Kordamisküsimused
I
Loeng
1Mis
on meditsiiniline keemia ja mida uurib?
Meditsiiniline
keemia on keemiline distsipliin, mis hõlmab füsioloogia, mikrobioloogia , rakubioloogia, farmakoloogia ja
farmaatsia
aspekte .Distsipliini
eesmärk on uute bioloogiliselt aktiivsete ühendite avastamine,
identifitseerimine ja süntees, metabolismiuuringud,
toimemehhanismide välja selgitamine molekulaartasandil,
struktuur-aktiivsus uuringud, ravimidisain struktuuri ja farmakokineetika seisukohast.Mis
on ravim ?
Ravim on iga valmistatud, turustatud või turustamiseks määratud aine,
mis on ette nähtud haigete ravimiseks, haigusseisundi
kergendamiseks, haiguste ärahoidmiseks või diagnoosimiseks inimesel
või loomal, inimese või looma elutalitluse taastamiseks,
korrigeerimiseks või muutmiseks (WHO).Ravimisarnasel
ainel on toimeainet koguses või vormis, mis ei luba teda pidada
ravimiks, või mis ei sisalda üldse toimeainet, kuigi tootja
kirjeldab tootel ravimile iseloomulikke kasutusalasid.Kuidas
ravimeid klassifitseeritakse?
Farmakoloogiline
efekt
Keemiline
struktuur
- Sarnase struktuuriga ühendid ühte gruppi: penitsilliinid, barbituraadid , opaadid, steroidid. Segane, kuna bioloogiline toime võib sarnastel ühenditel erineda.
Sihtmärksüsteem
- Antihistamiinid, koliinergikud. Täpsem kui struktuur, kuid ei täpsusta, mis punktis ravim reaalselt toimib.
Molekulaarne sihtmärk
Mida
väljendavad ravimi kemoterapeutiline ja terapeutiline indeks?Paul
Erlich postuleeris võimaluse defineerida ühendite toime
selektiivsust mingi parameetriga, võimaldamaks võrrelda ravimeid
toime-ohutus kategoorias:
Kemoterapeutiline
indeks
= Min.efektiivne
doos / Max.talutav doos;
väikseim
parim.Sel
teel leiti skriiningu tulemusena ülitoksiline Salvarsan, mis osutus
ohutumaks kui atoksüül. Enne penitsilliini. Tänapäeval alles
terapeutiline indeks.
Ravimi
suhteline efektiivsusTerapeutiline
indeks :
LD50 / ED50
Iseloomustab
ravimi ohutust, arvestades erinevusi indiviidide vahel,
suurim
ohutuim,
arvestades doosierinevusest tingitud
ohutusvaru raviva ja
toksilise doosi vahel.
Ravimidoosi
ohutusKus
ja kuidas ravim toimib?
Ravimi
toime põhineb organismi ülesehitusel ja molekulaartasandi
funktsioonidel. Manustatud ravim kui keemiline ühend siseneb
keemiliste reaktsioonide maailm. Signaaliülekande ahelas
signaal võimendub.
Ravimi
toime avaldub rakus, raku osadele-organellidele. Ravim mõjutab
rakkudel elutalitlust, muutes füüsikalis-keemilisi tingimusi
rakuvälises keskkonnas, rakumembraanide
ioonkanaleid , rakkude
metabolismi ensüümide/nukleiinhapete kaudu ning toimides
regulatoorsetele protsessidele.
Ravim,
retseptor,
efektor , ioontransport, ensüümide
aktiveerimine /pärssimine, ainevahetus, mediaatorite/hormoonide
vabanemine , raku funktsioonide elavdamine/pärssimine -> elundite
funktsiooni muutus.
Mis
on ravimi molekulaarsed sihtmärgid rakus?
Makromolekulid;
lipiidid (membraani lõhkumine),
süsivesikud (väga spetsiifiline),
nukleiinhapped (DNA protsessid),
valgud (enamik ravimeid),
struktuuri- ja transportvalgud (vähe ravimeid),
ensüümid (inhibiitorid),
retseptorid (suur hulk, mis interakteeruvad
retseptoritega).
Sidumise interaktsioon toimub sihtmärkmolekuli sidumistsentris, kus võib
toimud keemiline
reaktsioon ravimi ja sihtmärkmolekuli vahel. Selle
tulemusena võivad
moodustuda kovalentsed sidemed (200-400 kJ/mol).
Haruldane .
Tavalised
on aga nõrgad
interaktsioonid (VDW (2-4 kJ/mol), HB (10-30 kJ/mol),
IB (20-40kJ/mol),
dipool -dipool,
tõukejõud ).
Lipiidid
ravimi sihtmärginaAmfoteritsiin
-
seenhaiguste vastane preparaat, interakteerub membraaniga, rajades
sellest läbi ulatuvad
hüdrofiilsed tunnelid.
Tsütoplasma voolab
välja ja
rakk hukub.
Valinomütsiin
(ionofoor) ja gramitsidiin A (kanali moodustaja) rikuvad
ioontasakaalu.
Suhkrud ja suhkrukonjugaadid ravimi sihtmärginaSuhkrutel
on oluline roll raku molekulaarse äratundmise, regulatsiooni ja
kasvu protsessides, millega on seotud paljud haigusseisundid.
Bakterid ,
viirused kasutavad raku pinnal
asuvaid süsivesikuid, mis
retseptoritena
toimivad , peremeesraku äratundmiseks.
Äratundmises
ei osale vabad sahhariidid, vaid
glükokonjugaadid
- glükoproteiinid, proteoglükaanid, glükolipiidid. Suhkru osa
sellest ulatub membraanist välja, toimides markerina. Oluline roll
on glükosfingolipiididel, mis osalevad kasvu regulatsioonis
(vähkkasvajad), olles samuti ka inimese veregrupi
määrajad .
Suhkrud ravimina Paljude
antibiootikumide struktuuri osad.
Streptomütsiin.
Ebahariliku kujuga süsivesikud on tihti vajalikud aktiivse ravimi
saamiseks,
aeglustades sauti bakteriliini
resistentsuse teket.
Mehhanism tundmata.
Asidotümidiin
(HIV),
atsükloviir.
Digitoksiin
(
südamepuudulikkus ).
Loeng
IIValgu
struktuuritasandid1.
Primaarstruktuur – peptiidahel aminohapetest
2.
Sekundaarstruktuur – peptiidahela pakkimine sama peptiidahela
lähestikku asuvate piirkondade sidumine vesiniksidemetega
3.
tertsiaarstruktuur – sama peptiidahela kaugel asuvate piirkondade
sidumine disulfiidsidemetega
4.
kvaternaarstruktuur – ruumiliselt korrastatud mitme peptiidahela
kombinatsioon
Keemiliste
sidemete tüübidKovalentne side 250 kJ/mol S-S side Cys
Vesinikside 7-40
kJ/mol Cys, Ser, Tyr, Trp, Thr, Asn, Gln, His,
Ioonne side 20 kJ/mol Asp, Glu, Lys, Arg
Van
der Waalsi jõud 1,9 kJ/mol aromaatsed jäägid Gly, Ala, Val, Leu,
Ile, Pro, Met, Phe
Valgu
interaktsioonid veegaHüdrofoobsed grupid interakteeruvad omavahel, ning hüdrofiilsed. Kõrvuti
asetsemise korral toimub tõukumine eri gruppide vahel. Vees
interakteeruvad veega hüdrofiilsed rühmad, lahustumisele aitavad
kaasa
vesiniksidemed , samal ajal pöörduvad valgu hüdrofoobsed
piirkonnad sissepoole, et veega võimalikult vähe interakteeruda
(
hüdrofoobne tsenter).
Transportvalkudele
orienteeritud ravimid Pöörduv
seondumine on viis viia polaarseid
ravimeid läbi membraani rakku (ravim kui
agonist ).
Pöördumatu
seondumine tõkestab looduslike
külalismolekulide transporti (ravim kui antagonist).
Kokaiin,
tritsüklilised antidepressandid , mis
inhibeerivad nordadrenaliini transportvalku.
Prozac e fluoksetiin inhibeerib pöördumatult
serotoniini transportvalku.
Struktuurivalkudele
orienteeritud ravimidSuunatud
tsütoskeleti valgule tubuliinile ning selle polümeriseerumisele,
mis on oluline raku pooldumises.
Colchicine
- aas-safranist, seostub tubuliiniga ning blokeerib
polümerisatsiooni, mis hoiab ära raku sisenemise mitoosifaasi.
Muudab neutrofiilid liikumatuks, millest tuleneb selle
põletikuvastane toime. EI KASUTATA
Vinkristiin
- blokeerib samuti polümerisatsiooni. Vähiravim.
Taksool
-
jugapuu koorest, seostub
mikrotuubulitega ja stabiliseerib neid, takistades
depolümerisatsiooni, arestides rakutsükli mitoosifaasis. Vähiravim
kemoteraapias.
Peptiidid ja valgud ravimitenaHormoonid.
Kui organism neid ise toota ei suuda
vajalikus koguses.
Võimaldas
rekombinantne DNA tehnoloogia .Vajalikku
valku
kodeeriv inimese geen kloneeritakse ja ekspresseeritakse mõnes
kiirelt kasvavas kultuuris (
bakter , pärm,
kasvajad ).
Puuduseks
manustamise
keerukus , kiire
metabolism ja võimalik immuunvastus.
Insuliin
- diabeet.
Seondub lihas- ja rasvarakkude insuliiniretseptoritega
soodustades glükoosi omandamist ning inhibeerides glükoosi
eritumist maksast.
Interferoon
-
leukeemia vormid, B ja C hepatiit, rekombinantne 19kD valk, mida
toodetakse E.coli kultuuris.
Erütropoietiin
- aneemiaravi. Glükoproteiin, mis stimuleerib verepunaliblede
(erütrotsüüdid) teket tüvirakkudest. Toodetakse rekombinantselt
imetajarakkudes.
Antikehad ravimitenaValkude
grupp, mille ülesandeks on ära tunda ja hävitada organismi
sattunud võõrvalgud või võõrrakud.
Y-
kujulised , koosnevad kahest raskest ja kahest kergest
ahelast ,
mille aminoterminustes on väga varieeruvad AH-järjestused.
Võõrkeha
teatud keemilised grupid, millele antikehad seostuvad, on
antigeenid .
Need on enamasti plasmamembraanist väljaulatuvad glükokonjugaadid.
Kui
antikeha on
antigeeniga seostunud , käivitub organismi
imuunvastus, mille käigus võõrrakk hävitatakse.
Antigeenid
on kui molekulaarne allkiri, mille abil omi rakke võõrastest
eristatakse.
Väga
efektiivne autoimmuunsete haiguste ravis
nagu nt reumatoid-
artriit . NSAID-id, kortikosteroidid võitlevad vaid
tagajärgedega, mitte põhjustajate ning kõrvaltoimetega.
Probleem
on luua antikehi inimrakkudele olukorras, kus inimorganismis nad ei
teki. Need genereeritakse mõne looma organismis ja eraldatakse,
ravim seotakse antikehale ning antikeha kasutatakse ravimi
transpordiks sihtmärkrakuni või manustatakse infusioonina.
Esimesena:
lümfotsüütide antigeeni vastu loodud antikeha. Seostub
lümfotsüütidega, mis olid muutunud kasvajarakkudeks ning
mobiliseeris organismi immuunsüsteemi neid hävitama.
Ensüümkatalüüsi
olemusAktsivatsioonienergia
vähenemise saavutab
katalüsaator siirdeoleku stabiliseerimise teel,
tagades optimaalse keskkonna või pinna, viies reageerivad molekulid
kontakti, fikseerides aineid nii, et siirdeolek oleks kergesti
saavutatav, nõrgendades katkema pidavaid keemilisi
sidemeid ,
osaledes reaktsioonimehhanismis ning hiljem taastudes.
Kiirendab
reaktsiooni alandades aktivatsioonienergiat, kuid ei muuda tasakaalu.
Valksubstraatinteraktsioonid
ensüümi katalüütilises tsentris Püruvaat
–laktaadi dehüdrogenaas –>
laktaat Phe,
Ser, Lys katalüütiline triaad, toimub kabonüülse hapniku
taandamine hüdroksüülrühmaks.
Ensüümide
regulatsioonKonkurentne
pöörduv inhibeerimine - ravim seondub
paremini kui
substraat ning seetõttu ei ole substraadil enam
võimalik seostuda. Reaktsioon on blokeeritud
Konkureeriv
inhibeerimine - etanooli ja
etüleenglükooli koos esinedes eelistab keha etüleenglükooli, kui
etanooli hulga suurenedes eelistab jälle etanooli.
Mittekonkurentne
pöördumatu inhibeerimine -
aspiriin atsetüleerib tsüklooksügenaasi pöördumatult.
Mittekonkurentne
pöörduv inhibeerimine - inhibiitori
seondudes muutub ensüümi aktsiivtsentri konformatsioon ning
substraat enam sinna ei mahu. Nt merkaptopuriin, leukeemiaravim, mis
inhibeerib puriinide biosünteesiahela esimese ensüümi ja sellega
ka DNA sünteesi.
Ensüümide
inhibiitorid ravimitenaSildenafil
- viagra
toimeaine , kaitseb
cGMP -d lagunemise eest, tagades NO
initsieeritud regulatsiooni kestva toime.
Aspiriin
- inhibeerib pöördumatult nii COX-1 kui COX-2, pole selektiivne.
Tselekoksiib,
Celebrex inhibeerivad vaid COX-2 (ei
mahu COX-1), millel on pikaajalise ravi korral väiksemad
kõrvalmõjud.
Antibakteriaalsed agendid - eluvajalike ensüümide selektiivne inhibeerimine Sulfoonamiidid
- inhibeerivad
foolhappe sünteesi, takistades raku kasvu ja
pooldumist.
Penitsilliin
- inhibeerivad pöördumatult glükopeptiid-transpeptidaasi,
blokeerides raku seina sünteesi viimase astme. Rakk
sureb .
Tsefalosporiinid
-
bakteritsiidne ravim koos penitsilliiniga.
Antiviraalsed
agendidAtsükloviir
ja famtsükloviir - nukleosiiditüüpi
ained mittetäieliku desoksüriboosiringiga. Fosforüleerides end
aktiivsesse trifosfaati, inhibeerivad pöördtranskriptaasi ja on DNA
sünteesi terminaatoriteks.
Zidovudin
on esimene registreeritud AIDSi ravim, inhibeerides HIV
pöördtranskriptaasi, blokeerides viiruse DNA sünteesi. Tänu
asiidgrupile on DNA ahela sünteesi
terminaator .
Loeng
IIIGeeni
ekspressiooni kontrolli tasandid - DNA lugemine, transkriptsiooniline kontroll
- mRNA modifitseerimine enne tuumast välja transportimist ja translatsiooni, RNA- processing
- kontroll mRNA transpordi tasemel
- kontroll mRNA translatsiooni tasemel
- kontroll mRNA degradatsiooni tasemel, juhul kui temast valku ei sünteestia
- peale mRNA translatsiooni valgu post-translatoorne kontroll -> inaktiivne valk
DNA
struktuur ja selle tasandid1.
Primaarstruktuur – nukleotiididest moodustunud singletne ahel,
moodustudes nukleosiidist, mis on omavahel ühendatud fosfaatrühmaga
ning mis sisaldavad desoksüsuhkrut ning lämmastikalust (
guaniin ,
adeniin,
tümiin , tsütidiin).
2.
Sekundaarstruktuur – biheeliks,
keeru kõrgus 34 A, keeru laius 20
A. Vasakukäeline. Biheeliksit hoiavad koos komplementaarsete
lämmastikaluste vahelised vesiniksidemet; A=T ja G≡C.
3.
Tertsiaarstruktuur – keerdumine (
supercoiling).
Enne replikatsiooni tuleb lahti harutada. Seda teostavad
topoisomeraasid, ensüümid, mistõttu saab neid bakteritel
inhibeerida nt
tsiprofloksatsiiniga,
mis on kinolooni tüüpi
antibakteriaalsed ravimid, fluorokinoloonid.
4.
Kvateraarstruktuur -
kromosoom DNA
kui ravimi sihtmärk
Interkalaarsed
agendid tungivad nukleiinhapete
aluspaaride kihtide vahele, moonutades kaksikspiraali kuju, mis
takistab DNA replikatsiooni ja transkriptsiooni.
Alküleerivad
agendid on tugevad
elektrofiilid ,
mis nukleofiilidega moodustavad tugevaid kovalentseid sidemeid.
Ahelat lõikavad agendid on suured
glükoproteiinid, mis lõikavad DNA ahelat ning oma mahukusega
takistavad DNA ligaasil tekkinud katkestuskohti parandamast.
Antimetaboliidid
blokivad normaalsed metaboolsed
rajad .
Aminopteriin
- blokeerib DNA sünteesi dihüdrofolaadi reduktaasi inhibeerides.
Sellega on inhibeeritud tetrahüdrofolaadi süntees, mis on
kofaktor puriinaluste sünteesil.
6-merkaptopuriin
on eelühend 6-merkapto-guanosiinfosfaadi sünteesil, ning inhibeerib
mitmeid radu
puriin -nukleotiidide sünteesil.
Fluorouratsill
- arvatakse, et inhibeerib mitmeid ensüüme DNA sünteesil.
Ensüümide
inhibiitorid inhibeerivad otseselt
DNA sünteesil osalevaid ensüüme.
ElliptitsiinAmsakriin
- topoisomeraaside inhibiitorid. Põhjustavad DNA replikatsiooni ja
transkriptsiooni blokeerimist, kuna ei lase vastavatel ensüümidel
DNA-d lõigata ja taasühendada, mille tagajärjel rakk sureb.
Kemoteraapia .
Interkalaarsed
agendidMahutavad
end DNA biheeliksi vahele, muutes ahela kuju. Õhukesed ja
planaarsed, harilikult aromaatsed või heteroaromaatsed polütsüklid.
Proflaviin,
mahutub kahe paari vahele.
Hiniin
ja klorokiin - malaariaravimid,
hiniin on kiinapuust eraldatud alkaloid ning klorokiin on selle
sünteetiline
analoog . Toime põhineb malaariaparasiidi DNA
transkriptsiooni blokeerimisel. Iseloomulik struktuur võimaldab
interkaleeruda malaariaparasiidi DNA-ga.
Aktinomütsiin
D ja Adriamütsiin on
antibiootikumid, mis sisaldavad vastavaid rühmi, millega töötavad
interkalaarsete agentidena.
Alküleerivad
agendidTugevad
elektrofiilid. DNA-s on mitmeid
nukleofiilsed gruppe, millest olulisim on guaniini 7.
lämmastik .
Agendid on väga reaktiivsed ning kasutatakse vähiravimitena.
Ravimil
peab olema kaks alküleerivat gruppi, mis reageerivad kahe lähestikku
asetseva, kuid erinevates DNA ahelates paikneva G-ga, ristseovad kaks
ahelat, blokeerides seega replikatsiooni ja transkriptsiooni.
Samuti
võib ravim siduda kaks guaniini samast ahelast, mistõttu takistab
ka selline
klammerdumine ligipääsu DNA
ahelale . Alküleeritud
guaniini transkriptsioon võib põhjustada vigu nagu nt
mittefunktsionaalse valgu sünteesi.
Metkloretamiin
-> asiridiini
ioon (endisest kahest alküülahelast moodustub
kolmene heterotsükkel N-ga) -> guaniini 7.lämmastik ründab
tsüklit (moodustub kovalentne side C-N ning teine alküülahel
moodustab kolmese heterotsükli N-ga) -> teise ahela 7.lämmastik
ründab tsüklit -> ristseotud DNA.
Kõrvalreaktsiooni
vältimiseks saab reaktsioonivõimet alandada nt metüülrühma
aromaatse ringiga asendades. Samuti aitab nukleiinhapete ehituskivide
lülitamine ravimi struktuuri nagu
Uratsiil
" mustard "il, kasutusel
kroonilise lümfiteede leukeemia ravis. Teatud selektiivsus.
Tsisplatiin
on testise ja munasarja vähkkasvaja vastu. Seondub DNA-le
järjestuslõikudes, kus on palju guanosiinijääke, neid
ristsidudes, ning transkriptsiooni takistades.
Mitomütsiin
C - vähivastane ravim, mis kehas
alküleerivaks reagendiks muudetakse. Toimub kinooniringi taandamine
ensümaatiliselt hüdrokinooniks, misjärel
metanool lahkub ning
3-liikmeline asiridiiniring
avaneb . Järgneb alküleerimine.
DNA
ahelat lõikavad agendidBleomütsiin
A2,
B2-
arvatakse et tõmbab DNA molekulis H-aatomeid. Moodustunud radikaalid
reageerivad hapnikuga,
andes peroksüradikale, mis seejärel
fragmenteeruvad.
RNA
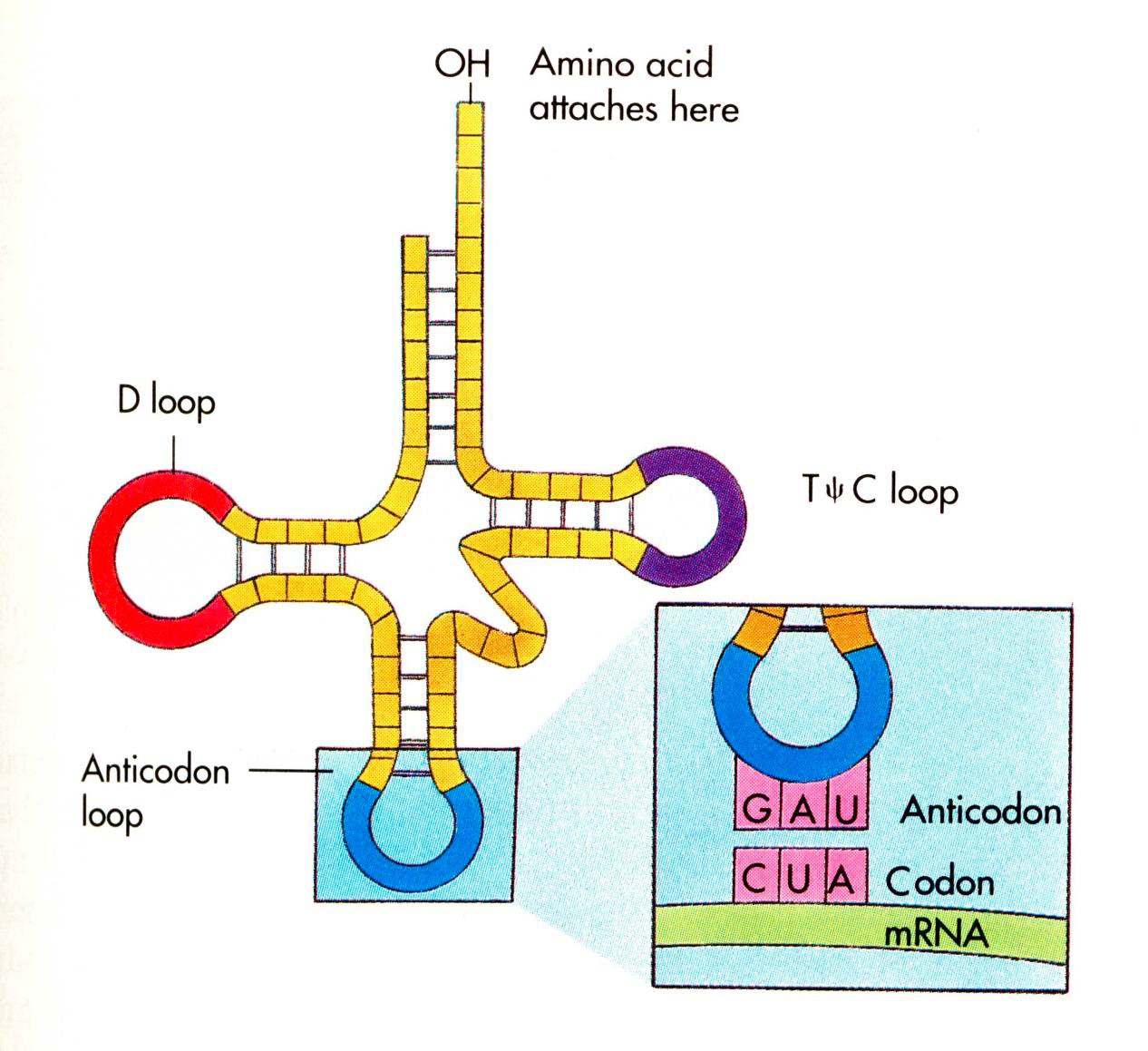
struktuur ja tüübidtRNA transport-RNA. tRNA on adaptor, mis ühendab
nukleiinhappe tripletset koodi vastava aminohappega. tRNA molekulil
on kaks sidumisregiooni. Üks seob transporditava aminohappe
kovalentselt terminaalse adenosüüljäägiga, teise otsa kolm
nukleotiidi (
antikoodon ) seostuvad komplementaarselt mRNA tripletiga.
mRNA
maatrits -RNA.
Vahendab valku kodeerivat
informatsiooni genoomselt DNA-lt valgu sünteesisaiti. DNA
kaksikspiraal hargneb, eksponeeritav DNA lõik (geen) toimib
matriitsina (
template ), millelt transkribeeritakse mRNA. Modimise
tagajärjel saab see tuumast väljuda, misjärel toimub valgu
süntees.
rRNA ribosoomne RNA. Ribosoomne RNA on valgu
sünteesisaidiks. Ta seostub mRNA molekuli ühte otsa, liigub
geneetilist koodi
lugedes kuni mRNA teise otsani.
rRNA
kaudu toimivad ravimidToimivad
bakteri RNA molekulidele, mõjutades translatsiooni.
Aminoglükosiidid
(gentamütsiin, kanamütsiin, neomütsiin) toime seisneb seondumises
bakteri 16S rRNA-le, millega kaasnevad vead koodi lugemises mRNA-lt
ning seeläbi bakteri eluliselt oluliste valkude sünteesi pärssimise
või mutantsete valkude akkumulatsioonis bakteri membraanis.
TUlemusena memrbaani barjäärifunktsioon häirub ning rakk lüüsub.
EI seostu inimese 18S-rRNA-le, siis on toime
selektiivne.
mRNA
kaudu toimivad ravimidAntisenss
ravimid seisnevad oligonukleotiidide kasutamises mRNA translatsiooni
takistamiseks.
Probleemiks
on asjaolu, et mRNA on suur
molekul , millel on sek ja
terts struktuur
- raske leida järjestuslõike, mis on eksponeeritud, lisaks sellele
oligonukleotiidi
viimine rakku. Potentsiaal on suur.
Esimene
tuli turule 1992.a papilloomiviiruse põhjustatud genitaaltüügaste
raviks.
Nukleosiiditüüpi
ravimidEi
ole algselt aktiivsed, kui peale fosforüleerimist trifosfaadiks võib
inhibeerida pöördtranskriptaasi ning viiruse DNA sünteesi.
AZT
e zidovudiin /retrovir
pürimidiinipõhine, AIDS-le, liitub kasvava DNA ahelaga, omades
suhkrujäägi OH asemel N3 rühma, katkestab DNA ahela edasise kasvu.
Atsükloviir/famtsüklovir
- puriinipõhine,
Herpes simplex,
vöötohatis . Töötab sarnaselt
tänu mittetäielikule suhkrujäägile.
Geenitehnoloogia ravimidisainisRekombinantsete
valkude ja peptiidide tootmine -
insuliin, kasvufaktorite
geenid viiakse
bakterirakku ning toodetakse
neis eesmärkühendit farmaatsiatööstuse
nõudlusele vastavalt.
Uute
valguliste ravimisihtmärkide tuvastamine
- eraldamine ja tuvastamine kloneerimisega. Isosüümide ja
retseptorite alatüüpide tuvastamine, mis viivad selektiivsemate
ravimite tegemiseni. Inimese geeniprojekt aitas kaasa sadade uute
valkude avastamisele.
Sihtmärkvalkude
molekulaarsete mehhanismide uurimine -
valkude modimine üksikute AH-de vahetamise teel, mis aitab selgitada
üksikute jääkide rolli ensüümkatalüüsis või retseptoriga
seostumises.
Somaatiline geeniteraapia - transportviiruse
kasutamine terve geeni sisse
viimisel rakkudesse, kus antud geen on
vigane . Viirusinfektsiooni järel ühineb terve geen peremeesraku
DNA-ga ning allub seejärel loomulikele protsessidele. Suur
perpektiiv .
Loeng
IVNeli
signaalmolekulide rühmaNeutrotransmitterid-
närviimpulsi ülekandeks närviraku poolt toodetud signaalmolekulid,
mis toimivad teise närvi-, lihas- või eritusrakule spetsiaalselt
välja arenenud kontaktkohtades - sünapsides.
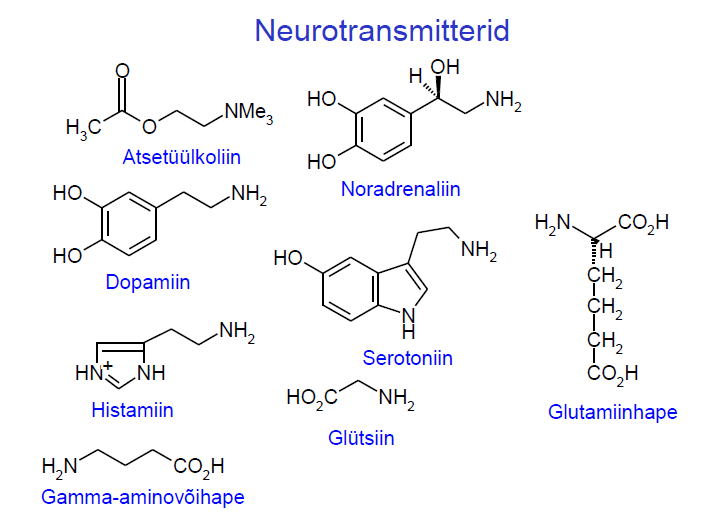
Atsetüülkoliin , noradrenaliin , adrenaliin , dopamiin , histamiin , serotoniin ,
glutaminhapeHormoonid-
bioaktiivsed
endogeensed ained, mida sünteesitakse
endokriinnäärmetes ja mis vere vahendusel reguleerivad metaboolseid
protsesse ja füsioloogilisi funktsioone.
Türoksiin,
kortikosteroonHormonoidid
e
koehormoonid e lokaalsed hormoonid
-
keemilised ained, mida toodavad mitmed kudede spetsialiseerunud
rakud ja millel on regulatoorne mõju metaboolsetele protsessidele.
Histamiin, prostaglandiinidNeurohormoonid-
hormoonid, mis produtseeritakse spetsiaalsete närvirakkude poolt.
OxytocinKuidas
tekib membraanipotentsiaal?
Ioonpumbad ja -kanalid tekitavad ioone sisse-välja pumbates
potentsiaalierinevuse membraani kummagi poole vahel, mis on üldjuhul
membraanist seespool negatiivne, eukarüootides -40 mV kuni -80 mV.
Puhkeseisundis
on raku kaaliumkanalid avatud ning K+,
mille konts rakust väljas on 30-40x seesmisest suurem, liigub raku
sisse, kuni on saavutatud raku K+
tasakaalupotentsiaal vastavalt Nernsti seadusele. Lisaks K+-le
osalevad ka Na+
ja Cl-.
Nernst’i
võrrandAktsioonipotentsiaal
( närviimpulss )Membraanipotentsiaali
kiire tõus ja langus pidevat trajektoori mööda. Tekib lihas-,
endokriin - ja närvirakkudes. Oluline roll neuronite rakk-rakk
suhtlemises. Aktsioonipotentsiaal on tuntud ka kui närviimpulss.
Aktsioonipotentsiaal
tekib pingetundlike ioonkanalite poolt. Puhkepotentsiaali ajal on
kanalid kinni, kuid avanevad kiirelt kui membraanipotentsiaal
suureneb teatud piirväärtuseni. Avanedes lasevad kanalid sisse Na+
ioone, mis muudavad elektrokeemilist gradienti, mis omakorda
potentsiaalierinevust suurendavad. Selle tulemusena avaneb veelgi
rohkem
kanaleid , suurendades
voolutugevust membraaniläbivas
elektrivoolus.
Sünaps Sünaps
on
närvisüsteemi struktuur,
mis lubab neuronil saata teisele neuronile keemilist või elektrilist
signaali. Sünapsis läheneb signaalisaatja neuroni membraan
(presünaptiline neuron)
vastuvõtja -neuronile (postsünaptiline
neuron), millel mõlemil on molekulaarsed vahendid membraanide
ühendamiseks ning signaliseerimiseks.
Närviimpulsi
ülekanne sünapsisNärviimpulss
kutsub esile närvilõpme membraani depolarisatsiooni, avanevad Ca2+
kanalid ja rakuväline Ca2+
suundub närvilõpmesse. Selle tagajärjel vabanevad närvilõpmes
vesiikulitest neurotransmitterid, mis satub sünaptilisse pilusse
ning seostub vastuvõtja-neuroni retseptoritega. Kompleksi
moodustamise tagajärjel avanevad retseptoriga seotud
ioonkanalid ning tekib ioonide vool läbi ioonkanali, mis kutsub ka
postsünaptilises rakus esile membraanipotentsiaalimuutuse -
postsünaptilise potentsiaali.
Agonisti ,
antagonisti ja osalise agonisti toime retseptorileAgonist - seostub ja aktiveerib
- peavad olema õiged keemilised rühmad
- rühmad peavad olema õiges kohas
- mõõtmed vastavad aktiivtsentri mõõtmetele
- dopamiini analoogid?
Optiline
isomeer võib anda vähem või rohkem interaktsioone.
Antagonist - seostub aga ei
aktiveeri - võib toimida nii aktiivtsentrisse kui
väljaspoole
- väljaspool aktiivtsentrit mõjub
allosteeriliselt või vihmavarjuna.
Vastab
aktiivtsentri konfiguratsioonile ja interakteerub selle
funktsionaalsete rühmadega, kuid retseptori konformatsiooni ei
muuda.
Allosteeriliselt
seostub allosteerilise tsentri, mitte aktiivtsentriga ning muudab
seostumise tulemusena retseptori aktiivtsentri konformatsiooni ning
agonist ei saa seonduda.
Vihmavarju
efekti näol seostub väljaspool
aktiivtsentrit olevate aminohappe jääkidega ning katab tsentri
osaliselt või täielikult.
Osaline
agonist aktiveerib retspeotir kuid avab
kanali väiksemal määral kui agonist. Kutsub esile ainult nõrga
bioloogilise vastuse. Osalist agonisti võiks käsitleda ka
antagonistina, kuna hõivad aktiivtsentri ja takistab
neurotransmitteril või agonistil suuremat efekti esile kutsumast.
Signaalmolekuli
ja retseptori vahelised interaktsioonidX
Retseptori
tundlikkuse muutumineTolerantsus võib olla seotud retseptorite sensibiliseerimisega, mistõttu läheb
varasemast rohkem ravimit uute retseptorite inhibeerimiseks. Kui
peatada ravimi võtmine järsku, on retseptoreid tavapärasest
rohkem, mistõttu on rakk neurotransmitteri suhtes ülitundlik ning
see kutsub esile
normaalsest tugevama reaktsiooni. Sellel perioodil,
kui retseptorid vähenevad, tekib patsiendil vajadus ravimi järele,
et ebameeldivaid nähtusi leevendada -
sõltuvus .
Desensibilisatsioon,
sensibilisatsioonKui
agonist toimib retseptorile pikemat aega, siis retseptor inaktiveerub
e desensibiliseerub. Põhjuseks on retseptori fosforüülimine, mille
tulemusel retseptor muutub konformatsioonilt inaktiivseks.
Kui
agonist veelgi edasi toimib, siis retseptorid viiakse tsütoplasmasse
ja hüdrolüüsitakse,
kusjuures uute retseptorite süntees
aeglustub.
Kui
antagonist retseptorile pikaajaliselt mõjub, siis retseptori
tundlikkus kasvab, see sensibiliseerub. Põhjuseks on kiirem
retseptorite süntees, kompenseerimaks inhibeeritud retseptorite
funktsioone.
Loeng
VRetseptorite
klassifikatsioonIoonkanaliga
seotud retseptorid - 2-, 3-, 4-TM
4-TMmõlemad
terminused on akust väljas. N-terminaalsel osal on signaalmolekuli
aktiivtsenter. Suletud olekus on J nurgad sees koos, avatud olekus
aga C kujuliselt, ning neist saab läbi minna ioonide
voog .
Avanemise põhjustab TM2 domeenide asendi muutus.
Nikotiini-,
serotoniini-, glütsiini- ja gammaaminovõihappe retseptorid.3-TMN-terminus
on rakust väljas, C-terminus on raku sees.
Ligandi sidumise kohas on
N-terminusel (A) ning teine (B) ekstratsellulaarsel domeenil TM3 ja
TM4 vahel.
L-glutamaadi
retseptori-Ca2+-kanali valgu subühik.2-TMMõlemad
terminused on rakus sees ning
ioonkanal kosneb viiest subühikust.
Kaks transmembraanset domeeni.
ATP
aktiveerib.G-valguga
seotud retseptorid - 7-TM
7-TMC-
term on raku sees, N-väljas. Ei ole seotud ioonkanaliga. Signaaliülekanne
toimub G-valgu kaudu, mis seostub C-terminaalse ahelaga pärast
retseptori aktiveerimist. Seostumine toimub C-terminuse ja VI ning V
subühikud ühendava ahela vahel.
Signaaliülekandjad:
monoamiinid (dopamiin, histamiin,
serotoniin, atsetüülkoliin, noradrenaliin), nukleotiidid, lipiidi,
neuropeptiidid, peptidohormoonid, glükoproteiinhormoonid, glutamaat,
Ca2+.Ligand
võib seonduda kas membraani sees (monoamiinid, nukleotiidid,
atsetüülkoliin, lipiidid), membraani ääres (neuropeptiidid),
eelpoolmainitud domeenide juures (glükoproteiinhormoonid) või
N-terminusepoolsele (glutamaat, Ca).
N:
Trombiini retseptor,
5-AH, ligand.
Trombiin lõikab N-terminaalset osa, mis
segab ligandi
sidumist aktiivtsentriga. Seejärel saab ligand aktiveerida
retseptorit.
Kinaasiga
seotud retseptorid - 1-TM
1-TMInaktiivsetele
EGF retseptori monomeeridele seondub EGF (bivalentne epidermaalne
kasvufaktor ), tekib dimeerne retseptor ja kumbki retseptori
türosiin -
kinaas aktiveeritakse ning nad fosforüülivad teineteist.
Mõlematele seotuakse kolm fosfaatrühma.
Insuliini
retseptor.Heterotetrameerne
kompleks . Samamoodi.
Intratsellulaarsed
retseptorid
Signaali
ülekanne ioonikanaliga seotud retseptori kauduSignaali
ülekanne G-valguga seotud retseptori kaudu - Neurotransmitter seondub retseptorile, mille tagajärjel toimub konformatsioonimuutus;
- G-valk membraanis moodustab retseptoriga kompleksi.
- G-valgu α-subühiku aktiivtsentrist vabaneb GDP
- GDP asemel seondub GTP
- α-subühik eraldub G-valgust ja inaktiveerib või aktiveerib membraanivalgu, vastavalt subühikule.
Adenülaadi
tsüklaasi rada - Kõigepelat toimub signaali ülekanne G-valguga seotud retseptori kaudu (GDP vabaneb, GTP seondub, α-subühuk lahkub).
- αQ/αS – subühik aktiveerib adenülaadi tsüklaasi; adenülaadi tsüklaasi aktiivtsenter avaneb ja toimub cAMP katalüütiline süntees ATP-st.
- Kuna α-subühik on GTPaasse aktiivsusega, hüdrolüüsib see lõpuks oma GTP, fosfaatrühm vabaneb ning alles jääb GDP. Kompleks laguneb ja adenülaadi tsüklaas inaktiveerub.
- Saadud cAMP võib aktiveerida proteiinkinaase, mis katalüüsivad ensüümide ja valkude fosforüülimist.
Fospolipaasi
C rada - αq-subühik (vabanedes G- valgult seoses GDP vabanemise ja GTP sidumisega) aktiveerib fosfolipaas C; selle aktiivtsenter avaneb ja fosfatidüül-inositool-4,5-bisfosfaat (PIP-2) hüdrolüüsitakse.
- Hüdrolüüsi tulemuel tekib kaks sek. signaaliülekandjast inositooltrifosfaadist (IP3) ja diatsüülglütseroolist (DG)
- α-subühik hüdrolüüsib GTP GDP-ks ning vabaneb fosfolipaas C-lt.
Signaali
ülekanne kinaasiga seotud retseptori kaudu - DG – diatsüülglütserool jääb lipofiilsena membraani
- DG aktiveerib proteiinkinaas C-d
- PKC fosforüülib tsütoplasma ensüümide Ser ja Thr jääke
- Fosforüülitud ensüümid katalüüsivad aktiivses olekus spetsiifilisi keemilisi reaktsioone
1
TMÜheaegselt
retseptor ja ensüüm. C-terminuse lähedal on katalüütiline
piirkond ja N-terminuse lähedal on ligandi sidumise piirkond.
Insuliini
retseptor.
Kasvofaktorite
retseptorid, tsütokiinide retseptorid. - IP3, inositooltrifosfaat toimib organellidele, mis seovad Ca+
- Ca vabaneb ja aktiveerib Ca-sõltuva proteiinkinaasi
- Proteiinkinaas fosforüülib spetsiifilisi ensüüme
- Vabanenud Ca2+ seondub kalmoduliiniga, mis aktiveerib Ca-kalmoduliin-sõltuvad proteiinkinaase
- Need proteiinkinaasid forsforüülivad edasi.
Signaali
ülekanne intratsellulaarsete retseptorite kauduAktiveeritud
retseptor käivitab rakus rea keemilisi reaktsioone, millede abil
signaal rakus liigub ja jõuab tsütoplasma valkudeni või tuuma.
Steroididele,
kilpnäärme hormoonidele, retinoididele.
Steroidhormoonide
retseptoridIntratsellulaarsed.
Steroidi sidumise koht ja DNA sidumisekoht. Tamoksüfeen on
östrogeeni antagonist, rinnavähi ravim.
Signaalmolekul
tuleb läbi membraani, moodustab retseptoriga ligand-retseptor
kompleksi millele seondub valk-koaktivaator, ning kogu komplekt saab
seonduda DNA-le.
Östrogeeni
retseptor seostub DNA järjesusega
5'-AGGTCA-3'.
14
































Kõik kommentaarid