3. VARS 3.1.
Morfogenees ja steeliteooriaVarre
varajane histogenees vastab
eespool kirjeldatule. Edasisel
diferentseerumisel, kasvukuhikust kaugemal on märgata rakkude
jagunemist nelja suuremasse rühma (joonis). Välimist, tuunikast
tekkinud kihti nimetatakse
protodermiks,
sellest kujuneb
epiderm . Protodermist seespool paikneb korpusest
pärinev
proparenhüüm,
millest tekivad kõik esikoore
koed , edasi tuleb
püsimeristeem,
mis areneb prokambiumiks ning
algsäsi, mis
muutub edasise arengu käigus säsiks.
Prokambiumist
tekivad kesksilindri välimised
kihid -- protofloeem, uus
prokambium ja protoksüleem.
Püsimeristeemi
ja algsäsi derivaate nimetatakse
kesksilindriks
ehk
steeliks.
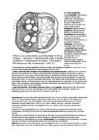
Steelitüüpe eristatakse floeemi ja ksüleemi paigutuse järgi,
nende evolutsioonist annab ülevaate joonis.
Kõik
steelid on evolutsioneerunud
protosteelist,
mis koosneb sambana kulgevast ksüleemist ja seda ümbritsevast
floeemist. Protosteel esineb tänapäeval üksikutel sõnajalgtaimedel
varre alumises osas ning vees kasvavatel katteseemnetaimedel.
Protosteelist
kujunes
aktinosteel
(kiirjas kesksilinder), kus ksüleem on sopistunud floeemi vahele.
Praegu elavatel soontaimedel esineb see
juurtes . Kui ksüleem esineb
kitsaste ribadena, nagu koldade (
Lycopodium )
vartes, räägitakse
plektosteelist
(põimjas kesksilinder).
Teine,
evolutsiooniliselt
edukam arengutee algab
sifonosteeliga
(torusteeliga), kus
juhtkude ümbritseb põhikudet -- säsi. Kui
floeem esineb ksüleemist ainult väljaspool, on tegemist
ektofloidse
sifonosteeliga, kui
aga mõlemal pool ksüleemi, siis
amfifloidse
sifonosteeliga.
Viimasest
on edasi arenenud
diktüosteel
(polüsteel). Siin on ebakorrapärase suurusega harusteelide ksüleem
ümbritsetud floeemist. Diktüosteel esineb sõnajalgtaimedel.
Kaasaegsete
kaheiduleheliste taimede vartes esineb kollateraalsetest
juhtkimpudest koosnev
eusteel,
üheidulehelistel aga korrapäratult paiknevatest juhtkimpudest
moodustunud
ataktosteel.
Artrosteel
on sisuliselt
eusteeli erijuht, siin on kollateraasete
juhtkimpude asemel bikollateraalsed.
Olulised
mõisted morfogeneesi käsitlemisel on
esikasv
ja
teiskasv
ehk resp.
primaarne
ja
sekundaarne
kasv.
Esikasvu
moodustuvad kõik koed, mis saavad alguse kasvukuhikust
äsjakirjeldatud viisil. Mitmeaastastel taimedel võib esikasvuks
pidada valdavat osa esimesel eluaastal tekkinud kudedest.
Teiskasv
seisneb juhtkimpude mahusuurenemises, mille tagajärjel vars jämeneb,
kuid ei pikene. Teiskasv algab harilikult teisel eluaastal (sõltub
kliimast ja taime liigilistest iseärasustest). Tekkivate kudede
nimetuse esimene sõna on teis-, näiteks teispuit, teissäsikiired.
3.2.
Varre esikasvPrimaarse ehitusega
varsi ümbritseb
epiderm,
mille
rakukestad on kutiniseerunud.
Epidermi välispinda katab
kutiikula. Õhulõhed ja
karvad on varre epidermile vähem
iseloomulikud kui lehtedele.
Epidermi
all on põhikoest koosnev
esikoor,
mida kloroplastide esinemisel nimetatakse
klorenhüümiks,
suurte rakuvaheruumide korral ka
aerenhüümiks.
Esikoore põhikude võib talitleda säilituskoena. Vahel esineb siin
kivisrakke ning kaltsiumoksalaadi kristalle. Esikoore välises,
vahetult epidermi all asetsevas kihis paikneb sageli kollenhüüm.
Enamasti ei moodusta kollenhüüm lausalist rõngast, vaid esineb
ribadena varre kantidel. Kõrrelistel ja mõnedel okaspuudel, harvem
teistel taimedel, on esikoores esinevaks tugikoeks sklerenhüüm.
Esikoore
sisemine rakkude kiht erineb ülejäänud esikoorest: selle
rakud on
pisut väiksemad,
tihedamalt üksteise kõrval ja neljakandilised.
Seda kihti nimetatakse
endodermiks,
rohke tärklisesisalduse tõttu ka tärklistupeks. Endodermis võib
esineda ka Caspary jooni.
Esikoorest
seespool asub varre
kesksilinder
(
steel ), mille tüüpide kirjeldus on toodud eelmises peatükis.
Steeli välimine
rakukiht on
peritsükkel
(perikambium). Siit algavad
lisajuured ja -pungad. Mitmekihilise
peritsükli rakkudest võib kujuneda
perivaskulaarne
tugikude,
(enamasti niinekiud) ning säsi. Sellise päritoluga
mehaaniline kude
paikneb
taimes enamasti üksikute kimpudena, harvem moodustub pidev
silinder. Piir esikoore ja kesksilindri vahel ei tarvitse olla
selgelt eristatav.
Juhtkimbud
asuvad kesksilindris kas ühe korrapärase ringina (kaheidulehelised)
või korrapäratult laialipillatuna (üheidulehelised). Nende arv
sõltub taimeliigist ning kasvutingimustest.
Üheiduleheliste
taimede korrapäratult
asetsevate juhtkimpudega kesksilindrit
nimetatakse
ataktosteeliks.
Varre välispinna lähedal on juhtkimbud ja põhikoe rakud väiksemad,
varre keskosas suuremad. Kui vars on õõnes, nagu enamikul
kõrrelistest (
Poaceae),
võivad ka üheidulehelistel juhtkimbud olla paigutunud suhteliselt
korrapäraselt.
Kaheiduleheliste
taimede juhtkimbud on avatud,
ksüleemi
ja
floeemi
vahele jääb ühe- või mitmekihiline
kambium .
Kambiumirakud on prosenhüümsed, ristlõikes lamedad, enamasti
teritunud tippudega, õhukesekestalised, üleni protoplasmaga
täitunud ja suuretuumalised. Juhtkimpudes esineb
kimbu-
ehk
fastsikulaarkambium,
juhtkimpude vahel, säsikiirtes võib tekkida
kimbuvahe-
ehk interfastsikulaarne
kambium.
Nende kahe
kambiumi servad liituvad ja nii moodustub varres pidev
kambiumsilinder (puittaimedel on see juba algusest peale).
Kambiumirakud
poolduvad nii tangentsiaalselt,
andes uusi floeemi ja
ksüleemirakke, kui ka radiaalselt, vältimaks kambiumirõnga
katkemist varre jämenemisel. Puhkeperioodide ajal on kambium
enamasti ühekihiline.
Ksüleem
sisaldab
trahheiide
ja
trahheesid.
Vanimad ksüleemi elemendid (
trahheiidid ) asuvad varre keskosas.
Kaheiduleheliste taimede ksüleem sisaldab ka tugikudet --
puidukiude
(libriformi). Lisaks esineb ksüleemis põhikude, mille rakud on
vähemal või rohkemal määral
puitunud . Seda nimetatakse
ksüleemi
põhikoeks.
Floeemi
olulisemateks elementideks on
sõeltorud
ja
saaterakud.
Harilikult paikneb iga sõeltoru kõrval üks saaterakk. Floeemis
leidub ka tugikudet -- niinekiude ning põhikudet (floeemi põhikude).
Niinekiud on enamasti puitunud.
Juhtkimbud
võivad olla ümbritsetud põhikoelisest, sagedamini aga
sklerenhüümsest
kimbutupest.
See on
paksem juhtkimbu sisemisel ja välimisel küljel, kambiumi
kohal on aga kimbutupp õhuke ja võib seal olla isegi katkenud.
Steelis
asuvat põhikudet, mis jääb juhtkimpudest sissepoole ja nende
vahele, nimetatakse
säsiks
(
medulla ).
Juhtkimpude vahele jäävad säsikiired. Esikasvu perioodil tekivad
esi-säsikiired, need ulatuvad steeli keskosa täitva säsini.
Säsile
on tunnuslik hästiarenenud rakuvaheruumide olemasolu. Rohttaimede
säsirakud on õhukesekestalised ja elusad, puittaimedel need aja
jooksul enamasti
puituvad , surevad ja täituvad õhuga. Säsikiire
rakud säilitavad varuaineid, läbi nende kulgeb ainete
radiaalsuunaline transport. Säsi välimises osas paiknevad rakud on
väiksemad ja pikaealisemad, seda piirkonda nimetatakse vahel ka
perimedullaartsooniks. Kui varre keskosas säsirakud surevad
(rohttaimede puhul tavaline nähtus), tekib sinna
õõnsus.
Primaarse
ehitusega on ka kaheiduleheliste ning üheiduleheliste taimede
maa-alused varred ehk
risoomid .
Nende siseehitusele on iseloomulik tüseda esikoore põhikoe kihi
esinemine, hästiarenenud
endoderm ja peritsükkel. Väliselt eristab
risoomi juurest soomusjate leherudimentide olemasolu.
3.3.
Varre teiskasvVarre
teiskasv (joonis) seisneb uute kudede tekkimises, mille tagajärjeks
on juhtkimpude mahu suurenemine ja seeläbi varre jämenemine, mitte
aga
pikenemine . Teiskasv on iseloomulik paljasseemnetaimedele ja
puitunud, harvem rohtsetele kaheidulehelistele. Viimastel esineb seda
harva ja piiratud ulatuses. Üheidulehelistel taimedel teiskasv
enamasti puudub või olemasolul erineb teiste soontaimede omast.
Üheiduleheliste teiskasvu siin ei käsitleta.
Kõik
koed, mida
kambium
toodab väljapoole, moodustavad
teiskoore,
sissepoole tekib aga
teispuit.
Vegetatsiooniperioodil esineb kambium enamasti mitme rakukihi paksuse
kambiaalse tsoonina,
puhkeolekus on kambium harilikult ühekihiline.
Parasvöötmealade taimede teispuidus võib eristada
aastaringe
(iga-aastast juurdekasvu). Kevadel toodab kambium enamasti suure
läbimõõduga trahheesid ja trahheiide, suvel ja sügisel aga
väikese diameetriga
kitsaid trahheiide ning rohkem puidukiude ja
põhikudet.
Teisksüleemi
moodustumisel lõpetab esiksüleem
funktsioneerimise , protoksüleem
hävib, kuid protoksüleemis leidunud säsirakud võivad
funktsioneerida veel aastaid. Varre keskosas asuv säsi võib olla
sekundaarsete kudede poolt kokku surutud.
Kõige
selgemini on aastarõngaste piirjooned nähtavad okaspuudel.
Lehtpuudest eristuvad aastarõngad hästi
rõngassoonelistel
liikidel. Aastarõngaste kevad- ja sügisosa erineb siin mõnevõrra
värvuse,
suuremal määral aga tiheduse poolest. Kevadosa koosneb
suure läbimõõduga trahheedest, mis ristlõigul on vaadeldavad
augukeste selge reana. Sügiseses osas on ülekaalus puidukiud ja
väiksema läbimõõduga trahheiidid.
Varjatud
rõngassoonelistel
liikidel paiknevad kevadosas rohkearvulised suured trahheiidid,
sügiseses osas on need väiksema läbimõõduga ning trahheiidide
vahele jääb rohkesti puidukiude.
Hajussoonelistel
lehtpuudel on aastarõngaid raske eristada, kuna sügis- ja
kevadpuit erineb üksteisest vähe.
Puidu
määramisel on olulisteks tunnusteks säsikiirte laius, värvus ja
hulk. Esineb üherealisi (okaspuudel, haaval), mitmerealisi (vahtral)
ning laiu (tammel) säsikiiri. Mitmerealised või
laiad säsikiired
on nähtavad puidus palja silmagagi,
kitsad säsikiired aga ainult
mikroskoobi abil.
Vanemate
puutüvede ksüleemis saab reeglina eristada kaht osa -- sisemist,
tihedamat, kõvemat ja värvuselt tumedamat
lülipuitu
ning välimist, heledamat
maltspuitu.
Lülipuit ainete
transpordis enam ei osale, sest siin on trahheed ja
trahheiidid ummistunud
tüllidega
-- ksüleemi põhikoe rakkude tsütoplasma väljasopistustega.
Lülipuidus on ka ksüleemi põhikoe rakud surnud. Lülipuidu rakkude
kestad on mõnedel liikidel läbi imbunud parkainetega, mis muudab
puidu tumedamaks, kõvemaks ja
lagunemisele vastupidavamaks.
Paljudel
kaheidulehelistel muutuvad esifloeemi rakud talitluse lõppemisel
niinekiududeks. Esikoor ja epiderm võivad eksisteerida veel aastaid,
juhul kui nende rakud jagunevad vajalikul määral.
Teiskoor
sisaldab samu rakutüüpe, mis primaarne niineosagi, kuid niinekiud
ja sõeltorud asetsevad siin rühmiti. Kambium toodab
vaheldumisi niinekiude ja sõeltorusid ning sõltub taimeliigist, mitu korda see
suve jooksul vaheldub. Niinekimpude rühmi moodustub rohkem kui
puidus aastarõngaid. Aastarõngad pole teiskoores nii selgelt
eristunud kui teispuidus. Okaspuude floeem sisaldab ainult
sõeltorusid ja floeemi põhikudet, saaterakke asendavad siin
erilised albumiini sisaldavad rakud. Floeemi ulatuv säsikiir koosneb
ainult
elusatest rakkudest.
Juhtkimpe
üksteisest
eraldavad säsikiired on teispuidus juhtkimpude poolt
kokku surutud ning tunduvalt kitsamad kui üheaastases oksas.
Juhtkimpudes võime näha ka
teissäsikiiri,
mis ei
alga varre keskosast, vaid mõne aastarõnga lähedusest.
Mitmeaastased
varred on harilikult kaetud ebaühtlase paksusega
felleemiga
(korkkoega), mis koosneb lapikutest, tihedalt ja kohakuti
asetsevatest korgistunud surnud rakkudest. Felleemist väljaspool
võib leiduda ka epidermi jäänuseid. Felleemi all asub õhuke
(enamasti
kahekihiline ) tsütoplasmarikas kude --
fellogeen
ehk korgikambium, mis väljaspoole toodab felleemi, sisse aga
esikoore põhikoele sarnanevaid rakke --
fellodermi.
Kõiki
fellogeenist tekkinud
kudesid nimetatakse
peridermiks.
Fellogeen lõpetab mõne aja pärast tegevuse, sügavamal tekib aga
uus fellogeeni kiht, mis toodab samuti felleemi ja fellodermi.
Felleem ei lase läbi vett ega seal lahustunud toitaineid, seetõttu
surevad kõik korkkoest väljapoole jäävad rakud. Nii tekib
enamikul puittaimedel sekundaarne
korkkude --
korp,
mis koosneb peridermi, koore- ja niinerakkudest. Sklerenhüümrõngas
rebeneb varre jämedamaks kasvades ribadeks ning korba tekkimisel
kaob hoopis.
Koos
epidermiga kaovad ka õhulõhed. Et
gaasivahetus saaks jätkuda,
tekivad korkkoesse lõved. Esialgu moodustuvad need endiste õhulõhede
kohal. Siinsed fellodermi rakud muutuvad
algkoe --
lõve
fellogeeni
-- rakkudeks. Lõve fellogeen toodab väljapoole kobedalt asetsevaid
põhikoerakke --
täiterakke,
sissepoole fellodermi. Lõve täiterakkude kest on õhem kui felleemi
rakkudel. Vanad täiterakud tõugatakse väljapoole,
meristeem kasvatab aga seestpoolt üha uusi täiterakke. Korba tekkimisel
hävivad ka senised lõved. Uued lõved tekivad sügavamal asuvatest
peridermi kihtidest. Lõved jagatakse ehitusest sõltuvalt kolme
tüüpi:
1)
täiterakud korgistuvad kohe; lihtsaim ehitustüüp, esineb näiteks
perekondades
pappel (
Populus)
ja
pirnipuu (
Pyrus);
2)
täiterakud asenduvad korgistunud rakkudega alles
vegetatsiooniperioodi lõpul, näiteks perekondades saar (
Fraxinus),
tamm (
Quercus),
pärn (
Tilia )
ja leeder (
Sambucus);
3)
kohevad korgistumata ja tihedad korgistunud rakkude kihid vahelduvad,
näiteks perekondades
kask (
Betula),
pöök (
Fagus),
ploomipuu (
Prunus )
ja robiinia (
Robinia).
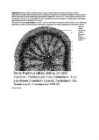
Kaheiduleheliste
taimede teiskasvu põhitüübid on järgmised (joonis):
Tilia-tüüp,
kambium asetseb algusest saadik pideva rõngana;
Aristolochia-tüüp,
algul esineb vaid kimbukambium, teiskasvu ilmudes tekib ka
inter-fastsikulaarkambium, mille talitlemisel moodustub küll pidev
kambiumirõngas, ent juhtkimbud jäävad eraldi;
Helianthus-tüüp,
sarnaneb eelmisele, kuid kimbuvahekambium tekitab uusi juhtkimpe;
Ricinus-tüüp,
esikasv sarnaneb
Aristolochia-tüübile,
kuid kimbuvahekambiumi tegevuse tulemusena muutub juhtkimpude
eristamine võimatuks.
4. JUUR 4
.1.
Juure esikasvJuure
tipus asuv
kasvukuhik
(joonis) sarnaneb ehituselt ja talitluselt varre kasvukuhikuga.
Erinevuseks on juure initsiaalrakkude omadus toota meristemaatilisi
rakke ka väljapoole. Need algkoe rakud moodustavad
kalüptrogeeni,
mille talitlemisel kujuneb õrnu initsiaalrakke vigastuste eest
kaitsev
juurekübar
ehk kalüptra.
Juurekübara
rakkudes leidub suuri amüloplaste --
statoliite,
mis
toimivad tasakaalukehadena. Statoliitide tõttu kasvavad juured
vertikaalselt allapoole (positiivne geotropism). Taim ei kasuta
statoliittärklist ka kõige suurema toitainete puuduse korral.
Initsiaalrakkudele
vahetult järgnevat juure osa nimetatakse
jagunemisvöötmeks.
Intensiivse ja kiire jagunemise tõttu jäävad siinsed rakud
väikesteks.
Järgneb
pikenemis -
ehk
kasvuvööde.
Selle vöötme rakud poolduvad harva, küll aga toimub nende
pikenemine. Tsütoplasmas tekivad esimesed
vakuoolid .
Edasi
tuleb juure
imamis-
ehk
diferentseerumisvööde,
kus kujunevad püsikoed ja tekib hulgaliselt juurekarvu.
Diferentseerumisvöötme
ja
juurekaela
(juure varreks ülemineku koht) vahelises osas --
juhtimisvöötmes
--
kujunevad külgjuured.
Juurekarvad on selles juureosas surnud,
mistõttu vööde ise vett ja mineraalsooli ei ima, vaid ainult juhib
neid edasi.
Primaarse
ehitusega juur on kaetud
epibleemiga
(risodermiga), mis
talitleb imikoena.Juurekarvadega epibleemi rakke
nimetatakse
trihhoblastideks.
Enamasti vahelduvad trihhoblastid epibleemi rakkudega, millel pole
juurekarvu.
Epibleemi
all asub isodiameetrilistest rakkudest
esikoor.
Esikoore väline kiht --
eksoderm
-- koosneb tihedalt üksteise vastu liibunud nurgelistest rakkudest,
mis vanemas eas sageli korgistuvad. Mõnikord võib siin näha ka
rakukestade erilisi paksendeid --
Caspary
jooni.
Arvatakse, et need aitavad reguleerida ainete transporti. Enamusel
soontaimedest on juure eksoderm hästi arenenud.
Eksodermile
järgneb
esikoore
põhikude.
Esikoore ja epibleemi põhiülesandeks on vee ja selles lahustunud
mineraalainete imamine ning transport steeli suunas. Vee- ja
sootaimedel ning teistel õhuvaesesse pinnasesse kinnituvatel
liikidel tekivad esikoores skiso- või lüsigeensed õhukambrid ning
moodustub
aerenhüüm.
Esikoore
sisemine kiht,
endoderm
(joonis), koosneb nii puitunud ja korgistunud paksukestalistest
rakkudest kui ka õhukesekestalistest elusatest
läbilaskerakkudest.
Viimased võimaldavad mineraalainetel ja veel kesksilindrisse
liikuda . Endodermi rakkude paksenenud kestadel võib leida
Caspary
jooni.
Kesksilindri
välimine kiht --
peritsükkel
(perikambium)
-- koosneb enamasti ühest, harvem mitmest reast elusatest,
meristemaatilistest rakkudest. Juure peritsükli üheks funktsiooniks
on külgjuurte moodustamine. Vaid üksikutel vee- ja parasiittaimedel
pole peritsüklit leitud.
Peritsüklist
seespool asub radiaalne
juhtkimp ,
kus ksüleem ja floeem on asetunud vaheldumisi, ribadena. Sellist ühe
radiaalse juhtkimbuga steeli nimetatakse
aktinosteeliks
(ka kiirjaks ehk radiaalseks steeliks). Sõltuvalt ksüleemi ja
vastavalt ka floeemi kimpude arvust võib aktinosteel (joonis) olla
kas
monarhne
(1 kimp), esineb mõnedel sõnajalgadel,
diarhne
(2 kimpu), sõnajalg- ja paljasseemnetaimedel, korvõielistel
(
Asteraceae),
ristõielistel (
Brassicaceae),
sarikalistel (
Apiaceae)
jt. õistaimede sugukondadel,
triarhne
(3
kiirt ),
tetrarhne
(4 kiirt),
pentarhne
(5 kiirt),
heksarhne
(6 kiirt) või
polüarhne
(ksüleemikimpe üle kuue); üheidulehelistel taimedel. Tetrarhne,
pentarhne ja heksarhne steel on iseloomulik enamikule
kaheidulehelistest.
Ksüleemis
on kerge eristada väikese läbimõõduga trahheiididest
protoksüleemi
(ksüleemikiirte tippudes) ja suurtest astrik-, võrk- ja
poortrahheedest koosnevat
metaksüleemi.
Floeemikimbud asuvad vaheldumisi ksüleemikiirtega.
4
.2.
Juure teiskasvSekundaarse
ehitusega juures on kambiumi tegevuse tulemusena moodustunud
teisksüleem ja -floeem. Kaheidulehelistel taimedel tekib kambium
floeemi ja ksüleemi vahele. Niinekimpudest seespool asuv kambium
funktsioneerib alguses aktiivsemalt ning seeläbi muutub laineline
kambiumikiht peagi ümaraks ja juure teiskasv toimub edaspidi
ühtlaselt (joonis).
Peritsüklist
kujuneb fellogeen, mis väljapoole moodustab felleemi.
Peridermi
tekkimise tõttu katkeb ainevahetus esikoore ja kesksilindri vahel;
esikoor
sureb ning
irdub . Teiskoore moodustavad koed, mida kambium
toodab väljapoole: floeem, teiskoore põhikude koos peridermiga
(joonis).
Juure
teisksüleem erineb varre teispuidust järgmiste iseärasuste
poolest:
1) säsikiired
on suuremad, need ei kulge
rangelt radiaalselt;
2) ksüleemi
põhikudet on rohkem, puidukiude vähem;
3) trahheesid
ja trahheiide on
arvukamalt ja need on suurema läbimõõduga ning
laiemate pooridega, tülle tekib haruharva;
4)
malts - ja
lülipuit ei erine teineteisest nii selgesti. Teisfloeemis on
niinekiude vähem ja sõeltorude läbimõõt on suurem.
Üheidulehelistel
taimedel teiskasv puudub, nende juured võivad aga puituda ja sel
teel väga tugevateks muutuda. Peridermi küll ei esine, kuid
endodermi ja esikoore põhikoe sisemised rakud korgistuvad ning
puituvad, moodustades
niimoodi juurt kaitsva katte.
4.3.
Juure muudendid Juure
muudendite
suurest hulgast
vaatleme praktikumides säilitusjuuri,
juuremügaratega juuri, mükoriisat, haustoreid ja õhujuuri. Lisaks
neile on
troopiliste alade taimedele väga olulised mitmesugused
tugijuured
mangroovisoodes ja
ronijuured.
Parasvöötmealade liikidel on laialdaselt levinud
tõmbejuured
(kontraktiiljuured,
radices
contractiles),
mis aitavad maa-alusel osal teatavas sügavuses püsida. Eriti
oluline on see taimede talvitumisel.
Säilitusjuur
võib tekkida ja pakseneda neljal
erineval moel:
1)
kambiumirõngas
toodab sekundaarset ksüleemi rohkem kui sekundaarset floeemi.
Sekundaarse ksüleemi juhtkudede ja puidukiudude vahele moodustub
rohkesti õhukesekestalisi, varuaineid sisaldavaid põhikoerakke.
Esineb näiteks redisel (
Raphanus
sativus).
2)
kambiumirõngas
toodab sekundaarset floeemi rohkem kui sekundaarset ksüleemi.
Sekundaarne floeem koosneb valdavalt põhikoerakkudest; esineb
porgandil (
Daucus
sativus).
3)
juures
tekib hulgaliselt sekundaarseid kambiumirõngaid (polükambiaalsus).
Varajane teiskasv sarnaneb tavalisele juure paksenemisele. Esimene,
algselt ühekihiline lisakambiumi rõngas tekib peritsükli ja
floeemi põhikoe rakkudest. Peagi jagunevad rakud tangentsiaalselt,
moodustub kahekihiline meristeemkude. Sisekiht hakkab floeemi ja
ksüleemi
tootma , välimine kiht muutub mõne aja pärast uuesti
kahekihiliseks, tekitades teise lisakambiumi rõnga. See jaguneb
omakorda kahekihiliseks: selle välimised rakud diferentseeruvad
kolmandaks lisakambiumi rõngaks,
sisemisest rakukihist tekib aga
teine juhtkimpude ring ja põhikude. Kolmas lisakambiumi rõngas
võtab osa
neljanda kambiumirõnga ja kolmanda juhtkimpude ringi
moodustamisest jne. Polükambiaalse tekkega on
peedi (
Beta vulgaris )
säilitusjuur.
4)
juur
pakseneb külg- või lisajuurte tüsenemise tõttu,
tekib
muguljuur.
Muguljuureks võib muutuda juur tervikuna või mingi osa sellest,
juur võib pakseneda ka katkendlikult. Esineb käpaliste
(
Orchidaceae)
sugukonnas, kanakoolmel (
Ranunculus
ficaria),
daaliatel (
Dahlia
sp.) jt.
Juuremügarad
esinevad kaunviljaliste ehk leguminooside seltsi (
Fabales)
kõikides sugukondades: liblikõielised (
Fabaceae,
peaaegu kõikidel liikidel), mimoosilised (
Mimosaceae,
enamikul liikidest) ja tsesalpiinialised (
Caesalpiniaceae,
vähem kui pooltel liikidel). Lisaks kaunviljalistele leidub
juuremügaraid veel kaheksa
sugukonna 140 liigil; näiteks porsal
(
Myrica gale ),
mõnel murakal (
Rubus
sp.), astelpajul (
Hippophaë
rhamnoides),
hõbepuul (
Elaeagnus
sp.), leppadel (
Alnus
sp.), drüüasel (
Dryas octopetala)
ja kasuariinil (
Casuarina
sp.).
Liblikõieliste
juuremügaraid põhjustavad mullas vabaltelavad õhulämmastikku
siduvad
bakterid enamasti perekondadest
Rhizobium
ja
Bradyrhizobium,
ülejäänud taimeliikidel kiirikseened (
Actinomycetes,
perekond
Frankia).
Põhjalikumalt
on uuritud liblikõieliste juuremügaraid. Bakterid, tunginud
juurekarvade kaudu koore põhikoesse, poolduvad jõudsalt ning
tungivad üha uutesse rakkudesse. Nakatunud
taimerakud suurenevad,
nende kromosoomide arv kahekordistub ning nii tekib
bakteroidkude.
Taim lagundab pidevalt bakterirakke ja omastab bakterite fikseeritud
lämmastikühendeid, bakterid aga kasutavad taimseid
sahhariide .
Mükoriisa
(
seenjuur )
on osa sugukondade, nagu kanarbikulised (
Ericaceae)
või käpalised (
Orchidaceae),
liikide jaoks obligatoorne, teistel võib esineda fakultatiivsena.
Mõne sugukonna taimed ei moodusta mükoriisat, seda ei esine ka vee-
ja üheaastastel taimedel. Hüüfide paigutuse järgi võib
mükoriisad jagada üldjoontes ka-
heks (mükoriisauurijad kasutavad
enamasti märksa detailsemat jaotust):
1)
vesikulaar-arbuskulaarne
(nimetatud ka endotroofseks mükoriisaks) -- levinuim tüüp, mis on
nime saanud juure esikoore põhikoe rakkudes asuvate põisjate
seenehüüfide järgi. Sellise mükoriisaga juuri pole väliselt
võimalik nakatumata juurtest eristada. Taimerakud võivad hüüfe
seedida, seda nähtust nimetatakse
mükofaagiaks.
Vesikulaar-arbuskulaarne
mükoriisa esineb obligatoorse mükoriisana peaaegu kõikides
katteseemnetaimede sugukondades, välja arvatud lõikheinalised
(
Cyperaceae),
ristõielised (
Brassicaceae)
ja maltsalised (
Chenopodiaceae).
Vesikulaar-arbuskulaarset mükoriisat moodustavad seened kuuluvad
sugukonda
Endogonaceae
(klass seigseened -
Zygomycetes);
Vesikulaar-arbuskulaarne
mükoriisa esineb peaaegu kõikidel käpalistel (
Orchidaceae),
kusjuures klorofüllita
heterotroofsed liigid -- pisikäpp (
Epipogium
aphyllum),
pruunikas
pesajuur (
Neottia
nidus-avis)
ja kõdu-koralljuur (
Corallorhiza
trifida)
-- saavad kogu vee ja
toitained seentelt.
2)
ektotroofne
-- hüüfid on juure koore põhikoe rakkude vahel (
Hartigi
võrk)
ning tiheda kattena juure välispinnal, moodustades juurt ümbritseva
mantli
(mükodermi). Ektotroofse mükoriisaga juured on tömbid, paksenenud,
kujult enamasti koraljad, keeljad või pintseljad; juurekarvad
puuduvad. Niisugune mükoriisa esineb 3% seemnetaimedest, enamasti
puudel. Mükoriisa moodustajateks on
kandseened (
Basidiomycetes),
lehikulaadsete (
Agaricales)
seltsi kuuluvad
puravikud (
Boletus),
pilvikud (
Russula),
riisikad (
Lactarius)
jt.
Ektotroofse
mükoriisa alatüübina võib eristada ekto-endotroofset mükoriisat,
kus osa hüüfe on tunginud rakkude sisse. See on ekto- ja
endomükoriisa vahepealne tüüp, esineb perekondades mänd (
Pinus )
ja
kuusk (
Picea )
ning
seltsis kanarbikulaadsed (
Ericales).
Parasiitide
imijuured ( haustorid ).
Pool-
ehk
hemiparasiitideks
nimetatakse taimi, kes on võimelised küll ise fotosünteesima, kuid
osa toitaineid võtavad need peremeestaimelt;
täielikud
parasiidid
(obligaat- ehk holoparasiidid) pole võimelised iseseisvalt
(auto-troofselt) toituma.
Parasiittaimede
juurtel arenevad
haustorid
ehk imitorud, millega need kinnituvad peremehe külge ning mille abil
omastavad temalt toitaineid. Nende keskosas on pikad
spiraalpaksenditega trahheiidid, mille abil vesi ja toitained
juhitakse
parasiidi ksüleemi. Tun-
ginud peremehe juhtkimpu,
limastuvad parasiidi hüüfitaoliste piklike rakkude
tipud , mille
tulemusena peremehe ja parasiidi tsütoplasmad puutuvad kokku.
Morfoloogiliselt on haustor lisajuur. Mõnel parasiidil (võrm --
Cuscuta)
areneb idanemisel kõigepealt lühike
peajuur , mis aga õige pea
sureb, teiste parasiittaimede (soomukas -
Orobanche)
seemned idanevad ainult pärast kokkupuudet
peremeestaime juurega.
Õhujuured
kitsamas mõttes on õhust niiskust koguvad juured, laiemas
käsitluses kuuluvad siia kõik õhkkeskkonnas paiknevad ja
talitlevad juured, sealhulgas assimileerivad ning
hingamisjuured .
Veeauru imavad juured esinevad troopilistel epifüütidel
sugukondadest käpalised (
Orchidaceae),
bromeelialised (
Bromeliaceae)
ja võhalised (
Araceae).
Lisaks õhujuurtele esineb epifüütidel ka tavalisi toitumis- ja
kinnitusjuuri. Kui õhujuur ulatub mulda, areneb selle tipust
tavaline juur.
Õhujuured
on kaetud
juurenahaga
(
velamen
radicum),
mis otsekui filterpaber endasse niiskust
imab . Juurenaha ülesandeks
peetakse ka juure kaitsmist mehaaniliste vigastuste eest ning
transpiratsiooni vähendamist juurest. Tekkelt vastab juurenahk
mullas arenevate juurte risodermile, kuid on mitmekihiline.





































Kõik kommentaarid