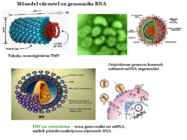
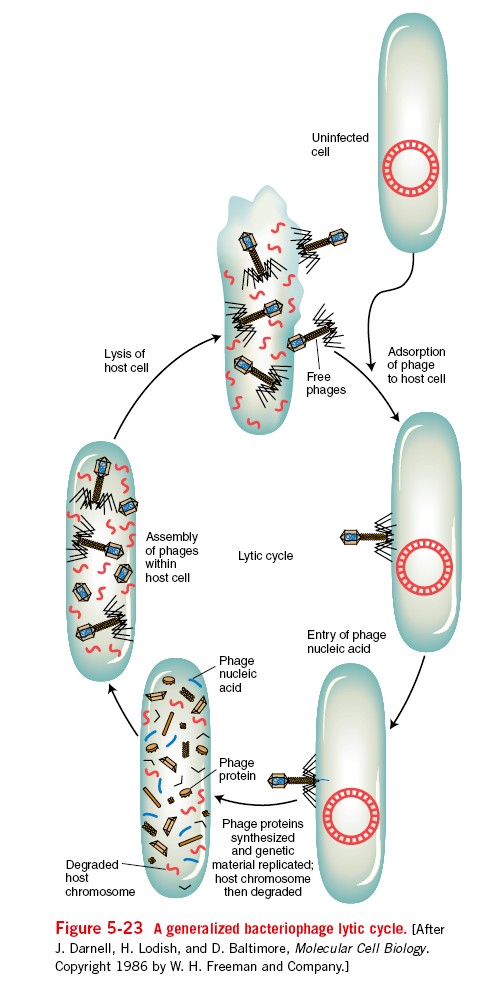
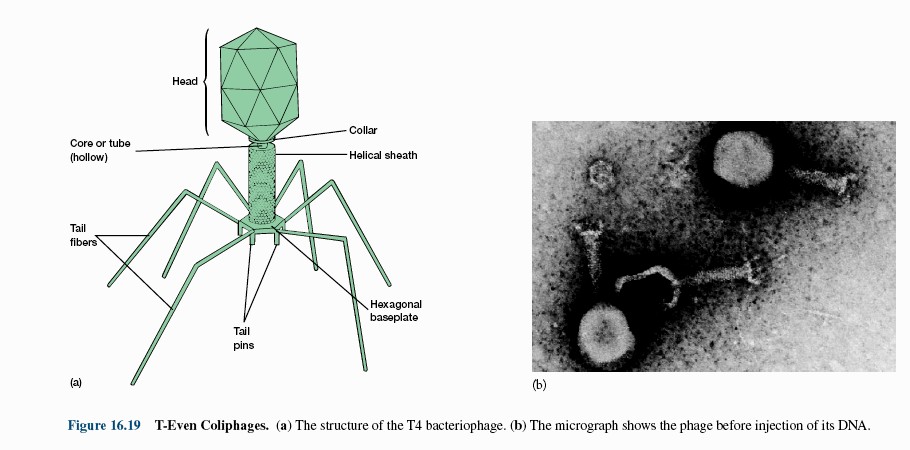
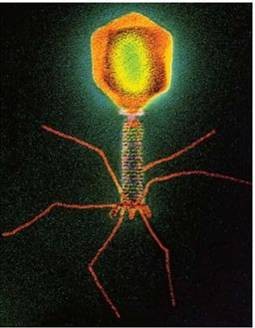
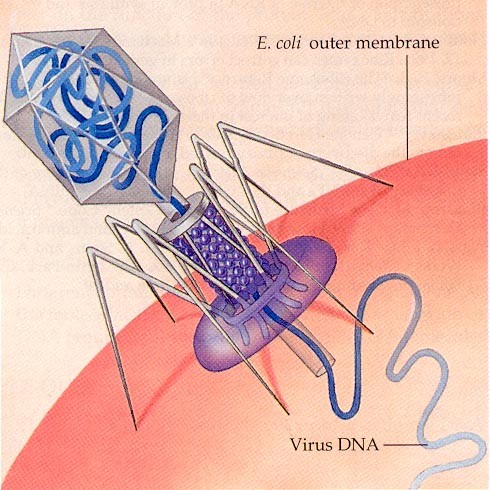
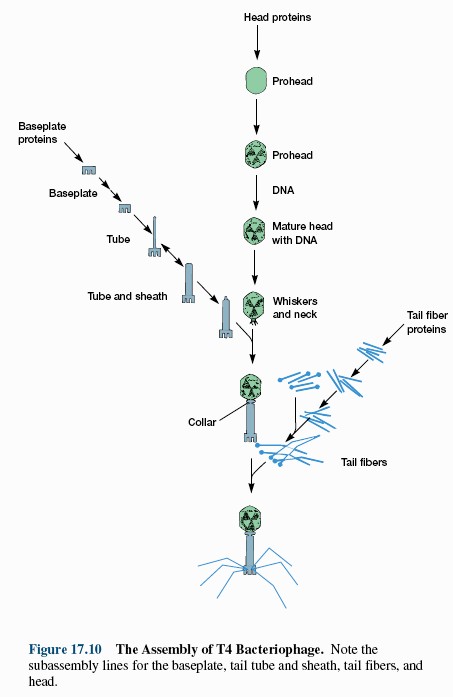
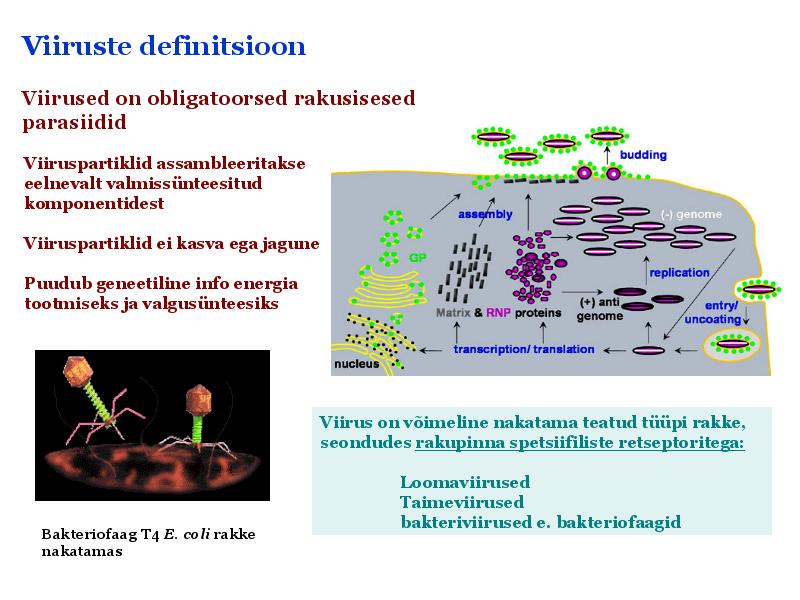
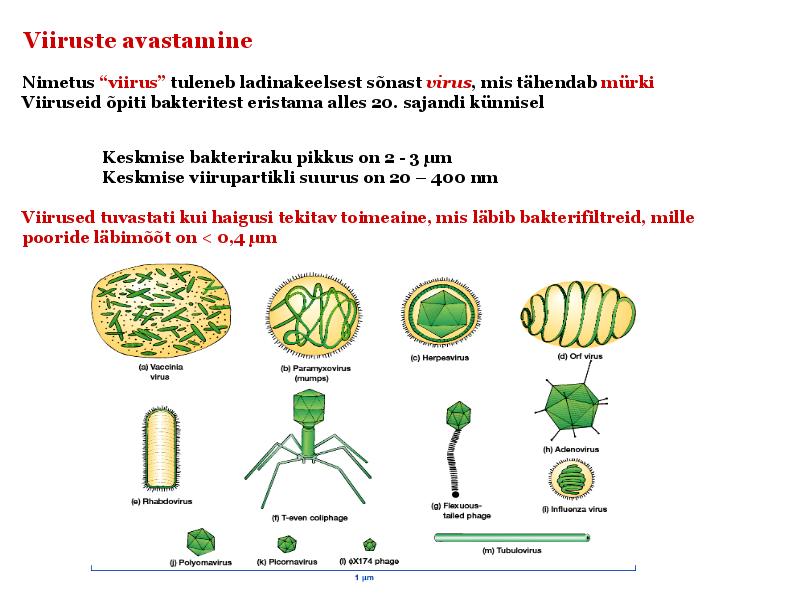
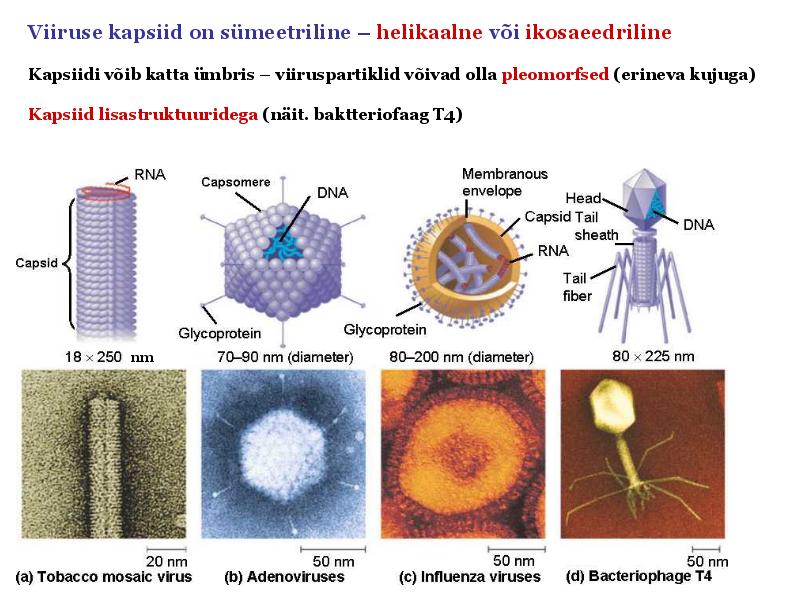
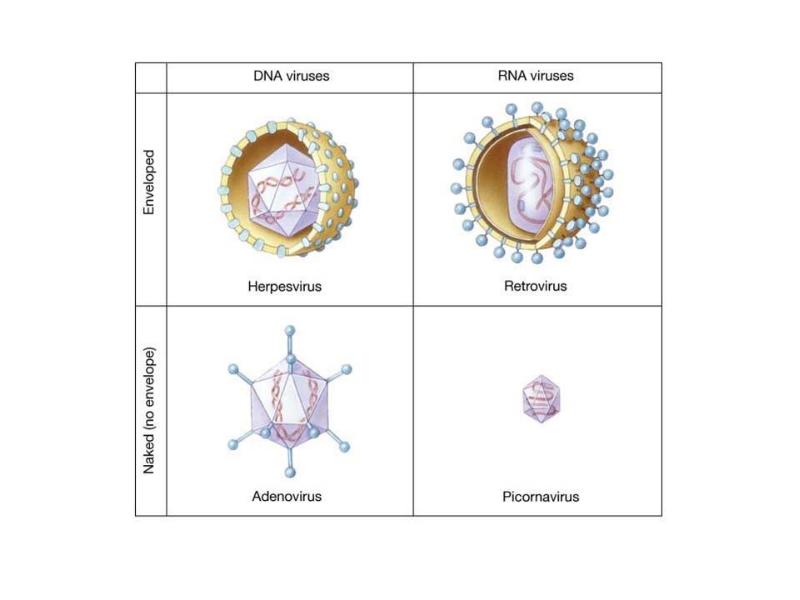
15. Viiruste geneetika Viiruste definitsioon. Viirused on obligatoorsed rakusisesed parasiidid. Samas on rakusisesed parasiidid ka mitmed prokarüootsed organismid, näit. bakterid perekonnast Rickettsiae ja Chlamydiae, mis suudavad rakuväliselt eksisteerida ainult väga lühiajaliselt. Seetõttu on viiruste defineerimiseks vaja juurde tuua veel lisaparameetrid: 1) Viiruspartiklid assambleeritakse eelnevalt valmissünteesitud komponentidest 2) Viiruspartiklid ei kasva ega jagune 3) Puudub geneetiline info energia tootmiseks ja valgusünteesiks. Viiruste spetsiifika: Iga viirus on võimeline nakatama ainult teatud tüüpi rakke. Vastavalt sellele klassifitseeritakse viiruseid loomaviirusteks, taimeviirusteks ning bakteriviirusteks e. bakteriofaagideks. Viirusega nakatamiseks on
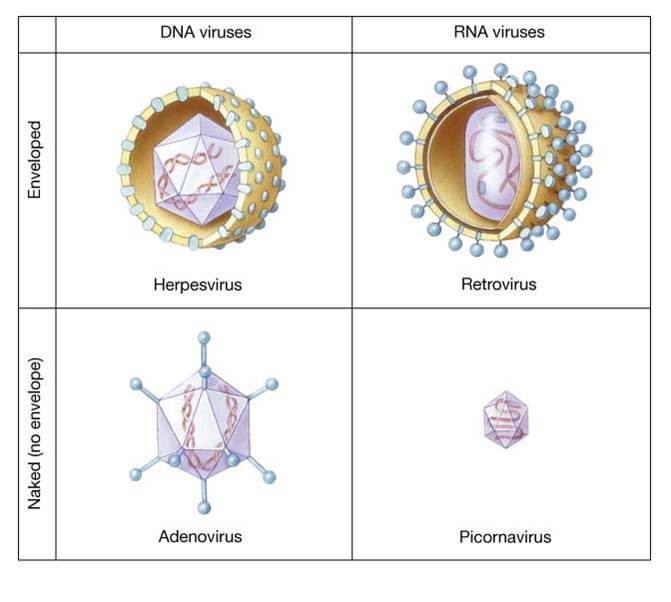
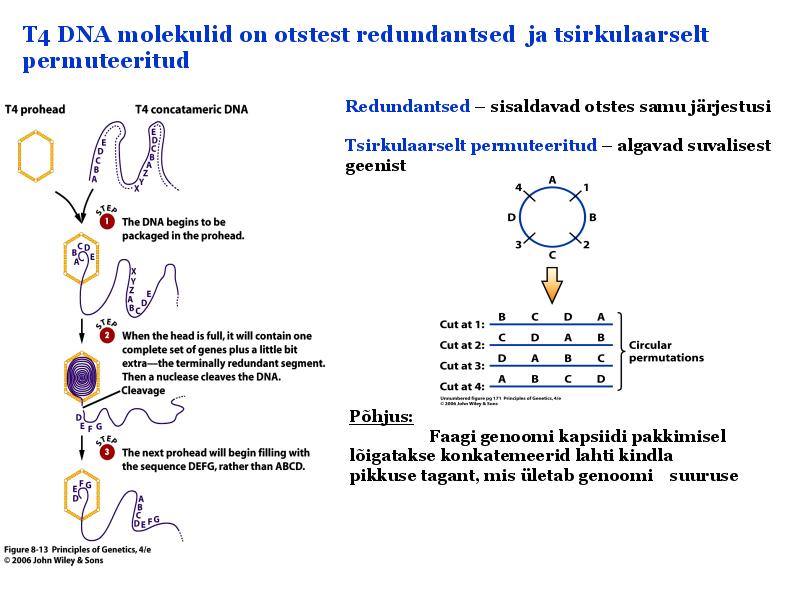
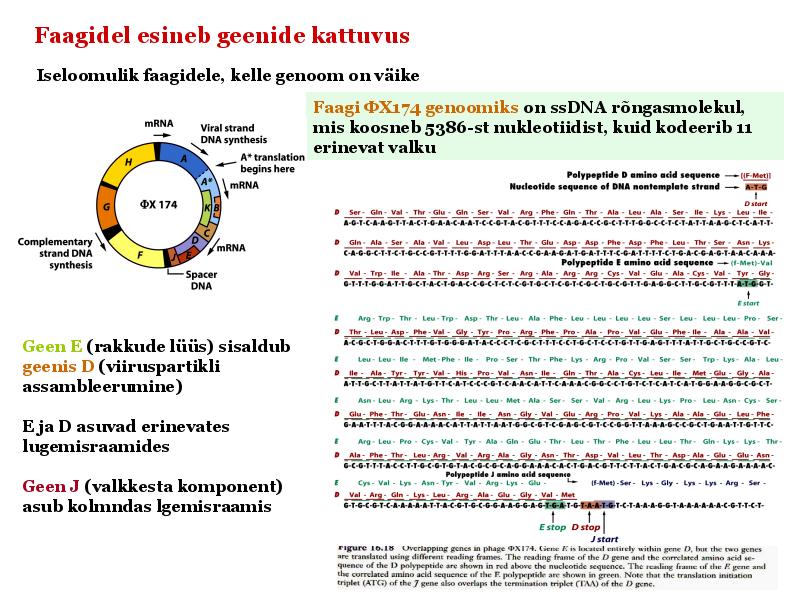
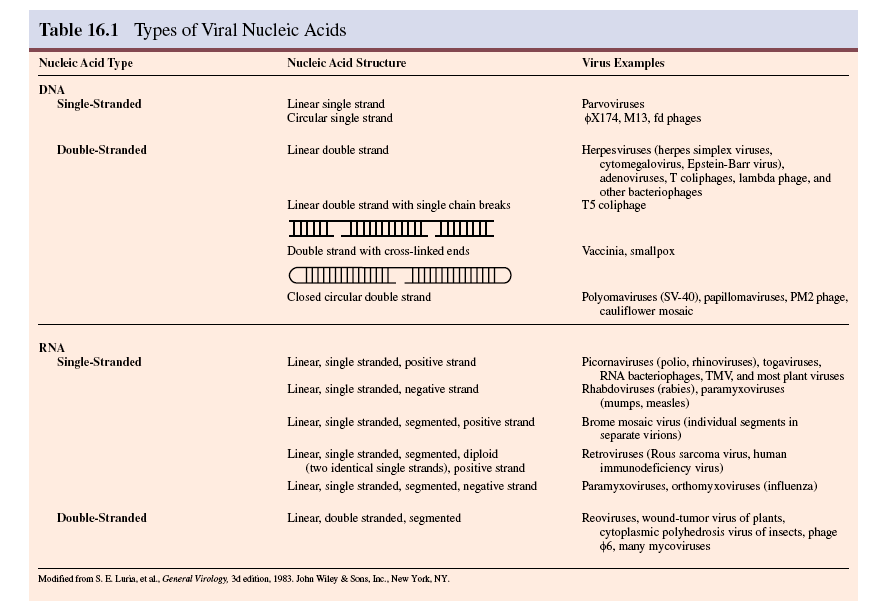
Kordamisküsimused geneetikas loeng 7 kohta: 1. Iseloomusta viiruste eripära? - Viirused on mitterakulise ehitusega obligatoorsed rakusisesed parasiidid. Definitsiooni juurde käib veel kitsendusena: viirusosad pannakse kokku eelnevalt rakus sünteesitud komponentidest viirused ei kasva ega jagune viiruse genoomis puudub geneetiline info energia tootmiseks ja valkude sünteesiks. Esineb DNA (kaksik- ja üksikahelalise) ja RNA (üksik- ja kaksikahelalise) genoomiga viirusi. viirustel on üks erineva suurusega kromosoom
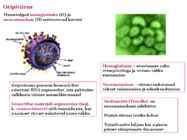
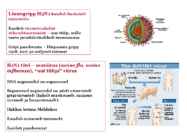
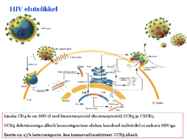
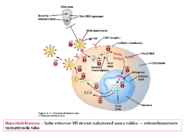
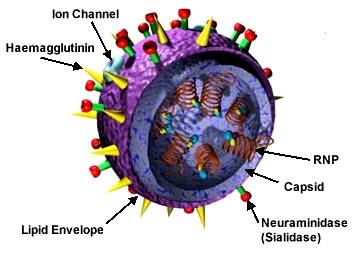
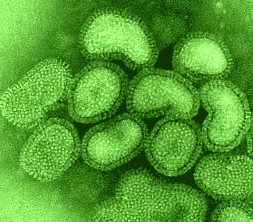
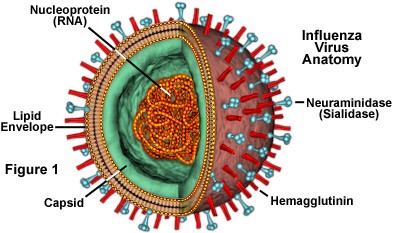
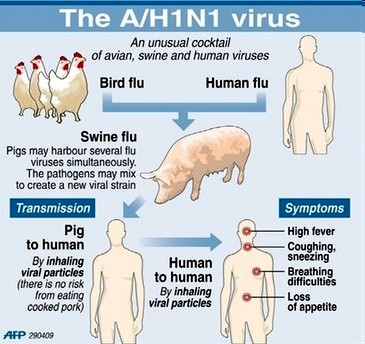
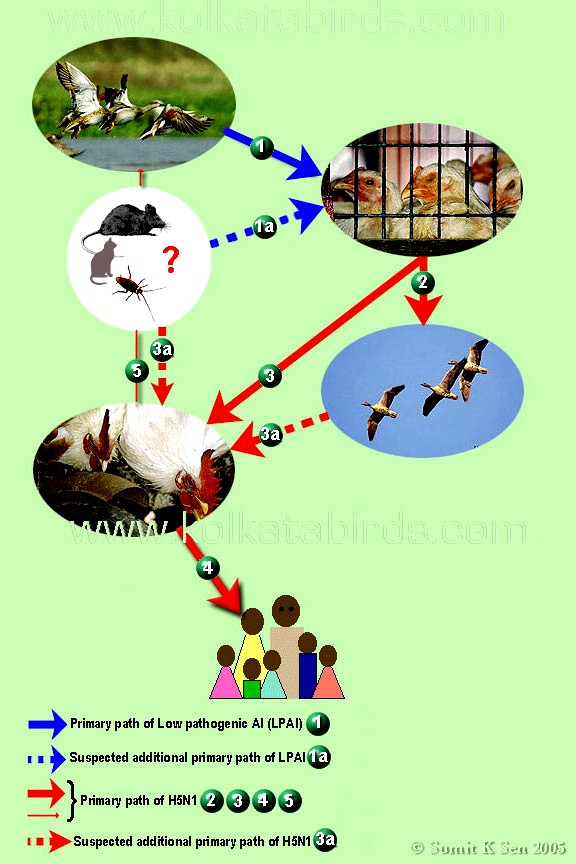
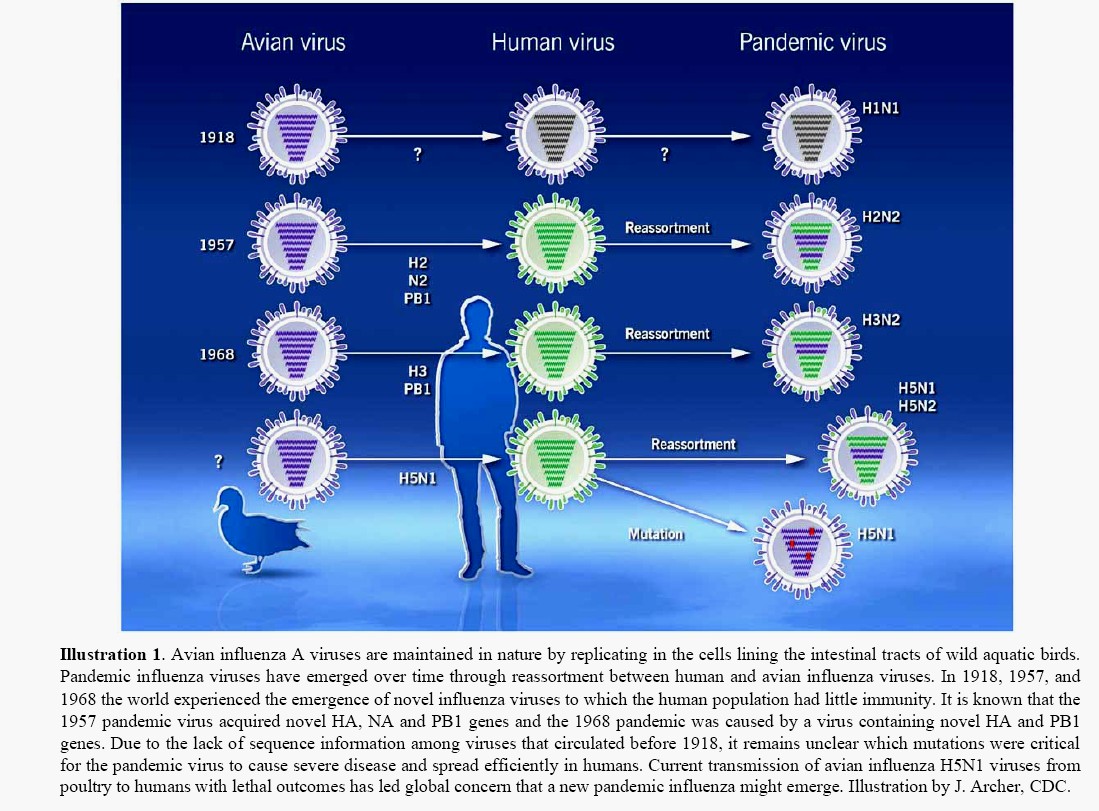
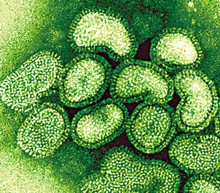
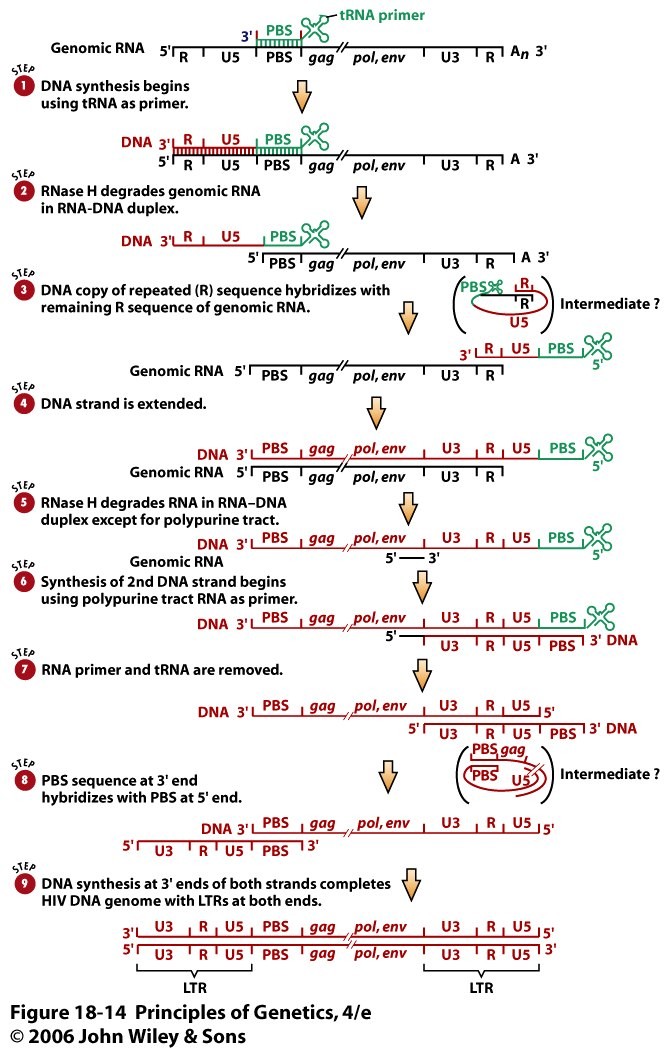
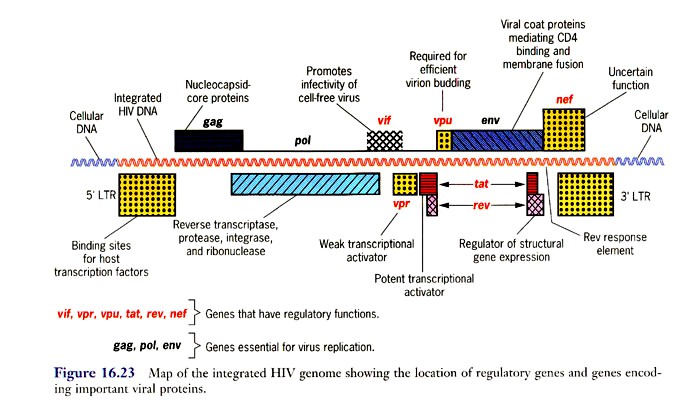
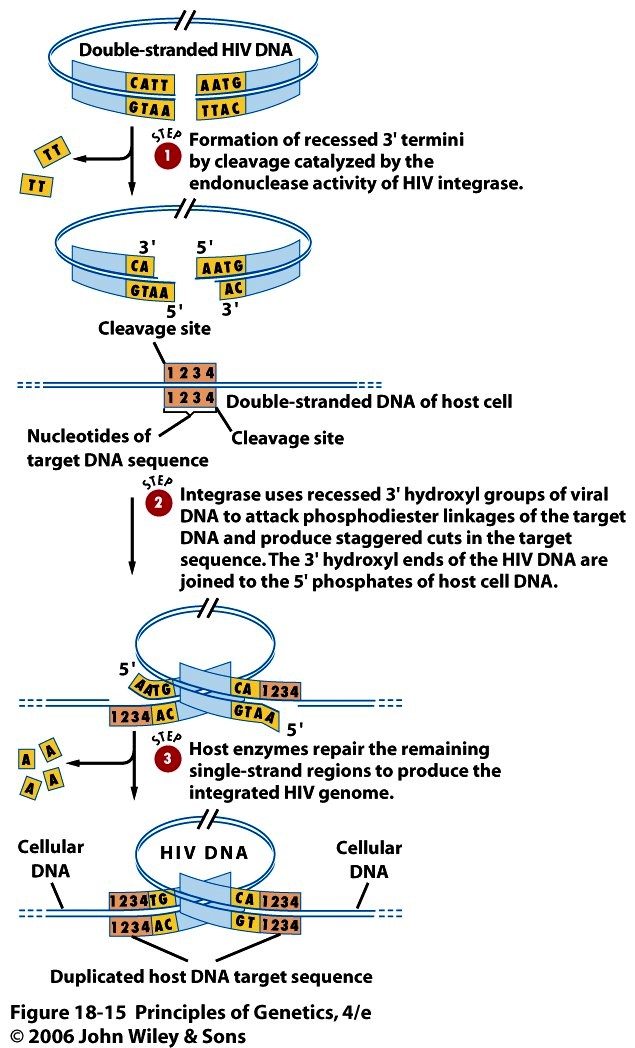
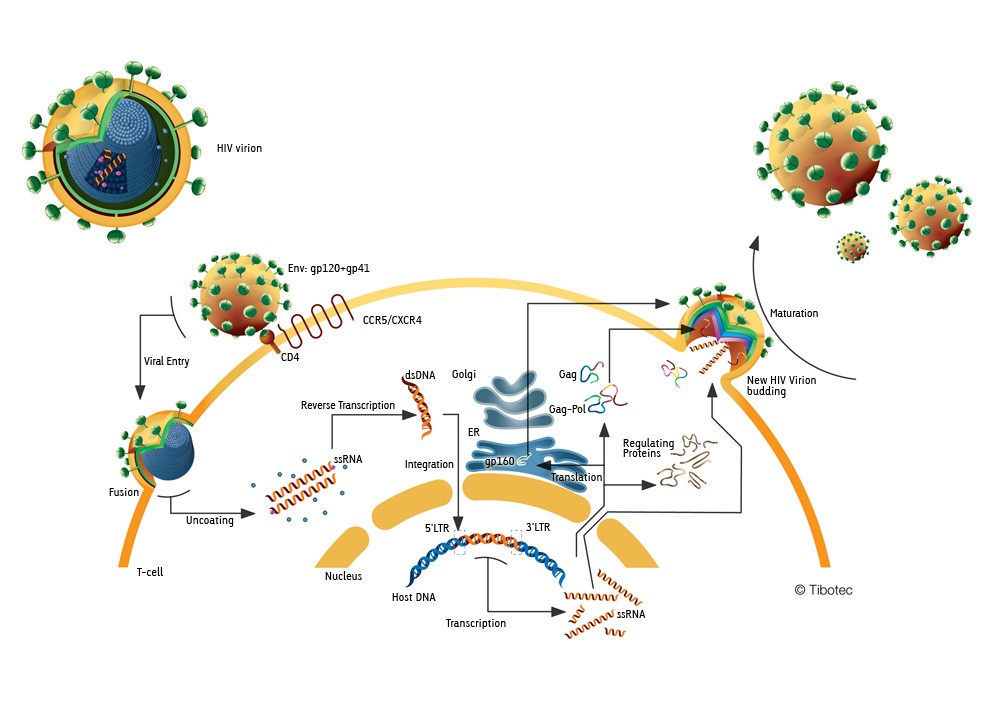
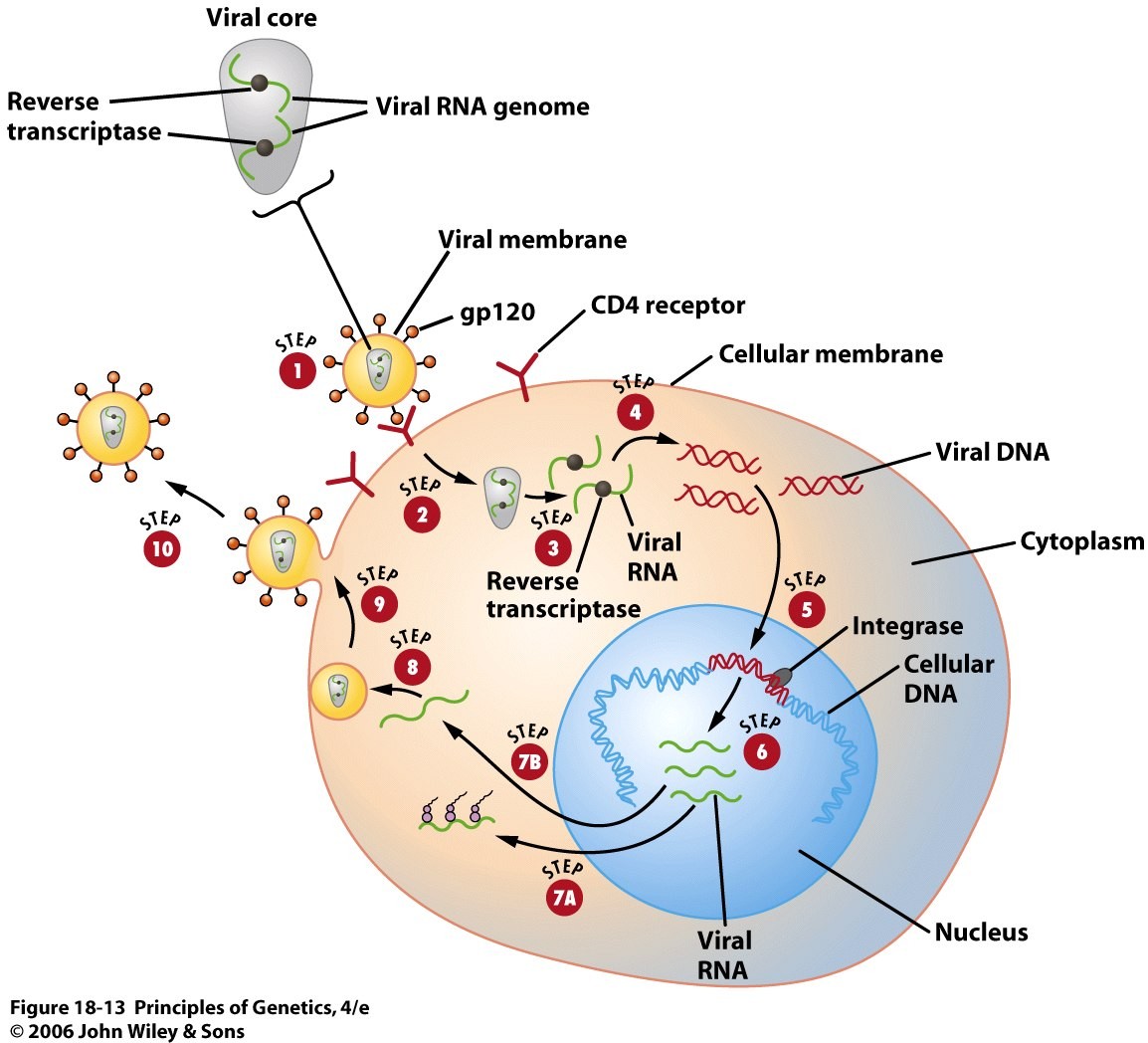
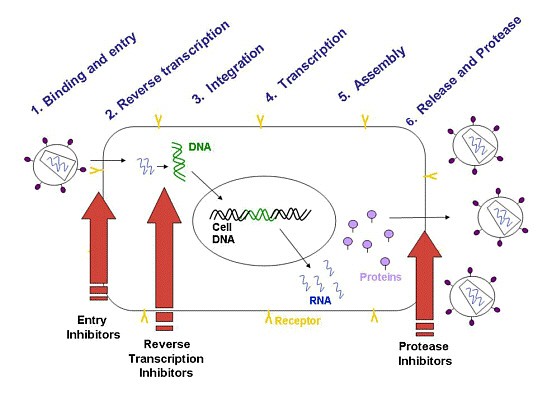
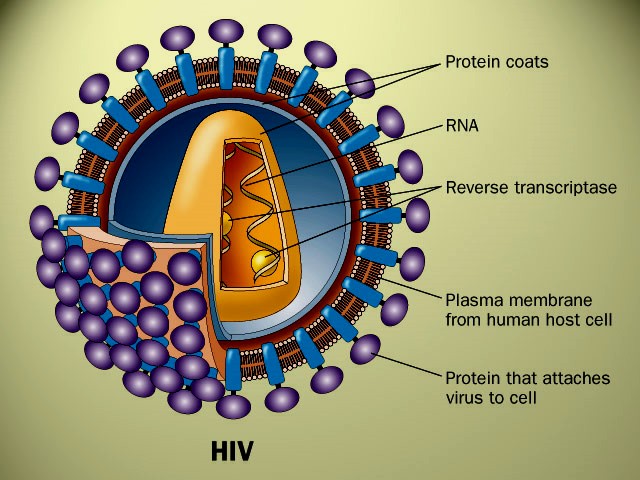
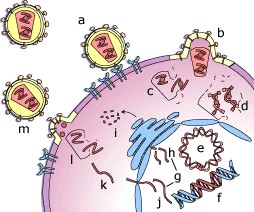
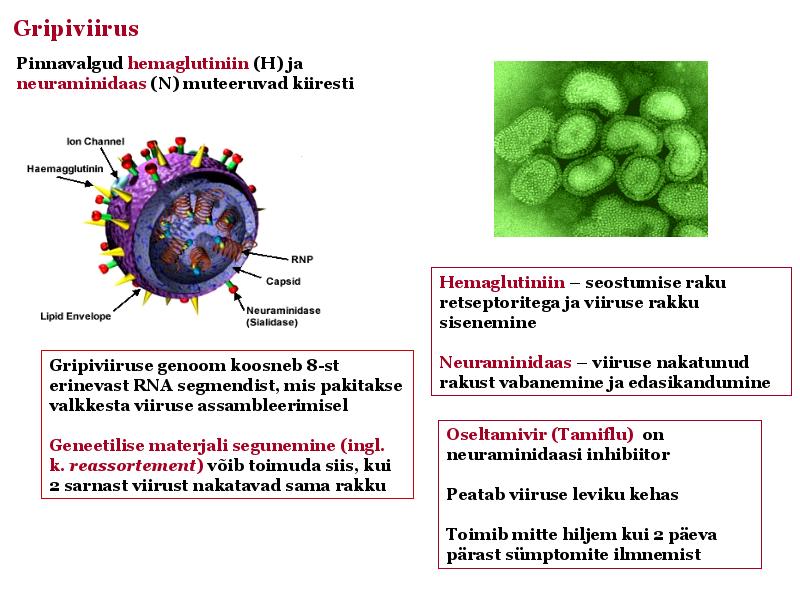
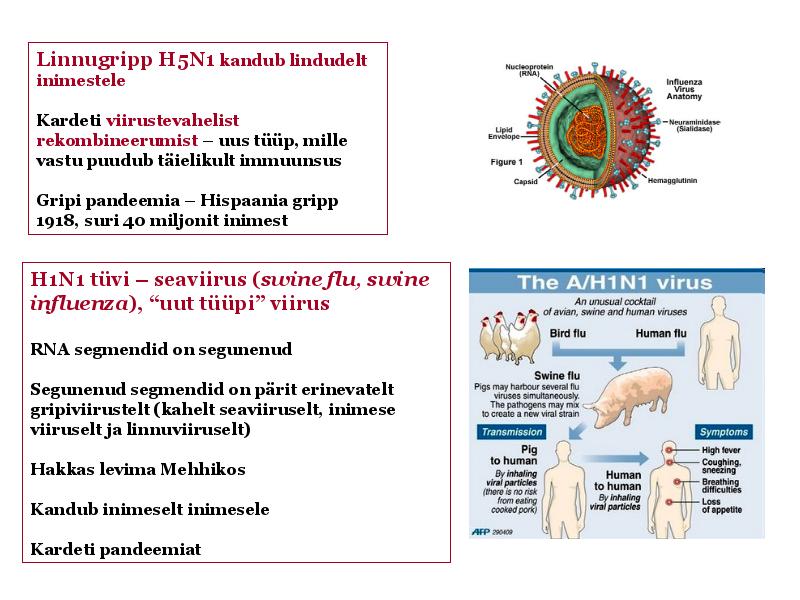
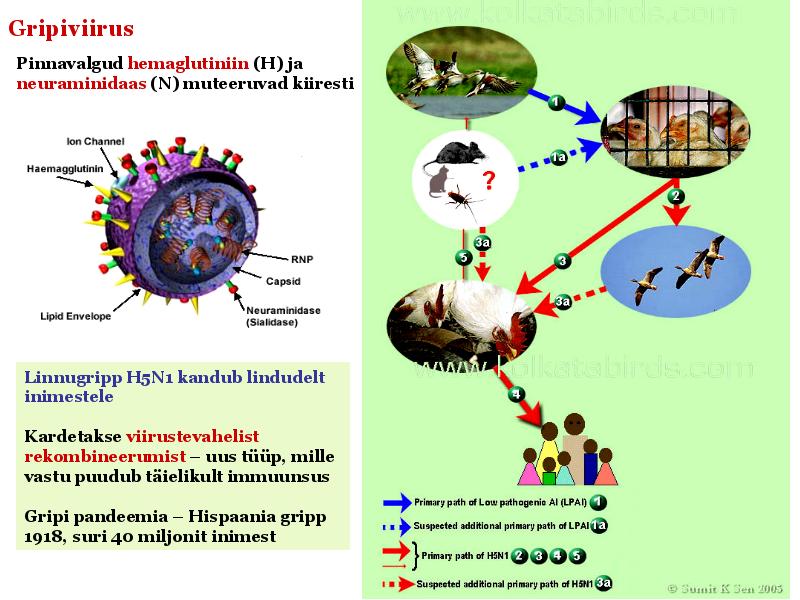
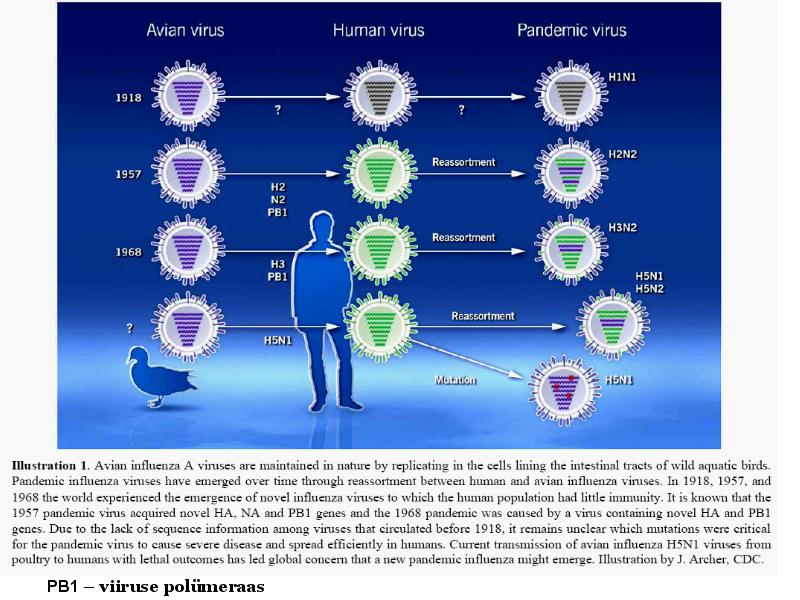
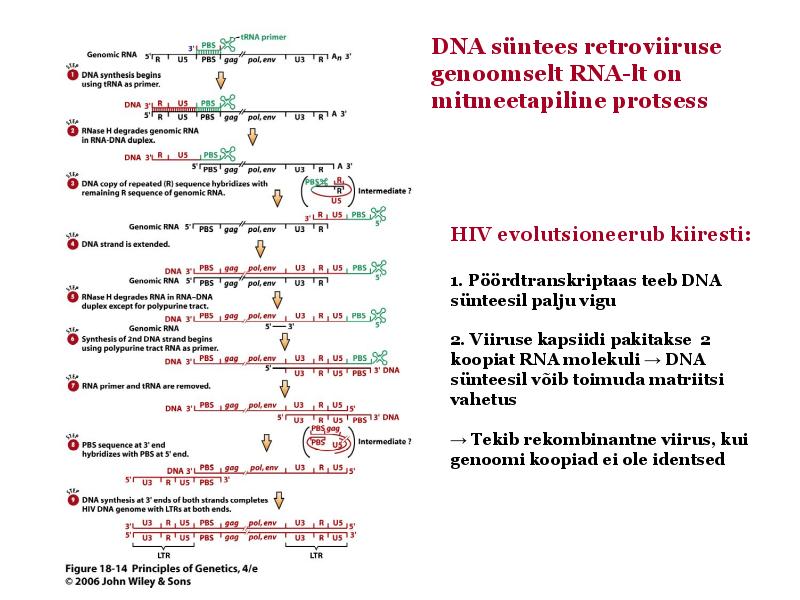
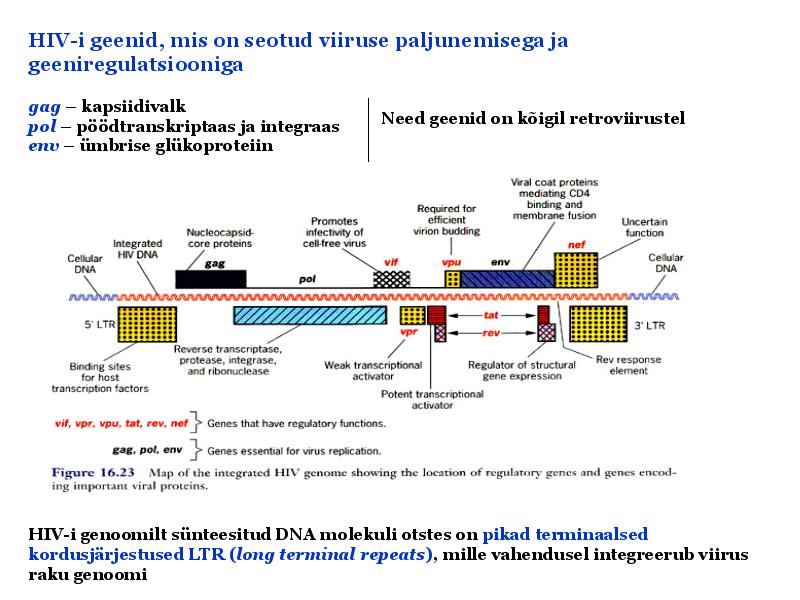
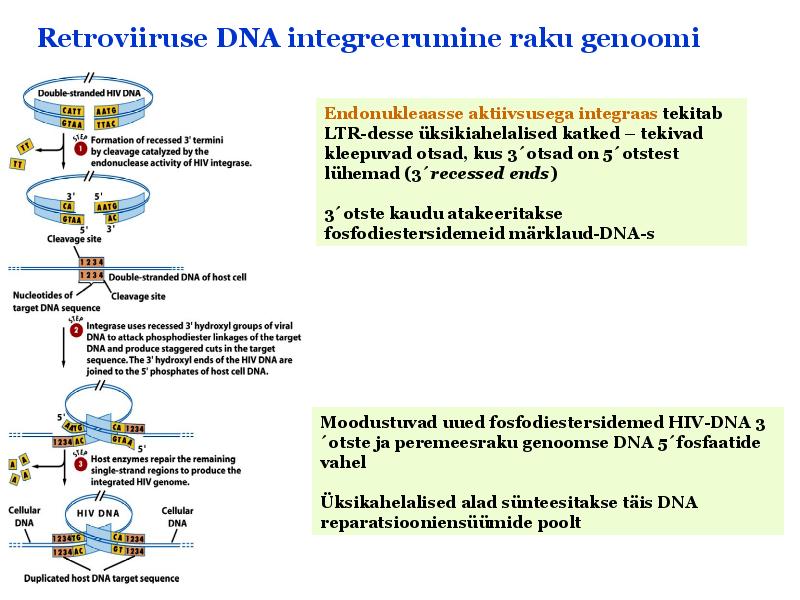
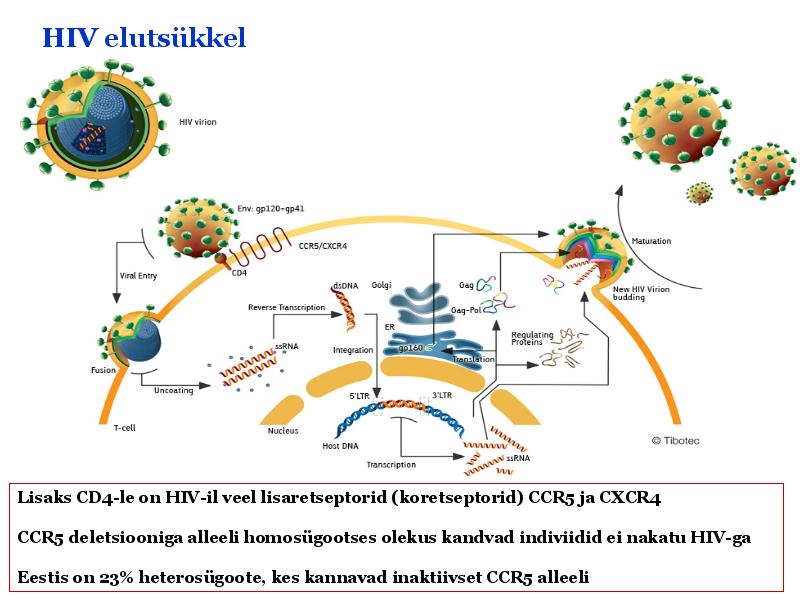
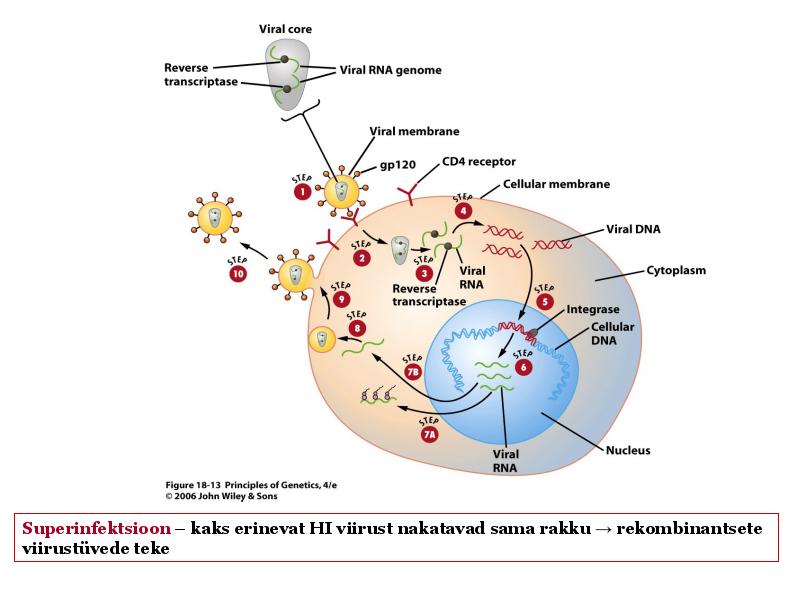
o Gripiviiruse genoom koosneb 8-st erinevast RNA segmendist, mis pakitakse valkkesta viiruse assambleerimisel o Geneetilise materjali segunemine võib toimuda siis, kui 2 sarnast viirust nakatavad sama rakku Rekombinantsed girpiviirused ja nendega seotud ohud: Linnugripp (H5N1): o Kandub lindudelt inimestele o Kardeti viirustevahelist rekombineerumist – uus tüüp, mille vastu puudub täielikult immuunsus o Hispaania gripp 1918 – pandeemia; viirus suutis välja lülitada RIG1 geeni, mis põhjustab immuunsüsteemi ülereageerimise H1N1 tüvi – seaviirus, „uut tüüpi“ viirus: o RNA segmendid on segunenud o Segunenud segmendid on pärit erinevatelt gripiviirustelt (kahelt seaviiruselt, inimese viiruselt ja linnuviiruselt) o Hakkas levima Mehhikos o Kandub inimeselt inimesele o Kardeti pandeemiat 3. Selgitage retroviiruse paljunemist HIV näitel. HIV-ga nakatunute ravi Retroviirused:
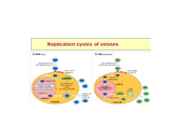
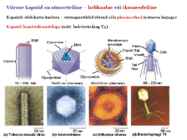
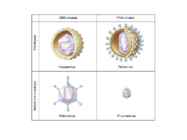
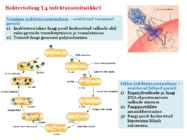
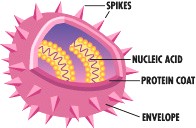
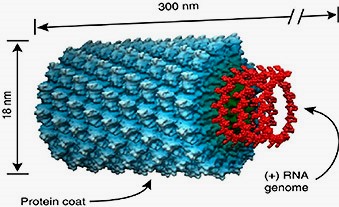
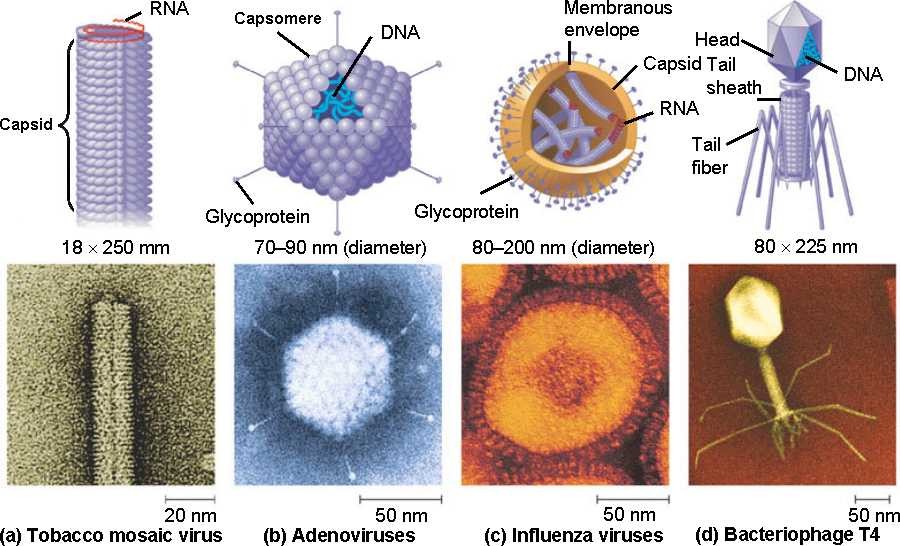
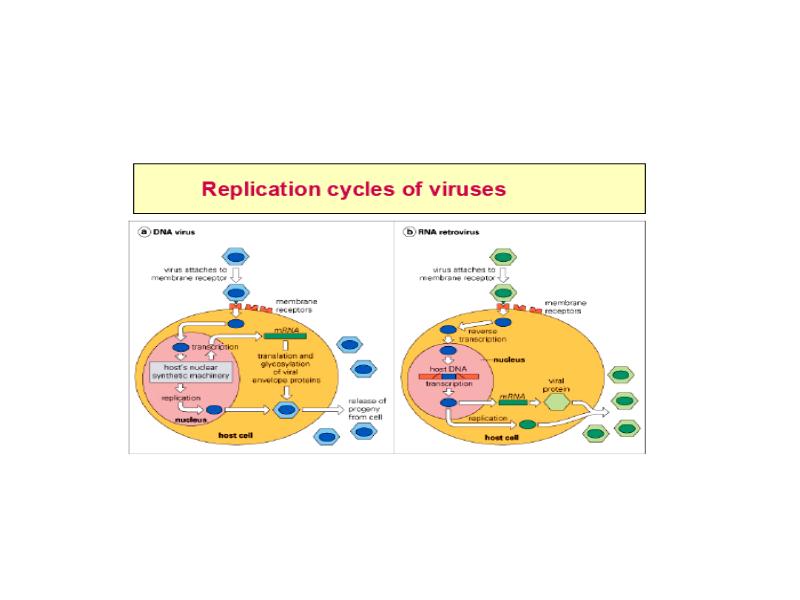
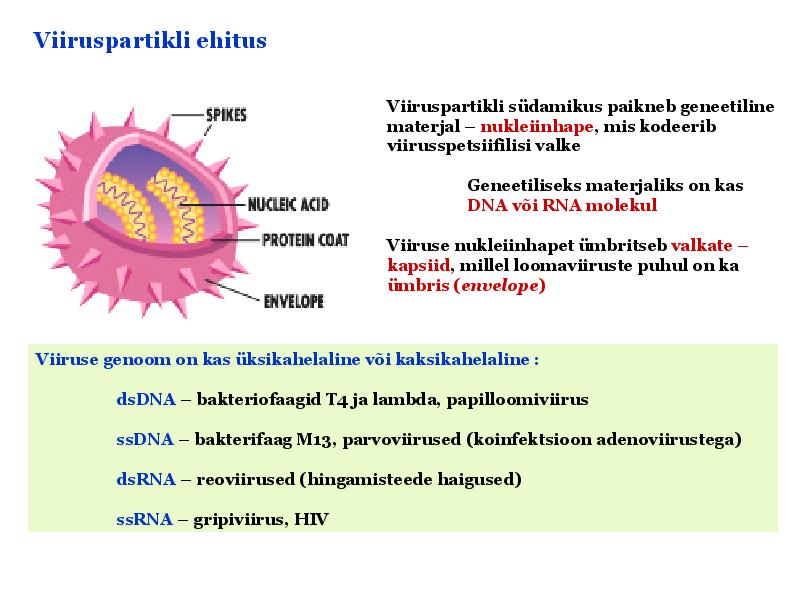
Viroloogia Viiruse definitsioon Viirused on rakulist ehitust mitteomavad obligatoorsed endoparasiidid. Erinevad rakulise ehitusega parasiitidest selle poolest, et : Viirus ei ole iseseisvalt aktiivse eluga väljaspool rakku. Viiruse bioloogiline aktiivsus avaldub ainult nakatunud rakus. Viiruse geneetiline materjal asub vahetult rakus, ei ole ümbritsetud mingi membraaniga. Viirus ei paljune pooldumisega vaid sünteesitakse ta struktuurseid osasid ja pakitakse jälle kokku. Ükski teadaolev viirus ei kodeeri tervikliku translatsioonisüsteemi. Viirus on võimeline peremeesrakust lahkuma ja nakatama teisi rakke. Virion- viiruse osake Virion- viirus-spetsiifiline struktuur , mis on ette nähtud viiruse genoomi toimetamiseks ühest rakust teise. Erinevused virioni struktuuris kajastavad enamasti erinevusi viiruste elutsüklites.
erinevate rakkude membraanile. Mikroobide tundlikkust MAC-ile nim seerumtundlikkuseks. Humoraalne immuunsus – antikehade poolt vahendatud immuunsus IMMUNOGLOBULIINID on antikehad, mille spetsiifiline antigeen on kindlaks tehtud (IgG, IgA, IgM, IgD, IgE). * Seotud B-lümfotsüütidega. (tüümusest sõltumatud lümfotsüüdid) Rakuline immuunsus – immuunvastus, mille tagavad peamiselt T8- lümfotsüüdid ja tsütokiinid; suunatud eeskätt viiruste ja kasvajate vastu Rakulises immuunvastuses osalevad lümfotsüüdid diferentseeruvad tüümuses – tüümusest sõltuvad Immunoglobuliinid - antikehad, mille spetsiifiline antigeen on kindlaks tehtud Opsonisatsioon - protsess, kus mikroob vm osake kaetakse antikehade või komplemendiga, tehes selle äratuntavaks ja fagotsüteeritavaks fagotsüütidele. kr keeles „söödavaks tegema“. Opsoneerivat toimet omavad osad IgG, IgM
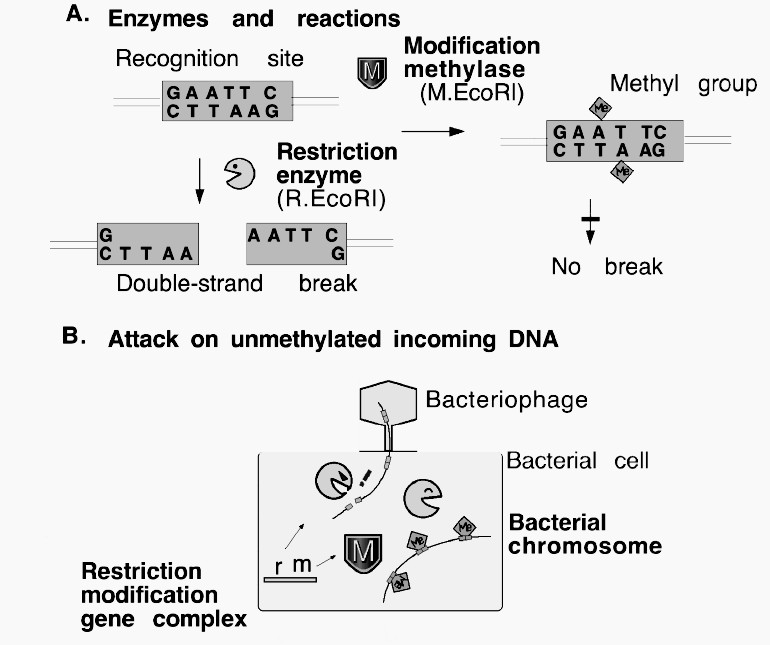
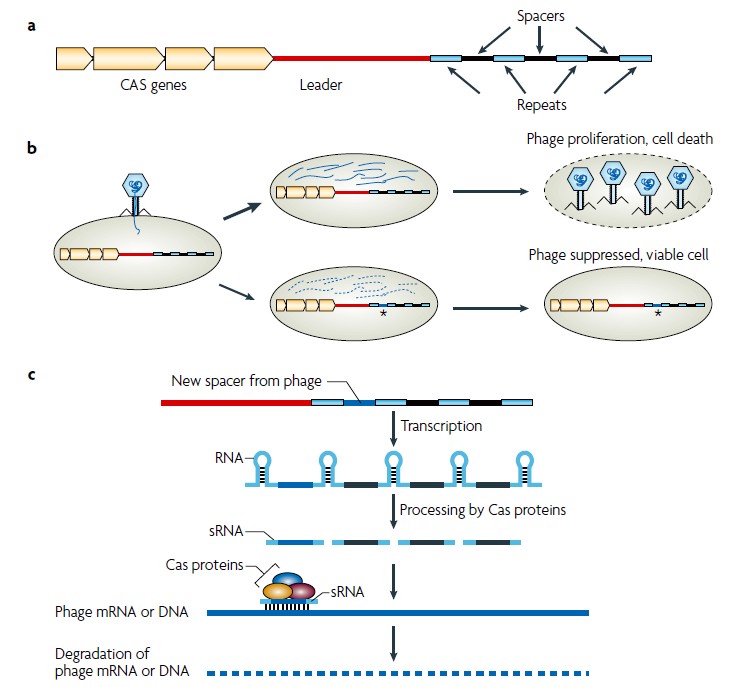
kiirguse. Kiirguse baktereid kahjustava toime mehhanismid § Adsorbeerunud valgus (peamiselt tsütokroomides) põhjustab liigse energia vabanemise tõttu molekulaarse hapniku tekke - tegemist on väga tugeva oksüdeerijaga, mis põhjustab kahjustusi. § UV kiirgus põhjustab DNA-s spetsiidilisi kahjustusi, tekivad tümiini dimeerid. § Ioniseeriv kiirgus põhjustab: H-sidemete katkemise, paljude ühendite oksüdeerumise, DNA katkemise. Balterite geneetika Bakterite geneetika § Haploidsed, enamasti tsirkulaarse genoomiga (E. coli 4.5 X 106 bp.) § Kiire kasv (E. coli 20 minutit generatsiooniaeg; seepärast, 1 rakust 1000000 7 tunni vältel.) § Vedelsöötmes ~109- 1010 küllastustasemel, bakterite sademes ~ 1012 rakku/g. § Rakutuuma puudumine; § Monokromosomaalsus ja haploidsus; § DNA on kogu oma pikkuses funktsionaalne; § Prokarüootsed rakud poolduvad mitoosita Nukleiinhapped DNA info kandja, säilitaja ja vahendaja; (bakterites,
Putukate, vetikate ja ainuraksete suured DNA genoomsed viirused Baculoviirused moodustavad suure viiruste sugukonna (teada üle 500 liigi). Neile kõigile on iseloomulikud suured tsirkulaarsed dsDNA genoomid, mis on virionis pakitud kepikesekujuline nukleokapsiid (lad. baculum – kepike, siit ka sugukonna nimetus). Baculoviirustele on iseloomulik nakatunud rakkudes spetsiifiliste inklusioonkehade moodustamine. Baculoviiruseid on kõige enam teada liblikalistel, kuid neid on leitud ka kahetiivalistel, ehmestiivalistel ja ka krevettidel. Baculoviirustel esineb kahte tüüpi virione:
Varased geenid stimuleerivad rakukasvu, mis võimaldab viiruse genoomi replikatsiooni peremehe DNA polümeraasi poolt, kui rakud jagunevad. Viirus-indutseeritud rakkude arvukuse tõus põhjustab naha basaal- ja ogakihi (stratum spinosum) paksenemist. Basaalrakkude diferentseerudes põhjustavad erinevates nahakihtides ja –tüüpides ekspresseeritavad tuumafaktorid erinevate viirusegeenide transkriptsiooni. Hiliseid geene ekspresseeritakse ainult lõplikult diferentseerunud pealmises nahakihis, viirus pakitakse kokku tuumas. Kasutades naharakkude küpsemist, saab viirus naha pinnale ja eritatakse/irdub koos nahapinna surnud rakkudega. Patogenees. Infitseerivad ja paljunevad lameepiteelis, indutseerides epiteeli proliferatsiooni: nahas tüükad, limaskestadel genitaalsed, oraalsed, konjuktiivipapilloomid. Tüügas areneb viiruse poolt indutseeritud raku kasvu ja basaal-, granuloos- ja ogakihi paksenemise tõttu












































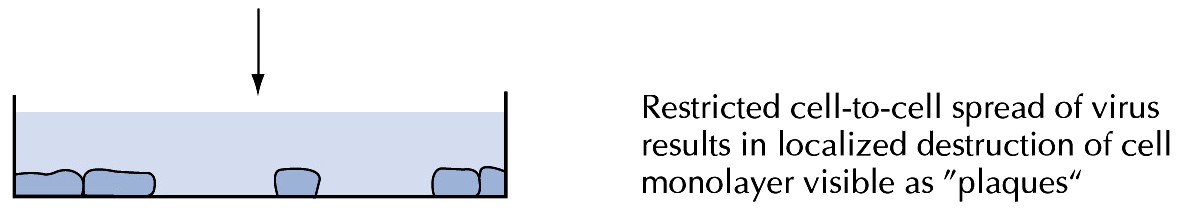
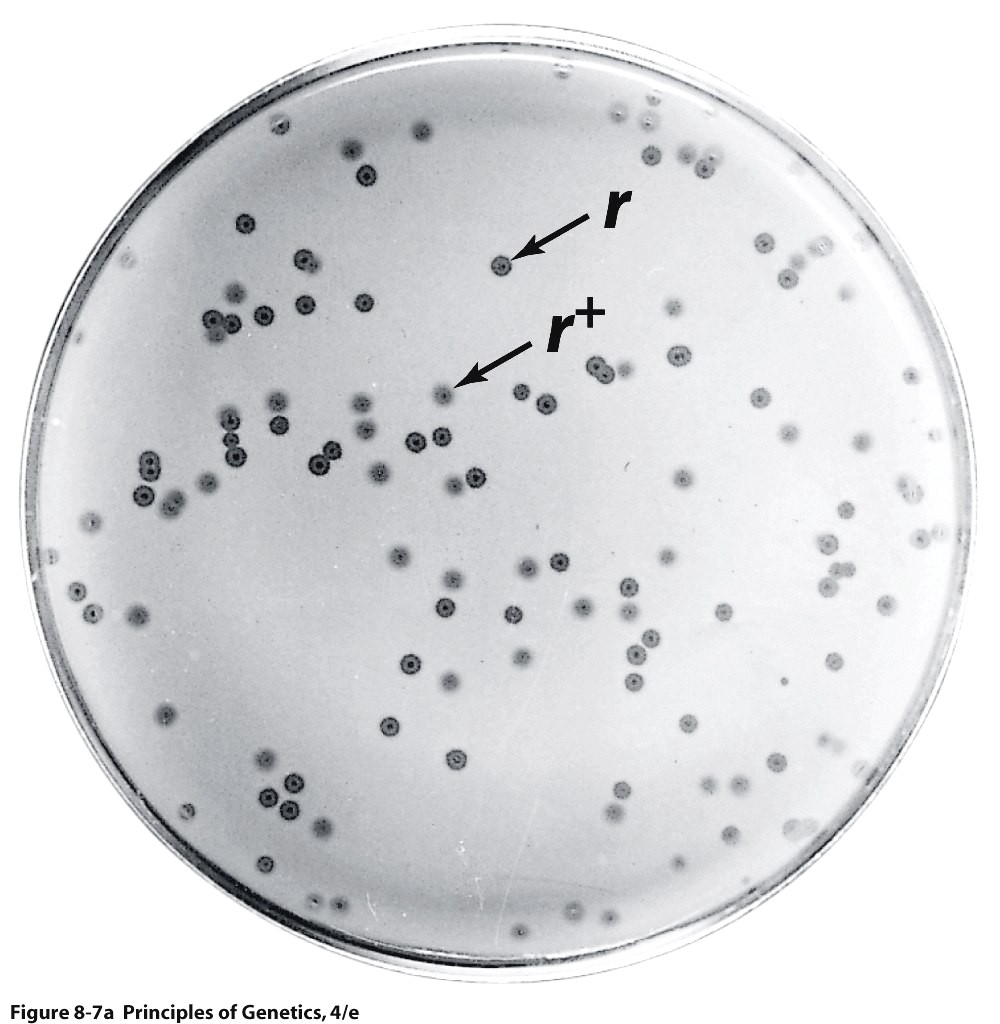
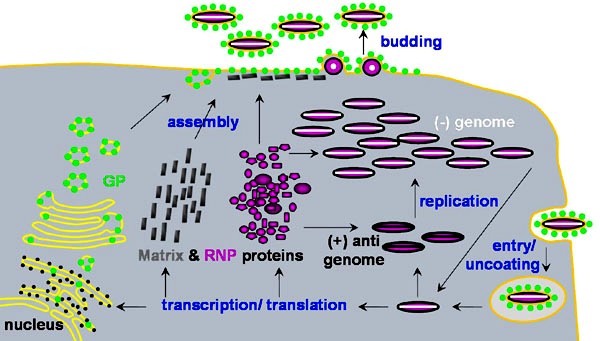

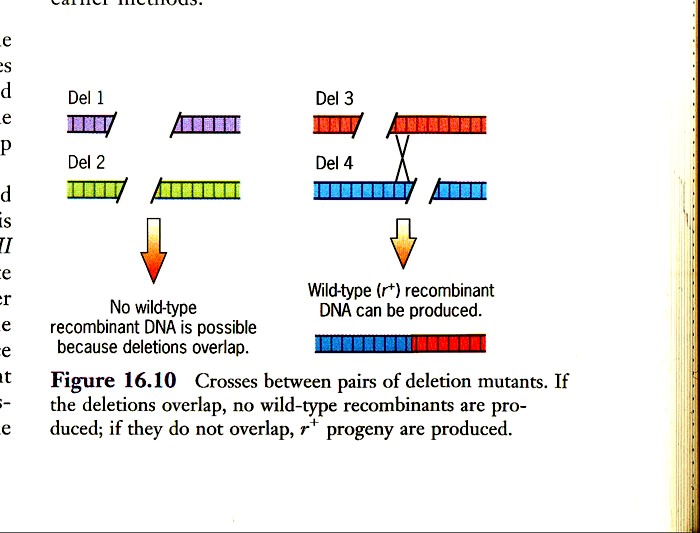

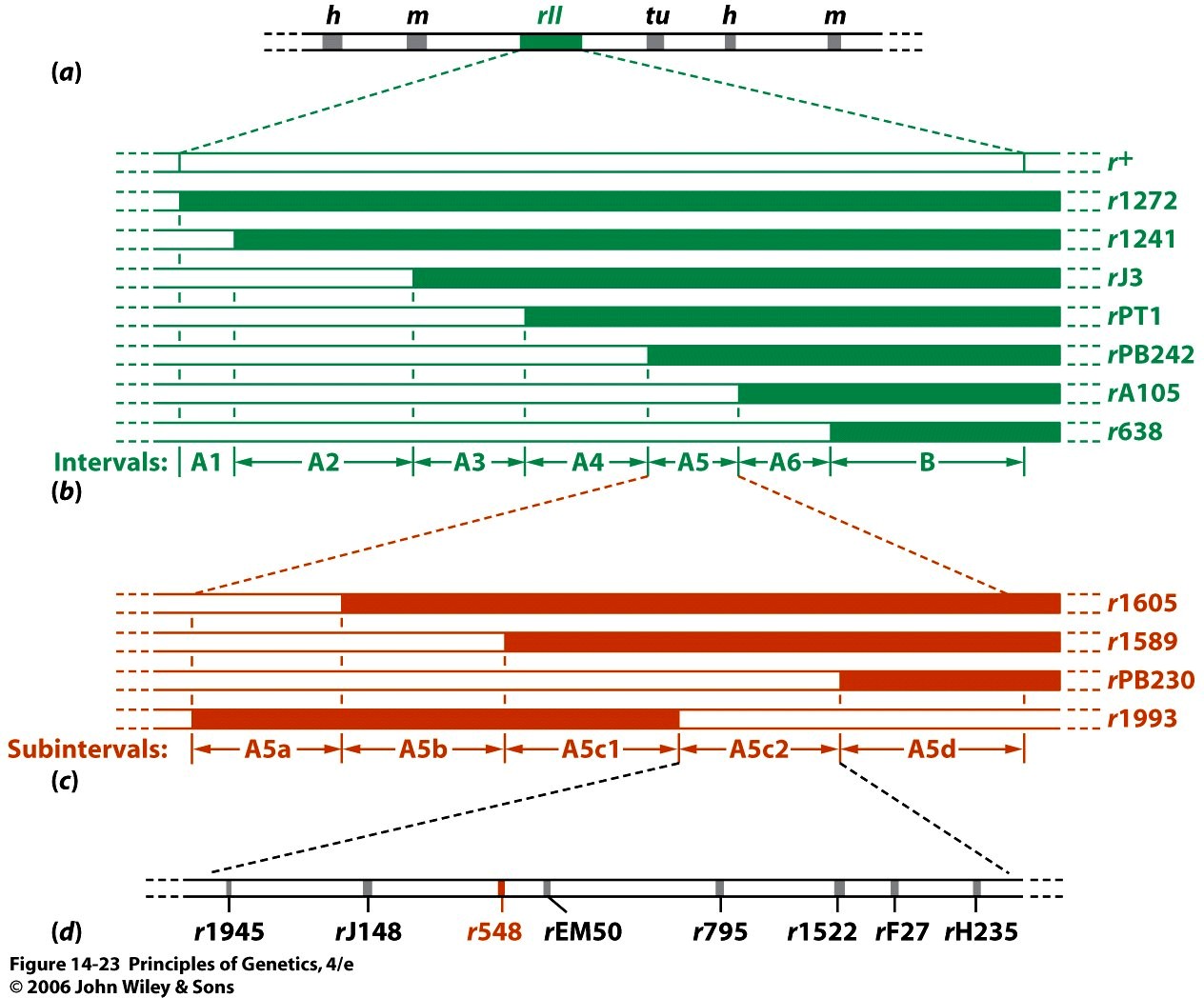
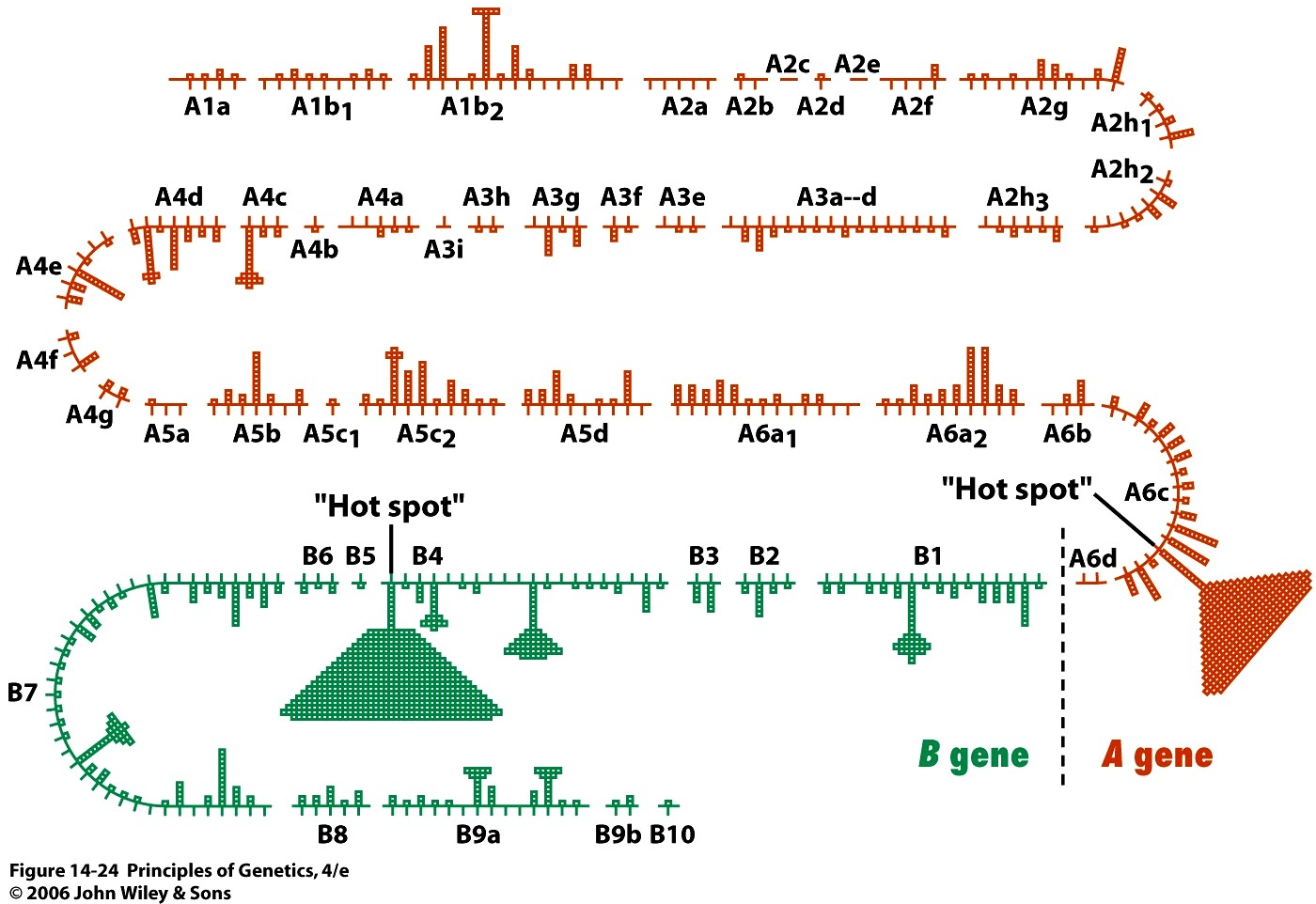
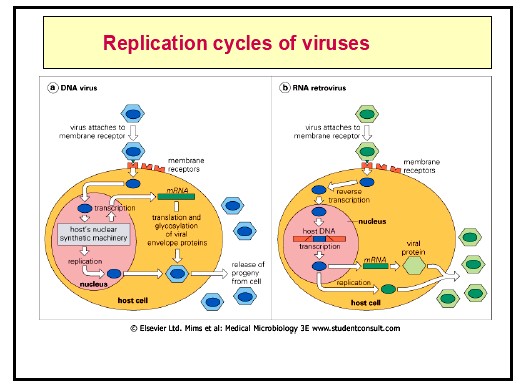


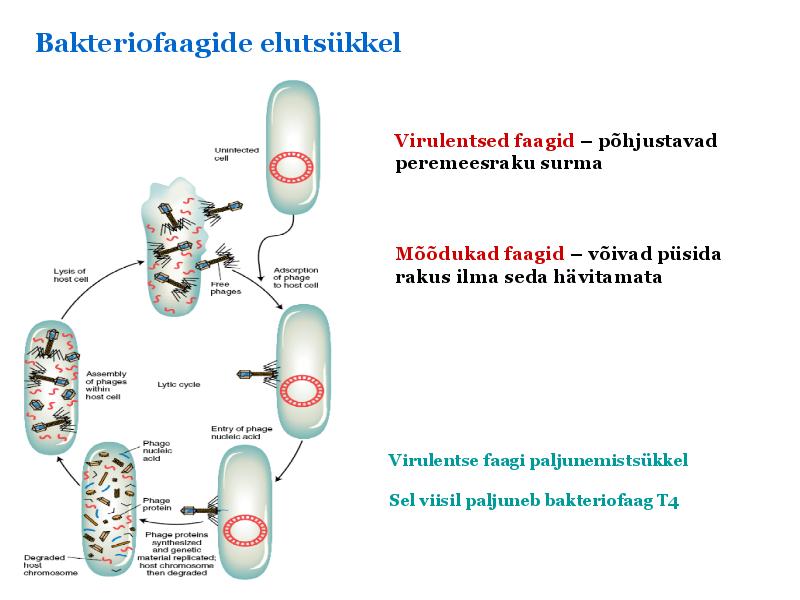
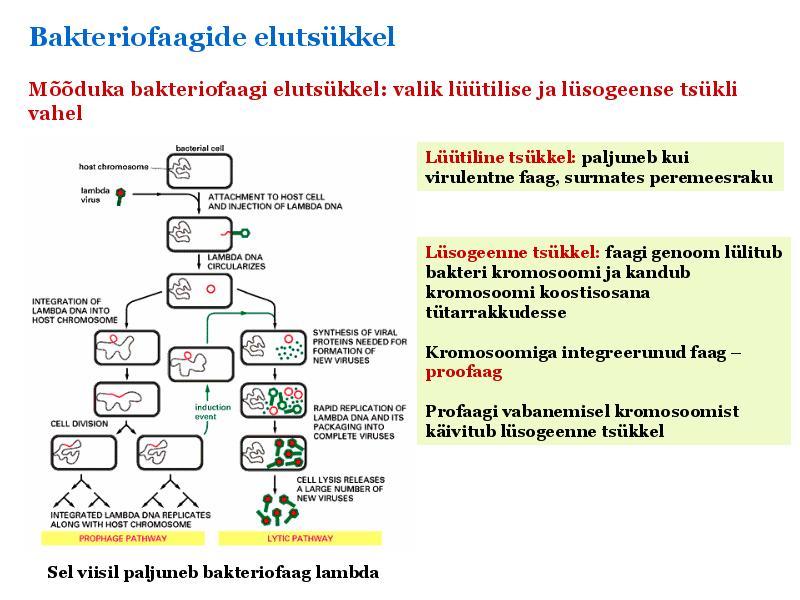
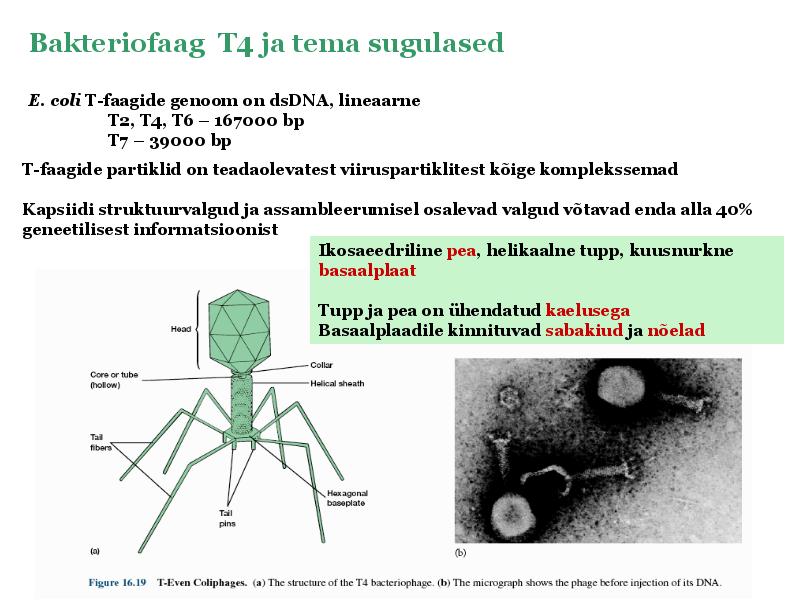
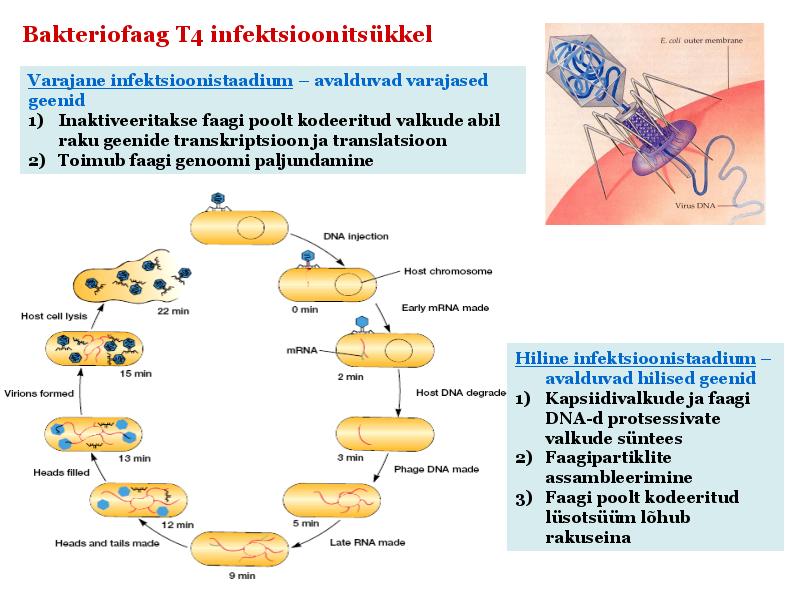
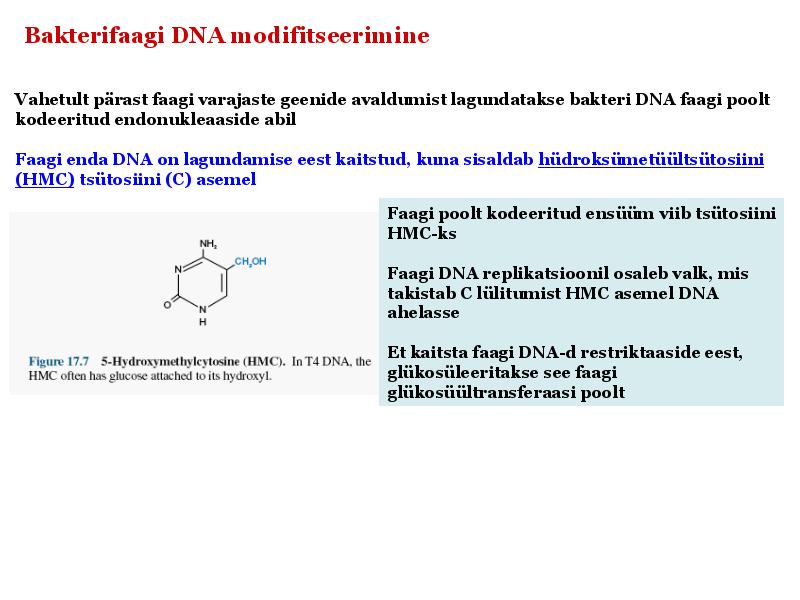
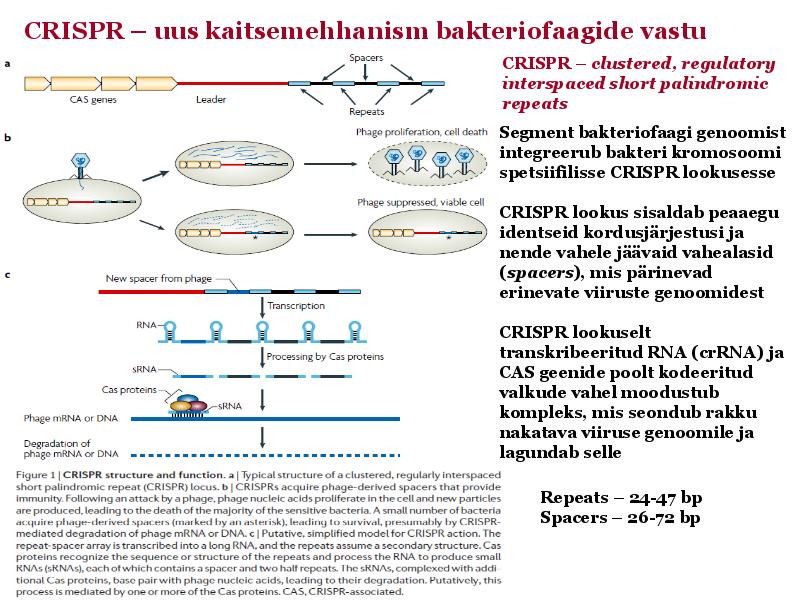
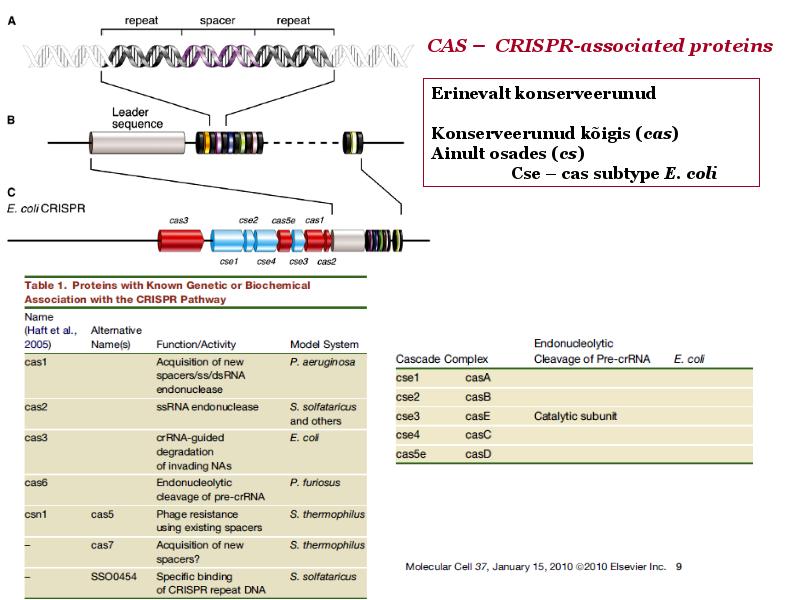
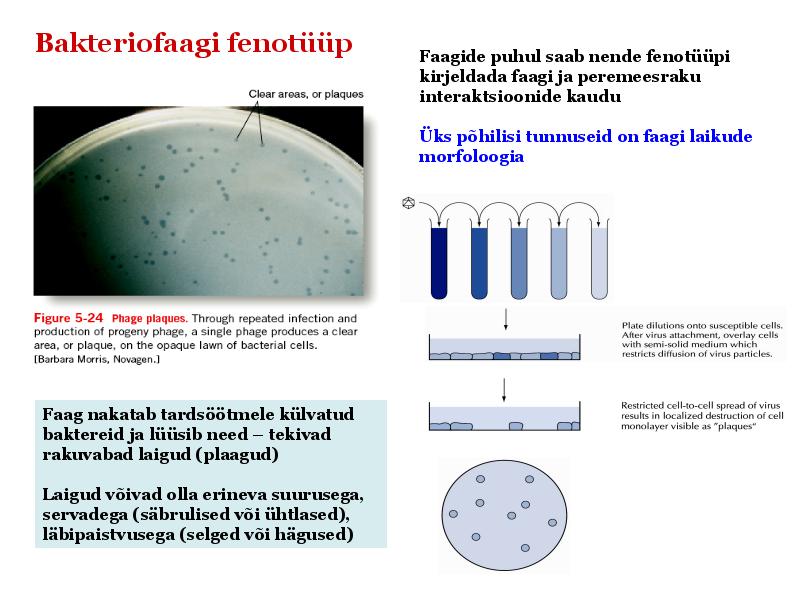
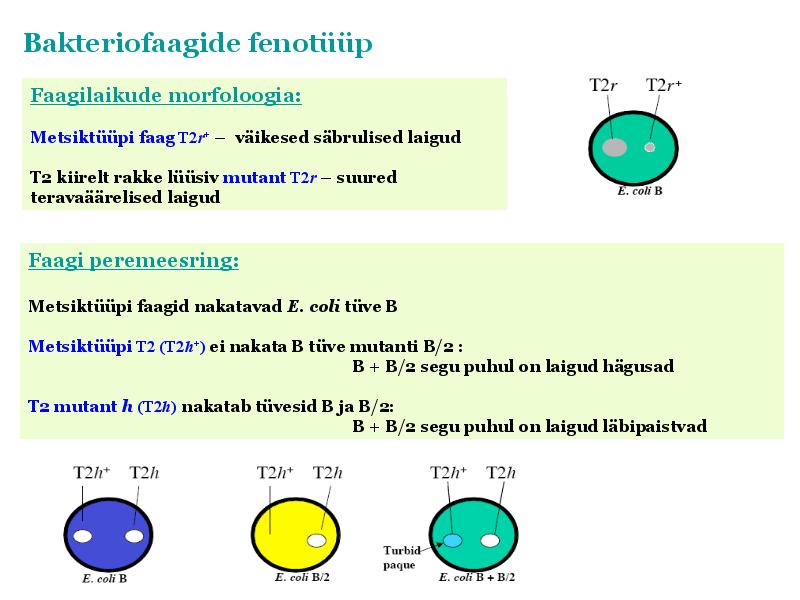
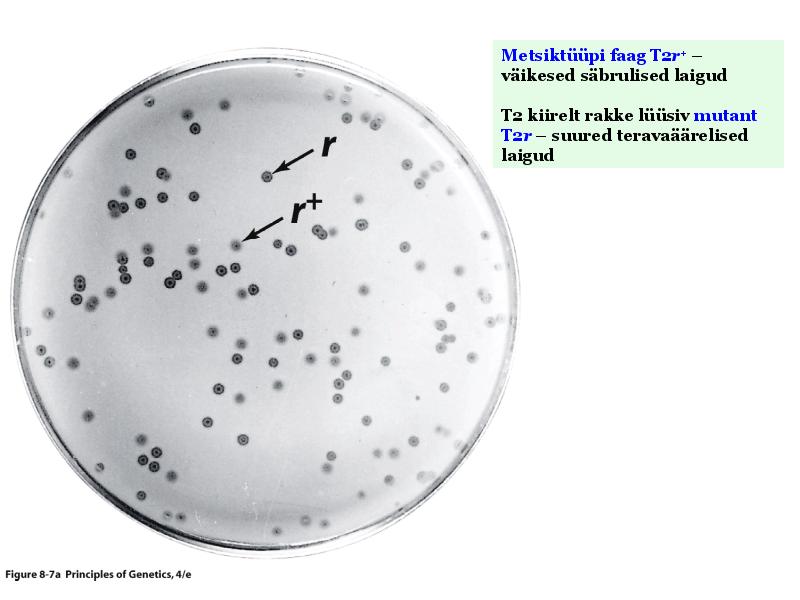
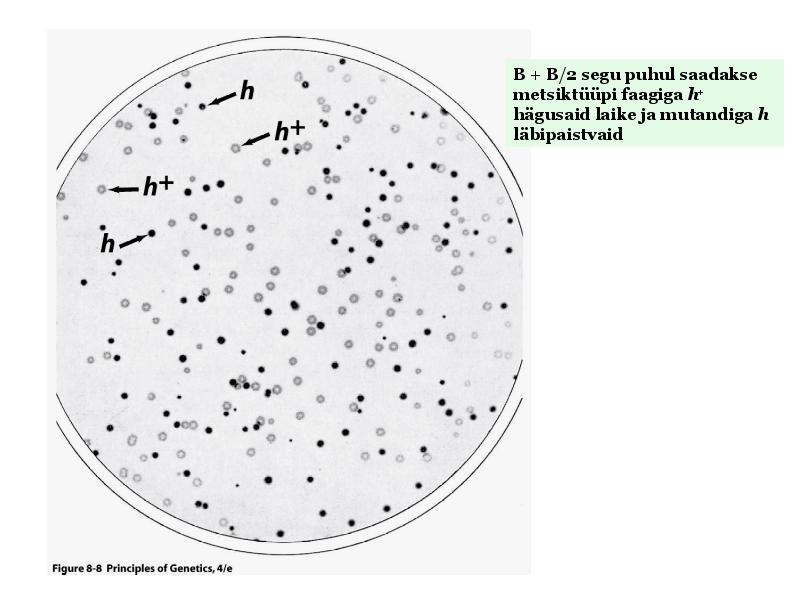
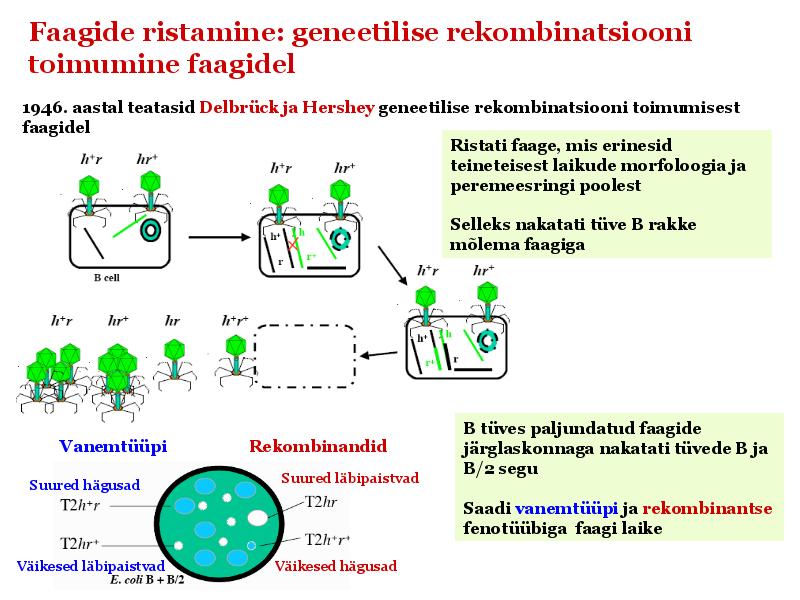
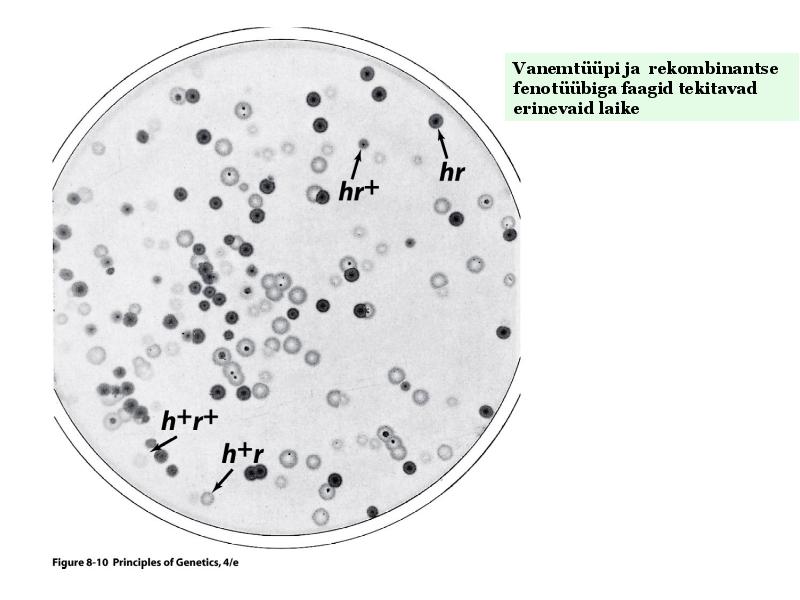
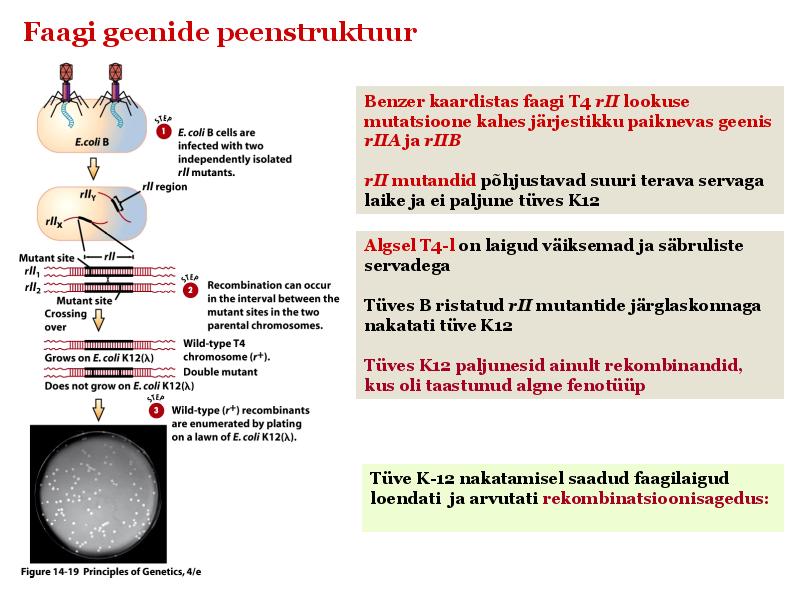
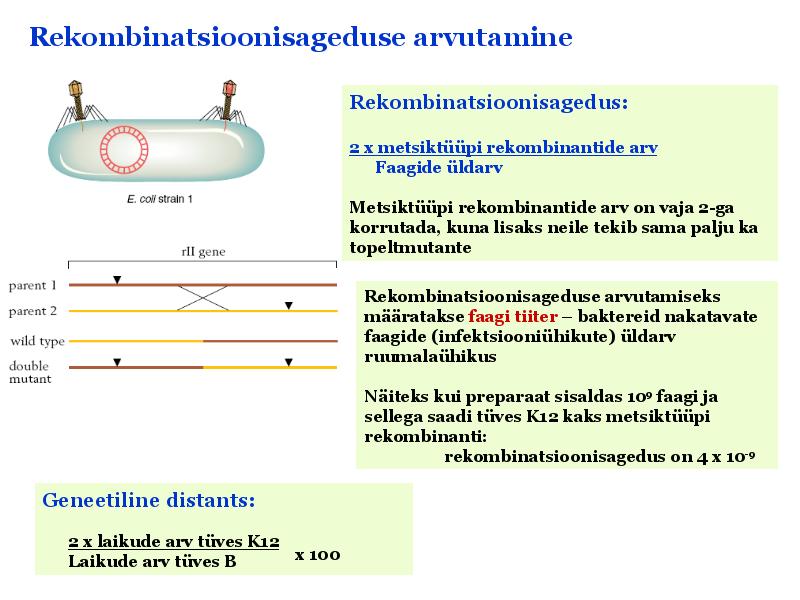


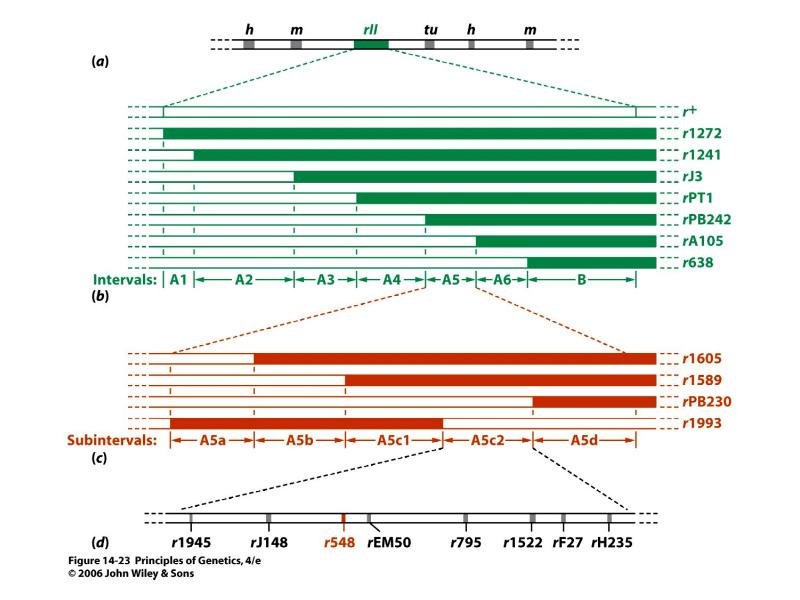
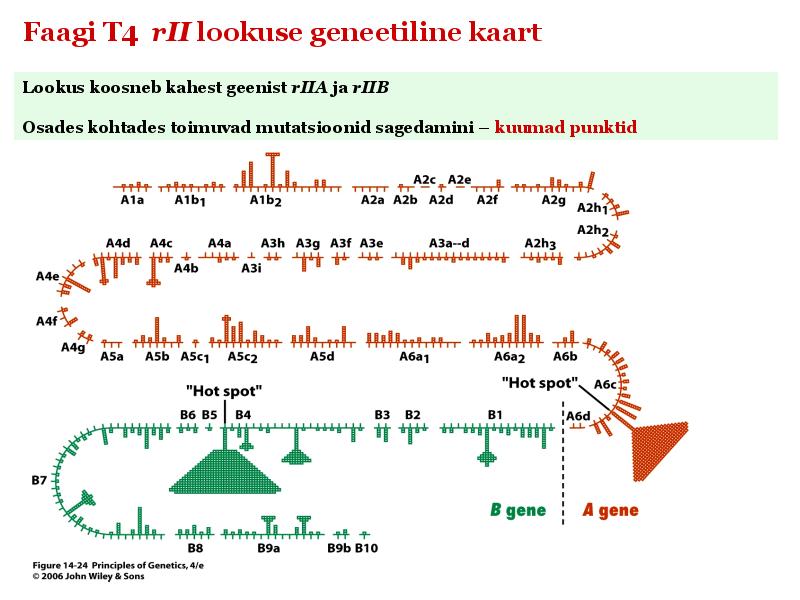
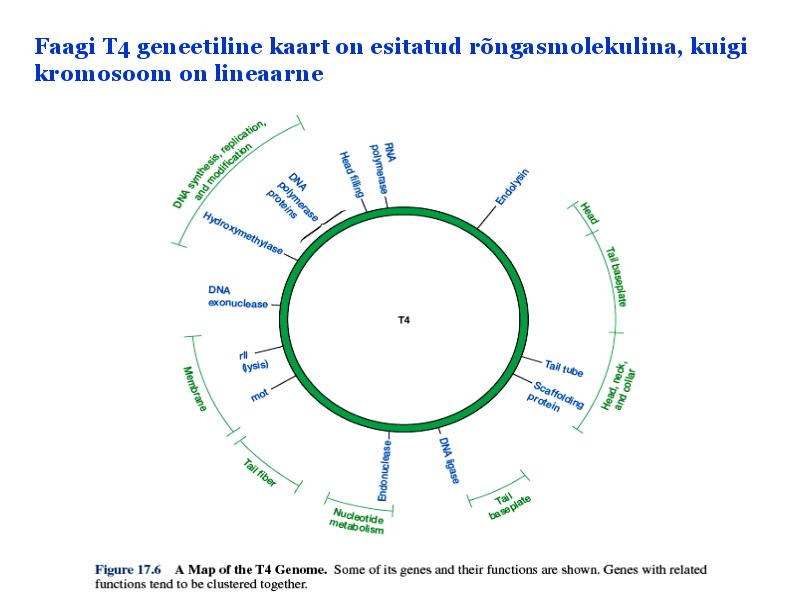
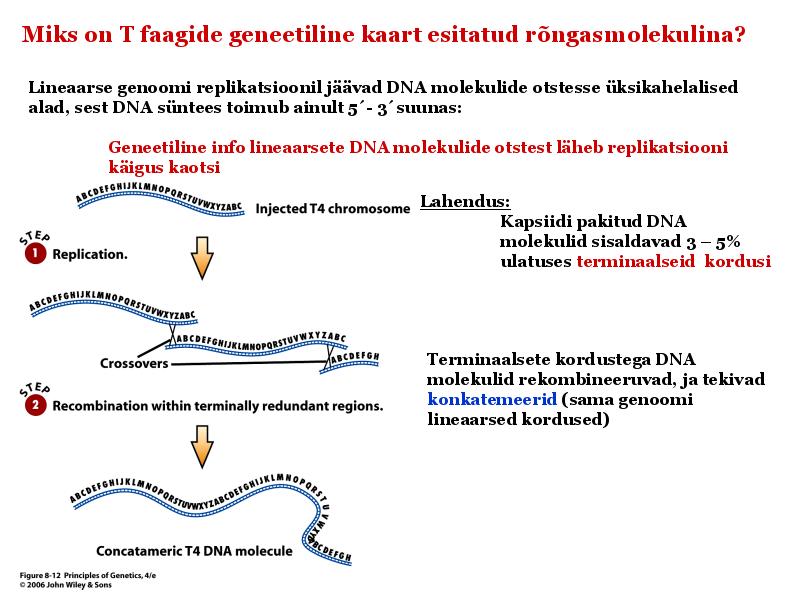


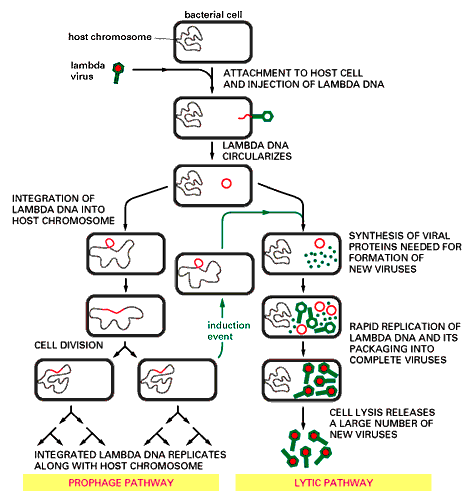
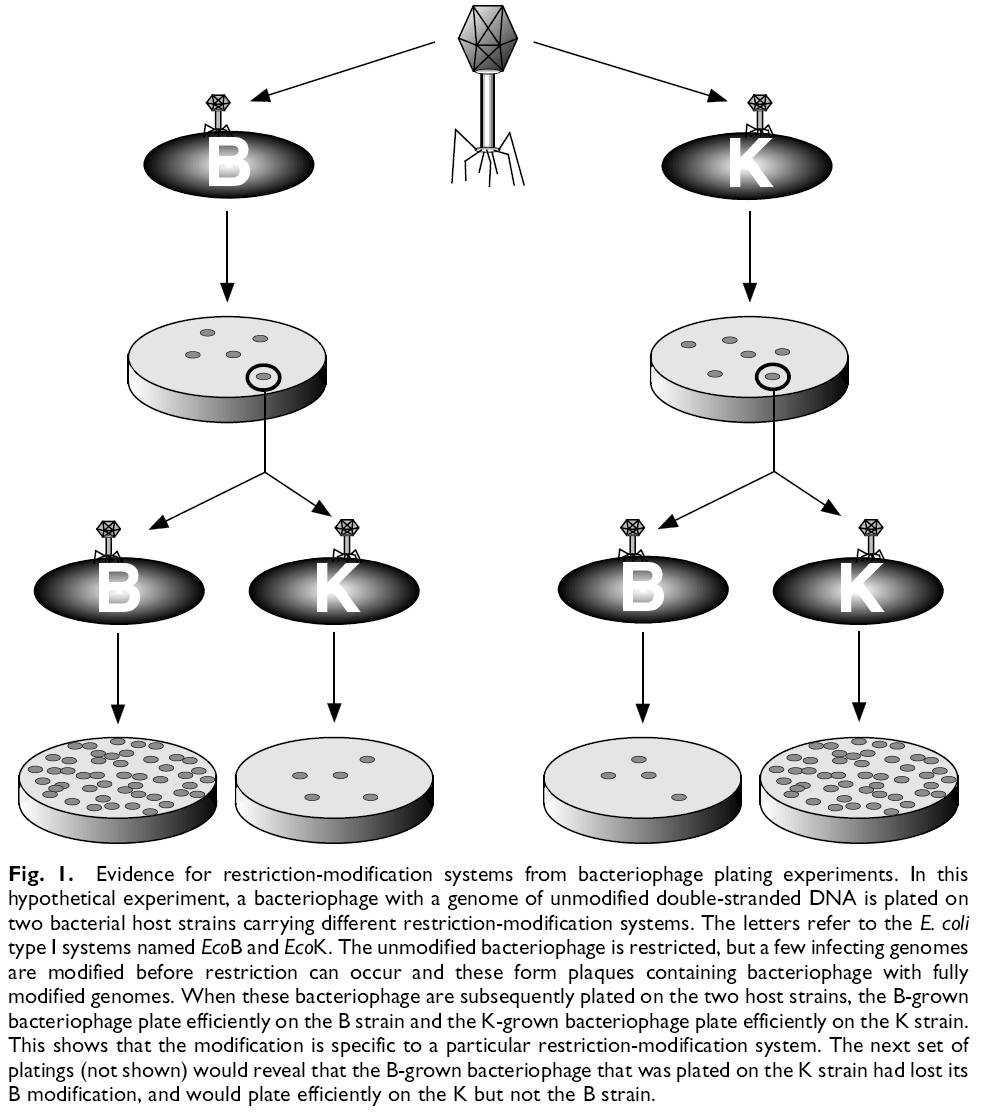
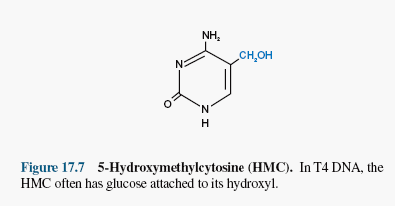

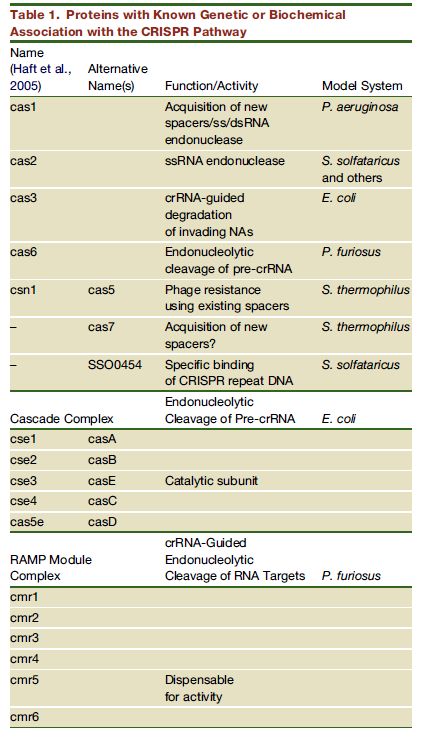
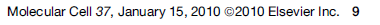

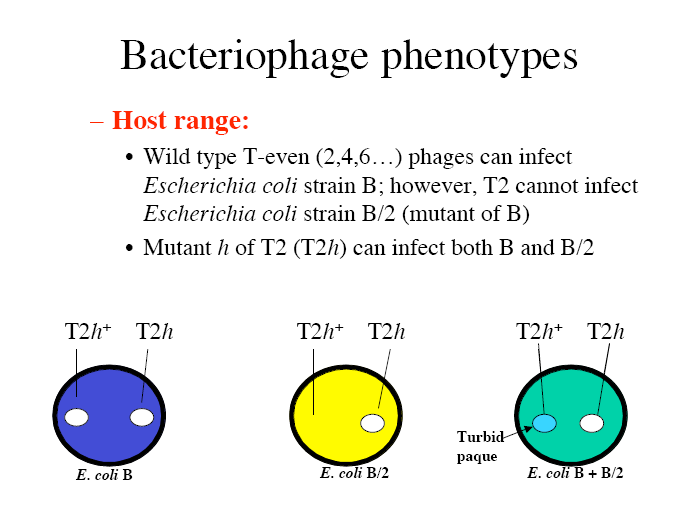
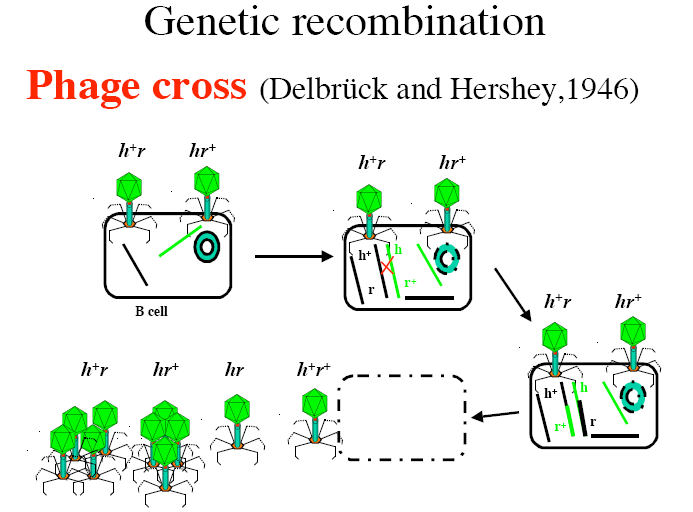
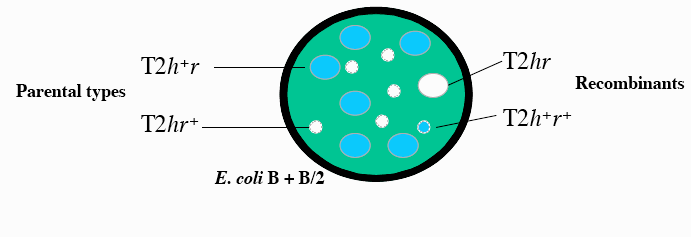
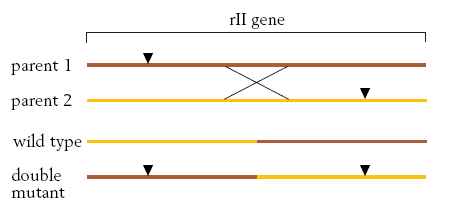
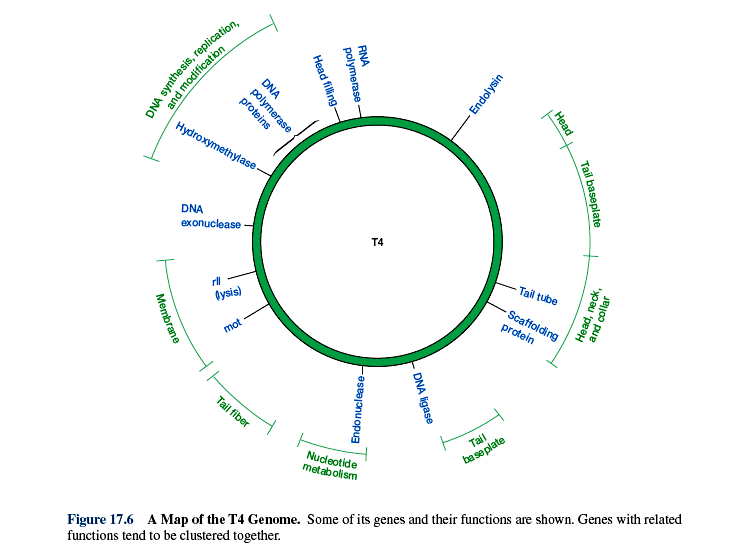




Kõik kommentaarid