BIOKEEMIA II TESTIKS | Mihkel
Heinmaa YAGB22 | TTÜ kevad 2010
XI SÜSIVESIKUD
1. Süsivesikuteks nim biomolekule, mis koosnevad vaid süsinikust, vesinikust ja hapnikust. Süsivesikuteks loetakse polühüdroksüaldehüüde ja ketoone või aineid, mis annavad hüdrolüüsi käigus vastavaid ühendeid. Nimetus tuleb empiirilisest
valemist Cn(H2O)n Süsivesikute bioloogiline roll. Väga mitmekesine ja looduses laialt levinud orgaaniliste molekulide klass; päikese energia salvestatakse fotosünteetiliste organismide poolt süsivesikutesse; paljude biomolekulide eelühendid; struktuuriline roll; molekulaarsed ja
rakk -rakk äratundmismehhanismid. Süsivesikute
multifunktsionaalsus põhineb struktuuri iseärasustel: asümmeetriliste tsentrite olemasolu; esinemine nii
lineaarses kui tsüklilises vormis; võime moodustada polümeere glükosiidsidemete abil; võime moodustada palju vesiniksidemeid nii keskkonna kui naabermolekulidega.
Nomenklatuur :
Monosahhariidid e monoosid.
Lihtsuhkrud , ei saa lõhustada lihtsamateks suhkruteks (C aatomeid 3...7) C-aatomite arvu järgi: trioosid (C3); tetroosid (C4);
pentoosid (C5);
heksoosid (C6) jne. Heksoosid on looduses levinuimad. Funktsionaalse rühma järgi:
Aldoosid sisaldavad aldehüüdrühma (nimetused lõpevad oos)
Ketoosid sisaldavad ketorühma. (vastava aldoosi nimetusele lisandum ul, ntks
riboos > ribuloos) Tsükli ehituse järgi Püranoosid (sarnasus püraani tsükliga) Furanoosid (sarnasus furaani tsükliga).
2.
Suhkrute ketorühma ja hüdroksüülrühm võivad molekulisiseselt reageerides anda tsüklilise hemiatsetaale või hemiketaalse. D/L
klassifikatsioon viitab suurima numbriga asümeetrilise tsentri klassifikatsioonile. Võrdleb vaadeldava stereotsentri konfiguratsiooni D/L-glütseraalaldehüüdi konfiguratsiooniga. Looduses domineerivad D-
suhkrud . S/R klassifikatsioon on absoluutne.
Kiraalne tsenter on asetatud nii, et kõige noorem asendaja oleks stereogeense tsentri taga. Kui ülejäänud asendajad saab ühendada vanuse alanemise järjekorras kellaosutis suunas, siis on R-konformatsioon, vastupidi S-konf. Anomeerid on monosahhariidide
isomeerid , mis erinevad teineteisesst ainult anomeerse süsiniku konfiguratsiooni poolest. Anomeersed süsiniku
aatomid on karbonüülsed süsiniku aatomid, mis hemiatsetaalsete/hemiketaalsete tsükliliste struktuuride moodustumisel muutuvad asümmeetrilisteks süsinikeks. D-suhkrud: kui anomeerse C aatomi juures paiknev OH-rühm on Haworth'i projektsioonis suunatud tsükli
tasapinna alla, siis -anomeer, kui üles, siis -anomeer. L-suhkrud:
vastupidine asetus . Esindajaid: riboos, glükoos,
fruktoos ,
galaktoos .
3. Monosahhariidide derivaadid.
Suhkurhapped : karboksüülrühm C-1
keto - või aldehüüdrühma asemel. karboksüül C-1 asendis > aldoonhapped (N: glükoonhape, galaktoonhape). Ainult lineaarne karboksüül C-6 asendis > uroonhapped (N: glükuroonhape, galakturoonhape). Tsükliline või lineaarne. Redoksreaktsioonide käigus suhkur oksüdeeritakse suhkurhappeks. Suhkuralkoholid: hüdroksüülrühm C-1 asendis (ntks glütserool, inositool lipiidides). Suhkuralkoholid e alditoolid on lineaarsed molekulid. Saab sünteesida aldoosidest ja ketoosidest karbonüülrühma taandamisel pehmete taandajatega. Desoksüsuhkrud: üks või enam hüdroksüülrühmi asendatud
vesinikuga (ntks 2-desoksü-D-riboos DNAs) Suhkruestrid: mõni hüdroksüülrühmadesr fosforüleeritud (ntks 5'-ATP). Suhkurestrid on olulised metaboolsed
intermediaadid .
Aminosuhkrud : aminorühm C-2 asendis hüdroksüülrühma asemel (leidub
oligo - ja polüsahhariidides, ntks kitiinis) 1
Atsetaalid, ketaalid ja glükosiidid: metüül-, etüülglükosiidid. Lehekülg 4.
Oligosahhariidid on liitsuhkrud, mis koosnevad väikesest arvust monosahhariidi molekuli jääkidest, mida seovad glükosiidsidemed. -glükosiidside tekib konfiguratsioonis C1 osalusel; -glükosiidside tekib konfiguratsioonis C1 osalusel. Glükosiidsidet tähistatakse: Glc1-4Glc; Glc14Glc; Glc4Glc. Glükosiidid on eetritüüpi ühendid, mis on moodustunud monosahhariidist ja alkoholist või mitmest monosahhariidist. Redutseeriva oligosahhariidi määrab vaba hemiatsetaalsne OH, st vaba
redutseeriv ots.
Sahharoos ei ole redutseeriv (taandav) suhkur. Olulisemad
disahhariidid : maltoos,
laktoos , sahharoos, tsellobioos, isomaltoos.
5. Polüsahhariidid e polüüsid koosnevad väga paljudest glükosiidsidemetega ühendatud monosahhariidi molekuli jääkidest. Funktsioonid: energiavaru: tärklis, glükogeen, dekstraan,
inuliin .
Struktuurne e ehituslik:
tselluloos ,
kitiin , ksülaan, galaktaan. Molekulaarne äratundmine: rakupinna polüsahhariidid. Tärklis on glükoosi varu taimedes. Sisaldab 2 tüüpi glükoosi polümeere, millest 10-30% on amüloos ja 70-90% amülopektiin. Glükogeen on glükoosivaru
loomades . Energiavaru, mis moodustab kuni 10% maksa ja 1-2% lihaste massist. Koosneb glükoosijääkidest, mis ühendatud (1,4)-glükosiidsidemetega. (1,6)-glükosiidsidemega hargnemised iga 8-12 jäägi järel. Amülaasid on ensüümid, mis katalüüsivad tärklise ja glükogeeni Tselluloos hüdrolüüsi. Tselluloos on looduses levinuim struktuurne polümeer. ,D- glükoosi jäägid ühendatud (14)-sidemetega lineaarseteks ahelateks Kitiin - eksoskelett koorikloomadel ja putukatel, esineb ka seente, pärmide rakuseinas; koosneb (14) seotud N-atsetüül-D- glükoosamiini monomeeridest. Risseotud dekstraanid on kasutusel geelidena kolonnkromatograafias. Need geelid sisaldavad 50-98% vett. Geelide mehaaniline tugevus ja vee
sidumise võime sõltub ristsidemete arvust.
6.
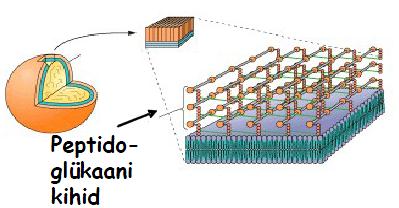
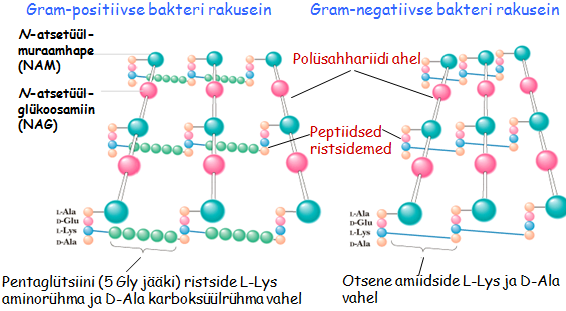
Gram -positiivne
bakter . Rakuseinas on palju peptidoglükaanikihte. Gram-positiivne bakter Gram-negatiivne bakter. bakteri rakuseinas on valine lipiidne
kaksikkiht (
membraan ), sisemine lipiidne kaksikkiht (membraan) ja nende vahel peptidoglukaani kiht (enamasti uks); raku pinnal paiknevad lipopolusahhariidid. Rakuseina peptidoglükogaane tuntakse ka mreiinide nime all.
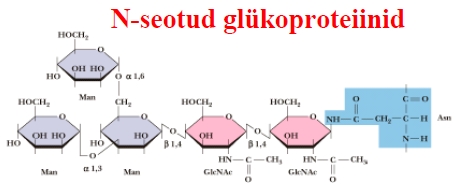
Gram-negatiivne bakter 7. Rakupinna süsivesikud kannavad endas olulist informatsiooni. Glükoproteiinid ja proteoglükaanid - tohutult palju erinevaid Glükoproteiinid = süsivesik (oligosahhariid) + valk Proteoglükaanid = heteropolüsahhariid (glükoosaminoaglükaan) + valk. Komponendid kovalentselt seotud Polüsahhariidne komponent reguleerib rakk-rakk äratundmist ja regulatsiooni Nende struktuuride "informatsiooni"
unikaalsus on tagatud ensüümidega, mis katalüüsivad nende polüsahhariidide sünteesi. Glükoproteiinid on
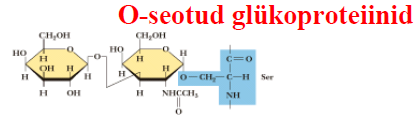
liitvalgud , mis sisaldavad mitte- aminohappelise osana süsivesikuid. Oligosahhariidid on 2
seotud ka N- või O-aatomi kaudu. O-seotud oligosahhariidid Lehekülg
on kinnitunud valgule Ser, Thr või hüdroksülüsiini OH-rühma kaudu. N-seotud oligosahhariidid on kinnitunud valgule aspargiini amiidrühma kaudu. Proteoglükaanid on hübriidsed makromolekulid, mis koosnevad heteropolüsahhariidist ja polüpeptiidist. Koosnevad tuumikproteiinist, millega on kovalentselt seotud ühte või kahte tüüpi glükoosaminoglükogaanid. Rakupinna ja ekstratselluaarse maatriksi komponendid. Ntks sündekaan. Funktsioonid: raku kasvu kontroll (kasvufaktorite sidumine proteoglükaanidega glükokalüüsis loob rakupinna kasvufaktorite reservuaari;
liigeste polsterdamine (kõhre maatriksi proteoglükaanid absorbeerivad suures koguses vett.
XII
LIPIIDID1. Lipiidid on
struktuurilt ja funktsioonilt
heterogeenne grupp biomolekule, mille ühiseks tunnuseks on lahustumatus vees. Struktuuri järgi jaotatakse:
rasvad , vahad, fosfo- ja glükolipiidid,
rasvhapped ja nende derivaadid, trepenoidid (sh
steroidid ). Funktsioonide järgi jaotatakse: varulipiidid (enam kui 80% adipotsüütide massist); struktuurilipiidid (5-10% raku kuivmassist); signaalmolekulid (hormoonid ja sekundaarsed ülekandjad); kofaktorid ja rasvlahustuvad
vitamiinid ; pigmendid. Rasvhapped on alifaatsed 4-24 (36) süsiniku aatomit sisaldavad karboksüülhapped. Küllastatud rasvhapete süsinikahelad pakitakse tihedalt üksteise kõrvale ning nad moodustavad organiseeritud jäiku agregaate. Vastavatel lipiididel on kõrgem sulamistemp. ning nad annavad membraanidele jäikuse. Küllastamata rasvhapete süsinikahelates on cis-kaksiksidemete juures jäigad painded, mistõttu ei saa neide tihedalt
pakkida . Vastavatel lipiididel on madalam sulamistemp ning nad annavad membraanidele elastsuse. Vt rasvhapete struktuuri esitamisviisid (12/5); süstemaatilised ja triv nimetused (12/6;7); ja levinuimad esindajad (12/8).
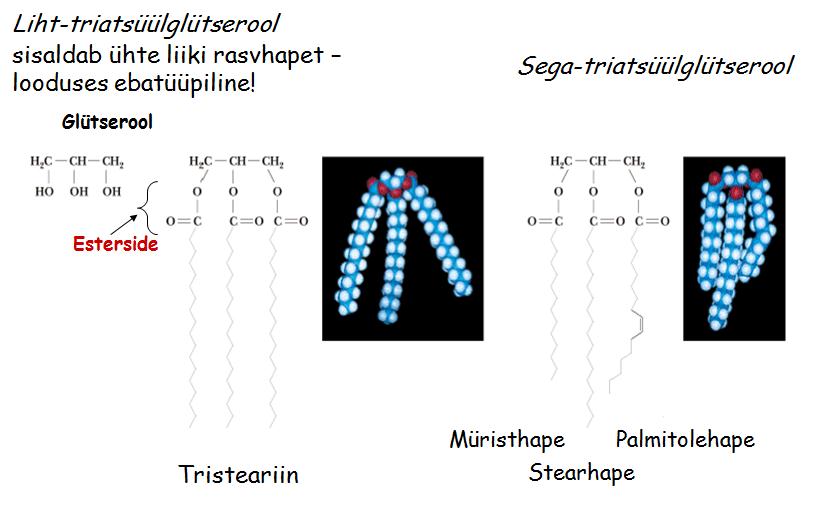
2. Triatsüülglütseroolid (triglütseriidid) = rasvad; on kolmealuselise alkoholi ja kolme rasvhappe
estrid . Peamine eneregeetiline varu organismides (sest: enimtaandatud süsiniku vorm looduses; ei vaja solvatatsiooni; efektiivselt pakitavad). Termoisolatsioon.
Metaboolne vesi (rasvhapete oksüdatiivsel lõhustamisel vabaneb vesi ja süsihappegaas). Rasvad ei kuulu membraanide koosseisu. Glütserofosfolipiid Vahad on pika C-ahelaga alkoholide ja pika C-ahelaga Vaha rasvhapete estrid. On vees
lahustumatud . Omavad kaitsefunktsiooni (vaha
kihiga on kaetud loomade nahk ja
karv , taimede lehed, lindude suled). Karnaubavaha palmilehtedest poleerimisvahad;
lanoliin lambavillast kosmeetika ja ravimid).
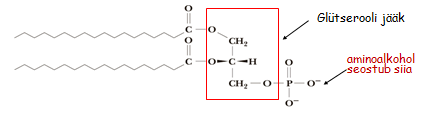
3. 3. Polaarsed lipiidid kus glütserooli C-1 ja C-2 juurde on estersidemega seotud rasvhapped ja C-3 juurde fosfodiestersidemega
polaarne alkohol (aminoalkoholid aminoetanool, trimetüülaminoetanool e koliin jt). Glütserofosfolipiidid. Kui esineb küllastamata
rasvhape , siis on see teises asendis. Levinuimad esindajad: fosfatidüületanoolamiin, fosfatidüülseriin, Singosiin fosfatidüülglütserool, difosfatidüülglütserool, fosfatidüülnositool. 3
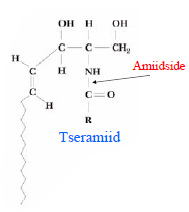
Sfingolipiidid e sfingosiidid. Struktuuri aluseks on sfingosiin - C-18 aminoalkohol. Lehekülg
Tseramiidid rasvhape on seotud sfingosiini C-2 asendi aminogrupiga amiidsidemega. Sfingomüeliinid on tseramiinid, mille C-1 asendi hüdroksüülrühm on esterdatud fosforüülkoliini või fosforüületanoolamiiniga. Glükosfingolipiidid on tseramiidid, mille C-1 asendi hüdroksüülrühma juurde on -glükosiidsidemega Sfingomüeliin seotud üks või enam suhkrujääki. Tsrebrosiidid sisaldavad monosahhariidi, gangliosiidid sisaldavad oligosahhariidi.
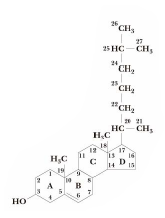
4. Trepenoidid on struktuurilt mitmeksine ja laialt levinud looduslike ühendite perekond, mille
molekulid koosnevad korduvatest C5 ühikutest, mida võib vaadelda isopreeni jääkidena. Steroidideks nim suurt gruppi tritrepenoide, kuhu kuuluvad paljud bioloogiliselt olulised looduslikud ühendid. Steroidid on polütsüklilised ühendid, mille molekulid sisaldavad steraani skleletti. Steroidide esindajate rühmad:
Steroolid (asendis C3 OH-rühm, asendis C17 kõrvalahel 8...10 süsinikuaatomit; leidub nii loomades kui taimedes); Sapphapped (asendis C17 ahel, milles COOH); Sugu-ja kortikosteroidhormoonid; D-rühma vitamiinid; Kolesterool Steroidalkaloidid; Südameglükosiidid. Kolesterool on kõige levinum steroid loomsetes organismides, olles ühtlasi lähteühendiks kõikidele loomsetele steroididele.
Steroidhormoonid reguleerivad mitmeid funktsioone loomades soolabilanss, metaboolne- ja sugufunktsioon.
XIII MEMBRAANID JA
MEMBRAANITRANSPORT1. Membraanid on raku välispiirdeks ning nad reguleerivad molekulide ja ioonide llikumist läbi selle piirde. Eukarüootides piiritlevad membraanid rakuorganelle, eraldades rakukomponente ja Membraani ehitus protsesse üksteisest ning reguleerides molekulide ja ioonide liikumist nende vahel.
Toimivad toksiliste molekulide vastaste barjääridena. Aitavad rakul omandada väliskeskkonnast lähteaineid biosünteesiks ja energeetilisteks
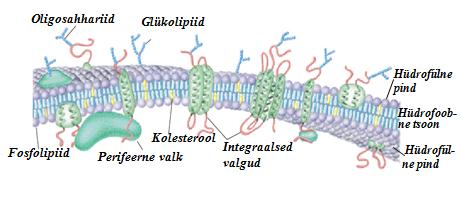
vajadusteks ning väljastada jääkaineid. Osalevad rakkude paljuenmises. Osalevad signaaliülekandes väliskeskkonnast rakku. Osalevad rakk-rakk interaktsioonides. Peamised koostisosad:
valgud (30-75%), fosfolipiidid (7-40%), steroolid (0-25%), süsivesikud. Membraanide lateraalne asümeetria: asümeetria monokihi piires. Valkude lateraalne asümmeetria: valgud võivad moodustada klastreid (kogumeid) membraani pinnal, nad ei ole ühtlaselt jaotatud. Lipiidide lateraalne asümeetria: lipiidid võivad moodustada klastreid membraani pinna, nad ei ole ühtlaselt jaotatud; lipiidklastrite teket saab indutseerida (faaslahutamine). Membraanide
transversaalne asümmeetria: asümeetria risti membraani suhtes. Valkude transversaalne asümmeetria: valgud ei paikne kaksikkihis sümmeetriliselt. Ntks glükoforiini N-terminus on ekstratsellulaarne ning C-terminus intratsellulaarne. Lipiidide transversaalne asümmeetria: paljude membraanide lipiidses kaksikkihis on sise- ja väliskihi koosseis erinev. Membraani faasiüleminekud. Üleminekutemperatuur e "sulamistemperatuur" (Tm) membraani iseloomustav suurus, sõltub membraanilipiidide polaarse peagrupi tüübist, atsüülahelate pikkusest ja küllastamatusest. Allpool Tm'i on 4
membraanilipiidid jäigad ja tihedalt pakitud, kõrgemal temp'l lipiidid muutuvad elastsemaks ja liikuvamaks. Temperatuuri Lehekülg
tõusul üle Tm lipiidi atsüülijääkide (hüdrofoob-
sete sabade) liikuvus kasvab drastiliselt. Membraani pind võib suureneda ja paksus väheneda. 2. Membraanivalkude tüübid:
perifeersed (välimised); integraalsed (sisemised);
lipiid -ankurdatud valgud. Perifeersed valgud. Harilikult
globulaarsed . Kinnituvad membraani integraalsetele
valkudele nõrkade jõudude abil. Neid on kerge membraanilt dissotsieerida kas pehme detergendi või kõrge soolakontsentratsiooniga lahustega töötlemisel. Integraalsed valgud. Tugevasti kinnitunud lipiidi kaksikkihti. Neid on võimalik membraanist eemaldada ainult membraani denatureerides. Sagel transmembraansed. Võimalik ka monotoopne sidumine membraanile (ulatuvad vaid ühte lipiidikihti). Lipiid-ankurdatud valgud. Ankurdatud membraani tänu kovalentsele sidemele lipiididega. Valkude pöörduv ankurdamine membraanile ja vabastamine membraanilt kontrollib signaaliülekannet. 4 ankru tüüpi: Amiid-seotud müristüülankrud; Tioester-seotud (rasvhappe-) atsüülankrud; Tioeeter-seotud prenüülankrud; Glükosüül-fosfatidüülinositoolankrud.
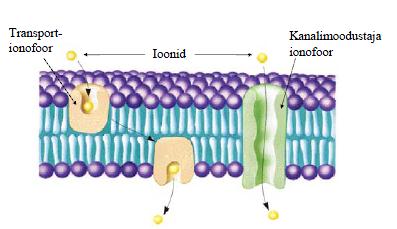
3. Membraanitransport. Vähesed apolaarsed ained lahustuvad membraani lipiidses kaksikkihis ning suudavad iseseisvalt läbida membraani. Polaarsed ja laetud ühendid ning
ioonid ei suuda läbida membraani ilma membraanivalkude abita. Tüübid: passiivne difusioon ja soodustatud difusioon ei vaja välist energiat, kulgeb kontsentratsiooni või laengu
gradiendi suunas. Aktiivne transport (primaarne ja sekundaarne) kulgeb vastu kontsentratsiooni gradiendi suunda, vajab ATP hüdrolüüsi või ioongradiendi energiat. ionofooride vahendatud ioonide transport, ioonkanalite vahendatud ioonide transport ning poriinide (välismembraani kanalvalgud) vahendatud transport kontsentratsiooni gradiendi suunas. Passiivne diffusioon. Laenguta osakeste. Ei vaja spetsiaalseid valke. Transporditavad osakesed liiguvad oma kontsentratsioonigradiendi suunas. Laetud osakeste. Sõltub lisaks osakeste kontsentratsioonidele kahel pool membraani ka veel osakeste laengust ja elektrokeemiliste potensiaalide vahest kahel pool membraani. Soodustatud difusioon. Lahustunud ained saavad liikuda ainult termodünaamiliselt eelistatud suunas. Valgud võivad hõlbustada molekulide või ioonide transporti, tõstes selle kiirust. Lahustunud aine liigub ainult eelistatud suunda. Valgud omavad afiinsust ja selektiivsust transporditava aine suhtes. Transport allub küllastuskineetikale. Ntks glükoosi transport erütrotsüütides.
4. Aktiivse transpordi süsteemid. Aktiivne transport kulgeb nii, et lahustunud ained liiguvad vastupidiselt elektrokeemilise gradiendi suunale. See vajab lisaenergiat, mille allikaks võib olla ATP, valgus või ioongradientides salvestatud energia. Primaarne aktiivne transport loob ja säilitab ioonide kontsentratsioonigradiente raku väliskeskkonna vahel, toimib ATP hüdrolüüsi energia arvel. ATPaasid katalüütilised valgud ("pumbad"), mis on võimelised ATP-d hüdrolüüsima ja vabanevat energiat kasutama, et + + + + + + muuta valgu konformatsiooni. Näiteid:* Na ,K -
ATPaas (Na/K-
pump ): väljutab rakust liigse Na ja sisestab K | * H ,K - + ATPaas (H/K-pump): mao limaskesta rakkude membraanis, tagab maos ülimadala pH; | * Ca -ATPaas (Ca-pump): väljutab + rakust liigse Ca . Sekundaarne aktiivne transport. Suhkrud ja
aminohapped akumuleeritakse rakku transpordiprotsesside abil, mis toimuvad ioongradientide toel. Sellisteks gradientideks on + + ATPaaside poolt genereeritud H , Na või teiste katioonide ning anioonide gradiendid. Liigid: sümport ioonid ja transporditavad aminohapped või suhkrud liiguvad samas suunas läbi membraani; antiport ioonid ja transporditavad osakesed liiguvad vastassuunades.
5. Ioonide transport ionofooride abil. Ionofoorideks nim ühendeid, mis seovad metalli-ioone ning transpordivad neid läbi membraani. Ionofoorid
XIV NUKLEIINHAPETE KOMPONENDID JA EHITUS
1. Informatsiooni ülekanne rakus. Informatsioon, mis on kodeeritud DNA molekulis transkribeeritakse RNA molekuli. RNA molekuli järjestus loetakse ja transleeritakse aminohappejärjestuseks valkudes. DNA
replikatsioon on matriitssüntees, mille tulemusena saadakse ühest DNA molekulist kaks ühesuguse nukleotiidse järjestusega DNA 5
molekuli. Lehekülg
RNA
transkriptsioon on matriitssüntees, mille käigus saadakse DNA molekuli ühe ahela nukleotiidse järjestusega komplementaarne mRNA
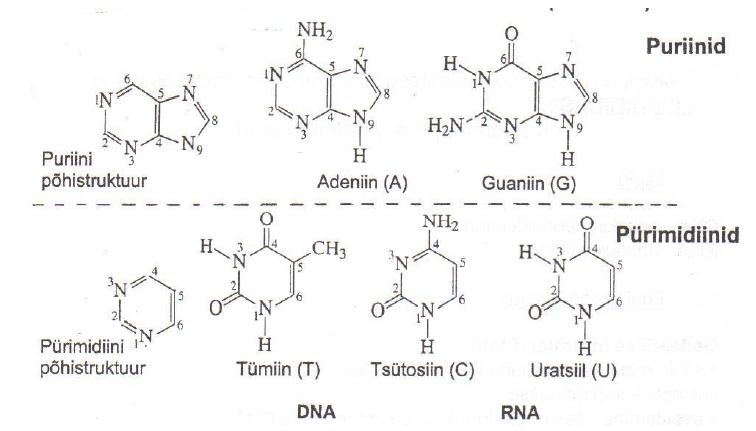
molekul Nukleotiid 2. Nukleiinhapete komponendid. Pürimidiinid: Tsütosiin (DNA, RNA);
Uratsiil (RNA); Tümiin (DNA)
Puriinid : Adeniin;
Guaniin . Suhkrud: Riboos/desoksüriboos.
Fosfaat .
Komplementaarsus on kindlate lämmastikaluste paardumine nukleiinhapete molekulides, mis põhineb
vesiniksidemete moodustumisel. Komplementaarsed aluspaarid on C-G ja A-T(DNA)/U(RNA).
3. 3.
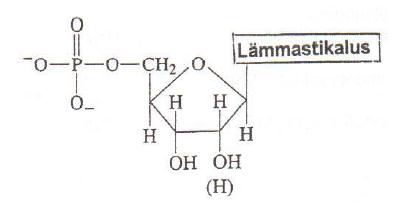
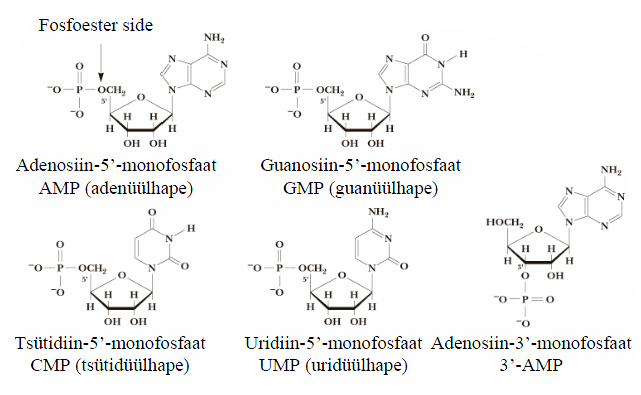
Nukleosiid = lämmastikalus + suhkur. Alus on seotud suhkru külge lämmastikglükosiidsidemega. Glükosiidsideme süsinik on anomeerne. Nukleosiidide nimetused saadakse, Lämmastikalused lisades nimetuse tüvele idiin (pürimidiinid) või osiin (puriinid). Nukleosiidid võivad esineda syn- või anti- konformatsioonis, mis on tingitud steerilisest takistusest pöörlemisel ümber glükosiidsideme. Puriinnukleosiididel on võimalikud mõlemad konformeerid, pürimidiinnukleosiididel on eelistatud anti-konformeer. Suhkrukomponendi tõttu lahustuvad nukleosiidid vees paremini kui vabad lämmastikalused. Nomenklatuur: adenosiin/desoksüadenosiin; guanosiin; tsütidiin/desoksütsütidiin; tümidiin; uridiin. Nukleotiid = nukleosiidfosfaat Ribonukleotiidide
struktuurid ja nomenklatuur = lämmastikalus + suhkur + 1-3 fosforüülrühma. Enamik vabu
nukleotiide on ribonukleotiidid, millele fosforüülrühm
riboosi 5'- asendis.
Nukleotiidid on polüprootsed happed. Fosforüülrühma esimene
prooton dissotseerub pH1 ja teine pH6 juures. Seega neutraalse pH juures on nukleosiidmonofosfaadi summaarne laeng -2. Nukleotiidide funktsioonid: nukleosiid-5'-trifosfaadid (NTP) on substraatideks nukleiinhapete sünteesil. NTPd on energiakandjad ATP on
keskne molekul energia metabolismis; GTP on peamine
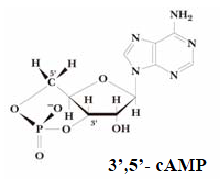
energiaallikas valgu sünteesis; CTP on oluline metaboliit fosfolipiidide sünteesis; UTP aktiveerib suhkrud polüsahhariidide sünteesis. Nukleotiidide koosseisu kuuluvad Tsükliline nukleotiid lämmastikalused äratundmisühikutena, ise nad metabolismis ei osale. Tsüklilised nukleotiidid on signaalimolekulid ja
regulaatorid raku metabolismis ja reproduktsioonis. Tsüklilised nukleotiidid.
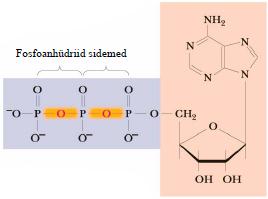
Nukleosiidmonofosfaadid , milles fosforhape on estersidemete kaudud seotud riboosi kahe hüdroksüülrühmaga. Raku metabolismi olulised regulaatorid. Nukleosiiddi- ja trifosfaadid. Nukleotiidi fosforüülrühmale võivad fosfoanhüdriidsideme 6
abil liituda täiendavad fosfaatrühma (tähistatakse , , ) Lehekülg Täiendava fosfaatrühma lisandumine fosfoanhüdriidsideme abil
4.
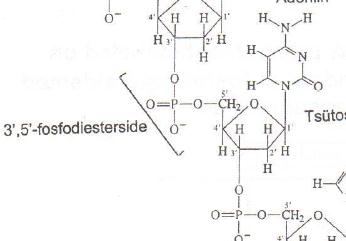
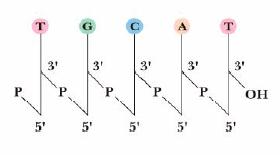
Nukleiinhape = polünukleotiid. Biopolümeerid, mille monomeersed ühikud, nukleosiidmonofosfaadid, on ühendatud 3'-5'- fosfodiestersidemetega. Monomeersete ühikute järjestust loetakse alati 5' 3' suunas. DNA on antiparalleelne kaksikheelik. Võib olla tsirkulaarne (prokarüootides) või lineaarne. Kompkaten ja tihedalt pakitud. Eukarüootne DNA kerituna ümber histoonvalkude moodustab nukleosoome. Eukarüootne mRNA (
maatriks -RNA)
vahendab geneetilist koodi. DNA transkriptsiooni käigus sünteesitakse heterogeenne tuuma RNA, kus vahelduvad
intronid (vahejärjestused) ja
eksonid (kodeerivad järjestused), sisaldab polü-A saba (seotud RNA stabiilsusega). Splaissimise käigus lõigatakse intronid välja ning saadakse küps mRNA. rRNA (ribosoomne RNA) ribosoomi struktuuri ja funktsiooni alus. tRNA (transpordi RNA) kannab
aminohappeid valgusünteesis. Lühikesed polünukleotiidahelad 73-94 jääki. Mitmed alused on metüleeritud. Iga aminohappe jaoks on vähemalt üks unikaalne tRNA. 3'-terminaalne järjestus on alati CCA.
snRNA (väike tuuma RNA) on oluline eukarüootsete geenide esmaste transkriptide protsessingul küpseks mRNA-ks enne eksporti tuumast Nukleiinhappe skemaatiline esitus. Suhkrud tsütoplasmasse. Koosneb 100-200 nukleotiidist, millest osa on on kujutatud püstjoontena, kaldjooned on fosfodiestersidemed. Lämmastikalused on metüleeritud. Esineb stabiilsete RNA-valk kompleksidena, nn väikeste tähistatud ringidena. tuuma ribonukleoproteiinidena (snRNP).
siRNA (väike interfeeriv RNA) osaleb transkriptisoonijärgses geenide vaigistamises.
5. Nukleiinhapete hüdrolüüs. RNA onvastupidav lahjades hapetes, hüdrolüüsitakse lahja
leelise poolt. DNA on vastupidav leeliste suhtes; lahjades hapetes depurineeritakse, st hüdrolüüsitakse puriinnukleosiidide N- glükosiidside. Polünukleotiidahelate lõikamine. Eksonukleaasid hüdrolüüsivad fosfodiestersidet terminaalsete jääkide juurest. Endonukleaasid hüdrolüüsivad nukleiinhapete sisemisi (mitte-terminaalseid) fosfodiestersidemeid. Restriktsiooniensüümid.
Bakterid kaitsevad ennast võõra DNA sissetungi eest restriktsiooniensüümide e restriktaaside abil. II ja III tüüpi restriktaasid lõikavad DNA ahelaid kindlates kohtades. Ensüümid võivad ära tunda 4-6 või rohkem aluseid potensiaalse lõikamissaidi juures.
XV NUKLEIINHAPETE STRUKTUURITASEMED
1. DNA sekundaarstruktuur
vesiniksidemed komplementaarsete aluspaaride vahel. Suhkur-fosfaat põhiskelett väljaspool; lämmastikalused seespool. Paremakäeline kaksikheeliks jätab aluspaaride omavaheliseks kauguseks 3,4 Å. Lineaarse biheeliksi vormid: A-vorm: paremakäeline, lühike ja lai 2,3 Å, 11 bp pöörde kohta. B-vorm: paremakäeline, pikem ja peenem 3,32 Å, 10 bp pöörde kohta. Z-vorm: vasakukäeline,
pikim ja peenim 3,8 Å, 12 bp pöörde kohta. DNA
denaturatsioon biheeliksi lahtikeerdumine. Struktuuri lõhuvad: ekstremaalne temp (suureneb UV
neelduvus ), keskkonna pH (10) ja ioonjõud ning tugevad vesiniksidemete moodustajad (formamiid,
uurea ). Kui temp alandada, siis aluspaaride valguse absorbeerimine nõrgeneb, mis näitab korrastatud struktuuri taastumist e renaturatsiooni. 7 Lehekülg
2. DNA tertsiaarstruktuurid. Superspiraalid. Ensüümid võivad tekitada või kõrvaldada superspiraale. Negatiivne superspiralisatsioon võib tekitada ristikujulisi struktuure. Tüübid: (a) DNA kaksikheeliks on
keerdunud spiraaliks ümber imaginaarse toroidi. (b) DNA on keerdunud ümber iseenda. (c) superspiraalid pikas ja lineaarses DNA ahelas (kromosomaalse DNA mudel). Kerimine ümber valgupooli (nukleosoomi) stabiliseerib DNA superspiraalset struktuuri. Kromosoomi struktuur. DNA kaksikheeliks keerdub 2x ümber histooni oktameeri moodustades nukleosoomid. Nukleosoomid on keerdunud solenoidideks (6 nukleosoomi pöördes), moodustades nii
filamente . Filamendid omakorda 6 moodustavad silmuseid, mis kinnituvad tuumamaatriksile. 18 silmust keerduvad rosettideks. Ligikaudud 10
rosetti eksisteerib inimese 4. kromosoomi igas kromatiidis. Nukelosoomi struktuur:
Kromatiin = nukeliinproteiin-
kompleks . Koosneb
DNAst , histoonidest ja mittehistoonsetest kromosoomsetest valkudest. Histoonid on valgud, mis seostuvad tugevalt DNAga ning esinevad kõikide eukarüootide kromosoomides. Mittehistoonsed valgud on geeni ekspressiooni regulaatorid.
o o o o 0 o 0 o o
3. RNAde struktuuritasemed: mRNA: 1 (oletatakse ka 2 ); tRNA: 1 , 2 , 3 ; rRNA: 1 , 2 , 3 , 4 . tRNA sekundaarstruktuur. Ristikheinaleht. Ulatusliku H-sidemete võrgustiku tekke tõttu tRNA palindroomsetes lõikudes moodustub 4 kaksikhelikaalset domeeni, millest 3 lõpevad silmusega ning neljas ,,tüvega", kus 3' ja 5'-
otsad . tRNA-s leidub palju ebaharilikke ribonukleosiide. Erinevate tRNA molekulide minimaalne arv rakus on 31. Ribosoomne RNA. Kõik
ribosoomid koosnevad suurest ja väikesest subühikust. Umbes 2/3 osas RNA-st, 1/3 osas valgust. rRNA toimib toena ribosoomsetele valkudele ja võib omada katalüütilist aktiivsust. Kõrge ahelasisene järjestuste komplementaarsus põhjendab (ilmselt) ulatuslikku aluspaaride teket ja kaheahelaliste lõikud moodustumist. Ribosoom koosneb kahest subühikust. Koostis ja suurus on pro- ja eukarüootides veidi erinev: Prokarüootne on väiksem ja sisaldab vähem valke ja nukleotiide. DNA ja RNA erinevused. DNA sisaldab tümiini kõik DNAs leiduvad uratsiiljäägid pärinevad mutatsioonidest ning kuuluvad asendamisele tsütosiinidega. RNA kõrvutiasetsevad 2' ja 3' OH-rühmad muudavad RNA vastuvõtlikuks hüdrolüüsile, DNA-l 2'OH puudub ja on seega stabiilsem RNA on
disainitud kasutamiseks ja seejärel hävitamiseks; geneetiline materjal peab aga olema stabiilne.
XVI METABOLISMI ÜLDPÕHIMÕTTED
1.
Metabolism . Keemiliste muutuste kogum, mis muundab
toitained energiaks ning keemiliselt ehituselt keerulisteks rakuproduktideks. Metaboolsed kaardid
esitavad reaktsioonide kaskaade integreeritud kujul. Sajad ensüümreaktsioonid on organiseeritud diskreetseteks metaboolseteks radadeks. Põhiliste metabolismiradade osas on organismidel märkimisväärne sarnasus (mis võib tõestada, et kogu elu pärineb ühes
eellasest ). Esineb ka suur mitmekesisus (auto- hetero-
kemotroofid jne). Organismid jaotuvad (1)
hapnikutarbe järgi: Aeroobsed: kasutavad hapnikku elektronide aktseptorina. Kui hapnik eluks välitimatu, siis oblikatoorsed aeroobid. Anaeroobid: võimelised
eksisteerima ilma hapnikuta. Kui ei talu üldse hapnikku, siis oblikatoorsed anaeroobid.
Fakultatiivsed anaeroobid on võimelised kohanema aeroobsete tingimustega, kasutades teisi elektronide aktseptoreid. (2) energia- ja süsinikuallika järgi
Fotoheterotroof : heterotroofne organism, kes kasutab valgust energia allikana, aga ei kasuta CO2 orgaaniliste ühendite moodustamiseks. Kasutavad keskkonnast saadavat orgaanilist ainet.
Kemoautotroof : ammutavad energiat teisi (peale hapniku) vaba elektronpaari
omavate ühendite oksüdeerimisest. Kogu vajalik orgaaniline aine sünteesitakse CO2.
Fotoautotroof : energia saamiseks viivad läbi fotosünteesi. Selle energiaga muudavad CO2 ja H2O orgaaniliseks aineks. Kemoheterotroof: kasutavad orgaanilist ainet keskkonnast.
2.
Katabolism on lagundav
ainevahetus , keerulisematest ainetest tekivad lihtsamad ja milles vabaneb energiat. degradatiivsed metabolismirajad.
Anabolism toimub biosünteesi
radadel . Lihtsamatest keemilistest ühenditest sünteesitakse
keerulisemad ühendid. Protsessi käigus vajatakse energiat. Katabolismiga (toitainete lõhustamisega) toodetakse keemiline energia, millega viiakse läbi anabolism (raku makromolekulide tootmine). Metabolismiradade organisatsioon:
rajad koosnevad järjestikustest ensüümireaktsioonidest. Ensüümid võivad esineda 8
eraldiseisvate valkudena, multiensüümsete kompleksidena, membraanseotud süsteemidena. Lehekülg
3. Et tagada raja
spontaansus peab vähmalt üks aste erinema. Erinevate
astmete esinemine võimaldab vastavate regulatsioonimehhanismide abil ühe- või teisesuunalise raja välja lülitada (metaboliidid on ensüümkatalüütiliste metaboolsete reaktsioonide vaheühendid). Katabolismi ja anabolismi sõltumatutel radadel toimub ühe poole aktiveerimisel teise poole inhibeerimine. Katabolismi ja anabolismi osaliselt kattuvatel radadel katalüüsivad pöördreaktsioone katabolismile või anabolismile vastav ensüüm, teist inhibeeritakse.
4.
Redoksreaktsioonid metabolismis. Katabolism on oksüdatiivne substraadid (valgud, suhkrud, süsivesikud) loovutavad redutseerivaid ekvivalente, harilikult + + hüdriidioone. NAD või NADP seovad kataboolsetes reaktsioonides vabanevad elektronid ja formeeruvad redutseeritud Süsiniku oksüdatsiooniastme järk-järguline muutumine vormid NADH või NADPH. Anabolism on redutseeriv NADH ja NADPH tagavad redutseeriva jõu anaboolseteks protsessideks.
5.
Eksperimentaalsed meetodid metaboolsete radade
uurimisel : (1) isotoopmarkerid sondidena (mõne keemilise elemendi isotoobid viiakse rakku); (2) inhibiitorite kasutamine (I'ga blokeeritakse üks või teine ensüümireaktsioon metaboolsel rajal.
Produkti järgi määratakse selle reaktsiooni asend metaboolsel rajal); (3) TMR
spektroskoopia ; (4) raku organellide lahutamine fraktsioonideks ja seejärel metaboolsete funktsioonide
uurimine . Peamised toitainete grupid: Valgud on lämmastikurikkad ning varustavad organismi aminohapetega. Süsivesikud annavad energia ja olulised komponendid nukleotiidide ja nukleiinhapete sünteesiks. Lipiidid varustavad rasvhapetega, mis on
rakumembraanide võtmekomponendid ning on eellasteks signaalmolekulidele. Kiudained stimuleerivad soolestiku ja absorbeerivad orgaanilisi molekule seedetraktis. Vitamiinid ja mineraalid.
6. Energia ülekanne: päigesevalgusest või toidust saadud energia tuleb kasutada energiat tarbivate protsesside käigushoidmiseks organismis. Selleks on võimelised kahte tüüpi
biomolekulid : (1) taandatud koensüümid (NADH, NADPH, - FADH2) kui kõrge e ülekandepotsensiaaliga ühendid ja (2) energiarikkad fosfaadid, mille hüdrolüüsi
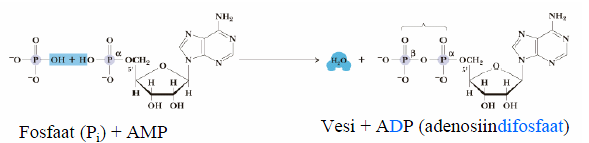
vabaenergia muut on suurem kui -25 kJ/mol.
Makroergilised biomolekulid on lühiajaliseks energiasalvestuseks ja ülekandeks kohandunud energiarikkad molekulid, peamiselt fosfaadid Adenosiin-5'-trifosfaat ATP on vahetu energia ülekande vahend. PEP + H2O Püruvaat + Pi; ADP + Pi ATP + H2O, ehk PEP + ADP Püruvaat + ATP. G=-23 kJ/mol. Suur hüdrolüüsi vabaenergia muut on võimalik tänu (1) reaktandi destabiliseerimisele ATP struktuur elektrostaatiliste tõukejõudude tõttu
osalise positiivse laenguga ATP trifosfaatahel sisaldab 2 fosfori aatomite vahel. (2) produktide stabiliseerimisele fosfoanhüdriidsidet, mille ionisatsiooni ja resonantsi teel. (3)
entroopia kasvule hüdrolüüsi ja hüdrolüüsil vabaneb palju energiat. ionisatsiooni tulemusel.
7. Rasvlahustuvad vitamiinid osalevad ainevahetuse regulatsioonis.
Retinool (vit A). Retinool, retinüülestrid ja retinaalid on vit A vormid. Retinooli omastatakse kas loomse toiduga või sünteesitakse -karoteenist. Retinool konverteeritakse retinaaliks silma võrkkestas, kus ta seotuna opsiinile moodustab rodopsiini valgustundliku pigmendi võrkkesta kepikestes ja koonustes tagab fotokeemiliselt nägemise. Maks, munad, piim, köögiviljad.
Kaltsiferoolid (vit D). Ergokaltsiferool (D2) ja kolekaltsiferool (D3) on hormoonide prekursorid. Kolekaltsiferool sünteesitakse nahas UV toimel. 9
Pole 'päris'
vitamiin , kuna aktiivne vorm, 1,25- Lehekülg
dihüdroksükolekaltsiferool, on
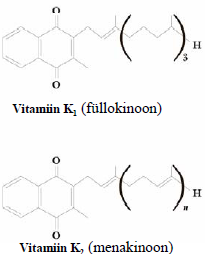
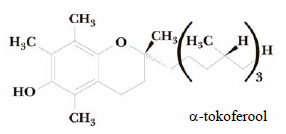
hormoon . Reguleerib Ca ja P metabolismi ja tagab luude normaalse arengu. Või, maks, kalarasv. Tokoferoolid (vit E). Vitamiin E (-tokoferool) on tugev antioksüdant. Kaitseb membraane lipiidide peroksüdatsiooni eest. Taimeõlid, seemned. Kinoonvitamiinid (vit K). Vitamiin K on oluline vere hüübimisel. Vastutab protrombiini aktiveerimise eest. Rohelised taimed, Vitamiin E tsitruselised,
seamaks Polüeenhapped. Taimeõlid, maks
XVII GLÜKOLÜÜS
1. Glükollüüs on ensümaatiliste reaktsioonide ahel, mille käigus glükoos muudetakse püruvaadiks. See on raku tsütoplasmas kulgev universaalne ainevahetusrada. Anaeroobsetes rakkudes ainus ATP
tootev rada. Aeroobsetes rakkudes esimene etapp süsivesikute oksüdatsioonil.
2. Glükolüüsi lähtesubsatraat on glükoos. Esimeses faasis glükoosi molekul muundatakse viie keemilise reaktsiooni abil kaheks glütseraalaldehüüd-3-
fosfaadi molekuliks. I faasis tarbitakse 2 molekuli ATP, et seda hiljem rohkem toota. Esimeses reaktsioonis heksokinaas fosforüleerib glükoosi glükoos-6-fosfaadiks ja hoiab teda rakus. Teises reaktsioonis glükoos-6- fosfaat fosfoglükoosi isomeraasi toimel fruktoos-6-fosfaadiks. Kolmandas reaktsioonis fruktoos-6-fosfaat fosfofruktoosi kinaasiga fosforüleeritakse fruktoos-1,6-difosfaadiks. Neljandas reaktsioonis fruktoos-1,6-
difosfaat lõhustatakse fruktoos- 1,6-difosfaadi aldolaasiga dihüdroksüatsetoonfosfaadiks (DHAP) ja glütseraalaldehüüd-3-fosfaadiks (GAP). Viiendas reaktsioonis dihüdroksüatsetoonfosfaat
konversioon trioosfosfaadi isomeraasi toimel glütseraalaldehüüd-3-fosfaadiks (GAP). Teises faasis tekib kaks molekuli püruvaati. Saadakse netona 4 ATP. Kuuendas reaktsioonis GAP oksüdeeritakse glütseraalaldehüüd-3-fosfaadi dehüdrogenaasi toimel 1,3-difosfoglütseraadiks (1,3-BPG). Seitsmendas reaktsioonis kantakse fosforüülrühm 1,3-GAP'lt üle ADP'le ja saadakse 3-fosfoglütseraat. Kaheksandas reaktsioonis 3-fosfoglütseraat konvergeeritakse fosfoglütseraadi mutaasiga 2-fosfoglütseraadiks. Üheksandas reaktsioonis 2-fosfoglütseraat dehüdraaditakse enolaasiga fosfoenoolpüruvaadiks (PEP). Kümnendas reaktsioonis püruvaadi kinaasiga kantakse fosforüülrühma PEP'lt üle ADP'le ja saadakse 2 püruvaat. ( ) ( ) ( ) ( ) ( ) + Summaarselt: Glükoos + 2 ADP + 2 NAD + 2 Pi 2 püruvaat + 2 ATP + 2 NADH + 2 H2O
0
3. Glükoosi energeetika. Standardolekui G' väärtused on
hajutatud . G rakkudes omab järgmisi väärtusi: enamik väärtusi on nulli lähedased. Kolm reaktsiooni kümnest (1, 3, 10) omavad suurt negatiivset G. Suure negatiivse G reaktsioonid on regulatsioonitsentrid. Ühe glükoosi molekuli kohta saadakse 2 ATP ja 2 NADH molekuli. ATP sünteesi mehhanism: PEP ja 1,3-BPG genereeritakse glükoosi lagundamise käigus. Nende (ja fosfaatide) Püruvaat energia kantakse üle ADP'le ATP sünteesiks: PEP + H2O Püruvaat + Pi; ADP + Pi ATP + H2O.
4. NADH transforamtsioon. NADH on potensiaalne energiaallikas. Aeroobsetes tingimustes NADH re-oksüdeeritakse elektronide transpordi ahelas oksüdatiivse fosforüleerimise käigus, tekib ATP. Anaeroobsetes tingimustes NADH re- + oksüdeeritakse laktaadi dehüdrogenaasi poolt,
andes täiendava NAD edasiseks glükolüüsiks. Püruvaadi
transformatsioon . Kannab endas samuti suurt energeetilist potensiaali. Aeroobsetes tingimustes siseneb 10
tsitraaditsüklisse, kus lõhstatakse CO2 ja H2O-ks ning sünteesitakse ATP. Anaeroobsetes tingimustes nim püruvaadi metabolismi fermentatsiooniks. Alkohoolne
fermentatsioon = käärimine: Lehekülg
Piimhappeline fermentatsioon (mikroorganismid ja loomad):
5. Enam-vähem vastaud 3.ndas.
6. Glükolüüsi teised substraadid. Fruktoos ja mannoos suunatakse glükolüüsi mööda üsna harilikku rada. Galaktoos konvergeeritakse enne glükolüüsi suunamist glükoos-6-fosfaadiks (Leloir'i rajal)
XVIII TSITRAADITSÜKKEL ehk TRIKARBOKSÜÜLHAPETE TSÜKKEL (TCA) ehk KREBS'i TSÜKKEL
1. Tsitraaditsükkel on universaalne ja keskne
kataboolne rada, milles süsivesikute, lipiidide ja valkude degradatsiooniproduktid oksüdeeritakse CO2-ks. Tsükli iga
ringiga siseneb 2 atsüülrühma süsinikku CoA kujul ning
lahkub 2 süsinikku CO2-na. Tsükli tinglikuks lähtepunktiks letakse atsetüül-CoA kondensatsiooni oksaalatsetaadiga, mille tulemusena moodustub tsitraat. Iga tsükkel produtseerib taandatud elektronikandjad, mille re-oksüdatsioon annab energia 1 ATP sünteesiks. Tsükli intermediaadid on biosünteesi lähteained. Eukarüootidel mitokondri maatriksis. TCA tsükli koht rakuenergeetikas. Universaalne katabolismirada, kus toimub süsivesikute, lipiidide ja valkude
degradatsioon toodetud atsetüül-CoA lõplik oksüdeerimine CO2-ks. Produtseeritakse elektronikandjad NADH ja FADH2, mille reoksüdatsiooni arvel taandatakse hapnik
veeks ning sünteesitakse ATP. Püruvaadi sisenemine TCA-sse. Seda katalüüsib püruvaadi dehüdrogenaasi kompleks, mis koosneb 3 ensüümist ja 5 + + koensüümist. Püruvaat + CoA + NAD Atsetüül-CoA + CO2 + NADH + H .
2. Tsükli keemiline loogika. TCA tsükkel võib paista liig keerulisna
atsetaadi oksüdeerimiseks, kuid normaalsed rajad C-C sidemete lõhustamiseks atsetaadi puhul ei tööta. Otstarbekam on kondenseerida
atsetaat oksaalatsetaadiga ja viia läbi - lõhustamine TCA ühendab endas sellise lõhustamise ja oksüdeerimise CO2-ks, regenereerib oksaalatsetaadi ja akumuleerib kogu energia NADH ja ATP kujul. Reaktsiooinid.
Reaktsioon 1:
tsitraadi süntees oksaalatsetaadist ja atsetüül-CoA tsitraadi süntaasiga (atsetüül-CoA karbanioon atakeerib nukleofiilselt oksalaatatsetaadi -keto rühma karbonüülset süsinikku, millele järgneb tioestri hüdrolüüs ja CoA
vabanemine ). Reaktsioon 2: tsitraadi isomerisatsioon isotsitraadiks akonistaasiga. Reaktsioon 3: esimene oksüdatiivne dekarboksüleerimine isotsitraadi dehüdrogenaasiga. Reaktsioon 4: teistkordne oksüdatiivne dekarboksüleerimine -ketoglutaraadi dehüdrogenaasiga (kompleks koosneb kolmest ensüümist, mis kokku kasutavad
viit koensüümi). Reaktsioon 5: Substraadi taseme fosforüleerimine suktsinüül-CoA süntetaasiga. Reaktsioon 6: suktsinaadi oksüdatsioon FAD osalusel suktsinaadi dehüdrogenaasiga (suktsinaadi oksüdeerimisega kaasneb FAD
taandamine FADH 2-ks, mille re-oksüdeerimine kannab elektronid koensüüm Q-le). Reaktsioon 7: katalüüsib fumaraadi transhüdratatsiooni L- + malaadiks fumaraasiga. Reaktsioon 8: malaadi NAD -sõltuv oksüdatsioon oksaalatsetaadiks malaadi dehüdrogenaasiga.
+ 3,4,6 ja 8. Raektsioonidel toimivad ensüümid vajavad koensüüme NAD või FAD.
3. Ühe atsetüül-CoA molekuli oksüdeerimisel ühe TCA tsükli jooksul saadakse kaks CO2, 1 ATP ja 4 taandatud koensüümi. Summaarselt on tsükkel eksergooniline.
4. TCA kui amfiboolne rada. -ketoglutaraat transamineeritakse glutamaadiks, mida kasutatakse Gln, Arg ja Pro ning puriinnukleotiidide sünteesis. Oksaalatsetaat transamineeritakse aspartaadiks ning kasutatakse mitmete aminohapete ja pürimidiinnukleotiidide sünteesis. Suktsinüül-CoA on kasutatav porfüriinide sünteesis. Fumaraat konverteeritakse aspartaadiks ning kasutatakse järgnevalt mitmete aminohapete ja pürimidiinnukleotiidide sünteesis.
5. TCA
regulatsioon . Tsitraadi süntetaas ATP, NADH ja suktsinüül-CoA inhibeerivad. Isotsitraadi dehüdrogenaas ATP 11
+ inhibeerib, ADP ja NAD aktiveerivad. -ketoglutaraadi dehüdrogenaas NADH ja suktsinüül-CoA inhibeerivad, AMP + aktiveerib. Püruvaadi dehüdrogenaasi regulatsioon ATP, NADH, atsetüül-CoA inhibeerivad, NAD , CoA aktiveerivad. Lehekülg
6. Glüoksülaadi eKrebs-Kornbergi tsükkel TCA tsükli variant taimedes ja
bakterites , mis võimaldab sünteesida 4C- dikarboksüülhappeid ja suhkruid atsetaadist. Atsetaadil, kui ainsal C allikal baseeruv kasv süsivesikute ja teiste vaheühendite süntees atsetaadist (mis ei ole võimalik (TCAs). Glüoksülaadi tsüklis jäetakse CO2 vabanemisega TCA tsükli etapid vahele. Glüoksülaadi tsükkel aitab taimedel kasvada pimedas. Toimib ka idanevates seemnetes, kus fotosüntees ei ole piisav.
XIX OKSÜDATIIVNE FOSFORÜÜLIMINE. ELEKTRONI TRANSPORDISÜSTEEM (ETS)
1. Oksüdatiivne fosforüülimine ADP ensümaatiline fosforüülimine ATP-ks, mis toimub konjugeeritult molekulaarse hapniku taandamisega veeks taandatud koensüümidelt pärit elektronide arvel. Elektronide transpordiahel elektronid, mida kannavad tsitraaditsüklis moodustunud taandatud koensüümid, läbivad valkude ja koensüümide ahela (hingamisaehla), et moodustada prootonigradient läbi mitokondri membraani. Protsessid toimivad mitokondri sisemembraanis või selle pinna. Mitokondrit ümbritseb kahekordne membraan. Sisemembraani sopistised moodustavad kurde e kristasid.
o
2. Redokspotentsiaal E väljendab seda, kui hästi on üks ühend võimeline teisi ' redutseerima (elektrone loovutama). Kõrge E0 näitab tugevat elektronide sidumise
tendentsi , st tendentsi saada taandatud. Elektronid loovutatakse negatiivsema redokspotensiaaliga reaktsiooni poolt ning võetakse vastu positiivsema redokspotensiaaliga. Vabaenergia muut reaktsiooni poolt. Elektronid
kaotavad energiat liikudes läbi hingamisahela kompleksidest I (või II) kompleks IV.
3. ETS 4 kompleksi. Kompleks I: elektronide ülekanne NADHCoQ, ensüüm: NADH-CoQ oksüdoreduktaas. Kompleks koosneb enam kui 30 subühikust ja töötab prootonite pumbana. Kompleks II: suktsinaadilt ubikinoonile, ensüüm: suktsinaadi dehüdrogenaas. 4 subühikut, neist 2 integraalsed membraanivalgud, 2 mitokondri maatriksis; 5 prosteetilist rühma (FAD, 3 Fe-S klastrit, CoQ). Suktsinaadi dehüdrogenaas on ühtlasi TCA tsükli ensüüm. Kompleks III: ubikinoonilt tsütokroom c-le, + ensüüm: CoQ-tsütokroom c oksüdoreduktaas. Ensüüm ühendab elektronide ülekande QH 2-lt tsütokroom c-le H pumpamisega maatriksist membraanidevahelisse ruumi. Peamine transmembraanne valk on tsütokroom b. Tsütokroom c on vesilahustuv elektronide kandja valk ja paikneb membraanidevahelises ruumis. UQH 2 on rasvlahustuv elektronide kandja. Kompleks IV: tsütokroom c-lt molekulaarsele hapnikule, ensüüm: tsütokroom c oksüdaas. Hapnik on terminaalseks elektronide aktseptoriks elektronide transpordiahelas. Tsütokroom c oksüdaas koosneb 13 subühikust ning kasutab + katalüüsis 2 heemi ja 2 Cu tsentrit. Samal ajal kompleks IV transpordib/pumpab ka H membraanidevahelisse ruumi. Tsütokroomid on väikesed valgud, sisaldavad tugevalt seotud kofaktorit heemi.
Heem - tetrapürrool, seotud koordinatiivselt Fe aatomiga. Fe iooni viies koordinatiivne side moodustub valguga, kuues tavaliselt substraadiga. 3 klassi heeme: (1) Fe- protoporfüriin IX: lihtsaim heem, biosünteesi raja esimene
produkt . Esineb b-tüüpi tsütokroomides. (2) Heem A: üks Me oksüdeeritud formüüliks, vinüülrühm seotud isoprenoidiga. Esineb a-tüüpi tsütokroomides. (3) Heem C: Mõlemad vinüülid on seotud tsõsteiinide kaudu valgu külge. Esineb c-tüüpi tsütokroomides. a ja b tsütokroomid on integraalsed membraanivalgud, c tsütokroom-
integraalne või perifeerne.
4. Ei tea
5. ATP süntetaasi ehitus? P/O suhe mitu ATP molekuli sünteesitakse iga ½ O2 taandamisel veeks (ühe elektronpaari ülekanne). Väärtus NADH reoksüdeerimisel=10/4=2,5; FADH2 reoksüdeerimisel 6/4=1,5.
6. Oksüdatiivse fosforüülimise protsessi regulatsioon.
Inhibiitorid : rotenoon inhibeerib kompleks I. Tsüaniid, asiid ja CO on mürgid, kuna inhibeerivad kompleks IV, seostudes tugevalt tsütokroom a3
vormiga . Oligomütsiin ja DCCD on ATP süntaasi inhibiitorid. Lahutajad on ained, mis lahutavad elektrontranspordi ADP fosforüülimisest, hajutades prootongradiendi. Lahutajad on hüdrofoobsed molekulid, millel on kergesti dissotseeruv prooton (dinitrofenool, dikumarool). Tsütosoolse NADH transport mitokondri maatriksisse on
lahendatud süstiksüsteemide abil, mis vahendavad elektronide 12
liikumist, kuigi tegelikult NADH läbi membraani ei kanna. Glütserofosfaadi süstik: 2 glütserofosfaadi dehüdrogenaasi funktsioneerivad koos kandes NADh-lt tsütosoolis saadud elektronid üle FAD-le
mitokondris . Malaat-
aspartaat süstik: Lehekülg
oksaloatsetaat taandatakse tsütosoolis NADH-i arvel malaadiks, mis transporditakse läbi sisemembraani maatriksisse, kus + reoksüdeeritakse konverteerides NAD NADh-ks.






































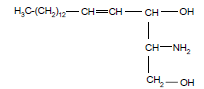




















Kõik kommentaarid