Eesti Maaülikool
Veterinaarmeditsiini ja loomakasvatuse instituut
Maari Aru
TOIDUAINETES SPOORE MOODUSTAVAD MIKROOBID
Referaat õppeaines Toiduainete mikrobioloogia
Tartu 2014
SISUKORD
Sissejuhatus ..................................................................................................................................... 2
1
Spoori moodustumine ............................................................................................................. 3
1.1
Endospoorid ...................................................................................................................... 3
1.2
Struktuur ........................................................................................................................... 4
1.3
Asukoht ............................................................................................................................. 4
1.4
Germinatsioon
Bacillus spp näitel .................................................................................... 5
2
Bacillus liigid .......................................................................................................................... 7
2.1
Kliinilised ilmingud .......................................................................................................... 7
2.2
Bacillus anthracis ............................................................................................................. 7
2.3
Bacillus cereus .................................................................................................................. 9
3
Clostridium liigid .................................................................................................................. 11
3.1
Kliinilised ilmingud ........................................................................................................ 11
3.2
Clostridium perfringens .................................................................................................. 11
3.3
Clostridium tetani ........................................................................................................... 12
3.4
Clostridium botulinum .................................................................................................... 13
Kokkuvõte ..................................................................................................................................... 15
Kasutatud kirjandus ....................................................................................................................... 16
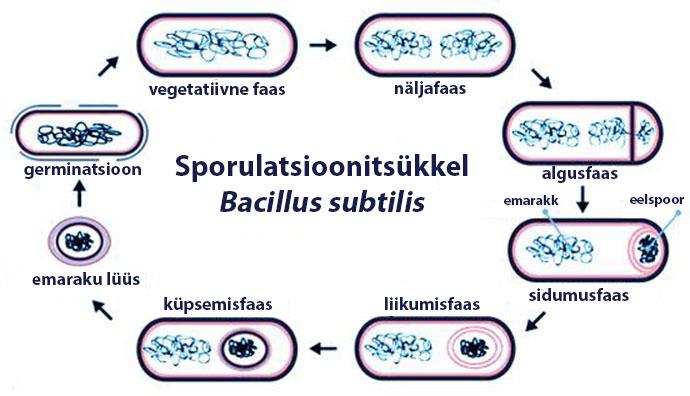
JOONISTE JA TABELITE LOETELU Joonis 1. Spoori
elutsükkel Bacillus subtilis’e näitel (Sporeweb, 2014) ........................................ 3
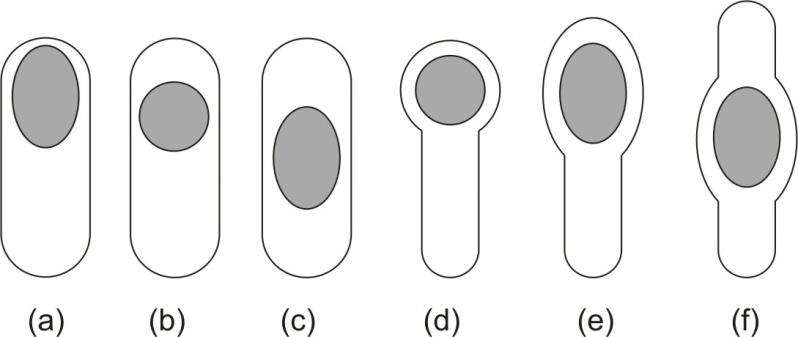
Joonis 2. Endospoori asukoht –
terminaalne (a, d, e), subterminaalne (b),
tsentraalne (c, f). (Tóth
et al., 2013) ...................................................................................................................................... 4
Joonis 3. Germinatsiooni
faasid (Setlow, 2014) ............................................................................. 6
Tabel 1.
B. cereus’e toidutekkeliste haiguste võrdlus (
Doyle et al., 2007) 10
SISSEJUHATUS Toiduainetes esinevad spoore moodustavad
bakterid on olulise tähtsusega, sest bakteri
moodustatud spoor on
resistentne kuumutamisele, külmutamisele, kemikaalidele ja teistele
ebasoodsatele tingimustele, mis toimuvad toidu töötlemise ning ettevalmistamise ajal. Nende
tingimuste juures vegetatiivne
rakk hävib, kuid
spoorid jäävad eluvõimelisteks ning vajavad
inaktiveerumiseks karmimaid tingimusi. Tähtsamad spoore moodustavad bakterid kuuluvad
perekonda
Bacillus. Nad on aeroobsed või fakultatiivselt anaeroobsed pulgakujulised mikroobid.
Bacillus liigid võivad põhjustada toidu riknemist või toidutekkelisi haiguseid. Teine oluline grupp
spoore moodustavaid baktereid on
Clostridium’i liigid. Enim tuntud
Clostridium’i põhjustatud
toidutekkeline haigus on
botulism . Kuigi igal aastal teatatakse väga üksikuid haigusjuhtumeid ning
inimesed tervistuvad õigeaegsel ravimisel, on haigus inimeste seas siiski väga kardetud (Cousin,
2003).
Piimatööstustes määravad
Bacillus’e ja
Clostridium’i liigid erinevate kuumtöödeldud
piimatoodete säilivusaja tingimusel, et töötlemisjärgne saastumine on madal (Giffel
et al., 2002).
Juba 20. sajandi alguses teavitati bakteriaalseid toidutekkelisi kõhulahtisusega kulgevaid
haiguseid. 1948. aastal tegi Norra
teadlane Steinar
Hauge kindlaks, et kohaliku toidutekkelise
haiguspuhangu põhjustajaks oli
Bacillus cereus. Erinevalt
Bacillus anthracis’est peeti
Bacillus perekonna liike mittepatogeenseteks kuni 1960.-ndate aastateni.
Clostridium’i perekonda on
patogeensusega rohkem seostatud. Need mikroorganismid on levinud kõikjal – õhus, pinnases,
vees, väljaheidetes, piimas ja teistes toitudes.
Bacillus liigid on fakultatiivselt anaeroobid, üldiselt
katalaas -positiivsed ja liikuvad.
Clostridium liigid on obligatoorselt anaeroobid ja katalaas-
negatiivsed ning nad võivad olla kas fermentatiivsed või proteolüütilised, või mõlemad korraga
(Baird-
Parker , 2000).
2
1 SPOORI MOODUSTUMINE 1.1 Endospoorid Kui keskkond ei ole arenguks soodne, on
Bacillus ja
Clostridium perekonna liikidel võime
moodustada spoore. Spoorid moodustatakse emaraku sees ja raku lüüsumise järel vabanevad nad
keskkonda. Joonisel 1 on toodud spoori elutsükkel
Bacillus subtilis’e näitel. Tegemist on
ellujäävate vormidega, mis on ekstreemselt resistentsed enamikele keskkonna stressifaktoritele.
Nende liikide spooridel on väike metaboolne aktiivsus. Vahetult pärast spoori moodustumist
esineb metaboolset aktiivsust vähesel määral, kui üldse (Setlow, 2014)
Joonis 1. Spoori elutsükkel
Bacillus subtilis’e näitel (Sporeweb, 2014)
Sporuleeruvad bakterid moodustavad kuumusele resistentseid endospoore, mis sisaldavad DPA-d
(
dipicolinic acid) ja on faas-kontrastsel mikroskoopial valgustmurdvad. Enamik uuringuid
sporulatsioonist, spooridest ja
spooride germinatsioonist on
viidud läbi kas aeroobsete batsillide
või anaeroobsete klostriididega (Doyle, 2007)
3
1.2 Struktuur Bakter toodab enda sees ühe endospoori. Mõnikord on spoor ümbritsetud õhukese
kattega , mida
nimetatakse eksospooriks ja mis katab spoori kesta. Spoori kest funktsioneerib sõelana, mis
elimineerib suured toksilised molekulid, näiteks lüsosüümid. Kest on resistentne
paljudele toksilistele molekulidele ning võib koosneda ensüümidest, mida on vaja germinatsiooniks.
(Wikipedia, 2014). Kesta all on
välismembraan , selle all peptidoglükaanist koosnev koor ja
seejärel iduraku sein ning sisemine
membraan (Setlow, 2014). Tuum sisaldab spoori
kromosomaalset DNA-d, mis on kapseldatud kromatiinilaadsetesse proteiinidesse, mis kaitseb
seda UV-kiirguse ja kuumuse eest. Tuum sisaldab ka tavalise raku struktuure, näiteks ribosoome
ja ensüüme, kuid ei ole metaboolselt aktiivne (Wikipedia, 2014).
Kuni 20% endospoori kuivainest moodustab tuumas asuv
kaltsium -dipikolinaat. Arvatakse, et see
aitab DNA-d stabiliseerida. Dipikoliinhape võib olla seotud spoori resistentsusega kuumusele.
Kaltsium võib aidata kaasa kuumaresistentsusele ja oksüdeerivatele teguritele. On isoleeritud
muteerunud spoore, mis on resistentsed kuumusele, kuid neil puudub dipikoliinhape. See võib
viidata, et resistentsust kujundavad ka teised tegurid (Wikipedia, 2014).
1.3 Asukoht Endospoori asukoht erineb bakteriliigiti ning on kasulik identifitseerimisel. Peamised endospoori
asukohad raku sees on: terminaalne, subterminaalne ja
tsentraalse asetusega. Terminaalsed
endospoorid on nähtavad raku poolustel, tsentraalsed on enam-vähem raku keskel. Aeg-ajalt on
leitud ka lateraalseid endospoore. Joonisel 2 on toodud endospooride asukohad. Terminaalsete
endospooridega on
Clostridium tetani, selle bakteri
endospoor on oma suuruse tõttu laiendanud
raku seina enda ümber.
Bacillus cereus’e endospoor on tsentraalse asetusega, subterminaalselt on
Bacillus subtilis’e endospoor (Wikipedia, 2014).
Joonis 2. Endospoori asukoht – terminaalne (a, d, e), subterminaalne (b), tsentraalne (c, f). (Tóth
et al.,
2013)
4
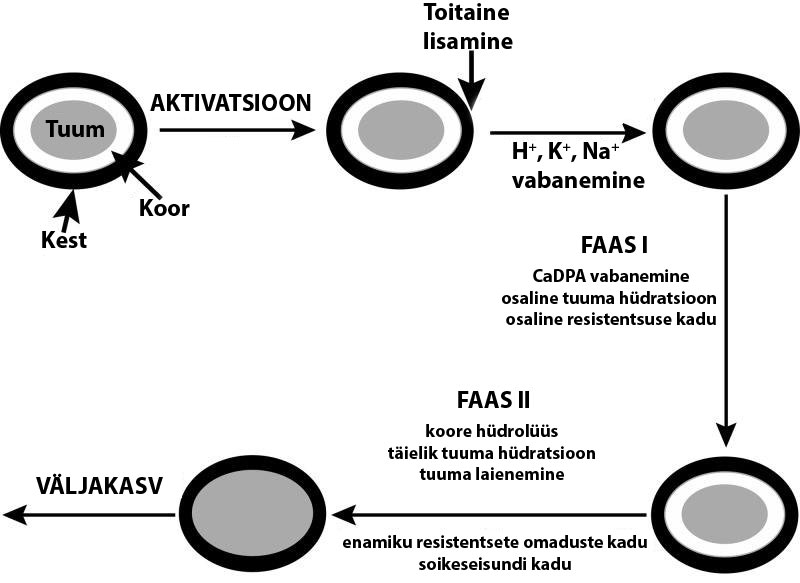
1.4 Germinatsioon Bacillus spp näitel Spoorid võivad jääda soikeseisundisse aastateks, aga kui saavad õige stiimuli (
germinant), algab
germinatsioon ning nad
kaotavad resistentsuse. Germinatsiooni protsessile järgneb väljakasv, mis
muudab spoori kasvavaks rakuks. Spoori germinatsiooni
kutsuvad esile mitmed tegurid, mõned
neist, näiteks CaDPA (Ca2+-
ioonid 1:1 suhtes dipikoliinhappega) ning katioonsed surfaktandid, on
olulised ainult laboratoorselt. Toiduainetööstuse suurenenud huvi on suunatud
kõrgele rõhule –
tuhanded atmosfäärid, eriti kui kombineerida kõrgete temperatuuridega. Looduses on tõenäolisem,
et spoori germinatsiooni kutsuvad esile kindlad
toitained (Setlow, 2014).
Pärast toitelise stiimuli saamist toimub viiteperiood (
lag period ), mis võib populatsiooni
individuaalsetel spooridel kesta mõnest minutist kuni 24 tunnini (Setlow, 2014).
Oletatakse, et enamik muutusi toimub sisemembraani läbilaskmisvõimes ja võimalik, et ka
struktuuris, toimub monovalentsete katioonide H+, K+, Na+ ja seejärel CaDPA
vabanemine .
Enamiku CaDPA vabanemine toimub mõne minuti jooksul ja kõige tõenäolisemalt läbi spoor-
spetsiifiliste SpoVa proteiinide moodustatud
kanalite . Need
proteiinid on kodeeritud ühe või
enama operoni poolt kõikidel spoore moodustavatel batsillidel ja klostriididel. CaDPA
vabanemine lõpetab germinatsiooni I faasi ning kutsub esile II alguse. Koore-lüütilised
ensüümid lagundavad peptidoglükaankoore. Koore lagundamisel saab iduraku sein laieneda ning tuum
suureneda , võttes endasse vett. Selle tulemusena saab läbi germinatsiooni II etapp (Joonis 3).
Tuuma
veesisaldus tõuseb kuni 80%, mis on võrdne kasvavate rakkudega. Veesisaldus võimaldab
tuumal metaboliseeruda. Algab väljakasv – toimub makromolekulaarsüntees, mis muudab
germineerunud spoori kasvavaks rakuks (Setlow, 2014).
5
Joonis 3. Germinatsiooni faasid (Setlow, 2014)
6
2 BACILLUS LIIGID 2.1 Kliinilised ilmingud Perekonda
Bacillus kuulub peaaegu 250 liiki. Meditsiiniliselt olulised on õnneks neist ainult
vähesed.
Bacillus anthracis põhjustab haigestumist antraksisse ning on
Bacillus perekonna
tähtsaim liige. Seda liiki peetakse kõige kardetumaks bioloogilises sõjas. 2001. aastal US
Postal Service kaudu levinud antraksi spoorid tõid uuesti esile selle bakteriga seotud
potentsiaalsed ohud.
Teine kliiniliselt tähtis liik on
Bacillus cereus, mis põhjustab gastroenteriiti, traumaatilisi
silmainfektsioone, kateetriga seotud sepsist ning harva tõsist pneumooniat (
Murray et al., 2012).
Bacillus cereus põhjustab kahte erinevat toidumürgistusega seotud sündroomi: äkktekkeline
emeetiline
sündroom koos iivelduse ja oksendamisega, ning
aeglase kuluga kõhulahtisuse
sündroom. Teised
Bacillus liigid, eriti
B. cereus, vähem ka
B. subtilis ja B. licheniformis, on
perioodiliselt seostatud bakterieemia/septitseemia, endokardiidi, meningiidi ja
haavade , kõrvade,
silmade, respiratoor-, urinaar- ning
seedetrakti infektsioonidega (Turnbull, 1996).
2.2 Bacillus anthracis B. anthracis bakteril on omadus moodustada
pikki pulgakujulisi ahelaid. Kultuurina on see
mitteliikuv ja mittehemolüütiline.
Kolooniad on kareda ja
ebaühtlase pinnaga. Bakteril on
polüpeptiidne
kapsel ainsa antigeeni tüübiga, millel on antifagotsütaarsed omadused.
Bacillus anthracis’e endospoorid on ekstreemselt tugevad ja võivad säilida keskkonnas kümneid aastaid.
Bakter toodab ka tõhusat eksotoksiini kompleksi, mis sisaldab kahte ensüümi - ödeemi
faktorit (
edema factor – EF) ja letaalset faktorit (
lethal factor – LF). Kui kumbki neist
seondub protektiivse
antigeeniga (PA), käitub PA kui translokaas, moodustades peremeesraku pinnale poorilaadse koha
ning
kompleks saab siseneda rakku. (Ray
et al., 2014).
Antraks on eelkõige herbivooride, näiteks hobuste,
lammaste ja veiste haigus.
Nakatumine toimub
karjamaalt
Bacillus anthracis’e spooride kaudu. Inimesed nakatuvad kontaktil haige
loomaga või
loomse toodangu kaudu, kui spoorid läbivad nahakahjustuse, süüakse toiduga või hingatakse sisse.
1920-ndatel aastatel esines USA-s rohkem kui 100 haigestumist aastas. Haigus levis
farmerite ,
veterinaaride ja lihakäitlejate seas, kuid loomade antraksi vaoshoidmine arenenud riikides on
muutnud inimjuhtumid harvaesinevateks. Põhja-Ameerikas esineb antraks üksikute endeemiliste
kolletena ning nakatumine on toimunud looduslikult. Teiseks allikaks on loomadest valmistatud
tooted – vill, nahk või
kondijahu väetis, mis on imporditud riigist, kus loomadel esinev antraks on
endeemiline (Ray
et al., 2014). Ligikaudu 95% inimeste antraksinakkused toimuvad
Bacillus 7
anthracis’e spooridega naha kaudu kas kontamineerunud pinnase või nakatunud loomade
toodangu kaudu (Murray
et al., 2012).
Nakatumine allaneelamise kaudu on inimeste seas väga harv, kuid tavaline
nakatumise tee
herbivooridel. Kuna organism moodustab vastupidavaid spoore, säilib ta nakatumisvõimelisena
pinnases või
loomses toodangus mitmeid aastaid (Murray
et al., 2012).
Inhalatsiooniantraksit on ajalooliselt kutsutud villa sorteerijate haiguseks, sest enamik
inimnakkuseid tulenes spooride sisse hingamisel
kitse karvade töötlemisel. Hetkel ei ole see
levinud nakatumise tee, kuid on kõige tõenäolisem võimalus nakatumiseks bioloogilises sõjas.
Usutakse, et bakteri nakatumisdoos on madal. Inimeselt-inimesele ülekannet ei esine, sest
bakteriaalne paljunemine toimub pigem keskseinandi lümfisõlmedes kui bronhopulmonaarkoes.
(Murray
et al., 2012).
Tüüpiline kutaanne antraks algab nakatumise kohas nahal
valutu paapuli tekkega. See
progresseerub haavandiks, mida ümbritsevad vesiikulid. Seejärel muutub nekrootiliseks koldeks.
(Murray
et al., 2012). Seotud süsteemsed sümptomid on tavaliselt kerged ning vigastus
paraneb tavaliselt väga aeglaselt pärast kooriku eemaldumist. Haigus progresseerub harvem massiivse
lokaalse ödeemi, tokseemia ja bakterieemiaga (Ray
et al., 2014).
Gastrointestinaalse antraksi sümptomid sõltuvad nakatumise asukohast. Kui bakterid tungivad
seedetrakti ülemisse
osasse , moodustuvad
haavandid suu või söögitoru pinnale, mis viib
regionaalse lümfadenopaatia, ödeemi ja sepsiseni. Kui organismid jõuavad pimesoolde või
niudesoole lõppu, järgneb
iiveldus ,
oksendamine ja ebamugavustunne, mis progresseerub kiiresti
süsteemseks haiguseks. Gastrointestinaalse antraksi
suremus on ligikaudu 100% (Murray
et al.,
2012).
Pulmonaarne antraks on seotud pika latentse perioodiga – 2 kuud või rohkem, mille jooksul on
nakatunud inimene asümptomaatiline. Spoorid võivad jääda latentseteks ninakäikudes või jõuda
alumistesse hingamisteedesse, kus alveolaarmakrofaagid nad alla neelavad ja transpordivad
keskseinandi lümfisõlmedesse. Lõplik kliiniline pilt on
mittespetsiifiline – lihasnõrkus,
palavik ,
köha ja ebamugavustunne. Haiguse teine faas on dramaatilisem, palavik tõuseb järsult,
keskseinandi
lümfisõlmed on suurenenud, järgneb
respiratoorne defitsiit ja
sepsis . Kopsupõletikku
tavaliselt ei teki. Peaaegu kõik
juhud lõpevad šoki ja surmaga 3 päeva jooksul pärast sümptomite
teket. (Murray
et al., 2012).
Antimikroobsel ravil on väike efekt kutaanse antraksi korral, kuid ennetab haiguse levikut.
Peaaegu kõik
Bacillus anthracis’e tüved on penitsilliinile, doksütsükliinile ja tsiproflokatsiinile
8
tundlikud. Kuigi
penitsilliin on olnud kaua aega kasutusel kõigi antraksi vormide raviks, jõuti
2001. aasta puhangu ajal järeldusele, et esimese valikuna tuleks eelistada doksütsükliini või
tsiproflokatsiini. (Ray
et al., 2014).
Kõige olulisem võimalus ennetamiseks on loomadel esineva antraks likvideerimine ja limiteerida
importimist endeemilistest piirkondadest. Kasutusel on
elusvaktsiinid atenueeritud tüvedest (Ray
et al., 2014).
2.3 Bacillus cereus Bacillus cereus on gram+ fakultatiivselt aeroobne spoore
moodustav pulgakujuline bakter. On
teada 6
Bacillus-e eri liiki:
Bacillus anthracis,
B. cereus,
B. mycoides,
B. pseudomycoides,
B. thuringiensis, ja
B. weihenstephanensis. Mitmed tüved on võimelised produtseerima
toksiine ning
põhjustama toidumürgistusi.
B. cereus on nõrk konkurentsmikrofloora esindaja, kuid kasvab hästi
pärast kuumutamist ja jahutamist (24 h)
0.5 – 5 h
Haiguse kestus 12 – 24 h (vahel mitu päeva)
6 – 24 h
Sümptomid Kõhuvalu , vesine diarröa,
Iiveldus, oksendamine,
vahel iiveldus
ebamugavustunne
Toidud, mis on sageli Lihatooted,
supid ,
juurviljad ,
Praetud või küpsetatud
riis ,
saastunud pudingid,
kastmed ja piim või pasta,
nuudlid või muud
piimatooted
küpsetised
Bacillus cereus’e põhjustatud gastroenteriidi kulg on lühike ning seepärast on adekvaatne ka
sümptomaatiline ravi. Toidumürgistust saab ennetada toidu söömisega kohe pärast selle
kuumutamist ning söömata toidu piisavalt madalal temperatuuril külmutamisega (Murray
et al,
2012).
Adekvaatsel temperatuuril toidu valmistamine hävitab enamuse toidutekkelistest patogeenidest,
sealhulgas
Bacillus cereus’e vegetatiivsed
rakud , kuid mitte spoore. Spoorid kaotavad
kuumaresistentsuse happelistes tingimustes (Cookingham
et al., 2004).
Soovitused
B. cereus’e hävitamiseks (Cookingham
et al., 2004):
aurutamine rõhu all, röstimine ja fritüürimine hävitab vegetatiivsed rakud ja spoorid;
kõhulahtisust põhjustav
toksiin inaktiveerub kuumutades 56 kraadi juures 5 minuti jooksul;
emeetilist toksiini sisaldavaid toite tuleb kuumutada üle 90 minuti 126 kraadi juures;
sooje toite tuleb hoida üle 60, jahutatud toite alla 4 kraadi juures.
10
3 CLOSTRIDIUM LIIGID 3.1 Kliinilised ilmingud Clostridium perekonda kuuluvad bakterid on levinud kõikjal – pinnases, vees,
kanalisatsioonis ning on inimeste ja loomade seedetrakti normaalmikrofloora asukateks. Enamik klostriide on
kahjutud
saprofüüdid , kuid mõned on hästi tuntud patogeenid. Ajaloost on teada hästi kirjeldatud
haigusjuhtumeid, näiteks
teetanus (
Clostridium tetani), botulism (
Clostridium botulinum, Clostridium baratii, Clostridium butyricum), müonekroos ehk gaasi gangreen (
C. perfringens, C. novyi, C. septicum, C. histolyticum) ja
kõhulahtisus ning jämesoolepõletik (
C. perfringens, C. difficile ). Valdav enamus tänapäeval levivatest nakkustest on naha ja pehmekoe haigused,
toidumürgistused
ning
antibiootikumidega
seotud
kõhulahtisus-
koliit .
Klostriidide
märkimisväärset võimet põhjustada haigestumist
seostatakse nende võimega säilida keerulistes
keskkonnatingimustes spooride moodustamisega; kiire kasvuga toitaineliselt soodsates
tingimustes; mitmete histolüütiliste, entero – ja neurotoksiinide tootmisega (Murray
et al, 2012).
3.2 Clostridium perfringens Clostridium perfringens on anaeroobne, kuid aerotolerantne gram-positiivne spoore moodustav
pulgakujuline bakter, mis toodab enterotoksiine. Bakter on suhteliselt külmatolerantne ja spoorid
kõrgele temperatuurile resistentsed. Mitte-
patogeenne C. perfringens on keskkonnas laialt levinud
ja seda on sageli isoleeritud inimeste ning loomade soolestikust. Bakteril on mitmeid isotüüpe,
tüüp A sisaldab peaaegu alati
cpe geeni – see on enterotoksiini geen, mis põhjustab
toidumürgistust. Tüübid B, C, D ja E võivad samuti vahel seda geeni
sisaldada .(Lampel, 2013).
Enamikel juhtudel põhjustab haigestumist liiga madalal temperatuuril
kuumutatud toitude
söömine . Kuumutatud toidus on sageli alles väikeses koguses baktereid. Valmistatud toidu
jahutamise ja säilitamise käigus võivad bakterid paljuneda koguseni, mis põhjustab mürgistust
(Lampel, 2013). Toidu uuesti
kuumutamine 74 kraadini hävitab kuumatundliku toksiini (Murray
et al., 2012).
Clostridium perfringens’i põhjustatud toidutekkeline haigestumine võib esineda kahe erineva
vormina. Gastroenteriidi vorm on väga tavaline, kulgeb kergelt ja on iselimiteeruv. Vesine
kõhulahtisus ja
krambid ilmnevad 8 – 16 h jooksul pärast söömist. Paljudel mööduvad sümptomid
iseenesest 24 h jooksul, kuid väga noortel, vanadel või immuunpuudulikkusega inimestel võib
haigus kesta kuni nädal. Sellised juhud vajavad sümptomaatilist ravi ja vedelikteraapiat. Teine
11
vorm nekrootiline enteriit on palju tõsisem ja sageli fataalne, kuid esineb harva. Sümptomiteks on
valu ja puhitus kõhus, kõhulahtisus ja oksendamine (Lampel, 2013).
Mõlemad haiguse vormid tulenevad suure koguse
C. perfringens bakterite allaneelamisel. On
oluline, et nad paljunevad palju kiiremini, kui enamik baktereid. Võrreldes teiste
bakteritega ,
saavutab
C. perfringens kiiremini patogeense taseme toidus, mida ei säilitada madalal
temperatuuril (Lampel, 2013).
3.3 Clostridium tetani Clostridium tetani on liikuv spoore moodustav pulgakujuline bakter. Erinevalt
C. perfringens-ist
on teda raske kasvatada, sest ta on tundlik hapniku toksilisusele ning moodustab söötmele pigem
kile, kui eraldiseisvad kolooniad (Murray
et al., 2012).
C. tetani on kõikjal keskkonnas. Vegetatiivsed vormid on hapnikule tundlikud, kuid nad
sporuleeruvad kergelt ning võivad jääda keskkonda pikaks ajaks. USA-s on haigus suhteliselt
haruldane , sest inimesed saavad immuunsuse vaktsineerides. Levinud on rohkem vanematel
inimestel, kelle immuunsus on langenud. Arengumaades esineb teetanust sagedamini, sest puudub
võimalus
vaktsineerida ja adekvaatse meditsiinilise abi järele. Arvatakse, maailmas esineb üle 1
miljoni haigusjuhu ning suremuse määr on 30 – 50%. Vähemalt pooled surmadest moodustavad
nakatunud vastsündinud (Murray
et al., 2012).
Inkubatsiooniperiood
varieerub mõnest päevast kuni nädalateni. See on otseselt seotud esmase
haavainfektsiooni kaugusest kesknärvisüsteemist.
Generaliseerunud teetanus on haiguse kõige
tavalisem vorm. See algab mälumislihaste kangestumisest, esineb suurenenud ilavoolu,
higistamist, ärrituvust ja
seljalihaste spasme. Haiguse tõsisema kulu korral on haaratud ka
autonoomne närvisüsteem – esineb südame arütmiaid, dehüdratatsiooni, tugevat higistamist ja
muutuseid vererõhus. Teine
teetanuse vorm on
lokaalne , mis püsib haavainfektsiooni piirkonna
lihastes. Vastsündinute teetanus on seotud nabaväädi infektsiooniga ning muutub
generaliseerunud vormiks. Vastsündinute
suremus ületab 90%, ellujääjatel esineb arengudefekte
(Murray
et al., 2012).
Teetanuse
ravis puhastatakse
haav , mille kaudu toimus nakatumine. Bakterite elimineerimiseks ja
toksiinide toime vähendamiseks kasutatakse penitsilliini või metronidasooli.
Sidumata toksiinide
neutraliseerimiseks kasutatakse passiivset immuniseerimist.
Vaktsineeritakse teetanuse
toksoidiga, sest nakatumine ei ole võrdne immuunsusega (Murray
et al., 2012).
12
3.4 Clostridium botulinum C. botulinum on samuti anaeroobne spoore moodustav pulgakujuline bakter, mis produtseerib
tugevat neurotoksiini. Tema spoorid on kõrgele temperatuurile resistentsed ja püsivad
eluvõimelisena toitudes, mida on minimaalselt või ebaühtlaselt töödeldud. Toksiini antigeense
ülesehituse alusel on teada
seitse erinevat botulismi põhjustavat bakteri tüve (A, B, C, D, E, F ja
G). Tüübid A, B, E ja F põhjustavad inimeste botulismi. Tüübid C ja D põhjustavad loomade ning
C ja E lindude haigestumist. Tüübi G haiguspuhanguid ei ole teatatud (Lampel, 2013). Kuigi
toksiinid on erinevad, on nad
struktuurilt sarnased ja omavad sarnast bioloogilist efekti. Lähtuvalt
füsioloogilistest eripäradest jaotatakse liigid veel nelja erinevasse gruppi (I – IV). See on kooskõlas
nukleiinhapete hübridisatsiooni ja 16S ribosoomi sekveneerimise uuringutega (Roasto
et al, 2011).
Organism ja selle spoorid on looduses laialt levinud. Neid on leitud nii kultiveeritud kui metsade
pinnasest, veekogude põhjasettest, kalade ja imetajate seedetraktist ning kalade ja koorikloomade
lõpustelt ja sisikonnast (Lampel, 2013).
Botulism on raskelt kulgev ja mõnikord fataalne haigus, mida põhjustab
C. botulinum bakteri poolt
toodetav toksiin. Nakatumisele järgneb lihaste, ka hingamislihaste lõtv
paralüüs . On teada 3
erinevat botulismi vormi: toidutekkeline,
väikelaste , mis on samuti toidutekkeline. Kolmas tüüp
on haava botulism, mida antud kontekstis ei käsitle (Lampel, 2013).
Toidutekkeline botulism tekib kõige sagedamini ebaühtlaselt või ebaadekvaatselt valmistatud
kodu-preservide söömisel. Kodus konserveeritud, harva ka tööstuslikult valmistatud toidud on
olnud seotud botulismipuhangutega USA-s. Nakatumise sagedus on küll madal, kuid kõrge
suremuse tõttu, ilma abi saamata, on murettekitav (Lampel, 2013). Nakatunud inimesed muutuvad
nõrgaks ja uimaseks 1 – 3 päeva jooksul pärast saastunud toidu tarbimist. Esimeste sümptomite
hulka kuuluvad hägune nägemine, fikseeritud
laienenud pupillid, suukuivus, kõhukinnisus ja
kõhuvalu. Järgneb
bilateraalne lihaste nõrkus (Murray
et al., 2012).
C. botulinum’i toksiin
põhjustab paralüüsi, blokeerides
motoorsed närvilõpmed
neuromuskulaarses ühenduskohas.
Paralüüs progresseerub sümmeetriliselt ülevalt alla, algab tavaliselt silmadest ja näost, järgneb
kõri , rind ja jäsemed. Kui häirub diafragma ja rinnalihaste töö, pidurdub hingamine ning ilma
sekkumiseta järgneb surm lämbumise tõttu (Lampel, 2013). Täielik
taastumine võtab aega mitu
kuud kuni aastaid, kuni haaratud närvilõpmed kasvavad uuesti (Murray
et al., 2012).
Väikelaste botulismi korral toimub spooridega nakatumine allaneelamisel. Seedetraktis hakkavad
spoorid arenema ning
tootma toksiine. (Lampel, 2013). Täiskasvanud inimestel ei püsi väiksed
kogused C. botulinum’i bakterid seedetraktis eluvõimelisena konkureeriva
mikrofloora tõttu
(Murray
et al., 2012).
13
Nakatunud inimestel tuleb anda ventilatoorset abi. Seedetraktist tuleb
elimineerida nakatumist
põhjustanud bakterid mao lavaaži kaudu ning manustada metronidasooli või penitsilliini.
Kasutatakse kolmevalentset antitoksiini (A, B ja E toksiini vastu), et inaktiveerida vereringes
sidumata toksiinid. Pärast haiguse läbipõdemist ei teki kaitsvaid antikehi ning on oht
mitmekordseks nakatumiseks (Murray
et al., 2012).
Haigestumist saab ennetada toidus spooride hävitamisega (praktilistel kaalutlustel võimatu),
ennetades spooride germinatsiooni
hoides toitu happelise pH juures või säilitada vähemalt 4 kraadi
juures. Kõik botulismi toksiinid inaktiveeruvad kuumtöötlusel 60 – 100 kraadi juures vähemalt 10
minuti jooksul. Väikelaste botulism on seotud mee tarbimisega, mis sisaldab
Clostridium botulinum’i spoore, seega alla 1-aastastele lastele ei tohiks mett sööta (Murray
et al., 2012).
Põhilised tegurid, mis mõjutavad
C. botulinum-i kasvu, on temperatuur, pH, vee aktiivsus,
redokspotentsiaal, lisatud säilitusained ja teised mikroorganismid.
Külmutamine toimib kasvu
pidurdavalt. Madalaid temperatuur on I grupile 10 ja teisele 3 kraadi. Need normid on
rakendatavad ainult vähestele tüvedele ja sõltuvad kasvukeskkonnast. Madalaim pH tase, mille
juures toimub veel I grupi tüvede elutegevus, on 4,6 ja II
grupil 5,0. Seega on paljud puu- ja
juurviljad inhibiitoriteks, kuna nende pH on piisavalt happeline. Happetolerantsed
mikroorganismid, nagu pärmid ja
hallitused , tõstavad oma
elukeskkonnas pH sellise tasemeni, mis
võimaldab
C. botulinum’il areneda. Optimaalsetes tingimustes on
C. botulinum’i kasvu pidurdava
soolvee kontsentratsioon I grupi tüvedele umbes 10% ja II grupi tüvedele 5%. Töödeldud
toiduainetele lisatakse nitriteid ja nitraate, et inhibeerida
C. botulinum’i elutegevust. (Roasto
et al.,
2011).
14
KOKKUVÕTE Toiduainetes spoore moodustavatest bakteritest on levinuimad
Bacillus’e
ja
Clostridium’i
perekonnad. Toidutekkelisi haiguseid põhjustavad sageli
nendest Bacillus cereus’e
ja
Clostridium perfringens’i
, C. botulinum’i liigid. Need liigid suudavad ebasoodsas elukeskkonnas moodustada
spoore, mis on resistentsed kuumutamisele, külmutamisele ja desinfitseerimisele. Vastavate
võtetega saab hävitada
vegetatiivse bakteriraku, kuid spoor jääb eluvõimeliseks. Soodsate
tingimuste või toitainelise stiimuli saamisel algab spoori germinatsioon ning ta areneb
metaboliseeruvaks bakterirakuks.
Inimeste haigestumist põhjustav mikrofloora satub toitu enamasti algtootmise ja –töötlemise ning
toiduainete käitlemisel: põllult
koristamisel , loomade/lindude farmis
kasvatamisel ja algtootmisel,
toodete pakendamisel ja ladustamisel ning
turustamisel (Roasto
et al,. 2011).
Spoore moodustavate bakterite tähtsus seisneb selles, et neid kõiki leidub pinnases. Seega on nad
otseselt seotud toiduainete saastumisega. Töödeldes või säilitades toitu
valedel meetmetel võivad
spoorid areneda vegetatiivseteks rakkudeks ja toota inimese tervisele ohtlikke toksiine. Oluline on
õigetel
temperatuuridel toitu kuumutada, kiirelt jahutada ja säilitada 4 kraadi juures.
Toiduainetööstused peaksid jälgima vee aktiivsust ja pH-d, millega on võimalik pidurdada
toksiinide moodustumist toidus.
15
KASUTATUD KIRJANDUS Baird-Parker, T. C., Gould, G. W., Lund, B. M. The Microbiological
Safety and
Quality of Food.
Volume I. Aspen Publishers, Inc. Gaithersburg, Maryland. 2000. lk 1401
Cookingham, T., Goodrich, R. M., Parish, M. E., Schneider K. R. Preventing Foodborne
Illness :
Bacillus cereus and
Bacillus anthracis.
University of Florida, USA. 2004
Cousin, M. A. Sporeforming Bacteria in
Foods . Department of Food Science, Purdue University,
West Layfayette, IN. 2003
Doyle, M. P., Beuchat, L. R. Food Microbiology: Fundamentals and Frontiers. 3rd
Edition . ASM
Press, Washington, 2007. lk 35-37; 445-452.
Giffel, M. C., Wagendorp, A., Herrewegh, A., Driehuis, F. Bacterial spores in silage and raw
milk .
Antonie van
Leeuwenhoek , 2002, 81, lk 625.
Lampel, K. A. Bad Bug Book, Foodborne Pathogenic Microorganisms and Natural Toxins. 2nd
Edition. FDA, USA. 2013, lk 83 – 86; 109 – 110.
Murray, P., Pfaller, M.,
Rosenthal , K.
Medical Microbiology. 7th Edition, Saunders, Elsevier.
2012, lk 209 – 215; 327 – 337.
Ray, C. G.,
Ryan , K. J. Sherris Medical Microbiology. 6th Edition. McGraw-
Hill Education, USA.
2014, lk 483 – 487.
Roasto, M., Breivel, M., Dreimann, P. „Toiduainetööstuse tootmishügieen“. Halo kirjastus, Tartu.
2001, lk 155; 279 – 285.
Setlow, P. Germination of Spores of
Bacillus Species : What We
Know and Do Not Know. Journal
of Bacteriology. Apr 2014, 196(7), lk
1297 – 1305.
Sporeweb -
http://ezines.tifn.nl/intouch/october2013/eijlander_sporeweb.html (23.11.14)
Tóth, E. M., Márialigeti, K.
Practical Microbiology, 2013 –
http://elte.prompt.hu/sites/default/files/tananyagok/microbiology/index.html (28.11.14)
Turnbull, P. C. B. Medical Microbiology. 4th edition. Galveston, University of Texas Medical
Branch at Galveston; 1996. -
http://www.ncbi.nlm.nih.gov/books/NBK7699/ (29.11.14)
Wikipedia.org -
http://en.wikipedia.org/wiki/Endospore (22.11.14)
16


































Kõik kommentaarid