Kullman, B. (2002). Horisontaalsest geeniülekandest seeneriigis. In: I. Puura & T.
Teder (toim), Võrkude teooria.
Schola Biotheoretica 28: 21-33. Tartu, Sulemees. 88 lk. (
ISBN 9989-9278-6-9)
Horisontaalsest geeniülekandest seeneriigis
Bellis Kullman
Intratsellulaarse sümbiogeneesi käigus on enamus protomitokondri geene transformeerunud peremees –tuuma
horisontaalse geeniülekande (HGT) käigus. Võrreldes erinevate seente mitokondriaalseid genoome (mtGenoom),
on jälgitav kodeerivate järjestuste asendumine mittekodeerivatega kuni nende kadumiseni. Pärmirakus ei muutu
mtGenoom ja tuumagenoom üksteisest sõltumatult. Võimalik, et HGT mitokondri ja tuuma vahel on siiani
käimasolev protsess. Seenerakus esineb palju viirusi ja plasmiide, mis võivad osaleda geenide ülekandel erinevate organismide vahel. Plasmiid võib olla mtGenoomi koosseisus või iseseisvalt paljuneda ja omada kindlaid
funktsioone (raku kasvu pidurdamine , vananemine, plasmiidis kodeeritud mürkide abil konkurentide
surmamine). Neid geene antakse edasi tsütoplasmaga seenehüüfide kokkukasvamisel (heteroplasmas). On
kirjeldatud HGT bakterilt seenele, seenelt seenele (eri liikide või sama liigi eri isendite mtGenoomide vahel,
seeneperekondade või ühe perekonna liikide tuumagenoomide vahel) ja seenelt taimele (sümbiontse või
parasiitsete seene mtGenoomist taime mtGenoomi). HGT on leitud rDNA I grupi intronite osas ja mitmete valke
kodeerivate järjestuste osas. Nii pärmidel kui hüüfe moodustavatel seentel on leitud mitootiline geenisiire, mis on sisuliselt sama, mis bakterite konjugatsioonil toimuv HGT. Kui haploidne tuum siseneb teise hüüfi, võib ta liikuda mitootiliselt
jagunedes, säilitades oma intaktsuse. Selliselt tekkivad dikarüootsete hüüfide tuumad võivad olla küllaltki
erinevad ( aneuploidsus , kromosoomi pikkuse polümorfism ). Tuumade liitumisel (konjugatsioonil? või/ja
kopulatsioonil) võib toimuda geenisiire sarnaselt bakteritega (HGT) või ka nii nagu tõelises meioosis
(genealoogiline, vertikaalne geenisiire - VGT). Seente tuumade jagunemisviiside ebastabiilsus ja mitmekesisus
võib olla põhjuseks, miks nii mõnelgi juhul pärilikkuse ‘võrku’ on ‘puust’ üpriski raske eristada. Kas ka
mitokondril on olnud roll homoloogiliste kromosoomide evolutsioonis , jääb mõistatuseks. Viimasel ajal on kogunenud üha enam andmeid horisontaalse geeniülekande (HGT –
horizontal gene transfer )
kohta seeneriigis. Nii leivapärmseene kui ka teiste seente
mitokondriaalne genoom (mtGenoom) on tugevalt
rekombinantne. Geneetilisi muutusi ja rekombinatsioone on kindlaks tehtud nii kunstlikul ristamisel (
Fincham ,
1979), kui ka looduslikes populatsioonides (
Saville , 1998). On isegi leitud, et klonaalselt
paljunevatel seentel on
raku tuumagenoomi muutumisele kaasnenud mtGenoomi rekombinantne (retikulaarne) muutumine (
Anderson ,
2001). Teadaolevalt paljunevad mitokondrid inimesel jt. imetajatel klonaalselt ning nende järgi on võimalik
määrata päritolu ‘mitokondriaalsest
Evast ’. On esitatud küll ka vastupidiseid seisukohi (Awadalla, 1999), mis
aga on osutunud ekslikeks andmete tõlgenduseks (
Kivisild & Villems, 2000).
Arvatakse, et HGT ja
endosümbioos on mänginud rolli nii valkude kui DNA mosaiiksuse tekkel (
Nara et al., 2000). Tõenäoliselt on HGT toimunud bakterite eellaste ja eukarüootide eellaste vahel väga varasest
organismide arengust alates mõlemas suunas ning võib toimuda mõnedes organismides tänapäevalgi. On
andmeid, et on toimunud HGT
bakterite eellastelt seente eellastele (Brinkmann
et al., 2001). Varakult pärast
Archaea ja eukarüootide divergeerumist võis aga toimuda vastupidine paljude geenide ülekanne
eukarüootidelt
bakteritele (Brown
et al., 1999). Hilisemast ajast on teada HGT
bakteritelt seentele (
eukarüoodid ). Näiteks
pärineb seene
Orpinomyces joyonii tselluloosi lagundamisel tähtsat rolli omav geen (celA) bakterilt
Fibrobacter succinogenes. See võimaldab seenel elada uues toitekeskkonnas, herbivoorsete imetajate vatsas
(
Garcia -Vallve
et al., 2000).
HGT prokarüootidelt eukarüootidele avastati 1990 aastal, kui võrreldi isopenitsilliin-N süntetaasi
(IPNS) geenide ja 5S rRNA geenide vastavaid evolutsioonilisi distantse.
Analüüsid näitasid
veenvalt , et IPNS
geen oli kahel korral
horisontaalselt üle
kantud bakteritelt hüüfe moodustavatele kottseentele (Panalva
et al., 1990).
Kuigi on teda, et DNA transport läbi membraanide ja rakkude vahel toimub, ei teata selle toimumise
mehhanismi.
Prokarüootidel toimub HGT peamiselt bakteriaalse konjugatsiooni käigus. Bakteritel on leitud
paljunemise ajal konjugatsiooni protsessis toimiv integraalne DNAd siduv valk TrwB (
Escherichia coli).
Arvatakse, et sellel
valgul võib olla roll geeni ülekandmisel bakterilt taimele ning bakterilt seenele (Comis-
Ruth et al., 2001).
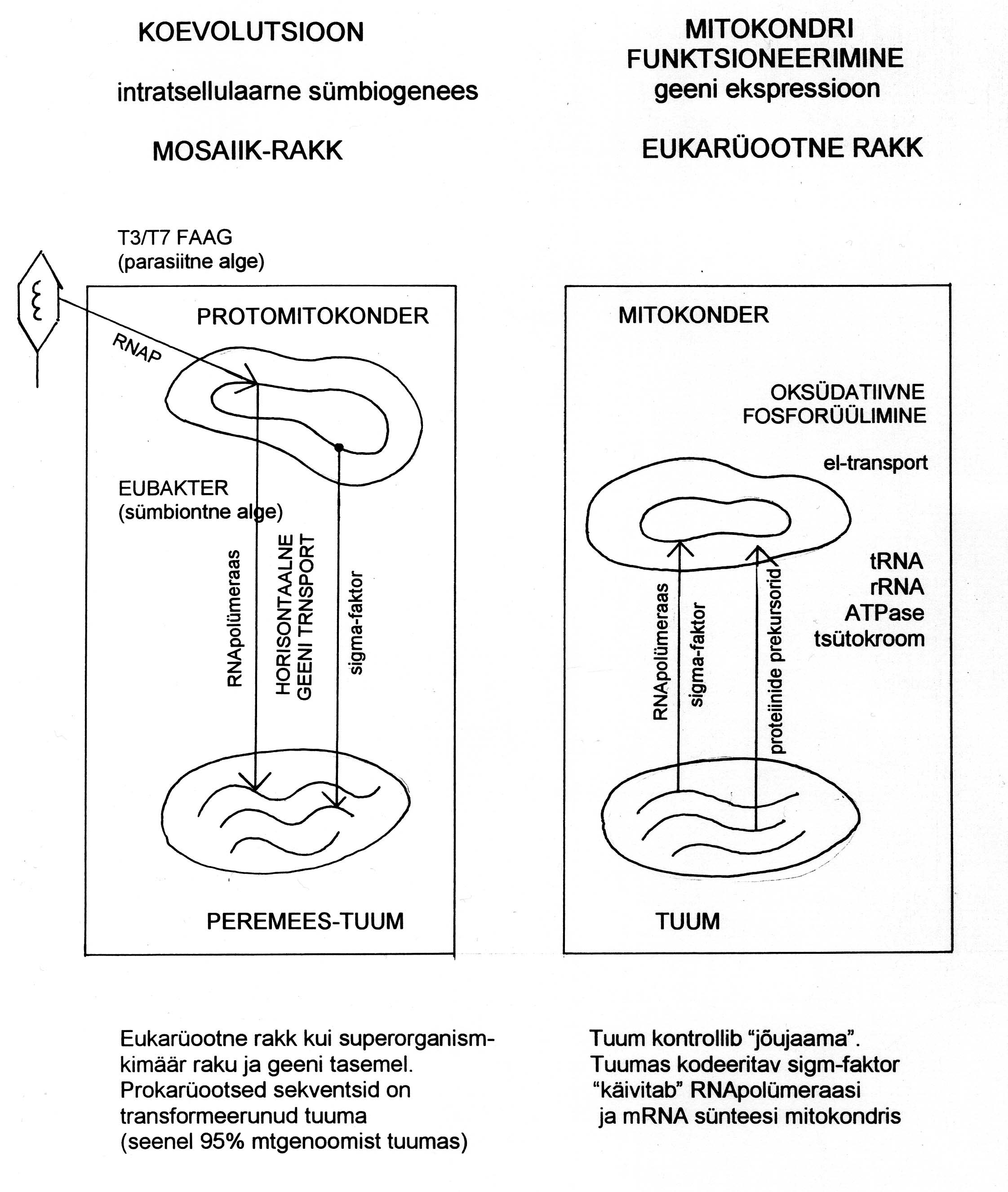
Joonis 1. Tuuma ja mitokondri koevolutsioon (vasakul). Eukarüootses rakus reguleerib tuum energia tootmist
mitokondris (paremal).
On andmeid ka
eükarüootide omavahelise HGT kohta. Nii on leitud HGT
seenelt taimele parasiitsete (Kim
et al., 2000) ja sümbiontsete seente puhul (Voughn
et al., 1995), kes elavad tihedas kontaktis
oma peremeestaimedega. Tegelikult on tegemist tsütoplasmaatilise pärilikkuse muutumisega. Nimelt on HGT
toimunud
seene mitokondrist taime mitokondrisse. Mitokonder on rõngasja kromosoomiga rakuorganell.
Mitokondri mtGenoom on haploidne, seetõttu kasutatakse tema kohta ka terminit haplotüüp. (Käesolevas töös
seda ei kasutata kuna hüüfe moodustavate seente tuumad on samuti valdavalt haploidsed). Mitokondril arvatakse
olevat eubakteriaalne päritolu (joonis 1). Kui oletada, et bakteriaalse päritoluga mitokondrid on säilitanud
bakteritele omaseid paljunemisviise, siis võib neil HGT aset leida konjugatsiooni käigus heteroplasmas
(piirkondades, kus
peremeestaime ja seene tsütoplasmad kokku puutuvad). Õistaimel
Peperoma polybotrya on
mitokondri coxI geenis
ekson ja
intron , milledel on selgelt erinev evolutsiooniline päritolu.
Eksoni DNA
järjestus on lähedane teiste õistaimede sama geeni vastavatele järjestusele, intronit aga teistel soontaimedel pole
leitud. Selle introni järjestus sarnaneb rohkem seente kui sammaltaime, helviku
Marchantia või rohevetika
Prototheca mitokondri vastavale geenijärjestusele. Arvatakse, et coxI introni doonoriks võis olla mingi
sümbiontne mükoriisaseen (Voughn,
et al., 1995).
On leitud HGT ka
seente endi vahel. Esmakordne otsene tõend HGT esinemisest mittesugulaslike
seente genoomide vahel saadi kahe kottsenne
Ascobolus immersus ja
Podospora anserina hüüfide
kontakteerumisel. Pärast seda leiti
esimese liigi lineaarne plasmiid teise liigi mtGenoomi koosseisust
(Kemken,
1995). Ka siin on tegu tsütoplasmaatilise pärilikkuse muutumisega. Plasmiidi insertsioon
mtDNA -sse võib
toimuda samal viisil kui bakteri DNA-sse (viimast kasutatakse
geenitehnoloogias ).
Mitmetes katsetes on saadud rekombinantseid mitokondreid (külmaseen, austerservik) sama seeneliigi
erinevate hüüfide liitumisekoha lähedusest. Ka külmaseene looduslikes populatsioonides esineb mtGenoomis
rekombinatsioone, nii et seente mitokondritele pole omane üksnes klonaalne paljunemine (Saville
et al., 1998).
Siiski, seentel pole HGT leitud üksnes erinevate liikide või sama liigi erinevate isendite mtDNA vahel,
vaid ka tuumageenide vahel. Tuumageenide HGT on leitud nii
seeneperekondade vahel kui ühe perekonna
liikide vahel. Liigi
Protomyces inouyei 18S rRNA geeni I grupi
intronid A ja B (mis sisaldavad tugevalt
konserveerunud nukleotiidse järjestusega elemente) asetsevad nii, et A on samas positsioonis nagu liigil
Pneumocystis carinii ja B nii nagu liigil
Ustilago maydis (Nishida,
et al.,1993). Uurides mitmete
taimeparasiitsete seente lähedasi perekondi (
Sclerotiniaceae) tuuma ribosomaalse DNA (SSU ja LSU rDNA)
järjestuste alusel leiti, et I grupi intronite osas on toimunud HGT (Holst-Jensen
et al., 1999). Samale järeldusel
jõuti parasiitse kottseene
Isaria japonica 18S rDNA I grupi intronite uurimisel (Ito
et al., 1999).
Jaapanis uuriti üherakuliste seente
Candida albicans ja
C. dubliniensis kliinilisi tüvesid ja leiti, et HGT
võib olla toimunud ühe perekonna liikide vahel. Kahel
C. albicans isolaadil 301 isolaadi seas tehti kindlaks
genotüüp , millises 25S rRNA geeni I grupi intronisse oli transponeerunud
C.
dubliniensis introni sarnane
insertsioon (Tamura
et al., 2001).
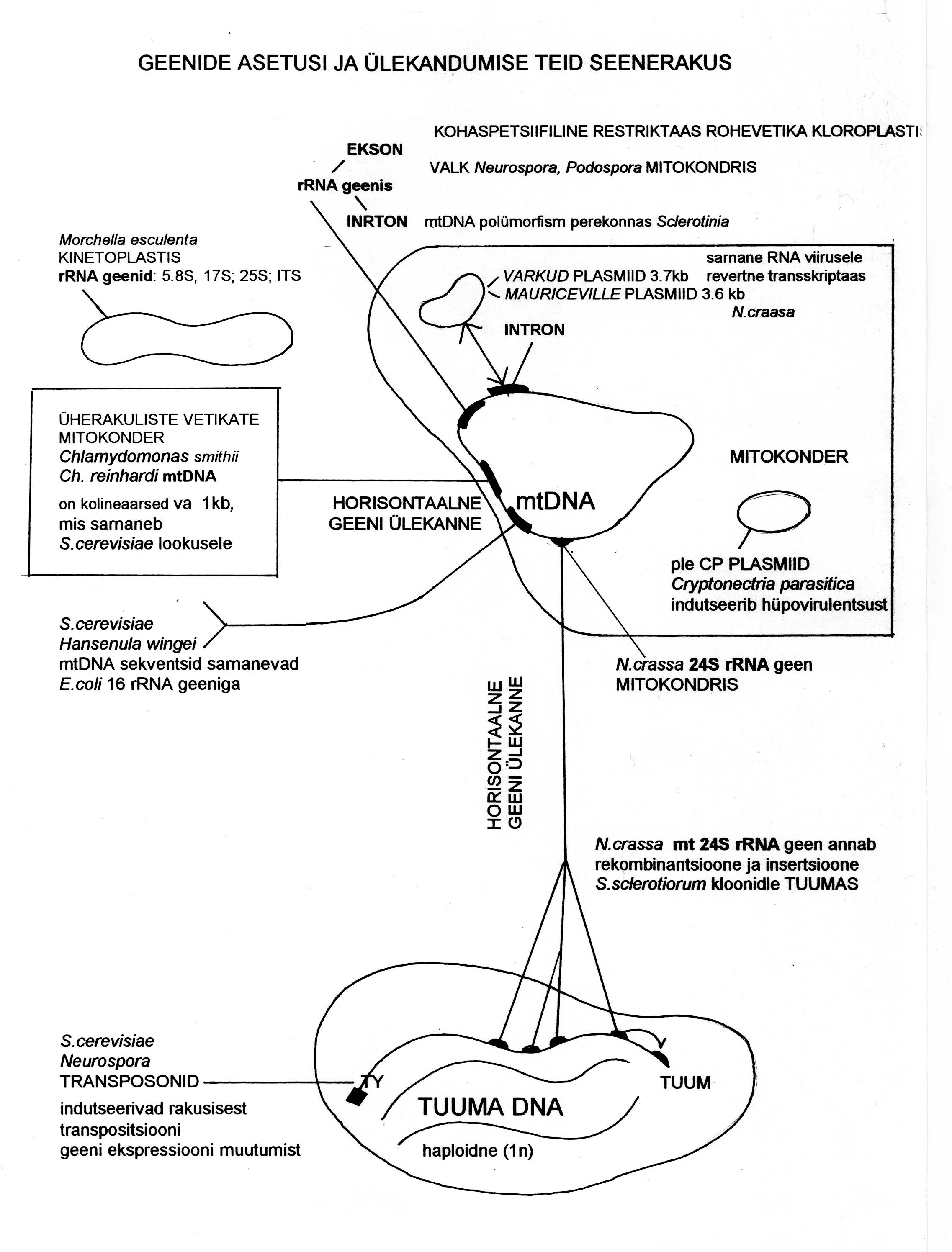
Transposoonid on geneetilised elemendid, mida on leitud nii pro- kui eukarüootides. Erinevalt teistest
geenidest on nad võimelised liikuma peremeesgenoomi sees uude asupaika. Transposoonide transpositsioon
kodeerivatesse DNA järjestustesse ja nende poolt algatatud kromosoomide ümberkorraldustel on suur mõju
geeniekspressioonile ja genoomi evolutsioonile. Kuigi hüüfe moodustavatel seentel avastati transposoonid alles
hiljuti, leitakse neid üha enam ja
seostatakse HGT-ga (Kempken & Kuck, 1998) (joonis nr 2 tähistatud TY).
Viimased uuringud on näidanud, et kuigi 20% kirjeldatud seentest on arvatud olevat mittesuguliselt ja
klonaalselt paljunevad, esinevad ka neil rekombinatsioonid (Taylor
et al., 1999). Selle üheks põhjuseks võiks
pidada meiootilistele ehk sugulistele rekombinatsioonidele alternatiivseid mitootilisi ehk somaatilisi
rekombinatsioone.
Paraseksuaaltsüklis toimuvat mitootilist geenisiiret on põhjalikult uuritud seoses penitsilliini
tootmisega. Pärast pintselhalliku
Penicillium chrysogenum hüüfide liitmist nende haploidsed tuumad liituvad
sagedusega 1 x 10–8 – 2 x 10-6. Nendes liitunud tuumades toimub mitootiline
ristsiire sagedusega 1 x 10-3 – 2 x
10-2 iga tuumajagunemise kohta (
Weber , 1993). Viimane arv võimaldab hinnata selle seene diploidina
jagunemise võimet.
Saades diploidseks, üks tuhandest kuni üks viiekümnest jaguneb rekombinantsena, oskamata
jaguneda nii, et tema
kromosoomid säilitaksid oma identsuse tütarkromatiidide lahknemisel. Ilmselt tuleneb see
metafaasiplaadi puudumisest seentel. Kromosoomid pole mitoosi ajal kondenseerunud (pole loetavad).
Tütartuumade laialitõmbamisel on tegev ainult seenele iseloomulik
tuumaga assotsieerunud
organell NAO
(
nucleous associated organelle), mis on tegev ka rakutuuma liikumisega interfaasis. Erinevalt kõrgemate
eukarüootide avatud mitoosist on seentele iseloomulik suletud
mitoos : jagunemine
tuumamembraani säilimisega.
Taime- ja loomarakus on mitoosi ajal kromosoomid tugevalt spiraliseerunud kromatiididega metafaasiplaadile
koondunud (nende arv on
loetav ). See tagab diploidsetel organismidel genoomi stabiilsuse tütarkromatiidide
lahknemisel. Neil säilib genoom tunduvalt stabiilsemana isegi diferentseerunud keharakkudes,
rääkimata sugurakkude tekkeks vajalikest tuumadest, mis on ‘konserveeritud’ suguorganites või meristeemkoes (suletud
paljunemissüsteem).
Seente diploidiseerumisel toimub midagi samaväärset kui bakterite konjugatsioonil. Bakterite
konjugatsioonil tekkivat rekombinantsust käsitletakse kui HGT tulemust. Aga kas seente puhul on õige
nimetada toimuvat HGT? Toimuv on seletatav kui algne variant meioosi esimesest jagunemisest, ajast, mil
polnud veel välja kujunenud täies pikkuses homoloogilisi kromosoome. Seene diploidses tuumas võib tõenäoselt
toimuda nii (1) kromatiidide vaheline ristsiire (mis toimub ka ilma kiasmideta) kui (2) kromosoomaberratsioonid
– seentele on iseloomulik kromosoomi pikkuse polümorfism (Zolan, 1995; Larraya, 1999) ja aneuploidsus
(Kullman, 2000).
Hüüfe moodustavatel seentel võib tänu seene hüüfi tipmisele kasvule võrrelda somaatilist geenivahetust
meioosiga (avatud paljunemissüsteem). Ega seene meioos spetsiaalsetes rakkudes (eoskotis, basiidis) sellisest
„somaatilises“ hüüfis toimuvast tuumade jagunemisest palju erinegi (sellest allpool). Haploidiseerumine
tuumade jagunemisel võib toimuda pintselhalliku hüüfides sagedusega 1 x 10-3 – 2 x 10-2 ühe tuumajagunemise
kohta (Weber, 1993). Sisuliselt näitab see meioosi teise jagunemise sagedust. Antud juhul langeb see kokku
mitootilise geenivahetuse sagedusega. Siin toimub meioosi esimesele jagunemisele tüüpiline geenivahetus
diploidse tuuma korduva (näivalt mitootilise) jagunemise käigus, enne kui jõutakse tuumade
haploidiseerumiseni. Meioosi teine jagunemine (haploidiseerumine) on vahepealsete jagunemistega ajaliselt ja
ruumiliselt eraldatud, edasi lükatud. Võib-olla annab see rohkem aega rekombinatsioonide tekkeks. Võiks öelda,
et suguline protsess on toimunud ilma nähtava sugulise paljunemiseta. Sellisel juhul on ilma viljakehade
moodustamiseta kasvava seene hüüfil justkui klonaalselt tekkivad
koniidid rekombinantsed venitatud ja varjatud
meioosi tõttu. Paraku ei allu rekombinatsioonide
lahknemine antud juhul populatsioonigeneetika
seadustele (rekombinantsete koniidide ebavõrdne arvukus? rekombinantide ebavõrdne lahknemine?).
Veelgi huvitavam on see, et
mitootiline geenivahetus võib toimuda ka klonaalselt paljunevas pärmirakus. Sugulise paljunemiseta diploidsed
pärmid on heterosügootsed (!). Mitootilised rekombinatsioonid
tekivad klonaalselt paljunevas pärmi
Candida albicans rakus kahe haploidse genoomi komponentide vahel
(Fincham
et al., 1979
). Sellel liigil haploidiseerumist ei toimu (meioosi teine jagunemine puudub). Küll aga
haploidiseerub sugulist paljunemist omav leivapärmseen, tehes
vahepeal oma rakust eoskoti, milles toimuvad
järjestikku mõlemad meioosi jagunemised ja moodustub neli eost.
On andmeid, et
rakutuuma ja mtGenoomi proliferatsioon ei ole sõltumatu. Klonaalsetelt paljunevatel
parasiitsetel kottseentel, nii üherakulisel diploidsel pärmil
Candida albicans (Xu
et al., 1999) kui hüüfe
moodustaval haploidsel taimede vahemädaniku tekitajal
Sclerotinia sclerotiorum (Kohli & Kohn,1998), esineb
nii mtGenoomis kui tuumagenoomis rekombinatsioone (retikulaarsus) (Kohli & Kohn,1998; Couch & Kohn,
2000; Anderson
et al., 2001).
Candida albicans kliinilistel tüvedel pole selliste rekombinatsioonide teke
juhuslik: väheste eranditega vastavad mtGenoomi genotüüpidele kindlad tuumagenoomi genotüüpide klastrid.
Leiti, et
tuumagenoomi muutumisele on kaasatud mtDNA muutumine. Mehhanismi ei teata. Sellised
retikulaarsed rekombinatsioonid võivad olla liigile adaptiivselt kasulikud ja omada olulist tähtsust tänapäevaste
kliiniliste, HIV viirust kandvatel patsientidel elutseva
C. albicans populatsioonide evolutsioonis (Anderson
et
al., 2001).
Vaatame, kuidas selline tulemus sobib kokku seene mitokondrite endosümbiontse päritoluga
eubakteritest (
Race et al., 1999) (võrdle päritoluga protobiontidest, Mikelsaar, 2000), mille tulemusel enamus
protomitokondri genoomist on üle kandunud tuuma ja tuum on saavutanud juhtrolli mitokondris toimuva
hingamise ja energia tootmise üle (oksüdatiivne fosforüülimine) (joonis 1). Näib, et seente puhul pole transport
mitokondritest tuuma suletud, nagu imetajatel, vaid on käimasolev protsess. Praeguse seisuga on teda, et 95%
mitokondriaalsest genoomist on esindatud ka tuumas (Weber, 1993). Milline ja kui suur oli aga algselt
sisserännanud “mitokondri” genoom võrreldes “tuuma” genoomiga, pole võimalik määrata, sest ei mitokonder
ega tuum ei eksisteeri enam iseseisvalt.
Võrreldes teiste eukarüootidega on seene mitokondril rohkem funktsioone ja tal on säilinud suurem
iseseisvus. Seentel leidub mitokondritega assotsieerunud mitmesuguste erifunktsioonidega plasmiide (esinevad
nii lineaarsete kui suletud DNA-ahelatena), mis reguleerivad kasvu ja vananemist, hüpovirulentsust jm. Samuti
esineb raku tsütoplasmas mitmesuguseid killer-faktoreid, mille abil mürgitatakse oma liigist või mõnest teisest
liigist pärit konkurente. Hulgaliselt esineb ka viirusi, mis teatud viiruste omavahelises kombinatsioonis võivad
osutuda seenele kahjulikuks (näiteks šampinjonikasvandustes). Kõiki neid geene antakse edasi tsütoplamaatilise
pärilikkusega (Weber, 1993).
Joonis 2. Horisontaalse geeniülekande võimalusi seentel.
Plasmiid võib olla kas vabas olekus mitokondri sees või insertsioonina mtDNA koosseisus. Selline
plasmiid meenutab bakteritel teadaolevaid tempereeritud viirusi. Viimased on bakteri koosseisus inaktiivsed ja
paljunevad koos temaga. Ontogeneesi tasemel näib selline funktsioon olevat kottseene
Podospora anserina seniilsusplasmiidil, mis vabaneb mitokondrist raku vananedes ja võib mängida rolli raku lagundamisel
(Jazwinski, 1996; Jamet-Vierny, 1997). Evolutsiooni tasemel näib toimuvat protsess, kus seene mitokondris
muutub tempereeritud viirusetaoline võõrgeen (ekson) inaktiivseks introniks. Sellisesse geneetiliste elementide
klassi, mis võisid olla või on mtDNA intronite eellased, võib kuuluda liigil
Neurospora crassa valku (710 AH)
kodeeriv plasmiid Mauriceville (3581 Np) (Nargang
et al., 1994) (joonis 2). Võimalik, et selles protsessis
omavad mutageenset rolli oksüdatiivsel fosforüülimisel elektronide transpordil ATP-genereerimisel
toimivad bioenergeetilised protsessid. Hingamisel on elektronide transport läbi membraanide võimas - aga ka ohtlik -
energia allikas (Race
et al., 1999).
Omavahel sarnased mtDNA järjestused SSU rRNA geenis esinevad rohevetika mitokondris ensüümi
kodeeriva eksonina, seentel
Neurospora ja
Podospora võimaliku valgu koodina ning perekonnas
Sclerotinia
intronina, mille erineva paiknemise alusel avaldub mtDNA järjestuse polümorfism.
Esmakordselt avastati mt
SSU rRNA geenis intron 1995. aastal
(Carbone
et al., 1995) (joonis 2). Need mitokondris introniteks
‘degradeerunud’ eksonid võivad anda tuumagenoomi insertsioone ja rekombinatsioone ning avalduda klonaalse
polümorfsusena, nagu monokultuuris kasvava valgemädaniku
tekitaja S. sclerotiorum klonaalselt paljunevatel
tüvedel. Antud juhul põhjustasid kloonide polümorfsuse mittesugulasliku liigi
N. carassa mtDNA järjestused
24S rRNA geenist
(joonis 2). Võimalik, et intronid (prügi
genoomis : endised
viirused , plasmiidid jt võõrgeenid)
võivad mingis positsioonis uuesti aktiveeruda. Toodud näidetest lähtuvalt võib oletada, et seente mitokondritele
on omased samasugused geeniülekande viisid nagu bakteritel - konjugatsioon,
transduktsioon ja
transformatsioon. Tuuma ja mitokondri kohta võiks veel esitada küsimuse, kumb on kumma ’prügikast’,
bakteritaolise käitumisega on mõlemad.
Mõeldes seene
tuuma DNA järjestuste HGT-st võiks spekuleerida järgmiselt. Seene
rakutuum on
põhiaja seene elutsüklist haploidne nagu mitokondergi (mõlemad koosnevad ühekordsest DNA kaksikahelast).
Seentel asuvad
homoloogilised kromosoomid eraldi tuumades, mis kumbki algul eraldi ja enne meioosi
sünkroonselt teineteise kõrval mitootiliselt jagunevad. Hüüfe moodustavate seente ja loomade põhierinevuseks
võibki pidada just seente “oskamatust” diploidsena mitootiliselt jaguneda. Enne haploidsete tuumade
liitumist diploidseteks võib toimuda kromosoomide replikatsioon, kuid sellele ei järgne homoloogiliste kromosoomide
tütarkromatiidide lahknemine (mitoosile omane) vaid tuumade ühinedes liituvad replitseerunud
(homoloogilised?) kromosoomid (meioosile omane). Nüüd peaks
järgnema meioosile iseloomulik
homoloogiliste tütarkromatiidide liitumine ja sünaptoneemsete komplekside teke, kuid näib, et mõnel juhul
tütarkromatiidid ei jõua teineteisest eraldudagi. Kuidas muidu seletada austerserviku viljakehast kahe erineva
tuumagenoomi suurusega eospopulatsiooni teket (Kullman, 2000). Meioos pole normaliseerinud kromosoomide
(genoomi) suurust, nagu näeb ette Zolan’i (1995) poolt välja
pakutud mudel.
Ilmselt paaruvad homoloogilised kromosoomid vaid osaliselt. Võimalik, et ka siin, nagu oletatud
mitokondrite puhul, toimub midagi taolist nagu bakterite konjugatsioonil. Meioosi teine jagunemine jätkab
vahepeal pisut segadusse
aetud mitootilist jagunemist. Kui arvestada seda, et lahknevad tuumad võivad säilitada
liitunud tuumade ebavõrdse suuruse, siis pole meioosi esimene jagunemine mitootilist jagunemist eriti
mõjustanud. Seentel on iseloomulik kromosoomi pikkuse polümorfism ja ka aneuploidsus (Larraya
et al., 1999;
Kullman, 2000). Sellisel juhul võivad
osalise homoloogilisusega tütarkromatiidid siduda oma vabaks jäävate
järjestustega teisi tsütoplasmas olemasolevaid geneetilisi elemente (viirusi, plasmiide, transposoone,
mitokondritest pärinevaid DNA-järjestusi). Võimalik, et see süsteem töötab kui “luud, mis pühib tsütoplasmast
prügi tuuma kokku” või “jäävad harja külge kahjulikud
pisikud kinni ja nad ei saa enam edasi tegutseda”. Kas
selline võikski olla üks mehhanismidest, mis tagab horisontaalsele geeniülekandele omase rekombinantsuse,
retikulaarse mosaiiksuse tuumas?
Seente hulgast võiks otsida vastust ka küsimusele
kuidas on tekkinud diploidsus. Näiteks
kandseentel (Coryolus versicolor) on ühe raku mõlemad haploidsed tuumad võrdväärsed. Eose idanemisel moodustub
ühetuumsete rakkudega haploidne
hüüf . Haploidsed tuumad lähevad vastakuti läbi hüüfide vahele tekkiva
anastomoosi, mille käigus mõlemad
hüüfid muutuvad kahetuumseteks tänu sisserändava tuuma järjepidevale
mitootilisele jagunemisele ja migreerumisele naaberrakku.
Seente oluline erinevus teistest eukarüootidest on see, et mitootilise jagunemise ajal säilib tuuma
membraan . Tütartuumade lahknemisel nöördub tuumamembraan nende vahel ja katkeb. Tuumamembraani
säilimine jagunemise ajal võib tagada tulnuktuuma suhteliselt suure autonoomsuse ja vähendab
tsütoplasmaatiliste geneetiliste elementide sattumist tema genoomi. Samas on see kaitse tuumamaterjali
kaotsimineku vastu. Tõenäoselt pärineb kromosoomide polümorfism (Zolan, 1995; Larraya, 1999) ja
aneuploidsus (Kullman, 2000) mitoosi vigadest.
Niisiis on seentele omane suur plastilisus, võime säilida ja säilitada reproduktsiooni käigus tekkinud
vigu. Seene hüüfile on iseloomulik
tipmine kasv ja avatud paljunemissüsteem (puudub embrüonaalne areng,
kudedeks diferentseerumine ). See, mis juhtub tipurakus, määrab temast edasikasvava hüüfi
saatuse .
Samast hüüfist varem hargnenud hüüfid kannavad edasi esialgset genoomi. Harude tekkemehhanismi ei teata.
Põhimõtteliselt on selliselt kasvav
isend igavene koos oma
vigade ja erinevate tuumapopulatsioonidega.
Külmaseene mitokondrid suudavad klonaalsel kasvamisel oma genoomi hästi säilitada. Nad ei muutu, kasvagu
seen kasvõi sadade (leitud kuni 900) hektarite ulatuses tuhandete (2400) aastate jooksul (Volk, 2002). Nii et
seeneriigist võib mõnel juhul leida ‘kasvava puu’ ja vahel isegi näha, kuidas ‘punutakse võrku’ selle okste vahel.
(Sugulise paljunemise, meioosi, tulemusel lahknevad vana ja uus tunnus haploidsetes organismides vahekorras
1:1, diploididele omane retsessiivsus puudub). Siiani on üles leitud ca
100000 seeneliiki, mis on vaid 5-10%
arvatavalt olemasolevatest seeneliikidest (Hawksworth
et al., 1995; Oliver & Schweizer, 1999).
Lõpuks tuleb rõhutada, et need haruldased
juhtumid , millal rolli mängib HGT, on olnud võimalik välja
selgitada genealoogilise pärilikkuse taustal, tänu valitsevale vertikaalse geeniülekande paradigmale.
Tänuavaldus: Palju tänu Mart Viikmaale redaktsiooniliste soovituste eest. Anderson, J. B., Wickens, C., Khan , M., Cowen, L. E., Federspiel, N., Jones, T. & Kohn, L.M. 2001. Infrequent genetic
exchange and
recombination in the mitochondrial genome of
Candida albicans.
Journal of Bacteriology 183: 865-872.
Awadalla, P., Eyre- Walker , A. & Smith, J. M. 1999. Linkage disequilibrium and recombination in hominid mitochondrial DNA.
Science 286: 2524-2525.
Brinkman, F. S., Macfarlane, E. L., Warrener, P. & Hancock, R. E. 2001. Evolutionary relationships among virulence-associated
histidine kinases.
Infection and Immunity 69: 5207-5211.
Brown, J. R. & Doolittle , W. F. 1999. Gene descent, duplication, and horizontal transfer in the
evolution of glutamyl- and glutaminyl-tRNA
synthetases.
Journal of Molecular Evolution 49: 485-495.
Garcia-Vallve, S., Romeu, A. & Palau, J. 2000. Horizontal gene transfer of glycosyl hydrolases of the rumen fungi.
Molecular Biology and
Evolution 17: 352-361.
Carbone, I., Anderson, J.B. & Kohn, L.M. 1995. A group I intron in the mitochondrial small subunit rRNA gene of Sclerotinia
sclerotiorum.
Current Genetics 27: 166-176.
Carbone, I. & Kohn L.M. 2001. A microbial population-species
interface : nested cladistic and coalescent inference with multilocus data.
Molecular Ecology 10: 947.
Couch, B. C. & Kohn, L. M. 2000. Clonal
spread of
Sclerotium cepivorum in onion
production with
evidence of past recombination events.
Phytopathology 90: 514-521.
Gomis-Ruth, F. X., Moncalian, G., Perez-Luque, R., Gonzalez, A., Cabezon, E., de la Cruz, F. & Coll, M. 2001. The bacterial
conjugation
protein TrwB resembles ring helicases and F1- ATPase.
Nature 409: 637-641.
Hawksworth, D. L., Kirk, P.M., Sutton , B.C. & Pegler, D.N. 1995. Ainsworth & Bisby’s Dictionary of the Fungi. CABI, 616 pp.
Holst-Jensen, A., Vaage, M., Schumacher , T. & Johansen, S. 1999. Structural characteristics and possible horizontal transfer of group I
introns
between closely related
plant pathogenic fungi.
Molecular Biology and Evolution 16: 114-126.
Ito, Y. & Hirano, T. 1999. A group I intron in the 18S ribosomal DNA from the parasitic fungus
Isaria japonica.
Journal of Molecular
Evolution 48: 337-340.
Jamrt-Vierny, C., Boulay, J. & Briand, J-F. 1997. Contribution of various classes of defective mitochondrial DNA molecules to
senescence in
Podospora anserina.
Current Genetics 31: 171-178.
Jazwinski, S.M. 1996. Longevity assurance
genes and mitochondrial DNA alterations: yeast and filamentous fungi. In:
Handbook of the
Biology of Aging (4th
edition ), p. 39. Academic Press, New
York .
Kamaletdinowa, F.I. & Vassilyev, A.E. 1982. Cytology of Discomycetes.
Alma -Ata, 176 pp.
Kempken, F. 1995. Horizontal transfer of a mitochondrial plasmid.
Molecular & General Genetics 248: 89-94.
Kempken, F. & Kuck, U. 1998. Transposons in filamentous fungi--facts and
perspectives .
Bioessays 20: 652-659.
Kim, N.S., Park, N.I., Kim, S.H., Kim, S.T., Han, S.S. & Kang, K.Y. 2000. Isolation of TC/AG repeat microsatellite sequences for
fingerprinting rice
blast fungus and their possible horizontal transfer to plant species.
Molecules and Cells 10: 127-134.
Kivisild, T. & Villems, R. 2000. Questioning evidence for recombination in human mitochondrial DNA.
Science 288:
1931 .
Kohli, Y. & Kohn, L.M. 1998. Detection of random
association among loci in clonal populations of
Sclerotinia sclerotiorum.
Fungal
Genetics and Biology 23: 139-149.
Kullman, B. 2000. Application of flow cytometry for measurement of nuclear DNA content in fungi.
Folia Cryptogamica Estonica 36: 31-
46.
Kullman, B. 2002. Nuclear DNA content, life
cycle and
ploidy in two
Neottiella species (Pezizales, Ascomycetes). Persoonia 18: 000-000.
Larraya, L.M., Pérez, G., Peòas, M.M., Baars, J.J.P., Mikosch, T.S.P., Pisabarro, A.G. & Ramírez, L. 1999. Molecular karyotype of
white rot fungus
Pleurotus ostreatus. Applied and Environmental Microbiology 65: 3413-3417.
Mikelsaar, R.H. 2000. Mitokondrite päritolu ja adapatsioon.
Schola Biotheoretica 26: 56-58.
Nara, T., Hashimoto, T. & Aoki, T. 2000. Evolutionary implications of the
mosaic pyrimidine-biosynthetic pathway in eukaryotes.
Gene 257: 209-222.
Nargang, F.E., Bell, J.B., Stohl, L.L. & Lambowitz, A.M. 1984. The DNA sequence and genetic organization of a
Neurospora mitochondrial plasmid suggest a
relationship to introns and
mobile elements.
Cell 38: 441-453.
Nishida, H., Blanz, P.A. & Sugiyama, J. 1993. The
higher fungus
Protomyces inouyei has two group I introns in the 18S rRNA gene.
Journal of Molecular Evolution 37: 25-28.
Oliver, R. & Schweizer, M. (eds) 1999. Molecular fungal biology.
Cambridge University Press, 377 pp.
Penalva, M.A., Moya, A., Dopazo, J. & Ramon, D. 1990. Sequences of isopenicillin N synthetase genes suggest horizontal gene transfer
from prokaryotes to eukaryotes.
Proceedings of the Royal Society of London, B 241: 164-169.
Race, H.L., Hermann, R.G. & Martin, W. 1999. Way have organelles
retained genomes?
Trends in Genetics 15: 364-370.
Saville, B.J., Kohli, Y. & Anderson, J.B. 1998. mtDNA recombination in a natural population.
Proceedings of the National Academy of
Sciences , USA 95:
1331 -1335.
Zolan, M.E. 1995. Chromosome-length polymorphism in fungi.
Microbiological Reviews 59: 686-698.
Tamura, M., Watanabe, K., Mikami, Y., Yazawa, K. & Nishimura, K. 2001. Molecular characterization of new
clinical isolates of
Candida albicans and
C. dubliniensis in Japan:
analysis reveals a new genotype of
C. albicans with group I intron.
Journal of Clinical
Microbiology 39: 4309-4315.
Taylor, J.W., Jacobson, D.J. & Fischer, M.C. 1999. The evolution of asexual fungi: reproduction, speciation and
classification .
Annual
Review of Phytopathology 37: 197-246.
Vaughn, J.C., Mason, M.T., Sper -Whitis, G.L., Kuhlman , P. & Palmer, J.D. 1995. Fungal
origin by horizontal transfer of a plant
mitochondrial group I intron in the chimeric CoxI gene of
Peperomia.
Journal of Molecular Evolution 41: 563-572.
Volk, T. 2002. The humongous fungus – ten
years later.
Inoculum (Supplement to Mycologia) 53: 4-8.
Weber, H. 1993. Allgemeine Mykologie. Gustav Fischer Verlag
Jena , Stuttgart, 541 pp.
Xu, J., Vilgalys, R. & Mitchell, T.G. 1999. Lack of genetic differentiation between two
geographic samples of
Candida albicans isolated
from patients infected with the Human Immunodeficiency
Virus .
Journal of Bacteriology 181:
1369 -1373.
























Kõik kommentaarid