steriilsed
XY naised steriilsed, neil ei teki munarakke
Kui testised on moodustunud, siis algab hormoon testosteroon sekretsioon. Testosteroon seondub paljude
rakutüüpide retseptoritega. Pärast seondumist kannab hormoon-retseptorkompleks signaali üle tuuma,
määrates sellega, kuidas rakk edaspidi diferentseerub. Eri tüüpi rakkude diferentseetumisega saavutatakse
isasorganismide esmalt primaarsete sugutunnuste (genitaalid) ja edasi sekundaarsete sugutunnuste areng
(tugev lihaskond, habe, jäme hääl). Kui testosterooni signaalsüsteem ei tööta, siis testosterooni poolt
indutseeritud tunnuseid ei moodustu ja organism areneb emasorganismiks.
Tfm on X-liiteline geen ja avaldub alles pärast testiste arengut, sest alles siis toimub testosterooni
sekretsioon. Testosterooni retseptori Tfm geeni defektsuse korral (ehk rakud ei moodusta
testosterooniretseptorit) ei kujune välja meestele iseloomulikud sootunnused (habe, muskulatuur, madal
hääl) vaid hoopis naiste sekundaarsed sootunnused.
Olukord: XY genotüübiga isenditel moodustuvad algul testised kui testosterooniretseptori puudumise tõttu
signaal edasi tuuma ei kandu ja organism areneb emasorganismiks – niisuguse pöördega tekib arengus
olukord kuis väljaareneval naisel munasarju ja emakat ei moodustu – nad on steriilsed. See on
androgeenitundetuse sündroom.
20. Kuidas on tagatud X-liiteliste geenide võrdne avaldumistase erinevast soost isenditel
imetajatel ja äädikakärbsel?
X-liiteliste geenide doosikompensatsioon:
1) Äädikakärbsel toimub isastel X-liiteliste geenide hüperaktivatsioon
2) X-liiteliste geenide inaktivatsioon imetajatel
Genotüübi XX korral on üks X kromosoomidest kondesneerunud ja nähtav Barri kehakesena. Barri
kehake kinnitub tuumamembraani sisepinnale.
Naise sugurakkude moodustumisel aktiveeritakse mõlemad X kromosoomid
X kromosoomi inaktivatsioon algab X-inaktivatsiooni keskusest XIC. Lisaks XIST RNA-le, mis on vajalik
inaktivatsiooniks, avaldub inaktiveeritud X kromosoomis >15% geenidest. Lisaks avaldub selles
kromosoomis veel 10% X-liitelisi geene ja seda erinevatel naistel erineval määral – suurendab
fenotüübilist varieeruvust
Regulatoorsete RNA-de roll X kromosoomi inaktivatsioonil
Xist RNA inaktiveerib X kromosoomi
Tsix, mis on komplementaarne Xist RNA-ga, paardub sellega ja surub maha Xist RNA ekspressiooni
21. Mitoosi- ja meioosikromosoomide uurimise tsütoloogilised meetodid.
15
Enamus uuringuid teostatakse mitoosi metafaasi kromosoomidega.
Rakkude töötlemine kromosoomide analüüsiks
Kromosoomide nähtavale toomiseks kasutatakse
erinevaid värve. Kromosoomide diferentsiaalvärvimise
meetodid põhinevad AT ja GC piirkondade erineval
afiinsusel värvainete suhtes ja võimaldavad eristada nii
üksikkromosoome kui ka kromosoomide eri osi mööda
nende pikitelge.
Feulgen’i reagent – reageerib DNA-s olevate suhkrujääkidega
DNA-ga interkaleeruvad, UV-s fluorestseeruvad (Quinakriin)
DNA-ga interkaleeruvad, mittefluorestseeruvad (Giemsa)
Erinevad värvid toovad nähtavale erinevad vöödid
Q-vöödid – Tekivad kromosoomide AT rikastes piirkondades töötlemisel kinakriiniga, mis on
malaariaravimi kiniini keemiline derivaat. See on fluorentseeruv ühend, mis UV valguses annab
helenduse. Värvainega mitteseondunud piirkonnad ei helendu.
G ja R-vöödid – Giemsa värv. Mittefluorestseeruv värv.
G vöödid: värv seondub eelistatult AT-rikastele aladele (heterokromatiin)
R- vöödid: preparaadi eeltöötlus kuumutamisega happelises keskkonnas, kus AT-rikkad alad
denatureeruvad; seejärel värvuvad paremini GC-rikkad alad
Igale kromosoomile on iseloomulik kindel vöödilisuse muster.
22. Inimese karüotüüp ja karüogramm.
Karüotüüp – liigiomane kromosoomistik. kromosoomistiku tunnustekogum, mida iseloomustab
kromosoomide arv, suurus, tsentromeeri asukohast olenev kuju ja vöödilisus
Vastavalt tsentromeeri olemasolule liigitatakse kromosoomid:
Tsentromeeri puudumisel – atsentrikud
Tsentromeeri olemasolul – monotsentrikud, ditsentrikud, jne
Tsentromeeri asukohast lähtuv liigitus:
Metatsentrikud
Submetatsentrikud
Akrotsentrikud
Karüogramm – kindlal viisil korrastatud ja süstematiseeritud kromosoomistiku fotokujutis. – indiviidi
karüotüübi uurimiseks kasutatav kromosoomistiku süstematiseeritud fotokujutis ühe raku metafaasi
kromosoomidest, kus kromosoomid on reastatud ja rühmitatud suuruse, kuju ja vöödimustri järgi
Inimesel on 46 kromosoomi – 44 autosoomi ja 2 sugukromosoomi.
Mitoosi metafaasi kromosoomid fotografeeritakse ja positiividelt saab nad välja lõigata ja järjestada, sest
igal kromosoomil on oma spetsiifilinekuju ja vöötmuster. Suurim kromosoom kannab järjekorranumbrit 1
ja vähenevas suuruse järjekorras kuni autosoomini nr 22. X-kromosoom on vahepealse suurusema ja Y-
kromosoom on umber nr 22 suurune.
16
A – suured metatsentrikud
B – suured submetatsentrikud
C – keskmised submetatsentrikud
D – suured akrotsentrikud
E – väikesed submetatsentrikud
F – väikesed metatsentrikud
G – väikesed akrotsentrikud
23. Polüploidsus ja selle fenotüübilised efektid.
Ploidsus – homoloogiliste kromosoomide kordsus
n = basaalarv, kromosoomide arv ühes kromosoomikomplektis
Polüploidsed rakud omavad mitut kromosoomikomplekti. 3n, 4n, 5n, ....
Polüploidsus:
1. Takistab soomääramise mehhanismi toimimist
2. Esineb peamiselt taimedel, mis paljunevad ka vegetatiivselt
3. Polüploidid on suuremad. Nt polüploidsed kultuurtaimed (maasikad nt)
Maasika kultuursordid on oktaploidsed (8 koopiat kromosoome). Metsmaasikas on diploidne.
Polüploidsus loomadel:
Harva esinev, põhjustab steriilsust
Sagedasem kaladel, amfiibidel, sisalikel, putukatel.
Nt: triploidne ahven on suurem, kiirema kasvuga, haigustele vähem vastuvõtlik
Nt: Triploidne auster on suurem ja haigustele resistentsem; paremad maitseomadused
Imetajatele on polüploidsus üldjuhul letaalne
Erandiks vizcacha rott (Tympnoctomys barrerae) Argentiinas. Arvati, et on tetraploidse genoomiga, sest on
poole rohkem kromosoome. 2017 leiti, et suurenenud on ainult kordusjärjestuste osa genoomis
Paljud polüploidsed liigid on steriilsed, kuna meioosi käigus lahknevad kromosoomid ebaregulaarselt –
tekivad aneuploidsed gameedid.
Triploidseid taimi paljundatakse vegetatiivselt (näit banaanid, õunapuud, tulbid)
24. Viljakate polüploidide saamine. Tooge näide.
Viljaka tetraploidi teke: kahe diploidi hübridiseerimisele järgneb nende kromosoomide kahekordistumine
e. Endoreduplikatsioon. Hübriidi kromosoomistiku kahekorditumine, kus mõlema liigi kromosoomid
moodustavad paarid ja nende lahknemine meioosi anafaasis on regulaarne ning taimed on fertiilsed.
Kromosoomikomplekti kahekordistumine saam toimuda nt juhul kui moodutuvad gameedid, kus
kromosoomid ei ole poolustele lahknenud. Hübriidsed tetraploidid sisaldavad seeda vanemorganismide
duplitseerunud kromosoomikomplekte.
Seega, viljakate allopolüoidide saamiseks on vaja kromosoomistik kahekordistada ning saavutada olukord,
kus iga liigi kromosoomikompleks oleks esindatud paarisarvulises korduses.
17
Allopolüploidid tekivad lähedaste liikide ristamisel (hübridiseerimisel) ja on enamasti viljakad
Autopolüploidid saadakse sama liigi siseselt ja on harva viljakad.
Krookuse allopolüploidi teke kahe eellase hübridiseerimisel, kellest ühel oli 4 ja teisel 3 paari
kromosoome
Fertiilne heksaploidne nisu on saadud erinevate metsikute eelasliikide järjestikulise
hübridiseerimise tulemusena. Kaasaegne nisu sisaldab kolme erinevat kromosoomikomplekti 7-st
kromosoomist, mille puhul toimus endoreduplikatsioon, nii et somaatilistes rakkudes on 42
kromosoomi
Lisaks kromosoomide duplitseerumisele liikidevahelistes hübriidides võivad polüploidsed taimed
areneda ka meristeemrakkude mitoosihäirete tagajärjel. Tütarkromatiidid ei lahkne mitoosi käigus
ja moodustuvad tetraploidsed rakud
25. Polüteenkromosoomide moodustumine ja omadused.
Polüteenkromosoomid – polüploidiseerumine viisil, kus DNA replikatsioonil tekkinud tütarkromatiidid
ei eraldu.
Osadel juhtudel tütarkromatiidid ei eraldu teineteisest vaid hoopis replitseeruvad ja korduvalt,
moodustades polüteenkromosoomid.
Polüteenkromosoome on kirjeldatud Drosophila vastsete süljenäärmetes, nähtavad interfaasi rakkudes.
Replitseerunud kromosoomid jäävad üksteise kõrvale, rakk ei pooldu. Geenide koopiaarv raku kohta
suureneb oluliselt, suurendades rakkude metaboolset aktiivsust. Rakud on suuremad. Iga kromosoom
replitseerub vähemalt 9 korda – tekib üle 500 koopia
Polüteenkromosoomide eripära:
a. Homoloogilised polüteenkromosoomid paarduvad ka somaatilistes rakkudes
18
b. Polüteenkromosoomid on jälgitavad ka interfaasi rakkudes
26. Aneuploidsus ja selle fenotüübilised efektid. Tooge näide.
Aneuploidid:
Ise ndid, kellel on lisakromosoom – hüperploidid
Isendid, kellel teatav kromosoom puudub – hüpoploidid
Teatava kromosoomi kolmekordistumine - trisoomia
Aneuploidsus annab tugeva fenotüübilise efekti.
Aneuploidsete gameetide moodustumine:
Monosoomia tekkimine:
1) Viljastamisel (kromosoomide ebavõrdne jaotumine gameetidesse)
2) Viljastamise järgselt (kromosoomide ebavõrdne jaotumine viljastatud munaraku jagunemisel)
NÄIDE: Downi sündroom
Downi sündroomi põdevad 21 kromosoomi trisoomiaga indiviidid on vaimselt alaarenenud
Downi sündroomiga laste IQ jääb 25 ja 50 vahele
Juhul, kui nad tänu pidevale hoolitsusele elavad üle 30-nda eluaasta, tabab neid Alzheimer’i tõbi.
NÄIDE: Edwardi sündroom: 18. Kromosoomi trisoomia
Tugev vaimse arengu peetus
Ainult 10% elavad aastaseks
27. Muutused kromosoomide struktuuris: deletsioonid, duplikatsioonid, inversioonid ja
nende ümberkorralduste fenotüübiline efekt.
Deletsioon – lõigu kaotsiminek kromosoomist
Cri-du-chat sündroom on põhjustatud 5-nda kromosoomi lühikesest õlast toimunud deletsiooni poolt
Duplikatsioon – kromosoomilõigu kahekordistumine
X kromosoomi keskmise segmendi duplikatsiooni (mutatsioon bar) kandval äädikakärbsel on väiksemad
silmad. Veelgi tugevam efekt ilmneb triplikatsiooni toimel
Inversioon – segment kromosoomist on ülejäänud osa suhtes 180° ümber pööratud
Peritsentrilise inversiooni tagajärjel võivad muutuda kromosoomi õlgade pikkused, akrotsentrilisest
kromosoomist tekib metatsentriline
Paratsentrilise inversiooni puhul tsentromeeri ei kaasata
Inversioonid põhjustavad meioosihäireid, mille tagajärjeks võib olla steriilsus
28. Translokatsioonid ja liitkromosoomide teke.
Translokatsioonide mõju geenide avaldumisele.
Translokatsioon – segment kromosoomist satub temaga
mittehomoloogilisse kromosoomi.
Retsiprookne translokatsioon - kui kaks mittehomoloogilist
kromosoomi vahetavad võrdsel hulgal geneetilist materjali
19
Translokatsiooni või inversiooni tulemusena võib geen sattuda uude kohta, kus ta avaldub erinevalt.
Nt: Geen white on X kromosoomis toimunud inversiooni tagajärjel sattunud heterokromatiimi sisaldava
tsentromeeri lähedale. Tulemus: Kärbse silma pigment on ebaühtlaselt jaotunud
Nt: 21. kromosoomi pika õla translokatsioon 14-ndasse kromosoomi võib põhjustada Downi sündroomi,
kui sugurakkude ühinemisel satuvad samasse rakku 14 -21 translokatsiooniga kroomosoom ja kaks 21.-st
kromosoomi.
Meioosis võivad translokatsioone sisaldavad, kuid muus osas mitte-homoloogilised kromosoomid
omavahel paarduda, moodustades ristikujulisi struktuure, kus on 4 tsentromeeri. Homoloogiliste
kromosoomide lahknemine on häiritud ja võivad tekkida aneuploidsed gameedid.
LIITKROMOSOOMID – erineb translokatsioonist. Homoloogilise kromosoomide endi vahel toimuv
liitumine.
Evolutsioonis on kromosoomide liitumisi sageli toimunud. Inimese 2. kromosoom on metatsentriline,
tema õlad vastavad kahele erinevale akrotsentrilisele kromosoomile ahvidel.
29. Mis on geenide aheldatus? Tooge näide.
Aheldunud geenid – samas kromosoomis paiknevad geenid. Samas kromosoomis paiknevad geenid
peaksid päranduma koos, sest nad on aheldunud, jäädes kokku ka pärast meioosi.
Teatud juhtudel ei jää geenid aheldatuks: meioosis võib toimuda homoloogiliste kromosoomide
kromatiidiosade vahetus e. ristsiire (crossing over).
NÄIDE:
Bateon ja Punnett ristasid kahe tunnuse poolest erinevaid lillherneid: punased/valged õied,
piklikud/ümarad tolmuterad.
Punaseõieliste piklike tolmuteradega taimede ristamisel valgeõieliste ümarate tolmuteradega taimedega
raade F1 põlvkonnas vaid punaseõielised piklike tolmuteradega taimed: punane värvus ja tolmuterade
piklik kuju peavad olema dominantsed tunnused.
Hübriidide iseviljastumisel saadi nelja fenotüübiga järglasi, nagu dihübriidsel ristamisel võib eeldada, kuid
fenotüübiline suhe erines oodatavast (9:3:3:1) suhtest. Tegelik lahknemissuhe oli 24,3:1,1:1:7,1. Kui jätta
välja rekombinantsed järglased, saaksime lahknemissuhte punaseõieliste piklite tolmuteradega järglaste ja
valgeõieliste ümarate tolmuteradega järglaste vajel 3:1 - nagu monohübriidsel ristamisel. Siit saab
järeldada, et vaadeldavad geenid kanduvad enamasti koos sest nad on aheldunud ja rekombinantsed
järglased peavad moodustama geenide ristsiirde tulemusel.
F1 põlvkonna gameetide moodustumisel toimus meioosis ristsiire, mille tulemusena tekkisid
rekombinatsed kromosoomid, kus alleelid homoloogilistes kromosoomides olid vahetunud.
Geenide aheldumisele viitab rekombinantide (vanemtüübist erineva fenotüübiga järglaste) madal sagedus
F2 põlvkonnas
Kui geenid asuvad samas kromosoomis, siis päranduvad nad koos samasse gameeti.
30. Geneetilise materjali rekombineerumine ristsiirde teel.
Teatud juhtudel ei jää geenid aheldatuks: meioosis võib toimuda homoloogiliste kromosoomide
kromatiidiosade vahetus e. ristsiire (crossing over). Ristriirde protsessis osaleb homoloogilistest
kromosoomidest 1 tütarkromatiid.
20
Ristsiirde toimumine põhjustab kõrvalekaldeid Mendeli sõltumatu lahknemise seadusest. Ristsiirde
tulemusena moodustuvad vanemtüüpi ja rekombinantseid kromosoome sisaldavad sugurakud.
Ristriirde ehk homoloogilise rekombinatsiooni käigus joonduvad isa kromosoomi sünaptoneemi abil. See
kompleks lahustub profaasi lõpuks ja ristsiidre ühendused saavad mikroskoobis nähtavaks kui X-kujulised
struktuurid, mida nimetatakse kiasmideks.
Kiasm – spetsiaalset kromatiini struktuurid, mis esinevad kohtades, kus rekombinatsioon on lõppenud.
Kiasmid hoiavad homoloogilised kromosoomid koos kuni järgmise anafaasini meioos ühes.
31. Kas kõrge rekombinatsioonisagedus viitab uuritavate geenide üksteisele lähestikku või
eemal paiknemisele? Põhjendage.
Kui geenid asuvad samas kromosoomis suhteliselt lähestikku, siis ongeeid aheldunud ja päranduvad edasi
järglastele enamasti koos.
Rekombinatsioonisageduse arvutamiseks jagatakse rekombinantide arv kogu järglaskonna arvuga.
Kui
rekombinatsioonisagedus ületab väärtuse 0,5 (so. 50%), pole geenid aheldunud vaid paiknevad erinevates
kromosoomides.
Kahe geeni vaheline rekombinatsioonisagedus ei saa olla kunagi kõrgem kui 50%, sest kõrgema sageduse
pjuhul ei ole geenid enam aheldunud, nad lahknevad juba sõltumatult ja peavad seeda paiknema erinevates
kromosoomides, või sama kromosoomi kaugetes lookustes.
Keskmine ristsiirete arv kromosoomi kohta võimaldab mõõta geneetilist distantsi
Kahe punkti vaheline kaugus kromosoomi geneetilisel kaardil kujutab nende punktide vahel toimuvate
ristsiirete keskmist arvu
Mida kaugemale jäävad geenid teineteisest, seda suurem on rekombinatsiooni- sagedus. Ristsiirete
toimumise sagedust mõõdetakse rakupopulatsioonis!
Rekombinatsioonisagedus mõõdab aheldatuse tugevust, ahelduse muudumisel on
rekombinatsioonisagedus 50% ja täielikul aheldatusel 0%.
32. Millest on tingitud erinevused kromosoomide geneetilisel ja füüsilisel kaardil?
Geneetilised kaardid on koostatud lähtudes rekombinatsioonisagedustest:
1 centiMorgan (cM) vastab geenidevahelisele kaugusele, mille puhul geenide rekombinatsioon toimub
sagedusega 1%.
Füüsiliste kaartide (restriktsioonikaart, DNA järjestus) puhul mõõdetakse vahemaid molekulaarselt –
aluspaaride (bp), kilobaaside (kilobase, kb) või megabaaside (megabase, Mb) kaudu.
Kuigi geenide molekulaarne distants üksteisest ei pruugi alati korreleeruda nende geneetilise kaugusega,
vastab 1 cM inimese kromosoomis eukromatiini sisaldavas alas ligikaudu 1 Mb-le.
1Mb = 1000 kb = miljon
aluspaari
Kaugused geneetilisel kaardil ei vasta täpselt kaugustele kromosoomi füüsilisel kaardil.
Rekombineerumine toimub väiksema tõenäosusega kromosoomi otste lähedal ning tsentromeeri
piirkonnas – geneetilisel kaardil on need alad kokku surutud.
21
Ülejäänud regioonid, kus ristsiirde toimumise tõenäosus on kõrgem, on geneetilisel kaardil välja
venitatud. Geneetiline ja füüsiline kaart on kolineaarsed – geenid paiknevad mõlemal kaardil samas
järjekorras. Rekombinantide analüüs võimaldab määrata geenide järjekorda kromosoomis, kuid mitte
nendevahelisi füüsilisi kaugusi
33. Pagaripärmi Saccharomyces cerevisiae elutsükkel. S. cerevisiae kasutamine ristsiirete
uurimisel.
Pagaripärmi elutsükkel:
Üherakuline haploidne organism paljuneb
pungumise teel
Sugulisel paljunemisel liituvad 2 erineva
ristumistüübiga rakku
Diploidne rakk läbib meioosi, mille
tulemusena tekib 4 haploidset askospoori, mis
jäävad kokku askusesse
Iga askus sisaldab ühe konkreetse meioosi
produkte! – suurepärane võimalus.
Saccharomyces cerevisiae
ristriisre uurimisel
Ühe ristsiirde tagajärjel on askuses 2 spoori vanemtüüpi ja 2 rekombinantsed, sest ristsiirdel
homoloogiliste kromosoomide kromatiidide vahel moodustub 2 rekombinantset ja kaks algset
kromosoomi.
Ühe rekombinatsioonisündmuse tulemusena sisaldub pagaripärmi askuses kaks rekombinantset ja kaks
mitterekombinantset askospoori. Järelikult toimub ristsiire pärast seda, kui homoloogilised kromosoomid
on duplitseerunud.
Homoloogiliste kromosoomide tütarkromatiidide vahel võib toimuda mitu ristsiiret, kuigi ühes kindlas
kohas osalevad korraga 2 kromatiidi.
Ristsiirde toimumist ei tuvastata, kui see toimub geneetiliselt identsete tütarkromatiidide vahel
Ristsiirde toimumise ajal koosneb kromosoom kahest tütarkromatiidist
Ristsiire toimub kahe homoloogilise kromosoomi kromatiidide vahel
Meioos II lõpuks satuvad 4 kromatiidi eraldi rakkudesse
34. Mida näitavad homoloogiliste kromosoomide vahelised kiasmid?
Kiasmid märgistavad ristsiirde toimumiskohta.
Kiasm – esimese meiootilise jagunemise profaasi diploneemastaadiumis kahe homoloogilise kromosoomi
neljast kromatiidist koosnevas grupis (bivalendis) nähtav kahe kromosoomi vaheline DNA lõikude
vahetuskoht, kus toimub ristsiire.
Iga homoloogiliste kromosoomide vahel jälgitav kiasm meioosi profaasis kajastab üht profaasi algusosas
toimunud ristsiiret. Kiasmide loendamine võimaldab määrata keskmist ristsiirete arvu kromosoomi kohta.
22
Kui kiasmide arv raku kohta on 2,15, siis kromatiidi kohta on see poole väiksem – 1,07. Sellest leiame, et
kromosoomi pikkus on 107 cM, sest kiasmide arv kromatiidi kohta väljendab kromosoomi geneetilist
pikkust.
Meioosi esimese profaasi lõpus on kromosoomid üksteisega kontaktis vaid kiasmide ja tsentromeeri
kaudu, mis võimaldab määrata kiasmide arvu. Kiasmide arv on proportsioonis kromosoomi pikkusega:
mida pikem kromosoom seda rohkem kiasme.
35. Millal toimub ristsiire? Rekombinatsiooni osa evolutsiooniprotsessis.
Ristsiire toimub meioosi esimeses profaasis. Profaasi varajase ja keskosa etappides (sügoteen – diploteen).
Rekombinatsooniprotsessi evolutsiooniline kasulikkus ilmneb näiteks mittesuguliselt ja suguliselt sigivate
organismide võrdluses. Oletame, et mõlematel toimub mingi kasulik mutatsioon ning aja jooksul ka teine.
Suguliselt sigvatel organismidel võivad need mutatsioonid sattuda samasse organismi ning meioosis
sugurakkude moodustumisel omavahel rekombineeruda. Võivad moodustuda üksikmutantidest edukamad
järglased, kelle enda järglased võivad saada mõne aja pärast populatsioonis ülekaalu. Mittesugguliselt
sigivatel organismidel kasulike mutatsioonide rekombineerumist toimuda ei saa, see tähendab, et eri
indiviididel tekkinud mutatsioonid ühte organismi rekombineeruda ei saa. Seega, evolutsioonilises
tähenduses võimaldab rekombinatsiooniprotsess viia erinevate geenide kasulikud alleelid kokku samasse
organismi.
Samas on aga evolutsiooniprotsessis välja kujunenud olukord, kus erinevatel organismidel toimuv
rekombinatsioonide sagedus on äärmiselt erinev. Juba väljakujunenud liigi geneetilise arenguprogrammi
säilitamise huvides pole kõrgsageduslik rekombinatsioon liigile kasulik. Äärmuslik näide on
äädikakärbeste isased, kel ristriiret üldse ei toimu.
36. Geenide kaardistamine Neurospora crassa askuste analüüsil.
Neurospora grassa – leivahallitus
Erinevalt pärmist on Neurospora’l askospooride kott piklik ja väga kitsas, nii et spooride reastumine
askuses kajastab seda, kuidas reastusid kromatiidid meioosis. Meioosi käigus rakud ei pooldu, tuumad
jäävad kõrvuti ning pärast meioosi toimub veel üks mitootiline jagunemine, nii et iga haploidne tuum
jaguneb veel omakorda. Lõpptulemuseks on kaheksa reas paiknevat tuuma, mis eraldatakse üksteisest
rakuseintega, nii et moodustuvad askospoorid.
Askospoorid askustes jäävad kindlasse ritta, kajastades olukorda, kuidas külgnesid meioosis üksteisega
neli kromatiidi. N. grassa askuste analüüs näitab, kas ristsiire uuritavate punktide (geen ja tsentromeeri
piirkond) vahel on toimunud.
Geeni ja tsentromeeri vahelise distantsi arvutamine erinevate N. grassa mutantide puhul võimaldab
uuritavad geenid reastada kromosoomi geneetilisel kaardil
37. Inversioone sisaldavate kromosoomide kasutamine geneetilises analüüsis.
23
Tasakaalustavate (inversioone sisaldavate) kromosoomide kasutamine äädikakärbse geneetilises analüüsis:
Rekombinatsioon inversiooni sisaldava kromosoomiga põhjustab aneuploidsust, järglased ei ole
eluvõimelised. Uuritav mutantne alleel on retsessiivne ja saab avalduda ainult homosügootses olekus, mis
on ristsiirde tagajärg. Seega ei saa mutantne retsessiivne alleel koos avalduda dominantse alleeliga, mis
asub paigalhoidvas kromosoomis, sest selle kromosoomiga rekombinante ei saada
Kuna mutantne fenotüüp avaldus F2 järglaskonnas koos mõlema tasakaalustava kromosoomi dominantse
alleeliga, paiknes mutatsioon hoopis 4-ndas kromosoomis
38. Geenide kaardistamise meetodid, mis põhinevad somaatiliste rakkude
hübridiseerimisel.
Somaatiliste rakkude hübridiseerimist saab kasutada inimese geenide kaardistamisel tingimusel, et uuritav
geen avaldub hübriidses rakus ja tema funktsioon on eristatav hiire omast.
Inimese ja hiire rakkude liitumist stimuleeritakse Sendai viirusega. Hübriidsete rakkude jagunemisel
lähevad inimese kromosoomid järk-järgult kaotsi.
24
Hübriidseid rakke selekteeritakse HAT (hüpoksantiin-aminopteriin-tümidiin) söötmel, kus aminopteriin
blokeerib põhilise nukleotiidide sünteesiraja
39. Geenide kaardistamine deletsioone ja duplikatsioone sisaldavate kromosoomide abil.
KAARDISTAMINE DELETSIOONIGA
Tehnika, mis võimaldab kindlaks määrata geeni asukohta kromosoomis.
Tsütoloogiliselt määratavat deletsiooni tähistatakse tavaliselt tähekombinatsiooniga Df. Näiteks
Drosophila X-liiteline geen, mille:
mutantne alleel w määrab valgesilmsuse
w+ määrab punasilmsuse tekke.
Järelikult on w mitteproduktiivne ehk nullmutatsioon.
Kui emastel kärbestel avaldub ühe X-kromosoomi retsessiivne alleel w heterosügootses olekus, siis peab
neil emastel teises X-kromosoomis olema samas piirkonnas deletsioon. Seetõttu võimaldavad w/Df-
heterosügoodid teha funktsionaalset testi, lokaliseerida geen w deletsiooni suhtes.
Nimelt on w/Df-heterosügoodid valgesilmsed, sest neil puudub w+-geeni funktsionaalne koopia mõlemas
X-kromosoomis. Sama deletsiooniga x-kromosoom koos metsiktüüpi x-kromosoomiga (w+) annab aga
punasesilmsed äädikakärbsed, sest dominantne alleel määrab pigmendi produktsiooni. Kui deletsioon ei
kata uuritavad mutantset alleeli w, siis on kõik järglased punasilmsed.
X-kromosoomi deletsioonid, mis ei kattu mutantse geeniga, ei võimalda avaldada uuritava mutantse alleeli
w mutantset fenotüüpi.
25
KAARDISTAMINE DUPLIKATSIOONIDEGA
Deletsioonikaardistamisele vastupidine meetod. Duplitaktsioonid, mis kattuvad retsessiivse mutantse
geeni alaga ning mis kürvaldavad mutantse fenotüübi tekke, peavad sisaldama mutantse geeni metsiktüüpi
alleeli.
Näites 1 duplikatsioonidest maskeerib ehk katab mutatsiooni w ja annab metsiktüüpi punasilmse
fenotüübi. Polüteensel x-kromosoomil lokaliseeriti geen w piirkondade 2D ja 3D vahele, ehk piirkonda
mis külgneb alaga C. Järelikult täpsustus duplikatsioonianalüüsil geeni w deletsioonanalüüsil määratud
lokalisatsioon.
26
Duplikatsiooni all on mõeldud segmenti X kromosoomist, mis on translokeerunud mõnda teise
kromosoomi. Uuritakse translokatsiooniga kromosoomi efekti white mutatsiooni avaldumisele
40. Geenide kaardistamine translokatsioone sisaldavate kromosoomide abil.
HPRT geeni täpsem lokaliseerimine X kromosoomi pikka õlga 14-X translokatsiooni abil. Translokatsioon
on toimunud x-kromosoomi ja 14. kromosoomi vahel: enamik x-kromosoomi pikast õlast on
translotseerunud 14.kromosoomi otsa ning väike segment 14.kromosoomi otsast on translotseerunud x-
kromosoomi otsa. Seda translokatsiooni sisaldavaid inimese rakke hübriiditi hiire rakkudega.
Peale hübriidrakkude selektsiooni HAT-söötmel saadi rakuliin, mis sisaldas vaid ühte inimese kromosoomi
– 14. kromosoomi kuhu oli liitunud enamus x-kromosoomi pikast õlast. Kui eelnevalt oli teada et HPRT
geen paikneb x-kromosoomis, siis käesolevad katsed näitasid, et ta asub x-kromosoomi pikas õlas.
Samasse õlga paigutusid ka fosfoglütseraadi kinaasi PGK ja glükoos-6-fosfaadi dehüdrogenaasi G6PD
geenid.
27
41. Millist tüüpi nukleiinhape võib olla päriliku informatsiooni kandjaks?
Nukleiinhapped jagunevad: DNA ja RNA. Mõlemad võivad olla geneetilise info kandjaks, kuid enamuse
organismide puhul on selleks 2-ahelaline DNA. Seda sellepärast et DNA on stabiilsem.
RNA on genoomiks osadel viirustel (nt HIV on retroviirus, kelle genoomiks on ssRNA, mille
pöördtranskriptaas sünteesib DNA)
42. Eksperimentaalsed tõendid selle kohta, et DNA kannab geneetilist informatsiooni.
1928 näitas Griffith bakteri
S.Pneumoniae transformatsiooni – DNA kandumist
väliskeskkonnast bakterirakku.
IIR tüüpi kapslita rakud – ei ole virulentsed. Väikesed karedapinnalised kolooniad.
IIIS tüüpi limakapsliga rakud – on virulentsed. Suured siledapinnalised kolooniad. Limakapsel takistab
peremeesorganismil bakterit hävitamast.
Griffith näitas, et kõrge temperatuuriga omandasid kapslita IIR rakud surmatud limakapsliga IIIS
rakukultuurist midagi, mis muutis mittevirulentsed elusad IIR tüüpi rakud virulentseteks. See tähendab, et
ta transformeeris IIR rakud patogeenseteks limakapsliga rakkudeks IIIS(kandis IIIS DNA
väliskeskkonnast IIR bakterirakku).
1944 näitasid Avery, MacLeod ja McCarty, et S.Pneumoniae mittevirulentsete IIR tüüpi rakkude
transformeerimist IIIS tüüpi virulentseteks rakkudeks põhjustas DNA.
Nad näitasid, et desoksüribonukleaas (Dnaas), mis lagundas DNA, kõrvaldas ka transformatsioonivõime.
Ribonukleaas (lagundab RNAd) ja proteaas (lagundab valke) transformatsioonivõimet ei mõjutanud.
1952 Hershey ja Chase tõestasid, et bakteriofaag T2 geneetiline info on talletunud DNA-s.
Eelnevalt oli näidatud, et T2 faagi elutsükkel tomub ainult E.Coli rakkudes. Järelikult sõltub faagi
paljunemine totaalselt E.Coli raku metaboolsest masinavärgist. Hersley ja Chase näitasid, et kui viirus
nakatab bakterirakku, siis viiruse valguline kest jääb raku pinnale ja rakku tungib vaid DNA. DNA
põhjustas uute viirusosakeske moodustumise rakud.
28 Griffith
44 Avery, MacLeod, McCarty
52 Hershey, Chase
43. Võrrelge DNA ja RNA koostist ning ehitust.
1. Erinevus 5-süsinikulise suhkru ehituses: DNAl on Desoksüriboos ja RNAl Riboos. Erinevus
seisneb selles, et DNAl puudub 3’C juures OH rühm.
2. DNA on võrreldes RNA-ga tunduvalt stabiilsem. Seda tänu OH rühma puudumisele.
3. RNA koostises on Tümiini asemel Uratsiil
4. RNA on tavaliselt üksikahelaline, DNA aga kaheahelaline molekul.
44. Selgitage DNA ahelate komplementaarsuse ja antiparalleelsuse põhimõtet.
DNA on paremalepöörduv 2-ahelaline heeliks.
KOMPLEMENTAARSUS: Tuleneb N-aluste spetsiifilisest paardumisest. Kui me teame DNA ühe ahela
nukleotiidset järjestust, saame üheselt määratleda teise ahela nukleotiidse järjestuse. Just DNA-ahelast
komplementaarsus võimaldab säilitada geneetilist infot muutumatult põlvkonnast põlvkonda.
ANTIPARALLEELSUS: DNA komplementaarsed ahelad on vastassuunalised, ehk komplementaarsed.
Ühe ahela nukleotiidide vahelised fosfodiestersidemed on nende suhkru süsinikuaatomite vahel suunaga
5’- 3’, teises ahelas aga 3’-5’. See tähendab, et DNA üksikahela ühes otsas on vaba 3’OH rühm (kust
toimub kasvamine), teises otsas 5’P-rühm. Suhkru 3´C küljes olev OH-rühm ühineb lisanduva nukleotiidi
suhkru 5´C-ga seotud fosfaadi vesiniku aatomiga; eraldub vesi H2O ja vabaneb pürofosfaat
28
45. Bakterikromosoomi struktuur.
Erinevate organismide genoomid on erineva suurusega. Sellest tulenevalt on ka erinev genoomi
kokkupakituse aste.
Bakterikromosoomiks on kaksikahelaline DNA. Bakterite geneetiline informatsioon asub ühes
rõngaskromosoomis, mis esineb rakus tuumapiirkonnas, mida nimetatakse nukleoidiks (tuumamembraan
puudub).
E.Coli kromosoomi kontuurpikkuseks on ca 1500 mikromeetrit, mis sisaldab 2500-3000 geeni. E.Coli
enda raku diameeter on aga 1-2 mikromeetrit. Seega peab olema bakteri kromosoom tugevalt
kokkukeerdunud ehk kondenseerunud. Kromosoom on 1000 korda lühemaks kokku pakitud, muudustub
400 lingu.
Iga ling on sõltumatult negatiivselt superspiraliseerunud. Kokkuvolditud kromosoom on kompleksis RNA
ja valkudega.
Üks DNA ahel on kaksikheeliksis roteerunud fikseeritud ahela suhtes kas parema- või vasakusuunaliselt.
Negatiivne superspiralisatsioon – vaba ahela vastassuunaline, fikseeritud ahela suhtes vasakule
roteerumine – DNA ahelad rohkem lahti keeratud (relaxed) ja võivad eralduda
46. Eukarüootsete kromosoomide koostis ja struktuur.
Eukarüootne kromosoom on bakteri DNAga võrreldes kompaktsem. Kromosoomi moodustab 1 DNA
molekul.
Eukarüootsete kromosoomide kondensatsioonil eristatakse 3 taset:
1.
DNA nukleosoomne struktuur
DNA on keritud ümber histoonidest moodustunud oktameeride. Oktameeri moodustab histoonivalkude
kogum, kus histoone H2a, H2b, H3 ja H4 on kahes korduses.
Nukleosoom sisaldab 146 aluspaari DNAd.
Nukleosoomid on omavahel ühendatud
linkeralade kaudu. Histoon H1 on seotud linkeralaga ja
see stabiliseerib struktuuri ning osaleb kromatiinikiudude moodustamisel.
29
DNA on nukleosoomis
negatiivselt superspiraliseerunud ja keritud 1,75 ringi ümber histoonse
oktameeri.
Nukleosoomi diameeter on 11nm.
2.
Kromatiinkiud
Kromatiin – interfaasi raku kromosoomimaterjal, mis koosneb valdavalt DNAst ja valkudest, väiksemas
hulgas ka RNAst.
Modifitseeritud histoonide tõttu moodustub kromatiini kokkupakkimise järgmine tase – kromatiinkiud.
Kromatiinkiu diameeter on 30nm.
Kromatiinkiu struktuur moodustub nukleosoomsest struktuurist H1 histoonide osalusel.
3.
Kromosoomi tase
Järgmisele tasemele pakivad kromatiini
mittehistoonsed valgud, mis moodustavad
kromosoomi toese,
ning sellele kondenseerub Kromatiinkiud. Sellega eralduvad ja eristuvad eukatüootses kromosoomis
superspiraliseerunud domeenid ehk lingud. Iga ling sisaldab 30 000 – 100 000 aluspaari.
47. Mis funktsioon on eukarüootsetes kromosoomides tsentromeeridel ja telomeeridel?
TELOMEERID – kromosoomiotstes asuvad kordusjärjestused.
1. Takistavad DNA molekulide otste lagundamist nukleaaside poolt – kaitsevad lineaarse DNA
lõppe
2. Takistavad erinevate DNA molekulide otste kleepumist – takistavad DNA otste ühinemist
3. Võimaldavad lineaarsete DNA molekulide otste replitseerumist, ilma et DNA molekulid
kaotaksid otstest geneetilist materjali – Võimaldavad replikatsiooni, ilma, et toimuks DNA kadu.
TSENTROMEERID - Tsentromeeri piirkonnas on DNA eriti tugevalt kondenseerunud
Tsentromeeridele kinnituvad mikrotuubulitest koosnevad kääviniidid, mis võimaldavad tütarkromatiididel
anafaasis liikuda raku vastaspoolustele.
48. Nukleiinhapete sünteesi suund ja nukleiinhapete sünteesi läbiviivad ensüümid.
DNA ahel kasvab 5´→ 3´suunas. Sünteesitavasse DNA ahelasse lülituvad nukleotiidid, mille
lämmastikalused on komplementaarsed matriitsahela nukleotiidide lämmastikalustega. DNA
replikatsioonil käituvad matriitsina mõlemad DNA ahelad; saadakse kaksikheeliksid, milles üks ahel on
uus ja teine vana
DNA polümeraas – sünteesib DNA ahelale komplementaarse ahela. vajab sünteesil praimerit.
RNA polümeraas – sünteesib DNA ahelale komplementaarse RNA ahela; transkriptsiooni initsiatsiooniks
seondub RNApolümeraas spetsiifiliselt promootorjärjestusega.
Pöördtranskriptaas e. revertaas – sünteesib RNA ahelale komplementaarse DNA ahela; vajab sünteesil
praimerit.
49. DNA replikatsiooni kolm mudelit – konservatiivne, dispersiivne ja semikonservatiivne.
Milline neist mudelitest leidis eksperimentaalselt kinnitust?
SEMIKONSERVATIIVNE MUDEL: DNA ahel kasvab 5´→ 3´suunas . Sünteesitavasse DNA ahelasse
lülituvad nukleotiidid, mille lämmastikalused on komplementaarsed matriitsahela nukleotiidide
lämmastikalustega. DNA replikatsioonil käituvad matriitsina mõlemad DNA ahelad; saadakse
kaksikheeliksid, milles üks ahel on uus ja teine vana.
Eksperimentaalselt leidis kinnitus semikonservatiivne mudel.
KONSERVATIIVNE MUDEL: algselt kaksikheeliksilt sünteesitakse uus; ühes DNA molekulis on koos
vanad ja teises uued ahelad
30
DISPERSIIVNE MUDEL: mõlemas DNA molekulis sisaldavad DNA ahelad segu vanadest ja uuesti
sünteesitud lõikudest
50. DNA replikatsiooni initsiatsiooni mehhanism.
DNA replikatsioon algab
oriC järjestustelt, kus esmalt moodustub DNA ahelate lokaalsel lahkuminekul
replikatsioonikahvel. Toimub DNA ahelate lahtikeerdumine ja praimeri süntees.
DNA replikatsioonikahvli avamine toimub A-T rikastest regioonidest initsiaatorvalgu
DnaA toimel.
Järgnevalt toimub DnaA-valkude
kooperatiivne liitumine, moodustub 20-40 polüpeptiidist koosnev
valguline kompleks, mille ümber seondub
oriC piirkond.
ATP juuresolekul toimub avatud kompleksi moodustumine. Ahelate lahknemine algab kolme tandeemse
13bp oriC kordusjärjestuste piirkonnas ja levib seni, kuni on avanenud kogu 13bp korduste piirkond.
DnaB helikaas ja
DnaC moodustavad denatureerunud DNA-ga prepraimingkompleksi. SSB ja güraasi
juuresolekul jätkub DNA ahelate lahtikeerdumine mõlemas suunas.
Järgneb praimeri süntees primaasi
DnaG poolt
DNA polümeraas III moodustab replikatsioonikahvli
51. Erinevate DNA polümeraaside funktsioonid bakterites. Mis mehhanismidega on tagatud
DNA replikatsiooni täpsus?
Bakteritel on lisaks DNA polümeraasidele I ja III veel vähemalt 3 DNA polümeraasi:
Pol I ja Pol II: DNA reparatsioonilised polümeraasid.
Lisaks 5'
3' polümeraassele aktiivsusele on Pol I ka 5' 3' ja 3´ 5' eksonukleaasne aktiivsus
DNA Pol I osaleb DNA sünteesil mahajäävalt ahelalt, kus see toimub Okazaki fragmentidena,
asendades seal RNA praimerid DNA-ga
DNA güraas (topoisomeraas II) on tetrameerne valk, mis koosneb kahest subühikust. DNA güraasi
on vaja E. coli DNA replikatsioonil vähendamaks replikatsioonikahvli ees tekkivat positiivset
superspiralisatsiooni
Pol III: põhiline DNA replikatsiooni läbiviija.
Polümeraasi õlad sünteesivad erinevaid ahelaid: vasak õlg – juhtivat. Parem õlg – Mahajääva ahela
Okazaki fragmente.
Pol IV ja Pol V: DNA reparatsioonilised polümeraasid. vigaderohke DNA süntees, kui Pol III töö on
blokeeritud DNA kahjustuse tõttu
.
Bakteris on põhiliseks DNA replikatsiooni läbiviivaks valguks DNA polümeraas Pol III.
DNA replikatsiooni täpsuse eest vastutab Pol III subühik epsilon. Epsilonil on 3’- 5’ suunaline
endonukleaasne aktiivsus, mis võimaldab valesti DNA ahelasse lülitatud nukleotiide kõrvaldada.
31
52. DNA replikatsioon juhtivalt ja mahajäävalt ahelalt.
DNA ahelate komplementaarsuse tõttu on DNA ahelad vastassuunalised. Replikatsiooni alguspunktis
moodustub 2 replikatsioonikahvlit, mille alusel toimub edasi mõlemasuunaline DNA replikatsioon.
DNA ahelad on antiparalleelsed. Teise ahela puhul, mis pikeneb 3’
5’ suunas, toimub tegelikult samuti
5'
3' suunaline süntees, kuid katkendlikult, lühikeste fragmentidena, mida nimetatakse Okazaki
fragmentideks.
Juhtiva ahela süntees saab olla pidev, teise ahela süntees on aga katkendlik. Okazaki fragmendid seotakse
üksteisega DNA ligaasi toimel.
DNA polümeraas on võimeline ainult olemasolevat nukleiinhappe ahelat pikendama, vajades seetõttu
sünteesi alustamiseks praimerit.
DNA liiderahela sünteesil on vaba 3’OH otsaga praimerit vaja vaid üks kord, sünteesi alustamiseks. Iga
viivisahela lõigu sünteesiks on vaja praimerit. Okazaki fragmentide sünteesi initsiatsiooniks on vaja
valkkompleksi, mida nimetatakse praimosoomiks (DNA helikaas + primaas). DNA praimeriks on
lühikesed RNA fragmendid, mille sünteesivad mahajääval ahelal
DNA praimaasid.
RNA praimeritelt jätkab sünteesi DNA polümeraas III.
DNA polümeraas I eemaldab RNA praimerid DNA-ahelast ja asendab need DNA lõikudega.
Ensüüm
DNA ligaas katalüüsib üksikahelaliste DNA katkete sulgumist.
DNA helikaas keerab DNA ahelaid lahti, kasutades ATP energiat
DNA topoisomeraas teeb DNA ahelatesse ajutisi katkeid, et soodustada DNA ahelate lahtikeeramist.
Üksikahelalist DNA-d stabiliseerivad sellele seonduvad SSB (single strand binding protein) valgud.
53. Võrrelge bakteri ja eukarüoodi kromosoomide replikatsiooni.
Eukrüoodil:
1. DNA süntees toimub ainult rakutsükli ühel etapil ja algab paljudest kohtadest korraga
2. Juhtiva ja mahajääva ahela sünteesiks on 2 erinevat DNA polümeraasi
3. Eukarüootne DNA on koos histoonidega nukleosoomideks organiseeritud: replikatsioonikahvli
läbiminekul jaotub nukleosoom ajutiselt kaheks alaosaks
4. Kromosoomid on lineaarsed DNA molekulid ja nende otstest lühenemist kaitsevad telomeerid.
Pärast RNA praimeri kõrvaldamist jääb mahajääv DNA ahel otsast lühemaks kui juhtiv ahel.
Telomeraas sisaldab RNA-d, mis on matriitsiks telomeeride pikendamisel telomeeri 3´
üksikahelalisest otsast. Kui telomeraas on telomeeri otsa piisavalt pikendanud, sünteesib DNA
polümeraas pikendatud ahelale komplementaarse ahela
54. DNA replikatsiooni veereva ratta mudel. Milliste DNA molekulide replikatsiooni puhul
seda on kirjeldatud? (õpik joonis 255)
Veereva ratta replikatsioon leiab aset rõngjas DNA-molekuli replikatsioonil.
DNA veereva ratta replikatsioon toimub järgmistel juhtudel:
1. Paljude
viiruste genoomi replikatsioon
2. Geneetilise informatsiooni ülekanne rakust rakku
bakterite konjugatsioonil
3. Amfiibide oogeneesis ribosoomi-RNA geenide klastreid kandva
kromosoomivälise DNA
amplifikatsioonil.
32
Üks algse DNA ahelatest jääb rõngaks ja on matriitsiks sünteesitavale komplementaarsele DNA ahelale.
Järjestuse-spetsiifiline nukleaas tekitab replikatsiooni alguspunktis ühte DNA ahelasse katke. DNA ahela
pikenemine algab vabast 3'-OH otsast ning 5'-fosfaadiga lõppev ahela ots eemaldub rõngast DNA sünteesi
käigus (See ots nagu "veereks" rõngalt maha). Teine ahel sünteesitakse Okazaki fragmentide abil
55. Molekulaarbioloogia põhidogma.
Molekulaarbioloogia põhidogma seisneb geneetilise info edastamisel.
Geneetiline info on salvestatud DNA nukleotiidses järjestuses. Geenide ekspressioon ehk avaldumine
realiseerub informatsiooni edastamise teel DNA nukleotiidselt järjestuselt aminohappelisse järjestusse:
1. Esmalt kandub info DNA-lt mRNA-le. Toimub transkriptsioon
See toimub rakutuumas (nukleoidis)
2. Edasi toimub mRNA nukleotiidses järjestuses salvestatud info põhjal tsütoplasma ribosoomidel
valkude süntees. Toimub translatsioon.
mRNA transporditakse tuumast välja tsütoplasmasse kus toimub tsütoplasmas ribosoomidel
valkude süntees.
DNA – RNA – valk.
RNA järjestuse põhjalt on võimalik sünteesida uuesti DNA – pöördtranskriptsioon
Valgu järjestuselt RNAd sünteesida ei ole võimalik!
Geneetiline info säilib, kandudes põlvkonnast põlvkonda edasi nukleiinhappelt nukleiinhappele.
Geneetiline info kandub edasi organismi geenide avaldumisel DNA-lt valkudesse.
56. RNA tüübid: tRNA, rRNA, mRNA, snRNA ja nende funktsioonid.
33
1.
mRNA – RNA molekulid, millelt toimub translatsioon. Messenger RNA
2.
tRNA – Osalevad polüpeptiidahela ehk valgu sünteesil (translatsioonil) mRNAs oleva
nukleotiidse järjestuse põhjal, kannavad aminohapped ribosoomi. Transfer RNA.
3.
rRNA - Ribosomaalsed RNA molekulid kuuluvad ribosoomide koostisesse. Ribosomaalse RNA
geenide transkriptsioonitase on rakkudes kõrge
4.
snRNA - Väikesed tuuma RNA-d osalevad intronite splaissingul. Small nuclear RNA
57. Mille poolest erineb transkriptsioni initsiatsioon replikatsiooni initsiatsioonist?
Transkriptsiooni initsiatsiooni võib omakorda jaotada kolmeks etapiks:
(1) RNA polümeraasi holoensüümi seondumine promootorile;
(2) DNA ahelate lokaalne lahtisulamine transkriptsiooni alguspunkti sisaldava ala piirkonnas, et
matriitsahel saaks paarduda RNA ahelasse lülitatavate ribonukleotiididega;
(3) Algab RNA ahela süntees, kus kasvavasse RNA ahelasse lülitatakse kuni 10 nukleotiidi, ilma et
RNA polümeraas DNA ahelal edasi liiguks (abortiivne transkriptsioon). RNA polümeraas on
DNA-l võimeline edasi liikuma alles siis, kui sigma faktor vabaneb. Siis algab RNA ahela
elongatsioon.
58. Võrrelge prokarüootset ja eukarüootset transkriptsiooni initsiatsiooni.
1. Eukarüoodil: Enne, kui eukarüootne RNA polümeraas seondub promootoralale ja algatab
transkriptsiooni, peavad sinna seonduma
basaalsed transkriptsioonifaktorid.
2. Nii nagu bakterirakus, on ka eukarüoodirakus transkriptsiooni initsiatsiooniks vajalik DNA
ahelate lokaalne teineteisest eemaldumine.
59. Transkriptsiooni elongatsioon ja terminatsioon.
ELONGATSIOON
RNA ahela elongatsiooni katalüüsib
RNA polümeraasi apoensüüm. Elongatsiooni käigus on DNA
ahelad transkriptsiooni toimumise kohas teineteisest 18 aluspaari ulatuses eraldunud.
RNA ahela süntees toimub kiirusega 40 nukleotiidi sekundis.
Sünteesitav RNA ahel eraldub DNA ahelast, vahetult transkriptsiooni toimumise kohas on DNA ja RNA
aga 12 nukleotiidi ulatuses paardunud.
Transkriptsioonikompleksi stabiilsuse määrab eeskätt ära siiski mitte see paardumine, vaid DNA ja
kasvava RNA ahela seondumine RNA polümeraasiga.
TERMINATSIOON
RNA ahela süntees lõpeb siis, kui RNA polümeraas kohtab terminatsioonisignaali. Seejärel
transkriptsioonikompleks dissotseerub.
Transkriptsiooni termineerivad signaalid on kas Rho valgust (
) sõltuvad või sõltumatud.
Rho-sõltumatud terminaatorid sisaldavad G:C-rikast regiooni, millele järgneb 6 või enam A:T paari (A
nukleotiidid asuvad matriitsahelas
Rho valgust sõltuva transkriptsiooni korral on terminatsioonijärjestused (samuti G:C-rikkad) pikemad.
60. Võrrelge eukarüootsete ja prokarüootsete geenide struktuuri. Eukarüootse RNA
transkriptsioonijärgne modifikatsioon.
Eukarüootidel enamasti paiknevad kodeerivate alade – eksonid vahel mittekodeerivad alad – intronid.
Intronite sisaldus ja pikkus varieerub geeniti
Algsest RNA-st lõigatakse tuumas teatud segmendid välja ja seejärel viiakse lõikusjärgne RNA
tsütoplasmasse.
34
Eukarüootide puhul ei ole transkriptsiooni käigus sünteesitud mRNA kohe lõplikult valmis, vaid vajab
mitmeid modifikatsioone:
1. CAP struktuur 5’ otsas.
Kaitseb mRNAd 5’otsa nukleaaside (lagundamise) eest
On seondumiskohaks valkudele
2. PolyA saba 3’ otsas.
Sinna seonduvad valgud mid aitavad vältida otste ladungamist ja osalevad mRNA transpordil
tsütoplasmasse.
3. RNA splaissing. Pre-mRNA sisaldab nii introneid kui eksoneid. Intronid eemaldatakse.
Splaissingus osaleb üle 200 erineva valgu, mis moodustavad splaissoomi. mRNA splaissimine on
mitmeetapiline protsess.
Splaissoomid: tuvastavad eksonite piirid, katalüüsivad introni välja lõikamist.
61. Intronite kõrvaldamine splaissingu teel.
Selleks, et splaissitud mRNA kodeeriks funktsionaalset valku, peab splaissingu protsess toimuma väga
täpselt.
Intronite väljalõikamine võib toimuda kolme erineva mehhanismi alusel:
1. tRNA prekursorite puhul teeb katked RNA ahelasse spetsiifiline
splaissingu endonukleaas ning
eksoneid sisaldavad RNA segmendid ühendatakse
splaissingu ligaasi abil.
2. Osade rRNA prekursorite puhul (paljudes madalamates eukaüootides, samuti ka rakuorganellides)
kõrvaldatakse intronid
autokatalüütiliselt, RNA molekuli enda poolt. Splaissingureaktsioon ei vaja
välist energiaallikat.
Kofaktorina on vaja vaba 3´-OH rühma nt GTP-lt ning monovalentset ja divalentset katiooni.
Splaissing toimub etapiviisiliselt: kõigepealt toimub fosfodiestersideme ülekanne ekson-intron
ühendusalalt G-OH-le (RNA ahela katkeb eksoni ja introni ühendusalas), seejärel katkeb
fosfodiesterside järgmise eksoni ja introni 3´-otsa vahel ning fosfodiesterside moodustub eksonite
vahel. Väljälõigatud intron tsirkulariseerub molekulisiseselt (toimub veel üks fosfodiestersideme
ülekanne).
3. Rakutuumas asuvate pre-mRNA molekulide splaissing toimub kahe-etapiliselt
splaissosoomides.
snRNA-d ei ole tumas vabalt, vaid kuuluvad väikestesse RNA-valk kompleksidesse, mida nimetatakse
snRNP.
62. mRNA molekulis asuva geneetilise informatsiooni muutmine – RNA editing.
Geneetilist informatsiooni RNA molekulis võidakse muuta kahel viisil:
1. lämmastikaluste asendamise teel
Lämmastikaluste asendamist on täheldatud peamiselt taimede mitokondrites ja valdavaks muutuseks on C
asendamine U-ga.
2. lisades või deleteerides U nukleotiide.
Nukleotiidide lisamine või deleteerimine on võrreldes lämmastikaluste asendamisega komplekssem protsess. mRNA
molekuli koostisesse lisatakse uridiinmonofosfaate. Nukleotiidide sisestamiskohti sisaldava mRNA piirkonnaga
paardub mRNA vastava kohaga osaliselt homoloogiline RNA molekul, mida nimetatakse
giid-RNA-ks (ingl. k.
guide RNA). Giid-RNA-s asuvad mRNA-ga mittepaarduvates alades A nukleotiidid ja just nende vastu lisatakse
mRNA-s uridiinmonofosfaadid.
63. Transkriptsiooni ja translatsiooni toimumise aeg ja koht bakterites ja eukarüootides.
Transkriptsioon ja valgusüntees (translatsioon) toimuvad bakterirakus korraga, eukarüoodis aga eraldi
etappidena ja erinevates kohtades.
35
Erinevalt bakterirakust toimuvad eukarüootses rakus transkriptsioon ja translatsioon nii ajaliselt kui ka
ruumiliselt lahus.
Bakterirakus algab translatsioon kohe kui konkreetse mRNA transkriptsioon on lõpule jõudnud.
64. Ribosoomide ehitus prokarüootses ja eukarüootses rakus.
Ribosoomid koosnevad suurest ja väikesest subühikust, milles sisalduvad rRNA ja valgud.
Ribosoomi alaosad eralduvad teineteisest, kui mRNA molekul on transleeritud ja ühinevad uuesti
translatsiooni initsiatsioonil.
Ribosoomide kolmemõõtmeline struktuur on üldjoontelt sarnane kõigis elusorganismides.
PROKARÜOOTNE RAKK
Bakterirakus asuvad ribosoomid on suurusega 70S
väike subühik (30S): 16S rRNA molekul + 21-st erinevat polüpeptiidi.
Suur subühik (50S): 2 RNA molekuli + 31 erinevat polüpeptiidi.
EUKARÜOOTNE RAKK
Eukarüootide tsütoplasmas asuvad ribosoomid on suurusega 80S.
väikesest subühik (40S): 18S RNA + 33 erinevat polüpeptiidi
suurest subühik (60S): 3 rRNA + 49 polüpeptiidi.
65. tRNA-de osalus translatsiooniprotsessis: tRNA laadimine aminohappega, tRNA seondumise
saidid ribosoomis.
tRNA laadimine aminohappega = tRNA aktiveerimine.
Aminohape seotakse tRNA molekuli 3´- OH rühma
külge aminohappe COOH rühma kaudu.
See toimub kahe-etapiliselt.
Esmalt aktiveerivad
aminoatsüül-tRNA süntetaasid aminohapped, kasutades selleks ATP energiat.
Seejärel seotakse aminohape tRNA molekuliga, moodustub aminoatsüül-tRNA.
Iga spetsiifiline aminoatsüül-tRNA-süntetaas tunneb ära teatud aminohappele vastavaid tRNA molekule.
tRNA seondumise saidid ribosoomis.
tRNA saab ribosoomis seonduda kolme saiti:
Esmalt seondub aminoatsüül-tRNA
aminoatsüül-saiti e.
A-saiti.
Peptidüül-saidis e.
P-saidis toimub aminohappe lisamine kasvavale polüpeptiidahelale.
E-saiti (exit site) liigub tRNA, millelt aminohape on seotud polüpeptiidahelasse.
Nende seondumissaitide põhiosa asub ribosoomi suures subühikus, samas kui mRNA on ribosoomi
väikese subühiku koostises.
Igale aminohappele vastab rakus üks kuni neli tRNA molekuli:
tRNA molekulid sisaldavad nukleotiidide tripleteid –
antikoodoneid
Translatsioonil paarduvad tRNA molekulid mRNA molekulis asuvate koodonitega antikoodonite
vahendusel
66. Võrrelge translatsiooni initsiatsiooni bakterites ja eukarüootses rakus.
Eukarüootides on translatsiooni initsiatsioon võrreldes bakterites toimuvaga komplekssem ja seda eeskätt
initsiatsioonifaktorite rohkuse tõttu. Üldiselt on initsiatsiooniprotsess aga sarnane, välja arvatud kaks
erinevust:
1) Polüpeptiidahelasse esimesena lülitatava metioniini aminorühm ei ole blokeeritud formüülrühmaga.
2) Initsiatsioonikompleks moodustub mRNA 5´-otsaga.
36
Ei ole vaja spetsiifilist järjestust mRNA molekuli alguses, mis paarduks rRNA-ga ribosoomis, vaid
initsiatsioonifaktorite hulgas on spetsiifiline valk
CBP, mis seondub mRNA 5´-otsas oleva CAP
otsaga ning edasi skanneerib initsiatsioonikompleks mRNA-d kuni esimese AUG koodonini.
Seetõttu algab eukarüootides translatsioon enamasti esimeselt AUG koodonilt.
67. Kirjeldage translatsiooni elongatsiooniprotsessi.
Translatsiooni elongatsiooni üldised printsiibid on eukarüootidel ja prokarüootidel sarnased, iga
aminohappe lisamine kasvavasse polüpeptiidahelasse toimub kolmeetapiliselt.
Bakteris E. coli toimub see järgmiselt:
1) Aminoatsüül-tRNA seondub ribosoomi A-saiti, paardudes antikoodonjärjestuse kaudu parasjagu A-
saidis asuva koodonjärjestusega mRNA molekulis. Selleks peab aminoatsüül-tRNA olema
assotsieerunud elongatsioonifaktoriga
EF-Tu, mis on seotud GTP-ga.
2) Peptiidsideme moodustumine ribosoomi A-saidis asuva aminoatsüül-tRNA aminorühma ja ribosoomi
P-saidis asuva tRNA-ga seotud kasvava polüpeptiidahela viimase aminohappe karboksüülrühma
vahel. Selle tulemusena vabaneb kasvav polüpeptiidahel tRNA-st P-saidis ja seotakse kovalentselt
tRNA-ga, mis asub A-saidis.
3) Selle tulemusena vabaneb kasvav polüpeptiidahel tRNA-st P-saidis ja seotakse kovalentselt tRNA-ga,
mis asub A-saidis
Polüpeptiidahela elongatsioon toimub kiiresti. Bakteris E. coli kulub eelpoolkirjeldatud kolme etapi
läbimiseks 0,05 sekundit. Seega kulub 300 aminohappe pikkuse polüpeptiidi sünteesiks ligikaudu 15
sekundit.
68. Kirjeldage translatsiooni terminatsiooniprotsessi.
Translatsioon termineerub, kui terminatsioonifaktorid tunnevad A-saidis ära stop koodonid
Polüpeptiidahela elongatsioon termineerub, kui ribosoomi A-saiti satub üks kolmest
terminatsioonikoodonist, UAA, UAG või UGA.
Stop-koodoneid tunnevad ära terminatsioonifaktorid, milleks on valgud tähistusega
RF (release factor).
Bakteris E. coli on kaks RF-i: RF-1 tunneb ära UAA ja UAG terminatsioonikoodoneid ning RF-2 UAA ja
UGA terminatsioonikoodoneid. Eukarüootides on ainult üks RF (eRF), mis tunneb ära kõiki
terminatsioonikoodoneid.
A-saiti sisenenud RF muudab peptidüültransferaasi aktiivsust, nii et peptidüültransferaas lisab
polüpeptiidahela viimase aminohappe karboksüülrühmale vee molekuli. Selle tulemusena vabaneb valmis
polüpeptiidahel P-saidis asuvalt tRNA molekulilt ning vaba tRNA liigub ribosoomi E-saiti. Translatsiooni
terminatsiooniprotsess lõpeb mRNA vabanemisega ribosoomilt ja ribosoomi jaotumisega kaheks alaosaks.
Seejärel on ribosoomi subühikud jällegi valmis ühinema järgmise polüpeptiidahela sünteesiks.
69. Geneetiline kood ja selle omadused.
Geneetiline kood – kood, mille järgi DNA-st RNA-sse kandunud geneetiline informatsioon tõlgitakse
nukleotiidide järjestusest polüpeptiidahela aminohapete järjestuseks.
Geneetilise koodi üksus: kolmest järjestikusest nukleotiidist moodustuv triplett; igale aminohappele vastab
üks triplett.
OMADUSED:
(1) Geneetiline kood
põhineb nukleotiidide triplettidel, kolm nukleotiidi määravad ära ühe aminohappe
polüpeptiidis
(2) Geneetiline kood
ei ole kattuv
37
(3) Geneetiline kood on komavaba, kõik mRNA-s asuvad koodonid loetakse translatsioonil järjest,
ühes
lugemisraamis (reading frame)
(4) Geneetiline kood on
degenereerunud e. kõdunud: peaaegu kõigile aminohapetele vastab enam kui
üks koodon
(5) Geneetiline kood on
seaduspärane (ingl. k. ordered)
(6) Geneetilisse koodi kuuluvad
spetsiifilised koodonid, mis on signaaliks translatsiooni
initsiatsioonile ja terminatsioonile
(7) Geneetiline kood on (v.a. mõned erinevused mitokondrites)
universaalne kõigile elusorganismidele
70. Koodon-antikoodon paardumine, selle täpsus. Mis on supressor-tRNA?
tRNA antikoodonjärjestus paardub mRNA-s asuva koodonjärjestusega koodoni kahe esimese nukleotiidi
osas väga täpselt, vastavuses lämmastikaluste komplementaarsuse põhimõttele.
Koodoni kolmandas positsioonis asuva nukleotiidiga paardumine on ebatäpne, mistõttu seda saiti
koodonis nimetatakse lõdvaks. Nii saab näiteks seriini tRNA antikoodoniga AGG seonduda nii
koodonitele UCU kui ka UCC.
Supressormutatsioonid – asendusmutatsioon ühes geenis surub maha e. supresseerib teises geenis tekkinud
mutatsiooni avaldumise: Näiteks mutatsioonid tRNA geenides, mille tulemusena mutatsioon tRNA
antikoodonis võimaldab tRNA-l paarduda mRNA-s oleva stop koodoniga ja taastada täispika polüpeptiidi
sünteesi. Stop koodoniga paarduvat mutantset tRNA-d nimetatakse supressor-tRNA-ks
71. Mutatsioonisagedust mõjutavad tegurid.
Mutatsioonisagedust tõstavad:
1. Kiirgus. (röntgenkiired, gammakiirgus, kosmiline kiirgus)
2. mitmesugused DNA-d kahjustavad ja modifitseerivad kemikaalid
Mõjutavad veel:
1. Ploidsus. Bakterites, kelle genoom on haploidne, on mutatsioonidel võimalus kohe avalduda.
Diploidse organismi puhul on mutatsiooni avaldumise seisukohalt määravaks see, kas tegemist on
dominantse või retsessiivse mutatsiooniga. Retsessiivne mutatsioon saab avalduda ainult
homosügootses olekus, dominantne avaldub aga koheselt.
2. Hulkraksetes organismides sõltub mutatsioonist põhjustatud fenotüübilise efekti avaldumine ka
sellest, millal ja mis tüüpi rakus mutatsioon on tekkinud. Näiteks kõrgemate loomade puhul
eristatakse
somaatilisi ja
sugurakkudes tekkinud mutatsioone (germinal mutations).
Järgmisesse põlvkonda kanduvad edasi ainult viimased.
Mutatsioonide tekkesagedust mõjutavad tegurid:
(1) DNA replikatsiooni täpsus
(2) DNA reparatsiooni efektiivsus
(3) Mutageensete faktorite olemasolu ja hulk keskkonnas
72. Spontaansed ja indutseeritud mutatsioonid. Mutatsioonitekke juhuslikkust tõendavad katsed.
Spontaanne mutatsioonisagedus rakus on madal. Bakterites ja bakteriofaagides (bakteriviirustes) tekib
spontaanseid mutatsioone sagedusega 10-8 kuni 10-10 nukleotiidipaari kohta generatsioonis, eukarüootse
raku puhul aga vahemikus 10-7 kuni 10-9 nukleotiidipaari kohta generatsioonis. Kui arvutada
mutatsioonisagedust geeni kohta, võttes keskmise geeni pikkuseks 1000 aluspaari, saame
mutatsioonisageduse, mis varieerub vahemikus 10-4 kuni 10-7 generatsiooni kohta.
Teatud füüsikalised tegurid nagu näiteks ultraviolettkiirgus ja kemikaalid, mis kahjustavad DNA-d,
teisisõnu
mutageenid, võivad tõsta mutatsioonisagedust rakus võrreldes spontaanse
38
mutatsioonisagedusega mitu suurusjärku. Mutageenide toimel tekkinud mutatsioone nimetatakse
indutseeritud mutatsioonideks.
KATSE: Kas antibiootikumile resistantsed mutandid on eelnevalt bakteripopulatsioonis olemas või
indutseerib vastava mutatsiooni teket antibiootikum?
jäljendkülvi võimaldab huvipakkuva tunnuse osas bakteripopulatsioonis korraga läbi testida palju
individuaalseid rakke. Kui külvata piisavalt lahjendatud bakterikultuur tardsöötmele, moodustuvad
sinna bakterikolooniad
Bakterikolooniate testimine jäljendkülvi meetodil näitas, et mutatsioonid tekivad juhuslikult.
Streptomütsiini suhtes resistentsed mutandid olid bakteripopulatsioonis olemas juba enne rakkude
kokkupuutumist antibiootikumiga. Sarnased tulemused saadi ka bakterifaagi resistentsete mutantide tekke
puhul.
73. Mutatsioonide mõju organismile. Mutatsioonide avaldumine haploidsetes ja diploidsetes
organismides.
Mutatsioonid – vead geneetilise materjali kopeerimisel viivad uute alleelide tekkele
Mutant – organism, kellel avaldub mutatsiooni tagajärjel uus fenotüüp
Mutatsioonid võimaldavad populatsioonisisest geneetilist varieeruvust ja on alusmaterjaliks
evolutsioonile. Mutatsioonisagedus ei tohi olla ei liiga kõrge ega liiga madal
Bakterites, kelle genoom on haploidne, on mutatsioonidel võimalus kohe avalduda. Diploidse organismi
puhul on mutatsiooni avaldumise seisukohalt määravaks see, kas tegemist on dominantse või retsessiivse
mutatsiooniga. Retsessiivne mutatsioon saab avalduda ainult homosügootses olekus, dominantne avaldub
aga koheselt.
Hulkraksetes organismides sõltub mutatsioonist põhjustatud fenotüübilise efekti avaldumine ka sellest,
millal ja mis tüüpi rakus mutatsioon on tekkinud. Näiteks kõrgemate loomade puhul eristatakse
somaatilisi ja
sugurakkudes tekkinud mutatsioone (germinal mutations). Järgmisesse põlvkonda
kanduvad edasi ainult viimased.
74. Punktmutatsioonid: transitsioonid, transversioonid ja raaminihkemutatsioonid. Kuidas
mõjutavad erinevat tüüpi punktmutatsioonid geeni poolt kodeeritud polüpeptiidi funktsiooni?
Punktmutatsioonid – muutused geeni tasemel.
Transitsioonid – puriin asendub puriiniga (A ↔ G) või pürimidiin pürimidiiniga (C ↔ T)
Transversioonid – puriin asendub pürimidiiniga või vastupidi (nt. T ↔ G ja A ↔ C asendused)
1.
Sünonüümsed mutatsioonid – koodon määrab sama aminohapet
2.
Missens mutatsioonid – muutub koodoni tähendus, määrab teist aminohapet
3. Nonsens mutatsioonid – viivad stoppkoodoni tekkele
4.
Raaminihke mutatsioonid – muutub lugemisraam ja seetõttu ka valgu aminohappeline järjestus
Enamus kahjulikke mutatsioone populatsioonis on retsessiivsed. Dominantseid kahjulikke mutatsioone
sisaldavad alleelid kõrvaldatakse loodusliku valiku teel kiiremini.
75. Ames´i test kemikaalide mutageensuse uurimiseks.
39
Bruce Ames ja kolleegid töötasid välja kiire, odava ja väga tundliku meetodi kemikaalide mutageensuse
testimiseks. See meetod põhineb bakterite histidiini suhtes auksotroofsete mutantide
reverteerumissageduste mõõtmisel tingimustes, kus bakterite kasvukeskkonda on lisatud uuritavaid
kemikaale. Mida mutageensem on kemikaal, seda suuremal hulgal tekib bakteripopulatsiooni revertante
(bakterirakke, kes on võimelised kolooniaid moodustama histidiini-vabas keskkonnas).
Kuna paljud kemikaalid on mutageensed vaid replitseeruva DNA korral, lisatakse bakterite
kasvukeskkonda vähesel määral ka histidiini, et rakud saaksid mõned korrad jaguneda, samas aga mitte
piisavalt vähe, et ei moodustuks söötmel nähtavaid kolooniaid. Nii testiti läbi tuhandeid erinevaid
kemikaale.
Osade potentsiaalselt kartsinogeensete kemikaalide puhul mutatsioonisagedus bakterirakus ei tõusnud.
Põhjuseks oli see, et need kemikaalid muutuvad mutageenseks alles eukarüoodi rakus toimuvate
biokeemiliste protsesside tagajärjel. Selleks, et tekitada algsetest kemikaalidest metaboliite, mis võiksid
olla potentsiaalsed mutageenid, töödeldakse uuritavaid kemikaale enne bakterikultuurile lisamist roti
maksa ekstraktiga.
Ames’i testi tulemusena selgus ka, et mitmed sigareti suitsus olevad komponendid võivad stimuleerida
raaminihke mutatsioonide teket.
76. Põhilised DNA reparatsioonimehhanismid rakkudes.
1. Valgusest sõltuv fotoreaktivatsioon: vea parandamine kohapeal
Fotoreaktivatsioon kõrvaldab tümiini dimeere ja vajab valgusenergiat.
2. Väljalõikereparatsioon: lämmastikualuste kõrvaldamine.
Glükosülaasid kõrvaldavad DNA-st kahjustatud lämmastikaluseid.
Eksonukleaas kõrvaldab lõigu DNA
ahelast ja tühiku sünteesib täis DNA polümeraas, kasutades vastasahelat matriitsina
3. Replikatsioonijärgne valepaardumisi kõrvaldav DNA “mismatch” reparatsioon MMR: Nukleotiidide
kõrvaldamine DNA ahelast.
UvrABC väljalõike-nukleaas kõrvaldab DNA ahelast DNA kahjustuse, mis on tekkinud UV-kiirguse
tagajärjel
4. Rekombinatsiooniline reparatsioon: rekombinantne reparatsioon DNA katkete puhul.
77. SOS vastus bakterites.
DNA kahjustuste või DNA replikatsiooni inhibeerimise tagajärjel tekib rakkudes SOS vastus.
Induktoriteks on UV-kiirgus, alküleerivad ühendid, tümiini vaegus ja ravimid. DNA replikatsiooni
blokeerimine kahjustuste kohas toob esile ssDNA, kuna DNA polümeraasi III peatumisel jätkab DNA
helikaas DnaB DNA ahelate lahtikeeramist. ssDNA aktiveerib RecA valgu, moodustub
nukleoproteiinne
filament. SOS vastusena käivitub DNA reparatsioon (rekombinatsiooniline reparatsioon) ja SOS
mutagenees. Bakteris E. coli põhjustab SOS mutageneesi vigaderohke DNA polümeraas V, mis jätkab
kahjustuse kohal peatunud DNA polümeraasi III asemel vigaderohket DNA sünteesi.
78. Ristsiirde toimumise mehhanism Holliday mudeli põhjal.
DNA ahelatesse tehakse katked ja DNA ahela ots ühest kromosoomist liidetakse teise kromosoomi DNA
ahela otsaga - toimub ahelate vahetus.
Homoloogiliste kromosoomide vahel toimuva rekombinatsiooni käigus moodustub struktuur, kus
tütarkromatiidide DNA ahelad on teineteisega risti – Holliday struktuur.
Tütarkromatiidide lahutamisel riststruktuur roteerub ja DNA ahelatesse tehakse kaks katket:
Ristsiire toimub juhul, kui roteerunud Holliday struktuuri tehakse katked vastasahelatesse
Samasse ahelasse katkete tegemisel moodustub vaid lühike heterodupleksi ala
79. Komplementatsioonitesti
rakendus,
selle
erinevus
rekombinatsioonitestist.
Komplementatsioonitesti piirangud.
40
Komplementatsioonitesti saab rakendada ainult retsessiivsete mutatsioonide testimisel.
Diploidsete organismide puhul (näiteks äädikakärbes) saadakse trans-heterosügoote sel viisil, et ristatakse
omavahel isendeid, kellest üks on homosügootne ühe ja teine teise mutatsiooni osas.
Lewis ristas erinevaid äädikakärbse mutante, kombineerides uuritavaid mutatsioone nii cis – kui trans-
konfiguratsioonis. Selgus, et tunnuse avaldumine (silma värvus) sõltus mutatsioonide asukohast, sellest
kas need paiknesid ühes või kahes erinevas X kromosoomis.
Komplementatsioonitest ja rekombinatsioonitest annavad vastuseid erinevatele küsimustele:
Komplementatsioonitest näitab, kas mutatsioonid paiknevad samas geenis, kas nad on alleelsed
(kumbki mutatsioon tekitab sama alleeli). Jälgitakse, kas ristamisel saadud heterosügoodid on
mutantse fenotüübiga või komplementeeritud, metsiktüüpi fenotüübiga
Rekombinatsioonitesti abil uuritakse, kas mutatsioonid on aheldunud (asuvad samas kromosoomis) ja
kui on, siis kui kaugel nad asuvad teineteisest kromosoomis
Komplementatsioonitesti piirangud:
1)Ei ole rakendatav dominantsete või kodominantsete mutatsioonide puhul, mis avalduvad
heterosügootses oleku
2)Tulemuste analüüs on raskendatud epistaatiliste mutatsioonide puhul, kus üks mutatsioon mõjutab teise
avaldumist. Epistaasi saab kindlaks teha cis-testiga: cis-heterosügoodid on metsiktüüpi fenotüübi asemel
mutantse fenotüübiga
3)Polaarsete mutatsioonide puhul mõjutab mutatsioon mitte üksnes selle geeni avaldumist, kus ta asub
vaid ka külgnevate geenide avaldumist.
Polaarsed mutatsioonid takistavad komplementatsioonitesti läbiviimist. Trans-heterosügoodi puhul ei
saada komplementatsiooni mutatsioonide asumisel erinevates geenides juhul, kui üks mutatsioonidest on
polaarne ja mõjutab mõlema geeni avaldumist.
80. Komplekssed seosed geenide ja polüpeptiidide vahel: alternatiivne splaissing, geenisegmentide
rekombineerumine antikehade puhul.
Geenide ja polüpeptiidide kolineaarsus ei pruugi selle algses tähenduses alati kehtida:
1. Geenide kattuvus
Geenide kattuvus – sama nukleotiidne järjestus kuulub kahe või enama arvu geenide koostisesse, neid
loetakse erinevates lugemisraamides. Esineb sageli viiruste puhul
2. Eukarüootsed geenide puhul võib toimuda alternatiivne splaissing
Alternatiivne splaissing võimaldab sünteesida polüpeptiide, mis on omavahel lähedases suguluses, kuid
erinevates valgu isovormides on teatavad lõigud aminohappelisest järjestusest puudu. Erinevad valgu
isovormid võivad avalduda organismi erinevates kudedes
3. Erinevaid antikehi (immunoglobuliine) kodeerivad järjestused saadakse erinevate
geenisegmentide kombineerumise tulemusena
Iga antikeha (immunoglobuliin Ig), mis seondub spetsiifiliselt ainult tema poolt äratuntava bioloogilise
võõrmaterjaliga (antigeeniga), sisaldab varieeruvaid regioone. Varieeruvaid geenisegmente on palju ja
need kombineeruvad rekombinatsioonil konstantsete regioonidega.
Varieeruvaid geenisegmente (V) on palju ja need kombineeruvad rekombinatsioonil konstantsete
regioonidega (C). DNA järjestused, mis kodeerivad antikehade segmente, ei vasta klassikalisele geeni
definitsioonile
Sel puhul räägitakse geeni segmentidest, mis liidetakse üheks geeniks rekombinatsiooni teel
41
Kaasaegse geeni käsitluse järgi on geen informatsiooniüksus, mis määrab ära ühe polüpeptiidi, struktuurse
või regulatoorse RNA molekuli sünteesi. Geeni koostisesse kuuluvad ka 5´ja 3´mittekodeerivad regioonid,
mis reguleerivad transkriptsiooni ja translatsiooni ning geen hõlmab ka intronjärjestusi.
42
Document Outline
- Bruce Ames ja kolleegid töötasid välja kiire, odava ja väga tundliku meetodi kemikaalide mutageensuse testimiseks. See meetod põhineb bakterite histidiini suhtes auksotroofsete mutantide reverteerumissageduste mõõtmisel tingimustes, kus bakterite kasvukeskkonda on lisatud uuritavaid kemikaale. Mida mutageensem on kemikaal, seda suuremal hulgal tekib bakteripopulatsiooni revertante (bakterirakke, kes on võimelised kolooniaid moodustama histidiini-vabas keskkonnas).
- Kuna paljud kemikaalid on mutageensed vaid replitseeruva DNA korral, lisatakse bakterite kasvukeskkonda vähesel määral ka histidiini, et rakud saaksid mõned korrad jaguneda, samas aga mitte piisavalt vähe, et ei moodustuks söötmel nähtavaid kolooniaid. Nii testiti läbi tuhandeid erinevaid kemikaale.
- Osade potentsiaalselt kartsinogeensete kemikaalide puhul mutatsioonisagedus bakterirakus ei tõusnud. Põhjuseks oli see, et need kemikaalid muutuvad mutageenseks alles eukarüoodi rakus toimuvate biokeemiliste protsesside tagajärjel. Selleks, et tekitada algsetest kemikaalidest metaboliite, mis võiksid olla potentsiaalsed mutageenid, töödeldakse uuritavaid kemikaale enne bakterikultuurile lisamist roti maksa ekstraktiga.
- Ames’i testi tulemusena selgus ka, et mitmed sigareti suitsus olevad komponendid võivad stimuleerida raaminihke mutatsioonide teket.























































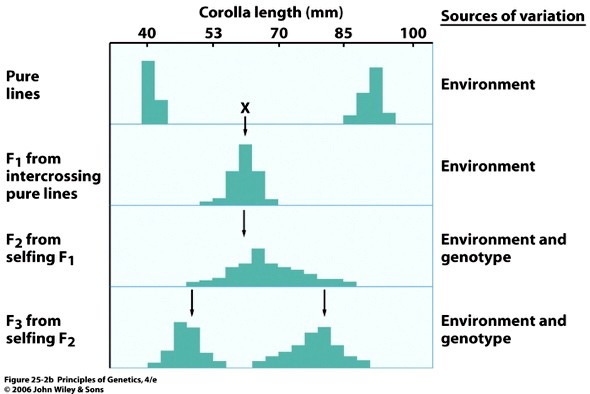
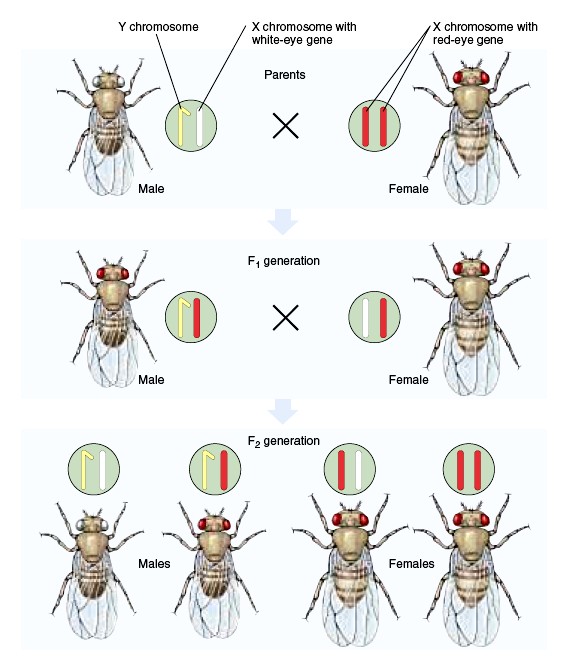
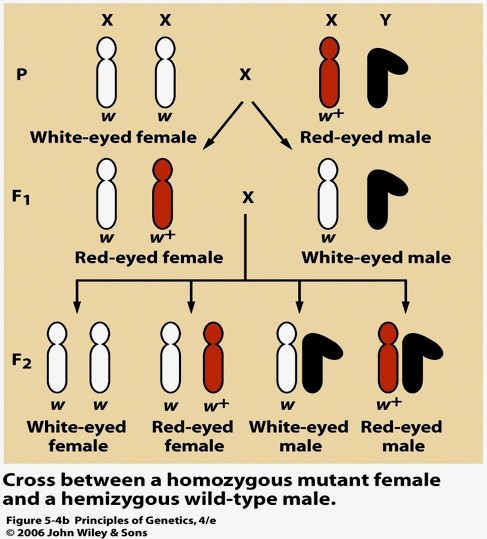
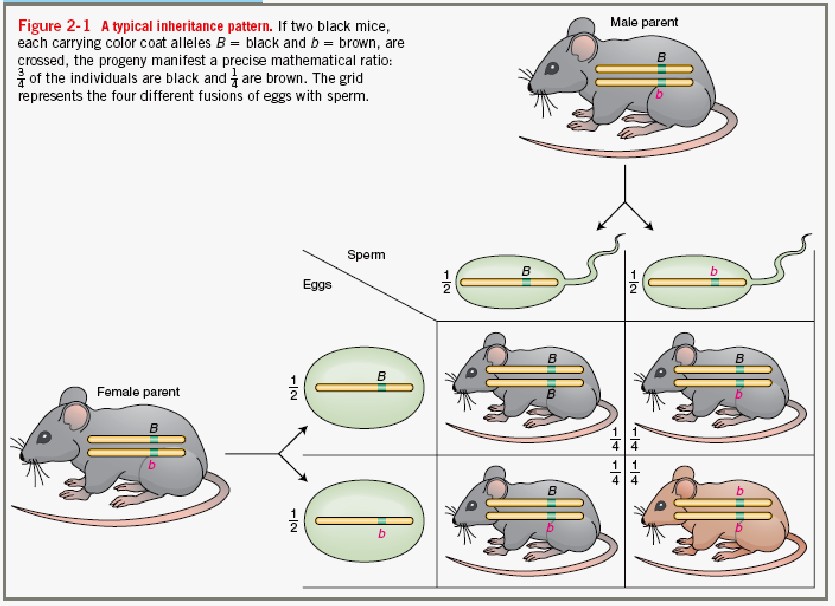
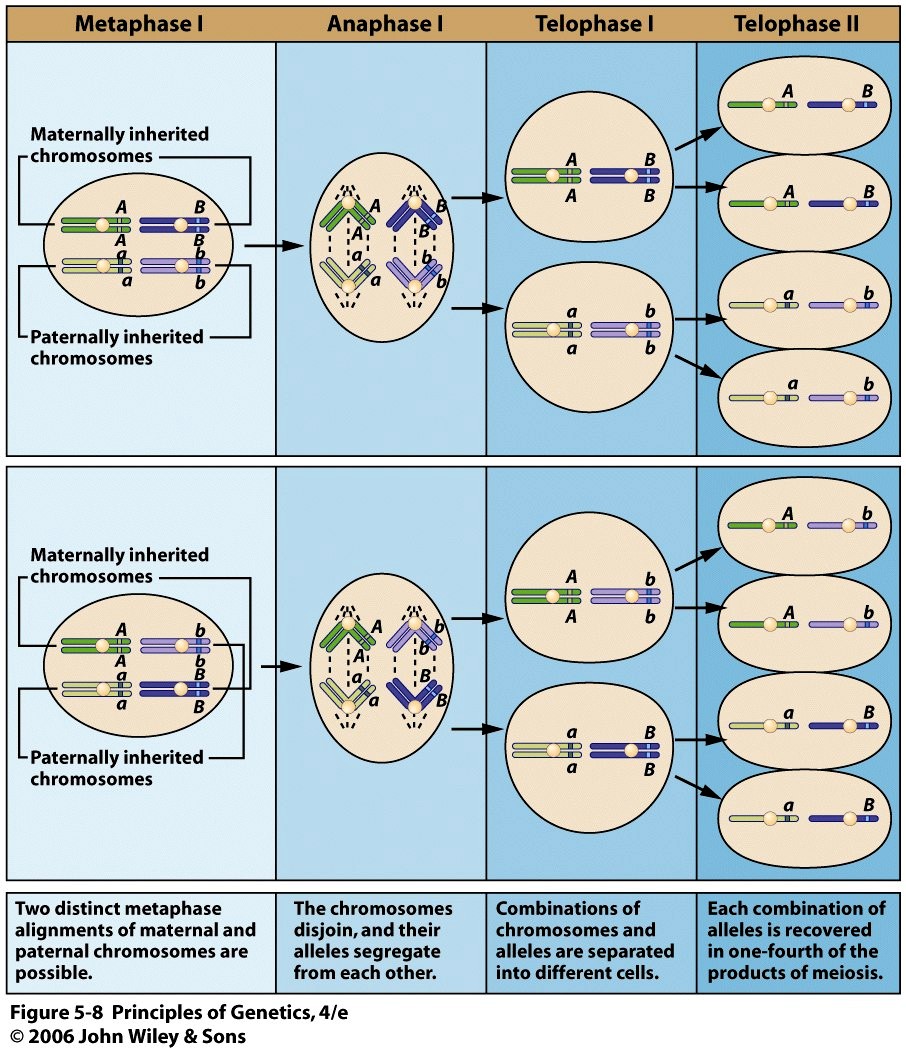
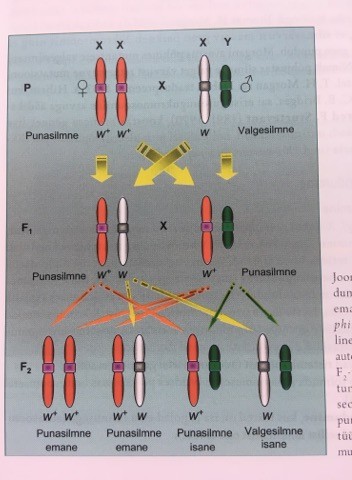
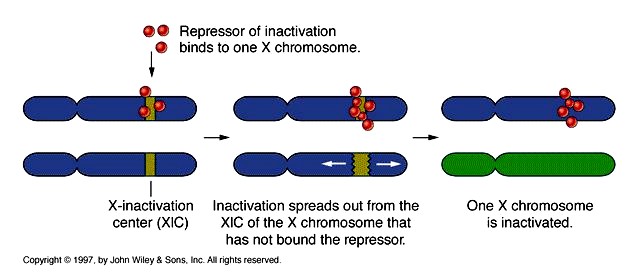
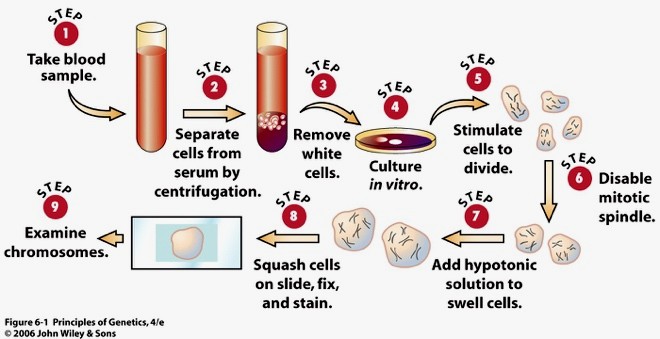
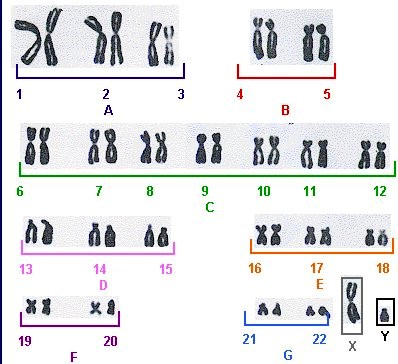
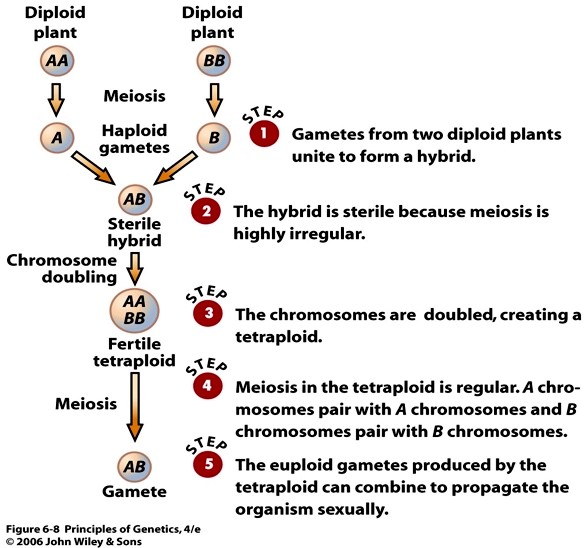
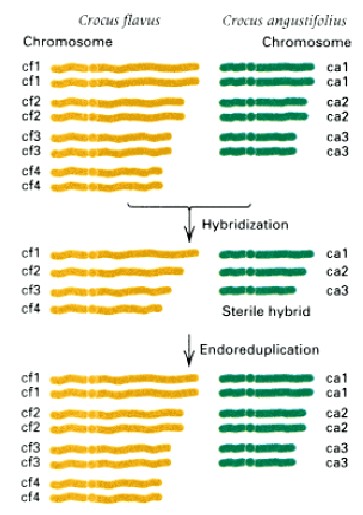
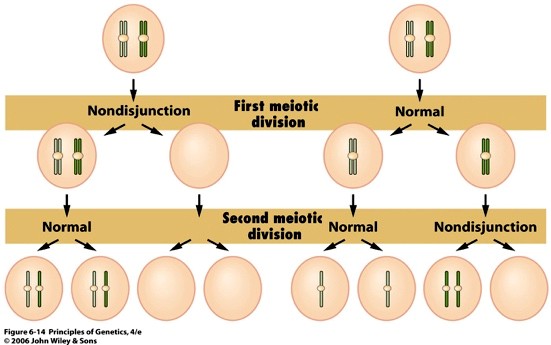
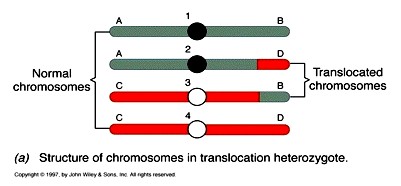
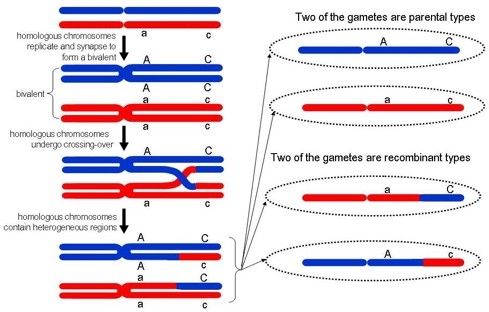
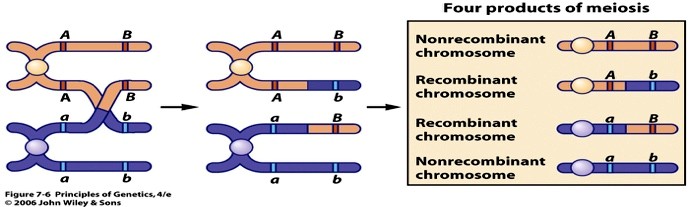
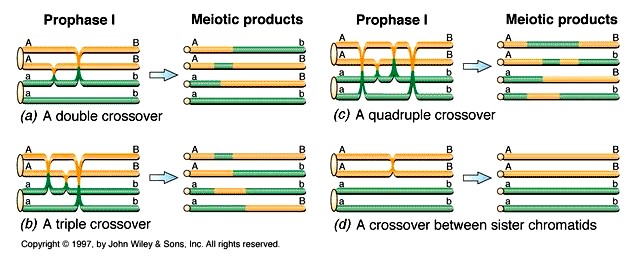
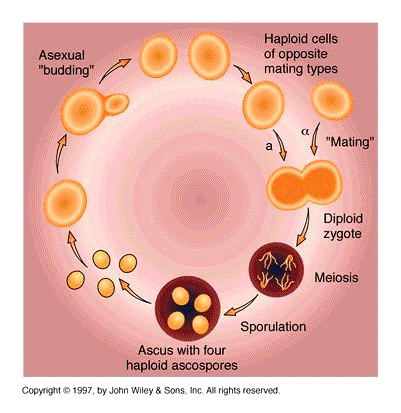
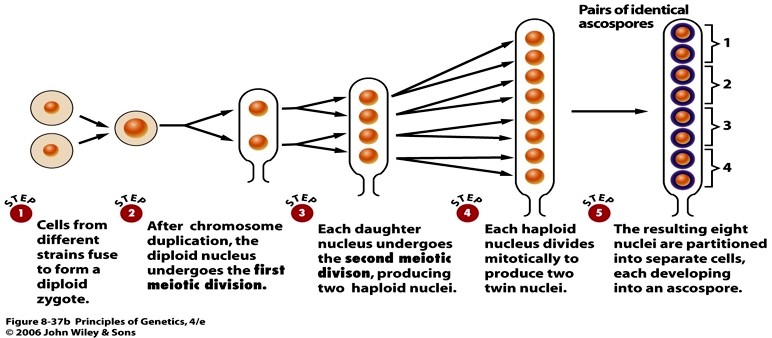
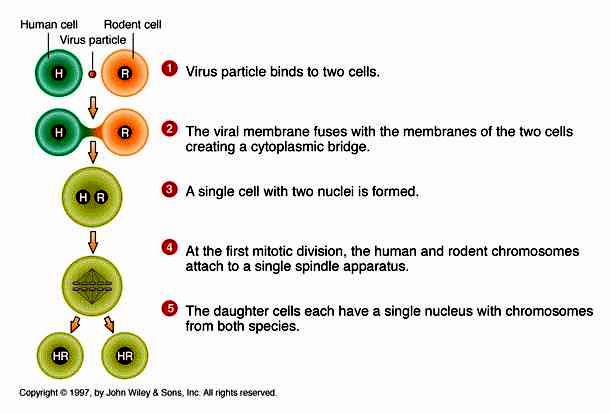
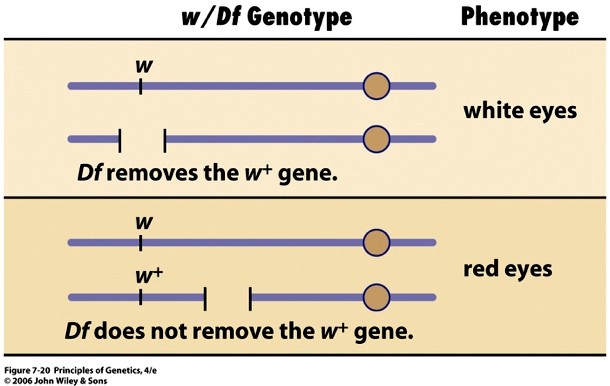
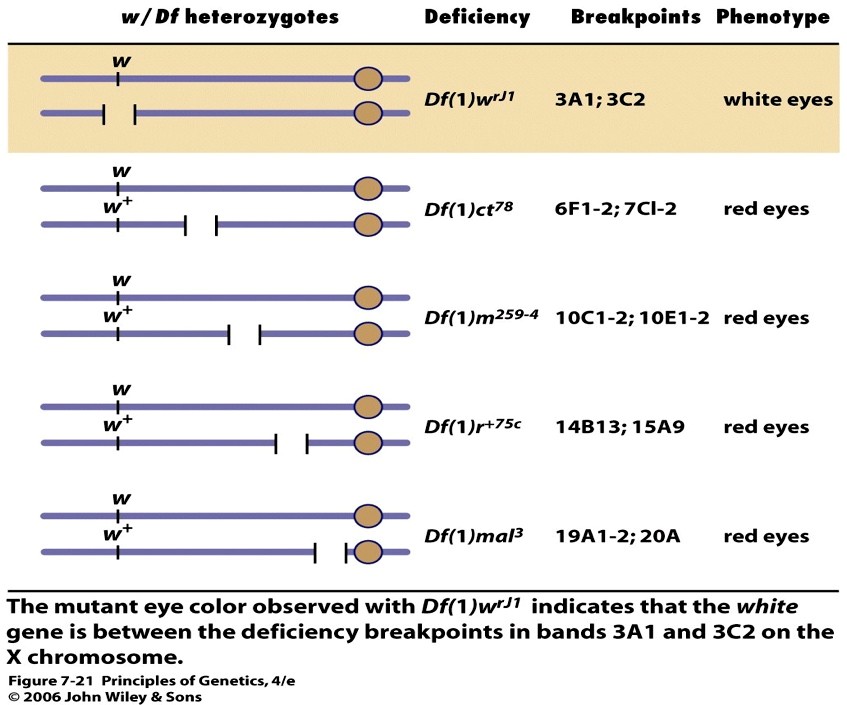
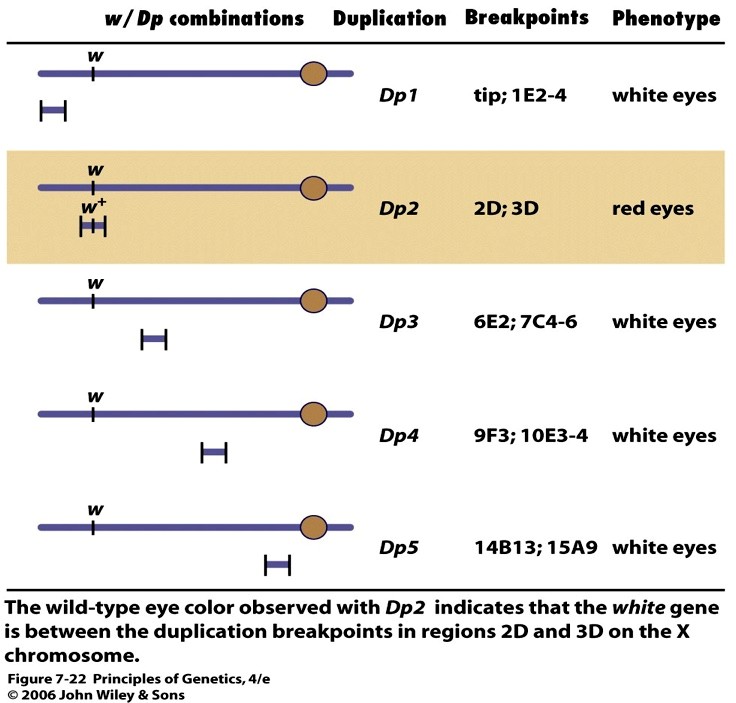
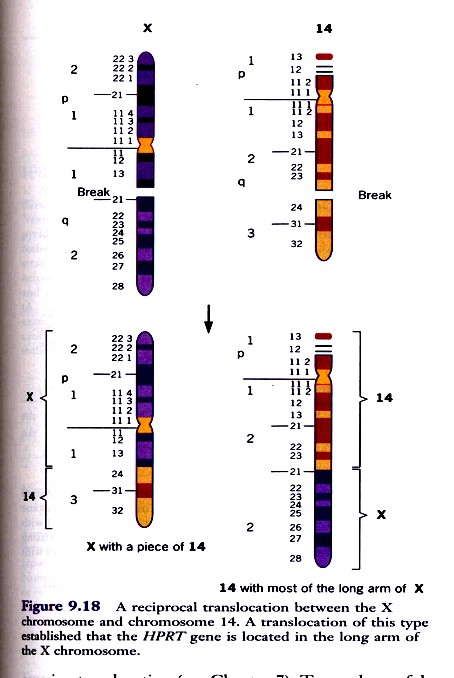
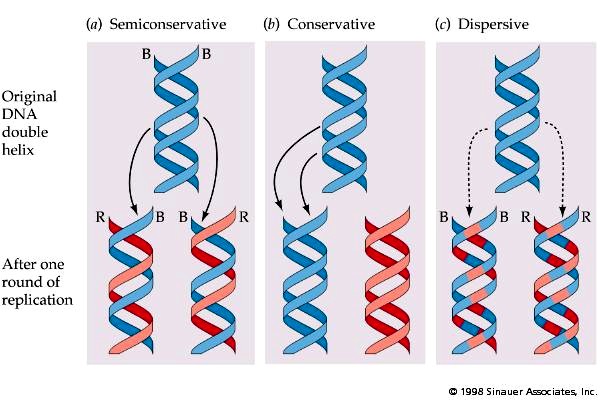
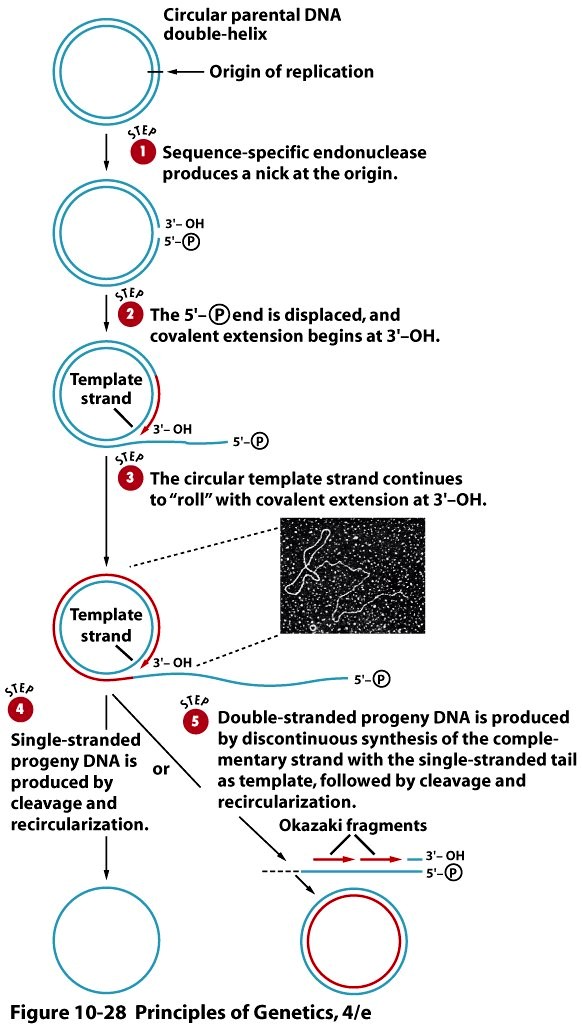
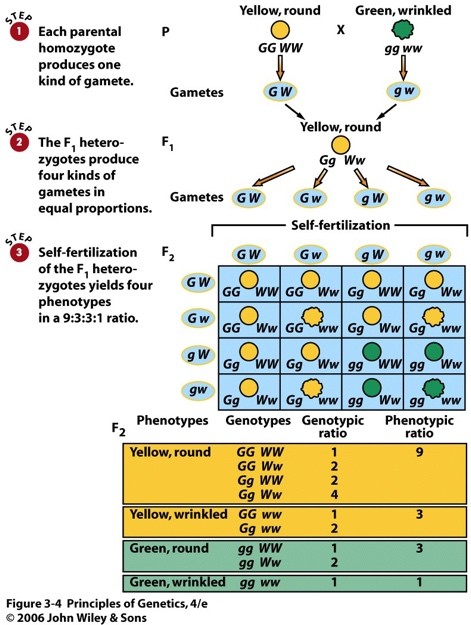
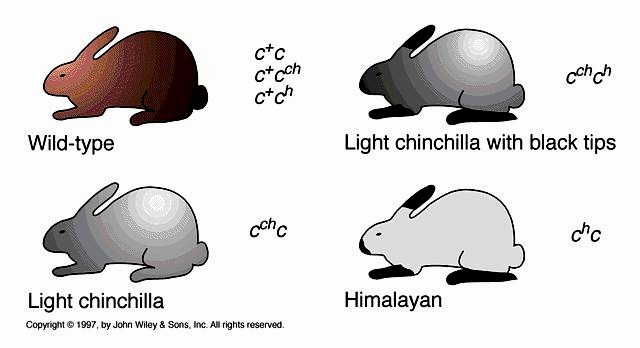
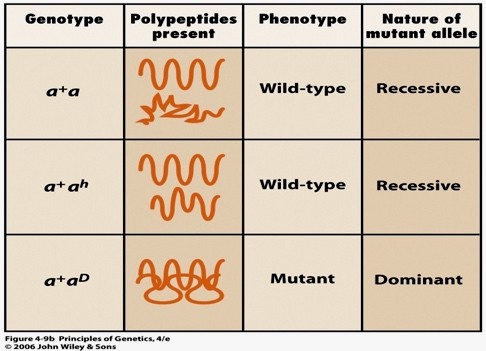
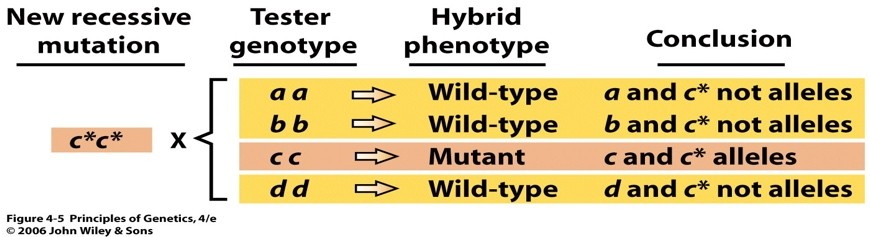
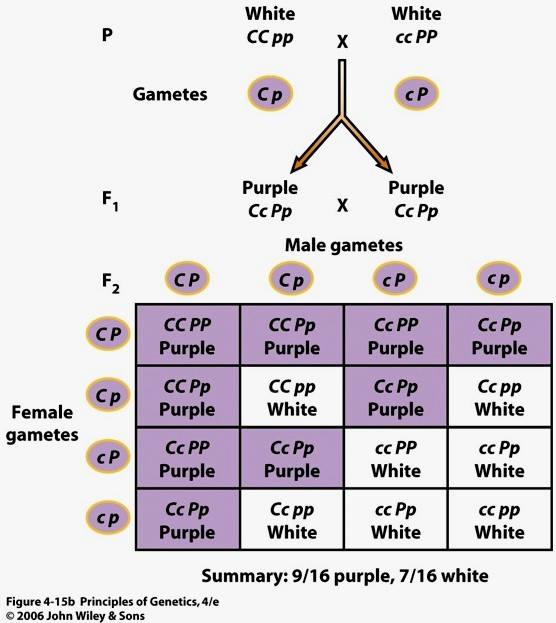
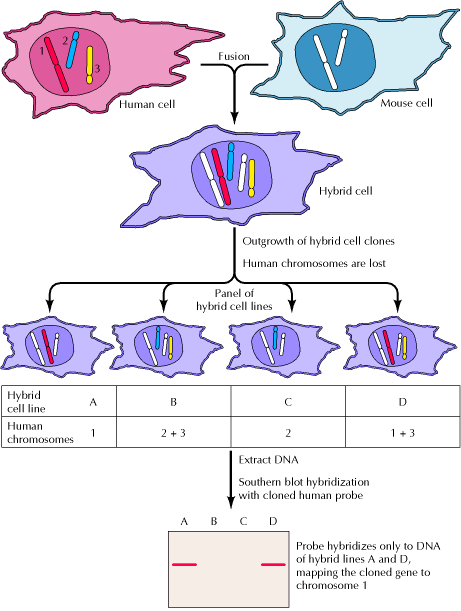
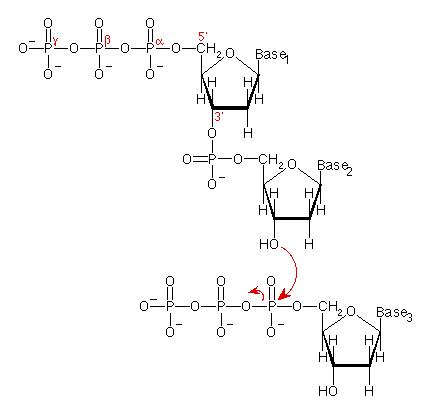
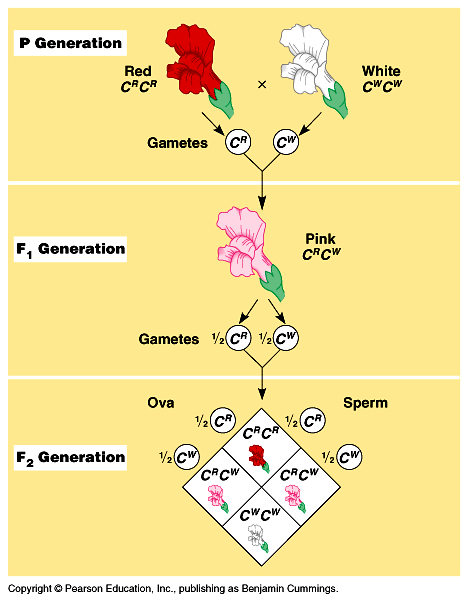




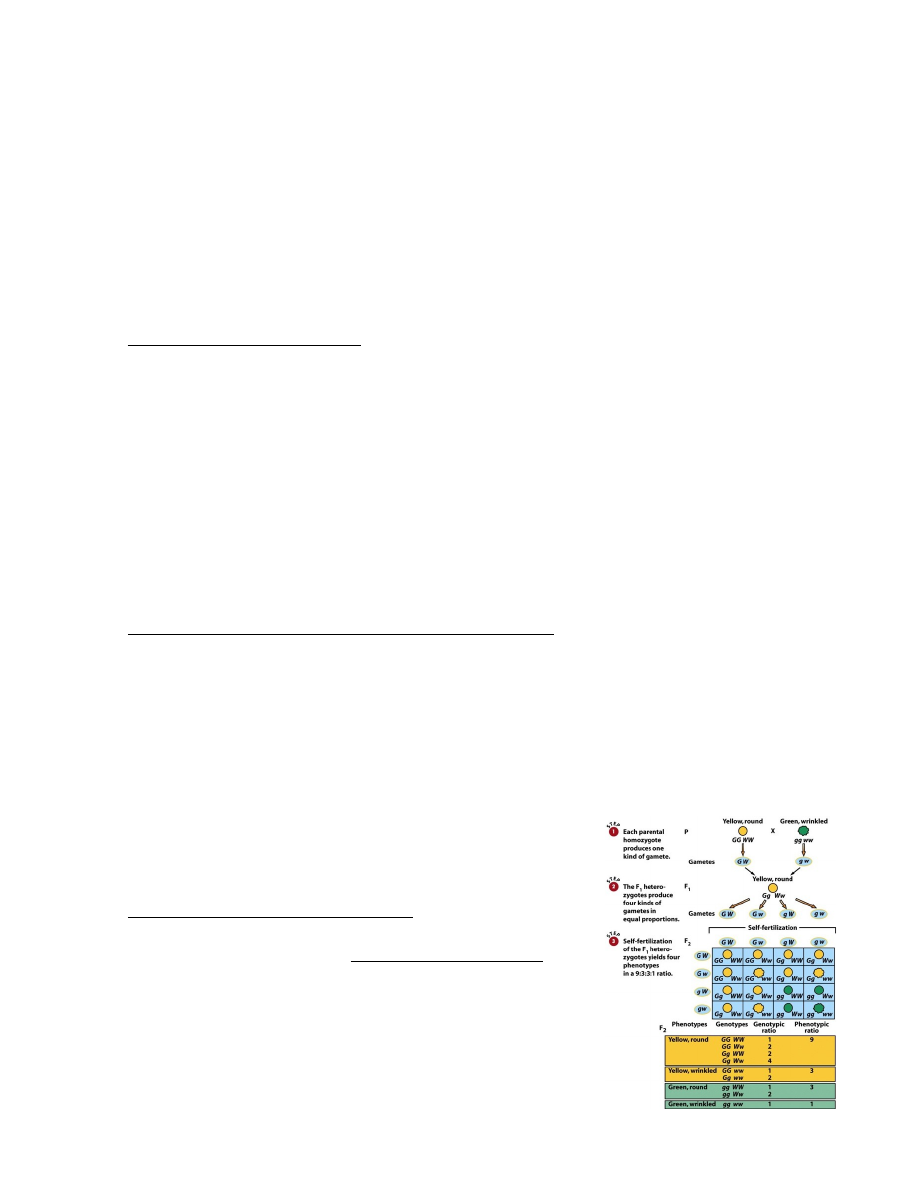
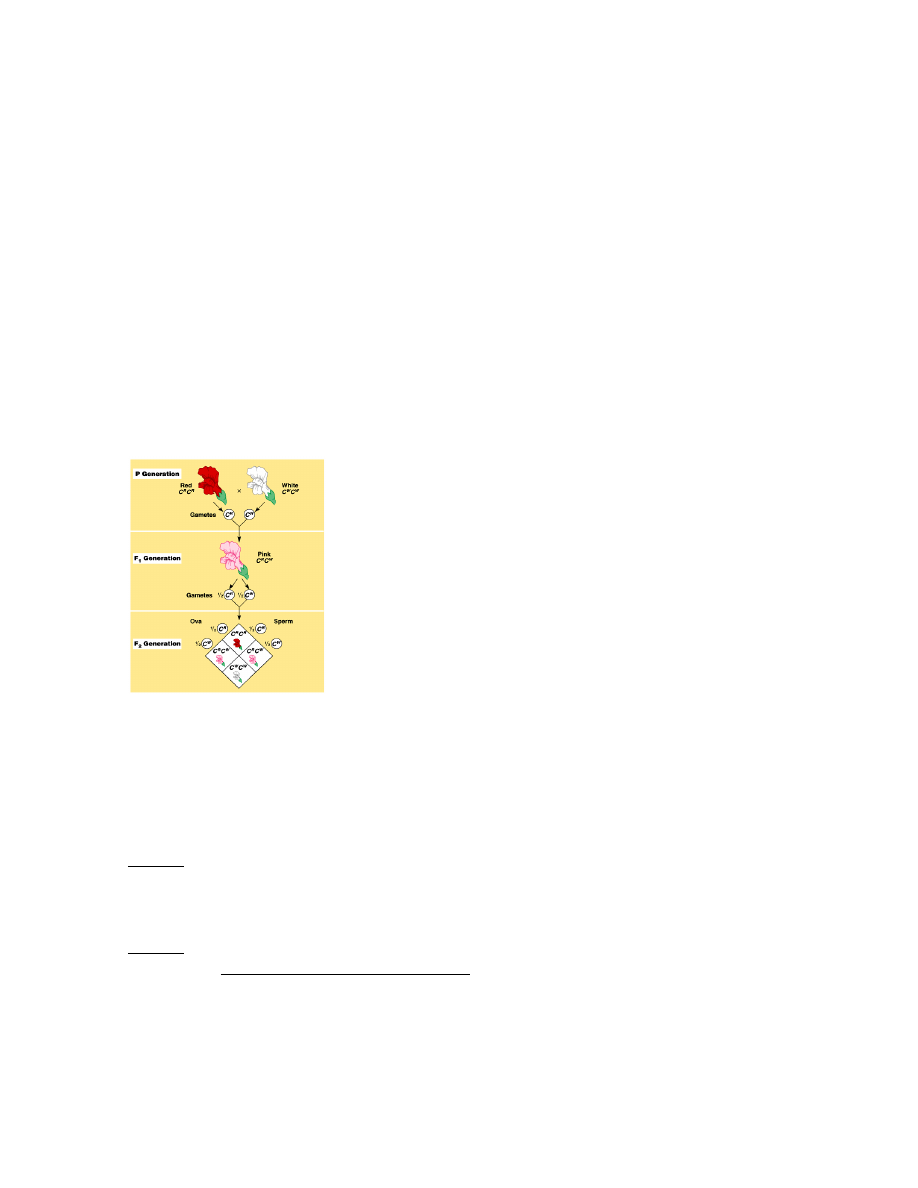

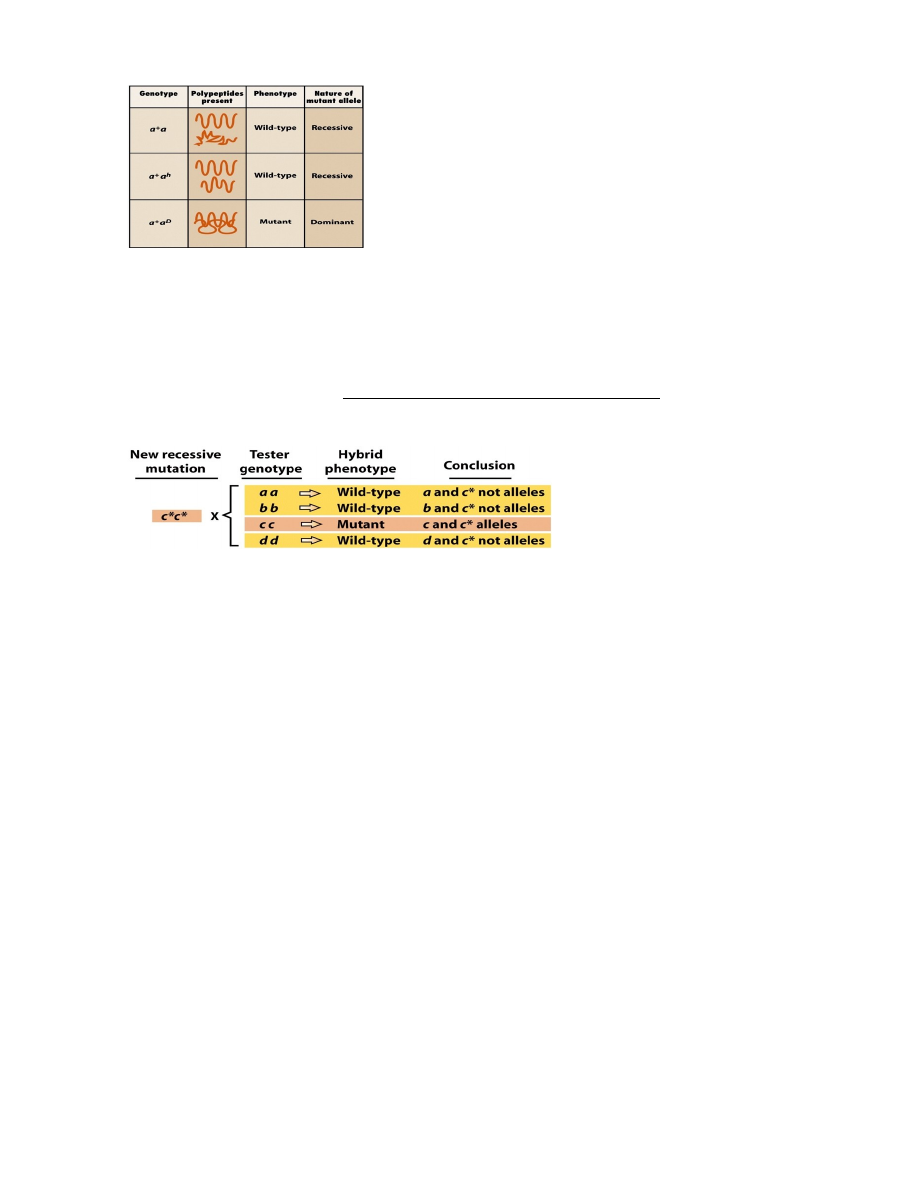
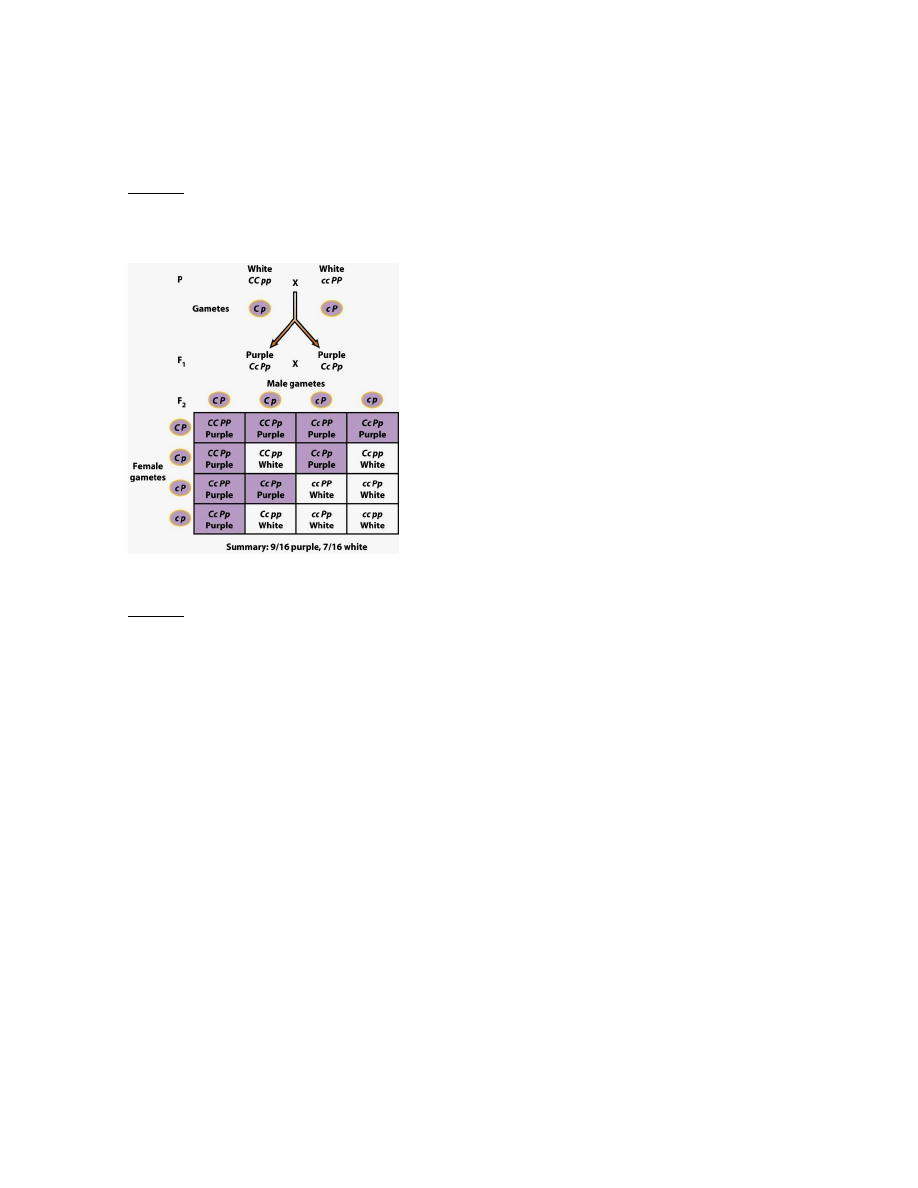












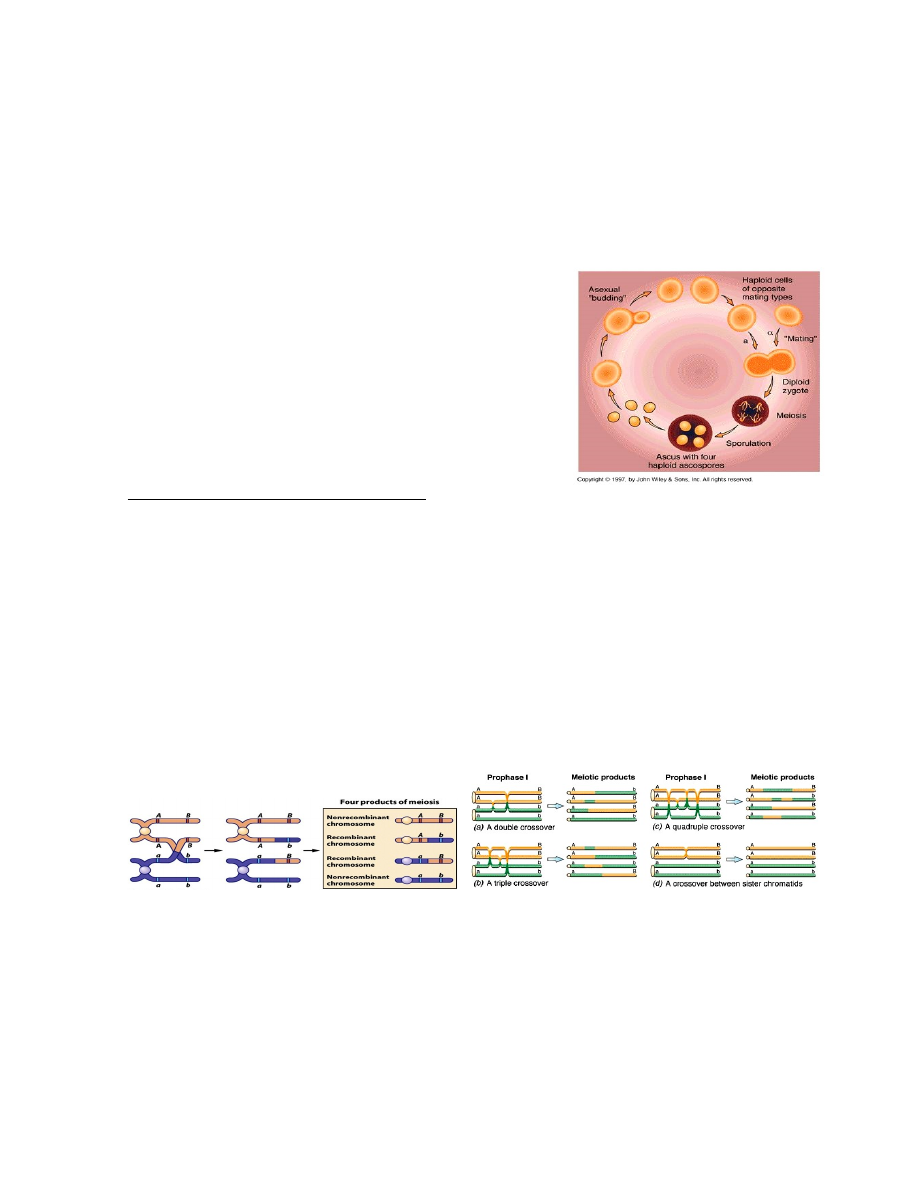


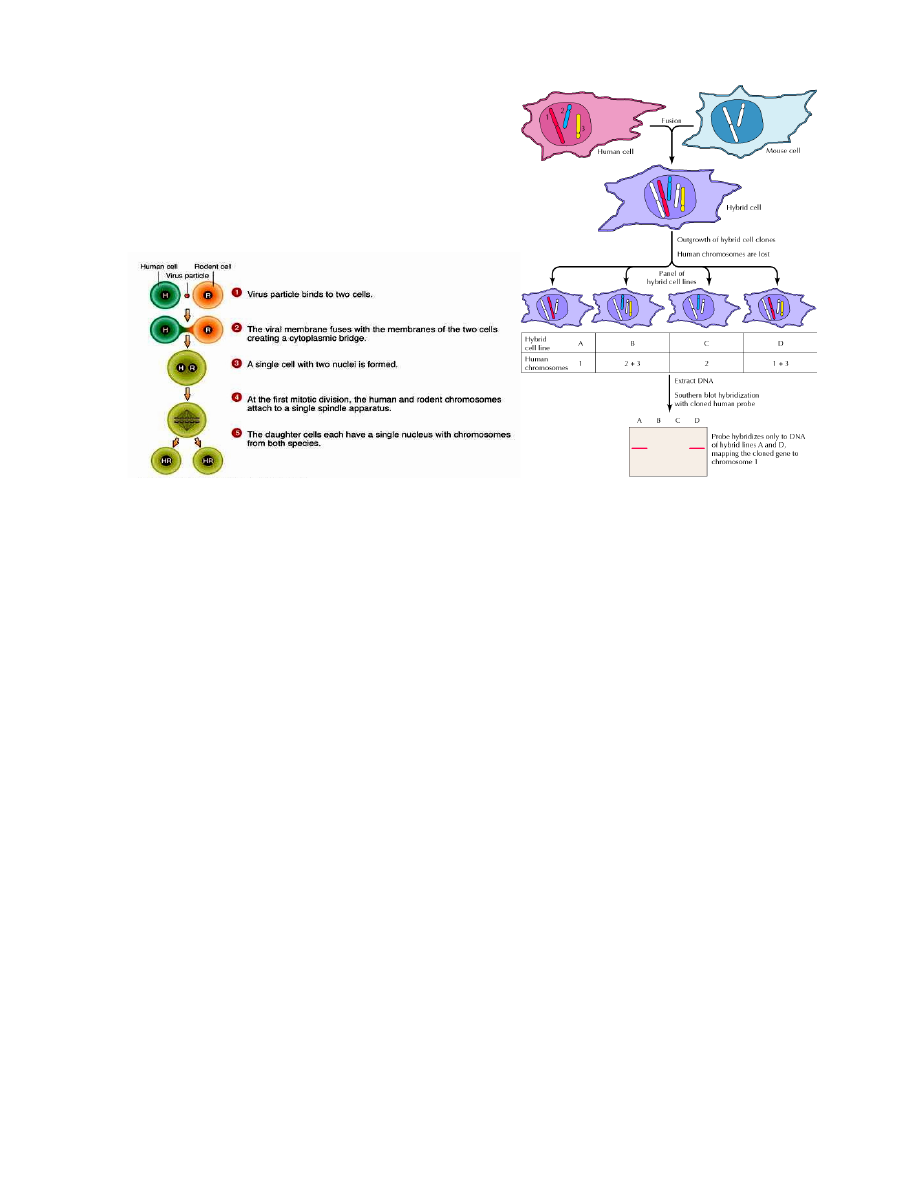
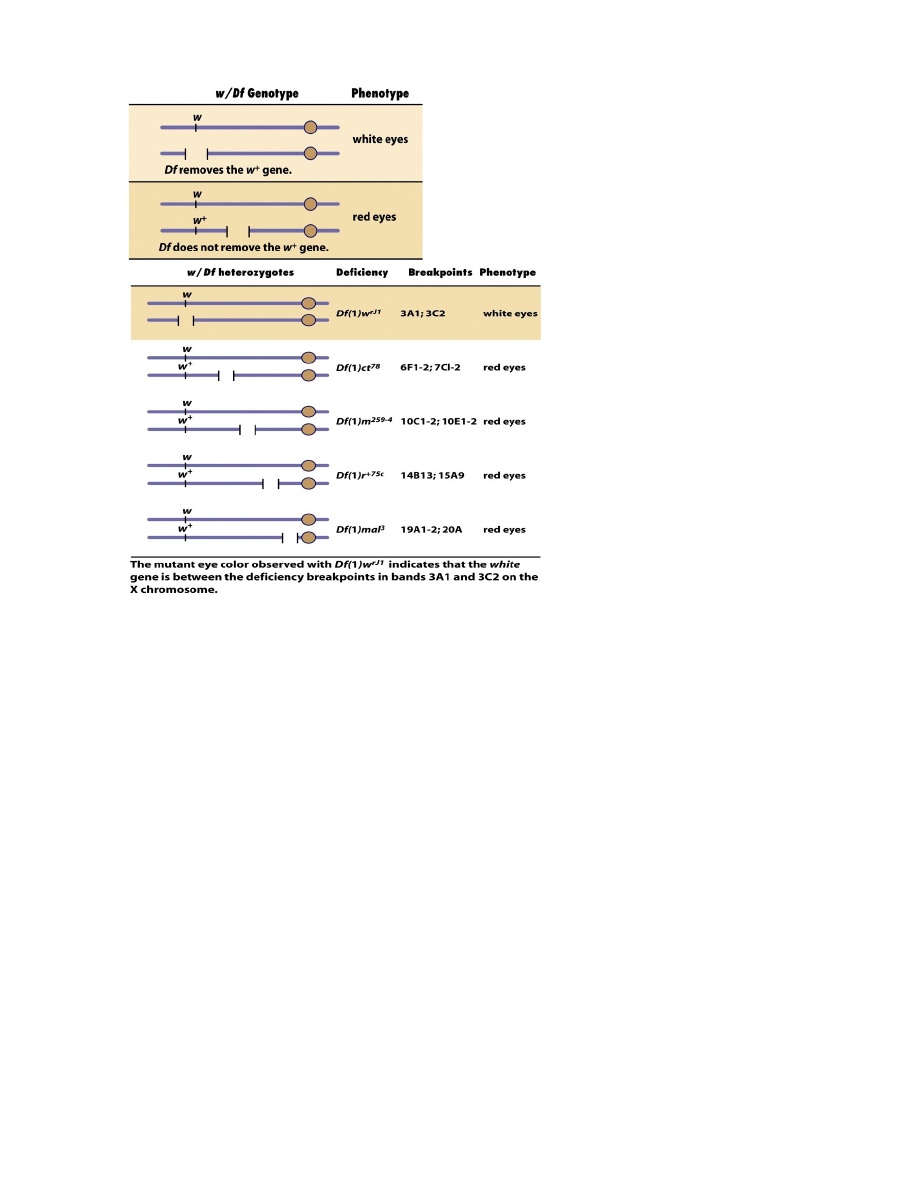
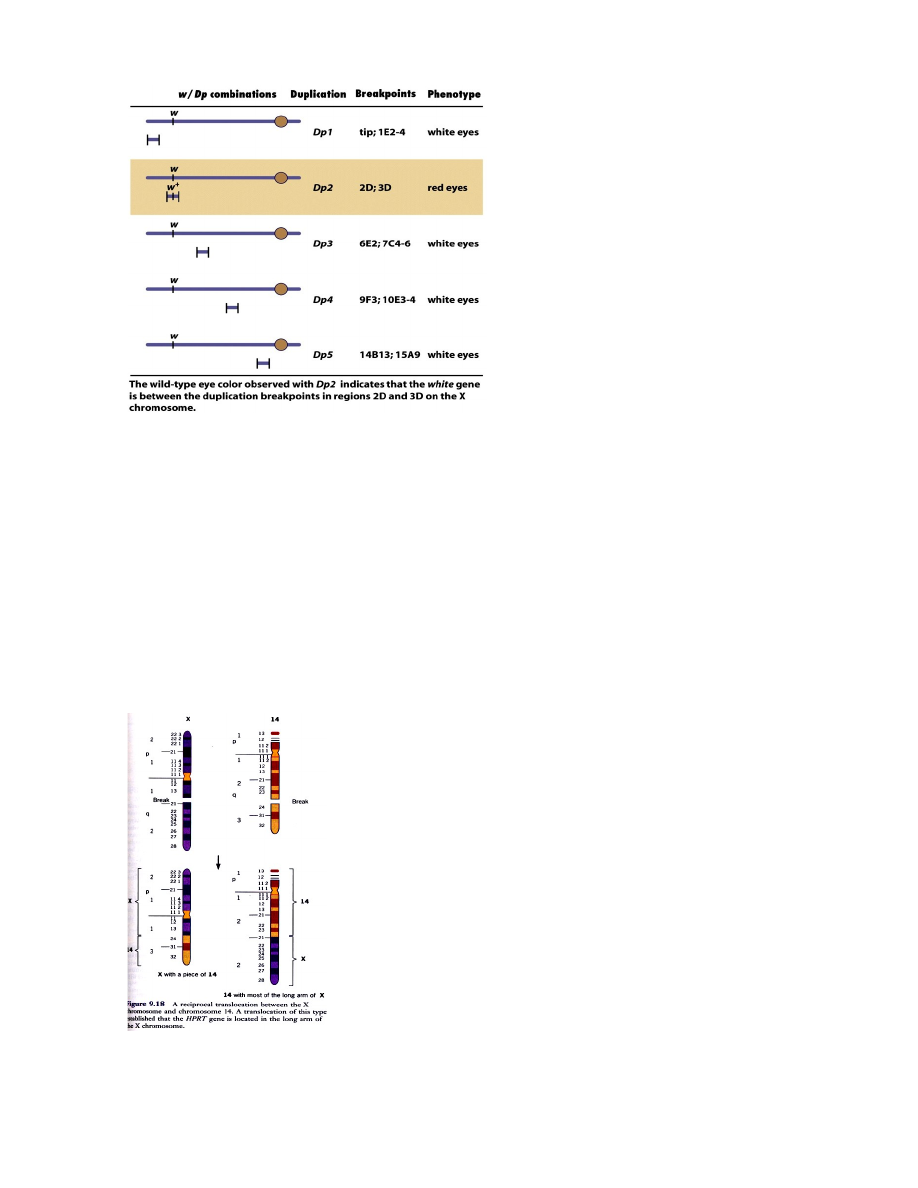




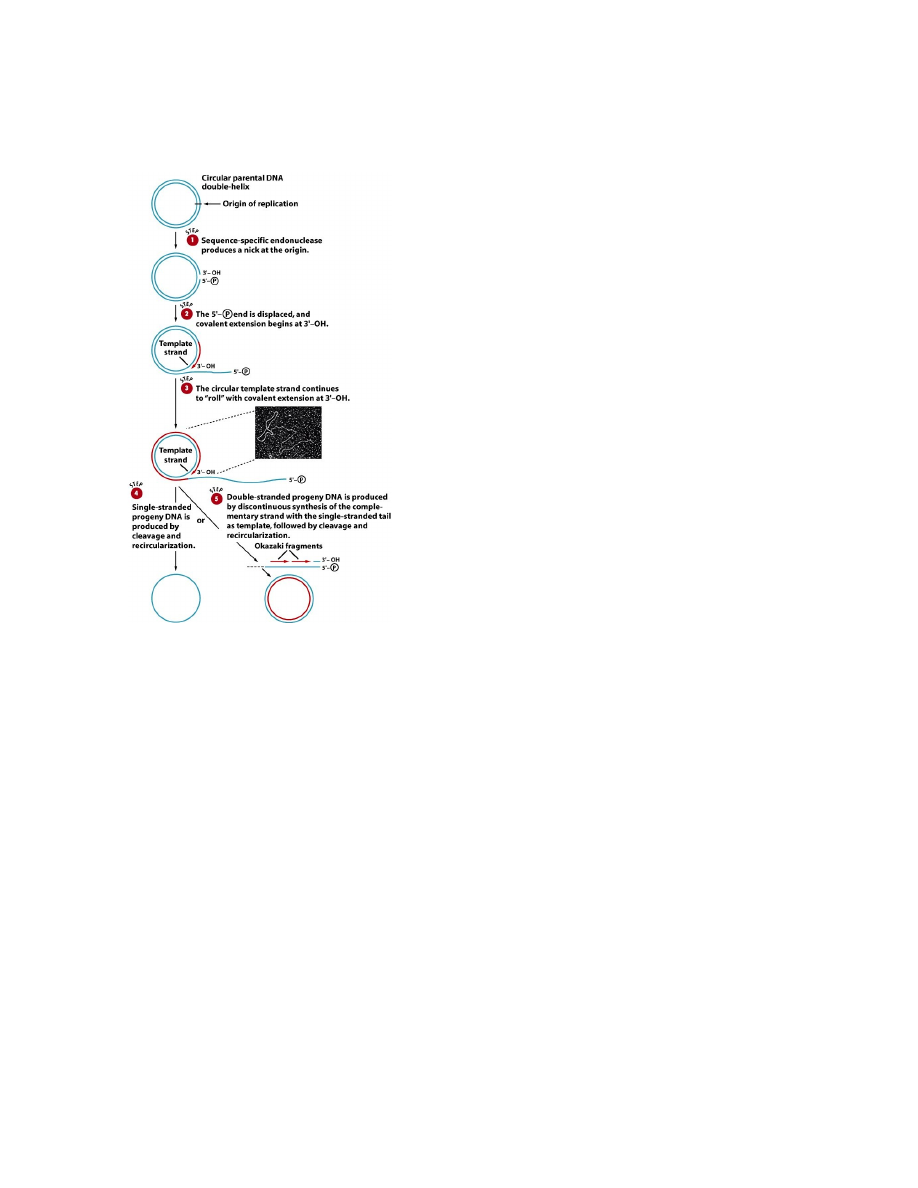












Kõik kommentaarid