Geneetika eksam (0)
Geneetika 2 kordamisküsimused
Lisaks tekstile ja õpikule vaadake kindlasti ka materjali slaididelt.
1. Võrrelge lüütilq
aaise ja mõõduka bakteriofaagi paljunemistsüklit
VIRULENTSED FAAGID – põhjustavad peremeesraku surma
MÕÕDUKAD FAAGID – võivad püsida rakus ilma seda hävitamata
o Lüütiline ja lüsogeenne fvgvb89htsükkel. Lüsogeenne tsükkel võib
keskkonnatingimuste muutudes üle minna lüütiliseks tsükliks
Lüütiline
: kinnitub peremeesrakule antiretseptori vahendusel; genoom sisestatakse rakku,
tehakse palju DNA/RNA koopiaid, viiruspartiklid pannakse kokku, rakk lüüsitakse
Lüsogeenne
: kinnitub peremeesrakule antiretseptori vahendusel; genoom sisestatakse
rakku, genoom integreerub peremehe genoomi ja kandub kromosoomi koostisosana
tütarrakkudesse. Keskkonnatingimuste muutudes võib lüsogeenne faag minna üle lüütilisse
tsüklisse, mille käigus sünteesitakse viiruse partikleid ning pannakse need kokku. Lõpuks rakk
lüüsitakse ning uued virionid vabanevad keskkonda, kus nad on võimelised minema
nakatama järgmisi peremeesrakke
2. Gripiviiruse muteerumine, rekombinantsed gripiviirused ja nendega seotud ohud
Gripiviirus:
o Pinnavalgud hemaglutiniin
(H)
ja neuraminidaas (N) muteeruvad kiiresti – RNA
polümeraas teeb palju vigu, mida ei parandata!
o Gripiviiruse genoom koosneb 8-st erinevast RNA segmendist, mis pakitakse valkkesta
viiruse assambleerimisel
o Geneetilise materjali segunemine võib toimuda siis, kui 2 sarnast viirust nakatavad
sama rakku
Rekombinantsed girpiviirused ja nendega seotud ohud:
Linnugripp
(H5N1)
:
o Kandub lindudelt inimestele
o Kardeti
viirustevahelist rekombineerumist – uus tüüp, mille vastu puudub täielikult
immuunsus
o Hispaania gripp 1918 – pandeemia; viirus suutis välja lülitada RIG1 geeni, mis
põhjustab immuunsüsteemi ülereageerimise
H1N1 tüvi – seaviirus, „uut tüüpi“ viirus:
o RNA segmendid on segunenud
o Segunenud segmendid on pärit erinevatelt gripiviirustelt
(kahelt seaviiruselt, inimese
viiruselt ja linnuviiruselt)
o Hakkas levima Mehhikos
o Kandub inimeselt inimesele
o Kardeti pandeemiat
3. Selgitage retroviiruse paljunemist HIV näitel. HIV-ga nakatunute ravi
Retroviirused:
o RNA genoomilt sünteesitakse pöördtranskriptaasi
(revertaasi)
abil DNA, mis
integreerub raku genoomi
o Raku kromosomaalse DNA koostises olevalt viiruse genoomilt toimub RNA süntees:
mRNA – viiruse geenide avaldumine; genoomne RNA – viiruse genoomi paljunemine
o AIDS – haigust põhjustab HIV, mis nakatab T-abistajarakke, mis aktiveerivad teisi
immuunrakke; HIV seondub T-abistajarakkude pinnal olevatele CD4/CCR5
retseptoritele
o Muteerub kiiresti: 1. Pöördtranskriptaas teeb DNA sünteesil palju vigu; 2. Viiruse
kapsiidi pakitakse 2 koopiat RNA molekuli → DNA sünteesil võib toimuda matriitsi
vahetus → tekib rekombinantne viirus, kui genoomi koopiad ei ole identsed
HIV on retroviirus – tema genoomiks on ssRNA, millelt pöördtranskriptaas sünteesib DNA.
RNA lülitub peremehe genoomi suvalisse kohta, kui on sünteesitud DNA, siis saab edasi
sünteesida viirusvalke jms.
HIV-ga nakatunute ravi:
o Pärast nakatumist järgneb akuutne faas – palavik, peavalu, veres palju HIV partikleid.
Järgneb latentne periood (8-10 aastat) – viirus lokaliseerub lümfisüsteemi. Lõppfaas –
T helperrakkude arv on langenud > 75%, immuunsüsteem ei toimi enam
o Ravimeid HIV-st vabanemiseks pole leitud
o Ravi toimub iga patsiendi jaoks individuaalse raviplaani alusel
o Raviks kasutatakse antiretroviiruseid
(aeglustab HIV paljunemist ning ei lase HIV-l
nakatada terveid rakke)
või siis HI-viiruse vastaseid ravimeid (viimasel juhul kestab
ravi elu lõpuni)
o Ravimid on mõeldud takistamaks HIV paljunemist, inhibeeritakse pöördtranskriptaasi
ja proteaasi tööd
Nt:
Epvir
ja Retrovir – pöördtranskriptaasi inhibiitorid; Crixivan – proteaasi inhibiitor
(primaarne translatsiooniprodukt lõigatakse proteaasi abil erinevateks viirusvalkudeks).
4. Bakterite kaitsemehhanismid bakteriofaagide vastu
Restriktsiooni-modifikatsiooni süsteem bakterites:
o Kaitseb bakterirakku bakteriofaagide eest
o W. Arber leidis, et T4 paljunemine on osades
E.coli
tüvedes restrikteeritud (piiratud)
o E.coli
tüve B rakkudes paljundatud faagi edasine paljunemine on takistatud tüve K
rakkudes ja vastupidi
o Rakus lagundatakse võõrast DNA-d, mis ei ole antud bakteritüve restriktaasi eest
kaitstud
o CRISPR –
clustered, regulatory interspaced short palindromic repeats
o Segment bakteriofaagi genoomist integreerub bakteri kromosoomi spetsiifilisse
CRISPR lookusesse
o CRISPR lookus sisaldab peaaegu identseid kordusjärjestusi ja nende vahele jäävaid
vahealasid (
spacers
), mis pärinevad erinevate viiruste genoomidest
o CRISPR lookuselt transkribeeritud RNA (crRNA) ja CAS geenide poolt kodeeritud
valkude vahel moodustub kompleks, mis seondub rakku nakatava viiruse genoomile
ja lagundab selle
5. Kuidas on võimalik uurida ja kirjeldada bakteriofaagide mutante? Põhilised
avastused bakteriofaagide geenide kaardistamisest
Mutantide uurimine ja kirjeldamine:
o Teostatakse geneetiline rekombinatsioon
(ristatakse erinevaid faage)
o Mutantidel on
wild-type
’ist erinev koloonia morfoloogia
o Mutantidel võib olla suurem või väiksem peremeestering
o Arvutatakse rekombinatsioonisagedust:
2 x metsiktüüpi rekombinantide arv
Faagide üldarv
Faagide geenide kaardistamine:
o Faagide geneetiline kaardistamine
– geeni peenstruktuuri määramine Benzeri
laboris:
1) Kõige väiksemaks mutatsiooniüksuseks on üks aluspaar (bp)
2) Rekombinatsioon võib toimuda kahe külgneva aluspaari vahel
3) Lükati ümber varasem seisukoht nagu oleks geen jagamatu ja seega kõige väiksem
mutatsiooni ja rekombinatsiooni üksus
o Deletsioonmutantide kasutamine geneetilises analüüsis
Kui mutatsioonikohad paiknevad geneetilsel kaardil lähestikku, on rekombinatsioonisagedus
madal:
1) Rekombinatsioonisagedusest tuleb maha arvata spontaansete revertantide
tekkesagedus
2) Kaardistatakse deletsioonmutante, mis ei ole võimelised spontaanselt
metsiktüübiks reverteeruma
6. Plasmiidide roll bakterite keskkonnaga kohastumisel. Plasmiidide klassifitseerimise
alused
Plasmiid:
PLASMIIDID – autonoomselt replitseeruvad ekstrakromosomaalsed DNA elemendid,
valdavalt rõngasmolekulid
o Suurus varieerub 1-200 kb ulatuses
o Megaplasmiidid > 200 kb
EPISOOMID – plasmiid, mis võib integreeruda kromosoomi (näiteks F plasmiid)
Plasmiidides sisalduv geneetiline informatsioon võimaldab bakteritel paremini kohastuda
keskkonnatingimustega
(näiteks resistentsus antibiootikumidele).
1) Resistentsus antibiootikumidele
– R-plasmiidid. Antibiootikumi modifitseerimine
(kloroamfenikooli atsetüül transferaas); antibiootikumi degradeerimine
(β-laktamaas degradeerib penitsilliine); antibiootikumi rakust väljapumpamine
(resistentsus tetratsükliinile)
2) Virulentsusfaktorid, toksiinid
3) Bakteriotsiinid
– nt: ColE1 poolt on kodeeritud E. coli
vastased kolitsiinid E1
(permeabiliseerib tsütoplasma membraani), E2 (DNA degradatsioon) ja E3
(ribosomaalse RNA degradatsioon)
4) Antibiootikumide produktsioon streptomütseetidel
5) Võime kasutada alternatiivseid kasvusubstraate
– mullabakterid kasutavad
süsinikuallikana fenoolseid ühendeid
6) Agrobacterium
'i Ti plasmiidid indutseerivad taimedel tuumoreid
Plasmiidide klassifitseerimise alused:
1) Funktsioonide alusel
2) Sobivusgruppide
alusel – samasse gruppi kuuluvad plasmiidid ei püsi koos
3) Peremeesring
o Kitsa peremeesringiga plasmiidid
– replitseeruvad üksnes lähedases suguluses
olevates bakterites
o Replitseeruvad väga erinevates bakteriliikides
– võimaldavad geenide
horisontaalset ülekannet erinevate liikide vahel
4) Konjugatiivsed ja mittekonjugatiivsed plasmiidid
7. Geneetilised elemendid, mis lisaks plasmiididele võimaldavad geenide
horisontaalset ülekannet bakteritel
Lisaks plasmiididele võimaldavad geenide horisontaalset ülekannet bakteritel:
1) Transposoonid
– kuna enamus transposoone kannavad antibiootikumide
resistentsusgeene, soodustab transposoonide hüppamine ühest DNA molekulist teise
(nt: plasmiidist kromosoomi ja vastupidi) resistentsuse kiiret levikut
bakteripopulatsioonis
Meditsiinis on suur probleem patogeensete bakteritega, kes on endale hankinud erinevaid
resistentsusgeene, olles sel viisil resistentsed mitmele antibiootikumile korraga.
Multiresistentsus levib näiteks R plasmiidide abil.
2) Integronid
– koht-spetsiifiline rekombinatsioon on vahendatud integraasi kaudu:
integraas lülitab geenikassette integroni, kasutades substraadina
attC
järjestust
3) Genoomsed saared
(GEI)
– horisontaalselt ülekandunud DNA regioonid; sisaldavad IS
elemente ja integraasi geene
4) Patogeensussaared
– sisaldavad virulentsusgeene
8. Bakterite mutandid ja nende kirjeldamine; mutantide isoleerimine
bakterigeneetikas
Bakterite mutandid ja nende kirjeldamine:
1) Antibiootikumile resistentsed mutandid
– võimelised kasvama ja moodustama
üksikkolooniaid söötmel, mis sisaldab teatavat antibiootikumi
2) Auksotroofsed mutandid
– ei kasva minimaalsöötmel, mis sisaldab ainult
süsinikuallikat ja mineraalsooli, kuna ei ole võimelised sünteesima mõnda
aminohapet või nukleotiidi
3) Süsinikuallika mutandid
– ei suuda kasvuks kasutada teatavat süsinikuallikat, näiteks
laktoosi
4) Muud tüüpi mutandid
– kolooniate morfoloogia; faagiresistentsus;
temperatuuritundlikkus
Mutantide isoleerimine bakterigeneetikas:
1) Lokaliseeritud mutagenees
o Uuritavassse geeni viiakse mutatsioonid
(ainult mingi lõik DNA-st)
ning
homoloogilise rekombinatsiooni teel asendatakse bakteri genoomis algse
geeni järjestus defektse geeni järjestusega
2) Negatiivne selektsioon
o Kasutatakse „rikastusmeetodit“, kus teatud söötmel kasvavad metsiktüüpi
rakud lüüsuvad nt penitsilliini juuresolekul
(algselt kasvatatakse aukso- ja
prototroofe rikkal mutageniseeritud söötmel, resuspendeeritakse söötmele,
kus on mingi aine puudu ning lisaks on penitsilliin; prototroofid suudavad
kasvada ning 99% neist hukkuvad penitsilliini tõttu; auksotroofid ei suuda
kasvada ja jäävad ellu; auksotroofid külvatakse järgmisele söötmele...)
3) Transposoonmutagenees:
o Transposoon
(mobiilne DNA element)
, mis kannab mingit geneetilist
markerit
, nt resistentsust AB-le, inserteerub kromosoomi erinevatesse
kohtadesse ning põhjustab insertsioonikohas asuvate geenide
inaktivatsiooni
o Paljudele bakteritele eluliselt tähtsate geenide inaktivatsioon on rakule
letaalse toimega
o Isoleeritakse ajutise fenotüübiga mutante, nt temperatuuritundlikke
(Ts)
mutante
9. Geneetilise informatsiooni ülekanne transformatsiooni teel: avastamine,
mehhanismid, rakkude kompetentsus. Kuidas on võimalik kindlaks teha, et DNA
kandus üle just transformatsiooni teel?
Avastamine:
1928 – F. Griffith avastas transformatsiooni
S. pneumoniae’s
. II R – kapslita rakud, ei ole
virulentsed, väikesed karedapinnalised kolooniad; III S – limakapsliga rakud, virulentsed,
suured siledapinnalised kolooniad.
1944 – Oswald Avery, Colin MacLeod ja Maclyn McCarty näitasid, et
S. pneumoniae
mittevirulentsete IIR tüüpi rakkude transformeerimist IIIS tüüpi virulentseteks rakkudeks
põhjustas DNA:
DNA preparaati töödeldi erinevate ensüümidega, mis lagundasid kas DNA,
RNA või valgud. Transformatsioonivõime kadus, kui preparaati töödeldi DNaasiga.
Geneetilise informatsiooni ülekanne bakteritel:
1) Transformatsioon
– doonori DNA satub retsipientrakku väliskeskkonnast (seostub
raku genoomi rekombinatsiooni teel)
2) Transduktsioon
– bakteri DNA kandub ühest rakust teise bakteriofaagide abil
3) Konjugatsioon
– DNA ülekanne doonorist retsipienti vajab rakkudevahelist kontakti;
üle võib kanduda plasmiid või ka kromosoom
(selleks on vaja kindlat plasmiidi, nt
F-plasmiid E.coli’l)
Transformatsioon:
TRANSFORMATSIOON
– protsess, kus retsipientrakk omandab vaba DNA ümbritsevast
keskkonnast
Enne, kui DNA siseneb rakku, lagundatakse üks ahel membraanseoselise eksonukleaasiga.
Rakku jõudnud ssDNA rekombineerub bakterikromosoomiga homoloogilise
rekombinatsiooni teel. Kui nukleotiidid ei ole täielikult komplementaarsed, parandatakse
vead
(mismatch repairation)
või asendatakse kas retsipient- või doonor-DNA lõik.
o DNA toiduks
o DNA reparatsiooniks
o DNA evolutsiooniks
Looduses kinnituvad bakterid pindadele ja moodustavad biofilmi ehk biokilet. Vaba DNA
(eDNA)
aitab moodustuda biofilmi.
Kunstlik transformatsioon:
E.coli
ei ole looduslikult kompetentne, kuid saavutab kompetentsuse rakkude töötlemisel
CaCl
2
lahusega. Töötluse tulemusena muutuvad rakud DNA suhtes läbilaskvaks ning
rakusisesed nukleaasid on ajutiselt inaktiveeritud.
Toimib efektiivselt plasmiidide korral
(dsDNA !!!!!)
, mis on võimelised retsipientrakus iseseisvalt replitseeruma. Lineaarse DNA
puhul võib väga madala sagedusega toimuda rekombinatsioon
(kui esineb homoloogia
doonori ja retsipiendi DNA vahel)
, kuid enamasti lagundatakse DNA fragment eksonukleaasi
toimel.
Transformatsiooni abil on võimalik uurida geenidevahelist distantsi:
Lähestikku asuvate geenide puhul on nende poolt kodeeritud markertunnuste
kotransformatsiooni sagedus kõrgem kui üksteisest kaugel paiknevate geenide puhul
(rekombinatsiooni sagedus on vastupidi)
.
Otsesed tõendid, et DNA on geneetiline materjal:
1928 – F. Griffith avastas transformatsiooni
S. pneumoniae’s
.
Streptococcus pneumoniae’
l on kahte sorti rakke:
1) II R – kapslita rakud, ei ole virulentsed, väikesed karedapinnalised kolooniad
2) III S – limakapsliga rakud, virulentsed, suured siledapinnalised kolooniad
*Elus S-rakkude hiirde viies hiir sureb; elus R-rakkude hiirde viies hiir jääb ellu; kuumusega
töödeldud S-rakkude hiirde viies hiir jääb ellu; kuumtöödeldud S-rakkud ja elus R-rakkude
hiirde viies hiir sureb, sest on toimunud transformatsioon*
Rakkude kompetentsus:
DNA sisenemiseks rakku peavad rakud olema eelnevalt kompetentsed.
o Loomulik kompetentsus
– esineb nt S.pneumoniae’l; B.subtilis’el
ja H.influenzae’l
(H.influenzae võtab vastu üksnes sama liigi DNA-d)
o Kunstlik kompetentsus
– saavutatud laboritingimustes
Kompetentsetel rakkudel on rakupinnal retseptorid, mis on vajalikud eksogeense DNA
esmaseks seondumiseks. Ekspresseeruvad DNA rakku transportimiseks vajalikud valgud.
S.pneumoniae
puhul saavutab esialgu kompetentsuse üksnes väike osa rakupopulatsioonist,
mis seejärel väljutab kasvukeskkonda kompetentsusfaktoreid. Kompetentsus saavutub
rakukultuuri teatavas vanuses, sageli eksponentsiaalses kasvufaasis.
Geneetilise info ülekandumisviisi kindlakstegemine:
U-toru katse:
Kasutatakse U-kujulist klaastoru, mille õlad on teineteisest isoleeritud klaasfiltriga. Filtrit
läbivad DNA molekulid ja bakteriofaagid, kuid mitte bakterid. Toru erinevates õlgades on
erineva genotüübiga bakteritüved ja jälgitakse rekombinantide teket.
1) Filti tõttu on välistatud erineva genotüübiga bakterite rakkudevaheline kontakt –
rekombinante ei saa tekkida konjugatsiooni tagajärjel
2) DNaasi lisamisel kasvukeskkonda lagundatakse rakuväline DNA – välistab
transformatsiooni toimumise
3) Rekombinantide teke DNaasi sisaldavas keskkonnas viitab transduktsioonile
10. Geneetilise informatsiooni ülekanne bakterite konjugatsiooni teel. Bakteri
kromosoomi geneetiline kaardistamine bakterite konjugatsiooni abil
Bakterite konjugatsioon:
Bakterite geneetilise informatsiooni ülekandumise rakust rakku
konjugatsiooni teel
avastasid 1946. aastal Lederberg ja Tatum.
o Konjugatsiooni toimumiseks
peavad rakud kontaktis olema
o Rakkudevahelist
kontakti aitavad saavutada doonorraku sugukiud e piilid
Bakterivaheliseks konjugatsiooniks on vajalikud
tra
geenid:
o Bakterirakkude liitumine ja DNA liikumine rakust rakku
Konjugatiivsed plasmiidid võivad mobiliseerida mittekonjugatiivsete plasmiidide ja
kromosomaalsete geenide ülekandumist.
Bakteris
E.coli
on kirjeldatud F-plasmiid e fertiilsusfaktor, mis sisaldab tra geene.
o DNA kandub üle doonorist retsipienti läbi konjugatsioonikanali
(konjugatsioonisilla)
F-plasmiidi ülekanne doonorilt retsipienti:
1) F-pilide kaudu luuakse kontakt
: doonori F-pili tunneb ära retsipiendi retseptorid
2) Plasmiidi ülekanne
: üks plasmiidi ahel doonoris (F
+) lõigatakse katki ning hakatakse
DNAd sünteesima veereva ratta mudelil
3) Replitseeritud DNA ahel liigub retsipienti
(F
-), kus sünteesitakse sellele DNA ahelale
komplementaarne ahel
4) Retsipient muutub F
+-ks (transkonjugant ehk rekombinant)
Hfr
(high frequency recombination)
bakteritüved: F plasmiid integreerub bakteri
kromosoomi.
E.coli
rakkude tähistus lähtudes F-plasmiidi olemasolust ja asukohast:
1) F-plasmiid puudub – F
-
2) F
+-- F-plasmiid olemas
3) Hfr – F-plasmiid integreerunud kromosoomi
F-plasmiidi integreerumine kromosoomi:
F-plasmiidi integreerumine toimub HR-i teel. Homoloogia aluseks on IS elemendid
(insertion sequences)
. Hfr tüvest toimub kromosomaalsete geenide ülekanne alates
integratsioonisaidist kindlas järjekorras. Enamus F-plasmiidist kandub üle kõige viimasena.
Erinevates Hfr tüvedes on F-plasmiidid integreerunud erinevatesse kohtadesse ja erinevas
orientatsioonis: kromosomaalsete geenide ülekanne neist tüvedest algab erinevatest
geenidest ja toimub erinevas järjekorras.
Konjugatsiooniprotsess bakterites:
o Konjugatsiooniprotsessi käigus toimub DNA replikatsioon veereva ratta mudelil
o Retsipientrakku kandub üle ainult ssDNA, millele komplementaarne DNA ahel
sünteesitakse retsipientrakus
o Kogu bakteri kromosomaalse DNA ülekandumiseks doonorist retsipienti kulub ~100
min
o Enamasti kaob rakkudevaheline kontakt enne protsessi lõpulejõudmist
o Ülekandunud DNA fragment rekombineerub retsipiendi kromosoomi homoloogilise
DNA järjestusega
o Toimub paarisarv DNA ahelate vahetusi
(2 ristsiiret)
o Transkonjugandi
(ehk rekombinandi)
järglaskonda kandub edasi ainult
kromosomaalne DNA; kromosoomiväline DNA fragment läheb kaotsi
o Hfr tüvede avastamine võimaldas välja töötada bakterikromosoomi geneetilise
kaardistamise metoodika, mis põhineb konjugatsiooniprotsessi katkestamisel
erinevatel ajamomentidel: vaja söödet, kus saavad kasvada ainult rekombinandid
(rekombinandil on mingi tunnus, mida doonoril ei ole! Nt on retsipiendid
streptomütsiin-resistentsed)
. Kaart on jagatud 100 minutiks – 0-punkti on paigutatud
thr
(treoniini süneesi) lookus: asus kõige esimesena kirjeldatud F-plasmiidi
integratsioonisaidi kõrval
F’ konjugatsioon e seksduktsioon:
1) F’-plasmiidi teke: F-plasmiid võib kromosoomist välja tulla ebatäpselt, haarates kaasa
külgnevaid geene
(nt lac geene – laktoosi lagundamine)
lac geenide sage ülekanne
2) Kromosomaalsete geenide ülekannet F’-plasmiidi koostises nim seksduktsiooniks:
seksduktsiooni tulemusena on rakud F-plasmiidis ja kromosoomis samaaegselt
asuvate geenide suhtes diploidsed.
Võimaldab hinnata kromosoomis ja plasmiidis
olevate alleelide dominantsust ja retsessiivsust
. F-plasmiidi lülitunud geenid peavad
asuma lähestikku.
Genoomi kaardistamine konjugatsiooni teel:
Hfr tüvede avastamine võimaldas välja töötada bakterikromosoomi geneetilise
kaardistamise metoodika, mis põhineb konjugatsiooniprotsessi katkestamisel erinevatel
ajamomentidel.
11. Kuidas toimub bakteriofaagi genoomi pakkimine valkkesta? Geneetilise
informatsiooni ülekandumine üldise ja spetsialiseeritud transduktsiooni teel
Faagi genoomi pakkimiseks valkkesta on faagi genoomis kindlad järjestused, mis on
markeriteks. Valkkesta pakitakse tavaliselt kuni 105% genoomist. Sõltuvalt sellest, kas faag
omab lüütilist või lüsogeenset tsüklit, võidakse valkkesta pakkida ka mingi osa peremehe
genoomist.
Üldine transduktsioon:
Üldine transduktsioon toimub
suvalise geneetilise markeri suhtes, kuna faagi valkkesta
võidakse eksikombel pakkida suvaline DNA fragment bakteri genoomist. Transduktsioonil
kasutatakse faagidest enim P1
(genoom: ~100 kb)
. Toimub madala sagedusega; moodustub
1-2% faagipartikleid, mis sisaldavad bakteri DNA-d.
Spetsialiseeritud transduktsioon:
Ülekantavad geenid pärinevad kromosoomi kindlast alast, mis külgneb sinna
inserteerunud faagi genoomiga
. Kui faag lambda on integreerunud E.coli
kromosoomi gal ja
bio
geenide vahele, võib ta kromosoomist välja tulla ka ebatäpselt. Ebatäpne väljatulek
toimub sagedusega 10
-4 ning sel juhul on osa faagi geenidest asendatud bakteri geenidega.
Hübriidse faagi genoom on sageli defektne
– kui faagi genoomi on lisandunud bakteri
geene, läheb hübriidse genoomi kapsiidi pakkimisel osa faagi geene kaotsi.
Kromosoomist ebatäpselt välja tulnud faag
(defektne!)
vajab paljunemiseks helperfaagi,
mis kompenseeriks neid geene, mis on defektsel faagil puudu!!!
Kui hübriidne faag nakatab
uut rakku ja integreerib oma genoomi peremehe genoomi, siis sisaldab peremehe genoom
nii bakteri enda kui ka faagi ning eelmise peremehe geneetilist materjali.
12. Bakteriaalsed IS elemendid ja transposoonid, nende koostisosad. Konservatiivse ja
replikatiivse transpositsiooni mehhanismid
Põhilised transponeeruvad elemendid bakterites – IS elemendid ja transposoonid:
1) IS elemendid
(insertion sequences)
– väikesed, 800-2000 bp pikkused DNA
elemendid.
Kodeerivad ainult transpositsiooniks vajalikke valke. Sama IS-i võib olla
rakus mitu koopiat.
Kui sama IS elemendi koopia on nii plasmiidis kui ka
kromosoomis, võib plasmiid homoloogilise rekombinatsiooni teel kromosoomi
integreeruda
. Sel viisil toimub F-plasmiidi integreerumine kromosoomi.
IS elementide otstes on pöördkordusjärjestused, tavaliselt 16-40 nt pikkused. Mutatsioonid
IR-ides muudavad elemendi transpositsiooniliselt inaktiivseks. Mõlemale poole IS
elemendist jäävad lühikesed, enamasti mitte üle 10 nt pikkused
otsekordusjärjestused
(direct repeats, DR)
. DR-id tekivad märklaud-DNA
duplitseerumise käigus
Otsekordusjärjestuste tekkimine transpositsiooni käigus: transpositsiooni käigus lõhutakse
fosfodiestersidemed märklaud-DNA-s nii, et tekivad lühikesed üksikahelalised otsad.
Mobiilne DNA element lülitub lõikekoha vahele ning üksikahelalistele märklaud-DNA
lõikudele sünteesitakse komplementaarsed DNA ahelad.
o DR-id esinevad nii transposoonide kui ka IS elementide puhul
2) Transposoonid
(Tn)
– IS elementidest suuremad. Kodeerivad lisaks transpositsiooniga
seotud valkudele ka teisi valke
(nt resistentsus AB-dele, raskemetallidele)
a) Komplekssed Tn-id
– sisaldavad mõlemas otsas IS elementi. Tn10 kannab endas
tetratsükliini resistentsusgeeni, sisaldab mõlemas otsas IS elementi IS10
b) Lihttransposoonid
– Tn3 perekonda kuuluvad Tn-id IS elemente ei sisalda.
Sarnaselt IS-idele on neil otstes lühikesed, sageli 38 bp pikkused IR-id
Konservatiivse transpositsiooni mehhanismid:
Konservatiivne e lõika ja kleebi transpositsioon
– Tn või IS lõigatakse doonormolekulist välja
ja viiakse märklaudmolekuli
(elemendi koopiaarv rakus ei suurene!)
. Toimub osade
IS-elementide ja komplekssete Tn-ide puhul.
Replikatiivse transpositsiooni mehhanismid:
Replikatiivne
transpositsioon (nt Tn3 tüüpi elemendid)
– transpositsiooni käigus toimub
transponeeruva elemendi replikatsioon, nii et lõpptulemusena jääb koopia transposoonist nii
doonori kui ka retsipiendi molekuli
(koopiaarv suureneb!)
.
Mehhanism
:
1) Kointegraadi moodustumine transposooni ja märklaud-DNA vahel; transposaas TnpA
lõikab doonormolekuli transposooni otste kõrvalt – vabanevad transposooni DNA
3’-OH otsad, mis atakeerivad fosfodiestersidemeid märklaud-DNA-s, tekitades
insertsioonisaiti üksikahelalised katked. Järgnevalt viiakse kokku transposooni ja
märklaud-DNA otsad. Tn3 replitseerub ning tekib kointegraat, kus mõlemad samas
orientatsioonis asuvad transposooni koopiad on ühenduses nii doonori kui ka
märklaud-DNA-ga
2) Kointegraadi lagunemine resolvaasi TnpR toimel, toimub sait-spetsiifiline
rekombinatsioon kahe Tn3 koopia vahel
res
saidis
13. Transposoonidest põhjustatud geneetilised ümberkorraldused. Millised
mutatsioonid võivad põhjustada transposoonide inaktiveerumist?
TE-d põhjustavad ümberkorraldusi kromosoomide struktuuris:
o HR kromosoomi erinevates osades paiknevate homoloogiliste elementide vahel
tekitab deletsioone, inversioone või duplikatsioone
o Kui 2 rekombineeruvat transposooni on kromosoomis samasuunaliselt, siis nende
vahele jääv ala deleteerub
o Kui 2 rekombineeruvat transposooni on kromosoomis vastupidises orientatsioonis,
siis nende vahele jääv ala inverteerub
(pöördub ümber)
o TE-d võivad indutseerida kromosoomidevahelisi ümberkorraldusi nii homoloogiliste
kui ka mittehomoloogiliste kromosoomide puhul. Tütarkromatiidide vahelise
ebavõrdse ristsiirde tulemusena tekib 2 produkti: 1) kromosoom, milles
transposoonidevaheline ala on duplitseerunud; 2) kromosoom, millest
transposoonidevaheline ala on kaotsi läinud
o Inimesel on
Alu
järjestuste poolt põhjustatud ebavõrdse homoloogilise
rekombinatsiooniga seotud >30 päranduvat haigust ja 16 erinevat vähijuhtu
o Glycogen storage disease
– L1 elementide vahelt on deleteerunud 7,5 kb DNA-d;
metabolismihäired
o Geenide duplikatsioon ja pseudogeenide teke LINE elementide kaudu.
Retrotransposoonid võivad transponeerumisel kaasa haarata külgnevaid alasid
Transposoonide inaktiveerumist võivad põhjustada:
Transpositsiooni viib läbi transposaas, mis seondub spetsiifiliselt elemendi otstes olevatele
pöördkordusjärjestustele
(inverted repeats IR)
. Mutatsioonid IR-ides või transposaasis
inaktiveerivad elemendi.
14. Tooge näiteid DNA transposoonidest eukarüootides. P-elementidest põhjustatud
hübriidne düsgenees, selle mehhanismid
1) RNA elemendid
– retrotransposoonid, transponeeruvad üle RNA vahevormi. Kui
transposoon paikneb genoomis, siis on sellel RNA vaheühend, millelt sünteesitakse
DNA, mis seejärel lülitub mingisse genoomi piirkonda
2) DNA transposoonid
Taimedel on lisaks klass I ja klass II elementidele ka helitronid.
o Maisi Ac ja Ds elemendid
o P elemendid äädikakärbestes
Hübriidne düsgenees:
äädikakärbse liinide M ja P ristamisel tekib järglaskond, mille puhul
ilmnevad katked kromosoomides ning on kõrgenenud mutatsioonisagedus
. F1 põlvkonna
järglased ei ole elujõulised, sest neil on palju mutatsioone.
Transponeeruv element esines
ainult P tüves.
P elemendi transpositsiooni võimaldas M tsütotüüp (munaraku tsütoplasma).
P elemendid transponeeruvad ainult sugurakkude eellastes
– F1 põlvkond on elujõuline.
P tsütotüüp pärsib
P
elemendi transponeerumist ja on iseloomulik liinidele, mis sisaldavad
genoomis
P
elemente. M tsütotüüp võimaldab P elementide transponeerumist, kuna
puudub transpositsiooni pärssiv faktor
. M tsütotüüp on iseloomulik liinidele, mille
kromosoomides
P
elemendid puuduvad. F1 põlvkonna somaatilistes rakkudes on P
elementide liikumine takistatud, kuna transposaasi mRNA alternatiivse splaissingu tõttu
sünteesitakse defektne valk. Et mutatsioonid avalduda saaksid, F1 hübriide ristatakse. F1
põlvkonna järglastes on mutantsed geenid
P
elemendi poolt märgistatud, lihtsustades
sellega nende geenide isoleerimist ja identifitseerimist
.
15. Retroviiruse-laadsed elemendid ja retroposoonid eukarüootides. Enimlevinud
transponeerivad elemendid inimese genoomis
Retroviiruse-laadsed elemendid:
Ehitus sarnaneb retroviiruste genoomile.
Elementide mõlemas otsas on LTR järjestused
(long terminal repeats)
, mis on orienteeritud samasuunaliselt ja mõnisada nukleotiidi
pikad
. Elemendi keskel on kodeerivad järjestused, mis on homoloogilised retroviiruste gag
(viiruse kapsiidi struktuurvalk)
ja pol (pöörtranskriptaas/integraas) geenidele.
Ty1 element on leitud pagaripärmist
(saccharomyces cerevisiae)
; Ty element on 5,9 kb
pikkune, sisaldab 340 bp pikkuseid LTR-e. Tekitab transpositsioonil 5 bp pikkuseid
otsekordusjärjestusi. Pärmi genoomis on leitud 35 koopiat Ty elementi. TyA ja TyB
(pöördtranskriptaas ehk revertaas, mis sünteesib Ty elemendi RNA-lt dsDNA)
on gag ja pol
geenide homoloogid. DNA molekul inserteerub genoomi, tekitades sel viisil Ty1 elemendi
koopia genoomi teises piirkonnas. Ty moodustab viiruselaadseid partikleid.
Retroposoonid:
Äädikakärbses on kirjeldatud F, G ja I retrosposonid. Imetajatel on levinud
LINE-d (long
inserted nuclear elements, 6-7 kb)
. Retroposoonide otstes pole LTR-e, kuid nende ühes
otsas on homogeenne A:T aluspaaridest koosnev järjestus
. A:T järjestus liitub retroposooni
koostisesse pöördtranskriptisooni käigus, kuna revertaas kasutab matriitsina RNA
molekuli, mille 3’-otsa on lisatud polü-A-saba
. SINE (short inserted nuclear elements)
RNA
pealt toimub pöördtranskriptsioon L1 poolt kodeeritud ensüümi abil
(siia kuuluvad Alu ja
MIR elemendid)
.
Nt: L1 inaktiveerib geene ja põhjustab ümberkorraldusi. L1 transpositsioonist teatati 1988.
aastal, kui hemofiiliat põdeval patsiendil kirjeldati transponeeruva elemendi inserteerumist
22. kromosoomist X kromosoomi lookusesse, mis kodeerib verehüübefaktorit VIII.
Enamlevinud transponeeruvad elemendid inimese organismis:
1) LTR elemendid
– HERV (endogenous retroviruses)
– 8% genoomist; lülitunud genoomi
varem kui 25 milj aastat tagasi; inaktiivsed
2) L1 elemendid
(LINE1)
– 17% genoomist; 500 000 koopiat genoomis; 150 milj aastat
tagasi; 100 koopiat veel aktiivsed; funktsionaalsed 6 kb pikad
3) Alu elemendid
– üle miljoni koopia; 65 milj aastat tagasi; 300 bp pikkused; sisaldavad
kahte 7SL RNA geeni
(signal recognition particle)
, mis võivad mõjutada teiste geenide
avaldumist
4) SVA elemendid
– 3000 koopia; 2 kb pikkused; sisaldavad Alu-laadset ja HERV-laadset
regiooni; 25 milj aastat tagasi
16. Transponeeruvate elementide osa genoomi evolutsioonis. Tooge näiteid
TE-d võtavad enda alla märkimisväärse osa genoomist
– äädikakärbsel nt 15-20% kogu
genoomist. TE-del on oluline roll mutatsioonide tekitamisel. Pooled äädikakärbsel kirjeldatud
spontaansetest mutatsioonidest on põhjustatud mobiilsete elementide poolt.
Nt: Erinevate TE-de insertisioonid geeni
white
regiooni. Paljudel äädikakärbestel on sellesse
geeni inserteerunud TE-d, mis võivad olla väga erinevad, mis põhjustavad mutatsiooni
white
.
TE-d põhjustavad ümberkorraldusi kromosoomide struktuuris:
o HR kromosoomi erinevates osade
s paiknevate homoloogiliste elementide vahel
deletsioone
, inversioone või duplikatsioone
o Kui
2 rekombineeruvat transposooni on kromosoomis samasuunaliselt, siis nende
vahele jääv ala deleteerub
o Kui 2 rekombineeruvat transposooni on kromosoomis
vastupidises orientatsioonis,
siis nende vahele jääv ala inverteerub
(pöördub ümber)
o TE-d võivad indutseerida kromosoomidevahelisi
ümberkorraldusi nii homoloogiliste
kui ka mittehomoloogiliste kromosoomide puhul
. Tütarkromatiidide vahelise
ebavõrdse
ristsiirde tulemusena tekib 2 produkti: 1) kromosoom, milles
transposoonidevaheline ala on duplitseerunud; 2) kromosoom, millest
transposoonidevaheline ala on kaotsi läinud
o Inimesel on
Alu
järjestuste poolt põhjustatud ebavõrdse homoloogilise
rekombinatsiooniga seotud >30 päranduvat haigust ja 16 erinevat vähijuhtu
o Glycogen storage disease
– L1 elementide vahelt on deleteerunud 7,5 kb DNA-d;
metabolismihäired
o Geenide duplikatsioon ja pseudogeenide teke LINE elementide kaudu.
Retrotransposoonid võivad transponeerumisel kaasa haarata külgnevaid alasid
17. Tooge näiteid tsütoplasmaatilisest pärilikkusest ja selgitage geneetilist tagapõhja
TSÜTOPLASMAATILINE PÄRILIKKUS – pärilikkuse ülekanne, mis sõltub tsütoplasmast või
tsütoplasmas olevatest struktuuridest, mitte aga tuumageenidest.
Nimetatakse ka
kromosoomiväliseks pärilikkuseks. Näiteks
mitokondriaalne DNA mitokondrites (
nt:
nägemisnärvi kahjustus; luuüdi-pankrease sündroom)
ja plastiidne DNA taimedel (taimede
lehtede mosaiiksus nt sügisel, kui lehed värvuma hakkavad)
, mis pärandub edasi sõltumatult
tuuma kromosoomide jagunemisest.
Tsütoplasmaatilise pärilikkuse geneetiline tagapõhi:
Edasi võivad kanduda haigused, mis on seotud emapoolsete rakuorganellidega
(nt
mitokondriaalsed haigused)
.
18. Võrrelge erinevate organismide mitokondriaalset genoomi. Mitokondriaalsete
geenide funktsioonid. Mitokondrite seos vananemisega
o mtDNA genoomi suurus varieerub organismiti:
selgroogsetel 16-17 kb; õistaimedel
kuni 2500 kb
o Mitokondrite genoomi koopiaarv raku kohta on kõrge: selgroogsete somaatilistes
rakkudes <1000 koopiat; ootsüütides 10
8 mtDNA koopiat raku kohta (mtDNA
pärandub emaliini pidi!)
o Enamasti on
mtDNA molekulid tsirkulaarsed
o Pärmidel
on lisaks tsirkulaarsele vormilse ka lineaarsed vormid (replikatsiooni
vaheproduktid)
o Mitokondritel on oma replikatsiooni-, transkriptsiooni- ja translatsiooniaparaat
Mitokondriaalsete geenide funktsioonid:
Mitokondriaalsed geenid kodeerivad ribosomaalset RNAd, tRNAd ning oksüdatiivsel
fosforüülimisel osalevaid ensüüme.
Mitokondrite seos vananemisega:
Kalorite hulga vähendamine katseloomade toidus pikendab nende eluiga: väheneb
elektronide pealevoog mitokondriaalsele hingamisahelale, seoses sellega tekib ka vähem
ROS-i ja DNA kahjustusi mtDNA-s
.
Lühenes hiirte eluiga, kus oli geneetiliselt inaktiveeritud MnSOD.
Inimese katalaasi geeni üleekspressioon hiirtes pikendas hiirte eluiga, nende loomade
lihasrakkudes olevate mitokondrite puhul akumuleerus elu jooksul vähem DNA kahjustusi
.
*Saadud on ka vastuolulisi tulemusi:
ROS-i hulga liiga tugev allasurumine antioksüdantidega
ei aktiveeri katseloomadel signaalradu, mis on vajalikud pikaealisuseks
.
Mutatsioonid vähendavad mitokondrite energiatootlikkust, mis viib vananemiseni –
rakkude apoptoos
: katse hiirtega, kus mtDNA replikatsiooni läbiviival DNA polümeraasil
puudus vigu korrigeeriv aktiivsus
(mutatsioonisagedus suurenes erinevates kudedes 3-11X)
.
mtDNA mutatsioonide hulga suurenemisega ei kaasnenud ROS-i hulga suurenemist.
Mitokondrite seos vananemisega:
mitokondriaalne ROS-i kahjustus mtDNA
punktmutatsioonid mtDNA mutantne amplifikatsioon mtPTP aktiveerumine
(mitochondrial permeabilitu transition...)
raku surm
19. Mitokondris ja rakutuumas asuvate geenide koostoime. Mitokondriaalse
valgusünteesi eripära
Enamus mitokondriaalsete geenide produkte toimivad mitokondris. Neist üksi mitokondrite
elutegevuseks ei piisa:
mitokondrite ribosoomid koosnevad mitokondriaalse päritoluga
RNA-st ja rakutuuma poolt kodeeritud valkudest
.
Ribosoomivalgud sünteesitakse tsütosoolis ja transporditakse mitokondritesse. Paljud
energiatootmisega seotud polüpeptiididest on tuumavalgud. Nt ATPaasi osa subühikuid on
kodeeritud tuumageenide, osa aga mtDNA poolt
.
Tuumageenid kodeerivad ~1500 mitokondrites toimivaid valke.
Mitokondriaalse valgusünteesi eripära:
Translatsioonimehhanismid mitokondrites ja tsütosoolis on põhimõtteliselt sarnased.
Erinevus on koodonkasutuses
(nt
AUA – üldiselt Ile koodon, aga pärmi mitokondrites
määrab ta Met
; UGA – üldiselt stoppkoodon, taimedes nt hoopis Trp koodon; AGA/AGG –
üldiselt Arg, pärmil ja selgroogsetel stoppkoodon
jne).
20. Mitokondriaalsed haigused ja nende pärandumine . Inimese evolutsiooni uurimine
mtDNA järjestuste võrdluse põhjal
Mitmed inimesel kirjeldatud haigused on seotud mutatsioonidega mtDNA-s ja päranduvad
edasi emaliini pidi
. Energiatootlikkuse langus põhjustab neuronite apoptoosi ja nekroosi.
Punktmutatsioonide ja deletsioonide akumuleerumine seoses Parkinsoni, Alzheimeri,
Huntingtoni tõve ja ALS-iga
(amytrophic lateral sclerosis)
.
Mitokondriaalse haiguse avaldumismäär sõltub mutantsete mitokondrite hulgast
munarakus
(keharakkudes on tavaliselt sadu mitokondreid, neis kõigis on 2-7 mtDNA
genoomi)
.
Haigustest saaks lapsi säästa, kui asendada ema kahjustatud mitokondrid
(three-parent
embryo; vajalik mitokondri doonor)
.
Nt:
Nägemisnärvi kahjustus LHON – avaldub täiskasvanutel ja lõpeb haigete
pimedaksjäämisega; mutatsioonid mtDNA-s muudavad hingamises osalevate ensüümide
AH-list järjestust. Kui mitokondrite energiatootlikkus väheneb, siis nägeminärv kärbub.
Müopaatia – lihaste degenereerumine
(lihaste nõrkus)
. Kardiomüopaatia
Pearsoni luuüdi-pankrease sündroom – avaldub lastel ja on tavaliselt surmaga lõppev
(väheneb luuüdi rakkude arv)
. Pearsoni sündroomi põhjustab suur deletsioon mtDNA-st
Osa mitokondriaalsetest haigustest on põhjustatud mutatsioonidest tuumageenides.
Mitokondriaalse hingamisega on seotud >70 tuumageeni, mis kodeerivad valkude
subühikuid, mis osalevad protsessis otseselt. Lisaks on sajad geenid, mis reguleerivad
oksüdatiivses fosforüleerimises osaleva valkkompleksi assambleerimist, funktsioneerimist ja
degradeerimist.
Haiguste korral on eelkõige mutantsed geenid, mis on seotud mtDNA
replikatsiooni ja transkriptsiooniga
.
Nt: koensüüm Q10
(ubikinoon)
defektsus – Q10 on hingamisahela lipofiilne komponent, mis
toimib ka kui antioksüdant ja membraanide stabiliseerija.
mtDNA-s tekkinud mutatsioonide analüüs võimaldab uurida inimese evolutsiooni:
o mtDNA evolutsionerub 10X kiiremini võrreldes rakutuuma DNA-ga tänu mtDNA
kõrgele koopiaarvule
o mtDNA analüüs võimaldab rekonstrueerida ka sündmusi, mis eluslooduse
evolutsiooni seisukohalt on toimunud alles suhteliselt hiljuti
Nt:
Leiti, et Neanderthali inimene ei ole kaasaegse inimese vahetu eellane – kaasaegse
inimese ja Neanderthali inimese liinide lahknemine toimus juba 550 000 – 690 000 a tagasi
(ehk siis varem, kui kujunes kaasaegne inimene)
.
21. Selgitage geenide kloneerimise põhimõtet (kasutatavad ensüümid,
klooneerimisvektori omadused). Rekombinantset DNA-d sisaldavate kloonide
selekteerimine
Geenide molekulaarseks analüüsiks kasutatakse rekombinantse DNA tehnoloogiat: DNA
fragmentide isoleerimine genoomist ning viimine replitseeruvatesse DNA molekulidesse –
kloneerimisvektoritesse.
Saadud rekombinantset DNA-d paljundatakse rakkudes – saadakse palju koopiaid ehk
kloone
. Vastavat protseduuri nimetatakse geenide kloneerimiseks (kloonimiseks)
.
Kloneerimisel on mitmeid rakendusi:
o DNA primaarjärjestuse määramine
o Geeni(de) avaldumise regulatsiooni uurimine
o Geeniproduktide funktsioonide uurimine
o Geenitehnoloogia
Lisaks geenide kloneerimisele on võimalik DNA-d paljundada nii
in vivo
kui in vitro – PCR.
Kasutatavad ensüümid:
o Nukleaasid – ensüümid, mis lagundavad NH-s fosfodiestersidemeid
*
Eksonukleaasid – lagundavad NH ahelat kas 5’- või 3’-otsast
*
Endonukleaasid – lagundavad NH-d, lõhkudes fosfodiestersidemeid ahelasiseselt (nt
restriktsioonilised endonukleaasid ehk restrikaasid)
. Restriktaasid tunnevad ära
spetsiifilisi DNA järjestusi, lõikeproduktid on kas kleepuvate või tömpide otstega.
Restriktaas2 tunneb ära palintroomseid järjestusi ja teeb katked DNA ahelatesse
o DNA ligaas – ensüüm, mis liidab DNA ahelate otsad, katalüüsides
fosfodiestersideme moodustumist 5’-fosfaadi ja 3’-OH vahel
o Restriktaas – ensüüm, mis lõikab DNA ahelaid katki
Rekombinantse DNA isoleerimiseks ja amplifitseerimiseks peavad kloneerimisvektoril olema
järgmised omadused:
1) Võime iseseisvalt replitseeruda
(sisaldab ori piirkonda (DNA replikatsiooni
alguspunkt)
ja replikatsiooni initsiaatorvalgu geeni)
2) Sisaldab dominantset selektiivset markergeeni
(AB resistentsusgeen; nt ampillitsiini
resistentsusgeen amp
r)
3) Sisaldab unikaalseid restriktaaside lõikesaite
Rekombinantset DNA-d sisaldavate kloonide selekteerimine:
22. Tooge näiteid plasmiididel ja viirustel põhinevatest klooneerimisvektoritest ja
nende rakendusest
Kloneerimisvektoritena kasutatakse plasmiidide ja viiruste genoome.
Paljudesse vektoritesse on viidud sünteetiline DNA järjestus, mis sisaldab unikaalseid
lõikesaite erinevatele restriktaasidele – polülinker ehk multikloneerimissait
(MCS)
.
Insertsioonilise inaktivatsiooniga vektorid:
Plasmiidid viiakse bakterisse transformatsiooni teel
(baktereid töödeldakse CaCl
2
-ga, et
permeabiliseerida rakku)
. Kloneerimissaidid on viidud vektoris olevasse geeni, mille
avaldumist on lihtne testida
:
1) Ravimiresistentsusgeenid
– pBR322-s asuva tet
geeni inaktivatsioon
2) Värvusreaktsiooni kaudu
– sini-valge test; testitav lacZ
geeni inaktivatsioon pUC18 ja
pUC19 vektorites. LacZ geen kodeerib beeta-galaktosidaasi
(lagundab laktoosi Glc-iks
ja Gal-iks)
, mis on võimeline lagundama substraati X-gal, mille tulemusena tekib sinist
värvi produkt bromokloroindool. Aktiivset beeta-galaktosidaasi sisaldavate rakkude
kolooniad on X-gal-i sisaldavatel söötmetel sinist värvi; rakud, milles on see ensüüm
inaktiivne, moodustuvad X-gal-i söötmetel valged kolooniad. Valged kolooniad
sisaldavad rekombinantset DNA-d, kus võõras DNA on inserteerunud lacZ geeni
järjestusse
Inserdi mittetoimumise põhjused: plasmiidi otsad kleepuvad tagasi kokku; restriktaas pole
kõiki plasmiide katki lõiganud.
Ekspressioonivektorid:
Inserteerunud DNA-s sisalduvaid geene transkribeeritakse vektoris oleva promootori alt.
pUC18 ja pUC19 ning pBluescript vektorid sisaldavad
lac
geenide (laktoosi metabolism)
promootorit. Enamasti on transkriptsioon promootoritelt reguleeritav induktori lisamisega
bakterite kasvukeskkonda.
Promootorite või transkriptsiooni terminaatorjärjestuste testimine: võimaldavad isoleerida
DNA fragmente, mis sisaldavad promootorit või transkriptsiooni terminaatorit
(geeni
maksimaalseks ekspressiooniks kasutatakse tihtipeale ka faagide promootoreid, nt T7)
.
Jälgitakse vastavalt kas testgeeni transkriptsiooni aktivatsiooni või terminatsiooni.
Kosmiidid:
KOSMIIDID – 4-6 kb suurused plasmiidid, mis sisaldavad faag lambda
cos
järjestusi.
Lisaks sellele sisaldavad nad:
1) AB resistentsusgeeni plasmiidi sisaldavate bakterirakkude selektsiooniks
2) Järjestusi plasmiidi replikatsiooniks
3) Unikaalseid restriktsioonisaite kloneerimiseks
Cos
järjestused tuntakse ära faag lambda pakkimisaparaadi poolt (pakkimissignaal).
Lõikamisel
cos
saidist tekivad 12 bp ssDNA otsad.
Kosmiidid võimaldavad kloneerida suuri, kuni 45 kb pikkusi DNA fragmente
.
Rekombinantsed DNA molekulised pakitakse
in vitro
faagi kapsiidi. Kuna pakkimisel on
olulised ainult
cos
saidid ja DNA molekuli pikkus, pakitakse ainult rekombinantsed DNA
molekulid kindla insertsiooni pikkusega
. Nakatatud rakkudes replitseerub vektor nagu
plasmiid, selektsiooniks on ravimi resistentsusgeeni avaldumine
.
Vektorid bakteriofaagide DNA-st:
M13 – filamentne faag. Filamentsed faagid nakatavad rakke F-pilide kaudu – nakatatavad
rakud peavad olema „isased“, st. sisaldama F-plasmiidi. Filamentsete faagide genoomiks on
ssDNA, mis on kodeeriv ahel ehk + ahel. ssDNA genoomi paljundamine toimub üle dsDNA
vahevormi, mida nimetatakse replikatiivseks vormiks
(RF)
.
M13 genoomi baasil on konstrueeritud vektorid M13mp18 ja M13mp19
(dsDNA!!!)
.
Sisaldavad lisaks faagi DNA-le multikloneerimissaiti ja
lacZ
geeni järjestust. Rekombinantsete
plasmiidide teket testitakse värvusreaktsiooni kaudu
(sini-valge test; lacZ pole aktiveerunud,
siis sinine laik?)
. Faagipartiklisse pakitakse alati + ahel. Sõltuvalt fragmendi orientatsioonist
vektori replikatsiooni alguspunkti suhtes on võimalik ssDNA-na amplifitseerida valikuliselt
kas ühte või teist kloneeritud DNA ahelatest. ssDNA isoleeritakse faagipartiklitest, dsDNA
(RF)
isoleerimine toimub aga rakkudest.
Fagemiidid:
Fagemiidide puhul on võimalik eraldada nii dsDNA-d kui ka ssDNA-d
(võimalik paljundada
nii plasmiidi kui ka faagi analoogina)
.
Fagemiidid sisaldavad:
1) Plasmiidi replikatsiooni initsiatsiooniks vajalikke järjestusi
2) Selektsioonimarkerit
3) Filamentse faagi genoomi
Rakkude nakatamisel sobiva helperfaagiga toimub ümberlülitumine plasmiidi
replikatsioonilt ssDNA replikatsioonile
. ssDNA-d eraldatakse faagipartiklitest. ssDNA-l on
mitu kasutust:
1) DNA nukleotiidse järjestuse määramine
2) DNA koht-suunatud mutagenees
3) Ahela-spetsiifiliste DNA proovide valmistamine NH-de hübridisatsiooniks
Faag lambda genoomi kasutamine kloneerimisvektorina:
Faag lambda genoomil põhinevad vektorid võimaldavad kloneerida suuri DNA fragmente.
Olemas on süsteemid, mis takistavad mitterekombinantide paljunemist.
Infektsioonivõimelised on ainult rekombinantset DNA-d sisaldavad partiklid, mis sisaldavad
~15 kb võõrast DNA-d.
o Kasutatakse DNA raamatukogude konstrueerimiseks
: rekombinantsed molekulid
sisaldavad erinevaid DNA fragmente, mis katavad kogu genoomi
o Rekombinantseid DNA molekule sisaldavaid faagipartikleid sisaldavat lüsaati
säilitatakse 4⁰C juures 0,3% kloroformi juuresolekul
Bakteris ja imetaja rakkudes replitseeruv vektor:
sacB
– levaan sukraas, katalüüsib fruktoosi rühma ülekannet erinevatele suhkrutele, letaalne
kui sööde sisaldab 5% sahharoosi.
Võimaldab selekteerida inserdiga vektorit (DNA lõik
inserteerunud BamHI
saiti). sacB-sse viiakse replitseeruv faktor; plasmiid peab sisaldama
eukarüootse viiruse genoomi
(genoomi osa)
.
Kunstlikud kromosoomid:
Plasmiidide või viiruste baasil konstrueeritud vektorid ei võimalda kloneerida üle 45 kb
pikkuseid DNA fragmente. Suuremate DNA segmentide kloneerimiseks kasutatakse pärmi
kunstlikke kromosoome YAC-e
(yeast artificial chromosomes)
, mis lasevad kloneerida
200-500 kb suuruseid fragmente.
YAC-id sisaldavad:
1) Pärmi replikatsiooni
ori
piirkonda (ARS – autonomously replivating sequence)
2) Pärmi tsentromeeri
3) Kahte pärmikromosoomi telomeeri molekuli mõlemas otsas
4) Selektiivset markerit – tavaliselt metsiktüüpi geen, mis taastab peremeesraku
prototroofsuse
5) Multiklorneerimissaiti
BAC – bacterial artificial chromosome.
BAC-id on konstrueeritud F-plasmiidi baasil ja on
YAC-idega võrreldes vähem komplekssed
. Replitseeruvad E.coli
rakkudes sarnaselt
plasmiididega.
23. DNA raamatukogude konstrueerimine. Võrrelge genoomset ja cDNA raamatukogu
DNA raamatukogude konstrueerimine ja analüüs:
1) Genoomse DNA raamatukogu
– sisaldab erinevaid DNA kloone, mis katavad
summaarselt kogu genoomi
Kasutatakse restriktaasi, mis tekitab kleepuvaid otsi
. Tömpide otstega DNA fragmentide
kloneerimine:
kleepuvate otste lisamine DNA fragmentidele ja vektorile toimub
terminaalse transferaasi abil. Vektorit ja kloneeritavaid DNA fragmente töödeldakse
esmalt faag lambda eksonukleaasiga
(DNA „süüakse“ ahelate 5’-otstest lühemaks)
. Seejärel
lisatakse 3’-üksikahelalistele otstele terminaalse transferaasi abil nukleotiide – vektori
DNA-le lisatakse polü-A sabad ja genoomse DNA fragmentidele polü-T sabad. Järgnevalt
liidetakse vektori DNA genoomse DNA-ga.
2) cDNA raamatukogu
– tahetakse saada ainult valku kodeerivaid järjestusi; rakkudest
isoleeritud mRNA-lt sünteesitakse pöördtranskriptaasiga komplementaarne DNA
ahel cDNA
(complementary DNA)
ja kloneeritakse. cDNA raamatukogu sisaldab
ainult ekspresseeruvate geenide kodeerivaid järjestusi
Kõrgemate loomade ja taimede genoomis on palju DNA järjestusi, mis on geneetiliselt
inaktiivsed.
Funktsionaalsete DNA järjestuste kloneerimiseks on otstarbekas sünteesida
rakus eraldatud mRNA-le komplementaarne DNA – cDNA
. cDNA sünteesil kasutatakse
praimerina polü-T oligomeere.
Pöördtranskriptaas ehk revertaas sünteesib mRNA-le
komplementaarse DNA ahela. Ribonukleaas H degradeerib RNA ahela. RNA degradatsioonil
tekkinud lühikesed fragmendid on praimeriteks DNA sünteesil. DNA teise ahela sünteesib
DNA pol I, mis ühtlasi kõrvaldab molekulist ka RNA praimerid, asendades RNA järjestuse DNA
järjestusega. DNA ligaas liidab DNA ahelate otsad. Kaksikahelalised cDNA molekulid
inserteeritakse vektorisse eelpool kirjeldatud strateegia alusel
.
DNA raamatukoguks vajalik rekombinantsete kloonide arv N: N sõltub genoomi suurusest.
Eeldades, et kõigil DNA fragmentidel on võrdne võimalus saada inserteeritud
kloneerimisvektorisse, saame arvutada kloonide hulga, mis on vajalik, et antud tõenäosusega
P isoleerida meid huvitav rekombinantse DNA kloon.
N =
ln(1−f)
ln(1−P ) f – näitab, kui suure fraktsiooni genoomist võtab enda alla keskmise pikkusega
DNA fragment DNA raamatukogus.
24. Selgitage, mis eesmärgil rakendatakse
Southern
, Nothern ja Western blot meetodit
o Southern blot:
kasutatakse DNA analüüsiks
On teada, et bakteri genoom (kromosoom või plasmiid) sisaldab mingit meid huvitavat geeni,
kuid ei teata, kus täpselt see genoomis asub. Selleks
lõigatakse DNA restriktaasiga ning
teostatakse geelelektroforees. DNA fragmendid lahutatakse geelelktroforeesil ning
voolutatakse geelist nitrotselluloosfiltrile. Geelile asetatakse nitrotselluloosfilter ja selle otsa
virn filterpabereid, mis imavad vedelikku üles, tõmmates sellega DNA geelist filtrile. Järgneb
filtrile kandunud DNA hübridisatsioon radioaktiivse prooviga, mis on komplementaarne
huvipakkuva geeniga või muu uuritava DNA piirkonnaga. Kui DNA on homoloogiline, siis
toimub hübridiseerumine proovis oleva DNA-ga ning UV-valguses muutub piirkond
nähtavaks
(on võimalik lokaliseerida meid huvitav piirkond genoomis)
.
Et hinnata meid huvitava DNA fragmendi suurust, kasutatakse geelelektroforeesil alati
suurusmarkerit.
Meditsiinis: kasutatakse nt tsüstilise fibroosi molekulaarses diagnostikas!
o Northern blot:
kasutatakse RNA analüüsiks
Meetod põhineb
RNA-de lahutamisel geelelektroforeesil ning nende hübridiseerimisel
radioaktiivse prooviga filtritel
. Meetod on oluline selgitamaks, millistes kudedes/rakkudes
meid huvitav geen ekspresseerub. Võimalik on kindlaks teha mRNA produkti suurus. Kui
DNA fragmentide lahutamine toimub neutraalse pH juures
(DNA on kaheahelaline)
, siis
kuna RNA on üheahelaline, kasutatakse denatureerivat keskkonda
(ei lase RNA-l
kaheahelalisi struktuure tekitada)
.
o Western blot:
kasutatakse valkude analüüsiks
Valgud kantakse geelilt membraanile. Filtrit töödeltakse lahusega, mis sisaldab analüüsitava
valgu vastaseid antikehi. Valguga seondunud antikeha lokaliseeritakse sekundaarse
antikehaga, mida on võimalik tuua nähtavaks nt ensümaatilise reaktsiooniga
. Uuritakse
seda, kas vastav geen valgu tasemel avaldub
(Kas ja millistes kogustes on võimalik valku
detekteerida? Millise suurusega valgud tekivad?)
.
Nt: glutamiini süntetaasi kloroplasti vormi tuvastamine taime lehtedest ja juurtest.
25. DNA nukleotiidse järjestuse määramine Sangeri meetodil ja selle edasiarendus
tänapäeval
DNA sekveneerimise ensümaatiline meetod
(Sanger et al)
– põhineb in vitro DNA sünteesil,
mis katkeb dideoksünukleotiidi lülitumise tagajärjel DNA ahelasse. DNA ahela
pikendamiseks on vaja vaba 3’-OH rühma, dideoksünukleotiidil
(ddNTP)
on 3’-OH asemel
3’-H
.
Reaktsioonisegusse lisatakse:
o DNA polümeraas
(tavaliselt faagi T7 polümeraasi modifitseeritud variant, mis peab
hästi vastu
in vitro sünteesil)
o 4 erinevat deoksüribonukleotiidi
o Radioaktiivselt märgitud
(või fluorestsentsmärkega)
deoksüribonukleotiid
o 4 spetsiifilist terminaatornukleotiidi
Reaktisooni tulemusena tekivad radioaktiivselt märgistatud DNA fragmendid, mis algavad
kõik samast kohast, kuid nende süntees on statistiliselt termineerunud erinevatest
kohtadest. Reaktsioonisegud denatureeritakse, et vabastada sünteesitud DNA fragmendid
matriitsahelalt ja fragmendid lahutatakse polüakrüülamiidgeelis. DNA fragmendid eristuvad
geelis üksteisest ühenukleotiidse täpsusega
.
DNA fragmendid tuuakse nähtavale
(kuna fragmendid piisavalt väikesed, siis
etiidiumbromiid ei värvi fragmente!)
:
1) Geelist autoradiograafia tegemisel
2) Spetsiaalse aparatuuri abil, mis detekteerib radioaktiivset fosforit või fluorestsentsi
*Selline meetod võimaldab määrata 300-400 nukleotiidi korraga (parematel juhtudel
500-600 nt).
Tänapäeval on DNA sekveneerimine automatiseeritud.
Reaktsiooniproduktid lahutatakse
üksteisest kapillaaris. Reaktsiooniproduktid on märgistatud 4 erineva fluorestsentsvärviga.
*96 kapillaariga masinad võimaldavad sekveneerida ~100 000 nt masina kohta päevas.
26. DNA lõikude paljundamine PCR meetodil (meetodi põhimõte ja rakendusalad)
Vaja on teada primaarjärjestust paljundatava DNA piirkonna mõlemas otsas – praimerite
järjestused
.
PCR toimub 3-etapiliselt:
1) Paljundatava DNA denatureerimine temperatuuril 92⁰C-95⁰C
2) DNA renatureerimine temperatuuri langetamisel
(~50⁰C)
ülehulgas lisatud
sünteetiliste praimerite juuresolekul, millest üks on komplementaarne
amplifitseeritava DNA ühele ahelale, teine selle vastasahelale – praimerid
hübridiseeruvad DNA ahelatega
3) DNA replikatsioon ahelatele kinnitunud praimerite 3’-OH otstest
(temp tõstetakse
72⁰C juurde)
Tsüklit korratakse vähemalt 20X.
DNA sünteesil kasutatakse kõrget temp-i taluvaid DNA polümeraase
. Nt: Taq polümeraas –
ensüümil puudub vigu korrigeeriv
(proofreading)
aktiivsus; Pfu polümeraas – omab 3’-5’ vigu
korrigeerivat aktiivsust jne;
Tfl
polümeraas – pikkade (kuni 35 kb) DNA lõikude paljundamine.
Rakendusalad:
PCR meetod on oluliselt lihtsustanud DNA kloneerimist ja DNA sekveneerimist, kuna
algmaterjaliks piisab väga väikesest DNA hulgast
. Rakendus diagnostikas ja isikute
tuvastamises, kuna võimaldab analüüsiks DNA-d amplifitseerida väga väikesest
koematerjali hulgast; evolutsioonis; DNA sõrmejäljed
(kriminalistika)
.
Järgmise põlvkonna sekveneerimismeetodid:
Pürosekveneerimine
– fragmenteeritud ja adapteritega liidetud DNA püütakse kerakestele.
Emulsioonis toimub PCR; DNA viiakse pikotiiterplaatide kannudesse, kus on ensüümid ja
praimerid. Lisatakse ühekaupa nukleotiide, nukleotiidi lülituse korral eraldub pürofosfaat,
mida detekteeritakse lutsiferaasi aktiivsuse kaudu
.
27. Kromosoomide geneetiline ja füüsiline kaart: selgitage geenide positsioonilise
kloonimise põhimõtet
Geneetilised kaardid
koostatakse lähtuvalt rekombinatsioonisagedusest. Kokkuleppeliselt
võetakse
ühikuks 1 sentimorgan (1cM), mis vastab geenidevahelisele kaugusele ühes
põlvkonnas, mille piires on rekombinatsiooni sagedus 0,01 e. 1%.
Geenide molekulaarne
kaugus üksteisest ei pruugi kaugeltki kokku langeda nende geneetilise kaugusega, kuna mitte
kõik genoomi DNA piirkonnad pole rekombinatsiooniliselt võrdväärsed.
Füüsilisel kaardil
, milleks on tavaliselt restriktsioonikaardid, on kauguste mõõtmise
mõõdupuuks aluspaarid, kilobaasid, megabaasid või gigabaasid
. Füüsilisel kaardil eristuvad
iseloomulikud unikaalsed nukleotiidijärjestused, mida nimetatakse järjestusspetsiifilisteks
saitieks
(STS). Neid lühikesi unikaalseid järjestusi nimetatakse ankurjärjestusteks, sest nende
kaudu ankurdatakse füüsilised kaardid geneetiliste kaartidega ja
vice versa.
Seega on
ankurmarkerid kaardistatud nii geneetiliselt kui ka füüsiliselt kaartidel
. Teiseks
ankurmarkerite tüübiks on avaldunud järjestuste märgised e. EST-id. Need on mRNA-st
pärinevad lühikesed cDNA koopiad, mida saab kasutada avalduvate geenide lokalisatsiooni
määramiseks.
Geenide identifitseerimiseks ja isoleerimiseks tuleb määrata nende asukoht
genoomis,
täpsemalt
kromosoomides. Geeni asukoha kindlaksmääramisel kromosoomis lähtutakse
tänapäeval
molekulaarsetest markeritest, milleks on tavaliselt spetsiifiline DNA-järjestus.
Molekulaarse markeri ülesleidmisele eelneb aga rida katseid.
Tavaliselt kaardistatakse geene
teatud kromosoomi piirkonna ristamiskatsete ja
geneetilise analüüsi alusel või lähtuvalt
inimese sugupuu analüüsist. Seejärel seostatakse uuritava geeni asukoht kindla molekulaarse
markeriga füüsilisel kaardil
. Selle leidmiseks on vaja üldjuhul isoleerida mutant, sest just
muutust saab seostada huvipakkuva geeniga. Edasi on vaja nii metsiktüüpi isendist kui ka
mutandist isoleerida DNA-segment, mis sisaldab huvipakkuva geeniga seonduvat
molekulaatset markerit, ning need sekveneerida
. Positsioonilisel kloonimisel saab isoleerida
geeni, mille asukoht on teada, kui mille funktsioon vajab selgitamist
.
Kui mutatsioon DNA-s on tekkinud restriktaasi lõikesaiti, siis seda järjestust restriktaas enam
ei lõika. Mutatsioonid võivad muuta DNA järjestust ka nii, et tekivad uued restriktaaside
lõikesaidid. Seega põhjustavad mutatsioonid DNA restriktsioonifragmentide mustris
erinevusi, kuna võrreldes algse DNA-ga tekib mutatsioone sisaldava DNA lõikamisel mingi
kindla restriktaasiga teistsuguse pikkusega fragmente. Nii võib erinevatest isolaatidest
pärinev DNA olla DNA restriktsioonanalüüsi põhjal restriktsioonisaitide suhtes polümorfne –
erinevate isolaatide puhul tekivad erineva pikkusega restriktsioonifragmendid.
Restriktsioonifragmentide pikkuse polümorfismi RFLP-d on võimalik detekteerida näiteks
“Southern blot” analüüsil. DNA restrikteeritakse, fragmendid lahutatakse geelelektroforeesil,
kantakse filtrile ja hübridiseeritakse spetsiifilise radioaktiivse DNA prooviga. RFLP-d
kasutatakse geneetilise markerina kromosoomide kaardistamisel. Samuti võimaldab RFLP
analüüs hinnata, kas ristamisel saadud järglased on vanemtüüpi või rekombinantsed. Juhul,
kui on tegemist rekombinantidega, saab rekombinatsioonisageduse põhjal arvutada RFLP-de
geneetilist distantsi. Sellist analüüsi on rakendatud näiteks taime
Arabidopsis
geneetilistes
katsetes.
RFLP markereid kasutatakse ka inimese kromosoomide kaardistamisel. Sel juhul on
vaadeldud näiteks markerite segregeerumist suguvõsas. Markerite aheldumise alusel on
koostatud kaardid. Nii esitati 1992. aastal esimene inimese kromosoomide RFLP-de kaart,
mis baseerus 2000-l RFLP-l.
Kui huvipakkuva geeni lähedal
(mille mutatsioon põhjustab näiteks mingit haigust)
on
identifitseeritud RFLP marker, on sellest markerist vaja liikuda edasi huvipakkuva geenini.
Kromosoomil “jalutamine”
: RFLP markerit sisaldava genoomse DNA raamatukogu klooni
baasil valmistatakse hübridisatsiooniproov ja identifitseeritakse sellega kloonid, mis
sisaldavad radioaktiivselt märgitud DNA-ga kattuvaid järjestusi. Seejärel koostatakse
hübrisiseeruvate kloonide restriktsioonikaart ning valmistatakse järgmine
hübridisatsiooniproov restriktsiooni-fragmendist, mille järjestus asub algse proovi
järjestusest kõige kaugemal. Identifitseeritakse uue DNA proovi järjestustega kattuvaid
järjestusi sisaldavad kloonid ja nii võidakse protseduuri korrata mitu korda, enne kui jõutakse
välja huvipakkuva geeni järjestuseni. Seda, et kätte on saadud huvipakkuva geeni järjestus,
on võimalik kontrollida mitmel viisil. Näiteks äädikakärbse puhul on näidatud metsiktüüpi
alleeli sisaldava DNA järjestuse viimisel mutanti metsiktüüpi fenotüübi taastumist. Inimese
geenide puhul analüüsitakse metsiktüüpi alleeli ja mutantsete alleelide nukleotiidset
järjestust ning näidatakse, et mutantsed geenid ei kodeeri funktsionaalset geeniprodukti.
Juhul, kui huvipakkuva geeni ja molekulaarse markeri vahemaa on suur, kasutatakse
kromosoomil “hüppamise”
meetodit. Iga hüpe võimaldab edasi liikuda veidi üle 100 kb
korraga. Stardipunktina lähtutakse jälle kloonist, mis sisaldab RFLP markerit ja mis isoleeriti
hübridisatsiooni tulemusena. DNA-le tehakse restriktaasiga osaline lõikus, mille tulemusena
saadakse suured DNA fragmendid. Fragmendi ühes otsas on algse prooviga hübridiseeruv
DNA ja teises otsas unikaalne järjestus. Fragmendi otsad ligeeritakse, tekib liitjärjestus.
Järgmise hübridisatsiooniproovi valmistamisel lähtutakse liitjärjestusest, märkides
radioaktiivselt unikaalsed DNA järjestused. Kuna rõngasmolekulid tekitatakse suurtest
restriktsioonifragmentidest, võimaldab liitjärjestuste kasutamine hübridisatsiooniproovide
valmistamiseks kiiremini huvipakkuva geenini välja jõuda.
28. Genoomide sekveneerimisprojektid: mida oleme teada saanud bakterite
genoomide sekveneerimisest? Minimaalse genoomi projekt
o Bakteritest on genoom sekveneeritud ~44 000-l
(1. märts 2015 seisuga)
o Mida stabiilsem on bakteri elukeskkond, seda väiksem on genoom
o Genoomi suurus varieerub suuresti
: al ~580 kb – 9100 kb
o Mida parasiitsem on bakter, seda väiksem on genoom, sest on võimalik kasutada
peremehe elutegevuse saadusi
o Genoomi suurus varieerub ka liigisiseselt
(nt: E.coli laboritüvi K12 ~4,6 Mb,
mittepatogeenne; E.coli O157 ~5,5 Mb, põhjustab verist kõhulahtisust)
o Rickettsia prowasekii
– tüüfusetekitaja, levib täide abil; genoom 1,1 Mb. Kõige
lähedasem mitokondrite sugulane
; puuduvad anaeroobse glükolüüsi geenid ja
geenid, mis on vajalikud AH-de ja nt-de sünteesiks; sisaldab ATP sünteesil osalevaid
geene; palju pseudogeene
o Pelagibacter ubique
– seni teadaolevalt kõige väiksema genoomiga väliskeskkonnas
elav bakter
; genoom ~1,4 Mb, 1354 geeni. Elab merevees, kus on väga vähe
toitaineid ja stabiilne keskkond
o Mükoplasma
– inimese genitaalide ja hingamisteede epiteelrakkude parasiit. Seni
kõige väiksema genoomiga rakuväliselt elav bakter
; genoom ~580 kb
Minimaalse genoomi projekt:
Kui palju on vaja geene, et tagada raku elutegevus?
Otsitakse kõige väiksema elu
võimaldava genoomiga organismi.
1) Võrdlev genoomika eesmärgiga leida ühisosa geenidest erinevatel bakteritel
o Probleemid:
bakterite erinev elukeskkond; samu fn-e võivad tagada
mittesuguluses olevad geenid
2) Oluliste geenide väljaselgitamine eksperimentaalselt
o Transposoonmutagenees
o Geeniekspressiooni inhibeerimine antisense RNA abil
o Süstemaatiline geenide inaktiveerimine
Võimalikud rakendused
: uute ja võimendatud omadustega bakterirakkude konstrueerimine.
o Biomeditsiin
(vähendatud genoomiga mükoplasmade kasutamine vektorina haiguste
ravis)
o Bioremediatsioon
o Bioenergeetika
(tsüanobakterid – H
2
produktsioon, CO2 kõrvaldamine)
29. Inimese genoomi uuringud: nende uuringute ajalugu ja tänapäev. Mida on teada
inimese genoomist?
Inimese genoomi projekt
(HGP) k
äivitus aastal 1990.
Eesmärk:
1) Kaardistada kõik inimese geenid
2) Koostada inimese genoomi füüsiline kaart
3) Määrata
(aastaks 2005)
inimese kromosoomide nukleotiidne järjestus
HGP-s osalevad
laborid sekveneerisid eelnevalt kaardistatud genoomseid kloone. Celeras
viidi kogu genoom väikesteks DNA fragmentideks, sekveneerides juhuslike fragmentide
otsi ning arvutianalüüsi abil ühedati kattuvad järjestused terviklikuks DNA järjestuseks
.
Selle strateegia rakendamisel tekitab
probleeme DNA kordusjärjestuste esinemine
eukarüootses DNA-s.
Võrreldes HGP projektiga võeti Celeras kasutusele uut tüüpi, võimsamad
automaatsekvenaatorid – ööpäevaga saadi sekveneerida kokku 100 miljonit nukleotiidi.
Leiutati meetod üle genoomi juhuslike fragmentide sekveneerimiseks: kogu genoom
fragmenteeritaks, igal fragmendil on ühisosa mõne teisega.
Inimese genoomi sekvenerimise edusammud:
o Konkurents kiirendas HGP tegevust
o Kogu genoomi DNA järjestuse mustandvariandi valmissaamisest teatati 2001.
aastal mõlema konkureeriva projekti poolt
o Saadud andmete põhjal
leiti, et inimesel on vähem geene kui algul arvati – ~25 000
piires
o Valke kodeerivaid järjestusi alla 2% genoomist; vähemalt 50% geenidest toimub
mRNA alternatiivne splaissing
o Geenide paiknemine üle genoomi on ebaühtlane: GC-rikastes piirkondades rohkem
geene ja lühemad intronid; AT-rikastes alades vähem geene ja pikemad intronid
o 2003
– DNA kaksikheeliksi kirjeldamise 50ndaks aastapäevaks esitas HGP inimese
genoomi nukleotiidse järjestuse, kus 98% geene sisaldavatest aladest oli
sekveneeritud 99,99%-lise täpsusega
Inimese genoomi info:
o Inimese genoomis on palju kordusjärjestusi:
transposoonid
o Genoom 3 Gb, millest eukromatiin 2,8 Gb
o SNiP-id; haplotüübid
o CNV
(muutus koopiaarvudes)
– hõlmavad ~12% genoomist; enamasti segmentaalsed
duplikatsioonid
Genoomi piirkonnad, mille nukleotiidne järjestus ei ole täielik:
1) Tsentromeeride ala
, mis sisaldab kõrgeltkorduvaid järjestusi
2) Kromosoomide otsad
(kõrgeltkorduv ala telomeeride piirkonnas)
3) Multigeensed lookused
(sama geeni peaaegu identsed koopiad)
30. Metagenoomika. Inimese mikrobioomi projekt
METAGENOOMIKA – kõigi antud keskkonnas elavate mikroobide genoomi
(mikrobioomi)
uurimine
. Võib pärineda nt mereveest, inimeste soolestikust, mullast jne.
1) Funktsionaalne metagenoomika
– analüüsib metagenoomi raamatukogus esindatud
geene seoses bakterite fenotüübiga
(nt soolatolerantsus, ensüümiaktiivsus)
ja
määrab nende geenide päritolu
2) Järjestusepõhine analüüs
– raamatukogus esindatud 16S rRNA geenide analüüs
eesmärgiga selgitada mikroobipopulatsiooni liigiline koosseis uuritavas keskkonnas
Inimese mikrobioomi projekt:
o 5-aastane projekt,
alustati 2008. aastal
o Inimese tervis sõltub mikroobikoosluse, peremehe ja keskkonna vahelistest
interaktsioonidest. Mikroobid mõjutavad inimese immuunsüsteemi, seedimist,
vitamiinide tootmist jne
o Projekti
eesmärgiks on mikroobikoosluse kirjeldamine ja sekveneerimine
inimorganismis
; leida seoseid mikoobikoosluste muutuste ja tervisliku seisundi
vahel
o 600 bakteri isolaadi sekveneerimine
o Uuritavad
mikroorganismid pärinevad suust, nahalt jne
o Leiti, et inimese mikrobioomis on 8 milj unikaalset geeni, mis on inimese
elutegevuseks vajalikud; inimese mikrobioom muutub elu jooksul
(haigused,
toitumine, AB-d)
31. Funktsionaalne genoomika (eesmärgid ja põhimeetodid)
FUNKTSIONAALNE GENOOMIKA – uurib RNA ja valkude ekspressiooni ning funktsioone
Genoomi primaarjärjestuse teadasaamine aitab oluliselt kaasa genoomi funktsioonide
uurimisele. Mis funktsioon on DNA järjestusel, selgub geneetilistest katsetest nende
mutantidega
.
Teades genoomi terviklikku järjestust, on võimalik uurida, millal ja kus erinevad geenid
avalduvad
:
o Üle genoomi transkriptsiooni uurimine
geenikiipide abil
Selleks, et uurida kogu genoomi või paljude geenide avaldumist,
seotakse kandjale
(nailonmembraanile või klaasile) fikseeritud asukohtadesse DNA lõigud, mis pärinevad
organismi erinevatest geenidest
. Saadakse geenikiibid, millele hübridiseeritakse rakkudest
eraldatud totaalselt mRNA-lt sünteesitud cDNA, mis on märgistatud
(tavaliselt
fluorestseeruva märgisega)
. Hübridisatsioonisignaale analüüsitakse arvutis. Signaali tugevus
viitab geeni avaldumismäärale rakkudes antud arenguetapil, koes või keskkonnatingimustel
.
Transkriptoomi analüüs RNA-seq meetodil: võrreldes mikrokiipidega on taustamüra väiksem.
Sekveneeritakse lühikesi DNA fragmente, mille külge on viidud adapterid
. Saadakse
kvantitatiivsed tulemused eksonite kohta ja saab analüüsida
(SNP)
infot eksonite ühenduse
kohta.
o Valkude ekspressiooni uurimine, analüüsides 2D geele
– proteoomika
PROTEOOMIKA – geeniekspressiooni uurimine valkude hulga määramise teel.
Valkude hulga hindamiseks kasutatakse totaalvalgu lahutamist kahedimensionaalsel (2-D)
polüakrüülamiid-geelelektroforeesil.
Igale valgule vastab geelis kindel koht. Mida suuremal
hulgal on teatud valku rakus, seda paremini on teda geelis näha
(tugevam signaal)
.
Mass-spektromeetria võimaldab määrata valgu järjestusi ka väga väikeste valgukoguste
korral.
o Metaboolne modelleerimine
32. Operoni mudel. Geenide positiivne ja negatiivne regulatsioon bakterites
o Bakterite geenid paiknevad regulatoorsetes üksustes, mida nim OPERONIDEKS.
o Operoni mudel loodi
1961. aastal Francois Jakob ja Jacques Monod poolt
o Operoni mudeli põhjal toimub
ühe või mitme järjestikku paikneva struktuurgeeni
(kodeerivad ensüümide või struktuurvalkude järjestusi)
transkriptsioon kahe
kontrolliva elemendi kaudu:
1) Regulaator- või repressorgeen, mis kodeerib repressorvalku
2) Teatud tingimustel on repressor seondunud teise elemendiga –
operaatoriga
o Promootor paikneb struktuurgeenidest eespool
(5´suunas). Operaatorala jääb
promootorist geenide poole
. Kui repressor on seondunud operaatoralaga, takistab
see RNA polümeraasi poolt läbiviidavat transkriptsiooni
Seda, kas repressorvalk seondub operaatoriga ja pärsib struktuurgeenide transkriptsiooni,
määrab
efektormolekulide olemasolu rakus. Efektormolekulideks on väikesed molekulid,
näiteks aminohapped või erinevad suhkrumolekulid. Indutseeritava operoni puhul
nimetatakse efektormolekuli
induktoriks, represseeritavate operonide puhul aga
korepressoriks
. Efektormolekul seondub repressoriga, muutes tema konformatsiooni ja selle
tagajärjel muutub valgu aktiivsus. Indutseeritavaid ja represseeritavaid operone eristab see,
kas repressori ja efektori kompleks on võimeline seonduma operaatorile või mitte:
1.
Indutseeritava operoni puhul on geenide transkriptsioon pärsitud seetõttu, et
induktoriga mitteseondunud repressor on seondunud operaatorile
. Efektormolekuli
seondumine repressoriga muudab selle konformatsiooni nii, et repressor vabaneb
operaatoralalt, võimaldades geenide transkriptsiooni. Seega on induktori olemasolul
rakkudes vastavate geenide transkriptsioon indutseeritud, sest repressori ja induktori
kompleks ei ole võimeline DNA-ga seonduma.
2.
Represseeritavate operonide puhul on vastupidi – vaba repressor ei ole võimeline
seonduma operaatoriga. Operaatoralale seondub ainult repressori ja efektori
(korepressori) kompleks. Seega on represseeritavas operonis paiknevate struktuurgeenide
transkriptsioon rakkudes efektormolekuli olemasolul pärsitud
.
Kui operonis sisaldub rohkem kui üks struktuurgeen, sünteesitakse mRNA molekul, mis
sisaldab informatsiooni kõigi nende geenide kohta
. Samas operonis paiknevad
struktuurgeenid on koordineeritult reguleeritud ja koos transkribeeritavad. Näiteks
trüptofaani operon sisaldab viit erinevat geeni trüptofaani biosünteesiks ja nende
transkriptsioon rakus on represseeritud trüptofaani kaudu.
Geenide negatiivne ja positiivne kontroll:
Kuna repressor pärsib struktuurgeenide transkriptsiooni, nimetatakse repressori poolt
vahendatud kontrollmehhanismi negatiivseks kontrollmehhanismiks. Lisaks negatiivsele
esineb ka positiivne kontrollmehhanism, kus regulaatorvalku on vaja geenide
transkriptsiooni aktivatsiooniks. Sellist regulaatorvalku nimetatakse transkriptsiooni
aktivaatoriks.
Transkriptsiooni aktivaatorite ja repressorite seondumisalad on erinevad:
o Repressorite seondumisalad jäävad enamasti promootorist allapoole või kattuvad
-10 elemendiga
o Aktivaatorite seondumisalad jäävad enamasti promootorist ülespoole või kattuvad
-35 elemendiga
33. Transkriptsiooni regulatsioon
E. coli
laktoosi operoni näitel. Millistel tingimustel
avalduvad laktoosi operoni geenid?
Laktoosi operoni regulatsioon:
Laktoosi operon sisaldab promootorit P, operaatorit O ja kolme struktuurgeeni lacZ
(beeta-galaktosidaas)
, lacY (beeta-galaktosidaasi permeeas; võimaldab suhkru transporti
rakku)
ja lacA (beeta-galaktosiidi transatsetülaas).
Laktoosi operoni puhul esineb nii negatiivne kui positiivne regulatsioon:
1) Negatiivne
– transkriptsiooni pärsib LacI repressor
Geen
lacZ kodeerib beeta-galaktosidaasi, millel on kaks funktsiooni:
o Lõikab laktoosi molekuli kaheks monosahhariidiks
, Glc-iks ja Gal-iks
o Konverteerib laktoosi operoni induktoriks allolaktoosiks
o Mutatsioonid
lacI
geenis võimaldasid lac operoni konstitutiivset transkriptsiooni:
trans-toime
(toime kaugelt
; seonduvad ühendid)
o lac
operaatori O mõju avaldub ainult cis-toimelisena (piirkond DNA-l), kuna
repressori seondumine operaatoralale peab füüsiliselt takistama struktuurgeenide
transkriptsiooni
. Mutatsioonid operaatoralas takistavad repressori seondumist –
operoni geene transkribeeritakse pidevalt, kui mutantne operaator paikneb geenide
suhtes
cis
-asendis ja funktsionaalne operaator trans-asendis
2) Positiivne
– transkriptsiooni aktiveerib CAP (CRP) aktivaatorvalk
o Selleks, et toimuks transkriptsiooni initsiatsioon
lac
promootorilt, on vaja, et
promootoralale seonduks aktivaatorvalk CAP
(catabolite activator protein
), mida
sageli nimetatakse ka CRP-ks (
cyclic AMP retceptor protein
)
o CAP valk aktiveerib transkriptsiooni
lac
promootorilt ainult siis, kui ta on
kompleksis cAMP-ga
CRP
(CAP)
valk on globaalne regulaator – kontrollib nii üksikute operonide (laktoosi ja
sorbitooli operonid)
kui ka regulonide (galaktoosi, glütseroolfosfaadi, maltoose regulonid)
transkriptsiooni. Kui bakterite kasvukeskkonnas on lisaks laktoosile ka glükoosi, on glükoosi
kasutamine eelistatud ning
lac
operoni geenide transkriptsioon pole aktiveeritud. Sellist
nähtust nimetatakse katabliitseks repressiooniks.
Kataboliitne repressioon
:
Nähtust, kus glükoosi juuresolekul kasvukeskkonnas eelistatakse teistele substraatidele
glükoosi, nimetatakse
glükoosi efektiks e. kataboliitseks repressiooniks. Lac
operoni ning ka
mitmete teiste operonide kataboliitne repressioon on vahendatud aktivaatorvalgu CAP (CRP)
ja tema efektormolekuli tsüklilise AMP (cAMP) poolt. Selleks, et toimuks transkriptsiooni
initsiatsioon
lac
promootorilt, peab promootoralaga seonduma CAP-cAMP kompleks.
Aktivaatorvalk seondub promootoralale dimeerina ning interakteerub otseselt RNA
polümeraasi a-subühikuga, soodustades sel viisil RNA polümeraasi seondumist promootorile
ning transkriptsiooni initsiatsiooni.
Rakusisene cAMP kontsentratsioon on tundlik glükoosi koguse suhtes. Kui rakus on kõrge
glükoosi kontsentratsioon, langeb cAMP tase järsult. cAMP madala kontsentratsiooni tõttu
pole aktivaator CAP võimeline seonduma
lac
operoni promootorile ning RNA polümeraas ei
ole võimeline transkriptsiooni initsieerima. Glükoosi olemasolul rakkudes jääb
lac
operoni
transkriptsiooni tase alla 2% tema aktiveeritud tasemest, mis ilmneb siis, kui bakterite
kasvukeskkonnas on laktoos, kuid pole glükoosi. Kataboliitne repressioon toimib sama
skeemi järgi ka arabinoosi (
ara
) ja galaktoosi (gal) operonide puhul.
34. Selgitage aminohapete biosünteesi operoni regulatsiooni
E. coli trp
operoni näitel
o Bakterirakus toimub raku kasvuks vajalike komponentide nagu aminohapped,
puriinid, pürimidiinid ja vitamiinid süntees
o Juhul, kui kasvukeskkonnas on piisavalt trüptofaani, toimub trüptofaani
biosünteesis osalevate ensüümide sünteesi mahasurumine vastavate geenide
väljalülitamise teel
o Geenide avaldumise pärssimist vastusena keskkonna signaalile nimetatakse
repressiooniks
o Geenide uuesti tööle lülitamist muutunud olukorras nimetatakse derepressiooniks
o Erinevad biosünteetilised rajad bakterirakus on enamasti represseeritavad. Nii nagu
ka induktsiooni puhul, toimub regulatsioon transkriptsiooni tasemel
Trüptofaani biosünteesi korismahappest kodeerivad viis geeni. Trüptofaani operoni
trp
avaldumine on reguleeritud transkriptsiooni initsiatsiooni repressooni ja transkriptsiooni
enneaegse terminatsiooni kaudu.
Repressioon:
Geen
trpR,
mis kodeerib trp operoni repressorit, paikneb operoni lähedal. Operaatorala O
paikneb tugeva promootori
P
1
regioonis. Lisaks P1–le on operonil ka teine, nõrk promootor
P
2
. Geeni trpA järel paiknevad kaks transkriptsiooni terminaatorjärjestust t ja t’. Järjestus trpL
kodeerib 162-nt pikkust mRNA liiderjärjestust.
Trüptofaani
(korepressori)
puudumisel seondub RNA polümeraas promootorregiooni ja
algatab
trp
geenide transkriptsiooni. Trüptofaani olemasolul rakkudes seondub trüptofaan
repressoriga ning trüptofaan-repressor kompleks seondub promootoralaga kattuvale
operaatorile, takistades transkriptsiooni initsiatsiooni
.
Võrreldes transkriptsioonitasemega
trp
promootorilt trüptofaani olemasolu korral, on
operoni transkriptsioonitase sellelt promootorilt derepresseeritud olekus 70 korda kõrgem.
TrpR mutantide korral on operoni ensüümide tase rakus trüptofaani juuresolekul aga ikkagi
10 korda alla surutud. Põhjuseks on
attenuatsioon, mis viib transkriptsiooni enneaegsele
termineerumisele.
Attenuatsioon:
Kui deleteerida
trp
operonist trpL regioon, tõuseb trp operoni ekspresioonitase. Samas
säilub operoni represseeritavus trüptofaani poolt
. Trp
operoni regulatsiooni järjestuse trpL
vahendusel nimetatakse
attenuatsiooniks ning järjestust trpL
attenuaatoriks. Attenuatsioon
kontrollib transkriptsiooni terminatsiooni mehhanismi operoni liiderjärjestustel.
trp
operoni transkriptsiooni terminatsioon liiderjärjestuselt toimub siis, kui tRNA
Trp on
trüptofaani poolt laetud
. Sel juhul sünteesib RNA polümeraas operonilt ainult 140 nt
pikkuse RNA molekuli
.
Attenuatsiooni piirkonnas asuvad DNA järjestused, mis sarnanevad transkriptsiooni
terminatsiooni signaalidele bakterites. Need järjestused koosnevad G:C rikkast
palindroomsest alast, millele järgneb A:T rikas ala.
Sellele piirkonnale vastav mRNA
moodustab juuksenõelastruktuuri, millega külgneb U – rikas järjestus.
DNA ja RNA vahelised
sidemed nõrgenevad ning RNA polümeraas dissotseerub DNA-lt.
Prokarüootses rakus
toimuvad transkriptsioon ja translatsioon samaaegselt – parasjagu sünteesitavalt mRNA-lt
algab kohe translatsioon
. trp
operoni liiderjärjestus on võimeline moodustama
alternatiivseid sekundaarstruktuure. Kui omavahel paarduvad nukleotiidid regioonidest 1 ja
2 ning 3 ja 4, moodustuvad sekundaarstruktuurid, mis termineerivad transkriptsiooni, kui
aga omavahel paarduvad nukleotiidid regioonidest 2 ja 3, on terminatsiooni põhjustavate
juuksenõelastruktuuride moodustumine takistatud ning transkriptsioon jätkub.
Trüptofaani operoni liiderjärjestuselt toimub ka translatsioon. Liiderpeptiidis on kaks
trüptofaani. Kaks Trp koodonit paiknevad
trpL
järjestuses kõrvuti. Kui rakus pole
trüptofaani, peatub valgusüntees sellel kohal ning omavahel paarduvad nukleotiidid
regioonidest 2 ja 3. Kuna sel juhul ei teki transkriptsiooni terminatsiooni signaali, jätkub
operoni transkriptsioon
. Kui aga rakkudes on trüptofaani, järgneb translatsioon Trp
koodonitelt ning ribosoom takistab regioonide 2 ja 3 omavahelist paardumist. Regioon 3 on
nüüd vaba paarduma regiooniga 4 ning moodustub transkriptsiooni terminatsiooni
struktuur.
Trüptofaani olemasolu korral rakkudes termineerub trp
operoni transkriptsioon
enneaegselt ligikaudu 90% juhtudest. Seega võib
trp
operoni avaldumistase muutuda ligi 700
korda
(10-kordne attenuatsiooniefekt + 70-kordne repressiooniefekt).
Regulatsiooni attenuatsiooni kaudu on kirjeldatud ka treoniini, isoleutsiini, leutsiini,
fenüülalaniini ning histidiini biosünteesi operonide puhul.
35. Geenide ekspressiooni regulatsioon bakterites translatsiooni tasemel.
Ribosoomivalkude sünteesi regulatsioon
Operonisiseselt tranksribeeritakse kõiki geene võrdselt, kuid sageli on raku elutegevuseks
vaja sama operoni poolt kodeeritud valke erineval hulgal. Bakteris toimuvad samaaegselt nii
geeni transkriptsioon, translatsioon kui ka vastava mRNA degradatsioon.
Sama operoni poolt kodeeritud valkude erinev hulk raku kohta võib olla tagatud 3 erineva
mehhanismi poolt:
1) Translatsiooni initsiatsiooni efektiivsus võib erinevate geenide puhul olla erinev
2) Ribosoomide liikumine mRNA molekulil toimub erinevate geenide puhul erineva
kiirusega. Ribosoomide liikumist takistavad mRNA sekundaarstruktuurid
3) Erinevate geenide mRNA degradatsioonikiirus võib olla erinev
Geenide avaldumise regulatsiooni translatsiooni tasemel
on põhjalikult uuritud näiteks
ribosomaalsete valkude puhul.
Kui rakud kasvavad optimaalsetes tingimustes, toimub
nende kiire kasv ning nad vajavad valgusünteesiks rohkem ribosoome kui rakud, mis
kasvavad kehvades tingimustes
. E. coli
ribosoomid sisaldavad kolme erinevat rRNA molekuli
ja 52 erinevat ribosoomi valku.
Enamus ribosomaalsete valkude geene paiknevad
operonides ja nende valkude süntees peab olema koordineeritud kolme erineva rRNA
sünteesiga, sest ribosoomide assambleerimiseks vajatakse neid kõiki kindlas koguses
.
Ribosomaalse valgu S10 geenide klaster sisaldab näiteks 10 geeni. Sellise regulatoorse
üksuse puhul on üks geenidest iseenda ja teiste negatiivseks regulaatoriks
. Geen rplD
kodeerib ribosoomivalku L4
. Kui rakus leidub vabu rRNA molekule, seondub L4 nendega ja
assambleeritakse ribosoomi. Vaba rRNA puudumisel seondub L4 aga S10 operoni mRNA 5’
otsaga, takistades sellelt translatsiooni. Selline mehhanism väldib ribosoomivalkude
asjatut sünteesi rRNA puudumisel
. Negatiivne autoregulatsioon esineb ka teiste
ribosomaalsete valkude operonide puhul.
Ribosoomivalkude translatsiooni kontroll:
o Ribosomaalse valgu
S10 geeni klastris on 10 geeni
o Üks geenidest on iseenda ja teiste negatiivseks regulaatoriks
o Geen
rplD
kodeerib ribosoomivalku L4
o Kui rakus leidub vabu rRNA molekule, seondub L4 nendega ja assambleeritakse
ribosoomi
o Vaba rRNA puudumisel seondub L4 S10 operoni mRNA 5’-otsaga, takistades sellelt
translatsiooni
o Negatiivne autoregulatsioon esineb ka teiste ribosomaalsete valkude operonide
puhul
36. Tooge välja põhilised erinevused prokarüootses ja eukarüootses
geeniregulatsioonis
Geneetilise info realiseerumise erinevus prokarüootidel ja eukarüootidel:
o Hulkraksete
eukarüootide puhul toimub geneetilise programmi realiseerumine –
hulkrakse organismi areng ja elutegevuse tagamine
o Bakterid
– kohanemine keskkonnatingimustega, individualism
Erinevalt bakterirakust toimuvad
eukarüootides transkriptsioon ja translatsioon nii ajaliselt
kui ka ruumiliselt lahus
.
Eukarüootsete geenide regulatsioon on võrreldes bakterigeenidega komplekssem:
1) Geenid paiknevad tuumas
– signaalid peavad esmalt jõudma raku pinnalt
tsütoplasmasse ja sealt läbi tuuma membraani kromosoomideni
. Eukarüootsed
rakud vajavad
transkriptsiooni regulatsiooniks sisemist signaalsüsteemi
2) Eukarüootsete organismide
hulkrakulisus – signaalid peavad liikuma läbi paljude
rakukihtide, enne kui nad jõuavad spetsiifilise koeni, kus konkreetsete geenide
avaldumist kontrollitakse
. Eukarüootse transkriptsiooni seisukohalt on väga oluline
rakkudevaheline kommunikatsioon
3) Eukarüootide geenide regulatsiooni komplitseerib
alternatiivne splaissing: kui geenis
sisaldub palju introneid, võidakse introneid RNA molekulist kõrvaldada kas eraldi
või kombineeritult
ALTERNATIIVNE SPLAISSING –
koos intronitega võidakse kõrvaldada ka nendevehelisi
eksoneid. Moodustuvad erinevad mRNA molekulid, mis kodeerivad erinevaid polüpeptiide
o Regulatsioon väikeste mittekodeerivate RNA-de (ncRNA) kaudu:
Eukarüootidel:
1) mikroRNA
– algne suurem juuksenõelastruktuur eksporditakse tuumast ja
protsessitakse 22-nt dsRNA molekuliks
2) siRNA
– dsRNA ahelad pärinevad kahest komplementaarsest transkriptist
Nii microRNA kui ka siRNA puhul seondub üks ahelatest RISC valkude vahendusel märklaud
mRNA-ga 3´UTR regioonis.
Prokarüootidel:
1) Ei allu protsessingule
, seonduvad märklaud-mRNA-le Hfq vahendusel ja enamasti
mRNA 5´otsale
, mis sisaldab translatsiooni initsiatsiooni saiti
37. Transkriptsiooni aktivatsioon eukarüoodi rakkudes: eukarüootsete geenide
regulatoorsed alad, transkriptsioonifaktorite osa transkriptsiooni aktivatsioonil
Eukarüootset transkriptsiooni mõjutavad:
1) Intronid – transkriptsiooni produktid ei avaldu translatsioonil. Intronid mõjutavad ka
nukleosoomi struktuuri. Immunoglobuliinide geenidel asuvad transkriptsiooni
regulaatorite seondumissaidid intronites. RNA editing on mõjutatud intronite poolt
2) Geenide avaldumist mõjutab ka DNA kondensatsiooniaste: Transkriptsiooni
initsiatsiooniks peab DNA olema kättesaadav RNA polümeraasile ja
transkriptsioonifaktoritele. DNA liiga tugev seondumine histoonidega ja teiste
struktuursete valkudega pärsib transkriptsiooni
3) Geenide avaldumises mängib olulist rolli geeni paiknemine kromosoomis
4) Histoonide atsetüleerimine/deatsetüleerimine – histoonide atsetüleerimine muudab
kromatiini avatumaks – võimaldab transkriptsiooni initsiatsioonil osalevate valkude
juurdepääsu DNA-le. Histoonide deatsetüleerimine suleb kromatiini – transkriptsioon
on pärsitud
Regulaatorvalgud interakteeruvad koaktivaatoritega ja korepressoritega:
o Korepressorid
– histoonide deatsetülaasi kompleksid
o Koaktivaatorid:
1) Histoonide atsetülaasi kompleksid
2) SWI/SNF kromatiini remodelleerivad faktorid – dissotseerivad DNA ajutiselt histoonidelt,
võimaldamaks transkriptsiooni initsiatsiooniks vajaliku kompleksi seondumist DNA-le
3) Mediaatorkompleksid – vahendavad transkriptsiooni aktivaatorite mõju
initsiatsioonikompleksile
Transkriptsiooni aktivatsioon eukarüootses rakus:
Esmalt on vajalik nukleosoomi struktuuri muutus – DNA histoonide ümbert lahtiharutamine,
et sellele pääseksid ligi polümeraasid.
o RNA polümeraas I – tuumakeses, viib läbi ribosomaalse rRNA (va. 5S rRNA) sünteesi
o RNA polümeraas II – rakutuumas, katalüüsib kõigi pre-mRNA-de sünteesi
o RNA polümeraas III – rakutuumas, katalüüsib 5S rRNA, tRNA-de ja snRNA-de sünteesi
Geenide aktiveerumine toimub mingitele signaalidele vastusena
(nt hormoonid,
väliskeskkonna stiimulid – nt valgus, temperatuuri muutused jne)
.
Erinevad transkriptsiooni aktivaatorid võivad seonduda promootorist erinevale kaugusele.
Kaugemale seondunud valgud tuuakse initsiatsioonikompleksi DNA lingu abil.
Osa transkriptsioonifaktoreid seonduvad geenist eemal asuvatele DNA järjestustele –
transkriptsiooni võimendajatele. Enamus transkriptsioonivõimendajaid toimivad
koespetsiifiliselt.
Transkriptsioonifaktorite osa trankriptsiooni aktivatsioonil:
Enamus TF-e sisaldab vähemalt kahte olulist domääni – DNA-ga seonduvat domääni ja
transkriptsiooni aktivatsiooni domääni.
o Tsink-sõrmed
– lühikesed lingud, mis moodustuvad siis, kui kaks tsüsteiini ühest
polüpeptiidi osast ja kaks histidiini teisest osast seovad koos tsingi iooni
– olulised
valgud DNA-ga seondumisel
o Heeliks-pööre-heeliks motiiv
– 3 lühikest AH-de heeliksit on üksteise suhtes
pööratud. C-terminaalsem kolmest heeliksist vastutab DNA-ga seondumise eest,
ülejäänud kaks aga valgu dimeriseerumise eest
. Olulisimad Homeobox
valgud –
määravad ära tulevase organismi kudede orienteerumise
(?)
o Leutsiini lukud
– rida AH-sid Leu-ga igas seitsmendas positsioonis ühest
polüpeptiidist dimeriseerub teise polüpeptiidiga samasuguse motiivi kaudu.
Polüpeptiidide interaktsioon toimub Leu-de kaudu.
Paiknevad positiivselt laetud
AH-de kõrval, mis dimeriseerumise järel pääsevad esile ja seonduvad negatiivselt
laetud DNA-ga
o Heeliks-ling-heeliks motiiv
– 2 helikaalset AH-de regiooni on teineteisest eraldatud
mittehelikaalse linguga
. Helikaalsed regioonid võimaldavad dimeriseerumist teise
polüpeptiidi samasuguse regiooniga. Motiivid külgnevad positiivselt laetud
aminohapetega – dimeriseerumine soodustab dimeeri seondumist DNA-le
38. Eukarüootsete geenide transkriptsiooni vaigistamise molekulaarsed mehhanismid
DNA metüleerimise kaudu. Imprinting
Kui DNA on metüleeritud, siis on geenide transkriptsioon rohkem mahasurutud
(halvemini
kättesaadav)
. DNA metülatsioon soodustab histoonide deatsetüleerimist.
Imetajate genoomis on keskmiselt 40% G:C paare ja neist 2-7%-l on C metüleeritud. C
metüleeritud piirkondi nim CpG saarekesteks
(1-2 kb pikkused)
, kus DNA on G:C rikkam.
Inimese genoomis on leitud 45 000 CpG saarekest. Enamus neist asuvad transkritpsiooni
alguspunktide läheduses.
Kui DNA on metüleeritud, viib see transkriptsiooni repressioonile.
CpG metülatsioon
indutseerib histoonide H3 ja H4 deatsetüleerimist.
Deatsetüleerimist soodustab
metüülrühmaga seonduv valk MeCP2
, mis moodustab multiproteiinse repressorkompleksi
koos deatsetülaasidega HDAC1 ja HDAC2. Repressorkompleks muudab kromatiini struktuuri.
DNA metülatsioonist tingitud geenide repressioon on stabiilne ning seda muudab ainult
kas DNA demetülatsioon või tugev transkriptsiooni aktivaator, mis suudab DNA-ga
seonduda, hoolimata selle metülatsioonist.
Kõige ulatuslikumalt on geenide transkriptsioon
DNA metülatsiooni kaudu pärsitud
imetajate X kromosoomi inaktivatsiooni puhul.
IMPRINTING
ehk vermimine:
o Imetjate
DNA metülatsiooniga seotud nähtus, kus teatavate geenide avaldumine on
kontrollitud selle kaudu, kummalt vanemalt geen on päritud
o Hüpermetülatsiooni tõttu avaldub vaid üks paar geenidest
Nt: hiire embrüos avaldub ainult isalt päritud kasvufaktorit kodeeriv
Igf2
. Emalt päritud geeni
ei transkribeerita, sest seda pärsivad Cpg saarekesed. Heterosügootsete hiirte kasv sõltub
sellest, kas normaalne Igf2 alleel on pärit isalt või emalt
(kui emalt, siis on normaalne kasv;
kui isalt, siis kääbuskasv)
.
Inimestel on samuti imprintingut kirjeldatud – nt PW-sündroom
(mõjutab lapse
rasvumist, hormoone, hingamise ja veresoonkonna arengut, neuroloogilised efektid)
, kus
mõjutatud on piirkond 15. kromosoomis
(emalt pärit kromosoomis on see piirkond alati
metüleeritud ja geenid ei avaldu)
.
39. Kromatiini struktuuri mõju geenide avaldumisele. Kromatiini remodelleerimine
Kromosoomidega seotud DNA pole transkriptsiooniks tavaliselt valmis.
Nukleosoomse
struktuuri muutuseid, mis toimuvad selleks, et geen transkribeeruks, nimetatakse
kromatiini modifitseerimiseks
. Kromatiini modifikatsioonilisi komplekse on kahte tüüpi:
1) Histoonide modifitseerimine
. Kompleksis on ensüümid histooni
atsetüültransferaasid (HAT), mis
atsetüülivad histoonid kindlates punktides
aminohappe lüsiini atsetüülgruppide ülekandega sama nukleosoomi piires
.
Kompleksis osalevad ensüümid
kinaasid, mis kannavad üle fosfaatgruppe, tehes
fosforüülimst
. Toimub ka teisi keemilisi modifikatsioone
2) Histoonide remodelleerimine e. ümberkujundamine
. On kirjeldatud mõnikümmend
eritüübilist kompleksi. Näiteks
SWI/SNF koosneb vähemalt kaheksast erinevast
valgust. Komlpeks reguleerib transkriptsiooni, liigutades histooni oktameere
assotsieerunud DNA suhtes. Sellega vabastatakse vastav DNA piirkond
transkriptsioonifaktorite seondumiseks
. Seesuguseid protsesse nimetatakse ka
nukleosoomide nihutamiseks.
Teisalt
aktiivset kromatiini saab muuta ka inaktiivseks kromatiiniks. Selline kromatiini
tagurpidi ümbermodelleerimine toimub läbi kahe nukleosoomi tänu histoonide
biokeemilisele modifikatsioonile
: deatsetüülimine histooni deatsetülaaside toimel ja
metüülimine histooni metüültransferaaside toimel.
Eu- ja heterokromatiin:
Enamik aktiivseid geene paikneb eukromatiinis.
Eukromatiinis on DNA vähem kokkupakitud
olekus.
Ligi 30% Drosophila
DNA-d on heterokromatiinne. Enamik Drosophila
heterokromatiinseid piirkondi sisaldab kordus-DNA-järjestusi, kus eristatakse nii
mõõdukalt kui ka sageli korduvaid järjestusi
. Teisalt, Drosophila
genoomis on teada ka ligi
20 valku määravat geeni, mis paiknevad heterokromatiinsetes osades, ning mitusada
rRNA-geeni, mis piknevad X- ja Y-kromosoomi tsentromeerses heterokromatiinses
piirkonnas
. Seega, mitte kogu heterokromatiin pole geneetiliselt inaktiivne. Kromosoomsete
ümberkorralduste tagajärjel, kui heterokromatiinse ala geenid liiguvad eukromatiinsesse
piirkonda, võib nende avaldumine muutuda
. Muutusi suurendavad supermutatsioonid, mis
pidurdavad samuti eukromatiinsete geenide avaldumist heterokromatiinses alas. Neil
geenidel ongi tendents avalduda ebanormaalselt. Erinevalt eukromatiinsetest geenidest on
heterokromatiinsete geenide avaldumisel asendiefekt. Seepärast võib samal organismil
tekkida segu normaalsetest ja mutantsetest tunnustest. Seda nähtust nimetatakse geeni
asendiefekti mitmekesisuseks.
40. X kromosoomi inaktivatsioon imetajatel. Miks erinevad 45, X genotüübiga naised
normaalsetest naistest?
o X kromosoomi inaktivatsioon algab kindlast kohast, mida nimetatakse X
inaktivatsiooni keskuseks XIC (
X inactivation center
) ning levib seejärel mõlemas
suunas
o Geen
XIST
(X inactive specific transcript), mis paikneb XIC regioonis, jääb
transkriptsiooniliselt aktiivseks
o Geenilt
XIST
sünteesitakse RNA molekulid on assotsieerunud inaktiivse X
kromosoomiga, takistades sealt transkriptsiooni
o Aktiivses X kromosoomis on geeni
XIST
transkriptsioon pärsitud
o Aktiivsete kromosoomide puhul on histoon H4 atsetüleeritud
o Inaktiivse X kromosoomi puhul on atsetüleeritud H4 leitud ainult vähestes
regioonides
o Inaktiveerunud X kromosoom on interfaasis nähtav Barri kehakesena
o Kumb X kromosoomidest inaktiveerub, on juhuslik
45, X genotüübiga naised erinevad normaalsetest naistest:
Turneri sündroomiga sünnib lapsi sagedusega 1:3000. Sellise kromosoomianomaaliaga
indiviididel ilmneb kasvupeetus, viljatus, kerged anatoomilised kõrvalekalded. Kui tervetel
naistel on erinevates rakkudes juhuslikult inaktiveeritud kas isalt või emalt pärti X
kromosoom, on siiski üksikuid geene, mis peavad olema aktiivsed mõlemas X kromosoomis.
Leitud on >20 geeni, mis on aktiivsed mõlemas X kromosoomis. Osa neist kodeerivad
transkriptsioonifaktoreid, mõned neist on olemas ka Y kromosoomis. See seletab, miks
Turneri sündroomiga (genotüüp 45, X) enamus embrüoid hukkub ning miks
45, X naised on
viljatud
. Neil puudub Barri kehake.
41. Selgitage DNA „sõrmejälgede“ meetodit ja selle rakendusi
DNA polümorfism võimaldab tuua esile indiviidile omapärase DNA fragmentide mustri,
mida nim DNA sõrmejälgedeks.
Testi läbiviimisel võrreldakse DNA piirkondi, mis sisaldavad kordusjärjestusi VTNR-id
(variable number tandem repeats)
. Erinevate indiviidide DNA töötlemisel restriktaasidega
saadakse erinev fragmentide muster
(kasutatakse Southern Bloti meetodit)
. Mida rohkem
erinevaid hübridisatsiooniproove kasutatakse, seda suuremat osa genoomist võrreldakse –
saadakse usaldusväärsemad tulemused.
Võimalik lähtuda väga väikesest bioloogilise materjali kogusest ja amplifitseerida sealt PCR
meetodil DNA järjestused, mida edaspidi analüüsitakse.
Rakendusi:
o Kohtumeditsiin
– isikute tuvastamine
o Isikute tuvastamine pärast katastroofi
– nt suured lennuõnnetused
o Ajalugu
– isikute tuvastamine matmispaikades jms
o Isadustestid
– isaduse tuvastamisel tuleb arvestada seda, et homoloogilistest
kromosoomidest pooled on pärit isalt, pooled emalt. Lapse DNA fragmentide mustris
sisaldub nii isale kui ka emale iseloomulikke fragmente
42. Tooge näiteid geneetilisi haigusi põhjustavatest mutatsioonidest ja selgitage
haiguste molekulaarset diagnostikat
Haiguste molekulaarne diagnostika:
Haiguste molekulaarne diagnostika eeldab konkreetse haigusega seotud geenide
nukleotiidse järjestuse ja haigust põhjustavate mutatisoonide teadmist
. Kui mutatsioonide
kohad on lokaliseeritud, saab vastavat DNA regiooni uuritavate indiviidide genoomist
PCR-i teel amplifitseerida ja seejärel juba täpsemalt analüüsida.
Võimalik on teostada ka
sünnieelset diagnostikat ja testida kromosoomide anomaaliaid.
PDG-d
(pre-implantation genetic diagnosis)
rakendatakse kunstlikul viljastamisel enne
embrüo viimist emakasse. Eraldatakse rakk 3-päevasest embrüost
(8-10 raku staadium)
.
Geneetilisi haigusi põhjustavaid mutatsioone:
1) Kolmenukleotiidse järjestuse liigne kordumine
– nt: Huntingtoni tõbi, fragiilne X,
müotoonne düstroofia jt
Huntingtoni tõbi:
Tegemist on dominantselt avalduva haigusega
(mutatsioon 4. kromosoomis paiknevas
geenis, mis kodeerib valku huntingtiini)
. Avaldub tavaliselt 30-50. eluaasta vahel tsentraalse
närvisüsteemi progresseeruva degeneratsiooni näol
(võnkuvad liigutused; dementsus)
ja
lõpeb 10-15 aastat hiljem patsiendi surmaga
. Varases staadiumis on võimalik kontrollida
motoorikat.
Huntingtoni tõbe on
võimalik diagnoosida, kui ülesamplifitseerida CAG järjestus.
Diagnostika on võimalik ka sünnieelselt.
Müotoonne düstroofia:
Tegemist on ühe lihasdüstroofia vormiga. See on
dominantne mutatsioon 19. kromosoomis
(DM1, CTG järjestuse kordistumine väljaspool DMPK geeni; häiritud on RNA protsessimine –
muudab kromatiini struktuuri ja mõjutab läheduses olevate geenide avaldumist)
või 3.
kromosoomis
(DM2, CCTG järjestuse kordistumine ZNF9 geeni intronis)
o Lihastegevus tugevalt häiritud
(avaldub nt näolihastes – nagu oleks mask ees; mõjub
südamelihasele jne)
Fragiilne X:
o Põhjustab vaimset alaarengut
o Haiged on heterosügootsed naised ja hemisügootsed mehed
o CGG järjestuse kordistumine
kuni 1000X
o Pärsib
FMR1 geeni avaldumise, mis mõjutab aju arengut juba sünnieelselt
o Premutatsioonilise
FMR1
alleeli puhul võib DNA polümeraas libiseda DNA-l tagasi ja
hakata sünteesima uuesti sama regiooni
2) Mutatsioonid ioonkanalites, 3-nukleotiidse järjestuse deletsioon
Tsüstiline fibroos:
o Autosomaalne retsessiivne mutatsioon
o Mutatsioonid geenis, mis kodeerib transmembraanset regulaatorit CFTR, mis asub
ioonkanalites, osaledes soolade transpordis rakust sisse ja välja
– häiritud on
klooriioonide transport hingamiselundites, pankreases, soolestikus ja teistes
organites
o Haigetel on
kõrge risk surra kopsupõletikku juba imikueas
o Diagnostika Southern Bloti meetodil –
potentsiaalset deletsiooni sisaldava DNA
piirkond amplifitseeritakse PCR teel. Amplifitseeritud DNA hübridiseeritakse
erinevate radioaktiivselt märgistatud oligonukleotiididega.
Hübridisatsioonitingimused on selektiivsed
: üks oligotest hübridiseerub ainult
algtüüpi DNA järjestusele, teine aga deletsiooni sisaldavale
o Mutantse
CF
lookuse isoleerimisel kasutati positsioonilist kloneerimist
3) Punktmutatsioonid
Retinitis pigmentosa – kanapimedus; pimedaks jäämine:
o Võrkkesta pigmendi kadumine
; lõppeb pimedaks jämisega
o Mutatsioonid rodopsiini geenis
Sirprakuline aneemia:
o Diagnoosimine DNA southern bloti meetodil
o Mutatsiooni tagajärjel on ära kadunud üks restriktaasi lõikesait beeta-globiini
geenist
4) Ühenukleotiidne polümorfism
– nt hemofiilia; kõrgenenud risk südame- ja
versoonkonna haigustele; muutused ravimi metabolismis
43. Selgitage geeniteraapia meetodeid ja rakendusi
GEENITERAAPIA – seisneb normaalse geeni viimises indiviidi genoomi, mis sisaldab defektset
geeni.
Juhul kui geeniteraapia on olnud edukas, sünteesib transgeenne organism varem puudunud
geeniprodukti ja selle tulemusena taastub tal normaalne fenotüüp.
Eristatakse 2 tüüpi geeniteraapiat:
1) Somaatiliste rakkude mittepäritav geeniteraapia
2) Sugurakkude geeniteraapia ehk päritav geeniteraapia
Inimese puhul on kasutatud ainult somaatiliste rakkude geeniteraapiat.
Geeniteraapias kasutatavad vektorid:
1) Retroviirustel põhinevad vektorid
: + Transgeen integreerub koos retroviiruse
DNA-ga raku genoomi ja kandub edasi rakuliini pidi; - Integratsioon võib
põhjustada kahjulikke mutatsioone
2) Adenoviirustel põhinevad vektorid: + Viiruse genoom koos transgeeniga
replitseerub autonoomselt ja püsib rakkudes seni, kuni immuunsüsteem selle
elimineerib; - Transgeen püsib rakkudes lühiajaliselt
Rakendusi:
Kasutatakse geneetiliste defektide parandamisega juhtudel, kus mõlemad geenikoopiad on
defektsed.
44. Geeniteraapia rakendamisega seotud riskid
Riskid:
o Translokatsioon
– põhjustatud retroviirustest
o Mõnede geenide üleekspressioon, mis võib viia mutatsioonideni
vms
o Liiga tugev immuunvastus
– viiruse genoomil põhinevate vektorite kasutamine
transgeeni rakku viimiseks võib põhjustada väga tugeva immuunvastuse viiruse
valkude vastu
45. Transgeensed organismid, nende konstrueerimise eesmärgid
TRANSGEEN – ühest organismist isoleeritud geen, mis on viidud teise organismi rakkudesse.
TRANSGEENNE ORGANISM – organismi, kuhu on viidud talle võõras geen
Eesmärgid:
1) Soovitavate tunnuste lisamine või võimendamine kultuurtaimedel ja koduloomadel
– nt suurem piimaand lehmal
2) Huvipakkuva produkti tootmine taimes või loomas
– nt ravimid
3) Transgeensete organismide konstrueerimine eesmärgiga
uurida bioloogiliste
protsesside toimumise molekulaarseid mehhanisme
Transgeeni rakku viimiseks on erinevaid võimalusi:
o Transformatsioon
o Rakku tulistamine volframi kuulikeste koostises
o Süstimine
o Viirused
46. Eukarüootsete valkude bakterites tootmiseks vajalikud geenimanipulatsioonid
o Bakterid on pandud sünteesima inimese insuliini, kasvuhormooni ja interferooni
o Tuleb lähtuda cDNA-st, kust on intronid kõrvaldatud. Geeni liidetakse ala, mis
omab translatsiooni initsiatsiooni signaali. Piirkond viiakse bakteri regulatoorsete
elementide kontrolli alla. Bakterisse viiakse see plasmiidi koosseisus või viiruste
abil. Valku on võimalik toota nii rakku sisse, kui ka rakust välja
(nt: somatotropiini
tootmine)
o Inimese kasvuhormooni ekspressioonikonstrukt bakteris
E. coli
o Ensüümide tootmine bakterites – nt: juustu valmistamisel vajalik renniin
o Võõrvalgu ekspressioon võib erineda erineva koodonkasutuse tõttu: ekspressioon
võib täiesti puududa või olle üleekspresseeritud
47. Transgeensete loomade ja taimede saamine. Tooge mõned näited
Transgeensete loomade saamine:
1) Embrüode nakatamine retroviiruste baasil konstrueeritud rekombinantsete DNA
molekulidega
2) Enamasti
süstitakse rekombinantne DNA viljastatud munarakku
Süstitav DNA võib olla kas lineaarne või rõngasmolekul
. Lineaarne DNA integreerub
genoomi ligi 5 korda efektiivsemalt kui tsirkulaarne DNA.
Ühte munarakku süstitakse sadu
kuni tuhandeid rekombinantse DNA molekule. Sisseviidud DNA
(transgeen)
integreerub
genoomi suvalistesse kohtadesse.
DNA süstimisel viljastatud munarakku toimub transgeeni integreerumine genoomi enamasti
embrüo arengu varajastel etappidel
. G
0
generatsioon on geneetilised mosaiigid. Et saada
transgeenseid loomi, kus tunnus on kinnistunud, ristatakse G0 generatsiooni isendeid
. G
1
generatsioonis on transgeensed need, kelle vanematel oli transgeen integreerunud
sugurakkudesse.
Nt:
Transgeense hiire saamine – embrüonaalsed tüvirakud ES isoleeritakse blastulast. Pärast
võõr-DNA sisestamist viiakse ES rakud mõnda teise arenevasse embrüosse. Osa neist on
aluseks organismi kudede moodustumisel. Sel viisil saadud organismid on kimäärsed,
sisaldades nii normaalseid kui ka mutantseid rakke
. Juhul kui mõni mutantne rakk on olnud
aluseks sugurakkudele kujunemisele, kandub mutatsioon ka järglastele.
Transgeensete hiirte kasutamine:
1) Imetajate geeniekspressiooni uurimine
2) Geenide ülekandevektorite ja meetodite katsetamine, et neid hiljem kasutada
inimese puhul
Lisaks on konstrueeritud veel transgeenseid sigu, kitsi, kanu, kalu.
Transgeensed taimed:
1) DNA tulistatakse volframi või kullapartiklite koosseisus taimerakku
2) DNA sisestamine elektrivoolu abil (elektroporatsioon)
3) Taimerakke transformeeritakse taimerakke
Agrobacterium tumefaciens
Ti
plasmiidide põhjal konstrueeritud rekombinantsete plasmiididega
Taimede geneetilisel manipuleerimisel on
suureks abiks taimerakkude totipotentsus – iga
üksiku taimeraku võime dediferentseeruda embrüonaalseks rakuks, millest seejärel
kasvatatakse uus taim
.
Nt
: herbitsiidide-resistentsed taimed, külmatolerantsed taimed, kahjurite-resistentsed
taimed, köögiviljade säilivustähtaja pikendamine, transgeenne riis
(vähendavad raua ja
vitamiin A defitsiiti)
.
48. Mille poolest erinevad vähirakud normaalsetest keharakkudest? Tooge tõendeid
vähi geneetiliste aluste kohta
o Vähirakkude
puhul toimub rakkude kasv ja jagunemine kontrollimatult. Normaalsed
keharakud
jagunevad harva ning kontrollitult
o Rakud jagunevad, kuhjudes üksteise otsa ning moodustavad tuumori
: healoomuline
kasvaja
(tuumori rakud ei levi ümbritsevatesse kudedesse)
; pahaloomuline kasvaja
(kasvaja levib teistesse kehapiirkondadesse, moodustades sekundaarseid tuumoreid –
metastaas)
. Keharakud jagunedes kuhilaid ei moodusta
o Vähirakud on surematud
. Normaalsed rakud ei suuda jaguneda rohkem kui 60-70
korda
o Vähirakkudes on aktiveerunud telomeraas
. Normaalsetes rakkudes on telomeraas
inaktiivne
o DNA kahjustusi ei parandata
o Vähirakud ei allu teiste rakkude signaalidele
*Vähirakke on võimalik saada normaalsete rakkude kultuurist, kui neid rakke töödelda
kartsinogeenidega
(teatud viirused, kiirgus või mutageensed kemikaalid)
.
Geneetilised alused:
o Vähirakkude kasv ja jagunemine ei allu teistelt rakkudelt saadetud signaalidele
o Nõrgema immunsusega organismidel võib kergemini tekkida vähkkasvaja
o Kontrollimatut jagunemist põhjustavad muutused geenide avaldumises
o Üleminek ühest rakutsükli faasist teise toimub rakuväliste ja siseste keemiliste
signaalide koostöös. Kui kontrollpunkt ei reageeri õigesti, võib DNA replikatsioon
toimuda ka DNA kahjustuste korral – suureneb mutatsioonide hulk, millest osa
võivad põhjustada vähki
Tõendid vähi geneetilistele alustele:
o Vähirakkude fenotüüp ilmneb ka tütarrakkudel
o Vähiteket võivad indutseerida teatud viirused
o Vähki indutseerivad kartsinogeenid põhjustavad DNA-s mutatsioone
o Mõnedes suguvõsades on vähk päritav
o Teatud verevähi vormid
(leukeemia, lümfooma)
on seotud kindlate kromosoomide
aberratsioonidega
Vähki põhjustavad mutantsed geenid jaotuvad kahte klassi:
o Onkogeenid
– geenid, mille mutantsed alleelid stimuleerivad rakkude jagunemist
o Tuumori supressorgeenid
– mutantsete geenide puhul ei suudeta pärssida raku
jagunemist. Mutatsiooni toime ilmneb juhul, kui mõlemad alleelid on mutantsed!
49. Võrrelge viiruslikke onkogeene ja nende rakulisi homolooge. Selgitage
mehhanisme, kus need geenid stimuleerivad vähki
Viiruslikud onkogeenid
(v-src)
:
o Tuumoreid indutseerivate retroviiruste puhul on kirjeldatud üle 20 viiruse onkogeeni
o Kodeerivad kasvufaktoritele ja nende retseptoritele sarnaseid valke, proteiini
kinaase, transkriptsioonifaktoreid
o Inimesel võivad vähki tekitada: HPV, adenoviirus, herpes, hepatiit B, hepatiit C,
inimese T-raku leukeemiaviirus 1 jt
o Puuduvad intronid
(c-onc geenilt transkribeeritud ja protsessitud RNA on lisandunud
retroviiruse genoomi)
o Retroviiruste onkogeenid on rakuliste geenide muteerunud variandid, mille
ekspressioon aktiveerub pärast viirusinfektsiooni. Need onkogeenid on rakud
omandanud eelmisest nakatunud rakust. Selline onkogeenide kaasavõtmine raku
jagunemisel on juhuslik ja seotud viiruse elutsükliga
o Onkogeenide aktivatsiooni mehhanismid:
onkogeeni kandva retroviiruse
transduktsioon; onkogeene mittekandva retroviiruse insertsioon raku genoomi
(kui
LTR inserteerub proto-onkogeeni lähedusse, siis määrab ta selle transkriptsiooni ning
rakulise onkovalgu ümberproduktsiooni)
; viiruse DNA genoomi replikatsiooniks
vajalike onkovalkude ekspressioon; viiruseline transkaktivatsioon retroviiruse
puhul
Rakulised homoloogid viiruslikele onkogeenidele – protoonkogeenid:
Viiruslike onkogeenide rakulisi homolooge nim protoonkogeenideks või normaalseteks
rakulisteks onkogeenideks.
o Rakuline homoloog geenile v-src on c-src
. Kodeerivad järjestused on mõlemal geenil
sarnased, erinedes teineteisest ainult 18 nt poolest
o c-ong geenide poolt kodeeritud valgud on olulised rakutsükli regulatsioonis.
Mutatsioonide tõttu nendes geenides on valgud kas üleekspresseeritud või pidevalt
aktiivses vormis: rakkude kontrollimatu jagunemine, vähi teke
o Olemas intronid
Rakuliste onkogeenide aktivatsioonimehhanisme on nelja tüüpi:
1. Punktmutatsioonid rakulistes proto-onkogeenides
2. Rakuliste proto-onkogeenide amplifikatsioon
3. Rakuliste proto-onkogeende translokatsioon aktiivse promootori/enhanseri alla.
4. Liit-onkovalkude teke translokatsioonil
Miks v-onc geenid indutseerivad tuumoreid, c-onc geenid aga mitte?
1) Viiruslike geenide ekspressioonitase on võrreldes rakuliste homoloogidega oluliselt
kõrgem
2) Viiruslikud onkogeenid võivad avalduda rakutsükli valel etapil
3) Viirusliku onkogeeni produkt võib olla rakulise homoloogi mutantne versioon ning
seetõttu on uute omadustega
Mutatsioonid, mis muudavad protoonkogeeni vähki stimuleerivaks:
o Geeni
translokatsioon või transpositsioon
o Geeni
amplifikatsioon (geeni koopiaarv suureneb)
o Mutatsioonid geeni regulatoorses alas
o Mutatsioon geenis
50. Tuumori supressorgeenide funktsioonid rakus. Selgitage rakutsükli
regulatsioonimehhanisme, mis käivituvad DNA kahjustuste korral
Tuumori supressorgeenid
– mutantsete geenide puhul ei suudeta pärssida raku jagunemist.
Mutatsiooni toime ilmneb juhul, kui mõlemad alleelid on mutantsed!
pRB kontrollib rakutsükli geenide transkriptsioonifaktoreit E2F-i:
Normaalse rakutsükli puhul, kui kontrollpunkti on jõudnud positiivsed signaalid selle kohta,
et rakk võiks jaguneda, seonduvad tsükliinid tsükliinidest sõltuvate proteiinikinaasidega ning
seejärel Rb fosforüülitakse. E2F-pRB kompleks dissotseerub ning E2F aktiveerib nende
geenide transkriptsiooni, mis on vajalikud raku jagunemiseks.
p53:
Üle 50% inimese vähijuhtudest on leitud mutatsioone tuumori supressorgeenis, mis kodeerib
tranksriptsiooni aktivaatorit p53.
p53 toimib transkriptsiooni-faktorina, indutseerides p21
sünteesi. p21 inhibeerib CDK-de fosforüleeriva aktiivsuse. pRB jääb E2F-ga kompleksi.
p53 roll rakutsükli regulatsioonis: 1) rakutsükli peatamine; 2) DNA
reparatsioonisüsteemide aktivatsioon; 3) apoptoos
Regulatsioonimehhanisme, mis käivituvad DNA kahjustuste korral:
o BRCA1 ja BRCA2 valgud osalevad DNA reparatsioonis, homoloogilises
rekombinatsioonis ja transkriptsiooni regulatsioonis
o Nad seonduvad transkriptsioonifaktoritega, osalevad kromatiini remodelleerimises.
Nende puudumisel akumuleeruvad rakkudes kromosoomide katked
o BRCA1 seondub dsDNA katketega ja initsieerib koos histoonilaadse valguga H2AX
reparatsiooni, modifitseerides lokaalselt kromatiini struktuuri
o DNA valepaardumisi kõrvaldav
mismatch
reparatsioon MMR
o Nukleotiidi väljalõike reparatsioon NER
– tuntakse ära piirkonnad DNA-s, mis on
kahjustunud
(nt: tümiini dimeerid)
; sinna tehakse katke ning kahjustunud koht
lõigatakse välja. DNA-le sünteesitakse vastu uus komplementaarne ahel
51. Selgitage geneetilist tagapõhja, miks on vähiteke astmeline protsess ning
geneetilist eelsoodumust vähitekkeks
Osa vähitüüpe areneb välja siis, kui teatav geen ei avaldu enam. Nt: retinoblastoomi
(võrkkesta vähks)
põhjustab mutatsiooni RB geenis.
Knudsoni kahe tabamuse hüpotees – vähi teke eeldab, et defektsed on mõlemad tuumori
supressorgeeni alleelid.
Vähiteke on astmeline protsess
, mille käigus akumuleeruvad somaatilised mutatsioonid
geenidesse, mis kontrollivad raku jagunemist. Nt: soolevähi kujunemiseks on vaja vähemalt 7
sõltumatut mutatsiooni: Esmalt tekib mutatsioon teises
APC
geeni alleelis, siis mutatsioonid
K-ras
proto-onkogeeni alleelides, 18-nda kromosoomi tuumori supressorgeenide alleelides ja
TP53
alleelides.
Kuna mutatsioonid peavad tekkima mõlemasse geeni alleeli, siis võib elu jooksul
muteeruda üks alleel, hiljem ka teine. Tavaliselt on vajalik veel mõne rakutsüklit
reguleeriva geeni muteerumine
.
Samuti ei pruugi alati välja areneda kohe pahaloomuline kasvaja.
Soolevähi eellaseks on
healoomuline kasvaja sooles, mis elu jooksul ning mutatsioonide kuhjumise tulemusena
võib välja kujuneda pahaloomuliseks. Pahaloomuline kasvaja hakkab levima ka teistesse
kudedesse – tekivad metastaasid
.
Päritavate vähihaiguste puhul
on laps vanematelt saanud enamasti ühe mutantse alleeli.
Rakupopulatsioonis tekib varem või hiljem spontaanne mutatsioon ka geeni teise alleeli
ning sellest rakust, milles on mutatisooni tagajärjel geeni mõlemad alleelid defektsed,
arenebki tuumor.
Päritavate vähivormide puhul on enamasti tegemist mutatsioonidega kas
tuumori supressorgeenides või DNA reparatsiooniensüüme kodeerivates geenides
.
52. Milline roll on emapoolse mõjuga geenidel hulkrakse organismi arengus? Selgitage
seda äädikakärbse mudeli näitel
Drosophila
munarakk on ellipsoidne, 1 mm pikkkune. Iga munarakk on ümbritsetud
spetsiaalse kihiga – koorioniga. Esimeses otsas on kaks filamenti, mis aitavad raku sisemusse
tuua hapnikku. Seemnerakk siseneb läbi teise esiosas paikneva struktuuri, mida nimetatakse
mikropüüliks. Vahetult peale viljastamist on sügoodi jagunemine nii kiire, et tütarrakkude
vahele ei jõua membraani moodustuda ja tekib sünsüütsium. Pärast 9-ndat jagunemist
sisaldab sünsüütsium 512 tuuma, mis seejärel liiguvad embrüo äärealadesse ja jagunevad
veel neli korda. Osa rakke liigub veel spetsiaalselt embrüo tagumisele poolusele. Pärast
13-ndat jagunemist eralduvad rakud tsütoplasmaatiliste membraanidega ning tekib rakuline
blastoderm, kus on üle 4000 raku. Blastodermi tagumisele poolele liikunud tuumadest
moodustuvad pooluse rakud, mis on aluseks sugurakkudele. Seega toimub somaatiliste ja
sugurakkude liinide eristumine juba väga varakult, blastula staadiumis.
Umbes päevaga areneb äädikakärbse embrüost vastne. Vastne kasvab väga kiiresti, vahetab
2 korda kesta ning moodustab viie päeva pärast nuku. Nukustaadiumis (neli päeva) hävivad
paljud vastsespetsiifilised koed ning moodustuvad täiskasvanud kärbsele iseloomulikud
organid nagu tundlad, silmad, tiivad ja jalad. Kuna täiskasvanud isendit nimetatakse
valmikuks, imaagoks (ingl. k.
imago
), nimetatakse vastse erinevates osades paiknevaid
rakukogumikke, millest arenevad organid, immaginaaldiskideks (
imaginal discs
).
Munaraku kasvamiseks ja valmimiseks ning viljastumisjärgseks embrüo arenguks
transporditakse munarakku teda ümbritsevatest rakkudest toitaineid ja komponente, mis on
olulised embrüo arenguks. Mõnede liikide puhul määravad need komponendid embrüo
põhiplaani, pea-saba ning kõht-selg eristumise.
Mutatsioone ema geenides, mis ei mõjuta ema enda eluvõimelisust ega munarakkude
valmimist, kuid mõjutavad järglaste arengut, nimetatakse emapoolse mõjuga
mutatsioonideks. Geene, mille muteerumine sellist efekti põhjustab, nimetatakse
emapoolse mõjuga geenideks.
Retsessiivne mutatsioon geenis
dorsal
(dl) ei võimalda eluvõimelisi järglasi saada siis, kui
ristata mutantseid homosügootseid emaseid metsiktüüpi homosügootsete isastega
. Dl
määrab selle, kuhu tekib embrüo kõhtmine ja kuhu selgmine pool. Mutatsiooni korral
kõhtmist poolt ei arene ja embrüo on kahe selgmise poolega.
Isapoolne metsiktüüpi alleel
embrüos ei avaldu.
Oogeneesi ajal sünteesitakse produkte, mis on folliikulirakkudele diferentseerumise
signaaliks – folliikulid diferentseeruvad dorsaalseks ja ventraalseks tüübiks.
Geeniproduktid ventraalsetest folliikulitest annavad signaali üle ootsüüdi ventraalsele
(kõhtmisele)
küljele. Signaal kandub membraanseoselisele retseptorvalgule (geeni toll
produkt), mis selle tagajärjel aktiveerub ning stimuleerib transkriptsioonifaktori sisenemist
.
Drosophila
embrüo arenemise kõige varajasemad etapid on kontrollitud ema poolt
sünteesitud faktorite kaudu. Hiljem aktiveeruvad selektiivselt embrüo geenid
- sügootiliste
geenide aktivatsioon.
Emapoolsete geenide
bicoid
ja nanos kontrolli all on ka embrüi gap geenid, mis määravad
segmentide piirkonna embrüos.
53. Organismi arengus osalevate geenide molekulaarne analüüs. Selgitage, milliste
meetoditega on võimalik tuvastada uuritava geeni rolli organismi arengus?
1) Drosophila
puhul on võimalik lokaliseerida kloneeritud geene, kasutades in situ
hübridiseerimist polüteenkromosoomidele
2) Spetsiifiliste RNA-de asukoha tuvastamine embrüo erinevatel arengujärkudel
3) Valkude lokaliseerimine huvipakkuvas kohas immunoloogilise reaktsiooniga
4) Geenide muteerimine
in vitro
ja nende mõju uurimine arenevas embrüos
54. Homeootilised geenid äädikakärbsel ja nende homoloogid eukarüootidel. Milliseid
väärarenguid põhjustavad mutatsioonid homeootilistes geenides?
HOMEOOTILISED GEENID – määravad selle, millised kehasegmendid arenevad, vallandades
nende poolt kontrollitavate geenide kaskaadse avaldumise
. Homeootilised geenid
kodeerivad heeliks-pööre-heeliks tüüpi transkriptsioonifaktoreid konserveerunud regiooniga
60 aminohappe ulatuses
.
Homeootilised geenid moodustavad ühes autosoomis kaks suurt klastrit:
1) Bithorax Complex
BX-C – 3 geeni. Mutatsioon: kärbse 3. rinnaku segment on
muutunud 2. segmendi sarnaseks – ilmneb teine paar tiivalaadseid moodustusi
2) Antennapedia Complex
ANT-C – 5 geeni. Mutatsioon: tundlad vahetuvad jalgadega
Kui diferentseerunud rakud on organiseerunud kindlal viisil, moodustavad nad organi.
Organite moodustumine õigetesse kehapiirkondadesse on väga täpselt geneetiliselt
kontrollitud. Nt: geen
eyeless
kodeerib homeodomääni sisaldavat transkriptsioonifaktorit,
mis lülitab tööle tuhandeid geene, et toimuks silma areng.
Äädikakärbse homeootiliste geenide homoloogid
(Hox
geenid) imetajatel:
Asuvad neljas erinevas klastris
HoxA, HoxB, HoxC
ja HoxD, mis paiknevad erinevates
kromosoomides. DNA järjestuse analüüs viitab sellele, et
4 Hox
geenide klastrit on tekkinud
algse klastri neljakordistumise tulemusena 500 – 600 miljonit aastat tagasi
.
Mutatsioonid
hox
geenides põhjustavad knock-out organismide arengut (tekivad kimäärid).
Nt: hox1b mutantidel ei ole närve, mis on vajalikud näoilmete väljendamise jaoks.
55. Mudelorganismid selgroogsete arengu uurimiseks. Selgitage hiire
knockout
mutantide konstrueerimist
Selgroogsete arengu geneetilist kontrolli aitab mõista selgrootute teadaolevate geenide
homoloogide isoleerimine ja iseloomustamine selgroogsetel organismidel.
Kui teatavat
arenguprotsessi mõjutav geen selgrootust organismist on identifitseeritud ja isoleeritud,
saab seda kasutada sondina tema homoloogide leidmiseks teistest organismidest.
Mudelorganismid:
o Hiir
, rott
o Konn
o Kana
o Sebrakala
Hiire
knockout
mutantide konstrueerimine:
o Huvipakkuva geeni funktsioon rikutakse ja viiakse inaktiveeritud geen hiire
genoomi normaalse geeni asemele
o DNA sisestatakse embrüonaalsetesse tüvirakkudesse ja kultuuris kasvatatud ES
rakud viiakse mõnda teise arenevasse embrüosse
o Sel viisil
saadud organismid on kimäärsed, sisaldades nii normaalseid kui ka
mutantseid rakke
o Seejärel
isoleeritakse järglased, kellel on mutatsioon kinnistunud
Kloneeritud
(uuritava organismi genoomist isoleeritud)
geenide abil on võimalik
konstrueerida
knockout
mutante. Sel juhul rikutakse kloneeritud geeni funktsioon ja
asendatakse normaalne geen hiire organismis rikutuga. DNA viimine hiire rakkudesse toimub
samal viisil nagu juba eelpool kirjeldatud. Defektse geeni asendumine normaalsega toimub
homoloogilise rekombinatsiooni teel. Heterosügootseid
knockout
mutatsiooni kandvaid hiiri
ristates saame homosügootsed järglased, kelle arengut jälgides on võimalik uurida mutantse
geeni mõju organismi arenguprotsessile.
56. Selgitage loomade kloonimise põhimõtet lammas Dolly näitel. Reproduktiivse ja
terapeutilise kloonimise eesmärgid
Organismi kloonimise tulemusena saadakse kaks või enam geneetiliselt identset isendit.
Sisuliselt on kloonid ka ühemunakaksikud, sest nad arenevad ühest viljastatud munarakust.
o Embrüokloonimine
: koduloomade puhul rakendatud varajase embrüo (2-8 raku
staadium
) rakkude lahutamist ning nendest identsete järglaste saamist
o Kloonimine lähtudes imetajate somaatilistest rakkudest
Lammas Dolly:
Kloonimisel lähtuti
doonori udararakust, kust võeti rakutuum. Teiselt utelt oli pärit
munarakk
, kust oli vaja kõrvaldada munaraku tuum ning asendada see doonorlamba
keharaku tuumaga.
Edasi stimuleeriti rakujagunemist ja arenes embrüo, mis viidi
kolmandasse lambasse
(asendusema)
. Sündis lammas Dolly, kes oli geneetiliselt identne
doonoriga, kellelt pärines tema DNA.
Reproduktiivne kloonimine:
Somaatilise raku tuum ühendatakse munarakuga, kust eelnevalt on tuum eemaldatud
,
kuid siin on
eesmärgiks saada arenevast diploidsest embrüost in vivo
täiskasvanud
organism, mis on doonori geneetiliselt identne koopia
Nt: väljasurevate või väljasurnud liikide kloonimine
Terapeutiline kloonimine:
Kasutatakse
peremehe somaatilisi rakke ES-rakkude populatsiooni moodustamiseks
kunstlikes tingimustes.
Somaatilise raku tuuma saab viia emasorganismi kui
doonorimunarakku, kust tuum on eelnevalt kõrvaldatud
. Ühendrakk on diploidne ning kui
ta areneb embrüoks, siis saab eraldada tüvirakke ja moodustada
in vitro
ES-rakkude liine,
et kasutada neid siis juba geneetiliselt identse materjalina tagasiviimisel
e.
transplantatsioonil
peremeesorganismi kudedesse või organitesse.
Nt: kudede regenereerimine, organite asendamine; aju või seljaaju kahjustuste
korrigeerimine; biokeemiliste defektide korrigeerimine
(diabeet)
.
57. Reproduktiivse kloonimisega kaasnevad probleemid. Kas teie arvates on mõtet
kloonida inimest?
Probleemid:
o Geneetilised ja epigeneetilised efektid kloonimisel
: kromosoomide aberratsioonid;
muutused geenide avaldumise mustris, mida mõjutab DNA metülatsioon ja
histoonide atsetülatsioon
o Kloonimisel ilmnevad arenguhäired
(hiire, sea ja kitse kloonidel vähem kui lamba ja
veise kloonidel)
: südame-veresoonkonna häired; organite patoloogia;
ebanormaalselt suur kasv sünnimomendil; kehaehituslikud defektid
o Erinevalt taimerakkudest tekivad
diferentseerunud imetajarakkude puhul probleemid
geenide avaldumise reprogrammeerimisega
o Pärast somaatilise raku geneetilise materjali ülekannet munarakku avalduvad
geenid, mis olid vaigistatud somaatilise raku tuumas; vaigistatakse geenid, mis olid
seotud raku diferentseerumisega
o Mittetäielik reprogrammeerimine viib embrüo arenguhäiretele
o Reprogrammeerimise efektiivsus varieerub sõltuvalt rakutüübist
Kas kloonida inimest?
Ei, sest liialt palju on defekte ning mutatsioone, mis võivad tekkida. Inimene ei pruugi üldse
eluvõimelisena sündida. See aitaks küll kindlasti uurida täpsemalt inimese geene, geenide
regulatsiooni, haiguste molekulaarseid tagamaid jne, kuid riskid kaaluvad üle õnnestumise
võimalikkuse.
Rahvastiku arv on maailmas ka juba piisavalt suur..
58. Selgitage antikehade spetsiifika geneetilist tagapõhja
Iga B rakk sünteesib ainult ühte spetsiifilist antikeha. Antikeha seondumine antigeeniga
stimuleerib B raku kiiret jagunemist, mille tulemusena tekib igast B rakust hulga sama
antikeha tootvaid plasmarakke. Maksimaalne antikehade produktsioon saavutatakse 2-3
nädala pärast. Antigeeni piirkonda, mida antikeha spetsiifiliselt ära tunneb ja seob,
nimetatakse epitoobiks. Antigeen-antikeha kompleksid degradeeritakse fagotsüütide poolt.
IgG ja IgM antikehad kutsuvad kohale komplemendivalke, mis degradeerivad võõrvalke
proteolüütiliselt ning hävitavad seente ja bakterite rakke nende lüüsimise teel. Kui võõras
antigeen on organismi kehast kõrvaldatud, pärsivad supressor T rakud antikehade tootmise.
Immuunsüsteemi iseloomustab tema spetsiifilisus.
Spetsiifilisus on tagatud tänu kolmele
põhikomponendile:
1) Koordineeritult töötavad spetsialiseerunud rakud
2) Antikehad ja T rakkude retseptorid, mis tunnevad ära võõraid substantse
3) Spetsiaalsed koesobivusantigeenid
, mis hõlbustavad rakkudevahelist
kommunikeerumist, võimaldavad immuunsüsteemi rakkudel ära tunda võõraid
substantse ning eristada organismile võõraid rakke omadest
Täiskasvanud inimese immuunsüsteemis osaleb ligi 10
12 valget vererakku ning 1015
spetsiaalset valgumolekuli
. Kui näiteks viirus või bakter satub inimese kehasse, tuntakse see
ära immuunsüsteemi rakkude poolt, mille tulemusena produtseeritakse kahte tüüpi valke –
antikehasid ning T rakkude retseptoreid, mis seonduvad spetsiifiliselt võõra substantsiga.
Antikehaga või T rakkude retseptoriga seondunud substantsi nimetatakse antigeeniks,
ning antigeeni, mis kutsub esile immuunvastuse, nimetatakse immunogeeniks
. Peaaegu
kõik bioloogilised makromolekulid (polüpeptiidid, polüsahhariidid, nukleiinhapped) on
võimelised toimima antigeenidena ning enamus neist käituma immunogeenidena.
Imetajate immuunvastust võib käsitleda kahekomponentsena:
1) Antikehade poolt vahendatud ehk humoraalne vastus
Humoraalse immuunvastuse korral
sekreteeritakse produtseeritud antikehad kehavedelike
ringlussüsteemi, kus nad seonduvad antigeenidega. Antikeha-antigeenkompleksid
haaratakse spetsiaalsete valgete vererakkude poolt ning hävitatakse
. Humoraalne
immuunsüsteem
kaitseb peamiselt organismi rakke võõraste rakkude (bakterid, seened) ja
vabade viiruspartiklite infektsiooni eest.
2) T rakkude poolt vahendatud ehk rakuline vastus
Rakulise immuunvastuse
korral sünteesitakse spetsiaalsete valgete vererakkude pinnale T
rakkude retseptoreid. Need retseptorid võimaldavad valgetel vererakkudel ära tunda ja
hävitada peremeesorganismi võõrmaterjaliga nakatanud rakke
. Seega kaitseb rakuline
immuunsüsteem organismi peamiselt viirusinfektsiooni vastu.
Spetsiaalsed rakud, mis osalevad immuunvastuses:
Immuunvastuses võõra substantsi vastu osalevad koordineeritult kõrgelt spetsialiseerunud
rakud
. Kõik need pärinevad mittediferentseerunud eellasrakkudest, mida nimetatakse
tüvirakkudeks (
stem cells
) ja mis asuvad luuüdis. Eristatakse kahte põhilist rühma:
1) B-lümfotsüüdid
, mida kutsutakse ka B rakkudeks, kuna nad valmivad luuüdis
Vastava signaali saabumisel
diferentseeruvad B lümfotsüüdid plasmarakkudeks, mis
produtseerivad antigeenidega seonduvaid valke
(antikehi)
ja B mälurakkudeks, mis on
sama antigeeniga uuesti kunagi hiljem kokkupuutudes võimelised väga kiiresti antikehi
produtseerima.
2) T-lümfotsüüdid
, mida kutsutakse ka T rakkudeks, kuna nende diferentseerumine
toimub tüümuses
Eraldi grupi moodustavad veel
fagotsüüdid (siia kuuluvad ka makrofaagid).
T-lümfotsüüdid diferentseeruvad nelja tüüpi rakkudeks:
1) Tsütotoksilised e.
tapjad T rakud (killer T cells
) – kannavad raku pinnal paiknevaid
antikehadega seonduvaid T rakkude retseptoreid ning hävitavad vastavaid
antigeene eksponeerivaid rakke
2) Abistavad T rakud
(helper T cells
) – stimuleerivad B- ja T-lümfotsüütide
diferentseerumist antikehasid tootvateks plasmarakkudeks ja tapjateks T
rakkudeks
3) Supressor T rakud
(supressor T cells
) – suruvad alla plasmarakkude tapjate T
rakkude aktiivsuse
4) T mälurakud
(memory T cells
) – „mäletavad“ antigeeni ning nende uuel kokkupuutel
sama antigeeniga moodustuvad kiiresti tapjad T rakud
Põhilised koesobivusantigeenid aitavad eristada võõrast omadest:
Omavahel mittesuguluses olevate indiviidide puhul ühelt teisele organi transplanteerimine
või naha ülekanne tavaliselt ebaõnnestub, kuna immuunsüsteem tõrjub transplanteeritud
koed kõrvale. Samas identsete kaksikute puhul transplantatsioon üldjuhul õnnestub. Samuti
võib samal indiviidil nahka ühest kohast teise istutada.
Võõraste kudede eristamine
organismi enda kudedest toimub tänu koesobivusantigeenidele
. Võtmeroll selles protsessis
on põhilise koesobivuskompleksi
(MHC)
valkudel. Inimesel nimetatakse MHC valke HLA
antigeenideks, kuna nad
avastati leukotsüütide pinnal. HLA antigeenid on kodeeritud suure
geenide klastri poolt, mida nimetatakse
HLA
lookuseks ja mis paikneb 6-ndas kromosoomis.
HLA
lookuse geenide polümorfism on väga suur, mis väljendub selles, et mõnel geenil on
kirjeldatud kuni sada või enam alleeli
. Seega on tõenäosus, et kaks omavahel
mittesuguluses olevat indiviidi kannaksid kõigi
HLA
geenide samu alleele, ülimalt väike ja
sobivate organidoonorite leidmine komplitseeritud
.
HLA
geenid kodeerivad kolme erinevat klassi valke:
1) Klassi I kuuluvad
HLA
geenid kodeerivad transplantatsiooni antigeene. Nimetus
tuleneb sellest, et just need valgud vastutavad transplanteeritud kudede
tagasitõrjumise eest. Neid nimetatakse
ka MHC klass I valkudeks. Need valgud on
oma
olemuselt glükoproteiinid, mis on ankurdunud rakumembraanile, nii et
antigeenne determinant ulatub rakust välja
. Transplantatsiooniantigeenid esinevad
praktiliselt kõigil rakkudel ning aitavad killer-rakkudel (tapjad T rakud) eristada
võõraid rakke omadest
. Sellesse klassi kuuluvad valgud võivad seonduda küllaltki
erinevate antigeenidega teiste rakkude pinnal ja esitada võõraid antigeene
eksponeerivaid rakke killeritele
2) Klassi II kuuluvad
HLA
geenid kodeerivad valke, mis asuvad peamiselt
B-lümfotsüütide ja makrofaagide pinnal
(need on rakud, mis interakteeruvad helper
T rakkudega). Need on heterodimeersed valgud, mis koosnevad kahest
polüpeptiidist. Polüpeptiidide polümorfsed N-terminaalsed domeenid
rakupinnal
moodustavad vao, kuhu võib seonduda küllaltki lai valik erinevaid antigeene
3) Klassi III kuuluvad
HLA
geenid kodeerivad komplement-valke, mis interakteeruvad
antikeha-antigeen kompleksidega ning aitavad neid proteolüütiliselt degradeerida
.
Komplement-valgud tapavad ka võõra bioloogilise materjaliga nakatunud rakke,
lõhkudes nende rakkude membraane
59. Antikehade klassi ümberlülitumise ja alleelse välistamise molekulaarsed
mehhanismid. Monokloonsete antikehade saamine
Antikehad grupeeruvad viide klassi: IgA, IgD, IgE, IgG ja IgM
. Antikeha funktsiooni ning
kuuluvuse ühte neist klassidest määrab ära raskete ahelate konstantsete regioonide
struktuur e. antikeha efektori funktsiooniga domääni struktuur
.
o Kui arenevates B-lümfotsüütides algab antikehade süntees, on kõik raske ahela
konstantse ala geenisegmendid
C
H
olemas – sünteesitakse IgM tüüpi raske ahelaga
antikehasid
, mis sisaldavad konstantse regioonina CH
µ
geeniprodukti
o Mõne aja pärast hakkavad osa plasmarakke tootma ka teistesse klassidesse
kuuluvaid antikehi, mis sisaldavad rasket ahelat IgA, IgD, IgE või IgG –
ümberlülitumine
o IgM ja IgG tootvates rakkudes toimub nii geeni
CH
µ
kui ka geeni CHδ
transkriptsioon ning RNA splaissingu käigus kõrvaldatakse üks järjestustest
o Üleminekul teistesse klassidesse kuuluvate antikehade tootmisele toimuvad
ümberkorraldused genoomis, mille käigus osa
C
H
segmentidest deleteeruvad ning
avaldub see raske ahela konstantset regiooni kodeeriv järjestus, mis külgneb vahetult
geeniga
Alleelne välistamine:
Iga plasmarakk toodab ainult ühte tüüpi antikeha
. Kuigi raku genoom on diploidne,
toimuvad
geneetilised ümberkorraldused ainult ühe alleeli ulatuses – alleelne välistamine.
Alleelne välistamine
toimub raskete ning kergete ahelate geenisegmentide puhul
üksteisest sõltumatult
. Kuna iga rakk produtseerib ainult ühte kindlat antikeha, mis seondub
ainult ühe epitoobiga, võimaldab see toota monoklonaalseid antikehi. Monoklonaalseid
antikehi kasutatakse valkude tuvastamiseks ja lokaliseerimiseks.
Monokloonsete AK-de saamine:
MONOKLOONSED ANTIKEHAD – kasvajaraku ja B-lümfotsüüdi hübriidimisel saadakse
sellised hübriidsed immortaliseeritud rakukloonid, mis on
in vitro
tingimustes võimelised
tootma vaid üht kindlat tüüpi antikeha, mis on ülimalt spetsiifiline mingi kindla aine suhtes
ja mis seondub vaid ühe epitoobiga – sellega, mille vastu ta on saadud
.
1) Katselooma
(hiir)
immuniseerimine meid huvitava antigeeniga (mingi valk,
hormoon, ensüüm; immuniseerida vôib ka tervete rakkudega, näit kasvajarakkudega,
et saada kasvajarakule iseloomulike antigeenide vastu antikehi jne)
2) Immuniseeritud hiire põrnarakud
(B-lümfotsüüdid)
liidetakse (hübridiseeritakse)
kasvajarakkudega
3) Hübridoomrakkude selektsioon HAT-selektiivsöötmel
. Selektiivsöötme põhimõte:
kasvajarakud, mis ei hübridiseerunud põrnarakuga, surevad, sest nad on defektsed
DNA sünteesi varutee eest vastutava ensüümi - HPGRT - suhtes
.
DNA sünteesi peatee
(
de novo süntees) on aga blokeeritud selektiivsöötmes oleva aminopteriini poolt.
Seetõttu antud söötmes saavad ellu jääda ainult hübridoomrakud, mille üks vanem
oli põrnarakk
(
sealt tuleb hübridoomile puuduv ensüüm HPGRT, ning ta kasutab DNA
sünteesiks varuteed, milleks vajalikud ained - hüpoksantiin ja tümidiin, on söötmesse
juurde lisatud)
4) Antikehade sünteesi testimine
– tuleb üles leida see rakukloon, mis sünteesib
tõepoolest teid huvitavaid antikehi
5) Soovitud antikehasünteesiga rakuklooni rekloneerimine, osa materjali
külmutamine
6) Hübridoomi kasvatamine masskultuuris või hiire kõhuõõnes
astsiitkasvajana
7) Antikeha puhastamine, kasutamine soovitud otstarbeks
60. Päritavad ja omandatud immuunsüsteemi haigused.
Selgitage, kas ja kuidas on neid
võimalik ravida?
Päritavad immuunsüsteemi haigused:
o Autoimmuunhaiguste puhul toodab indiviidi organism antikehi tema enda
antigeenide vastu
– kahjustavad organismi enda kudesid ja organeid
Nt:
Juveniilne diabeet – antikehad ründavad pankreases insuliini tootvaid beetarakke.
Insuliini puudumine põhjustab
pimedaks jäämist, haigestuvad ka süda ja neerud. Juveniilset
diabeeti ehk tüüp I diabeeti pole täielikult võimalik ravida, kuid on võimalik sümptomeid
leevendada. Selleks manustatakse haigele insuliini, pankrease transplantatsioon, neerude
transplantatsioon.
o Kombineeritud immuundefektsuse sündroom
Nt:
ADA-defektsuse tagajärjel akumuleerub organismi desoksü-ATP ning tema liigkõrge
kontsentratsioon on T helper-rakkudele toksiline
. Ilma T helper-rakkudeta ei toimu B- ja T-
lümfotsüütide diferentseerumist antikehi tootvateks plasmarakkudeks ega T
killer-rakkudeks.
Haigust on võimalik ravida, kasutades geeniteraapiat või luuüdi siirdamist
o X-liitelise agammaglobulineemia
puhul pole indiviidi organism võimeline sünteesima
antikehi
. X-liiteline haigus, kus B rakkude diferentseerumist ei toimu. Haigetel toimib
T rakkude poolt vahendatud immuunvastus normaalselt,
nad on kaitstud
viirusinfektsioonide vastu, kuid väga tundlikud bakteriaalsete ja seeninfektsioonide
suhtes
. Tänu luuüdi transplantatsioonidele (doonori luuüdi sisaldab normaalseid
tüvirakke, mis on võimelised diferentseeruma ja tootma antikehi)
on tänapäeval
võimalik seda haigust ravida
Omandatud immuunsüsteemi haigused:
Immunodefektsus võib kujuneda välja ka loote arengu käigus või sünnijärgselt toksiliste
kemikaalide või patogeensete mikroorganismide toimel.
Nt:
DiGeorge sündroomi puhul ei ole organismi loote arengu käigus saadud kahjustuste
tulemusena arenenud normaalselt funktsioneerivat tüümust, mistõttu
haige organismis ei
moodustu T rakke
. Haigust pole võimalik ravida, kuid haigel peab silma peal hoidma, et oleks
võimalik kiiresti ravida kaasuvaid defekte
(nt bakteriaalsed infektsioonid, südame
abnormaalsused jne)
.
Omandatud immunodefektsuse sündroom (
acquired immunodefeciency syndrome
) AIDS
on põhjustatud HIV (
human immunodeficiency virus
) poolt. HIV nakatab ja hävitab T
helperrakke – organismis väheneb T ja B lümfotsüütide diferentseerumine killer T rakkudeks
ja plasmarakkudeks. Immuunvastus nõrgeneb järk-järgult, kuni organism ei suuda enam
toime tulla infektsioonidega ja seejärel haige sureb
. Haigusele täielik ravi puudub, kuid on
olemas erinevaid ravimeid, mis leevendavad sümptomeid ja võimaldavad patsiendil elada
täisväärtuslikku elu. Ravi kestab terve elu.
61. Biorütmide uurimine ja biorütme mõjutavad geenid
Biorütmid ehk bioloogilised rütmid on organismide füsioloogiliste, emotsionaalsete,
intellektuaalsete ja sotsiaalsete omaduste ja võimete korrapärane perioodiline korduvus
mingis ajaühikus.
Biorütmide geenid
o Tsirkaadirütmid (ööpäevarütmid) seentel ja bakteritel
– bakteritest on
tsirkaadirütme kirjeldatud
tsüanobakteritel lämmastiku fikseerimist ja fotosünteesi
määravate geenide avaldumisel
.
Tsüanobakterite
bioloogilise kella toimimise mudel põhineb keskkonnainformatsiooni
ülekandel ööpäevaselt histidiini proteiinikinaasilt CikA signaali ülekandeahela väljundahela
kaudu tsüanobakterite adaptiivsele sensorile SasR
. Selles protsessis stimuleerivad teatud
valgud SasA-valgu autofosforüülimist. Viimane kannab fosforüülgrupi üle
regulaatorvalgule SasR, millega see regulaator mõjub omakorda kogu genoomi
avaldumisele. Õhtupimedas hakkab toimima valk KaiB, mis seondudes KaiC-ga, põhjustab
SasA toime blokeerimise ja hiljem kompleksi dissotseerumise algkomponentideks
.
Seentel avalduvad tsirkaadirütmid nt
filamentse seene Neurospora sp. a
seksuaalsete
spooride rütmilise moodustumise käigus
. Nimetatud nähtust kontrollib geen frq
.
o Äädikakärbse perioodilisuse geenid –
Drosophila
X kromosoomis paikneb geen per
(
period
), mis on funktsionaalselt ekvivalentne Neurospora geenile frq. Äädikakärbse
käitumises täheldatakse 24-tunnist rütmi, mida nimetatakse
tsirkaadseks rütmiks.
Tsirkaadne rütm kajastub näiteks kärbse valmikute nukust koorumisel.
Äädikakärbsed
on kõige aktiivsemad koiduajal
. Kärbeste aktiivsust saab mõõdeta nende
liikumisaktiivsuse kaudu klaastorus. Klaastoru on pimedas, ning ühest kohast läbib
teda kitsas punane valguskiir, mida kärbsed ei näe. Iga kord, kui kärbsed valguskiire
läbivad, see registreeritakse
. Geenil per
on kirjeldatud mitu alleeli: alleeli per0 puhul
on isendite käitumine arütmiline, alleeli
per
S puhul lüheneb tsükkel 19 tunnile ning
alleeli
per
L puhul pikeneb 29 tunnile. per lookus kontrollib nukkude koorumise aega.
Tavaliselt toimub see hommikul (päikesevalguse käes kuivavad valmiku tiivad
kiiremini ning ta saab varem lendama hakata).
per
0 mutantide puhul toimub
valmikute nukust koorumine suvalisel ajal ning alleelide
per
S ja perL puhul muutub
tsükkel vastavalt eelpoolkirjeldatule
. Geen per
kontrollib ka kärbeste
paaritumisrituaali
. Isased kärbsed esitavad pulmatantsu, mille võib jagada mitmeks
etapiks. Liigiti on tants erinev. Põhimomendid on aga järgmised: isane läheneb
emasele nurga all ning liigub emase tagakeha suunas; ta tõstab oma tiivad 90° üles ja
vibreerib nendega, tekitades tiibadega heli, mis peaks emast stimuleerima; seejärel
puudutab ta emase genitaale ja üritab emast viljastada. Alati ei pruugi ta eesmärki
saavutada ning siis tuleb rituaali korrata. Tiibade vibratsiooni teel tekitatud “laul” on
liigiti erinev, erinedes pulsside intervalli poolest (15 kuni 100 millisekundit). Tiibade
vibreerumises on teatav rütm – ääikakärbsel toimub see 60-sekundiliste perioodide
tagant ning vibratsioonikiirus on erinevatel etappidel erinev (algab ühtlase suminaga,
siis järgneb kiirem vibreerimine, seejärel on pulsside vaheaeg pikem, siis aga jälle
kiirem). Geen
per
kontrollib perioodide pikkust. Alleeli perS puhul väheneb see 40
sekundini ning alleeli
per
L puhul pikeneb 90 sekundini. Nende alleelide osalust
vibratsiooniperioodide pikkuse kontrollil on näidatud ka, kasutades molekulaarseid
meetodeid. Geen
per
kodeerib transkriptsioonifaktorit nimetusega PER. See valk
transporditakse rakutuuma ning nii
per
geeni mRNA kui ka PER valgu enda tase rakus
kõiguvad rütmiliselt. PER valku on leitud eeskätt aju rakkudes, kuid ka teistes
rakkudes. Kuna PER valgul pole DNA-ga seondumise domääni, mõjutab ta
transkriptsiooni kaudselt, interakteerudes teiste transkriptsioonifaktoritega. PER
valgu puhul esineb autoregulatsioon. PER represseerib teda kodeeriva geeni
transkriptsiooni. Selle tulemusena väheneb rakkudes vastava mRNA hulk ning
seejärel ka valgu hulk. Kui PER valgu hulk väheneb, toimub geeni derepressioon ning
mRNA süntees algab uuesti (tavaliselt toimub see koiduajal). PER valk toimib
repressorina, komplekseerudes valguga
TIM (timeless
), mille tase on samuti kõrge öö
saabudes ja langeb hommikuks. Leiti, et valgus inaktiveerib valgu TIM ning see
põhjustab ka TIM-PER kompleksi lagunemist. Kui kärbseid öösel perioodiliselt
valgustada, läheb nende elurütm sassi.
o Biorütmid taimedel
– taimede vegetatiivsetel osadel on ainuvalitsevad arusaadavalt
tsirkadiaansed rütmigeenid, mis avalduvad päeval ja on välja lülitatud öösel. See rütm
säilib ka siis, kui hoida taimi pikemalt pimedas.
Taimede biorütmi uurimise
mudelorganismiks on olnud müürlook
. Transgeensetes taimedes, kuhu on viidud
näiteks helendumist määrav lutsiferaasigeen, on näidatud biorütmide toimumist
koos taime lehtede orientatsiooniga maksimaalse valguse suunas
. Üheks
võtmegeeniks on klorofülliga seonduvat valku kodeeriv geen
. Taimsed
regulaatorained fütokroomid aktiveeritakse punase lainepikkusega valguse poolt.
Aktiveeritud fütokroomid lülitavad sisse biorütmide toimimise eest vastutavad
geenid, milleks on tavalislet transkriptsioonifaktorieid määravad geenid.
Moodustunud transkriptsioonifaktorid seonduvad omakorda märklaudgeenide
regulatoorsetesse piirkondadesse, muutes vastavate geenide avaldumist.
Valgusega seotud taimevalkude füsioloogiliseks vastuseks on taime
fotomorfogenees
(lehtede ja varte areng ning kolroplastide moodustumine)
o Bioloogilised rütmid loomadel
– hiirte puhul on uuritud tsirkaadset rütmi üsna
täpselt.
Kui hoida hiiri pimedas, jälgivad nad seda rütmi veidi üle 23 tunni pikkuste
intervallide kaupa. Mutatsiooni
clock
puhul pikeneb rütm homosügootidel 27-28
tunnini, heterosügootidel 25 tunnini. Pärast kahenädalalist pimedas hoidmist kaob
homosügootsetel mutantidel rütm täiesti
. Vastavat mutatsiooni sisaldav geen on
lokaliseeritud 5-ndasse kromosoomi
62. Tooge näiteid ühe geeni mutatsioonidest, mis mõjutavad äädikakärbse käitumist
Geeni
per (period)
mutatsiooni puhul muutub äädikakärbse tsirkaadne tsükkel
arütmiliseks
: alleeli per
S puhul lüheneb tsükkel 19 tunnile, alleeli perL puhul pikeneb tsükkel
29 tunnile.
per
lookus kontrollib nukkude koorumisaega – tavaliselt kooruvad valmikud hommikul. Per0
mutantide puhul toimub valmikute nukust koorumine suvalisel ajal. Alleelide
per
S ja perL
puhul tsükkel vastavalt kas lüheneb või pikeneb.
Tiibade vibratsiooni teel tekitatakse pulmalaul
: alleeli per
S puhul lüheneb periood 40
sekundini ning alleeli
per
L puhul pikeneb 90 sekundini. Tavaliselt on see 60-sekundilise
perioodi tagant.
Mutatsioonid geenis
fru
põhjustavad homoseksuaalsust. Fru geeni isaste splaissinguvariant
muudab emased kärbsed homoseksuaalseks.
63. Neuroloogilised haigused inimesel, mis on seotud ühe geeni mutatsiooniga
o Alkoholism – mutatsioonid alkoholi dehüdrogenaasi geeni alleelis
o Fenüülketonuuria – autosoomne retsessiivne tunnus, põhjustatud fenüülalaniini
hüdroksülaasi (PAH) defektsusest
o Huntingtoni tõbi – mutantset
HD
alleeli kandvatel haigetel hävivad järk-järgult
peamiselt aju keskosas asuvad neuronid, mis paiknevad ajutuumades. Need
piirkonnad kontrollivad liikumise koordinatsiooni
o Jne
64. Käitumuslikud geenidoosimutandid: kõrvalekalded inimese kromosoomide arvus
o Downi sündroom
– 21. kromosoomi trisoomia; madal IQ, ahvivagu, mongoliidsed
silmad, väike kasv
o Klinefelteri sündroom
– XXY mehed; raskusi eneseväljenduses, keelte omandamises,
düsleksia, depressioon, ei suuda toimida stressisituatsioonis
o XYY mehed
– kontrollimatum demperament, agressiivsus, hüperaktiivsus
o XXX naised
– käitumuslikult üsna sarnased normaalsete XX naistega, nende
verbaalne IQ on keskmisest veidi madalam, suhtlemisraskused
o Turneri sündroom
– X0 naised; kuulmishäired, kehv ruumitaju, raskused täpseid
liigutusi nõudvate töödega, raskusi matemaatikaga
65. Kvantitatiivsete käitumistunnuste uurimine katseloomadel
Kvantitatiivsed tunnused
on need, mida saab kaaluda ja mõõta. Nad on määratud paljude
geenide üheaegse mõjuga või koostoimega.
Kui tunnust määravad paljud geenid, siis on ka
keskkonnategurite toime vastava tunnuse avaldumisele suurem
. Enamik organismide
füüsilisi, aga ka käitumistunnuseid on komplekssed. Erit just komplekssete tunnuste puhul on
võimalik, et suur osa organismi geneetiliste faktorite avaldumisest jääb kasutamata.
Seepärast sõltub organismi geneetilise potentsiaali avaldumise ulatus mõnes osas indiviidide
elukeskkonnast.
o Uuritakse hiire jooksmist rattal
. Hiir jälgib ööpäevast rütmi veidi üle 23 h pikkuste
intervallide kaupa
o Süüria hamstril kirjeldati mutatsiooni geenis
tau
: homosügootidel lühenes 24h-ne
tsükkel 20h-seks; muutus hormoonide sekretsiooni rütm; paljunemistsükkel ei
sõltunud enam aastaajast
o fosB mutantsed hiired ei imeta oma järglasi
o Rottide õppimisvõime uurimine labürintides, vesivannis platvormi leidmisel
o Ristates erinevaid koeratõuge, on võimalik jälgida, millised käitumisjooned on
dominantsed, millised retsessiivsed
o Erinevatel koeratõugudel esinevad erinevad haigused
(narkolepsia, pimedaks
jäämine)
– võimaldab uurida haiguste geneetilist tausta ka inimesel
66. Intelligentsuse ja isiksuse omaduste geneetilise faktori osakaalu uurimine inimesel
Intelligentsust mõjutavad nii indiviidi genotüüp kui ka keskkonnategurid
. Intelligentsust
kirjeldab erinevate vaimsete võimete kogum:
1) Sõnade defineerimine ning mõistmine
2) Verbaalse mõtlemise kiirus
3) Matemaatiline mõtlemine
4) Ruumiline mõtlemine
5) Meeldejätmine ning informatsiooni taastamine
6) Objektidevahelise sarnasuse ja erinevuse hindamine
7) Seaduste, põhimõtete või kontseptsioonide formuleerimine probleemide
lahendamiseks ning erinevate olukordade hindamiseks
Keskmine IQ
(intelligence quotient
) – keskmine intelligentsuse kvoot on 100. IQ arvutatakse
sel viisil, et jagatakse indiviidi vaimne iga tema tegeliku vanusega ning korrutatakse
100-ga. Kui näiteks 10-aastase lapse testi tulemused vastavad 12-aastase keskmistele testi
tulemustele, on selle lapse IQ 120.
Lihtsam on identifitseerida ühe geeni mutatsioone, mis põhjustavad silmatorkavaid
neuroloogilisi kahjustusi ning sellega kaasnevaid muutusi käitumises.
Paljud neuroloogilised haigused on komplekssed:
o Sümptomite väljakujunemises osalevad korraga mitmed erinevad geenid ning
erinevad keskkonnategurid
o Näiteks võib tuua skisofreenia ning maniakaalse depressiooni, millele on küll
geneetiline soodumus, kuid haigus avaldub teatavate keskkonna mõjutuste toimel,
näiteks väga tugeva stressi korral
67. Evolutsiooniteooria kujunemine. Selgitage Darwini evolutsiooniteooriat
1859
– Darwini “The Origin of Species...“
(“Liikide tekkimine”)
Idee eluslooduse evolutsioonist pakuti välja sadakond aastat enne Darwini teose
avaldamist
. Tekkis tugev vastasseis teoloogilise maailmapildi näol:
o Katstrofismi doktriin
o Tüpoloogiline mõtlemine e. essentsialism
Viiskümmend aastat enne Darwinit avaldas
Lamarck teooria liikide tekkest: Liikide areng
uuteks liikideks toimus graduaalselt, organismide elu jooksul omandatud tunnuste
kumuleerumise tagajärjel
. Lamarck ei arvestanud looduslikku valikut, mis viib
vähemkohastunute hukkumisele
.
Darwini evolutsiooniteooria:
Aastatel 1831-1836 sooritas Darwin reisi Lõuna-Ameerikasse, kus ta kogus põhilised
andmed, mis said aluseks evolutsiooniteooriale
:
1) Darwin kirjeldas Lõuna-Ameerikas väljasurnud organismide luid, mis sarnanesid
kaasajal elanud organismide luustikega
2) Looduslikud tingimused Maal on olnud pidevas muutumises – Darwin leidis
Andidest kunagi meres elanud organismide jäänuseid
3) Darwin kirjeldas Galapagose saartel 13 vindi liiki, mis olid sarnased Lõuna-Ameerika
läänerannikul elavate vintidega
Vindid erinesid keha suuruselt ja noka kujult ning asustasid ökoloogiliselt erinevaid piirkondi.
Darwin oletas, et vindid tulid Galapagose saartele mandrilt ning seejärel populatsioon
mitmekesistus, asustades erinevate looduslike tingimustega piirkondi
.
Evolutsiooniteooria väljaarendamiseks
sai Darwin inspiratsiooni Malthuse ideedest:
toiduressursid on limiteeritud, samas suureneks inimpopulatsioon geomeetriliselt, kui
seda ei piiraks konkurents
.
Darwin:
organismide paljunemine populatsioonisiseselt toimuks eksponentsiaalselt, kui
seda ei piiraks ressursside vähesus. Konkurentsi tingimustes jäävad ellu vaid vähesed.
Darwini loodusliku valiku teooria järgi tekib populatsiooni paljunemise käigus uusi
variante ning edukamad variandid asendavad vähem edukamad variandid.
Darwini evolutsiooniteooria muutis oluliselt seni lääne ühiskonnas valitsenud mõttemaailma.
See tõrjus kõrvale katastrofismi idee ning liikide loomise idee. Darwini teooria lükkas kõrvale
antropotsentristliku mõttemalli nagu oleks inimliik universumi keskpunkt. Ka osutus Maa
palju vanemaks kui piibliõpetuse järgi pakutud 4000 kuni 6000 aastat. Tüpoloogiline
mõtlemine asendus populatsioonide tasemel mõtlemisega ning enam ei käsitletud
organismide arengut eesmärgipärasena.
Darwini teooria ei käsitlenud mehhanisme, kuidas kujuneb geneetiline variantsus, mil viisil
päranduvad tunnused järglastele ning kuidas populatsiooni geneetiline koostis aja jooksul
muutub. Kuigi Darwini evolutsiooniõpetus käsitles pärilikkuseühikute – gemmulate –
kandumist järglaskonda, sarnanes ettekujutus gemmulate muutlikkusest Lamarcki poolt
esitatud organismi elu jooksul omandatud tunnuste ideele. Alles Darwini õpetuse
kombineerimine Mendeli pärilikkuse teooriaga võimaldas luua kaasaegse pildi looduslikes
populatsioonides toimuvast evolutsiooniprotsessist.
68. Selgitage Hardy-Weinbergi populatsiooni geneetilise tasakaalu seadust. Millisel
juhul see rakendub?
POPULATSIOONIGENEETIKA – uurib alleelide sagedusi populatsioonides
Hardy-Weinbergi populatsiooni geneetilise tasakaalu seadus:
Hardy ja Weinberg leidsid seaduspärasuse, kuidas
alleelide sagedus populatsioonis
võimaldab ennustada genotüüpide sagedust juhuslikult ristuvas populatsioonis
.
Kui oletada, et populatsioonis on kaks alleeli
A
ja a ning alleel A esineb sagedusega p ja
alleel
a
sagedusega q, siis saab nende alusel arvutada genotüüpide sagedused.
Genotüüpide sageduse arvutamine:
AA
genotüübi tõenäosus populatsioonis on alleeli A sageduse korrutis p x p = p2. Samal
põhimõttel arvutatakse ka genotüübi
aa
tõenäosus populatsioonis (q x q = q2).
Heterosügootide
Aa
moodustumise tõenäosus on 2pq, sest on võimalik, et alleel A
munarakust kombineerub alleeliga
a
seemnerakust ja vastupidi.
Genotüüpide sagedust populatsioonis saab esitada valemi näol: (
p
+ q)2 = p2 + 2pq + q2 = 1
H-W seaduspärasus kehtib suurte populatsioonide puhul, kus populatsioonisiseselt toimub
vaba ristumine
– panmiktiline populatsioon. Panmiktilises populatsioonis ei toimi
evolutsioonitegurid
(mutatsioonid, valik, migratsioon) ning tasakaalulised alleeli- ja
genotüübisagedused säilivad muutumatuna põlvkonnast põlvkonda
.
69. Populatsioonis esinevate alleeli- ja genotüübisageduste muutuste põhjused
Alleeli- ja genotüübisageduste muutuste põhjused:
1) Ristumine pole juhuslik
o Omavahel ristuvad sugulased või fenotüübilt sarnased isendid – selle
tulemusena kasvab homosügootide arv populatsioonis
2) Erineva genotüübiga isendite edukus järglaste saamisel on erinev
3) Toimub populatsioonisisene eraldumine
(näiteks geograafiline isolatsioon)
4) Migratsioon – osa populatsioonist liigub uude elukohta
70. Genotüüpidele rakenduv suhteline kohasus ja selektiivne surve populatsioonis.
Millisel juhul on selektiivne surve genotüübile maksimaalse väärtusega? Tooge
näide
Organismi kohasuse puhul näitab
w
= 1, et antud isend on andnud ühe järglase; w = 2, kui on
kaks järglast
jne. Enamasti kasutatakse matemaatilist mõistet suhteline kohasus, kus
kohasuse maksimaalne väärtus
w
= 1,0.
w
= 1 – s; w– suhteline kohasus, s – selektiivne surve
Erinevatele genotüüpidele rakendub erinev selektiivne surve
s
. Kohasuse ja selektiivse surve
väärtused on vastandlikud.
Mida tugevam selektiivne surve rakendub antud genotüübile,
seda väiksem on kohasuse väärtus. Äärmuslik olukord, kus genotüübi kohasuse väärtus
w
on null
(steriilsus)
ning selektiivne surve s väärtus üks: teatava genotüübiga indiviidid ei ela
nii kaua, et jõuaksid paljunemisikka.
Nt: Tay-Sachs tõbi, Lesch-Nyhani sündroom, Duchenne
lihasdüstroofia.
71. Miks on mõne kahjuliku alleeli sagedus populatsioonis oodatust kõrgem? Selgitage
mehhanisme ja tooge näide
Mõne kahjuliku alleeli sagedus populatsioonis on oodatust kõrgem juhul, kui valik toimub
retsessiivse alleeli vastu
. Kui selektsioon toimub retsessiivse alleeli vastu, hakkab selle alleeli
sagedus populatsioonis langema.
Valik retsessiivse alleeli vastu toimub homosügootses
olekus. Kui retsessiivne alleel osutub võrreldes dominantsega kasulikumaks, hakkab selle
sagedus populatsioonis suurenema. Võrreldes vastuselektsiooniga retsessiivsele alleelile
toimib positiivne selektsioon retsessiivsele alleelile oluliselt kiiremini, kuna sel juhul
kõrvaldatakse dominantne alleel.
Nt:
pöidlaküüdi efekt – kuna erinevate geenide erinevate alleelide puhul on neile rakenduv
selektiivne surve erinev, siis nende geenide aheldumise korral ei pruugi kahjulik alleel
populatsioonist elimineeruda
. Tsüstilist fibroosi (CF)
põhjustav alleel püsib valge rassi
populatsioonis heterosügootses kõrge sagedusega
(üks indiviid 25-st on CF-i suhtes
heterosügootne)
. Eeldatavast kõrgemat sagedust aitab selgitada nn. “pöidlaküüdi efekt”
(“
hitchhiker effect
”), kus jälgitav alleel on tugevalt aheldunud teise geeni alleeliga, mis
suurendab indiviidi kohasust populatsioonis
. Pöidlaküüdi efekti põhjustab CF alleeli kandvate
indiviidide kõrgem vastupanu tüüfusele
(tüüfusetekitaja nakatab heterosügootide rakke
väiksema edukusega)
.
72. Loodusliku valiku kolm viisi: stabiliseeriv, suunav ja lõhestav valik. Tooge näiteid
Stabiliseeriv ehk tasakaalustav valik:
Selektsiooni kaudu on eelistatud optimaalne fenotüüp, kõik äärmused elimineeritakse
.
Homosügootidel on võrreldes heterosügootidega väiksem kohasuse väärtus, kuid samas
säilivad populatsioonis mõlemad alleelid tänu heterosügootide suuremale kohasusele.
Nt:
Punaste vererakkude sirprakset aneemiat põhjustava alleeli püsimine malaaria
levikuga piirkondi asustavas inimpopulatsioonis
. Heterosügoodid, kes kannavad ühte
normaalset β globiini alleeli ja teist mutantset, ei haigestu malaariasse – eelisolukord
võrreldes ainult normaalseid alleele kandvate homosügootsete indiviididega, kes on
malaariaparasiidile vastuvõtlikud.
Näiteks, kui vastsündinud beebide kaal erineb oluliselt optimaalsest väärtusest,
vähenevad nende võimalused ellu jääda.
Suunav valik:
Selektsiooni kaudu on eelistatud üks äärmuslik fenotüüp ning selle tulemusena muutub
populatsiooni keskmine kindlas suunas
.
Leiab aset siis, kui keskkonnas toimuvad pidevad muutused.
Nt:
hobuse kehaehituse ja suuruse muutumine võrreldes tema eellastega; suunav valik
rakendub populatsioonile ka
tõu- või sordiaretuse käigus (nt: kultuurtaimed)
.
Lõhestav valik:
Eelistatud on äärmused ning lõpptulemusena jaotub populatsioon kaheks või enamaks
alampopulatsiooniks, mis erinevad üksteisest teatava tunnuse suhtes oluliselt.
Ristumine toimub eelistatult liinisiseselt, mis erinevad üksteisest teatava tunnuse suhtes
oluliselt ja nii muutuvad kaks alampopulatsiooni teineteisest isoleerituks
.
Nt: Ameerikas elav grisli ning Euroopas elav pruunkaru on isoleeritud ning arenenud nii, et
nad vabalt ristuda enam ei saa.
Lõhestavat valikut tõestati ka katseliselt:
Drosophila
populatsioonist valiti välja isendid,
kellel oli keskmistega võrreldes kas oluliselt rohkem või vähem harjaseid ning lasti neil
omavahel ristuda. Populatsioon lahknes kaheks alampopulatsiooniks, kus olid kas väheste
harjastega isendid või paljude harjastega isendid. Katsetulemused näitasid, et ristumine
toimus eelistatult liinisiseselt ja nii muutusid kaks alampopulatsiooni teineteisest
isoleerituks
.
73. Selgitage inbriidingu mõju populatsioonile. Tooge näiteid ja selgitage geneetiliselt
inbriidingu kahjulikku mõju
Populatsioonisisese vaba ristumise piirangud võivad viia inbriidingule, mille kõige
äärmuslikum võimalus on iseviljastumine taimedel.
Inbriidingu tulemusena jääb alleelide
sagedus populatsioonis küll samaks, kuid muutub genotüüpide sagedus: tõuseb
homosügootsuse aste ja väheneb heterosügootsuses aste. Inbriidingu tulemusena võivad
homosügootsesse olekusse sattuda ka muidu harva esinevad geneetilisi haigusi
põhjustavad retsessiivsed alleelid.
Nt:
Albinismi sagedus varieerub 1:10000 Põhja-Euroopas kuni 1:30000 Lõuna-Euroopas, kuid
USA-s Arizona osariigis elavatel Hopi indiaanlastel esineb albinism sagedusega 1:200. Hopi
indiaanlaste populatsioon on suhteliselt isoleeritud ning seal on sageli sugulastevahelisi
abielusid. Albiinod ei talu palju päikest, mistõttu pole neist põllu- ja muudel välitöödel suurt
kasu.
74. Selgitage alleelisageduste juhuslikku muutust populatsioonis geenitriivi tagajärjel.
Tooge näiteid geenitriivist inimpopulatsioonide näitel
GEENITRIIV – populatsiooni paljunemise käigus võib alleelide sagedus muutuda juhuslike
sündmuste tulemusena
. Geenitriiv leiab aset väikeste populatsioonide puhul.
Heterosügootide sagedus populatsioonis väheneb geenitriivi tulemusena seda kiiremini,
mida väiksem on populatsioon.
Geenitriiv
sõltub sellest, milliseid alleele sisaldavad gameedid järglaste saamisel osalevad.
Alleelide sagedust võib mõjutada ka see, et osadel vanematel on tunduvalt rohkem järglasi
kui teistel, kuid see ei pruugi olla seotud nende parema kohastumusega. Teiste vanemate
järglaskonda võivad vähendada näiteks õnnetusjuhtumid, ootamatu surm
keskkonnakatastroofide tõttu, halvad ilmastikutingimused.
Olukorras, kus populatsioonist on ainult väga vähesed ellu jäänud ja järglasi andnud,
räägitakse pudelikaela efektist.
Inimpopulatsioonide puhul on järgnevate põlvkondade
genofondi oluliselt muutnud näiteks suured epideemiad ja vallutussõjad
.
Geenitriivi üks mehhanisme on rajaja ehk asutaja efekt: emapopulatsioonist irdunud üksikud
isendid panevad aluse uuele populatsioonile, mille geneetiline struktuur erineb oluliselt
lähtepopulatsiooni omast.
Geenitriivi on kirjeldatud ka inimpopulatsioonide puhul:
18. sajandi algul rändas Saksamaalt
Ameerikasse rühm inimesi, kes kuulusid saksa baptistide usulahku, mille liikmeid nimetati
dunkeriteks. Kuni tänaseni elavad dunkerid USA-s isoleerituna. Kuna dunkerite
populatsioon on väike, on seal AB0 vererühmasid määravate alleelide sagedus võrreldes
ülejäänud USA elanikkonnaga ning Saksamaa populatsiooniga, millest nad pärit olid,
silmatorkavalt erinev
. Nt: vererühm B või AB esineb neil sagedusega 5%, teistel on vastavate
vererühmade sagedus 15%; vererühma A esineb dunkeritel sagedusega 60%, teiste 40-45%.
75. DNA ja valkude polümorfism populatsioonis. Miks võimaldab DNA järjestuse
analüüs võrreldes valkude järjestuse analüüsiga tuvastada rohkem mutatsioone?
Valkude aminohappelise järjestuse ja DNA nukleotiidse järjestuse võrdlustest saadud
andmete põhjal on selgunud, et organismide liigisisene alleelne varieeruvus on suur
.
Geneetilise muutlikkuse säilimise eest populatsioonis vastutavad nii stabiliseeriv valik kui
ka neutraalsete või peaaegu neutraalsete mutatsioonide korduv tekkimine.
Looduslikes populatsioonides esinev fenotüübiline muutlikkus ei kajasta alati
populatsioonides esinevat geneetilist muutlikkust. Selleks, et määrata geneetilise
muutlikkuse osa kogu muutlikkuses, analüüsitakse juhuslikult valitud geenide alleelset
varieeruvust molekulaarsel tasemel. Mutatsioonid konkreetses geenis ning sellest tulenevad
muudatused polüpeptiidi aminohappelises järjestuses on tuvastatavad mutantsete
polüpeptiidide erineva liikuvuse kaudu geelelektroforeesil.
Polümorfismi uurimine DNA järjestuse tasemel:
Valkude analüüs geelelektroforeesil ei võimalda tuvastada kõiki mutatsioone:
1) Mõnede alleelide puhul võivad mutatsioonid olla toimunud väljapool polüpeptiidi
kodeerivat ala, nt: intronis, promootoralas või kodeerivatest järjestustest
tahapoole jäävates alades
2) Mutatsioon kodeerivas järjestuses võib muuta koodoni viimast nukleotiidi, kuid
mõlemal juhul kodeerivad koodonid sama aminohapet
(nt: UUA ja UUG kodeerivad
mõlemad leutsiini)
3) Toimub ühe aminohappe asendumine teisega, millel on eelmisega võrreldes sama
laeng
(nt: valiini asendumine alaniiniga)
76. Liigi fenotüübiline ja bioloogiline kontseptsioon. Millal ei saa liiki defineerida,
lähtudes liigi bioloogilisest kontseptsioonist?
Liigi fenotüübiline kontseptsioon:
Käsitleb liiki organismide fenotüübiliste omaduste kaudu. Selle kontseptsiooni põhjal on
ühte liiki kuuluvate isendite fenotüübilised omadused sarnased
. Fossiilide
süstematiseerimise puhul ainuvõimalik meetod.
Liigi bioloogiline kontseptsioon:
Defineerib liike kui erinevaid organismide gruppe, populatsioone, kus ristumine toimub
ainult nende gruppide siseselt, gruppide vahel esinevad aga ristumisbarjäärid. Geenide
ülekannet erinevatesse liikidesse kuuluvate populatsioonidesse üldjuhul ei toimu.
Liiki ei saa liigi bioloogilisest kontseptsioonist lähtuvalt defineerida, kui liikide vahel
puudub ristumisbarjäär
. Nt: Bakterite puhul toimub geneetilise info ülekanne nii
erinevatesse liikidesse kui ka erinevatesse perekondadesse
. Geenide horisontaalne levik
erinevate bakteriliikide vahel toimub kas konjugatsiooni, transformatsiooni või
transduktsiooni teel, ähmastades oluliselt liikidevahelisi piire.
77. Tooge näiteid liikidevahelistest ristumisbarjääridest?
Presügootiline isolatsioon
– takistatud on erinevatesse liikidesse kuuluvate isendite
paaritumine
1) Ökoloogiline isolatsioon
– populatsioonid asustavad erinevaid elukeskkondi ega
puutu kokku
. Nt: lõvi ja tiiger asustavad ökoloogiliselt erinevaid piirkondi
2) Ajaline isolatsioon
– loomade paaritumise või taimede õitsemise ajad on erinevad.
Nt:
konnade paaritumisajad on liigiti erinevad; ahvenate kudemisajad on liigiti
erinevad
(ühtedel kevadel, teistel talvel?)
3) Käitumuslik isolatsioon
– erinevatesse liikidesse kuuluvate isendite vahel puudub
külgetõmme, pulmarituaalid on erinevad
. Nt: erinevad pulmarituaalid Hawaiil
elavatel äädikakärbestel
4) Isolatsioon erinevate tolmuterade edasikandjate tõttu – esineb taimedel
. Nt:
mõndasid taimi suudavad tolmeldada kimalased, kuid mitte tavalised mesilased
5) Isolatsioon sugurakkude sobimatuse tõttu
Postsügootiline isolatsioon
– hübriidne järglaskond on vähese eluvõimega või steriilne
takistades sellega hübriidide paljunemist
. Nt: leopardkonnade hübriidide embrüotel tekivad
väärarengud; hobuse ja sebra hübriidid on tihti steriilsed.
78. Liigitekke mehhanismid (allopatriline ja sümpatriline liigiteke)
Allopatriline liigiteke:
Populatsioonisisene diferentseerumine leiab aset siis, kui muutuvad elukeskkonna
füüsikalised või biootilised faktorid
. Populatsiooni alaosades akumuleeruvad erinevad
mutatsioonid, tekivad rassid, mis on kohastunud erinevate keskkonnatingimustega
. Kui
populatsioonidevahelised geneetilised erinevused suurenevad veelgi, tekib
ristumisbarjäär. Isegi siis, kui populatsioone varem eraldanud geograafilised barjäärid
edaspidi kaovad, jäävad need populatsioonid
(liigid)
paljunemise seisukohalt isoleerituks.
Sümpatriline liigiteke:
Populatsiooni ühes osas tekkinud unikaalsed mutatsioonid levivad teatavat ökoloogilist
keskkonda asustavate isendite vahel
. Geneetiliste erinevuste suurenemisel selles
ökoloogilises tsoonis väheneb ristumine teisi elupaiku asustavate isenditega, mis
soodustab geneetilist divergeerumist, kuni moodustub uus liik, mis on paljunemise
seisukohalt emapopulatsioonist täielikult isoleeritud
.
79. Evolutsiooni uurimine molekulaarsel tasemel. Mis on molekulaarne kell?
Motoo Kimura neutralistliku teooria põhjal on muudatused polüpeptiidide
aminohappelises järjestuses leidnud aset neutraalsete või peaaegu
neutraalsete
mutatsioonide juhusliku fikseerumise tulemusena erinevate liikide genoomi.
Erinevatesse liikidesse kuuluvate isendite polüpeptiidides toimunud muutuste analüüs
võimaldab hinnata evolutsioonilisi sündmusi. Mida kaugemad liigid, seda enam on muutusi
DNA-s
– molekulaarne kell.
Molekulaarse evolutsiooni neutralistlik teooria ei arvesta seda, et mõned mutatsioonid
võivad olla organismile kasulikud: neutralistliku teooria põhjal saab valik toimida ainult
kahjulike variantide kõrvaldamise kaudu
– erinevalt sünteetilisest evolutsiooniteooriast on
kõrvale jäetud darvinistlik positiivne valik
.
DNA järjestuse võrdluse põhjal on võimalik joonistada fülogeneesipuu.
Juureta puu –
näitab ainult erinevate liikide sugulusastet. Juurega puu – näitab lisaks sugulusele ka nende
liikide pärinemist ühest eellasest ja lahknemisaega
(mitu aminohappe asendust on
toimunud)
.
80. Inimese evolutsiooni uuringud DNA järjestuste analüüsi põhjal
Inimese lähievolutsiooni kohta on kogutud hulgaliselt andmeid tänu mitokondriaalse DNA
analüüsile
. Mitokondriaalse DNA analüüsi tulemusena pärineb meie kõigi esiema Aafrikast.
Ainult poegi sünnitanud naiste mitokondrid ei kandu järglastele
: arvutused taandusid
lõpuks ühele liinile, mille alguses olevat naist hakati nimetama Aafrika Evaks.
Tänapäeval Aafrikat asustav inimpopulatsioon on mitokondriaalse DNA suhtes kõige
heterogeensem – see populatsioon on kõige vanem.
Sarnased õppematerjalid
![GENEETIKA]()
34
docx
GENEETIKA
sõltumata eri gameetide vahel.
Tõenäosus, et kaks üksteist välistavad sündmust toimuvad on võrdne kummagi nähtuse
tõenäosuste summaga.
Geneetiliste andmete statistiline analüüs:
1. Mendeli suhteid saab ennustada matemaatiliselt null hüpotees.
2. Null hüpotees = erinevused on juhuslikud.
3. Võrdleme null hüpoteesi saadud andmetega, kuidas need sobituvad eeldatavate
tulemustega.
Kokkuvõte:
Mendeli geneetika ehk mendelismi aluseks on geenide ülekanne vanematelt järglastele ehk
põlvkonnast põlvkonda. Geenide ülekande mehhanism põhineb alleelipaari lahknemisel ehk
�segregatsioonil erinevatesse gameetidesse ning wrinevate alleelipaaride sõltumatul jaotumisel
sugurakkudesse.
Mendelismi printsiibid ja seaduspärasused on universaalsed kõikide suguliselt paljunevate
organismide korral. Mendeli seaduspärasused on oma loomult statistilised. Paljud
![Geneetika I kordamisküsimused-2012]()
18
doc
Geneetika I kordamisküsimused (2012)
tekib peptiidside. Polüpeptiidahela ühes otsas on alati amiinogrupp (algus) ja teises
karboksüülgrupp (lõpp).
Valkude struktuurid- primaarstruktuur- lineaarne a-h järjestus
Sekundaarstrukuur- alfa heeliksi või beeta-heeliksi sheet struktuur
Tertsiaarstruktuur- funktsionaalsetel valkudel 3D struktuur
Kvaternaarstruktuur- mitme valgumolekuli kompleks
Alguskoodon- AUG, lõppkoodon- UGA, UAA, UAG
7. Geneetika mõisted (alleel, monogeenne tunnus jne.)
Geen- Täielik nukleiinhapete järjestus, mis on vajalik funktsionaalse geeniprodukti(polüpeptiid,
RNA) sünteesiks.
pärilikkuse elementaarüksuse, DNA lõik, mis määrab ära RNA molekuli sünteesi: määrab ühe valgu
� järjestuse.
Struktuurgeenid- info RNA ehituse kohta (mRNA, tRNA, rRNA)
Regulaatorgeenid- kontrollivad struktuurgeenide aktiivsust ja avaldumist
Kromosoom- pärilikkuse salvestaja, geenide materiaalne kandja
![Rakutuum]()
12
doc
Rakutuum
Rakutuum
Rakutuum esineb ainult eukarüootsetes rakkudes. Üks suurematest organellidest (5-25 µm).
Ümbritsetud kahekordse membraaniga nn tuumaümbrisega. Välimine membraan on sageli
seotud ER membraaniga ja kahe membraani vaheline ruum on ER luumeni jätkuks.
Tuumamembraanid koosnevad lipiidsest kaksikkihist, milles esinevad teatud tüüpi valgud.
Tuumas on eristatav tuumakese piirkond. Tuuma sisemist osa, mis ei ole tuumake,
nimetatakse nukleo- e. karüoplasmaks. Tuuma struktuur ja koostis on määratud tuuma
funktsioonidega. Tuumas paikneb DNA ja toimub DNA replikatsioon. Tuumas paikneval
DNA-l sünteesitakse mRNA, tRNA, rRNA, samuti toimub tuumas ribosoomide subühikute
teke. mRNA, tRNA ja ribosoomide subühikud peavad liikuma tuumast tsütoplasmasse.
Tsütoplasmast tuuma peavad liikuma tsütoplasmas sünteesitud valgud (histoonid,
regulaatorvalgud, DNA ja RNA polümeraasid jt) Seega läbi tuuma membraani peavad
liikuma kõrgmolekulaarsed
![Geneetika I vastused]()
42
docx
Geneetika I vastused
GENEETIKA I KORDAMISKÜSIMUSED EKSAMIKS
1. Kaasaegse geneetika rakendusalad meditsiinis ja kohtumeditsiinis.
MEDITSIIN
Geneetilised uuringud on alati olnud suures ulatuses seotud meditsiiniga ja nende eesmärgiks on olnud
meditsiiniprobleemide lahendamine. Need uuringud on võimaldanud leida viise võitluses
nakkushaigustega ning kindlaks teha geene, mis on otsustavad pärilike haiguste tekkel. Geneetikute töö
tulemuseks on ka efektiivselt töötavad vaktsiinid.
1
![Geneetika kordamisküsimuste vastused 2013]()
22
doc
Geneetika kordamisküsimuste vastused 2013
ja tuvastamine
Meditsiin Geeniteraapia haigust tekitav geen on isoleeritud, teatakse selle geeni
poolt kodeeritud valkude biokeemilisi funktsioone organismis. Geenidefekt
kompenseeritakse normaalse, funktsionaalse geeni viimisega haige rakkudesse.
Molekulaarne diagnostika võimalik tuvastada haigust tekitavaid mutantseid geene,
siis toimub ravi või hooldus selle põhjal. Eriti oluline on sünnieelne diagnostika.
2. Kaasaegse geneetika rakendusalad põllumajanduses. Transgeensed organismid.
Organismi kloonimine.
Sordiaretus põllumajanduses mais(viljakam), nisu (stressikindel), tomatid (suurus,
värvus, kuju), koduloomade tõuaretus (rohkem piima või liha). Kunstlik
seemendamine.
Transgeensed organismid:
· soovitavate tunnuste lisamine või võimendamine taimedel ja loomadel nt
Bacillus thuringiensis genoomis geen, mis kodeerib putukatele toksilist valku,
![Sissejuhatus geneetikasse]()
96
doc
Sissejuhatus geneetikasse
http://www.tymri.ut.ee Õppetöö Geneetika 1
1. Sissejuhatus geneetikasse. Klassikalise ja molekulaargeneetika kujunemine. Geneetika tänapäeval:
rekombinantse DNA tehnoloogia; genoomide sekveneerimine; globaalne geeniekspressiooni uurimine,
geenikiibid. Kaasaegse geneetika rakendusalad; geneetika ja meditsiin (haigust põhjustavad
mutatsioonid geenides, geeniteraapia, molekulaarne diagnostika); geneetika kaasaegses
põllumajanduses; organismide kloonimine. Geneetika väärkasutused: eugeenika; lõssenkism.
2. Reproduktsioon kui pärilikkuse alus. Rakk kui elusorganismi ehituskivi. Eukarüootne ja
prokarüootne rakk Kromosoomid. Rakutsükkel, selle toimumist mõjutavad kontrollpunktid. Raku
jagunemine mitoosi teel. Raku jagunemine meioosi teel. Meioosi häired. Meioosi evolutsiooniline
tähtsus. Gameetide moodustumine erinevatel organismidel: oogenees; spermatogenees; sugurakkude
moodustumine taimedel.
3
![Klassikaline ja molekulaargeneetika-geneetika rakendus kaasajal]()
94
doc
Klassikaline ja molekulaargeneetika, geneetika rakendus kaasajal
1. Sissejuhatus: klassikaline ja molekulaargeneetika, geneetika
rakendus kaasajal
Klassikalise ja molekulaargeneetika kujunemine
Geneetika on suhteliselt noor teadus. Kuigi pärilikkuse põhilised seaduspärasused esitas Gregor Mendel
aastal 1865, tuleb geneetika sünniks lugeda siiski 20-nda sajandi algust. Alles siis taasavastati Mendeli
ideed, mis said aluseks klassikalisele geneetikale. Tõendid selle kohta, et DNA kannab geneetilist
informatsiooni, saadi 20-nda sajandi keskel. 1944. aastal kirjeldasid Avery ja ta kolleegid katseid, kus nad
uurisid bakterite (Streptococcus pneumoniae) transformatsiooni rakkudest isoleeritud DNA-ga. Hersey ja
Chase poolt aastal 1952 avaldatud tulemused kinnitasid seda, et DNA on pärilikkuse kandja. Nad näitasid,
![Üld- ja käitumisgeneetika kordamisküsimused-kevadsemester 2015]()
68
docx
Üld- ja käitumisgeneetika kordamisküsimused, kevadsemester 2015
On
olemas kindlaks tehtud individuaalsed geenid, kaksikute uurimisel on
pärilik 75%.
Autism ja ADHD on ilmselt kaks kõige tugevama päriliku
määratlusega psüühilist haigust.
4. Haploidne ja diploidne rakk, kromosoomid
Kromosoomid on raku struktuurid, mis koosnevad peamiselt DNA-st
ja valkudest. Haploidses eukariootses rakus on üks, diploidses kaks
komplekti kromosoome.
5. Geneetika arenguetapid
Klassikalise geneetika perioodil selgitati välja kromosoomide
seos pärilikkusega.
Populatsioonigeneetika liitmisel Darwini
evolutsiooniteooriaga tekkis tänapäevani toimiv sünteetiline
evolutsiooniteooria, mis andis ka selgituse organismide
muutlikkuse põhjuste kohta.
Molekulaargeneetika peerioodil selgitati välja geneetilise
Üld- ja käitumisgeneetika
Meedia
Kommentaarid (0)


















































































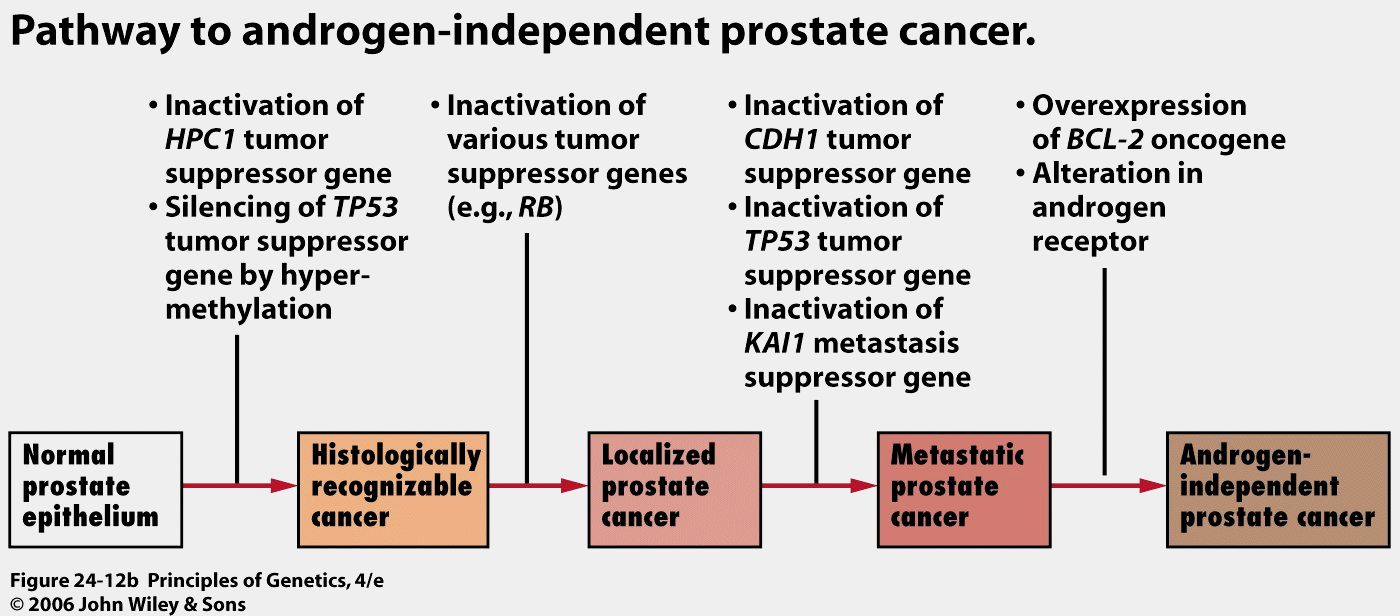































































Kõik kommentaarid