Energia
on keha võime teha tööd. (vaja vähemalt teist keha, mille suhtes
tööd tehakse)
Töö
on füüsikaline suurus, mida möödetaks jõu ja jõu suunas läbitud
teepikkuse korrutisega
(A=fs
cosα) f on jõud, s on
teepikkus ja α jõu ja
liikumissuuna vaheline nurk. Töö ühik on
džaul (J)
Džaul
(J) on töö, mida teeb jõud üks Njuuton (N) ühe meetri
pikkusel teel.
Võimsus
on füüsikaline suurus, mida mõõdetakse ajaühikus tehtud töö
hulgaga . N=∆A/∆t
Energia
on keha võime teha tööd, kuid millegipärast ta seda veel ei tee.
Tööd ei tehta veel, sest puudub üks kahest vajalikust
komponendist , kas jõud või teepikkus, kuigi teine kahest on olemas.
Nt kui keha asetseb gravitatsiooniväljas, kuid veel ei liigu, siis
jõud mõjub, kuid läbitud teepikkus puudub. Niisugusel kehal on
potentsiaalne energia, nagu nt seda on
kõrgele tammi taha
paisutatud veel, mis veel alla ei lange.
Kineetilise
energia puhul on liikumine olemas, kuid jõudu vee mitte. Keha liigub
ühtlaselt ja sirgjooneliselt kui temale ei mõju jõud. Ehk keha
liigub inertsiaalselt, kuid ei kohta mingit teist keha. Niisugusel
kehal on kineetiline energia.
Energia
ei teki ega kao, vaid muundub ühest liigist teise. Ehk looduses
toimub kineetilise energia
muundumine potentsiaalseks ja
potentsiaalse energia muundumine kineetiliseks. Nt kui auto sõidab
vastu puud, siis kineetiline energia liigutab
plekke paigast ja
murrab luid. Kineetiline energia muutub potentsiaalseks, kui liikuvat
keha peatab jõuväli.
Kulon
(C) on laeng mis tõmbab teist samasuurt vastasmärgilist laengut 1 m
kauguselt jõuga 1N.
Plancki
konstant seob minimaalse võimaliku energia ja võnkesageduse.
h=6,626*10-34Js.
Potentsiaalide
vahe mõõtühikuks on volt (V). Elektriväljas kahe punkti
potentsiaalide vahe on 1 Volt kui laengu 1 Kulon üleviimisel ühest
punktist teise tehakse tööd 1 Džaul. Max potentsiaalide vahe mis
saab keemiliselt olla on 13V.
Elektronvolt
on töö, mida tuleb teha, et üks
elektron viia ühe voldi võrra
negatiivsemale potentsiaaline.
Mis
on
faraday arv? F=96500. Ühe mooli elektronide liikumisel
läbi potentsiaalide vahe 1 V tehakse tööd 96500 J. Faraday arv
tähistab tööd, mida tuleb teha, et üks
mool elektrone viia läbi
potentsiaalide vahe 1V.
Kvandi energia E=h
v. Kus
v=c/λ. Ehk E=hc/λ.
Nähtava
valguse
lainepikkus on vahemikus u
400-700 nm
Punase
valguse lainepikkus
625-740 (680) nm
Sinise
valguse lainepikkus
440-486 nm
Lainepikkus
antud, arvuta
footoni sagedus v
= c/λ. Nt λ=680nm(tee m-
deks ). c=3*108m/s.
v=4,4*1014s
Kui
suur on sinise valguse energia?
1,8 eV on punase kvandi energia. kui punane 680 nm. Kui sinine
valguse lainepikkus on 450nm, siis 680/450=1,5 ; 1,8*1,5=2,7. Ehk
sinisel kvandil oleks vastavalt suurem energia, nii mitu korda
suurem, kui palju tema lainepikkus on väiksem.
Iga
mitme vee molekuli kohta tuleb üks lahustunud aine molekul ? Ühes
liitris vees on 1000g/18gmol-1 = 55.6 mooli, st vesi ise
on 55.6 molaarne. Ühe
molaarses lahuses on seega 1 lahustunud aine
molekul ca iga 55 vee molekuli kohta. seega 55 vee molekuli vahel on
1 molekul ainet.
Miks
õhumolekulid 10km kõrguselt
alla ei kuku?
Kukuvad koguaeg,
aga juhuslikult
põrkuvad jälle üles. Kõik sõltub kuidas teised
põrgatavad üles või alla poole. Kogu
atmosfäär on molekulide
põrkumine , millest juhuslikult mõned satuvad kõrgemale, kukuvad
jälle alla, teised lähevad asemele jne.
Mool
on aine hulk, milles on
avogaadro arv molekule. NA
= 6.022*
1023 . Ehk üks mool ainet sisaldab endas Na
aineosakest.
Mooliks kutsutakse
gramm -molekuli. Gramm-molekul
on aine hulk
grammides , mis on arvuliselt võrdne selle aine
molekulmassiga Daltonites. Nt 1 mool vett on 18g.
Rõhk
on füüsikaline suurus, mida mõõdetakse pinnaühikule mõjuva
jõuga. Rõhuühikuks on N/m2=Pa. Gaasi rõhk tuleneb
sellest, et molekulid põrkuvad nõu
seintelt tagasi.
Normaalrõhk on
101300 Pa.
Temperatuur
iseloomustab gaasi molekulide liikumise kineetilist energiat. temp on
0 kui molekulid on paigal ja see kasvab võrdselt molekulide
kineetilise energia suurenemisega, st võrdeliselt kiiruse
ruuduga .
0oC=273K (norm temp).
Gaasi
olekuvõrrand pV=nRT. R=8.314 J/K*mol. 1 mol gaasi ruumala
normaaltingimustel (273K, 101300 Pa) on 22.4 l.
V=1*8,314*273.16/101300=22.4. toatemp ja normrõhul on mooli ruumala
24.
Molaarsus
– ainete kontsentratsioone väljendatakse ühikutes mooli/liitris
(molaarsus, M). Atmosfääriõhu kontsentratsioon toatemperatuuril on
1/24=42mM. Hapniku kontsentratsioon seejuures on 0,21*42=8,82mM. CO2
kontsentratsioon on 0.000365*42=15µM. (N2 78%, O2
21%, Ar 1%, CO2 0,037%)
RT
– näitab kui palju energiat tuleb kulutada ühe mooli gaasi
temperatuuri tõstmiseks ühe kraadi võrra. Sõltuvalt
vabadusastmete arvust (molekuli ehitusest) on see kas 3/2R∆T või
5/2R∆T. tegu on mooli soojamahtuvusega ehk erisoojusega konstantse
ruumala puhul. R on ühe mooli gaasi energia ühe kraadi kohta, RT on
ühe mooli gaasi energia T kraadi kohta. ehk RT on keskmise
soojusliikumise energia.
RTln
V2/V1. Kui mingisugune gaas kokku suruda või lasta paisuda, siis
mooli kohta tehtud töö on võrdeline logaritmiga
suhtest . Ei ole
absoluutsuurustest vaid suhtest, mitu korda me gaasi kokku surume.
Seega kui surusime kokku
hõreda gaasi, siis mooli kohta on töö üks
ja seesama. kui tihe siis kerge üks mool kätte saada. Niisuguste
masinate võimsus on väike kui on hõre faas ehk kui
kontsentratsioonid on väikesed siis selle masina võimsus on väike.
Kui kontsentratsioonid on suuremad siis võimsus on suurem.
Niisuguseid
protsesse gaasidega, kus väline soojusvahetus on täielikult
välditud, nimetatakse
adiabaatseteks. Lahustes
toimuvates protsessides, mis on bioloogias peamised, on temperatuur
tavaliselt
konstantne ja adiabaatseid nähtusi esineb harva.
Kuidas
mõistate entroopiat? Ebamäärane suurus, mille sisu
kokkuvõtlikult ei ole midagi muud kui muundumine korrastamatuseks..
kõrgema temperatuuriga väiksem hulk molekule kui panna meie heaks
midagi tegema siis tööd tehes see energia mis oli väiksemal hulgal
kiiremalt liikuvatel molekulidel jaguneb suurema hulga aeglasemalt
liikuvate molekulide vahel.Või suurtes sirgjooneliste liikumiste
portsjonites olev energia hajub paljudeks väikesteks ebamäärases
suunas liikuvateks portsjoniteks.
Membraanide
lipiidkihid ei juhi elektrit (elektrone ega ioone) ega ole
läbitav isegi vee molekulidele (vee jaoks valgud ja veekanalid et
membraani läbida). Kuid see ei võimalda elektrijuhtivust, sest
ioonid, kuigi ise mõõtmetelt väikesed, on ümbritsetud polaarsete
vee molekulide mitmekordse
kihiga ja seetõttu liiga suured, et
valkudes
liikuda . Na+, K+ ja H+ diffundeeruvad läbi membraani läbi
juhtivuskanalite. Suhkrud on neutraalse laenguga ja seega neid ei
pumbata ega saa sisse vedada.
Millest
tekib membraanil potentsiaalide vahe? Membraanis on ATP
seoselised ioonpumbad, mis ATPd hüdrolüüsides võivad H ioone
rakust välja pumbata. Ioonide liikumine tekitab kohe membraanile
potentsiaalide vahe. ATP seoselised pumbad on võimelised ioone
ainult rakust välja pumpama. H välja pumbates tekib rakku
neg potentsiaal. K
kanal avaneb ja K liigub sisse negatiivse
potentsiaali tõmbejõu mõjul. See neutraliseerib küll membraani
potentsiaali, kuid H+ pumpamist jätkates saab rakus luua
suteliselt kõrge (kümnetesse mM ulatuva) K+ ioonide
kontsentratsiooni. See rakusisene kõrge K+
kontsentratsioon ongi rakumembraanil toimuvate protsesside peamiseks
energeetiliseks
kandjaks .
Jaotusfunktsioon
– mis on Y
teljel ? X-teljel on molekulide kiirus m/s. y-
telg omab
dimensiooni dn/ndv. See
dimensioon võimaldab leida molekulide
suhtearvu, mille kiirused asuvad vahemikus v kuni v+dv, kui
tulba pindala, mille laius on dv ja kõrgus f(v). On mõistlik küsimus,
kui suur osa molekulidest omab kiirusi mingis vahemikus, nt
400-410m/s. ebamõistlik küsimus, kui suur osa molekulidest omab
kiirust täpselt
400m /s, sest vastus sellele on: niisuguseid molekule
ei ole.
Bolzmanni
faktor näitab kui suur osa molekulidest on võimeline mingiks
keemiliseks reaktsiooniks.
Ensüümid
ei muuda reaktsiooni
kulgemise energeetilist lõppefekti, küll aga
vähendavad oluliselt vahepealset energiabarjääri,
aktivatsioonienergiat.
Keemiline
tasakaal on olukord, kus reaktsiooni kiirused edasi- ja
tagasisuunas on võrdsed. Tasakaalu minek on seda kiirem mida väiksem
on aktivatsioonibarjäär, aga
barjääri suurusest ei muutu
lõpptulemus.
Aine
potentsiaalne keemiline energia ehk
keemiline potentsiaal on
suurus mida mõõdetakse reaktsiooni käigus vabanenud energiaga
mooli kohta. lahuses tulevad sisse ka teised jõud. Seda komponenti
nimetatakse standardseks keemiliseks potentsiaaliks µ0.
Kui aine A muutub aineks B, siis
summaarne energeetiline efekt oleks
.
Kui reaktsioonis osalevate ainete kontsentratsioonid on võrdsed,
ainult siis on reaktsiooni energeetiline efekt võrde molekulisisese
muutuse poolt põhjustatuga µ0 [J/mol]. Reaktsiooni
energeetiline efekt läheneb
nullile , kui
reaktsioon toimub keemilise
tasakaalu lähedal.
Kui
reaktsioonis toimuv keemiline potentsiaali muutus oleks 96500 J/mol,
võiks öelda, et reaktsioonis olevate ainete redokspotentsiaalide
vahe oleks 1V.
ATP
laialdane kasutatavus metabolismis tuleneb energiarikka sideme
ülekandevõimalusest, fosfaatrühma ülekande teel. ATPs on rida O
aatomeid paigutatud lähestikku
lineaarses struktuuris. O aatomite
vahel valitseb tugev
tõukejõud , mis püüab fosfaatrühmi
üksteisest lahku tõugata.
Fotokeemiliselt
on aktiivne ainult
punase kvandi energia. fotosünteesi
mehhanism lõikab lühemalaineliste kvantide energia maha ja muudab
selle soojuseks.
Fotosünteesis
eraldub O2 veest, mitte CO2-st. CO2 + 2H2O
+ 8hv ->
CH2O + H2O + O2.
Kogu
fotosünteesi protsessi jagatakse kaheks osaks, “valgusreaktsioonid”
ja “pimereaktsioonid”. “Valgusreaktsioonides” lagundatakse
vesi, kuid CO2 taandava jõuna ei eraldu mitte vesinik,
sest see lenduks, vaid universaalne bioloogiline e- kandja
NADPH . Fotosünteesi “pimereaktsioonides” NADPH ja ATP
kasutatakse CO2 taandamiseks.
Calvini tsükli vaheproduktid on
fosfaadid .
CO2
taandamiseks kasutatakse 4 elektroni energia, mis eralduvad veest ja
mille abil kahe kaksiksidemega CO2 struktuur ümber kujundatakse
nelja üksiksidemega struktuuriks. Tekkiva suhkru koosseisu jääb
aga neljast elektronist kaks.
Tülakoidi
membraansüsteem moodustab sisemise ruumiosa, luumeni, mis on
stroomast membraaniga eraldatud. Luumeni ja
strooma vahel tekib H
kontsentratsiooni
elektrokeemiline gradient, mis energiseerib ATP
sünteesi. Tülakoidi membraanides asuvad valgust neelavad
fotosüsteemid I ja II ja nendevahelised elektronikandjad
plastokinoon, tsütokroom b6f, plastotsüaniin, samuti ka NADP
reduktaas ja ATP süntaas. CO2
sidumise ja taandamise reaktsioonid
aga toimuvad tülakoidide vahel asuvas
stroomas .
Valgusreaktsioonideks
nimetatakse valguse neeldumist ja elektronide ülekandega seotud
reaktsioone. Valguse abil toimub tegelt ainult elektroni ergastumine
pigmendil. NADPH on universaalne elektronikandja. H-
ioonidel on
eriline roll ATP sünteesis, kus need, liikudes läbi membraani
kontsentratsioonigradiendi ja elektripotentsiaalide vahe mõjul,
annavad ATP sünteesiks vajaliku energia. valguse neeldumise
tulemusena tõuseb elektron klorofülli molekulis kõrgemale
energianivoole.
Klorofüllid a ja b neelavad peamiselt punases ja
sinises spektriosas. Kloroplastis abipigmendid,
karotenoidid ,
rohelise ja kollase valguse kasutamiseks. Abipigmendid annavad oma
ergastuse edasi klorofüllile. Vett lagundav
kompleks on
fotosüsteemiga ühendatud tülakoidi luumenipoolsel küljel. selle
koosseisus on tähtsad neli Mn aatomit. Lõppaktseptoriks on
kinoon QA.
Trap limited mudel – eksiton liigub pidevalt
antenni ja tsentri vahet.
Elektron
eraldub tsentripigmendis, mis PSIIs on P680 ja
PSIs P700 (tähistatud
vastavalt lainepikkusele). Tsentripigment on klorofüllide dimeer.
Lisaks on läheduses veel
PSII feofütiin. Lõpuks elektron
stabiliseerub aktseptorpoolel kinooni
molekulil . PSIIs
redutseerunud plastokinoon QB eraldub ja difundeerub membraani sees,
seda asendab teine, oksüdeerunud kinoon. Redutseerunud kinoon
difundeerub Cyt b6f kompleksini kus oksüdeerub
andes elektroni edasi
Cyt b6f kompleksis olevatele heemidele Cyt f ja Cyt b. PSI liigub
elektron edasi seotud FeS kandjatel kuni jõuab ferredoksiinile. See
on esimene stroomas lahustuv elektronkandja. Lõplikuks elektroi
aktseptoriks on NADP.
Fotosüsteemid
moodustavad tandemi. PSII
lagundab vett, kannab elektronid
plastokinoonile, see neutraliseerub sidudes prootoni, difundeerub
tsütokroom b6f kompleksile, kus oksüdeerub andes elektroni Cyt f
kaudu plastotsüaniinile, viimane difundeerub PSI juurde ja annab
elektroni valgusenergia abiga ferredoksiinile ja sealt edasi NADP-le.
Vee
lagundamise aparaat – elektronid eemalduvad ühekaupa iga
valgusvälgu ajal niisugusest 4Mn kompleksist, mis akumuleerib 4
positiivset vakantsi, mis siis korraga täituvad 2 H2O
oksüdeerimisel. Fotosüsteem II aktseptorpoolel on kaks kinooni Qa
ja Qb. Qa on valgus kinni ja annab elektroni Qb-le. Kui Qb-l koguneb
kaks elektroni, lisanduvad stroomast
prootonid ,
neutraalne kinoon
vabaneb ja difundeerub membraani lipiidfaasis tsütokroom b6f
kompleksile, kus oksüdeerub, andes elektronid tsütokroomi
rauale ,
prootonid vabanevad luumenisse.
Kinooni
eripära – et ATPd on võimalik sünteesida.
Q- tsükkel
– tsütokroom võtab korraga vastu vaid ühe elektroni. Tsütokroom
b6f kompleksis olev tsütokroom b6 redutseeritakse. Tsütokroom f
asub membraani luumeni külje lähedal ja tsütokroom b6 asuvad
strooma poolsel küljel. kui kinoonilt esimene elektron liigub Cyt
F-le, siis kohe teine elektron redutseerib ühe Cyt b6, mõlemad
prootonid aga eralduvad luumenisse. Sama juhtub järgmise kinooniga,
sest cyt f on
vahepeal osküdeerunud, andes elektroni
plastotsüaniinile, teine kahest cyt b6 –st on aga veel elektronita
ja redutseerub nüüd. Kaks cyt b6 koos redutseerivad he osküdeeritud
kinooni. See kinoon vabaneb ja sarnaselt iga teise
topeltredutseeritud kinooniga oksüdeerub cyt b6f andes üle
elektroni cyt f-le ja teise jällegi cyt b6-le. Kokkuvõttes käib
üks elektron ühe, teine kaks ja kolmas kolm korda läbi cyt b6f
kompleksi, kandes iga kord kaasa prootoni.
ATP
süntaas – prootoneid pumbatakse stroomast luumenisse
elektrontranspordiga paaris, tagasi stroomasse ringlevad nad aga läbi
ATP süntaasi, mis kasutab prootongradiendi energiat ATP sünteesiks.
Sünteesib ühe täispöörde kohta 3 ATP molekuli.
Membraanis on neli
suuremat kompleksi: Fotosüsteemid I ja II, nende vahel elektrone
piki membraani edasiandev ja samal ajal prootoneid risti membraani
läbi kasndev tsütokroom b6f ja ATP süntaas, mis prootonid uuesti
välja laseb ja nende energia arvel ATPd sünteesib. PSII ja Cyt b6f
vahel kannab elektrone membraani sees difundeeruv plastokinoon, Cyt
b6f ja PSI vahel aga kannab elektrone luumenis difundeeruv
plastotsüaniin. Kokku jääb iga elektroni kohta luumenisse kolm
prootonit (3H+/e-), millest üks eraldus vee
lagundamisel ja kaks seoses plastokinooli oksüdeerimisega Cyt b6f
kompleksil. Nelja elektroni kohta transporditakse 12 H+,
mis võimaldavad sünteesida 3 ATP. Niiviisi, koos nelja elektroni
jõudmisega transportahela lõppu (2 NAPH sünteesiga) sünteesitakse
paralleelselt ka 3 ATP, stöhhiomeetriliselt vajalik hulk ühe CO2
sidumiseks.
NADPH
ja ATP sünteesiga lõppevad valgusreaktsioonid.
Pimereaktsioonideks
nimetatakse CO2 sidumise ja taandamise reaktsioone. CO2 taandatakse
suhkruks. Paralleelselt süsiniku taandamisega suhkruks tarbitakse
3ATP iga taandatud C kohta.
CO2
siseneb lehte difusiooni teel õhulõhede kaudu. Lehe sisse jõudes
difundeerub CO2 kiiresti laiali rakkudevahelistes õhuruumides. CO2
liikumise viimane etapp hõlmab lahustumise rakuseina
vedelikus ja
difusiooni vedelas faasis kuni ensüümini, mis CO2 seob ja muudab
taandamiseks valmisolevateks
vaheproduktideks .
Rubisco on CO2
siduv
ensüüm . Väga aeglane ensüüm. Ensüümi peab palju olema
mõistliku CO2 sidumise kiiruse saavutamiseks. Karboksüülimine
tähendab CO2 sidumist substraadile, milleks on ribuloosbisfosfaat.
Tekib kuuesüsinikuline
produkt , mis laguneb kohe kaheks
kolmesüsinikuliseks. Need ongi elektrone vastuvõtvaks substraadisk,
et saada muudetud kolmesüsinikuliseks suhkruks. Ühekaupa ei saa CO2
taandada, mürgise sipelghappe tekkimise pärast. Seetõttu CO2
taandatakse 3-5 süsinikuliste vaheühendite kaudu. CO2 sidumise
tsüklit nn
Calvini tsükliks. Seda võib jagada kolmeks: CO2
aktseptori karboksüülimine(CO2 sidumine), taandamine (suhkru
moodustumine) ja aktseptori regeneratsioon. Lisaks NADPH-le vajatakse
ka ATPd, mis valmistab karboküülimise
produktid ette taandamiseks
ja regenereerib kaks korda fosforüülitud aktseptori.
Regeneratsiooniosas tehakse viiest kolmesüsinikulisest kolm
viiesüsinikulist suhkrut.
Fotohingamine – lisaks CO2-le võib Rubisco abil
ribuloosbisfosfaadile liituda ka O2. Tekib viiesüsinikuline produkt,
mis laguneb 2 ja 3-süsinikuliseks. 3-süsinikuline taandatakse,
2-süsinikuline viiakse metabolismiahelasse, kus kaks molekuli
liidetakse kokku, saadakse 4C ühend, siis eraldatakse CO2 ja järgi
jääb 3C ühend. See muudetakse jälle fotosünteesimehhanismis
taandatavale kujule, CO2 aga väljub lehest. Fotohingamise üks mõte
on et kaitsta ülereduktsiooni eest.
C4 fotosüntees i on mitu tüüpi. NADP-
malic enzyme tüübi
puhul PEP karboksüülimisel tekkiv oksaalatsetaat taandatakse
malaadiks, mis difundeerub pärjarakkudesse ja dekarboksüülitakse
seal NADP seoselise malic ensüümi abil. Sel juhul CO2 eraldumist
saadab NADP taandamine, niiet koos CO2ga imporditakse pärjarakkudesse
ka NADPH. Seda tüüpi taimedes toimub pärjarakkudes ainult ATP
süntees tsüklilises elektrontranspordis, vee lagundamist ja NADPH
sünteesi ei toimu. PSII puudub pärjarakkudes üldse.
CAM metabolism – kaktustes. Öösel glükolüüsi raja kaudu
sünteesitakse PEP, mis karboksüülitakse sidudes CO2 ja tekkinud
malaat salvestatakse vakuoolis. Õhulõhed on avatud. Päeval malaat
dekarboksüülitakse ja tekkinud CO2 kasutatakse tavalise C3
fotosünteesi substraadina. Õhulõhed on päeval suletud,
fotosüntees toimub sisemise CO2 arvel.
Kirjeldage
tärklise ja sahharoosi sünteesi erinevusi. Tärklise süntees
toimub kloroplastis, kus päeval kogunevad
tärkliseterad . Öösel
need hüdrolüüsitakse ja vabaneb
glükoos , mis väljub
kloroplastist. Tärklise süntees vajab ATPd ja selle ahela tähtsaim
ensüüm ADP glükoosi pürofosforülaas on aktiveeritav PGA, F6P ja
trioosfosfaatide poolt. sahharoosi süntees toimub tsütosoolis
P-translokaatori kaudu väljatoodud trioosfosfaatidest. Sahharoosi
sünteesi kiirust reguleerib tsütosooli F6Bpaas, mis blokeeritakse
F2, 6BP
molekuliga . Viimase süntees ja lagundamine on
reguleeritavad trioosfosfaatide ja Pi tasemega. Kõrge T3P ja madal Pi vivad
inhibiitori taseme alla, madal T3P ja kõrge Pi aga tõstavad seda.
Nii hoitakse sahharoosi sünteesi kiirus tasakaalus fotosünteesi
kiirusega. Sahharoosi sünteesil kasutatakse UDP-glükoos
pürofosforülaasi.
Tärklis
on sahharoosi varuprodukt mida on vaja ööseks
tarbimiseks alles
hoida. Varud saab säilitada polümeeridena, väikeste molekulide
säilitamine võimatu, kuna tekitab suure
osmootse rõhu.
Hingamine on fotosünteesi pöördreaktsioon produktide ja substraatide
vahetuse mõttes.
CH2O + O2
-> CO2 + H2O.
Nagu fotosünteesi puhul, ka hingamise puhul on tegu nelja
elektroni liikumisega CH2O ühikult hapnikule. Suhkru ühe C-ühiku
oksüdeerimisel vabaneb 4 elektroni energia, mis
kanduvad üle O2-le
moodustades vee. Suhkru koosseisust otse võetakse kaks elektroni,
kaks aga hüdrolüüsil veest.
Glükolüüs
– nii nagu sahharoosi süntees, nii ka glükolüüs toimub
tsütoplasmas. Glükolüüsi käigus poolitatakse 6-süsinikuline
molekul. Molekuli poolitamiseks on vaja äärmised C-d fosforüülida,
kulutades selleks 2ATP-d. Selleks tuleb glükoos viia enne lineaarse
fruktoosi kujule.
Niisiis algab glükolüüs ATP tarbimise mitte
selle sünteesiga. Kahest otsast fosforüülitud fruktoos bisfosfaat
laguneb vastava heksokinaasi toimel kaheks trioosfosfaadiks. Need
oksüdeeritakse, viies 2 elektoni üle NADP+-le, NADH moodustumiseks.
Toimub substraatne fosforüülimine. Fosfaadijääk seotakse
ensüümiga, kust see koos elektroni ülekandega NAD+le kantakse üle
trioosfosfaadile, seda nüüd kahekordselt fosforüülides. Siin
elektroni ülekandel vabanevat energiat kasutatakse ATP sünteesiks
otseselt, ilma membraani ja prootongradiendi vahenduseta.
Bisfosfoglütseerhape aga on
sedavõrd tugeva sisemise pinge all, et
fosfaatrühm võib üle kanduda ADP-le, jättes järele
3-fosfoglütseerhappe (3-PGA). Sellega oleme raja alguses kulutatud
ATP tagasi saanud, aga on järel veel kaks monofosfaati (3-PGA). See
aine ise pole ATP substraatseks fosforüülimiseks võimeline, kuid
selle enoolvorm on. Seega, nüüd viiakse fosfaatrühm üle molekuli
keskele (3-PGA +> 2-PGA) ja viimane keeratakse enoolvormi,
moodustades fosfoenoolpüruvaadi (PEP). Viimase defosforüülimisel
vabaneb sedavõrd palju energiat (produkt püruvaat on kompakne
ühend), et
fosfaat saab ADP le üle kanduda, andes nüüd
kummagi trioosi kohta ühe ATP puhaskasuks. Seega on glükolüüsi
lõpptulemus ühe glükoosi (6 C) kohta 2 ATP ja 2 NADH. Hapniku kui
NADH oksüdeerija olemasolu korral kantakse elektronid sellele,
moodustades vee ja aidates kaasa suurema hulga ATP sünteesile, koos
Krebsi tsüklist vabaneva NADH ga.
Krebsi
tsükkel – püruvaadi oksüdeerimine. 3C happe oksüdeerimine,
nii et vähem kui 4C ühendit kunagi ei teki. Püruvaat siseneb
mitokondrisse ja reageerib koensüüm A-ga. Selle reaktsiooni käigus
vabaneb üks süsinik CO2 kujul ja kaks elektroni regutseerivad ühe
NAD. Atsetüüljääk koensüümilt ei vabane vaid kandub üle 4C
aktseptorile(oksaalatsetaat) moodustades 6C
tsitraadi . Tsitraadi
edasisel oksüdeerimisel vabaneb veel 2 CO2 ja tekib 2 NADH. Tsükli
teises
pooles veel NADH ja FADH2. Kogu hindamisahela kohta tervikuna
eraldub 4 elektroni CO2 kohta. koensüümA on ATP
derivaat , milles
kolmas fosfaat on asendatud pika ahelaga mille lõppu väävli kaudu
ühildub atsetüülijääk. Kõik vaheproduktid võivad
ahelast väljuda, pannes aluse aminohapete, rasvhapete ja nukleiinhapete
sünteesile. Osmootse rõhu konstantsuse tagavad translokaatorid.
Mitokondriaalne
elektron- prooton -transport ja ATP süntees - Aeroobsetes
tingimustes (O2 olemasolu) oksüdeeritakse tekkinud NADH (ka FADH2)
mitokondrite elektrontransportahela poolt, mis viib elektronid
hapnikule, olles aga seejuures paardunud transmembraanse
prootontranspordiga. Viimase poolt genereeritud energiat kasutatakse
ATP sünteesiks, lastes prootonid läbi ATP süntaasi maatriksisse
tagasi voolata.
Fotosüntees
teeb ATPd enda jaoks, hingamine teeb teiste jaoks, ise ei vaja.
Mitokondri membraanis samasugused
kompleksid nagu kloroplastis. Sealt
kust NADH sisse lähevad nim kompleks I, kompleks II vahepeal, edasi
kompleks III ning lõpuks kompleks IV (kus toimub el liikumine
hapnikule ja vee tekkimine). el antakse kinoonile, tema annab
tsütokroomile. Toimub Qtsükkel. el liigub veefaasis (ennem
lipiidfaasis membraanis). Lõpuks el hapnikule ja tekib vesi.
Mitokondriaalses elektrontranspordiahelas on neli peamist kompleksi.
Kompleks I kaudu sisenevad elektronid NADH –lt. Kompleks II kaudu
sisenevad elektronid FADH2 –lt (suktsinaat-fumaraat üleminekul).
Kompleks III
teostab Q-tsüklit (tsütokroom b6c kompleks) ja
Kompleks IV redutseerib hapnikku, moodustades vee. Lisaks võivad
elektronid siseneda veel otse rasvhapetest ja taimedes ka NADPH-lt.
Pange tähele, et prootoneid transporditakse maatriksist välja ja
need
voolavad tagasi läbi ATP süntaasi. ATP sünteesitakse
maatriksis.
Kompleks
I oksüdeerib NADH, transportides samal ajal 4H+/2e- , nii nagu selle
sees töötaks veel seniavastamata Q-tsükkel (punktiirjoon).
Elektronid kanduvad ubikinoonile, mis haarab samuti 2H+/2e-. Joonisel
on näitamata, et NADH oksüdeerumisel eraldub selle koosseisust H+,
mis vähendab üldise prootontranspordi suhte Kompleks I juures
6H+/2e-, asemel 5H+/2e-
tasemele . Prootonid pumbatakse mitokondri
kristast välja, membraanide vahelisse ruumi, ning see muutub
aluselisemaks. ATP tuleb välja viia ja ADP sisse tuua. Kui nad
üksteise suhtes vahetada, siis välja viiakse rohkem ioone kui sisse
tuuakse ning seega tuleb ka fosfaat selle sissetoomisega ühendatud.
Kompleks
II saab elektronid otse suktsinaat-fumaraat reaktsioonist, mis toimub
membraanile ankurdatud Kompleks II juures, mitte vabalt difundeeruva
NADH-ga, nagu Kompleks I juures. Elektronid kanduvad FAD ja
kompleksile seotud ubikinooni kaudu vabale ubikinoonile, mis seob
maatriksist 2H+/2e- (võrdle, ka PSII
juurtes kanduvad elektronid
vabale kinoonile seotud kinooni kaudu) . Siin rohkem prootoneid üle
ei
kanta kui vaba ubikinooniga seostuvad.
Kompleks
III on
vaste tsütokroom b6f kompleksile kloroplastides. Erinevus on
see, et tsütokroom f asemel on tsütokroom c, aga mõlemad on ühte,
c-tüüpi. Cyt f
tähistus tuleneb sellest et tema avastati taimedes.
Q-tsükkel töötab siin samamoodi kui kloroplastis (siin ta
avastatigi). Aga järgnevaks vabaks e- kandjaks ei ole mitte
plastotsüaniin, vaid tsütokroom c.
Kompleks
IV (tsütokroom c
oksüdaas ) saadab elektronid hapnikule, moodustades
vee. Kuna energiahüpe on suur, ei ole siin 4e- Mn kompleksi, nagu
fotosünteesis ja hapniku taandamise
molekulaarmehhanism on erinev
vee oksüdeerimise molekulaarmehhanismist.
Erinevused
ATP tootlikkuse vahle hingamises ja fotosünteesis - Elektronide
liikumise põhimõtted on samad. (väidetavalt hingamises
produtseeritakse ühe glükoosi kohta (6C) 36 ATP, ehk 6ATP eralduva
CO2 kohta. seda on 2 korda rohkem kui fotosünteesis). Hindamisel
saadakse substraatse fosforüülimisega 2 glükolüüsis ja 2 Krebsi
tsükklist ehk 4 ATP-d. Iga NADH kohta saame 9 ja iga FADH2 kohta 6
ülekantud H+. kokku u 100 ATP sünteesi tarbeks. ATP süntaas nõuab
kas 4 või 3 H+ iga ATP kohta ja ADP ja Pi sissetransport ühe H+.
seega ühe ATP sünteesiks kulub kas 6 või 5 H+. kokku tehakse umbes
16ATPd, lisades substraatsel teel sünteesitud, saame kokku 18.
Arvestades ka lisaprootoneid on summaks umbes 22. Kui prooton kulu
on 5H+ ATP kohta, saame kokku 22. Kui kompleks IV kannab
lisaprootoneid läbi membraani, on see number pisut suurem u 26.
Fotosünteesis toodetakse 4 elektroni kohta 3 ATPd.
Kloroplast sünteesib ATP ainult endajaoks.
Närvirakus
Na
kanalite avanemisega põhjustatakse
depolarisatsioon , mis lõpeb
väga kiiresti. Repolarisatsioonil osalevad K kanalid, mis
avanevad depolarisatiooni ajal pingetundlikult. Toimub nagu Na-K kanalite
omavaheline võitlus, kuid mõlemad korraga laialt avatud ei ole.
Selle kindlustab Na kanalite ehitus, mis avab kanalid
pingetundlikult. Pingetundliku kanali suleb
stopper -
segment ja kanal
pöördub algasendisse.
Na
ja K kanalite tähtsaim omadus on nende ioonide vahel vahet teha.
Ioonide
mõõdud väga sarnased, kuid siiski väiksem
ioon ei läbi
suuremale mõeldud kanalit. Moodustuvad koorinatsioonisidemed metalli
ja poori seinte aatomite vahel, nende sidemete täpne pikkus
kindlustab selektiivsuse.
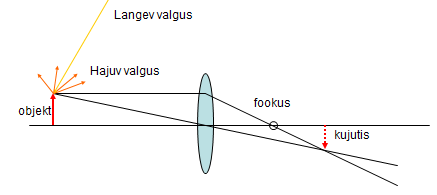
Joonista optilist kujutist.Lääts
on sellise kujuga, kui tema peale langevad paralleelsed kiired, mis
tulevad otste poolt, murduvad rohkem, mis rohkem keskelt poolt
murduvad vähem ja täpselt keskelt ei murdu üldse. Punane valgus
murdub vähem, seega o keskelpool ja sinine murdub rohkem ja on otste
poolt. kõik paralleelsed kiired lõikuvad
fookuses .
Silma
valgustundlikkus põhineb retinaali cis-trans isomerisatsioonil.
Alguses cis-retinaal opsiiniga kompleksis. Valguse
neeldumine ergastab pii-sideme, see hetkeks katkeb. Tekib trans vorm. Nüüd on
rodopsiin aktiveerunud ja edastab sidnaali membraani Na-kanalitele.
Trans-retinaal eraldub opsiinist, ebsüümi abil pöördub cis-vormi
tagasi ja liitub uuesti opsiiniga, olles valmis järgmiseks
impulsiks .
Tavaseisus
kepikeste membraanis palju avatud Na
kanaleid , mis depolariseerib
rakumembraani. Aktiveerunud rodopsiin
algatab GTP hüdrolüüsiga
seotud ahela, mis katalüüsib cGMP ülemineku GMPks, mis põhjustab
K kanali
juhtivuse suurenemise ja membraani polarisatsiooni.




























Kõik kommentaarid