Ensüümid .. on
bioloogilised katalüsaatorid , mille peamiseks
ülesandeks elusorganismis on keemiliste reaktsioonide kiirendamine.
.. on
valgud ..ei saa käivitada termodünaamiliselt
võimatut protsessi
.. ei mõjuta reaktsiooni
kulgemise suunda
Ometi ensüümid kontrollivad ainevahetusprotsesside üldist suunda,
sest nende aktiivsus sõltub organismi vajadusest ja ühed
reaktsioonid ei kesta kogu aeg vaid muutuvad.
Ensüümide katalüüsivõime aluseks on nende omadus alandada
reaktsioonide
aktivatsioonienergiat. Aktivatsioonienergia on
energia, mis on vajalik reageerivate ainete ergastamiseks.
Ensüümidele on iseloomulik
spetsiifilisus:
- Stereokeemiline spetsiifilisus (eristatakse D- ja L-isomeere)
- Sidemespetsiifilisus (ensüümid võivad katalüüsida ainult teatud sidemete tekkimist ja lagunemist nt a1,4 glükosiidside )
- Rühmaspetsiifilisus (kindla funktsionaalse rühmaga toimuvad reaktsioonid)
- Absoluutne spetsiifilisus (eelnimetatud aspektide kominatsioon)
Ensüümide klassifikatsioon:
- Oksüdoreduktaasid -> katalüüsivad biokeemilisi redoksreaktsioone
- Transferaasid -> funkts rühmade ülekanne ühelt molekulilt teisele
- Hüdrolaasid –> katalüüsivad hüdrolüüsi (C-N, C-O, C-P, C-S sidemete lõhustamist vee toimel)
- Lüaasid -> katalüüsivad C-C, C-O, C-N,C-S jt mittehüdrolüütilist lõhustamist
- Ligaasid -> katalüüsivad sünteesireaktsioone
- Isomeraasid
Ensüümi toime:
E+S - > ES E-
ensüüm S-
substraat ES-substraadi
kompleks ES->EI EI- üleminekukompleks
EI- >EP EP- produktikompleks
EP-> E+ P P – produkt
Selle asemel , et teha kõrge akt.energiga reaktsiooni EP ,
sooritatakse mitu madalama akt.energiaga reaktsiooni , mis kulgevad
kiiremini.
Ensüümi
molekulaarne aktiivsus – üksiku ensüümimolekuli
poolt kindla ajaühiku vältel muundatud substraadimolekulide arvu
näitamine
Kõige aktiivsem karboanhüdraas (süsihappegaasi ja vee reageerimine
süsihappeks – CO2 eemaldamine kudedest, transport verega
kopsudesse ja organismist väljutamine)
Aktiivtsenter- piirkond, millel on võime siduda substraadi
molekuli ensüümiga ning mis omab katalüütilist toimet. Ruumiline
komponent , mis moodustub lähestikku paiknevate aminohappejääkide
külgahelatest. NB! Ruumiline
lähedus , mitte keemiline!
Aminohappejäägid jagatakse nelja kategooriasse:
- Siduvad aminohappejäägid -> paiknevad aktiivtsentris ja funktsioon on substraadi molekuli sidumine
- Katalüütilised aminohappejäägid -> paiknevad aktiivstsentris, avaldavad katalüütilist toimet
- Struktuuri säilitavad ahj -> tagavad valgu tertsiaare (v kvaternaarse) struktuuri , seega ka aktiivtsentri formeerumise ja püsimise
- Väheolulise ahj -> paiknevad nt ruumilise struktuuri pinnal, suht mõttetud
Liitensüümide puhul kuulub molekuli koostisesse ka teatav
mittevalguline komponent. (
apoensüüm – ensüümi valguline
osa;
kofaktor – mittevalguline osa)
Koensüüm – juhul kui mittevalguline osa kujutab endast
suhteliselt suurt orgaanislist molekuli (mitte lihtsalt mõnd iooni),
siis nimetatakse sellist osa koensüümiks.
Holoensüüm – terviklik ensüümimolekul, mis sisaldab
endas nii apoensüümi kui ka kofaktorit või koensüümi
Koensüümide
põhilisteks struktuurseteks ja funktsionaalseteks
komponentideks on erinevad veeslahustuvad
vitamiinid nt NAD ja NADP -
> vitamiin B5
derivaadid .
NB! Vesiniku aatomi
liitmine või loovutamine tähendab
redoksreaktsiooni!
Orgaanilised ühendid võivad dehüdrogeenimisel vabastada vesiniku
aatomeid, vesinikioone ja ka hüdriidioone.
Peamised koensüümid, mis on seotud redoksidega on NAD+
ja FAD. FAD seob vesiniku aatomeid. NAD+ seob
hüdriidioone. FAD redutseeritakse FADH2. NAD+
redutseeritakse NADH+H+. Seega NAD+ ja FAD
võivad eksisteerida nii
oksüdeeritud kui redutseeritud
koensüümina.
Transferaaside koensüümidest Co A (ainevahetusrajad!) +
regulatoorne funktsioon. Tema kõrge konts rakus tähendab energia
küllastatust , mis tähendab et energeetiliste protsesside
aktiivsust tuleb alandada.
Ensüümi aktiivsus: 1
katal (kat) on võrdne ensüümihulgaga, mis
on võimeline muundama 1mmol substraati produktiks 1 sekundis.
Aktiivsust iseloomustatakse reaktsiooni kulgemise kiiruse alusel.
Seda saab mõõta substraadi hulga vähenemise või produkti hulga
suurenemise määraga ajaühikus.
Vmax - >
maksimaalne reaktsiooni kiirus.
Substraadi lisamisega reaktsiooni kiirus enam ei suurene.
Km ->
Michaelise konstant (poolele maksimaalsest
kiirusest vastav substraadi konts)
Vmax ja Km on kineetilised
parameetrid . Km
peegeldab ensüümi afiinsust (keemilise sideme tugevust) substraadi
suhtes. Mida madalam on selle väärtus, seda kõrgem on ensüümi
afiinsus .
Ensüümi aktiivsus on otseses sõltuvuses ensüümi enda
kontsentratsioonist uuritavas materjalis.
Inhibiitorid on ühendid, mis mõjutavad ensüümi aktiivsust
alandamise suunas, aktivaatorid suurendavas suunas.
Mittespetsiifilised inhibiitorid (tugevad happed ja
leelised )
alandavad praktiliselt mistahes ensüümi aktiivsust.
Spetsiifilised inhibiitorid
toimivad teatud ensüümide suhtes. Füsioloogilistes
tingimustes on tähtsamad just spetsiifilised
inhibiitorid.
Inhibeerimine võib olla pöörduv või pöördumatu - >
pöörduval inhibiitor ensüümiga kov.
sidemeid ei moodusta ning
ensüümi aktiivsus on taastatav, pöördumatul moodustatakse kov
sidemed ninh langus on pöördumatu (mittespets inhibiitorite
korral).Konkureerivad ja mittekonkureerivad inhibiitorid.
Konkureerivad inhibiitorid on substraadiga sarnase struktuuriga ja
nad seonduvad hõlpsasti ensüümi aktiivtsentriga.
Mittekonkureerivad ei ole struktuurselt sarnased, aga on võimelised
selle aktiivsust pidurdama (ensüüm säilitab võime substraati
siduda, aga tema katalüütiline võime langeb).
Fosforüülimine ja defosforüülimine. Fosforüülimist
katalüüsivad proteeini kinaasid. Defosforüülimist proteiini
fosfataasid.
Allosteeriline regulatsioon . Allosteeriline ehk regulatoorne
tsenter. Allosteeriline tsenter on võimeline siduma talle
keemiliselt
struktuurilt sobivaid ühendeid. Seal ei toimu aga mingit
katalüüsiprotsessi, seega neid ühendeid, mis seonduvad ensüümiga
selle tsentri kaudu ei nimetada substraatideks vaid allosteerilisteks
regulaatoriteks, sest nende sidumine mõjutab aktiivtsentri seisundit
ja ensüümi aktiivsust.
Vesinikioonide kontsentratsioon. Ensüümid on tundlikud
vabade vesinikioonide kontsi suhtes keskkonnas, sest säilitavad
katalüüsivõime suhteliselt kitsas pH vahemikus. Ensüümi
aktiivsuse languses
vales keskkonnas kaks põhjust: esiteks võib
ensüümi molekuli struktuur muutuda ja lakata olemast ning teiseks võib pH nihe muuta happeliste ja aluseliste aminohappejääkide
külgahelate seisundit (ionistatsiooni) ensüümi aktiivtsentris.
Temperatuur. Suhteliselt madala tempratuuri korral kaasneb
tempi tõusuga ensüümi aktiivsuse suurenemine, optimaalse tempi
ületamisega kaasneb aktiivsuse järsk langus. Temperatuuri
stimuleeriv toime on seotud molekulide seisundi muutumisega,
aktiveerumisega. Toime täielik lakkamine on seotud
denatureerumisega.
Isoensüümid ehk isosüümid . Isoensüümid on ensüümide
erivormid, mille erinevad molekulaarsed vormid katalüüsivad sama
keemilist reaktsiooni sama mehhanismi kaudu, ent erinevate
kineetiliste parameetritega.
Hormoonid
... on bioloogilselt aktiivsed ühendid, mille pamine ülesanne
paljurakulises organismis on erinevate rakkude talitluse
koordineerimine .
Hormoone produtseeritakse
sisenõre - ehk endokriinnäärmetes ning
eritatakse otse verre. Verega transporditakse hormoonid kudedesse,
kus nad mõjutavad rakkude ainevahetusprotsesse. On ka
erandeid .
Mõnede hormoonide toime ei ulatu tekkekohast eriti kaugele ja
mõjutavad vaid neid produtseerinud raku kõrval või lähikonnas
paiknevate rakkude ainevahetust või siis toimimad vaid neid
sünteesinud raku siseselt.
Kõik hormoonid ei teki sisenõrenäärmetes. Ka teistest kudedes on
rakkusid, mis sünteesivad hormoonide funktsioone täitvaid aineid.
Neid nimetatakse
koehormoonideks. (nt
gastriin , sekretiin)
Vastavalt tegevusraadiusele saab hormoone klassiitseerida järgnevalt:
- Endokriinsed hormoonid – produtseeritakse sisenõrenäärmetes, läbivad vereringes pikki vahemaid ja mõjutavad ka kaugel paiknevaid rakke.
- Parakriinsed hormoonid – eemalduvad tekkekohast vähe, toimivad naabruses olevate rakkude suhtes.
- Autokriinsed hormoonid – toimivad peamiselt samas rakus, kus nad on sünteesitud.
Hormoonide keemiline struktuur. Hormoonid võivad olla peptiidid ja
valgud (nt insuliin,
glükagoon , kasvuhormoon), aminohappe türosiini
derivaadid (
katehhoolamiinid –
epinefriin (adrenaliin) ja
noradrenaliin ; türoksiin ja
trijoodtüroniin ), steroidid
(
neerupealise koore hormoonid ja suguhormoonid) ja arahhidoonhappe
derivaadid (
arahhidoonhape on polüküllastumata
rasvhape ). NB!
Steroididel on steraantuum.
Hormoonide transport veres. Valdava enamuse hormiinode transport
toimub vere kaudu. Hormoonide transpordiviis veres sõltub nende
keemilisest ehitusest. Nt valgulise ehitusega ja katehhoolamiinid toimetatakse edasi molekulidena.
Steroid - ja türeoidhormoonid
moodustavad komplekse spetsiaalsete transpordivalkudega.
Bioloogiliselt aktiivne on ainult vaba
hormoon st hormoon, mis ei ole
seotud teiste ainetega.
Hormoonide
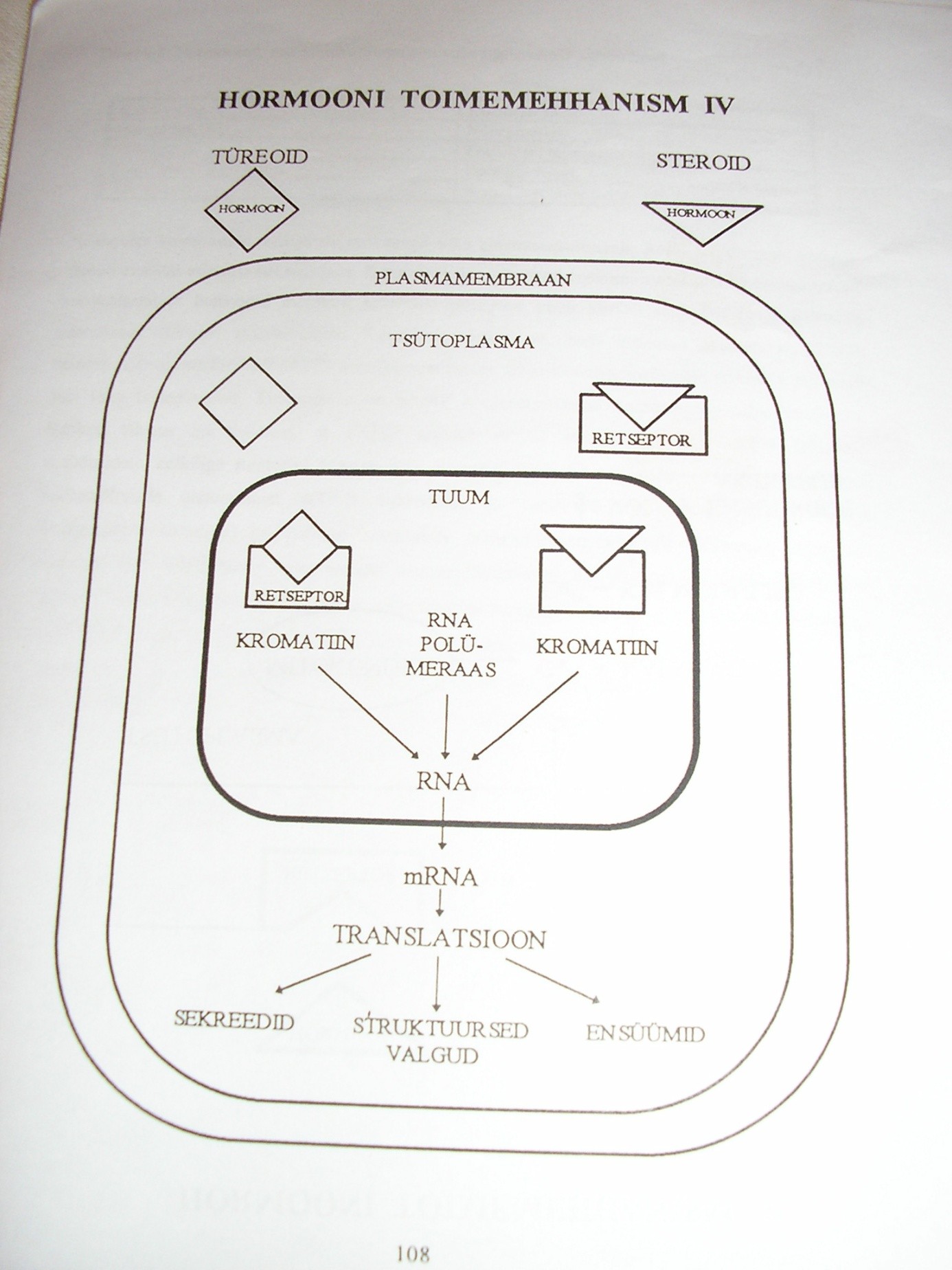
retseptorid . Retseptorid kujutavad endast suuri
valgumolekule, mis on võimelised neile spetsiifilisi hormoone endaga
siduma. Erinevate hormoonide retseptorite
paiknemine rakus on erinev.
Plasmamembraanis on retseptorid katehhoolamiinidele, peptiididele ja
valgulistele hormoonidele. Tsütoplasmas ja tuumas paiknevad
steroidhormoonide, valdavalt tuumas aga
türeoidhormoonide retseptorid. Retseptorite arv rakus varieerub. Lihasrakus võib
hormooni retseptorite arv muutuda ka treeningkoormuste mõju. Lisaks
arvule võib muutuda ka retseptorite afiinsus (st võime siduda
endaga hormoone. Retseptorite aktiivsust mõjutavad nt pH, ioonide
kontsentratsioon jne. Keemiliselt struktuurilt on enamus retseptorid
glükoproteiinid.
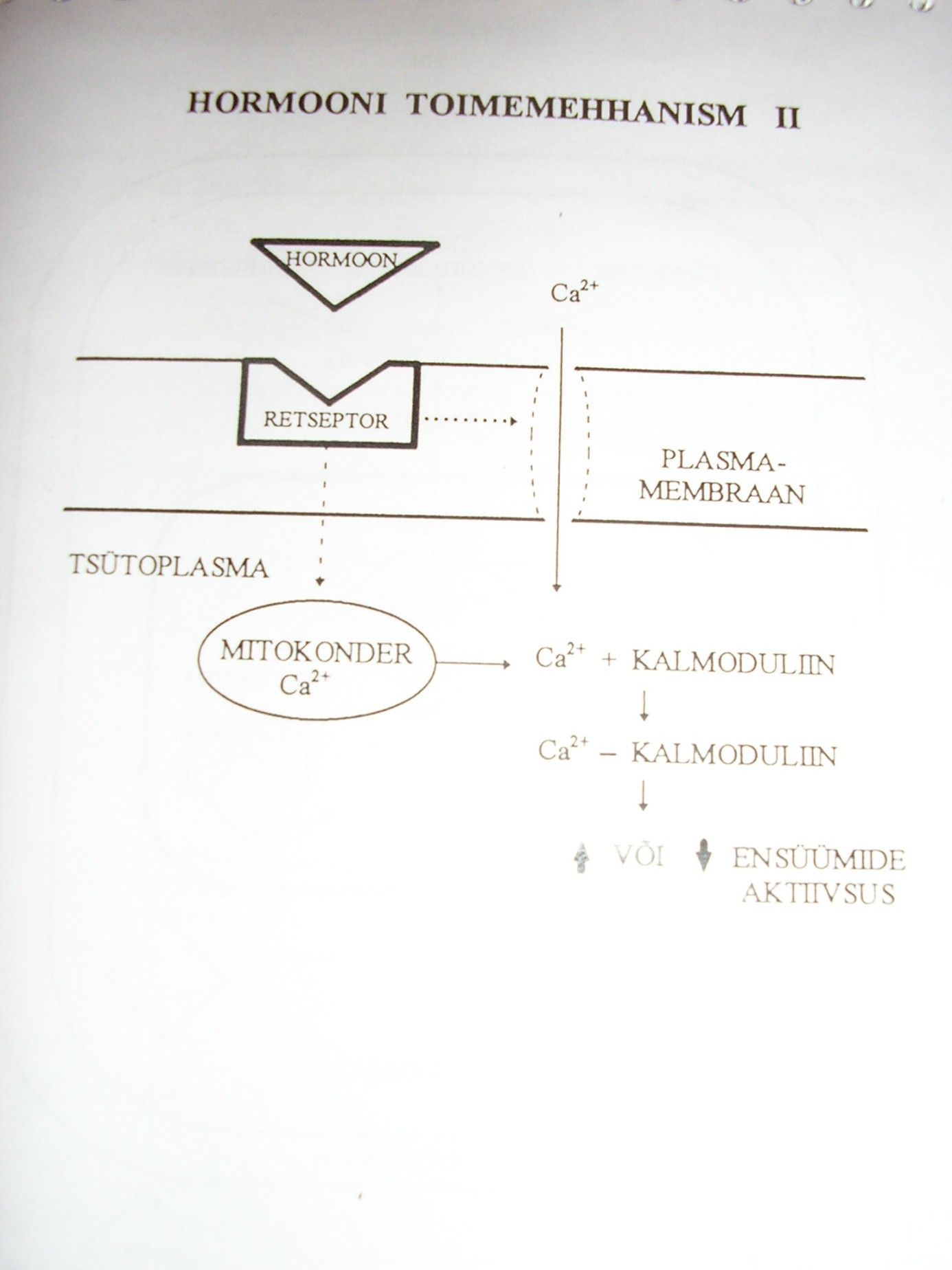
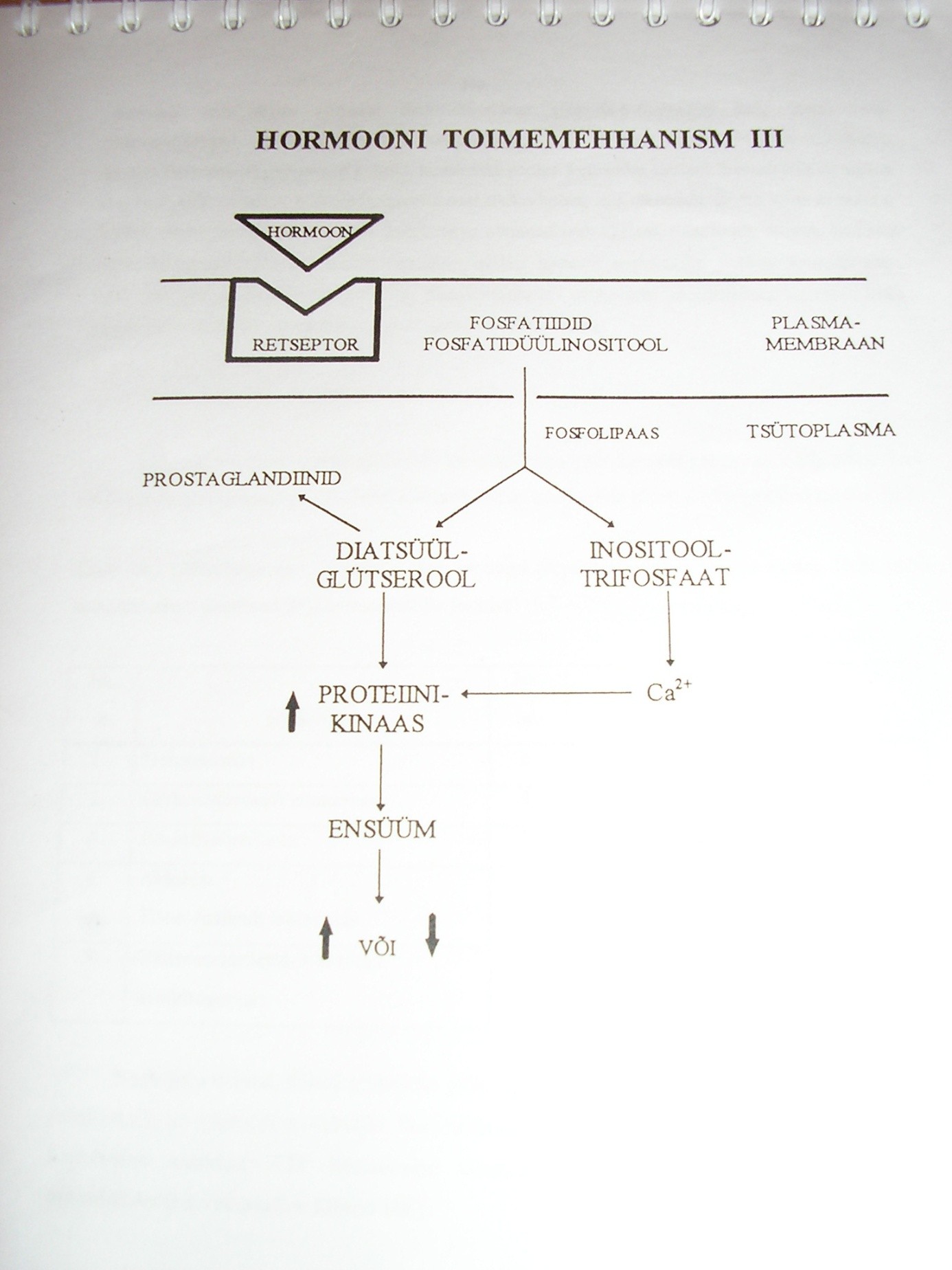
Hormoonide toime raku ainevahetusele.
Hormoonide toime seisneb selles, et nad muudavad erinevate ensüümide
aktiivsust rakus. Seda tehakse:
- Olemasolevate ensüümimolekulide struktuuri muutmisega, kutsudes sellega esile inaktiivse ensüümi transformeerumise aktiivseks vormiks
- Ensüümimolekulide arvu muutmisega rakus nende sünteesi stimuleerides.
- Substraatide kättesaadavuse muutmisega ensüümimolekulidele nt rakumembraani läbitavuse muutmise kaudu.
Hormoonide toimemehhanismid. Õp 103-108
Ainevahetusrajad
Glükolüüs Glükolüüs on glükoosi
anaeroobse lagundamise protsess, mille
tulemusel ühest glükoosi molekulist tekib kaks molekule laktaati.
Iga glükoosi lagundamise käigus vabaneb energia, mille arvelt
toodetakse 2 molekuli ATP-d.
Kokkuvõtlikult :
GLÜKOOS + 2ADP + 2P ->2
LAKTAAT + 2H+
+
2ATP Glükolüüsi käigus vabaneb vaid 7% energiast ning sellest 60%
kasutatakse vaid ATP sünteesiks.
Glükolüüsi protsessis võib eristada üldistatult peamiselt kahte
staadiumit. Esiteks glükoosi fosforüülimine glükoos-6 fosfaadiks
ning jagunemine kaheks nii et tekib kaks molekuli
glütseeraldehüüd-3-fosfaati. Selle käigus kasutatakse ATP-d ning
2ATP-d muundatakse 2ADP-ks. Teises staadiumis toimub
glütseeraldehüüd-3-fosaadi ümbertöötlemine kaheks laktaadi
molekuliks, millega kaasneb 4ADP molekuli fosforüülimine ATP-ks ehk
siis kogu protsessi summaarne energeetiline efekt on 2ATP-d.
Skemaatiliselt:
glütseeraldehüüd-3-
fosfaat 2ADP->2ATP laktaat
GLÜKOOS 2ATP->2ADP
Glütseeraldehüüd-3-fosfaat 2ADP-2ATP laktaat
Kahjuks nii lihtne asi ei ole.
Pikemalt siis.
Algab kogu protsess glükoosi molekuli fosforüülimisega (glükoos -
> glükoos-6-fosfaat). Vajalik fosaatrühm saadakse ATP
lõhustamise tulemusena (ATP->ADP). Sellega tegeleb
heksokinaas.
Järgnevalt toimub isomerisatsioon (glükoos-6-fosfaat ->
fruktoos -6-fosfaat). Sellega tegeleb
glükoosfosfaadi isomeraas.
Seejärel see ühend fosforüülitakse uuesti lisandub veel üks
fosfaatrühm ning tekib fruktoos-1,6-bisfosfaat, mille käigus
lõhutakse jälle ATP-d (ATP->ADP). Seda reaktsiooni katalüüsib
fosfofruktokinaas. Seejärel saab tööd
aldolaas, mis
lagundab fruktoos-1,6-bisfosfaadi glütseeraldehüüd-3-fosfaadiks ja
dihüdroksüatsetoon fosfaadiks. Viimane muundub
trioosfosfaadi
isomeraasi mõjul samuti glütseeraldehüüd-3-fosfaadiks ning
glükolüüsi rada jätkub kahe identse rajana. Siis
glütseeraldehüüd-3-fosfaat oksüdeeritakse
glütseeraldehüüd-3- fosfaadi dehüdrogenaasi poolt
1,3-bisfosfoglütseraadiks. Selles reaktsioonis osaleb koensüümina
NAD+, mis redutseeritakse NADH+H+.
Seejärel kantakse 1,3-bisfosfoglütseraadilt
fosfoglütseraadi
kinaasi toimel üks fosfaatrühm üle ADP-le, mille tulemusena
tekib ATP ja 3-fosfoglütseraat. Sellega on
reaktsioon energeetiliselt tasakaalustunud (kulutatud on 2 molekuli ATP-d ja
sünteesitud samuti 2 molekuli ATP-d). See fosfaatrühm kantakse
fosfoglütseraadi kinaasi toimel üle teisele süsinikule,
tekib 2-fosfoglütseraat, millest
enolaasi toimel saadakse
fosfoenoolpüruvaat. Sellest võtab
püruvaadi kinaas ära
fosfaatrühma, mille ta seob ADP-ga (tekib ATP) ning
fosfoenoolpüruvaadist saab lihtsalt
püruvaat . Püruvaat
redutseeritakse laktaadiks laktaadi
dehüdrogenaasi abil ning
samal ajal oksüdeeritakse NADH +H+ ning selle reaktsiooni
produkt NAD+ saab osaleda juba järgmise molekuli
lagundamisel.
Et kogu see pikk jutt kokku võtta toimub glükolüüs peamiselt
kolme liiki reaktsioonidena. Esiteks glükoosi süsinikahel
lammutatakse,
fosfaatrühmad kantakse ühenditelt üle, et
produtseerida ATP-d ning sobstraadi molekulidelt võetakse vesinikke
ja seotakse need NAD+ abil püruvaadile.
Fosforüülitud ühendid on sellepärast olulised, et fosfaatrühmad
on tavaliselt ioniseeritud ning selle tõttu ei saa läbida
rakumembraani ning seega hoiavad fosforüülitud ühendid neid aineid
raku sees ega lase neil imbuda väliskeskkonda. Enamjaolt on kõik
vajalikud ensüümid raku tsütoplasmas ning see võimaldab
reaktsioonil väga kiiresti toimida.
GlükogenolüüsKui glükolüüs oli ainevahetusrada, mis lähtus glükoosist, siis
glükogenolüüs on ainevahetusrada, mis lähtub glükoosi
polümeerist glükogeenist. Valdavalt need reaktsioonid kattuvad, aga
glükogenolüüsi tulemusel sünteesitakse 3ATP molekuli iga
glükoosijäägi kohta, mis teeb selle energeetilisemalt
kasulikumaks.
Kõigepealt glükogeeni fosforülaas katalüüsib
a-1,4-glükosiidsidemete hüdrolüüsi, mille käigus vabanev
glükoosijääk ühtlasi ka fosforüülitakse ning produktiks on
glükoos-1-fosfaat. Fosfaatrühm ei võeta mitte ATP-lt vaid
ortofosforhappelt ning seetõttu ongi energeetiliselt reaktsioon
kasulikum. Glükogenolüüsirajal saadakse glükoos-6-fosfaat
lihtsalt fosfaatrühma ümberpaigutamise tulemusena. Seda katalüüsib
fosfoglükomutaas. Edasi toimub kõik analoogselt
glükolüüsirajaga.
Kuna glükoos on inimese keha rakkudele esmajärgulise tähtsusega
energiaallikas , siis on organismi normaalse talitluse tagamiseks
oluline kindlustada stabiilne glükoosi kontsentratsioon veres. Eriti
sõltuvad sellest erütrotsüüdid. Vere glükoositaseme säilitamisel
on oluline roll maksarakkudel. Teatavasti asuvad just maksas
glükogeeni varud ning glükogeenist glükoosi saamine põhineb
glükoos-6-fosfataasi olemasolul. Tema ülesanne on võtta
glükoos-6-fosfaadist ära fosfaatrühm ning seega on produktiks
glükoos. Seega, kui tekib glükoosi puudus, siis on võimalik
glükoos-6-fosfaati glükogenolüüsi
rajalt eemale juhtida ja
sellest glükoosi sünteesida.
Glükolüüsi ja glükogenolüüsi regulatsioonGlükolüüsi ja glükogenolüüsi intensiivsust reguleeritakse
vastavalt raku energiavajadusele. Regulatsioon realiseerub osaliselt
reaktsioonidele vajalike substraatide kontsentratsiooni muutuste
kaudu. (meaning mida rohkem substraati, seda suurem on reaktsiooni
intensiivsus, sest kõik tuleb ju ära kasutada). Enim muutub
reaktsiooni kiirus aga erinevate ensüümide aktiivsuse muutumisel.
Kõige rohkem kontrollitakse fosfofruktokinaasi ning heksokinaasi
aktiivsust.
Praktiliselt kõikide glükolüütiliste ensüümide (va aldolaas)
ühiseks tunnuseks on see, et nad vajavad toimimisega Mg2+
ioonide juuresolekut.
Süsivesikute
aeroobne
oksüdatsioon Tsitraaditsükkel ja hingamisahela ensüümidGlükolüüsi protsessis vabaneb glükoosi molekulist ainult 7%
võimalikust energiast ehk siis püruvaat (või laktaat) hoiab endas
ülejäänud energiat. Siis kui hapnikupuudust rakus ei teki, ei ole
vaja ka püruvaadist tingimata laktaati moodustada ning püruvaat
transporditakse hoopis mitokondrisse, kust toimub tema oksüdatiivne
dekarboksüülimine (meaning hapniku juuresolekul võetakse talt ära
karboksüülrühm). Seda protsessi katalüüsib ensüümide kompleks
püruvaadi dehüdrogenaas. Selle tulemusel eraldub üks
molekul CO2 ning ülejäänud osa liidetakse koensüüm
A-ga, mille tulemusel tekib
atsetüül CoA. Ehk siis:
PÜRUVAAT + NAD+ + CoA - > ATSETÜÜL CoA + NADH + H+
+ CO2
Atsetüül CoA suundub aga tsitraaditsüklisse ehk Krebsi tsüklisse.
Tsitraaditsüklis lammutatakse lõplikult algselt glükoosist pärinev
süsinikahel. Iga atsetüül CoA molekuli kohta eraldub 2 molekuli
CO2, samuti vabaneb energia, mille arvel sünteesitakse
üks molekul GTP-d, mis on energeetiliselt väärtuselt samaväärne
ATP-ga. Siis eemaldatakse veel erinevatelt vaheproduktidelt 4 paari
vesiniku aatomeid, mis seotakse NAD+ või FAD-i poolt
(suhtes 3:1). Nemad loovutavad oma vesinikut hingamisahela
ensüümidele, mis genereerivad neist 3ATP molekuli iga NAD+ ja 2ATP
molekuli iga FAD-i poolt hingamisahelasse kaasa toodud vesiniku
aatomite paari kohta.
NB! Aeroobsetes tingimustes siseneb hingamisahelasse ka algselt
glütseeraldehüüd-3 fosfaadi dehüdrogenaasi poolt katalüüsitavas
reaktsioonis NAD+-ga seotud vesiniku aatomite paar.
Vesinikud võtab endale hapnik ning üheks lõpp-produktiks on vesi.
Pikemalt tsitraaditsüklist...
Atsetüül CoA siseneb tsitraaditsüklisse ning reakeerib
oksaalatsetaadiga, reaktsioon toimub
tsitraadi süntaasi
toimel ning tekib tsitraat. Tsitraadis tõstab
akonitaas ümber
hüdroksüülrühma ning läbi vaheühendi cis-akonitaadi tekib
isotsitraat. Seejärel toimub isotsitraadi oksüdatiivne
dekarboksüülimine, mida katalüüsib
isotsitraadi dehüdrogenaas.
Reaktsiooni käigus eemaldatakse isotsitraadi molekulist vesiniku
aatomite paar, mis seotakse NAD+-ga ning eraldub üks
atsetüülrühma
süsinik CO2-na. Produktiks on
α-ketoglutaraat, millest
α-ketoglutaraadi dehüdrogenaasi
toimel tekib suktsinüül CoA. Selle protsessi käigus eemaldatakse
veel üks vesinikupaar, mis seotakse NAD+ ning eraldub
CO2.
Suktsinüül CoA süntaasi toimel lagundatakse
suktsinüül CoA suktsinaadiks ja koensüüm A-ks, millega kaasneb
GDP fosforüülimine GTP-ks.
Suktsinaadi dehüdrogenaas
oksüdeerib suktsinaadi fumaraadiks. Selles reaktsioonis võtab
vesinikepaari endale FAD. Fumaraat hüdratiseeritakse(!)
fumaraasi
toimel malaadiks, mis
malaadi dehüdrogenaasi toimel
oksüdeeritakse oksaalatsetaadiks. Seega on oksaalatsetaat nii
tsitraaditsükli lähteühend kui lõpp-produkt.
Elektronide ülekanne hapnikule hingamisahelasKogu selle eelneva jama energeetiline väärtus on 2GTP molekuli iga
glükoosi molekuli kohta, mida on vähe.
Peamine energiat tootev protsess on aga oksüdatiivne fosforüülimine,
mis toimub hingamisahelas elektronide ülekande vaba energia arvel.
Oksüdatiivne fosforüülimine tähendab seda, et midagi tuleb
oksüdeerida ja midagi fosforüülida.
Oksüdeerimine tähendab seda,
et vaheproduktidelt eemaldatakse vesiniku
aatomid ja kantakse need
üle hapnikule. See on järk-järguline protsess, milles esinevad
järgmised staadiumid:
- Primaarne oksüdeerimine - > dehüdrogenaaside toimel eemaldatakse vesinikud ning seotakse NAD+ ja FAD poolt. Seega nad redutseeruvad vastavald NADH+ H+ ja FADH2
- Intermediaarne oksüdeerimine - > koensüümide oksüdeerimine ehk neilt võetakse vesiniku aatomid ära ja antakse need üle hingamisahela ensüümide süsteemile.
- Terminaalne oksüdeerimine - > vesiniku aatomid kantakse üle hapnikule, mille tulemusel moodustub vesi
Fosforüülimine on ADP molekulile fosfaatrühma lisamine kasutades
selleks ortofosforhapet ja hindamisahela vesinike aatomite (õigemini
nende elektronide) ülekande vaba energiat. Elektronid transpordituna
ensüümilt ensüümile jõuavad järjest madalamale energiatasemele
kuni lõpuks seotakse hapnikuga ning iga NADH + H+ poolt
transporditud elektronipaari ülekandel sünteesitakse kuni 3 ATP ja
FADH2 transpordil 2ATP-d, sest FADH2 loovutab
enda elektronpaari energeetiliselt madalamal
nivool .
Kõiki neid
seoseid pole selgeks tehtud, aga Peter Mitchelli teooria
järgi kasutatakse vaba energiat prootonide
pumpamiseks sisemembraani
välisküljele, aga enamus polaarseid molekule ja ioone ei saa läbida
sisemembraani ning sellepärast tekib
elektrokeemiline gradient
membraani sise- ja väliskülje vahel. See avaldub selles, et
välisküljel on suurem positiivne laeng (
gradiendi elektriline
komponent) ja suurem vesinikuioonide kontsentratsioon (gradiendi
keemiline komponent). Mitokondri sisemembraanis on aga valke, mis
suudavad neid prootoneid jälle gradiendi surve toimel sisse tuua ja
selle prootonite voo energia arvelt toimubki
ATP süntaasi
abil ADP fosforüülimine. ATP süntaas koosneb kahest subühikust,
üks neist (F0) paikneb sisemembraanis ja moodustab poori, mille
kaudu
prootonid saavad maatriksisse sisenega ning F1, mis katalüüsib
ATP sünteesi.
Praktiliselt saab hingamisahelat jagada
neljaks kompleksiks. I
kompleks võtab NADH + H+ elektronid ja annab need
koensüüm Q-le. See on NADH dehüdrogenaas. Prootonid lasevad
alga sisemembraani välisküljele ja coQ redutseerub coQH2. II
kompleks (flavoproteiini hüdrogenaas) võtab elektronidepaari FadH2
ja annab selle ka coQ-le. Selle tulemusel prootoneid ei väljutata.
III kompleksis kannab reduktaas elektronid coQH2 kahele tsütokroom
c-le ja IV kompleksis võtavad kaks tsütokroomi a ja a3 elektronid
vastu tsütokroom c-lt ning annavad hapnikule, mille mõlemad aatomid
võtavad 2 elektroni.
ATP, ADP ja Pi transport läbi membraaniATP ja ADP transport toimub mitokondri sisemembraanis paikneva
ATP-ADP translokaasi toimel üheaegselt erinevates suundades.
Ortofosforhappe transport seostatakse aga prootonide liikumisega ja
seda transportvalku nimetatakse
Pi-H+
translokaasiks. ATP on negatiivsema laenguga (4-)ja sellepärast
liigub positiivsemale alale ehk membraani välisküljele ning ADP
(3-) liigub sellest vabaneva energia tulemusel sisse. Pi
transportimiseks vajalik energia pärineb prootonite voost.
Rasvhapete
oksüdatsioon
Rasvhapete energiat saab kasutada ainult aeroobsetes tingimustes.
Kuna kõik aeroobsed ainevahetusprotsessid toimuvad mitokondrites
(sest ainult seal on sobivad ensüümid), siis tuleb rasvad ka
transportida kõigepealt mitokondrisse. Selleks moodustub
tsütoplasmas rasvhappe jäägi ja CoA kompleks – atsüül CoA. Iga
molekuli atsüül CoA tekkeks kulutatakse 2ATP hüdrolüüsi jagu
energiat. Mitokondri sisemembraani läbimiseks moodustub veel
transpordikompleks karnitiiniga, atsüülkarnitiin, millest
mitokondri maatriksis taastatakse atsüül CoA.
Rasvhappe molekuli (atsüül CoA) lagundamine toimub järk-järgult
ning selle protsessi käigud eemaldatakse rasvhappe süsinikahelast
kahe C aatomi pikkused
fragmendid ning töötatakse need ümber
atsetüül CoA ühikuteks. Need fragmendid eraldatakse β-süsiniku
juurest ja seda protsessi nimetatakse
rasvhapete
β-oksüdatsiooniks. Tekkinud atsetüül CoA lagundamine toimub
analoogselt süsivesikute aeroobse oksüdatsiooniga tsitraaditsüklis.
Iga atsetüül CoA molekuli produtseerimisega β-oksüdatsiooni
käigus kaasneb kahe paari vesiniku aatomite eemaldamine rasvhappe
molekulist, millest üks seotakse FAD-iga ja teine NAD+-iga
ja need elektronid antakse üle hingamisahelasse ja nende arvelt
toodetakse ATP-d nagu süsivesikutegi puhul.
Rasvhappe molekuli täielikul
oksüdatsioonil vabaneva energia arvel
produtseeritava ATP hulk sõltub rasvhappe süsinikahela pikkusest.
Pikemalt β-oksüdatsioonist
Esimest reaktsiooni katalüüsib
atsüül-CoA dehüdrogenaas,
mis paikneb mitokondri sisemembraani sisepinnal. Selle kigud
eralduvad atsüü-CoA kaks vesiniku aatomit, mis seotakse FAD-iga
ning atsüülrühma α ja β süsinike vahele tekib kaksikside.
Saadud ühendit nimetatakse trans-enoüül-CoA-ks. Teise
reaktsioonina hüdratiseeritakse trans-enoüül-CoA
enoüül –CoA
hüdrataasi toimel 3-hüdroksüatsüül-CoA-ks. Järgnevalt
oksüdeeritakse 3-hüdroksüatsüül-CoA 3-ketoatsüül-CoA-ks. Seda
katalüüsib
3-hüdroksüatsüül-CoA dehüdrogenaas .
Koensüümina toimib siin NAD+, mis redutseeritakse.
Seejärel
atsüül-CoA tiolaas katalüüsib reaktsiooni, mille
käigus liidetakse β-süsinikule 3-ketoatsüül-CoA molekulis CoA,
mille tulemusel vabaneb kahe süsiniku pikkune
fragment atsetüül-CoA
ning tekib uus lähteühendist 2 süsiniku võrra lühem atsüül-CoA
molekul ning reaktsioon hakkab uuesti.

































Kõik kommentaarid