Nädal 1 1. Mis tüüpi küsimustele populatsioonigeneetika vastust otsib. Kirjelda üldiselt põhilist töövõtet. Too näiteid erinevatest populatsioonigeneetika mudelitest. Mis on mudeli parameeter ja tema hinnang? Mis tegurid võivad viimast mõjutada. (Sille) Populatsioonigeneetika uurib populatsioonide varieeruvuse muutumist ajas: selgitab, kuidas praegune varieeruvus tekkinud on, ja ennustab seda, milliseks varieeruvus tulevikus saab. Uurib erinevaid tegureid, mis varieeruvust mõjutavad (nagu loodusliku valiku toimumine, geneetiline triiv, mutatsioonid, geenisiire, Mendeli pärilikkuse seadused, paarumismustrid, populatsioonide struktureeritus). Varieeruvus kui alleeli/genotüüpide/haplotüüpide sageduse muutumine
EESTI MAAÜLIKOOL VETERINAARMEDITSIINI JA LOOMAKASVATUSE INSTITUUT LOOMAGENEETIKA JA TÕUARETUSE OSAKOND ARETUSÕPETUS I OSA (POPULATSIOONIGENEETIKA) LOENGUKONSPEKT G P Koostaja: dots. E. Orgmets TARTU 2008 1 KORDAMISKÜSIMUSED 1. Populatsioonigeneetika. (populatsiooni mõiste, panmiksis, biotsönoos, populatsiooni evolutsioonitegurid, suletud ja avatud populatsioon, populatsioonimaht, valimi mõiste). 2. Juhuslikkus ja tõenäosus. Tõenäosuste korrutamise seadus. 3. Genotüübi sagedus ja selle arvutamine. 4. Geeni- ehk alleelisagedus ja selle arvutamine. Geenisageduse arvutamine alleeliseeria korral. 5. Hardy-Weinbergi seadus (definitsioon, tingimused, valem p2+2pq+q2=1). POPULATSIOONI GENEETILINE DÜNAAMIKA 6
mitme SNP tõttu DNA kodeerivas piirkonnas (ka indel-id). Polümorfisme piimavalkude geenides saab tuvastada restriktaaside kaasabil . Muutliku DNA lõigu paljundamine. Restriktsiooni reaktsioon. Restriktsioonifragmentide tuvastamine elektroforeesiga seos jõudlusega (piima kg, v% ...) ·mõju piima tehnoloogilistele omadustele ·mõju inimtoidule (sh allergeene) 2. Põlvnemisandmete kontrollimine (ülesanne) 3. Populatsioon. Panmiktiline/ideaalne/geneetilises tasakaalus (Hardy-Weinbergi tasakaaluseadus). Panmiktilises populatsioonis, mis on geneetilise tasakaalu seisundis, püsivad alleeli- ja genotüübisagedused põlvkonniti konstantsed. Alleeli- ja genotüübisagedused on omavahelises sõltuvuses. Populatsioon püsib geneetilises tasakaalus seni, kuni talle ei toimi geeni- või genotüübisagedusi muutvad faktorid. - a. Isoleeritud- geneetiline tasakaal on püsiv, b. Panmixis c
juhuvaliku tõttu. Järglase alleelide kombinatsioon on vanematelt juhuslikul moel pärandunud, mistõttu see, kas üks või teine isend jääb ellu ning saab järglasi, oleneb mingil määral juhusest. Mingi alleeli sagedus (alleelisagedus) populatsioonis on selle alleeli eksemplaride osatähtsus selle alleeli eksemplaride hulgas. Geneetilise triivi tagajärjel võib osa alleele (geenivariante) kaduda, nii et geneetiline mitmekesisus väheneb. Geenitriivi mõju on seda suurem, mida väiksem on populatsioon. Teadlased vaidlevad ägedalt selle üle, kui suur roll võrreldes loodusliku valikuga on evolutsioonis neutraalsetel protsessidel, sealhulgas geenitriivil. Ronald Fisher oli seisukohal, et geenitriivi roll evolutsioonis on tagasihoidlik või olematu, ja see vaade jäi mitmeks aastakümneks valdavaks. Vaidlus läks uue hooga käima, kui 1968. aastal esitas Motoo Kimura oma molekulaarse evolutsiooni neutraalse teooria, mille väitel geneetiline muutus (mis ei pea tingimata
Kordamisküsimused 1. Geneetika põhietapid 1.1. Eelteaduslik periood Geneetika eelteaduslikule perioodile on iseloomulikud üksikud õiged ja objektiivsed tähelepanekud, mida varjutavad aga tol ajal massiliselt levinud spekulatsioonid ja filosoofilised targused. · Hippokrates (V-IV saj. ema.) - lapsed arenevad algmetest, mis tekivad kogu kehas. Selle tõttu sarnanevad lapsed vanematele ja omandatud tunnused päritakse. Pärilikkust võivad mõjutada isegi mõtted. Seisukoht tuntud pangeneesi hüpoteesina. Darwin arendas seda omandatud tunnuste päritavuse põhjendamiseks (gemmulad). · Demokritos (V-IV saj. ema.) - inimeste võimed arenevad peamiselt harjutamise, mitte kaasasündinud eelduste tõttu. Koos Empedokelesega preformatsiooniprintsiibi pooldaja ja propageerija. · Pythagoras (V saj ema.) isaslooma kehas (närvid, aju jne.) tekkiv fluidum koituse ajal kondenseerub emasloom
vahel isendite migratsiooni ja ristumise teel. 33. Ringliigid (mõiste ja näide) Olukord, milles 2 reproduktiivselt isoleeritud populatsiooni, kes elavad samas regioonis, on seotud omavahel ristuvate populatsioonide geograafilise ringiga. . Salamandrid Californias on põhja pool ühe asurkonnana, mägedes (Sierra ja Kaljumäestik) teise asurkonnana 34. Liik (mõiste ja näide) organismide populatsioon, millel on kõrgtasemeline geneetiline sarnasus. 35. Alamliik organismide kogum populatsioonis, mille liikmetel on teatud erilised tunnused. Omavahel liikmed ristuvad vabalt. 36. Pleiotroopsus (mõiste ja näide) ühe (mutantse) geeni samaaegne fenogeneetiline toime mitmele erinevale tunnusele. Näiteks kollast karvkattevärvust põhjustav geen hiirtel kutsub samal ajal esile ainevahetuse aeglustumise, mistõttu hiired rasvuvad kergesti 37
evolutsioonilisi muutusi, uurib evolutsiooniprotsessi käimalükkavaid molekulaarseid mehhanisme ja geenide, genoomide ja nende produktide (sh valkude) muutusi evolutsiooniprotsessis. Peamisteks aladeks on makromolekulide evolutsiooni uurimine, geenide ja organismide evolutsioonilise ajaloo uurimine ehk molekulaarne fülogeneetika, elu tekke ja päritolu uurimine. Molekulaarne evolutsiooniga seotud teadusharudeks on molekulaarbioloogia (andmed) ja populatsioonigeneetika (teooria). Molekulaarset evolutsiooni uuritakse liikidevaheliste erinevuste ja liigisiseste polümorfismide alusel (DNA või valk). Põhiprobleemid, millega mol evo tegeleb: Uuritakse liikidevahelisi erinevusi ja polümorfisme. Põhjustajateks on mutatsioonid – evolutsiooni seisukohalt on olulised ainult pärilikud mutatsioonid. Kui selline mutatsioon tekib ja on kahjulik, kõrvaldatakse see
17. Vali 2 darvinismi ja mendelismi peamist ühendajat 1930. 40. aastatest ja kirjelda kummagi panust eraldi? R.A. Fisher pidevate tunnuste pärandumist seletab Mendel, eeldades, et geenidel on polümeerne koostoime. Kuidas LV väikese efektiga geenide puhul mõjutab populatsiooni aeglast muutumist. S. Wright inbriiiding ja juhuslik geneetiline triiv. Kohastumine on seda efektiivsem, mida väiksemateks alampopulatsioonideks populatsioon jaguneb valiku ja triivi koostöö. Rääkis seostest geno/fenotüüpide ja kohasuse vahel. 18. Mida kujutab endast evolutsiooniteooria moodne süntees? Millised olid põhisaavutused/põhiseisukohad? Millised olulised küsimused jäid vastamata? Moodne süntees ühendab darvinismi ja mendelismi. Põhiseisukohad: mendellik geneetika on LV alus, adaptsioone teeb ainult LV, varieeruvus säilib ja ei ühtlustu, keskkond ei mõjuta varieeruvust otse, mutatsioonid on juhuslikud
















































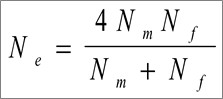
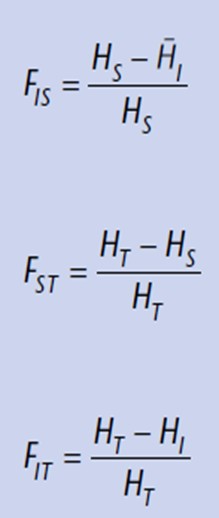

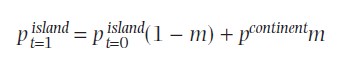
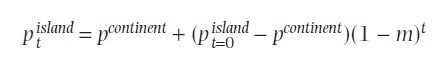
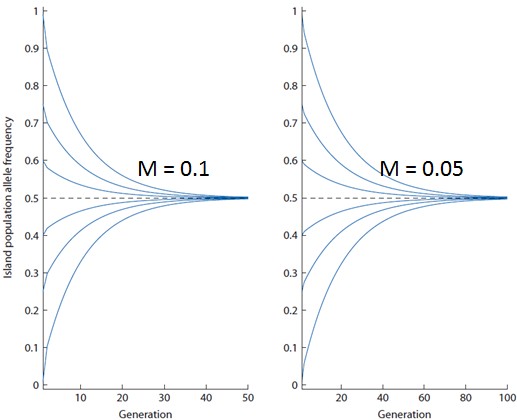
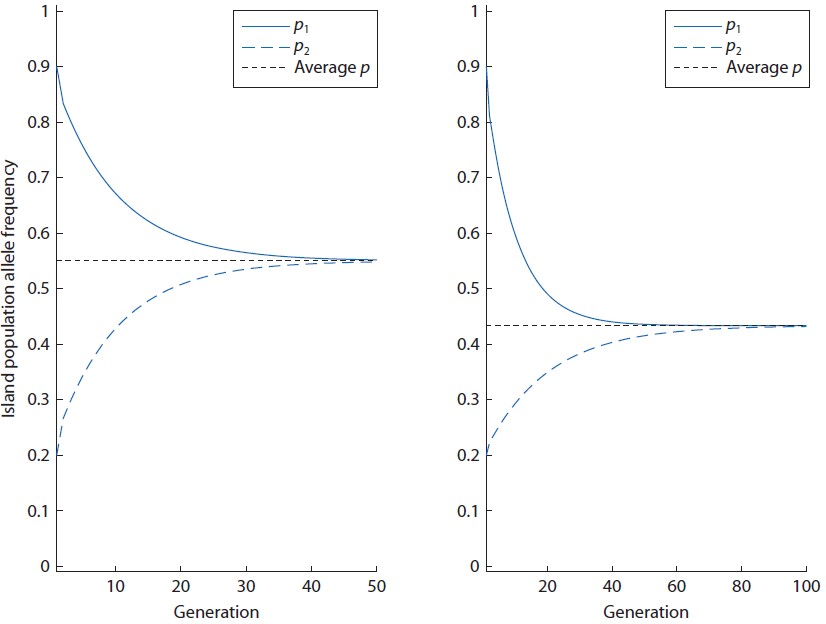
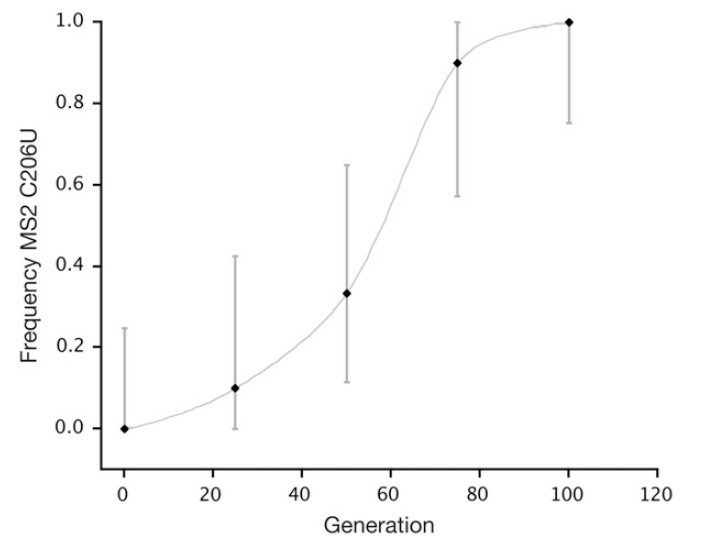

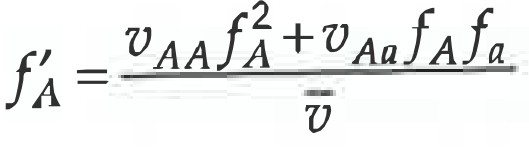
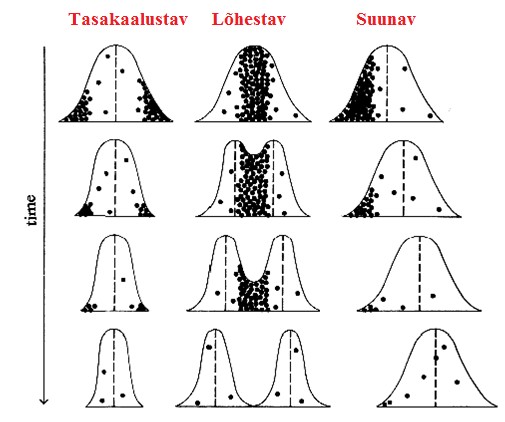
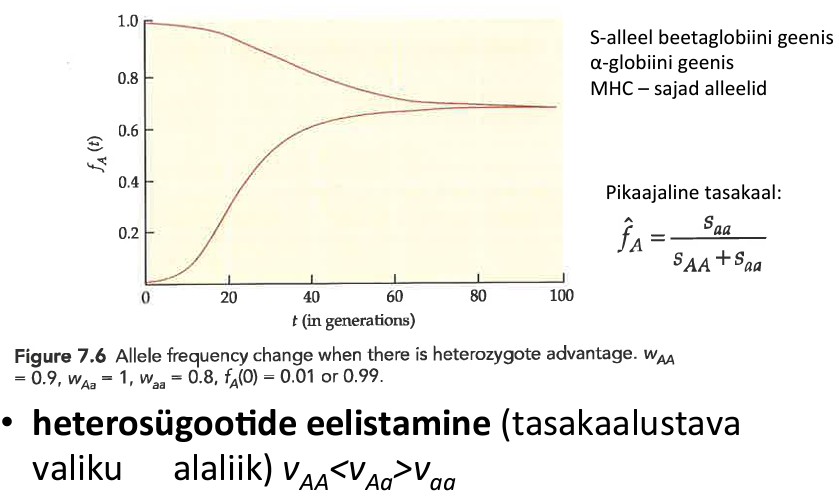
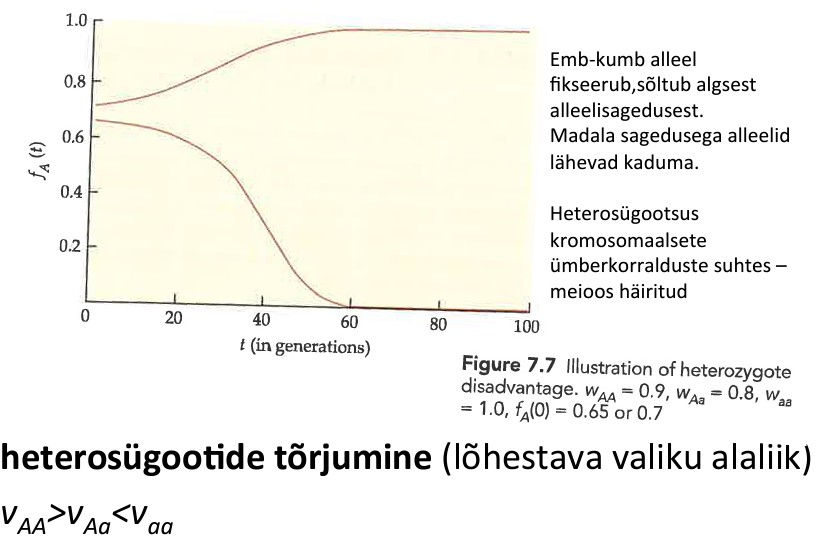
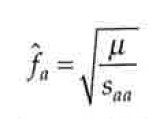
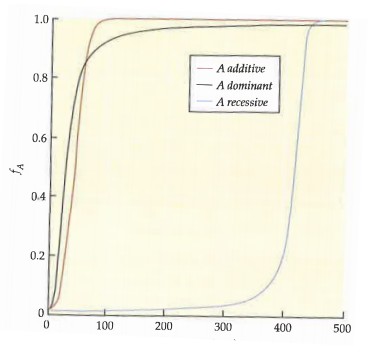
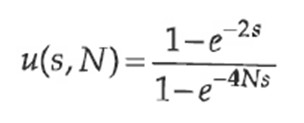
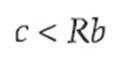
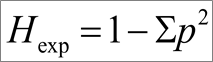
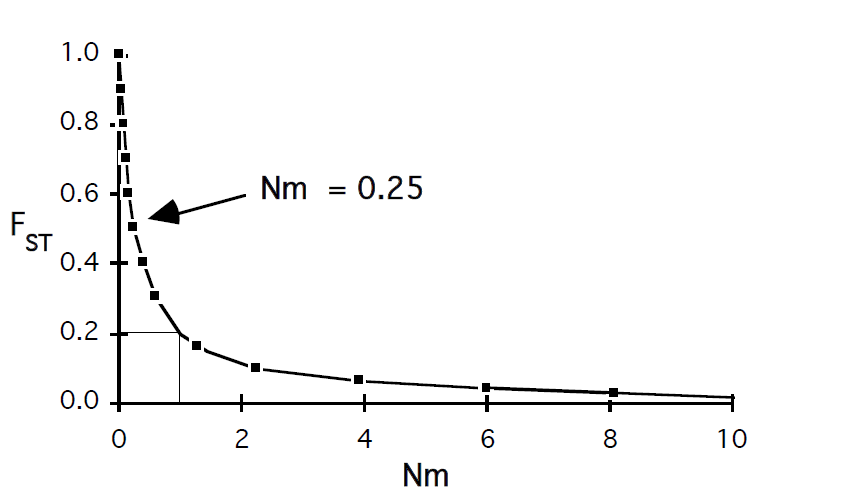
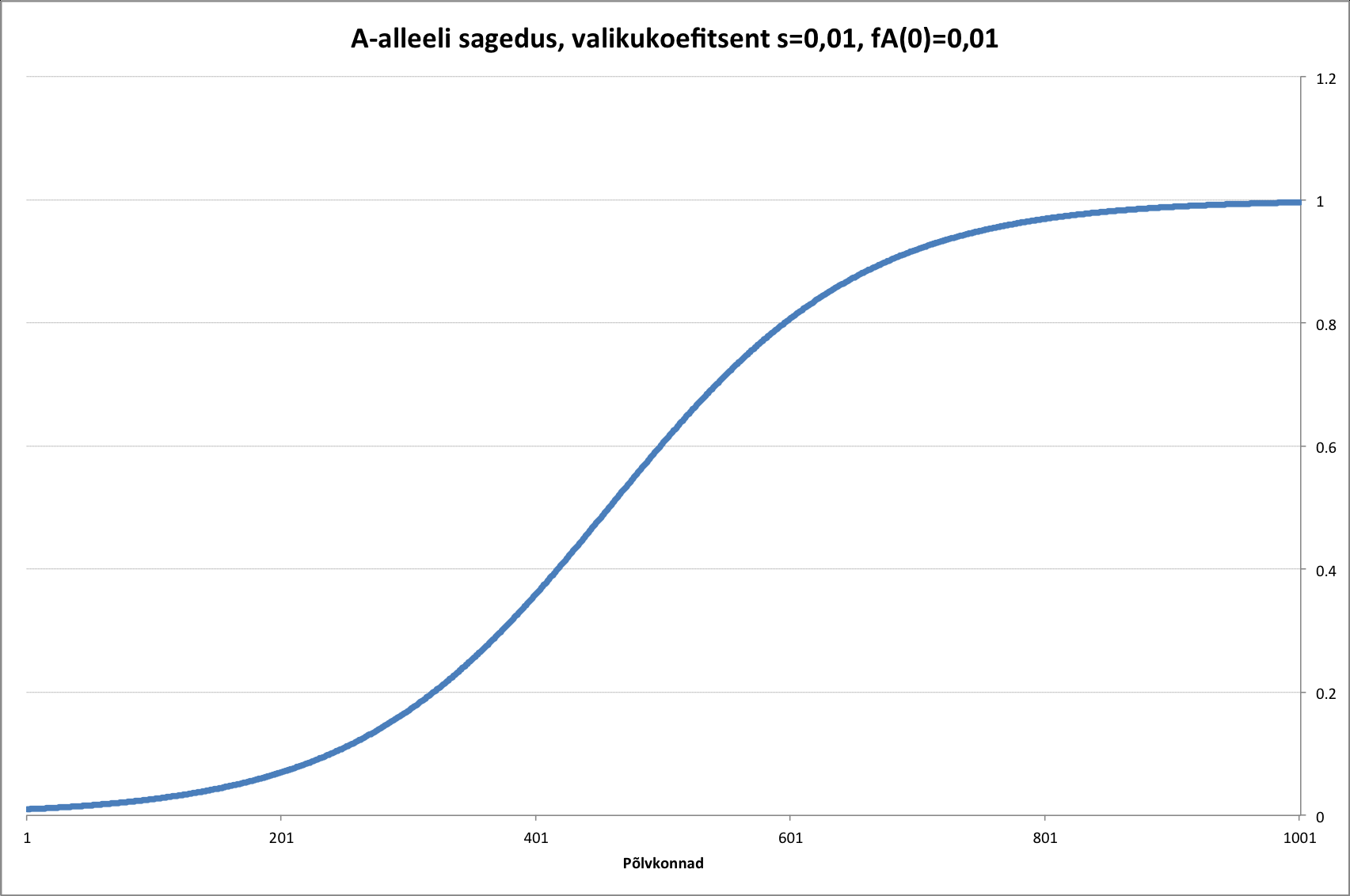
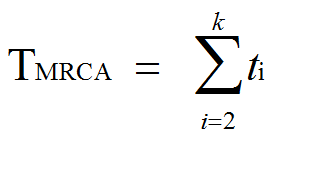




Kõik kommentaarid