komponente. YAC on valmistatud pärmi kromosoomi spetsiifiliste piirkondade põhjal ning BAC toodetud F-plasmiidi põhjal. YAC on lineaarne fragment, kuid BAC sirkulaarne. YAC and BAC vectors are two types of artificial vector systems designed to clone large genomic DNA fragments. They have multiple applications in the preparation of genomic and cDNA libraries. 15. Kirjelda kloneerimise protseduuri. Mis on kloneerimiseks hädavajalikud komponendid ning tingimused? Geeni kloonimine (ingl. gene cloning) eeldab mitut järjestikust protseduuri, milledeks on uuritava geeni: 1) isoleerimine; 2) viimine isereplitseeruvasse geneetilisse elementi (plasmiidi või viirusgenoomi); 3) amplifikatsioon ehk paljundamine; 4) uuritava geeniga rekombinantsete kloonide selektsioon. 16. Mis on geeniraamatukogud? 1:43. Geeniraamatukogu on bakteri või faagikloonide kogumid millest iga kloon sisaldab meid huvitavast geneetilisest materjalist ühte konkreetset alamosa. 17
Alusta PCR'i. (5) Samal ajal valmista ...% geel. 40 ml TBE puhvri kohta on vaja kaaluda ... g agaroosi. Valmistamist vaata tööst nr 3. (6) PCRi lõppedes (<1 tund), analüüsi 1/5 saadud produktist agaroosgeelil. Selleks pipeteeri 10 l 1 x TBE foreesipuhvrit (lilla) + 2 l PCR tuubist kokku 12 l, kanda kõik geelile. Geelile lisada kindlasti molekulmassi marker ja kontroll PCR (amplifikatsioon tühjalt pBS SK+ plasmiidilt). Interpreteeri tulemus. Saadud rekombinantsed plasmiidid on valmis inserdi sekveneerimiseks T7 praimeri ja "kit"i abil (järgmises praktikumis). Restriktsiooni tulemused: Minu minipreparatsioonis tuli välja 2 inserti. Üks umbes 2,5 kb pikkune ja teine umbes 6 kb pikkune. Võib olla see on tingitud sellega, et ensüüm Cfr421 ei töötanud väga hästi. 10 Lisaülesanded: 4.1) Leia Wallace reegli abil (4x GC + 2x AT) T3 ja T7 praimerite sulamistemperatuurid. Kui
Valmistamist vaata tööst nr 3. (6) PCRi lõppedes (<1 tund), analüüsi 1/5 saadud produktist agaroosgeelil. Selleks pipeteeri 10 l 1 x TBE foreesipuhvrit (lilla) + 2 l PCR tuubist kokku 12 l, kanda kõik geelile. Geelile lisada kindlasti molekulmassi marker ja kontroll PCR (amplifikatsioon tühjalt pBS SK+ plasmiidilt). 12 Interpreteeri tulemus. Saadud rekombinantsed plasmiidid on valmis inserdi sekveneerimiseks T7 praimeri ja "kit"i abil (järgmises praktikumis). PCRi tulemused. Marker 100 bp pBS SK+ Kristine Arvidas Hannes Mihkel Heleen Ksenia Marko Okana Jelena Doris Mikk Anna Alex Aare Irina Inna Jana Karl Suure pildi peal insertsioon puudub, restriktsioonanalüüsiga ilmus siiski välja 800-900 nukleotiidi pikk järjestus. Lisaülesanded: 4
Tallinna Tehnikaülikool Matemaatika-loodusteaduskond Geenitehnoloogia Instituut Molekulaar- ja rakubioloogia praktikum YTM0012 MOLEKULAARBIOLOOGIA PRAKTIKUM Pipeteerimine Automaatpipetiga pipeteerimise põhitõed Pipeteerimisel on kõige olulisem võtta õige suurusega pipett Pipeteeritav vedelik peab olema homogeenne. Seintelt ja korgi küljest tuleks tilgad ja kondensaat põhja fuugida Enne pipeteeritavasse lahusesse viimist vajuta kolb esimese astmeni alla. Pipeti tühjenemiseks on vaja vajutada kolb teise astmeni. Viskoosse lahuse pipeteerimine Vajalikud materjalid: pesuvahendi kontsentraat, vesi, pipetid koos otsikutega, falcon tuub, aukudega plaadike Töö käik: 1. Kasutades 1 ml pipetti tegin 15-ml falconisse 5,5ml 30% homogeenset pesuvahendilahust (1,65 ml peduvahendit ja 3,85
On DNA kloneerimise teel saadud valk. Selliseid valke saab toota nt bakterite abil või transgeensete loomade abil. Mis on cDNA? cDNA (komplimentaarne DNA) on pöördtranskriptsiooniga saadud DNA koopia. Pöörd-transkriptaas on ensüüm, mille abil sünteesitakse mRNA’lt cDNA’d. cDNA’d kasutatakse eukarüootsete geenide kloonimiseks prokarüootsetes organismides. Bakteri rakkudesse geenide viimisel peame me mRNA alusel tegema DNA. Millised on kloneeritud DNAde kasutusalad tänapäeval? Rekombinantsed valgud on meile väga praktilised, kuna saame neist toota erinevaid ravimeid nt. insuliini. Kasutatakse veel toidutööstuses ensüümid (piim, juust), keskkonnakaitses (toksiliste jääkide likvideerimine bakterite abil). Kloonitud geenid on kasulikud tegemaks uuritavast geenist kergesti hallatavaid koopiaid ja tootmaks selle põhjal vajaminevat valku. Sangeri sekveneerimise põhimõte. Kasutatakse didesoksüribonukleotiide, mis erinevad desoksüribonukleotiididest selle
Kontrolltöö 1. 1. Milline järgnevatest vektorsüsteemidest kasutab peremeesrakku sisestatud DNA konstrukti tsirkulariseerimiseks cre rekombinaasi ? a) Plasmiid b) Mitte ükski c) Kosmiid d) YAC e) P1 2. Üheks väga levinud viisiks reaalaja kvantitatiivse PCR-i (qPCR-i) tegemisel on TaqMan proovide kasutamine. Millisel DNA polümeraasi omadusel, lisaks 5'-3' sünteesi aktiivsusele, põhineb antud tehnoloogia? a) DNA polümeraasi 3'-5' eksonukleaassel aktiivsusel b) DNA polümeraasi 5'-3' eksonukleaassel aktiivsusel c) DNA polümeraasi termostabiilsusel 3. Milliseid mehhanisme kasutatakse rakus geeniekspressiooni inhibeerimisel antisense DNA oligonukleotiididega? a) Moodustuva antisense oligonukleotiid/mRNA kompleksi katkilõikamine restriktsioonilise endonukleaasiga b) Translatsiooni blokeerimine oligonukleotiidi poolt c) DNA oligonukleotiid/mRNA kompleksi degradatsioon Dicer ensüümi poolt d) RNaasH vahendatud
rakku, genoom integreerub peremehe genoomi ja kandub kromosoomi koostisosana tütarrakkudesse. Keskkonnatingimuste muutudes võib lüsogeenne faag minna üle lüütilisse tsüklisse, mille käigus sünteesitakse viiruse partikleid ning pannakse need kokku. Lõpuks rakk lüüsitakse ning uued virionid vabanevad keskkonda, kus nad on võimelised minema nakatama järgmisi peremeesrakke 2. Gripiviiruse muteerumine, rekombinantsed gripiviirused ja nendega seotud ohud Gripiviirus: o Pinnavalgud hemaglutiniin (H) ja neuraminidaas (N) muteeruvad kiiresti – RNA polümeraas teeb palju vigu, mida ei parandata! o Gripiviiruse genoom koosneb 8-st erinevast RNA segmendist, mis pakitakse valkkesta viiruse assambleerimisel o Geneetilise materjali segunemine võib toimuda siis, kui 2 sarnast viirust nakatavad sama rakku Rekombinantsed girpiviirused ja nendega seotud ohud:
Molekulaar- ja rakubioloogia praktikum ARUANNE Ave Tüür 155356 YAGB41 Kevadsemester 2017 Tallinna Tehnikaülikool Pipeteerimine 8. veebruar I HARJUTUS Eesmärk: Õppida õigesti pipeteerima. Materjalid: Arvutused: Pesuvahendi kontsentraat Pesuvahendit on vaja: 5,5 ×30 Vesi 100 = 1,65ml Pipetid ja otsikud 5,5−2 ×1,55=2,4 ( ml ) 15ml falkon tuub 2,4 ÷ 0,2=12 1,5 ml tuubid PCR plaadi tükk Töö käik: Teen 15-ml falkon tuubi 5,5ml 30% homogeenset pesuvahendilahust Võtan kaks 1,5ml- tuubi ning pipeteerin mõlemasse 1,55ml lah


























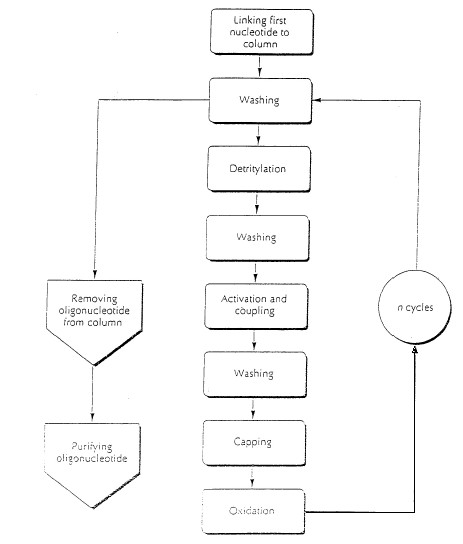
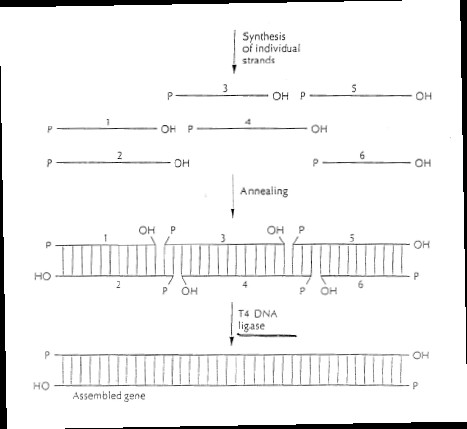
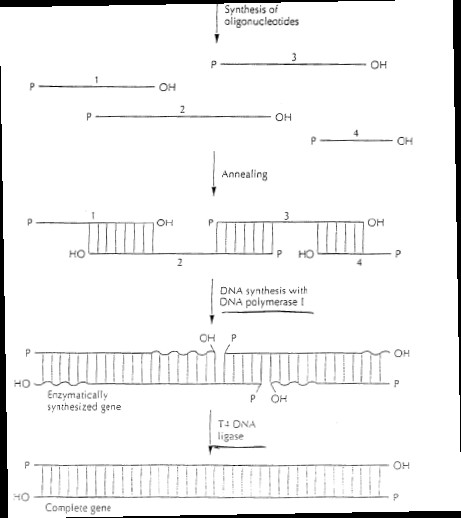
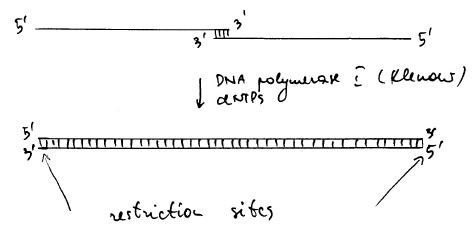
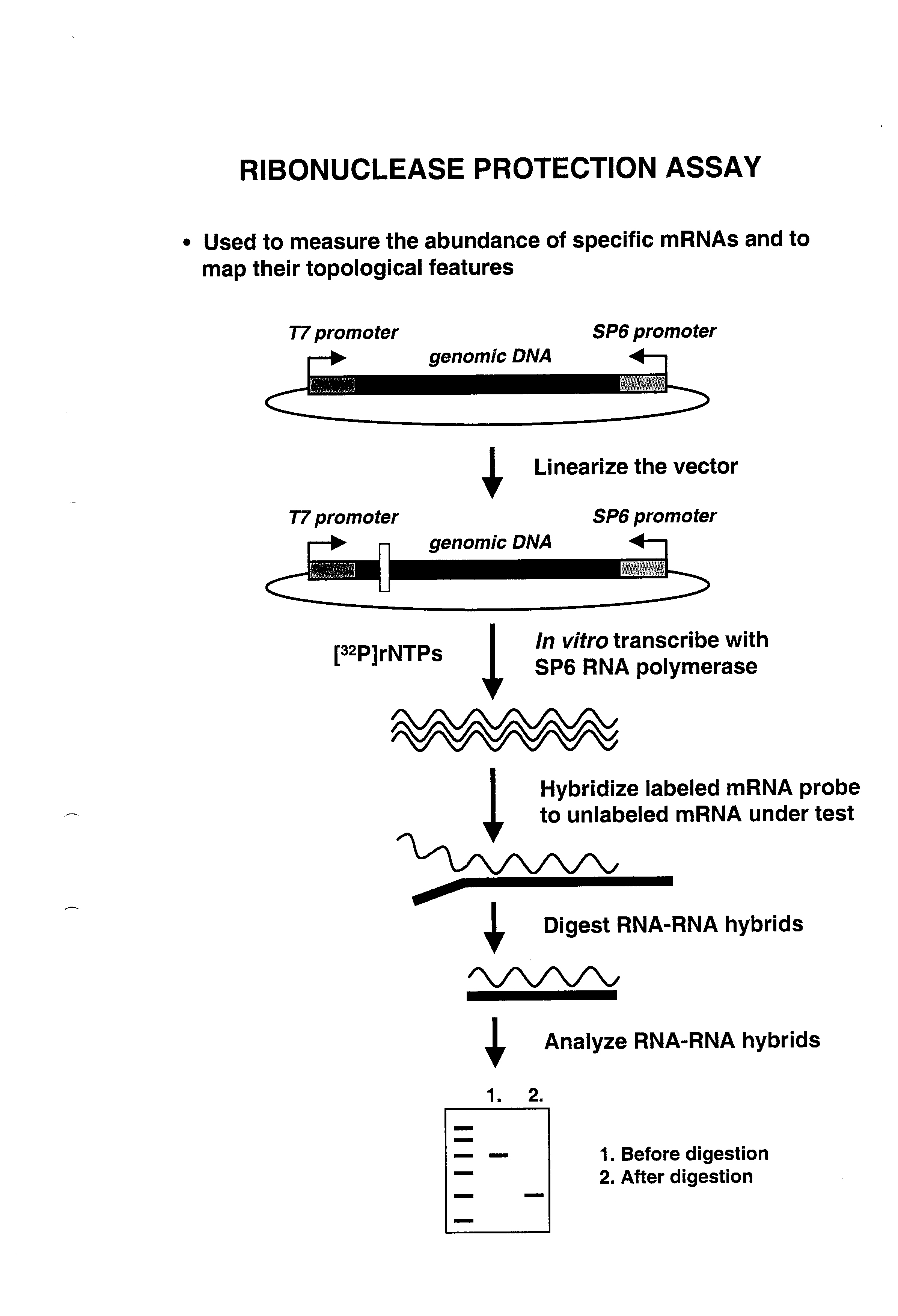
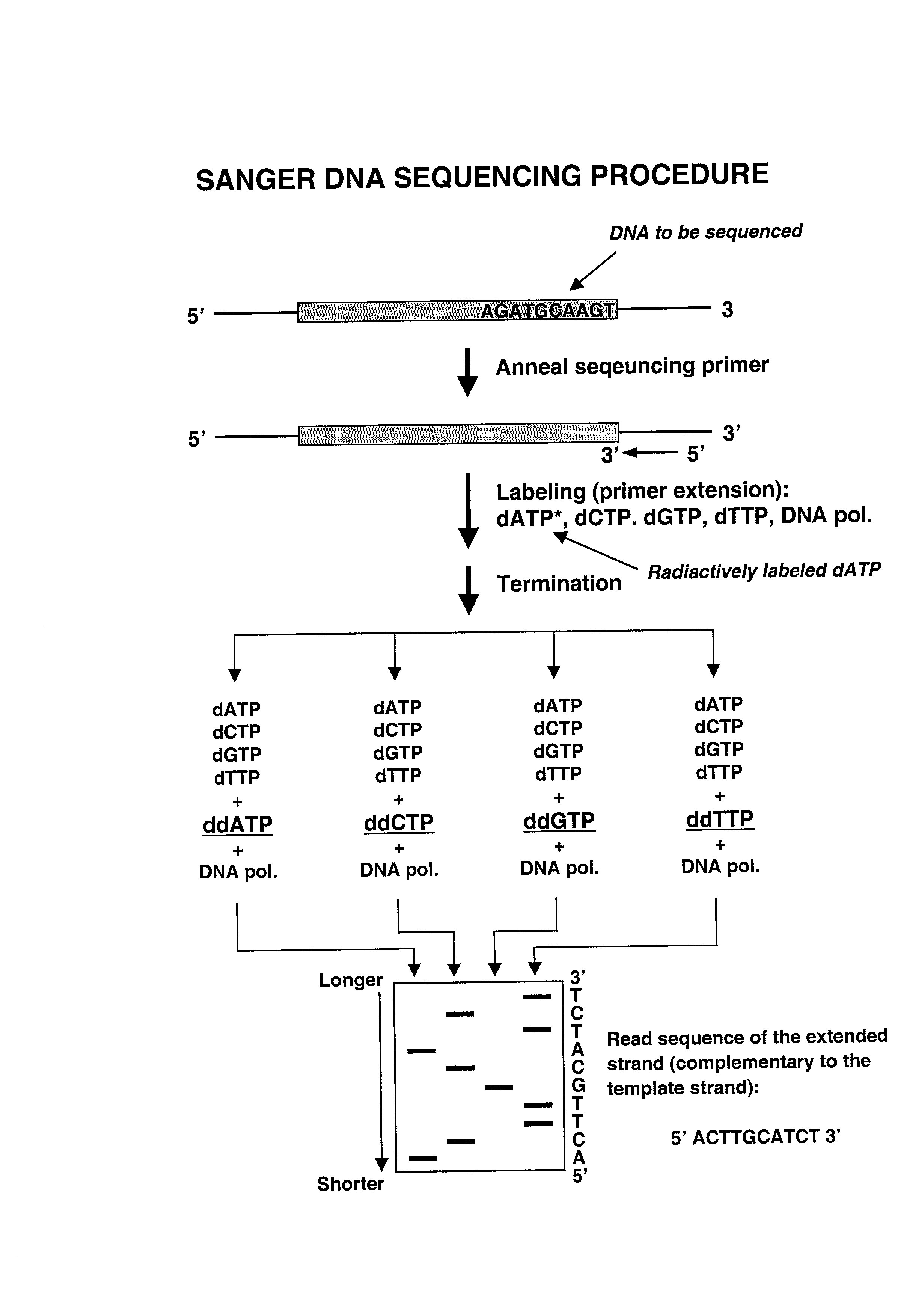
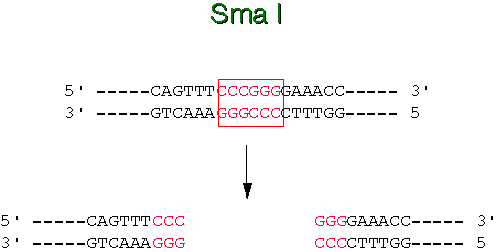
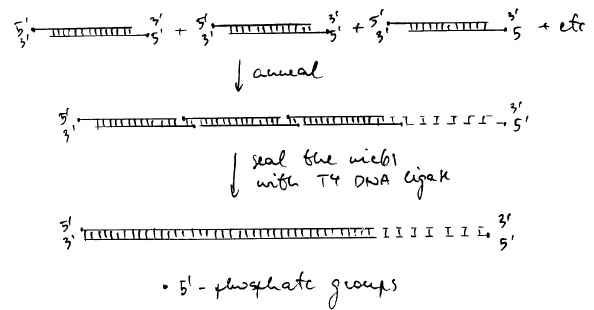




Kõik kommentaarid