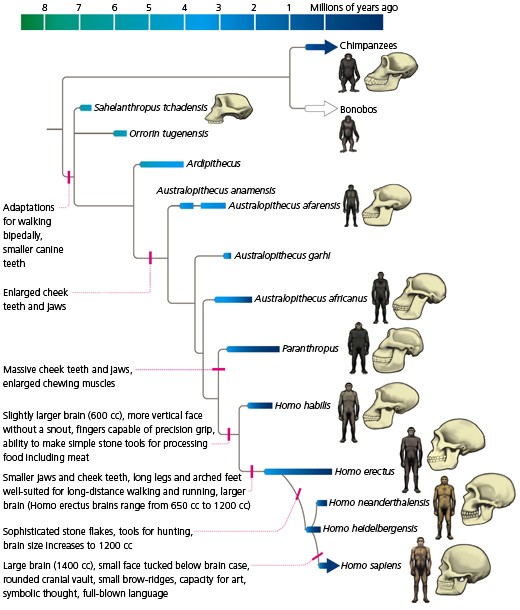
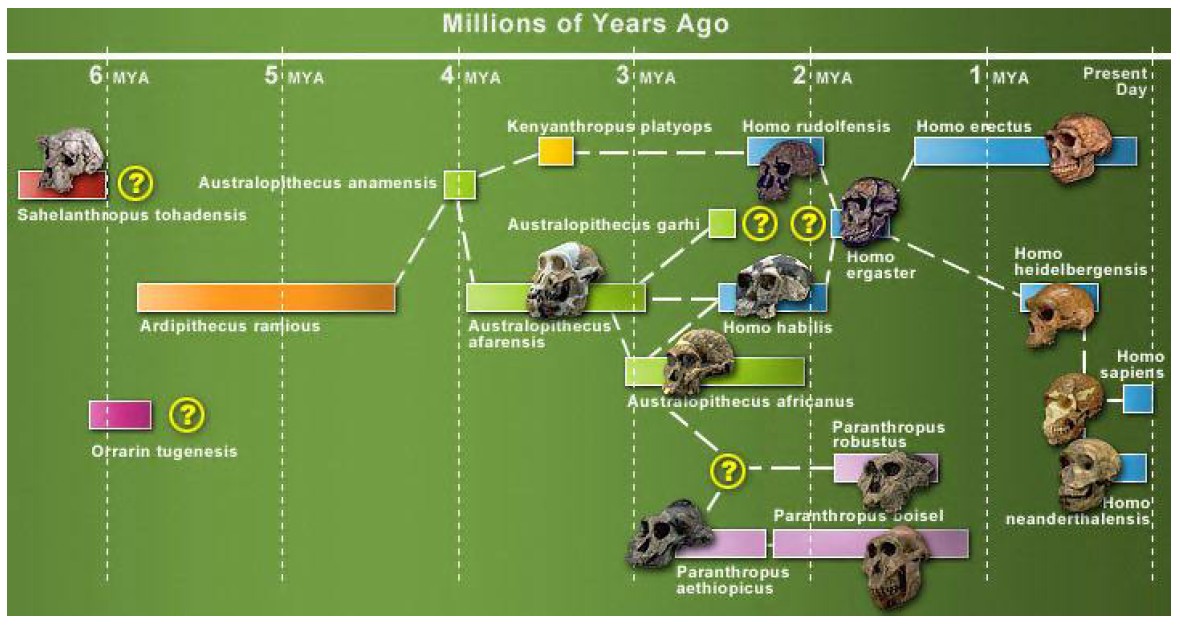
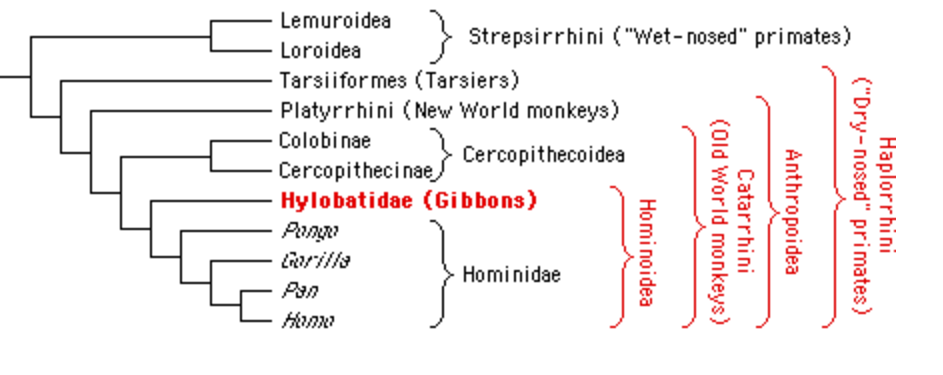
-ka simpans, vares oskavad riistu kasutada HOMO ERECTUS -hakkas liha sööma -suutis ületada ka merd, kus teine kallas näha ei olnud -siiski ei jõudnud Ameerikasse ega Austraaliasse -võttis kasutusele efektiivse kivikirve -võib-olla suri välja kõigest 30-50 KAT (Erectusest arenes ka selline loom nagu H. Floresiensis 80000-20000 a tagasi. 1m pikk, aju maht 300 cm3 EVOLUTSIOON ei pea alati olema progressiivne!!!) NEANDERTAALLANE -avastati 1856, Neandertal (Neanderi org), Saksamaa -tõenäoliselt ei olnud ristumisbarjääri inimesega -lahknemine inimese eellasest ca 600 KAT -suutis tõenäoliselt rääkida (anatoomia nagu lubaks) -tööriistad olid inimesega samad kuni ca 40-50 KAT -tegi ka kaelakeesid jms asju, aga augustas merekarpe oma tehnikaga erinev H. Sapiensist -pea täielikult lihasööjad
EVOLUTSIOONITEOORIA KUJUNEMINE Elu ajaloolist arengut liikide üksteisest põlvnemise ja muutumise kaudu nim elu evolutsiooniks e bioloogiliseks evolutsiooniks. Muutused on kindlasuunalised ja pöördumatud. Elu tekkis Maal u 3,7-4 miljardit a tagasi. Füüsikaline evolutsioon Keemiline evolutsioon Bioloogiline evolutsioon Sotsialne evolutsioon Arenemislugu: elu algus VEES! 1. nn ürgpuljongis isepaljunevad biomolekulid (geenide esivanemad) 2. biomolekulid koondusid pikemateks ahelateks (viirused ja praeguste kromosoomide esivanemad) 3. need koondusid bakteriteks 4. bakterite sümbioosi tulemusena eukarüootsed ainuraksed (u 2 miljardit a tagasi). 5. need ühinesid kolooniateks ja hulkrakseteks organismideks (vetikad 1 miljard a tagasi) 6
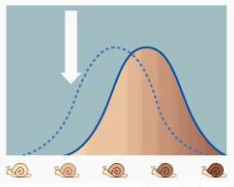
Sellega sai evolutsiooniteooria aluseks populatsioonigeneetika. Neodarvinismi kohaselt on Darwini poolt võimalikuks peetud pangeneetiline pärandumine (s.t., et ka elu jooksul omandatud tunnused võiksid päranduda järglastele) võimatu. Geneetiline tekib muutlikkus populatsioonides juhuslikult mutatsioonide tõttu ja rekombinatsiooni tõttu (see on homoloogiliste kromosoomide ristsiire meioosi ajal). Evolutsioon seisneb põhiliselt alleelisageduste muutumises põlvkondade lõikes geenitriivi, geenisiirde ja loodusliku valiku tagajärjel. Liigiteke leiab aset järk-järgult, kui populatsioonid geograafiliste barjääride tõttu reproduktiivselt isoleeruvad. Postulaadid: Populatsioonid sisaldavad geneetilisi variante, mis tekivad juhuslikult mutatsioonide ja rekombineerumise tulemusel (st. mitte adapteerumisest tingituna);
Statistilised meetodid, et hinnata LV mõju pidevatele tunnustele populatsioonis. Saltatsionistid eitasid LV-d ja arvasid, et muutused pidid toimuma hüppeliselt, kuna vahepealsed vormid ei olnud nende meelest kohased ja LV peaks neid siis elimineerima. Lamarkism oli valdav omandatud tunnused on päritavad, liigid muutuvad, kuid ei jagune ega kao. Primitiivseid liike tekib juurde. Sisemine täiustumistung, sunnib saama järglasi, kes on temast paremad. Darwinil oli kaks seotud teooriat evolutsioon (liikide muutumine ja jagunemine, paljude liikide väljasuremisega) ja LV (kui evolutsiooni mehhanism). Evolutsioon on hargnemine vastavalt kohanemine uute keskkonnatingimustega. Puudub vajadus hierarhiale, lihtsalt keerukale. 9. Miks ei leidnud Darwini idee LV-st tema kaasajal toetajaid? Usuti, et liigid muutuvad, aga ei usutud, et need toimuvad sammhaaval. Kuna erinevused eri kehaosade vahel on liiga erinevad, vahepealsed vormid ei saa olla kohased. Kuna pärilikkust ei
interpreteerida võimena muutuda. Kuid on ilmne, et ideed muutuvast loodusest hakkasid elujõudu koguma alles 18 sajandi teisel poolel. Dogmaatiline kirik ja tähttähelt võetav Vana Testament on valdavalt võõras ka tänapäeva (katoliku) kirikule. Paavst astus siin hiljaaegu paar otsustavat sammu. Galileo mõisteti õigeks ja seejärel sõnastati ka k. kiriku uus seisukoht evolutsiooni suhtes: bioloogiline evolutsioon, k.a. inimese kui ühe imetajaliigi teke on bioloogia probleem, mida uuritakse teaduse vahenditega. Hinge teke on kiriku probleem. See on äärmiselt tervitatav interpretatsioon ja tõeline revolutsioon evolutsiooni käsitlusel. Paraku ei ole kõik kirikud nii targad kui Püha Peetruse tooli poolt juhitav. Agressiivne krieitsionism on küll rohkem ameeriklaste probleem - Euroopa on sellest vähem puudutatud, kuid Ameerika (USA) mõjujõudu (rikkust) silmas pidades pole põhjust neid tendentse
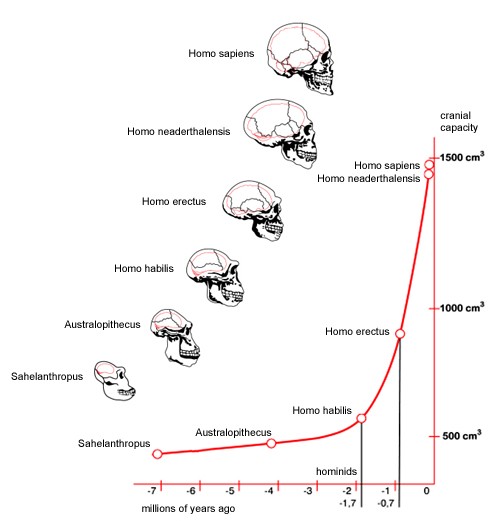
1. evolutsiooni kulg ja globaalsed katastroofid, nende mõju- Dateerimine süsinikumeetod, mis vanade fossiilidega ei tööta (kasut. teisi isotoope ja nt magnet) 3,5 mlrd a.t. üsna kindlad mikrofossiilid, hiliseim elu algus (areng protobiontidest). Tsüanobakterid (2,5) asusid fotosünteesima, hapnik kulus algul raua oksüdeerumiseks. 1,8 mlrd a.t eukarüoodid, sest genoom oli suurenenud ja rõngaskromosoom ei olnud otstarbekas. Endosümbioos. 1,2 mlrd a.t mingisuguse hulkraksuse teke. Kambriumi alguses toimus ,,Kambriumi plahvatus", milles kujunesid välja peaaegu kõik tänapäevased hulkraksete rühmad. Asustati maismaa. Devoni ajastul ilmusid kahepaiksed, neile järgnesid roomajad ja putukad. Keskaegkonnas said valitsevaks rühmaks just roomajad, kes suutsid saada domineerivaks kõikides nissides, mistõttu imetajate areng mõneks ajaks aeglustus ja nad jäid tahaplaanile. Ontogenees peege
PRODUTSENDID Ökoloogilise püramiidi reegel: Iga järgmise troofilise taseme biomass on ~10% eelneva taseme biomassist. Biomass ja eneriga vähenevad kõrgemate troofiliste tasemete suunas. EVOLUTSIOONITEOORIA KUJUNEMINE Elu ajaloolist arengut liikide üksteisest põlvnemise ja muutumise kaudu nim elu evolutsiooniks e bioloogiliseks evolutsiooniks. Muutused on kindlasuunalised ja pöördumatud. Elu tekkis Maal u 3,7-4 miljardit a tagasi. Füüsikaline evolutsioon Keemiline evolutsioon Bioloogiline evolutsioon Sotsialne evolutsioon Arenemislugu: elu algus VEES! 1. nn ürgpuljongis isepaljunevad biomolekulid (geenide esivanemad) 2. biomolekulid koondusid pikemateks ahelateks (viirused ja praeguste kromosoomide esivanemad) 3. need koondusid bakteriteks 4. bakterite sümbioosi tulemusena eukarüootsed ainuraksed (u 2 miljardit a tagasi). 5. need ühinesid kolooniateks ja hulkrakseteks organismideks (vetikad 1 miljard a tagasi) 6
KÜSIMUSI 1. J-B Lamarcki ja C. Darwini seisukohtade kokkulangevused ja erinevused Lamarck arvab, et elu omandatud tunnused päranduvad edasi aga Darwin arvas teistpidi. Lamarck ei arvestanud looduslikku valikut. Lamarck arvas, et liigid on püsivad, aga Darwin ütles, et liigid tekivad üksteisest ja ka kaovad(looduslik valik). Lamarck väitis,et liigid evolutsoneeruvad eesmärgipäraselt-pm evolutsioon on eesmärgiks, tglt toimub evolutsioon kuna keskkond muutub ja olen peab kohaneme, et ellu jääda/püsida konkurentsivõimelisena ja sellest tulenevad muutused põhjustavad evolutsiooni. 2. looduslik valik Loodusliku valiku teooria esitas esimesena C. Darwin. Tema raamat ,, The origin of species" ilmus 1859. aastal ja müüdi välja esimese päevaga. Raamatu sisu täienes tänu Wallace'ile ja ilmus kiiresti tänu Darwini tuntusele tollases ühiskonnas.




























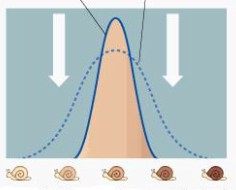
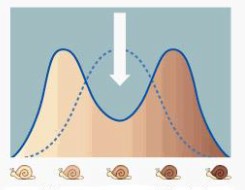
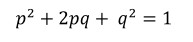




Kõik kommentaarid