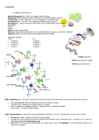
järjestus, mis esineb helix-pööre-helix motiivina . 17- aminohappeline kolmas heeliks ulatub DNA suurde vakku. Seondumine VÄGA SPETSIIFILINE . Juba 1 aminohappe muutus mõjutab DNA-ga seondumist (seon dub mõne teise promootori/enhanceriga). 10. Põhjenda miks ja kirjelda kuidas toimub X kromosoomi inaktivatsioon imetajatel? Kahekordne X-kromosoomide geenide produktid oleksid rakkudele letaalsed. Igas imetaja rakus inaktiveeritakse üks X-kromosoomidest. Barri kehake replitseerub hilises S-faasis ja valdavat osa DNA- st ei transkribeerita. See, kas arengu käigus inaktiveeritakse isalt või emalt saadud X-kromosoom, on juhus. Inaktiveeritud X-kromosoom inaktiveeritakse ka tütarrakkudes inaktivatsiooni pöördumatus . CpG-saared inaktiivses X-kromosoomis on suuremas osas metüleeritud. Metüleerimine on mehhanism transkriptsiooni peatamiseks. XIC X-kromosoomi inaktivatsiooni tsenter
*ensüümi kovalentne modifotseerimine (fosforüleerimine), *sümogeenid, isosüümid, modulaatorvalgud. Ensüümide kovalentne modifitseerimine ensüüme, mida nii reguleeritakse nimetakse interkonverteeritavateks ensüümideks. Ensüüme, mis katalüüsivad interkonveeritavate ensüümide kahe vormi vahelist konversiooni, nim. konverterensüümideks. Fosforüleerimine fosforhape jäägi lisamine ensüümile. Ensüümvalgus teatud aminohape fosfoleeritakse, selle tulemusel ensüüm kas inaktiveeritakse või aktiveeritakse. (Vaata loeng 9 slaid 6 joonist) Defosforüleerimine fosforhappe jäägi eemaldamine ensüümilt. Temagi roll on ensüüm kas aktiveerida või inaktiverida. Seega fosforüleerimise vastand reaktsioon. 3) Proensüümid (sümogeenid) ja isoensüümid (isosüümid) mõisted, nende roll katalüüsi regulatsioonis. Proensüümid e sümogeenid valkude inaktiivsed prekursorid, mille proteolüütilise lõikamise tulemusel saadakse aktiivne valk
mRNAd 2.kontroll mRNA töötlemise tasemel - millised muutused toimuvad mRNA-ahelas enne translatsiooni algust 3.kontroll RNA transpordi tasemel - millised mRNA molekulid viiakse tsütoplasmasse, kus toimub valkude süntees 4.kontroll translatsiooni tasemel - millised mRNA molekulid kaastakse valgusünteesi 5.kontroll mRNA lagundamise tasemel - millised mRNA molekulid suunatakse lagundamisse 6.translatsiooni-järgne kontroll - millised toodetud valkudest aktiveeritakse/inaktiveeritakse, millistesse raku osadesse valgud transporditakse 16)Välis- ja sisekeskkonna tegurid, mis mõjutavad geenide avaldumist. keskkonnatingimused nt välistemperatuur, õhusaastatus, toitained, kemikaalid
mRNAd 2.kontroll mRNA töötlemise tasemel - millised muutused toimuvad mRNA-ahelas enne translatsiooni algust 3.kontroll RNA transpordi tasemel - millised mRNA molekulid viiakse tsütoplasmasse, kus toimub valkude süntees 4.kontroll translatsiooni tasemel - millised mRNA molekulid kaastakse valgusünteesi 5.kontroll mRNA lagundamise tasemel - millised mRNA molekulid suunatakse lagundamisse 6.translatsiooni-järgne kontroll - millised toodetud valkudest aktiveeritakse/inaktiveeritakse, millistesse raku osadesse valgud transporditakse *Rakkude erinevused tulenevad geenide valikulisest avaldumisest. *Geenid avalduvad kui nende pealt sünteesitakse RNAd. *Geenide avaldumise lõppeesmärgiks võib olla vajaliku valgu süntees. Kõik geenid ei sisalda infot valgusünteesiks, nende pealt sünteesitakse rRNAd, tRNAd. *Kõik peab olema kindla kontrolli all: rakkude jagunemine, geenide avaldumine.
läikimine, paksenemine ja ninde tagajärjel kergesti tekkivad vigastused. Paljud veenilaiendite ravimid sisaldavad hepariini, mis see on? Hepariinid on segu antikoagulantsete omadustega, erineva molekulide ahela pikkusega glükoosaminoglükaanidest, mida produtseerivad nuumrakud ja basofiilid. Hepariinid võivad pärssida trombotsüütide funktsiooni ning põhjustada trombotsütopeeniat. Hepariinid inaktiveeritakse maksas ning eritatakse uriiniga. Kokkuvõte Veenilaiendid on haigus, mis kimbutab väga paljusid inimesi. Kahjuks pööratakse sellele küllaltki vähe tähelepanu, mis aga ainult süvendab haigusekäiku. Kahjuks ei ole sellele ka 100%-st ravi. Vaevusi saab leevendada erinevate sukkade ja käsimüügi ravimitega, veidi tõhusamad on spetsialisti käe all veenidesse süstimine ning veenioperatsioon. Need võivad aga paljudele tunduda hirmutavad ja
6. Millised on retseptorid sümpaatilises närvisüsteemis ja milline on seal neuromediaator neuroni ja sihtmärkorgani vahel? Sümpaatilise närvisüsteemi retseptorid on α1, α2, β1, β2. Neuromediaatoriks esimese ja teise neuroni vahel on AK (atsetüülkoliin), teise neuroni ja sihtmärkorgani vahelises sünapsis on neuromediaatoriks NA (noradrenaliin). 7. Millised ensüümid lammutavad sümpaatilises närvisüsteemis noradrenaliini? Väiksem osa noradrenaliinist inaktiveeritakse mitokontrites paikneva MAO (monoaminooksüdaasi) ja tsütoplasmaatilise ensüümi COMT (katehhool-o- metüültransferaasi) poolt. 8. Mis juhtub selle noradrenaliiniga, mida ensüümid ei lammuta? Presünaptilisest osast vabanenud noradrenaliin transporditakse suures osas tagasi presünapsisse ja hoiustatakse uuesti vesiiklites. 9. Sümpaatilise närvisüsteemi retseptorite tüübid, asukohad ja mis toime aktivatsioonil. 10
suundub abil...Kovalentsed proteiinhormoonid- närvilõpmesse.Siis sidemed ei nende vabaneb teki.Desensibilisatsioo sidumiskohad)On neurotransmitter,mis n-rets tundlikuse erinevad G-valgu sattub sünaptilisse muutmine kui agonist subühikud.Liigist sõltub pilusse ning seostub toimib rets-le pikemat mis valki aktiveeritakse rets-ga.Tekib iooonide aega,siis toimub rets või inaktiveeritakse ja vool läbi inaktiveerimine.Kui sellele järgnev signaali ioonkanali,toimub toimib veel,siis ülekande biokeemiline membraanipotents eemaldatakse rets rada.Adenülaadi muutus.Agonist membraanist,aeglustub tsüklaasi aktiveerib G- seostub rets-ga ja uute rets valguga,as subühik aktiveerib seda(peavad süntees.Sensibilisatsi aktiveerib fosfolipaasi
3. Kontroll RNA transpordi tasemel: millised tuumas toodetud mRNA-molekulid viiakse tsütoplasmasse, kus toimud valkude süntees. 4. Kontroll translatsiooni tasemel: millised tsütoplasmas leiduvad mRNA- molekulid kaasatakse valgusünteesi ribosoomides. 5. Kontroll mRNA lagundamise tasemel: millised tsütoplasmas leiduvad mRNA- molekulid suunatakse lagundamisse. 6. Translatsiooni-järgne kontroll: millised toodetud valkudest aktiveeritakse või inaktiveeritakse, millistesse raku osadesse valgud transporditakse. Organismi geenide avaldumist mõjutab keskkond: nii väline keskkond, milles organism areneb ja elab, kui ka organismi sisekeskkond: hormoonid ja ainevahetus. Organismiväliselt mõjutavad geenide avaldumist näiteks erinevad kemikaalid, ravimid, temperatuur, valgus ja toitainete kättesaadavus. Keskkonnatingimuste mõjul võivad geenid lülituda sisse või välja, mõjutades nii tunnuste avaldumist.
3) Kontroll RNA transpordi tasemal: millised tuuma toodetud mRNA-molekulid viiakse tsütoplasmasse, kus toimub valkude süntees. 4) Kontroll translatsiooni tasemel: millised tsütoplasmas leiduvad mRNA –molekulid kaasatakse valgusünteesi ribosoomides 5) Kontroll mRNA lagundamise tasemel: millised tsütoplasmas leiduvad mRNA –molekulid suunatakse lagundamisesse 6) Translatsiooni-järgne kontroll: millised toodetud valkudest aktiveeritakse või inaktiveeritakse , millistesse raku osadesse valgud transporditakse 1.5 Geenide avaldumise häired Healoomuline kasvaja- kontrollialt väljunud rakud ei tungi ümbritsevatesse kudedesse Pahaloomuline- rakud tungivad ümbritsevatesse kudedesse, välja arenenud vähk. Prioonhaigused- tekitavad aminohappejärjestuselt normaalsed, kuid vigase struktuuriga valgud- prioonid Tekitavad valesti voltunud valgud, mis hakkavad muutma ka teisi. Creutzfeldti-Jakobi tõbi
Kapsiidi struktuurvalgud ja assambleerumisel osalevad valgud võtavad enda alla 40% geneetilisest informatsioonist Ikosaeedriline pea, helikaalne tupp, kuusnurkne basaalplaat Tupp ja pea on ühendatud kaelusega Basaalplaadile kinnituvad sabakiud ja nõelad Bakteriofaag T4 infektsioonitsükkel Varajane infektsioonistaadium – avalduvad varajased geenid 1) Inaktiveeritakse faagi poolt kodeeritud valkude abil raku geenide transkriptsioon ja translatsioon 2) Toimub faagi genoomi paljundamine Hiline infektsioonistaadium – avalduvad hilised geenid 1) Kapsiidivalkude ja faagi DNA-d protsessivate
◦adenülaadi tsüklaasi inhibeerimise ◦K+-kanalite läbilaskvuse suurenemise (hüperpolarisatsioon) ja postsünaptilise neuroni pidurdus ◦Ca2+-kanalite läbilaskvuse vähenemise (ACh, NA, DA vabanemine sünapsipilusse väheneb) Kodeiini farmakokineetika Köha pärssiv toime 10-15 mg Imendub peensoolest ja nahaalusest koest kiiresti Kodeiin konverteeritakse morfiiniks maksasensüümi CYP450 CYP2D6 vahendusel ning CYP3A4 vahendusel tekib norkodeiin Tekkinud metaboliidid inaktiveeritakse metaboliseerides kodeiin-6-glukuroniidiks(C6G) valdavalt ensüüm uridiindifosfaatglükoronosüül transferaasi (UGT2B7)vahendusel ◦6–10% valgest rassist ja 2% asiaate on “aeglased metaboliseerijad“ ◦0.5-2% populatsioonist on “ülikiired metaboliseerijad“ (polümorfismi tõttu toodetakse palju CYP2D6) Dekstrometorfaan Levometorfaani derivaat Toime nõrgem kui kodeiinil Dekstrometorfaan suurendab morfiini jt agonistide toimet retseptoritel
enne translatsiooni algust 3)kontroll RNA transpordi tasemel :millised tuumas toodetud mRNA-molekulid viiakse tsütoplasmasse, kus toimub valkude süntees 4)kontroll translatsiooni tasemel : millised tsütoplasmas leiduvad mRNA-molekulid kaasatakse valgusünteesi ribosoomides 5)kontroll mRNA lagundamise tasemel : millised tsütoplasmas leiduvad mRNA- molekulid suunatakse lagundamisse 6)translatsiooni-järgne kontroll : millised toodetud valkudest aktiveeritakse või inaktiveeritakse, millistesse raku osadesse valgud transporditakse *Geenide avaldumise esimene etapp on transkriptsioon. RNA-polümeraas seostub promootoriga ja hakkab sünteesima mRNAd. KESKKOND MÕJUTAB GEENIDE AVALDUMIST *Nii väline keskkond kui ka sisekeskkond: hormoonid ja ainevahetus. Organismiväliselt mõjutavad geenide avaldumist nt kemikaalid, ravimid, temp, valgus ja toitainete kättesaadavus.
somaatilistes rakkudes 37. X-kromosoomi inaktivatsioon X-i inaktivatsioon toimub astmeliselt ja järgib "n-1" reeglit, mis tähendab, et kõik X- kromosoomid peale ühe diploidse genoomi kohta inaktiveeruvad ühtlustub X-i geenide doos emas- ja isasisendite somaatilistes rakkudes Lyoni hüpotees - "ühe aktiivse X-i hüpotees", 1961 Lyoni hüpotees 1. üks imetajate emasisendite X-kromosoomist inaktiveeritakse varases embrüonaalse arengu staadiumis 2. inaktiveeritakse juhuslikult emalt või isalt saadud X- kromosoom 3. kõik antud raku järglased säilitavad inaktivatsiooni mustri, st. sama X on inaktiivne 4. kui rakus on enam kui kaks X-kromosoomi, on kõik peale ühe inaktiivsed X- kromosoomi inaktivatsioon inimesel sõltub kromosoomipiirkonnast, mida nimetatakse X-i inaktivatsiooni keskuseks (X inactivation center, Xic)
Kasutatakse radioaktiivseks märgistamiseks, kloonimisel (RACE tehnika), et lisada homopolümeersed osad DNA molekuli 3' otsale kloonimiseks. Alkaalne fosfataas ,,BAP": bakteriaalne alkaalne fosfataas, mis eemaldab 3´ ja 5´ fosfaatrühmad DNAst ja RNAst. Tähtis roll kloonimisel: defosforüülib 5´-fosforüülitud otsa selleks, et takistada vektori ise-ligeerimist. BAP aktiivne 65°C juures for at least 1 h ja seda inaktiveeritakse phenol extraction. Alkaalne fosfataas ,,CIP": fosfataas, mis eemaldab 3´ ja 5´ fosfaatrühmad DNAst ja RNAst. Hüdrolüüsib NTPd ja dNTPd. Kasutatakse kloonimisel, et vältida korduvat ligeerimist (Aga seda raske väljalülitada, võib häirida ligeerimist). Võib kasutada ka dNTP lõhutamiseks PCR reaktsioonis, et valmistada DNA sekveneerimiseks või SNP analüüsiks.
Plaadis toimuvad konformatsioonilised muutused ning ta avaneb DNA väljutamiseks. ATP-d tarbiv tupe kontrakteerumine surub faagi pead basaalplaadi ja kiudude suunas. Saba südamikus olev toru läbib rakukesta, kuid mitte rakumembraani ning valkkatteta DNA siseneb läbi rakumembraani. Selleks, et faagi DNA oleks kaitstud rakusiseste nukleaaside eest, on ta modifitseeritud. Faagi paljunemistsükli erinevatel etappidel avalduvad erinevad geenid. Varajases infektsioonistaadiumis inaktiveeritakse faagi poolt kodeeritud valkude abil raku geenide transkriptsioon ja translatsioon. Bakteri RNA polümeraas modifitseeritakse nii, et ta tunneb ära faagi geenide promootoreid. Toimub ka intensiivne faagi genoomi paljundamine. Hilised geenid kodeerivad faagi kapsiidi valke ja faagi DNA-d lahtilõikavaid ensüüme. Lisaks basaalplaadis sisalduvale lüsotsüümile sünteesitakse ka lahustuvat lüsotsüümi, mis hävitab rakuseina.
· Patau 13 triploidne · Edward 18 triploidne · Turner üks X kromoosom puudu/vigane · Kleinfelter XXY · Triplo-X XXX 5. Kirjuta doosikompensatsiooni mehhanisme Geenide avaldumise regulatsioonimehhanism, mis tagab geenide võrdväärse avaldumise X-liiteliste geenide puhul homo- ja heterogameetsel sugupoolel. Erinevatel organismirühmadel on vastav mehhanism erinev. M. Lyon avastas, et emasimetajate somaatiliste rakkude kahest X- kromosoomist inaktiveeritakse üks varases embrüogeneesis fakultatiivse heterokromatinisatsiooni teel (rakutuumas tekib Barri kehake e. sugukromatiin). See inaktivatsioon on igas rakus homoloogiliste X- kromosoomide suhtes juhuslik, nii et emasisendi kudede erinevates rakkudes talitlevad erinevad homoloogid (seda nähtust nim. laionisatsiooniks). Selline mehhanism tagab seda, et emasorganismi rakkudes talitleb samamoodi üks koopia X-liitelisi geene, nagu hemisügootses isasorganismis, kuid emasisend
annustes, mis põhjustab regulaarseid koordineerituid kontraktsioone, mis liiguvad emaka kehast kaelani. Kontraktsioonide amplituud ja sagedus on sõltuvuses manustatud doosiga. Väikestes doosises emakas jõuab lõõgastuda kontraktsioonide vahe peal, suurtes doosides mitte. Suured annused on aga ohtlikud lootele. Oksütotsiin mõjutab ka rinnapiima ekspressiooni alveoolidesse ja juhadesse. Sellel on ka vasodilatoorne mõju. Mängib rolli ka mating ja parenting. Oksütotsiin inaktiveeritakse maksas ja neerudes ja veres ringleva platsentaalse oksütoksinaasiga. Kõrvalmõjud: Doosist sõltuv hüpotensioon, reflektoorne tahhükardia. Veepeetus ja hüponatreemia kui vee tarbimist ei jälgi. Dinoprostoon on PGE2 ja misoprostool PGE1 analoog. Prostaglandiinid soodustavad koordineeritud kontraktsioonide teket raseda emaka kehas ja samal ajal lõõgastab emakakaela. Raseduse varases faasis põhjustavad ka aborti. Dinoprostooni manustatakse geelina intravaginaalselt.
näpunäiteid. 2 Lisaülesanded (teoreetilised): 1.1) Vaata Fermentase kataloogist, milline on Cfr42I jaoks kõige sobivam puhver (viie puhvri süsteemis). Millistes puhvrites see ensüüm veel töötab? Lisaks tutvu kataloogis olevate tingmärkidega (millisel temperatuuril ensüümid töötavad, kuidas neid inaktiveeritakse, milline on antud restriktaasiga lõigatud fragmentide ligeerimise efektiivsus jne). Kõige sobivamad on puhver B, G ja Tango. Ensüümid töötavad 37 kraadi juures, inaktiveeruvad 65 kraadi juures, efektiivsus 95%, tundlik CpG metülatsioonile. 1.2) Kui restrikteerimiseks kasutada üheaegselt kahte erinevat ensüümi (näiteks Cfr42I ja PstI), siis milliseid puhvreid oleks kõige mõistlikum kasutada (vt Fermentase kataloogist restriktaaside tabelist)?
Piima koostisosade muutmine tootele soovitud omaduste saavutamiseks (kalgendi tugevus, sünerees); Bakterite arvu vähendamine loob eeldused piima edasiseks töötlemiseks erinevateks toodeteks. Piima koostiosade muutus kuumtöötlemisel Lipiidid suhteliselt termostabiilsed Laktoos- moodustub laktuloos; muudustuvad erinevad happed; toimub maillardi reaktsioon; moodustuvad lenduvad ühendid Vitamiinid- enamus vitamiine on suhteliselt termolabiilsed Ensüümid- enamus ensüüme inaktiveeritakse pastöriseerimisel Mineraalained muutused toimuvad vaid karbonaatide ja kaltsiumfosfaatidega Vadakuvalgud- madala termostabiilsusega; kõige termostabiilsem on a- laktalbumiin; kõige termostabiilsemad on immunoglobuliinid Kaseiinid- termostabiilne valgufraktsioon Pästoriseeritud piima tehnoloogiline skeem Toorpiima vastuvõtt ja säilitamine Puhastamine ja separeerimine Standardiseerimine (voolus või tankis)- Homogeeniseerimine 45-70 C Pastöriseerimine Jahutamine
Reaktsiooni võib alustada ka madalamal temperatuuril (näiteks 15 oC). Inkubeerimisaegadel lahendada allpool toodud lisaülesanne, vt lisamaterjale ja kuula juhendaja näpunäiteid. 2 Lisaülesanded (teoreetilised): 1.1) Vaata Fermentase kataloogist, milline on Cfr42I jaoks kõige sobivam puhver (viie puhvri süsteemis). Millistes puhvrites see ensüüm veel töötab? Lisaks tutvu kataloogis olevate tingmärkidega (millisel temperatuuril ensüümid töötavad, kuidas neid inaktiveeritakse, milline on antud restriktaasiga lõigatud fragmentide ligeerimise efektiivsus jne). Kõige sobivam puhver viie puhvri süsteemis Cfr 421 jaoks on : B(blue) 100 ja Tango (yellow) 50-100. Ensüüm veel töötab G(green) puhvris. Ensüümid töötavad 37 kraadi juures,lõigatud fragmentide ligeerimise efektiivsus on 95%. 1.2) Kui restrikteerimiseks kasutada üheaegselt kahte erinevat ensüümi (näiteks Cfr42I ja PstI), siis
● 45;X0 - monosoomia, kus X kromosoom on puudu - Turneri sündroom. 69. SRY geen Y- kromosoomi aheldunud geen, mis kodeerib valgulis testise moodustamise faktorit (SRY-valk), millel on võtmeroll isasorganismi arenemises ja suguorganite kujunemises. Asub Y- kromosoomi lühikeses õlas. Määrab suure osa testosterooni tootmise nii lootes kui ka hilisemas elus. 70. X - kromosoomi inaktivatsioon X kromosoomi inaktivatsioon on protsess, mille käigus inaktiveeritakse emastel imetajatel üks X kromosoom, võimaldades X-liiteliste geenide doosikompensatsiooni sugude vahel. Selle tulemusena avalduvad X-liitelised geenid võrdses koguses nii emastel kui isastel. Inaktivatsiooni protsess on suhteliselt universaalne kõigis imetajates, ent teatavad erinevused liigiti eksisteerivad.Barri kehake ehk sugukromatiin on tume keha inimese rakus, mis moodustub ühest kahest X-kromosoomist, mis inaktiveeritakse imetajatel embrüonaalse arengu väga varajases etapis
Raku pinnapealsed retseptorid-hüdrofiilne signaalmolekul Rakusisesed retseptorid- väikehüdrofoobne signaalmolekul kandja valgul liigub rakku ja seal tuuma,kus on intratsellulaarne retseptor. Nt steroidhormoonid Proteiinkinaaside ja -fosfataaside ning GTP seostumise ja hüdrolüüsi osa signaalide sisse ja väljalülitamises. Proteiin kinaasi poolt lisatakse fosfaat valgus oleva aminohappe hüdroksüülrühmale ning aktiveeritakse. Proteiin fosfataas eemaldab fosfori valgul ja inaktiveeritakse Valgul on endal GTPaasne aktiivsus –Valgul on GDP,kui sinna seondub GTP, sktiveerub valk. GTP hürolüüs inaktiveerib valgu GTP-d siduvate valkude e. GTPaaside toimemehhanism. GTPaas katalüüsib GTP hüdrolüüsi,mis inaktiveerib valgu. GTPaasi aktiveeriva valgu ja guaniin nukleotiidi vahetava faktori toimemehhanism. GAP – GTPaasi aktiveeriv valk suurendab GTPaasi hüdrolüütilist aktiivsust seega tekib GDP ning valk inaktiveerub
ning seal on mitmeid regulatoorseid geene, millest paljud on essentsiaalsed eluks. Y kromosoomil on aga vähe kodeerivaid geene, millest ükski pole essentsiaalne. Seepärast on X kromosoom vajalik kõikidele inimestele, mitte ainult naistele. Kuid naised ei saaks funktsioneerida, kui neil oleks topelt valke, mis tehtud kokku kahe X kromosoomi poolt, seega tuleb üks X kromosoom inaktiveerida (seda tehakse varases embrüogeneesis). Iga rakk valib ise juhuslikult, kumb X kromosoom inaktiveeritakse. Tihedalt kokku pakitud inaktiivses olekus X kromosoom on Barri kehake. Avaldumine fenotüübis – kuna nad sisaldavad võrdsel hulgal mõlemat tüüpi rakke – olles seetõttu X kromosoomi suhtes geneetilised mosaiigid, on nt emane, kes on heterosügootne mingi x-liitelise geeni suhtes võib omada kahte erinevat fenotüüpi – kass on ühest kohast valge ühest must ja ühest punane. 12. Tuumake: ehitus (piirkonnad, tuumakese organisaatori piirkonnad), koostis ja funktsioonid
Algab tärklise seede, sest süljes on amülaas (omnivooridel, lihtmaoga herbivooridel, mõnedel lindudel). Lipaas esineb noorloomadel (nt vasikal) – rasvade seede, see kaob vanusega. Sülje amülaasi ja lipaasi toime avaldub mao proksimaalses osas, sest suus ei ole toit piisavalt kaua. Mao proksimaalses osas pole näärmeid – pH on sobiv amülaasi jaoks ja see ei inaktiveerita. Mao distaalses osas on pH üsna madal ja süljeensüümid inaktiveeritakse. Ehk kohe kui toit makku jõuab, ei segata maonäärmetega. Liigilised erinevused Mäletsejalistel tugevalt aluseline - bikarbonaat (HCO3 - ) neutraliseerib suuõõnes ja vatsas bakterite toodetud happeid (kaitseb hambaemaili). Mäletsejalistel lisaks bikarbonaadile ka palju fosfaatioone – vatsakeskkonna puhverdamine ja toitaine mikroobide. Mäletsejaliste sülg sisaldab ka uureat – proteiini sünteesivate vatsabakterite toitaine. Ruminantide sülg on isotooniline, kõrge bikarbonaatide ja
oksüdatsiooni käigus vabaneb energia ning jääkainetena tekivad CO2 ja H2O. 3. Maksa – ja lihasrakkudes aktiveeritakse ensüümid, mis muudavad osa glükoosist glükogeeniks. See käib ka insuliini mõjul. 4. Aktiveeritakse ensüümid, mille toimel glükoosist tekkinud rasvhapped ja glütserofosfaadid muudetakse triglütseriidideks e varurasvaks. See on protsess on väljendunud eriti rasvkoe rakkudes. 5. Inaktiveeritakse need ensüümid, mis soodustavad triglütseriidide ja glükogeeni lammutamist. Reserve selles faasis ei võeta kasutusele (3-4 h pärast söömist). 6. Insuliini mõjul rakus suureneb aminohapetest valgusüntees. Insuliini sekretsioon hakkab langema siis kui vere glükoosi tase hakkab normipiiridesse jõudma. Kui glükoosi tase hakkab piirinormidesse jõudma või lähenema alumistele väärtustele (3,3 kanti), siis suureneb teise hormooni glükagooni produktsioon (glükagooni
tööd. Ajuripats toodab kasvuhormooni. Kui lapseeas eritub seda liiga palju, on tagajärjeks hiidkasv, kui aga liiga vähe, jääb inimene kääbuseks. Hormoonide toimemehhanism võib olla erinev: · nad võivad mõjutada rakumembraanide läbitavust; · rakusiseseid ensümaatilisi protsesse, neid aktiveerides või pärssides; · ensüümide sünteesi . Hormoonide toime sõltub nende kontsentratsioonist toimimispaigas. Pärast oma toime avaldamist hormoonid inaktiveeritakse. Neurohormoonidel toimub see sekundite jooksul, insuliinil paari tunniga ja kilpnäärme hormoonil türoksiinil nädala jooksul.
AA-st prostaglandiine ja tromboksaane. Lipooksügenaasne rada kasutab seda HPETE-de, leukotrieenide ja HETE-de biosünteesiks. (joonis, õpik II, lk 131, joonis 94). Prostaglandiinid (PG) on lokaalsed keemilised signaalmolekuulid, neid nimetatakse ka põletikumeidiaatoriteks. Nende toime on retseptor-vahendatud. PG puhul G- valkudega interakteeruvate retseptorite avil. PG-d mõju piirdub suhteliselt väikse rakurühmaga sest peale vabanemist haaratakse nad kiiresti rakkude poolt ja inaktiveeritakse. Keskne prostaglandiin on PGE2. Teda produtseeritakse inimorganismis praktiliselt kõikjal. Üldistatult võib PG-de füsioloogilised toimed kokku võtta järgmiselt: · Veresoonte permeaabluse regulatsioon ja seega ka vererõhu regulatsioon · Trombotsüütide agregatsiooni regulatsioon (seotud verehüübimisega). · PG osalevad põletikulise vastuse ning valuärrituse tekkes ja infektsioonide kulu moduleerimises.
tööd. Ajuripats toodab kasvuhormooni. Kui lapseeas eritub seda liiga palju, on tagajärjeks hiidkasv, kui aga liiga vähe, jääb inimene kääbuseks. Hormoonide toimemehhanism võib olla erinev: nad võivad mõjutada rakumembraanide läbitavust; rakusiseseid ensümaatilisi protsesse, neid aktiveerides või pärssides; ensüümide sünteesi . Hormoonide toime sõltub nende kontsentratsioonist toimimispaigas. Pärast oma toime avaldamist hormoonid inaktiveeritakse. Neurohormoonidel toimub see sekundite jooksul, insuliinil paari tunniga ja kilpnäärme hormoonil türoksiinil nädala jooksul.
[10] Tõhusalt funktsioneerivad parabeenid toodetes, mis on valmistatud vee baasil. Õlidel baseeruvate toodete puhul ei ole parabeenid nii efektiivsed (taimsetes õlides siiski efektiivsemad). Seega, parabeenid on pH tundlikud ja toimivad paremini happelises 12 Eesti Esimene Erakosmeetikakool Kristiina Meentalo Rahvusvaheline CIDESCO-kool 52. grupp keskkonnas. Parabeenid inaktiveeritakse osaliselt või täielikult tselluloosi derivaatide, proteiinide ja letsitiini poolt. [10] Negatiivne pool nende puhul on see, et nad põhjustavad allergiat ning kogunevad organismi, mõjutades inimese hormoonsüsteemi ning soodustades vähki. Parabeenid võivad avaldada ka östrogeenset mõju ning tekitada tundlikkust. Koostisainete nimekirjas märgitakse neid nimetusega, mille lõpus on “parabeen“. [12]
tööd. Ajuripats toodab kasvuhormooni. Kui lapseeas eritub seda liiga palju, on tagajärjeks hiidkasv, kui aga liiga vähe, jääb inimene kääbuseks. Hormoonide toimemehhanism võib olla erinev: · nad võivad mõjutada rakumembraanide läbitavust; · rakusiseseid ensümaatilisi protsesse, neid aktiveerides või pärssides; · ensüümide sünteesi . Hormoonide toime sõltub nende kontsentratsioonist toimimispaigas. Pärast oma toime avaldamist hormoonid inaktiveeritakse. Neurohormoonidel toimub see sekundite jooksul, insuliinil paari tunniga ja kilpnäärme hormoonil türoksiinil nädala jooksul.
Geenid mida nad mõjutavad on tuumas Ei ole seotud geenide või genoomi muutustega, vaid on puhtalt geeni ekspressiooni tulemus. Kui munarakk küpseb foliikulas, siis munaraku valgud, mida tsütoplasmas on terve rida, moodustuvad sellest munaraku genoomist endast. Need valgud, mis on sinna ennem tulnud, mõjutavad väga varast embüronaalset arengut ja selle tõttu võib tekkida muutus, mis ei allu II põlvkonnas Mendeli seadusele. Genoomne imprinting Inaktiveeritakse kogu eluks teatud geenid ühes homoloogis, et vältida liigset geeni ekspressiooni nn. monoalleelne ekspressioon. See toimub juba gameedis selle küpsemise käigus. Ei ole tegemist ema efektiga . Üks kromosoom inaktiveeritakse ära, et ei oleks meil üledoosi. Genoomne imprinting tähendab seda, et lookuses ühe gameedi alleel on inaktiveeritud ja seda tehakse gameedi küpsemise käigus. Imprintingu erinevus geeni ekspressioonist on see, et imprinting jääb organismile kuni elu lõpuni.
neuroni ja sihtmärkorgani vahel? Sümpaatilise närvisüsteemi retseptorid on (vereooned, seedetrakti silelihased, urotrakti silelihased, maks, silm), (presünaptiline membraan, trombotsüüdid, veresoonte silelihased) , ß(süda, neerud), (bronhioolid, veresooned, seedetrakt, maks, emakas). Neuroni ja sihtmärkorgani vaheliseks neurumediaatoriks on NA. Millised ensüümid lammutavad sümpaatililises närvisüsteemis noradrenaliini? Väiksem osa noradrenaliinist inaktiveeritakse mitokontrites paikneva MAO (monoaminooksüdaasi) ja tsütoplasmaatilise ensüümi COMT (katehhool-o-metüültransferaasi) poolt. Mis juhtub selle noradrenaliiniga, mida ensüümid ei lammuta? Presünaptilisest osast vabanenud noradrenaliin transporditakse suures osas tagasi presünapsisse ja hoiustatakse uuesti vesiiklites. Sümpaatilise närvisüsteemi retseptorite tüübid, asukohad ja mis toime aktivatsioonil?
b – altruisti sugulase ellujäämisvõimaluse suurenemine R – suguluskoefitsient, ühtlasi tõenäosus, et sugulane kannab seda altruistlikku mutatsiooni 27. Tooge selgitustega näiteid isekast DNA-st (meiootiline surve, transposoonid). Isekad geenid on geenid, mis moodustavad erandi Mendeli seadustele – heterosügootsed isendid ei tooda võrdsel hulgal kumbagi alleeli kandvaid sugurakke (st toodavad, aga üht alleeli kandvad rakud inaktiveeritakse). Meiootiline surve (meiotic drive) – toodetakse rohkem mõne geeni teatud alleele kandvaid sugurakke. Transposoonid – lühikesed genoomsed regioonid, mis võivad genoomis teistesse kohtadesse liikuda. Iseka DNA hulka kuuluvad need, mis teevad endast koopiaid, mis liiguvad mujale, ega kustuta algset koopiat, nt retrotransposoonid. 28. Milles seisneb valiku kinnitamise (reinforcement) mõju liigitekke puhul?
b – altruisti sugulase ellujäämisvõimaluse suurenemine R – suguluskoefitsient (õdedel-vendadel ½, poolõdedel-vendadel ¼ jne), ühtlasi tõenäosus, et sugulane kannab seda altruistlikku mutatsiooni 27. Tooge selgitustega näiteid isekast DNA-st (meiootiline surve, transposoonid). Marian ISEKAD GEENID: geenid, mis moodustavad erandi Mendeli seadustele – heterosügootsed isendid ei tooda võrdsel hulgal kumbagi alleeli kandvaid sugurakke (st toodavad, aga üht alleeli kandvad rakud inaktiveeritakse). Meiootiline surve (meiotic drive) – toodetakse rohkem mõne geeni teatud alleele kandvaid sugurakke. Nt koduhiire (Mus musculus) kromosoom 17 t-haplotüübi suhtes heterosügootsed isased toodavad u 90% t-haplotüüpi kandvaid sperme. t-haplotüüp peaks fikseeruma → ära hoiab see, et t-homosügoodid on steriilsed. Transposoonid – lühikesed genoomsed regioonid, mis võivad genoomis teistesse kohtadesse liikuda. Iseka
Kontroll RNA transpordi tasemel kontrollitakse, milliseid mRNA molekule viiakse tsütoplasmasse, kus toimub valkude süntees. Kontroll translatsiooni tasemel kontrollitakse, milliste tsütoplasmas leiduvate mRNA molekulide abil toodetakse valke ribosoomides. Kontroll mRNA degradatsiooni tasemel selleks lagundatakse selektiivselt mõnesid mRNA molekule tsütoplasmas. Posttranslatsiooniline kontroll selleks selektiivselt aktiveeritakse või inaktiveeritakse toodetud valke või transporditakse neid erinevatesse raku osadesse. 26. Vähirakud. Põhjused, miks rakkude jagunemine ei ole enam kontrollitav. Embrüo rakkudes ja mõnedes tüvirakkudes muudab telomeraas telomeeri struktuuri sedavõrd, et rakk saab jaguneda kauem. Ent ka neil juhtudel aja jooksul telomeraasi hulk väheneb ja lõpuks rakkude jagunemine peatub. Vähirakkudes aga jätkab telomeraas telomeeride struktuuri muutmist palju pikema perioodi jooksul kui
A nukleotiidi analoog, seondumine toimub suurele subühikule. Antibiootikumiresistentsuse kolm mehhanismi 1. Kui meil mingi AB märklaud (ribosoom, topoisomeraas jne), kuid selle märklaua geenis toimub mingi muutus AB ei saa enam oma märklaulale seonduda. Tekib geeniresistentne variant. 2. Bakter omandab pumba, millega AB bakterirakust välja pumpab ja vähendab AB-de kontsentratsiooni rakus. 3. AB lihtsalt inaktiveeritakse. Nt penitsiliini puhul lõigatakse AB hüdrolüütilise ensüümi poolt katki. Mõnede teiste puhul ka nt fosforüleerimine, metüleerimine jne. Geneetiline kood. Koodoni wobble positsioon (inosiin ja tRNA modifitseerimine). Kolmandas positsioonis on koodoni kasutus lõdvem, tänu millele võib tekkida seal rohkem vigu. Koodoni 1 ja 2 täht olulisemad (seos ribosoomi dekodeeriva tsentriga). Kolmandas positsioonis on puriin või pürimidiin
Ajuripats toodab kasvuhormooni. Kui lapseeas eritub seda liiga palju, on tagajärjeks hiidkasv, kui aga liiga vähe, jääb inimene kääbuseks. Hormoonide toimemehhanism võib olla erinev: · nad võivad mõjutada rakumembraanide läbitavust; · rakusiseseid ensümaatilisi protsesse, neid aktiveerides või pärssides; · ensüümide sünteesi . Hormoonide toime sõltub nende kontsentratsioonist toimimispaigas. Pärast oma toime avaldamist hormoonid inaktiveeritakse. Neurohormoonidel toimub see sekundite jooksul, insuliinil paari tunniga ja kilpnäärme hormoonil türoksiinil nädala jooksul. 5
transmembraanse poori, mis viib märklaudraku lüüsini. · Alternatiivne - ei vaja antikeha aktivatsiooniks. Antigeenid on vajalikud selle tee aktivatsiooniks. Nagu ka klassikaline nii ka alternatiivne tee produtseerib C3 ja 5 konvertaasi, mis viib C5b ja seejärel MACi seondumiseni. Komplement C3 jaguneb spontaalselt, mis tähendab, et C3b ja a hulbivad vabalt seerumis. Seega C3b võib rünnata erinevaid pindu, kaasaarvatud peremehe omi. C3b inaktiveeritakse kiirelt. Membraani küge seotakse Cb3 komplement faktor B abil, mis seejärel jaguneb faktor D-ks. Ba hulbib minema, Bb jääb seotuna C3b. See kompleks on alternatiivse raja C3 konvertaas, ja ei ole väga stabiilne, vajab properdiini, mis siis seondub selle kompleksiga. Konvertaas käitub analoogselt klassikalise raja konvertaasiga, lõigades C3 molekulid C3aks ja bks. Moodusutb C5 konvertaas ja samamoodi sarnaselt klassikalise rajaga moodustub lõpuks MAC. · letiini tee.
Kontroll RNA transpordi tasemel kontrollitakse, milliseid mRNA molekule viiakse tsütoplasmasse, kus toimub valkude süntees. Kontroll translatsiooni tasemel kontrollitakse, milliste tsütoplasmas leiduvate mRNA molekulide abil toodetakse valke ribosoomides. Kontroll mRNA degradatsiooni tasemel selleks lagundatakse selektiivselt mõnesid mRNA molekule tsütoplasmas. Posttranslatsiooniline kontroll selleks selektiivselt aktiveeritakse või inaktiveeritakse toodetud valke või transporditakse neid erinevatesse raku osadesse. 26. Vähirakud. Põhjused, miks rakkude jagunemine ei ole enam kontrollitav. Embrüo rakkudes ja mõnedes tüvirakkudes muudab telomeraas telomeeri struktuuri sedavõrd, et rakk saab jaguneda kauem. Ent ka neil juhtudel aja jooksul telomeraasi hulk väheneb ja lõpuks rakkude jagunemine peatub. Vähirakkudes aga jätkab telomeraas telomeeride struktuuri muutmist palju pikema perioodi jooksul kui
implantatsiooni. Imprinditud geenid säilitavad oma metülatsiooni märgiseid läbi kogu reprogrammeerimise lubades vanema spetsiifilist ekspressiooni pärinemist somaatilistes kudedes. • • Geenidoosi kompensatsioon epigeneetiliste mehhanismide abil: X- inaktivatsioon. • Geenidoosi kompensatsioon tagab X-liiteliste geenide võrdväärse avaldumise emastel ja isastel – emaste üks X kromosoom inaktiveeritakse ning tekib Barri kehake. X kromosoomi inaktivatsioon toimub läbi CpG metülatsiooni ja histoonide modifikatsioonide, need viivad kromatiini kondenseerumiseni, inaktivatsioon on juhuslik (kõik emased mosaiiksed). X inaktivatsioon toimub Xist transkripti kaudu, mis on lcnRNA – Xist katab terve inaktiveeritud X-kromosoomi. • • Geenide vanema-spetsiifiline ekspressioon ja vaigistamine (imprinting): levimine organismides, mõistete sisu ja mehhanismid.
käigus ulatuslikke geneetilisi ümberkorraldusi. Mu faagi välja tulekul haaratakse faagi genoomiga kaasa ka külgnevat DNAd. Mu faagiga nakatunud bakterid sisaldavad palju erinevaid mutatsioone. T4 nukleiinhape on modifitseeritud, kaitsmaks teda faagi-spetsiifiliste nukleaaside ja rakuliste restriktaaside eest. Struktuursed osad (pea, saba, fiibrid) moodustuvad teineteisest sõltumatult. Infektsiooni tagajäjel inaktiveeritakse raku geenide transkriptsioon ja translatsioon. Vastavalt avaldumise ajale klassifitseeritakse geenid: varajased, keskmised ja hilised geenid. Levioni geenid: kapsiidivalk, RNAst sõltuv polümeraas, replikaas, lüüsivalk, A-valk (antiretseptor)
Serooskest Välimiseks kestaks moodustab sidekoeline proopria, mida katab ühekihiline lameepiteel – mesoteel Epiteelirakud on madalad, sakiliste rakupiiridega ning rakus võib olla kaks tuuma MAKSA EHITUS. MAKSASAGARIK. Punakaspruuni värvi serooskestaga kaetud kompaktne organ Seedesüsteemi kõige suurim organ ja loomaorganismi suurim nääre, funktsioonid väga laialdased Osaleb organismi ainevahetuses, kaitsereaktsioonides, seal inaktiveeritakse hormoone ja ravimeid Polüsahhariid glükogeeni moodustumine, milline mängib olulist rolli stabiilse glükoosi taseme hoidmisel veres Seal salvestatakse ka rasvu, samuti erinevaid vitamiine Maksa katab pealt serooskest, selle alla jääb fibrooskihn, mis ümbritseb maksaväratis paiknevaid veresooni, sapijuhasid ning maksavärati kaudu kulgevad väga õhukesed sidekoelised väädid parenhüümi Organi parenhüümi moodustavad maksa sagarikud
märklaudraku lüüsini. Alternatiivne tee ei vaja antikeha aktivatsiooniks. Antigeenid on vajalikud selle tee aktivatsiooniks. Nagu ka klassikaline nii ka alternatiivne tee produtseerib C3 ja 5 konvertaasi, mis viib C5b ja seejärel MACi seondumiseni. Komplement C3 jaguneb spontaalselt, mis tähendab, et C3b ja a hulbivad vabalt seerumis. Seega C3b võib rünnata erinevaid pindu, kaasaarvatud peremehe omi. C3b inaktiveeritakse kiirelt. Membraani küge seotakse Cb3 komplement faktor B abil, mis seejärel jaguneb faktor D-ks. Ba hulbib minema, Bb jääb seotuna C3b. See kompleks on alternatiivse raja C3 konvertaas, ja ei ole väga stabiilne, vajab properdiini, mis siis seondub selle kompleksiga. Konvertaas käitub analoogselt klassikalise raja konvertaasiga, lõigades C3 molekulid C3aks ja bks. Moodusutb C5 konvertaas ja samamoodi sarnaselt klassikalise rajaga moodustub lõpuks MAC. 30
1h. Bakterid jagunevad kiiremini kui 1 h. Mitmekorruseline replikatsioon tagab selle, et DNA süntees jõuab jagunemisele järele. DNA süntees sõltub de novo valgu sünteesist. Eukarüootide replikatsiooni initsiatsioonil osalevad valgud. Kui DNA polümeraas kohale jõuab, hakatakse DNA-d sünteesima. Cdk on oluline regulaator – stimuleerib ja pidurdab replikatsiooni algust. Kui esineb Cdk aktiivsus, siis pre-RC kompleksi ei aktiveerita. Eukarüootide replikatsiooni käigus ORI-d inaktiveeritakse. Esineb palju alguspunkte (ORI-sid). ORI-d on seotud DNA järjestusega, iseseisvalt ei toimi. DNA replikatsioon toimub vaid ühe korra, vajatakse ainult ühte koopiat. DNA alguspunktide inaktiveerimiseks peab valk olema seondunud ja see eemaldab alguspunktid. Pre-RC – pre-replikatsiooni kompleks, mis koosneb ORC-dest. Moodustab selle 6 eri polüpeptiidist. Eukarüootide replikatsiooni lõpetamine on seotud terminatsioonisaidiga – järjestus on polaarne
..60% kogu ööpäevasest energiakulust. Polüsahhariidid (tärklis, glükogeen, tselluloos), disahhariidid (sahharoos, laktoos, maltoos), monosahhariidid ( glükoos, fruktoos). Süsivesikud imenduvad monosahhariididena. Kiudained lõhustatakse osaliselt alles jämesoole bakterite poolt. Kiudained seovad rohkesti vett, stimuleerivad soole motoorikat. Sülje ja kõhunäärme - amülaas lõhustab polüsahhariidid kuni disahhariidi maltoosini. Amülaas inaktiveeritakse maos soolhappe toimel, disahhariidid jõuavad lõhustamatult peensoolde. Süsivesikute lõhustamise lõpetavad peensoole limaskesta epiteelirakkudes tekkivad ensüümid disahharidaasid: maltaas, sahhraas, laktaas. Aineid transporditakse läbi raku, s.o transtsellulaarselt. Enterotsüüdi plasmamembraani apikaalsetes ja basolateraalsetes piirkondades on erinevad kandurid e transporterid. Monosahhariidide kergendatud difusioonis osalevad kandurid kuuluvad GLUT perekonda.
riini poolt valguse produktsioonil, mis võimaldab ATP olemasolu ning kogust registreerida. Reaktsioon toimub minutitega ja seda võib kasutada kiire hügieeni- testina, kus ATP esinemine toidutöötlusruumides on ebaadekvaatse sanitatsiooni (puhastamine, pesemine, desinfitseerimine) selgeks näitajaks. ATP sondeerimist on 38 võimalik kasutada ka bakterite arvukuse määramisel. Selle meetodi puhul lüüsi- takse kõigepealt loomse või taimse päritoluga rakud ning inaktiveeritakse vabane- nud ATP. Seejärel lüüsitakse bakterirakud ning määratakse kindlaks nendest vaba- nenud ATP, mille kontsentratsioon on proportsionaalne analüüsitavas piirkonnas olnud bakterite arvuga. Seda meetodit kasutatakse ka rümpade ja lihatoodete hügieenilise seisundi hindamiseks, kaasa arvatud juhtudel, mil tegemist on väga madala bakterite arvukusega. Et tegemist on ATP määramisega, siis on tulemuste reprodutseeritavus selle meetodi kõige suuremaks puuduseks
Raku sees aktiveeritakse ensüümid, mis soodustavad glükoosi oksüdatsiooni ja selle oksüdatsiooni käigus vabaneb energia ning jääkainetena tekivad CO2 ja H2O. 3. Maksa ja lihasrakkudes aktiveeritakse ensüümid, mis muudavad osa glükoosist glükogeeniks. See käib ka insuliini mõjul. 4. Aktiveeritakse ensüümid, mille toimel glükoosist tekkinud rasvhapped ja glütserofosfaadid muudetakse triglütseriidideks e varurasvaks. See on protsess on väljendunud eriti rasvkoe rakkudes. 5. Inaktiveeritakse need ensüümid, mis soodustavad triglütseriidide ja glükogeeni lammutamist. Reserve selles faasis ei võeta kasutusele (3-4 h pärast söömist). 6. Insuliini mõjul rakus suureneb aminohapetest valgusüntees. Insuliini sekretsioon hakkab langema siis kui vere glükoosi tase hakkab normipiiridesse jõudma. Kui glükoosi tase hakkab piirinormidesse jõudma või lähenema alumistele väärtustele (3,3 kanti), siis suureneb teise hormooni glükagooni produktsioon (glükagooni
ELISA), PCR ja DNA sondid. Seroloogiat kasutatakse harva, ainult epidemioloogilistel kaalutlustel. Isoleerimiseks rakukultuurid. Histoloogiliselt rakkudes iseloomulikud tuumasisesed inklusioonid (esinevad harva, vaja eristada CMV-st). Ravi ja profülaktika. Ravi puudub. Olemas suukaudne elusvaktsiin, mida USA sõjaväes kasutatakse, tsiviilidel mitte. Adenoviiruseid kasutatakse geenisiirdamisvektoritena mitmete inimese haiguste raviks. Viirus inaktiveeritakse E1 ja teiste viirusgeenide muteerimisega, siis sisestatakse vajalik geen, mis viiakse viirusega inimese raku genoomi, kus ta õigesse kohta sisestub. Aga vaatamata muteerimisele, võib ikka haigust põhjustada. Viirused mõmm :) 05/06 Herpesviirused: VZV, EBV, CMV, HHV 6 ja HHV 7, HHV 8 Struktuur. Suured ümbrisega kaksikahelalise DNA-ga viirused. Virion 150 nm
rakutsükli kulgemises. CDK katalüütiline kompleks on iseenesest inaktiivne ning peab ühinema tsükliiniga ja vajab teroniinjäägi fosforüliseerimist, et täielikult aktiviseeruda. CDK-tsükliinkompleks desaktiveeritakse kas türosiinjäägi, mis asub adenosiintrifosfaatsideme piirkonnas, fosforüliseerimisega või ühinemisel tsükliinkinaasi inhibeerivate proteiinidega. Peale täielikku rakutsüklit kompleks inaktiveeritakse pöördumatult mediaator ubikitiini poolt degradeerides kompleksist tsükliini. S-faasi minekut kontrollitakse tsükliinide D; E; ja A poolt. Molekulaarsete kontrollpunktide geenid p34 Kiirguskahjustuse liigid Surmav kahjustus subletaalne kahjustus potensiaalselt letaalne kahjustus Doositaseme toime Kiirguse bioloogilise toime tagajärjed ja risk Keskmine aastane kiirgusdoos Radoon 50% Maakoor 14%
FSH produktsioon aga langeb sellest momendist, kui algab hCG süntees ja jääb ülimalt madalatele väärtustel kogu raseduse vältel. Prolaktiini produktsioon tõuseb kogu raseduse vältel pidevalt. Hüpofüüsi tagasagaras toodetava oksütotsiini hulk suureneb raseduse lõpul ja sünnituse ajal. Oksütotsiinil on emakakontratsioone stimuleeriv toime. Oksütotsiini väärtused jäävad suhtelisel kõrgeks ka imetamisperioodil. Raseduse ajal inaktiveeritakse oksütotsiin platsenta ensüümide toimel. Ovaariumid Raseduse ajal folliiklite arengut lõpliku küpsuseni ei toimu. Ovulatsioonid on lakanud. Raseduse esimesed kolm kuud funktsioneerib raseduspuhune kollaskeha, mis kindlustab vajaliku progesterooni sünteesi. Edaspidi jätkub selle hormooni süntees platsentas. Neerupealised Neerupealiste verevarustus raseduse ajal paraneb ning kortikosteroidide süntees suureneb. See on vajalik valgu- ja süsivesikute ainevahetuse reguleerimiseks