katalüüsivad ensüümid. LIPOGENEES LIPOLÜÜS - rasvade hüdrolüütiline lagundamine LIPOGENEES - eeskätt uute rasvhapete süntees Loomadel toimub aktiivselt rasvkoe-, piimanäärme- ja maksarakkude tsütoplasmas, taimedel - kloroplastides. RASVHAPETE SÜNTEESI LÄHTEAINED · Atsetüül-KoA CH3CO~SkoA - süsinikuallikad · HC03-(C02) · NADPH+H+ - redutseerija · ATP - energiakandja · Rasvhapete süntaas - multiensüümkompleks Rasvhapete süntaas koosneb kuuest erinevast ensüümvalgust ja ACP-st (bakteritel ja taimedel) või ühest suurest multifunktsionaalsest valgust (selgroogsetel), mis omab seitset erinevat ensüümaktiivsust; sisaldab kahte -SH rühma, mis seovad makroergilise tioestersidemega rasvhapete atsüüle. ACP ehk ACP-SH (acyl carrier protein) - atsüülikandev valk; väikese molekulmassiga valk, mis toimib kui sünteesi vaheühendite kandja. LIPOGENEESI STAADIUMID
Koosneb dinitrogenaasist ja dinitrogenaasi reduktaasist. Taandab õhus sisalduva N2 ammooniumiks. Kasutab ATP hüdrolüüsil vabanevat energiat N 2-s Sisalduva kolmiksideme lõhustamise reaktsiooni aktivatsioonibarjääri ületamiseks. Inaktiveerub õhuhapniku toimel. 3. Selgitage milline on glutamaadi ja glutamiini funktsioon ammooniumi assimileerimisel. Kirjeldage reaktsioonid, mida katalüüsivad glutamaadi dehdrogenaas, glutamiini süntetaas ja glutamiini süntaas. Nende reaktsioonidega toimub anorgaanilise lämmastiku sisenemine orgaaniliste ühendite koosseisu. Glutamaadi dehüdrogenaas kasutab NADPH redutseerivaid ekvivalente ammooniumi sidumiseks - ketoglutaraadiga. Glutamiini süntetaas on põhiline ensüüm mida kasutatakse ammooniumi fikseerimiseks. Kasutab ATP energiat reaktsiooni läbiviimiseks. Substraadiks glutamaat. 4. Asendatavad ja hädavajalikud aminohapped. Asendatavad aminohapped on sellised mille biosünteesi rajad on loomadel olemas
ee mudelitega. (Otsing: “Rakumaailm”) Fotosüntees. 1. Ava mudel 1, lahenda see ja vasta küsimustele. Kasuta ka teooria lehekülge. a) Millise molekuli lisasid valgusstaadiumis esimesena? H20 b) Mis aine molekulid on suured lillad? NADP c) Mis Miki-Hiire taoline ühend tekib lillale molekulile siniste kõrvade lisamisel? NADPH2 d) Vesinikioonid liiguvad raku välispinnale läbi ensüümi ATP süntaas. Mis ülesanne on sellel valgulisel ainel? Salvestab nende väljumisega vabaneva energia ATP molekulidesse. e) Pimedusstaadiumis lisasin CO2 molekuli. See vallandas reaktsiooni, mille tulemusena tekib glükoos. f) Mis juhtub pimedusstaadiumis ATP molekulidega? Eraldub üks fosfaatrühm, tekib ADP. 2. Ava mudel 2, lahenda see. Mida pidid selles mudelis tegema teisiti võrreldes esimese mudeliga
reoksüdeeruvad). 1. Andke lühike seletus järgmistele mõistetele a. Mitokondri krista - sisemembraani sopististe kurrud(sisemembraanistik) b. mitokondri maatriks - ruum, mis jääb sisemembraanistiku vahele c. intermembraanne(membraanide vaheline) ruum - ruum, mis on sise ja välismembraani vahel d. tsütokroomid - valgud, mis paiknevad membraanidevahelises ruumis. Käituvad elektronide ülekandjatena. [membraanvalgu kompleks, mis vastutab fotosünteesi eest sini-rohebakterites] e. ATP süntaas - ensüüm, mis sünteesib ATP-d 2. Selgitage hingamisahela e. elektronide transpordisüsteemi (ETS) kohta järgmist: a) Millises raku kompartmendis ETS lokaliseerub ja funktsioneerib? Toimub eukarüootsetes rakkudes mitokondri sisemembraanisvõi selle pinnal (membraaniprotsess). b) Millistest komponentidest ETS koosneb? Dehüdrogenaasid, tsütokroomid, Fe-S valgud (viimased kaks on elektronide edastajad).
Fotosüsteem II nimetatakse tema ensüümide järgi veel plastokinooni oksidoreduktaasiks. Oksidoreduktaas on valgussõltuv ensüüm, mis kasutab valgusenergia footoneid elektronide ergastamiseks. Seejärel transporditakse need üle erinevate koensüümide ja kofaktorite, et redutseerida plastokinoon plastokinooliks. Vee molekuli lagundamisel saadud vesinikioonid e prootonid aitavad tekitada prootongradienti, mida kasutab teine tülakoidide membraanis paiknev valkkompleks ATP süntaas, mis genereerib ATPd ADPst ja anorgaanilisest fosforist. ATP süntaas pumpab prootoneid kloroplastide stroomasse. Eraldub hapniku molekul. On teada, et fotosüsteem II asub tülakoidide membraanis nii, et vett oksüdeeriv sait ehk doonorsait on suunatud tülakoidide sisemuse ehk luumeni poole ja plastokinooni reduktaasi sait ehk aktseptorsait asub stroomapoolsel küljel. FSII absorbeerib kiirgust max 680nm juures (klorofüllid a ja b ning lisapigmendid: P680). Terminaalsed
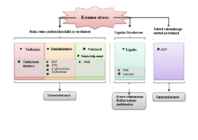
Kuuma stress Teised rakuseinaga Raku seina polüsahhariidid ja ensüümid Ligniini biosüntees seotud proteiinid Tselluloos Hemitselluloos Pektiinid Ligniin Pektiini külg ahelad Tselluloosi EXP süntaas XTH β-glükosidaas Ksülosidaas Termotolerants Kasvu vähenemine Termotolerants Rakke kaitsev mehhanism Tabelis: ! - Suureneb orgaanilise ühendi molekulide sisaldus rakus. " - Väheneb molekulide sisaldus rakus.
pumbad) ensüümid aktiveeruvad – sekundaarne lüsosoom. Haaravad endasse: Võõrstruktuure (viirused, bakterid) ja sisse toodud osiseid (fagotsütoosi ja pinotsütoosi põiekesed, tahked aineosakesed) = fagolüsosoomid ehk heterolüsosoomid. Raku enda lammutamisele määratud komponente = autolüsosoomid. Järele jäävad jääkkehad. ! 5. Kus, kuidas ja kui palju tehakse ATP-d? ! Pikk variant I: Üldiselt sünteesib ATPd ensüüm ATP süntaas, mis asub mitokondri sisemembraanis (läbib seda) ja katalüüsib reaktsiooni ADP + Pi -> ATP. Energia antud reaktsiooni läbiviimiseks tuleb prootonite ehk vesinikioonide gradiendist. Prootonite gradient tekitatakse redoksreaktsioonides, kus osalevad hapnik, redutseeritud NAD (see tähendab NADH), koensüüm Q10 derivaadid jne. Summaarselt kutsutakse seda redoksreaktsioonide jada hingamisahelaks. Hingamisahela kompleksid: . I NADH dehüdrogenaas . II suktsinaadi dehüdrogenaas
TKT tähtsus Energia kasutamine: (1 ATP, 3 NADH, 1 FADH2)= 12 ATP TKT talitluse ja häirete meditsiinilised aspektid TKT funktsioneerimishäired organismile ohtlikud ja kriitilised väga intensiivse aeroobse metabolismi kudedes. TKT vajadus Tiamiin, riboflaviin, pantoteenhape, nikotiinhape, lipoehape Regulatoorsed ensüümid TKT talitluses ATP/ADP/AMP, GTP/GDP/GMP, NADPH/NADH/NAD, sukstinüül-CoA, Ca2+, atsüül-CoA, hormoonid TKT kontrollpunktid Tsitraadi süntaas- ATP, NADH, sukstinüül-CoA ja atsüül-CoA allosteeriline inhibeerimine Isotsitraadi DH- ATP ja NADH allosteeriline inhibeerimine. (Mg ja Ca defitsiit-> ensüümi töö) AKGDH-(3 ensüümi+ 5 vitamiini)- ATP, NADH, GTP ja sukstinüül-CoA allosteeriline inhibeerimine TKT respiratoorne kontroll NAD ja FAD pidev varustamine hingamisahela poolt Oksüdatiivse fosforüülimise sõltuvus ADP/ATP suhest TKT anaboolne (biosünteetiline) tähtsus Tsitraat- rasvhapete ja steroolide süntees
Glükolüüsi ja glükogenolüüsi energeetiline efektiivsus. Glükolüüsi kaks peamist etappi. Glükolüüsi vaheühendite fosforüülituse tähtsus. Glükolüüsi tulemusena tekkinud püruvaadi edasine metaboolne saatus sôltuvalt raku hapnikuga varustatusest. Glükolüüsi raja vôtmeensüümid - heksokinaas, fosforülaas, fosfofruktokinaas, püruvaadi kinaas. Nende aktiivsust môjutavad faktorid - hormoonid, Mg2+. Laktaadi dehüdrogenaasi funktsioon. Glükogeeni süntees. Glükogeeni süntaas - glükogeeni sünteesi vôtmeensüüm, glükogeeni "juuretise" olemasolu tähtsus rakus. Glükoneogeneesi môiste. Glütserool, aminohapped, laktaat kui glükoneogeneesi peamised substraadid. 1 9. Süsivesikute aeroobne oksüdatsioon. Atsetüül-CoA olemus ja teke püruvaadist (laktaadist), tsitraadi (Krebsi) tsükkel, elektronide transport hingamisahela ensüümide vahendusel
GTPlt. Millise üldnimetusega ensüümid vastavaid reaktsioone katalüüsivad, kui on teada, et reaktsiooni tulemusel toimub fosforüüli ülekanne? Kinaasid (Transferaasid) 9. DNA-sse kuuluvad desoksüribonukleotiidid sünteesitakse ribonukleotiidi reduktaasi toimel vastavatest ribonukleotiididest. Milline koensüüm varustab sünteesi katalüüsivaid ensüüme elektronidega? NADPH 10. DNA sünteesiks vajalike tümiin-nukleotiidide sünteesi eest vastutab ensüüm tümidülaadi süntaas. Mis on selle ensüümi substraadiks? dUMP (desoksüuridiinmonofosfaat) 11. Katabolismi lõpp-produktina tekib primaatide, lindude ja mitmete teiste organismide rakkudes puriinidest kusihape, pürimidiinidest aga -alaniinina, mis organismist väljutatakse. O H H N N O + O H3N C C C O N N H2 H2 O H
NT: fotosüntees, valkude süntees, DNA süntees NT: glükoosi lagundamine (rakuhingamine) Ülekaalus: lapsel, rasedal, sportlasel Ülekaalus: haigel, näljutajal, vanainimesel ATP ehk AdenosiinTriFosfaat Universaalne energia talletaja ja ülekandija Osaleb kõigi rakkude ainevahetuses (metabolismis) Moodustub glükolüüsi, käärimise ja hingamise käigus Toodetakse mitokondrite membraanis paikneva ensüümi, ATP-süntaas, abil RAKUHINGAMINE ehk glükoosi lagundamine Dissimilatsiooni protsess, mis toimub loomades, seentes ja taimedes RAKUHINGAMISE 3 ETAPPI: 1. GLÜKOLÜÜS - toimub tüstoplasmavõrgustikul 2. TSITRAADITSÜKKEL - toimub mitokondri sisemuses 3. HINGAMISAHELA REAKTSIOONID - toimub mitokondri harjakeste membraanil 1. ETAPP - GLÜKOLÜÜS (tsütoplasmavõrgustik) Glükoos lõhustatakse 2 püroviinamarihappe molekuli C6H12O6 2 C3H4O3 + 4 H
kromosoomide arvu püsivus tütarrakkudes. Interfaasiks nimetatakse kahe mitoosi vahele jäävat raku eluperioodi. Rakutsükliks nimetatakse raku eluringi ühe mitoosi lõpust läbi interfaasi järgmise mitoosi lõpuni. Mitoosi ja interfaasi kestus sõltub rakutüübist, vastava koe füsioloogilisest aktiivsusest ja keskkonnatingimustest. Raku ainevahetuse põhiprotsessid (organellide arvu suurenemine, ATP ja teiste makroerigiliste ühendite süntaas, tsentrioolide ja DNA kahekordistumine) toimuvad interfaasis, raku mõõtmed suurenevad. Interfaasi lõpus toimuva DNA kahekordistumise tagajärjel on kromosoomid rakujagunemise alguseks kahekromatiidilised (nende koostisse kuulub kaks identset DNA molekuli). Mitoosi alguseks on ühe kromosoomi kromatiidid omavahel ühendatud tsentromeeri abil, mis jagab iga kromatiidi kaheks osaks (kromosoomi õlgadeks). Rakujagunemisel lahknevad kõigi kromosoomide kromatiidid ja
7. Ammooniumi lülitumine orgaanilistesse molekulidesse toimub põhiliselt kolme tüüpi reaktsioonide abil. Otsustage, milline ensüüm millist reaktsiooni katalüüsib. Reaktsioon Ensüüm A. -ketoglutaraat + NH4++ NADPH glutamaat + NADP+ I. Karbamoüülfosfaadi süntetaas B. HCO3- + NH4+ +2 ATP H2N-COO-PO32- II. Glutamiini süntetaas C. Glutamaat + NH4++ATP Glutamiin + ADP+Pi III. Glutamaadi süntaas 8. Ammooniumi lülitumisel orgaanilistesse molekulidesse on võtmepositsioonis kaks aminohapet. Nimetage, millised ja kirjutage nende struktuurivalemid. Glutamiin (C5H10N2O3) ja glutamaat (C5H9NO4). 9. N tasakaalu tagamiseks organismis peab inimene toiduga saama ööpäevas ~60-70g valke. Mitut proteogeenset aminohapet ei suuda inimese organism piisavas koguses sünteesida ja miks? 10, sest imetajatel puudub vastav ensüüm sünteesimiseks. Kas võite nimetada need aminohapped
Transgeenide vaigistamine Taime viidud transgeenid on sageli siRNA poolt vaigistatud Vaigistamise lülitab sisse: Väga kõrge ekspressiooni tase dsRNA, mis tekib transgeenilt Hälvik RNA-d, mida transgeen kodeerib Transgeene vaigistatakse post-transkriptsionaalselt ja ka transkriptsiooni ajal. 1980-tel lõid teadlased meetodi, et viia geene taime genoomi, kasutades taime patogeeni Agrobacterium tumefaciens (bakter). Sisestatud geene kutsutakse transgeenideks. Kalkooni süntaas on ensüüm, mis on oluline antotsüaanide biosünteetilise raja alguses. Fenomen, kus mõlemad geenid (transgeen ja oma geen) on vaigistatud, nimetatakse `koos- allasurumiseks. Translatsioon- RNA ahelast saadakse geneetilise koodi kaudu valgu aminohappeline järjestus. Toimub ribosoomidel. mRNA- RNA molekul, kus toimub translatsioon geneetiline kood- 1 koodon (3 nukleotiidi) kodeerib 1 hapet. Olemas on 20 aminohapet.
üksiksidemega struktuuriks. Tekkiva suhkru koosseisu jääb aga neljast elektronist kaks. Tülakoidi membraansüsteem moodustab sisemise ruumiosa, luumeni, mis on stroomast membraaniga eraldatud. Luumeni ja strooma vahel tekib H kontsentratsiooni elektrokeemiline gradient, mis energiseerib ATP sünteesi. Tülakoidi membraanides asuvad valgust neelavad fotosüsteemid I ja II ja nendevahelised elektronikandjad plastokinoon, tsütokroom b6f, plastotsüaniin, samuti ka NADP reduktaas ja ATP süntaas. CO2 sidumise ja taandamise reaktsioonid aga toimuvad tülakoidide vahel asuvas stroomas. Valgusreaktsioonideks nimetatakse valguse neeldumist ja elektronide ülekandega seotud reaktsioone. Valguse abil toimub tegelt ainult elektroni ergastumine pigmendil. NADPH on universaalne elektronikandja. H-ioonidel on eriline roll ATP sünteesis, kus need, liikudes läbi membraani kontsentratsioonigradiendi ja elektripotentsiaalide vahe mõjul, annavad ATP sünteesiks vajaliku energia
- Rasvhapete vajadus kaetakse kahel viisil o Suurel määral normaalse söömisega o Endogeense sünteesiga (ei sünteesi vaid LA ja ALA) - Toimub maksas ja lakteerivas piimanäärmes - Tsükliline protsess - 2-süsinikuline fragment sisestatakse ahelasse iga tsükli ajal - De novo süntees pole B-OX pöördprotsess kuna o Toimub tsütoplasmas o Protsessi viib läbi ekstramitokondriaalne süntaas o Lähteühend on atsetüül-CoA, ja süntees toimub läbi võtmeühendi MALONÜÜL-CoA o Vajab redutseerivat energiat (NADPH) ning ATP, bikarbonaati, biotiini, Mn o On intensiivne süsivesikuterikka toidu tarbimisjärgselt, st teda soodustab INS o Sünteesi aktiveerib tsitraat (kuna see on TKT esiprodukt palju aktiveerib sünteesi) ja pärsib atsetüül-CoA
moodustades mikrofibrille Tselluloos on D-glükoosi jääkidest koosnev polümeer. Monomeerid on omavahel seotud -1,4- glükosiidsideme kaudu. Tselluloosiahela minimaalseks kordusühikuks on tsellobioosi jääk. Mitteredutseerivas otsas on glükoos alati tsüklises vormis, redutseerivas otsas esineb tasakaal avatud aldehüüdse ja tsüklilise poolatsetaalse vormi vahel. Looduses vastutab tselluloosi sünteesi eest plasmamembraanis paiknev tselluloosi süntaas. Aktiveeritud monomeerina kasutatakse UDP-glükoosi, mis lülitatakse kasvava tselluloosiahela mitteredutseerivasse otsa ning glükosiidse sideme tekkega vabaneb UDP. Sõltuvalt tselluloosi päritolust võib glükoosijääkide arv ahelas ehk polümerisatsiooni aste (DP) varieeruda vahemikus 2000-20 000. Taimede plasmamembraanis paiknevad tselluloosi süntaasid rosettidena, mis tagab kasvavate tselluloosiahelate õige orientatsiooni teineteise
millsit reaktsiooni loetakse tsükli esimeseks? C) millise metaboliidi järgi on tsükkel nime saanud? D)millised energiakandjad (mitu?) tsükli iga ring produtseerib. 18.Mõtestage lahti mõiste oksüdatiivne fosforüülumine. Kirjeldage, millistes rakustruktuurides (organell, kompartment) oksüdatiivse fosforüülumise protsess toimub ja mida selles produteeritakse? 19.Millist rolli täidab oksüdatiivse fosforüülumise protsessis ensüüm ATS süntaas? Iseloomustage a) kus see ensüüm paikeb, b) milline on tema ehitus, c) millise energia arvel ensümaatiline protsess käivitub. 20.Arvutage, milline on energiasaagis ATP-na 1 glükosimolekuli aeroobsel oksüdatsioonil, kui ka tsütosoolis tekkiv NADH siseneb mitokondrisse ja reoksüdeerub oksüdatiivse fosforüleerumise protsessis. Biokeemia test II variant 2 1. Kirjutage mingi aldopentoosi molekul a) lineaarses vormis, b)tsüklilises furanoosi vormis
betaiin 2) Etanoolamiini 2) Letsitiini komponent metüülimine 2) Atsetüülkoliini süntees 3) Koliini oksüdeerimine 3) Metüülrühmade doonor Kreatiin, fosfokreatiin, Skeletilihastes ja (CK) isoensüümne kreatiniin ning kreatiniini südamelihastes markerensüüm süntaas Histamiin Histidiini dekarboksüülimine Lokaalne signaalmolekul nuumrakkudes ja basofiilides Maohappe sekretsioon Allergiline reaktsioon Põletik KNS neurotransmitter
Refsum haigus- pärilik mittekoordineeritud polüneuriit, mida põhjustab fütanaadi kuhjumine maksas ja veres A-OX-raja defektsuse tõttu. Sümptomiteks on neuroloogilised häired, võrkkestapõletik, naha ja luude anormaalsus. Rasvhapete Oomega-oksüdatsioon Toimub ER-s cytP450 ja NADPH ning hapniku toimel. Rasvhapete de novo süntees Palmitaadi süntees, mis toimub maksas ja lakteerivas piimanäärmes. Süntees toimub tsütoplasmas, sünteesi viib läbi multiensüümsüsteem rasvhappe süntaas Palmitaadi sünteesi võtmesündmus on atsetüül-CoA karboksüülimine malonüül-CoA-ks biotiini osalusel. Süntees vajab veel NADPH, bikarbonaati, mangaani, pantoteenhapet. Sünteesi aktiveerib tsitraat ja pärsib atsetüül-CoA ning soodustab kõrge INS/glükagooni tase, sest INS on antilipolüütilise toimega. Üldised regulatsioonifaktorid: ATP, vitamiinide tase, nälgimine (stimuleerib rasvhapete oksüdatsiooni ensüümide stressi) Rasvhapete oksüdatsiooni regulatsioon:
eluks vajalikku energiat ja on mitmete oluliste anabolismireaktsioonide eelduseks. 8. Selgitage: a) milliseid protsesse tähistab termin oksüdatiivne fosforüleerimine üks olulisimaid biokeemilisi protsesse, mis võimaldab enamikul biosfääri organismidel biosünteesida orgaanilistest ainetest vabanenud vaba energia arvelt ADP-st ja ortofosforhappest ATP. b) kus need protsessid aset leiavad mitokondri membraanis c) mis on ATP süntaas ja milline on tema roll ATP-d sünteesivat ensüümi nimetatakse ATP süntaasiks ja see koosneb membraanis olevast kompleksist Fo ning membraanist väljaulatuvast kompleksist F1. Kasutab vesinikioonide gradienti, et teha ATP'd. d) milline on prootonigradiendi roll protsessis kuidas formeerub, milleks ta on vajalik. Mitokondrite ja kloroplastide organiseeritus ning nende ATP tekitamise mehanism on sarnane.
Lüaaside süstemaatilised nimetused: Näiteks: tsitraadi hüdro-lüaas, kus tsitraat on substraat ning hüdro (H2O) on eralduv rühm. Lüaas Reaktsioon dehüdrataas vee elimineerimine hüdrataasid vee liitmine dekarboksülaas CO2 elimineerimine süntaas kondensatsioonireaktsioonid, mis ei vaja ATP, GTP jt energiat EC 5. Isomeraasid ja nende süstemaatilised nimetused: Isomeraasid katalüüsivad muutusi ühe molekuli piirides ehk isomerisatsiooni. Isomeraaside reaktsioonides on üks substraat ja üks produkt. Isomeraasid jagunevad: ratsemaasideks, epimeraasideks, mutaasideks ning cis-trans-isomeraasideks.
pärmirakud ja tekib sültjas mass, mis ujub vedeliku pinnal. · Seal on hapnikku ja saab toimuda pärmide poolt suhkru kääritamisel moodustunud etanooli oksüdeerimine hapnikuga äädikhappeks. · Acetobacter'i rakuväline tselluloos · Enim on rakuvälise tselluloosi sünteesi uuritud A. xylinumil. · Tselluloosi moodustub enam seisukolvis kasvatamisel, kui loksutil. · Tselluloosi süntaas on membraani välispinnal paiknev ensüüm. Läbi pooride kestas eritatakse tselluloosikiud, mis moodustavad väljaspool rakku lindi. · Kasutatakse tsellulaaside uurimisel puhta tselluloosse substraadina. Selline tselluloos ei sisalda hemitselluloosi jm saastaineid. · Acetobacter diazotrophicus on suhkrurootaime juhtkudedes elav N2 siduv bakter. Vajab ja talub väga kõrget suhkrusisaldust. Toodab sahharoosist levaansukraasi abil
Glükogenolüüs on maksa ja lihaste glükogeeni mobiliseerimine Kesksed momendid: Ahelate lühendamine Hargnemispunktide elimineerimine Glükoos-1-P konversioon glükoos-6-P-ks On vajalik: Veresuhkru taseme hoidmiseks Glükogenees on glükogeeni biosüntees glükoosist tsütoplasmas, mis algab glükoosi aktiveerimisega ATP energia arvel Glc-6-P tekkega. Glükogeeni süntaas liidab glükoosijääke sidemega α (1,4) tsütoplasmas asuvale „juuretisele“, millega pikendatakse ahelaid ning sidemete α (1,6) abil luuakse hargnemispunktid. Glükoneogenees on glükoosi biosüntees mittesahhariidsetest eelainetest (laktaat, püruvaat, glütserool, ossa aminohappeid). Peaaegu 90% toimub maksas ja u 10% neerukoores. Nt glükoneogenees on vältimatu ära hoidmaks hüpoglükeemiat. 36. Käärimised, sahhariidide ainevahtuse eripärad mäletsejatel
Glükogenolüüs glükogeeni lagundamine püroviinamarihappeks Kesksed momendid: Ahelate lühendamine Hargnemispunktide elimineerimine Glükoos-1-P konversioon glükoos-6-P-ks On vajalik: Veresuhkru taseme hoidmiseks, maksas talletub varu glükogeen, kust organism saab varu Glükogenees on glükogeeni biosüntees glükoosist tsütoplasmas, mis algab glükoosi aktiveerimisega ATP energia arvel Glc-6-P tekkega. Glükogeeni süntaas liidab glükoosijääke sidemega (1,4) tsütoplasmas asuvale ,,juuretisele", millega pikendatakse ahelaid ning sidemete (1,6) abil luuakse hargnemispunktid. 9 Glükoneogenees on glükoosi biosüntees mittesahhariidsetest eelainetest (laktaat, püruvaat, glütserool, ossa aminohappeid)
elektrontranspordi ahel - koosneb elektrone liitvatest (aktseptor) ja loovutavatest (doonor) molekulidest e. kandjavalkudest(plastokinoon, plastotsüaniin, ferredoksiin). Tsütokroom b6f kompleks plastokinoon annab plastotsüaniinile elektroni, prootonid pumbatakse läbi selle kompleksi elektroni ülekandel vabanenud energia arvel luumenisse Ensüümkompleks NADPH2 sünteesiks ferredoksiin NADP + reduktaas ATP süntaas - ensüümkompleks ATP sünteesiks 8. Peaksite membraani valgusreaktsiooni osasid kujutaval joonisel ära tundma FS I ja FS II, vett lagundava kompleksi, elektrontranspordiahelaga seotud kandja valgud, ATPaasi, NADPH reduktaasi PS = FS Oxygen-evolving complex = vett lagundav kompleks Elektrontranspordiahelaga seotud kandja valgud on plastokinoon(plastoquinone), plastotsüaniin(plastocyanin) ja ferredoksiin(ferredoxin) ATPaas = ATP synthase FNR - ferredoksiin NADP+ reduktaas
organismides on nikkel tähtis katalüsaator erinevatele bioloogilistele reaktsioonidele tänu oma mitmekülgse kordinatsiooniga geomeetriale, mis muudab metallid üleüldse olulisteks osadeks igas elussüsteemis. Nikkel on vajalik element mitmetes essentsiaalsetes metalli-ensüümide aktiivtsentrites bakteritele ja madalamatele-eukarüootidele. Avastatud on 8 mikroobset niklit sisaldavat ensüümi: ureaas, hüdrogenaas, CO dehüdrogenaas, atsetüül-CoA süntaas, metüül- CoM reduktaas, Ni-superoksiidi dimutaas, happereduktsiooni dioksügenaas ja glüoksalaas I, lisaks on neid veel avastamisel. Taimedel on kindlaks tehtud niklit vajavaks ureaas, mõned taimed on arenenud ka hüperakumuleerima nikli ioone, luues komplekseid süsteeme metalli mürgisuse vähendamiseks ja homöostaasiks, mis on uurimise all, et kunagi kasutada reostuste puhastamiseks.
05 ja -1,3 V Liikumine toimub alati väiksemat suuremale: -1,3, -0,05 ja siis +0,82 Mida negatiivsem redokspotentsiaal, seda parem elektronide doonor 3. Nimetage mõni fotosünteesi valgusreaktsioonides osalev ühend/kompleks mis paikneb kloroplastide luumenis Tülakoidi membraanides asuvad valgust neelavad fotosüsteemid I ja II (PSI ja PSII) ja nendevahelised elektronikandjad plastokinoon, tsütokroom b6f, plastotsüaniin, samuti ka NADP reduktaas ja ATP süntaas 4. Taimed kasutavad fotosünteesis millist valgust? (Kirjutage lainepikkuste vahemik) Fotosünteesil kasutatakse nähtavat valgust 400 700 nm punases (650-680 nm) ja sinises valguses (430-470 nm, nn Soret riba). Sinise valgusega ergastumisel kukub elektron kiiresti punase riba energiale ja ainult seda saab kasutada fotosünteesiks. 5. Loetlege viis sinist valgust absorbeerivat pigmentide rühma taimedes
Toimub kahesüsinikulise atsetüüljäägi ülekanne neljasüsinikulisele ühendile (oksaloatsetaat) ja tekib kuue süsinikuline tsitraat. See dekarboksüülub, eraldub CO 2, mille käigus tekivad viie ja neljasüsinikulised metaboliidid, millest viimane konvereerub oksaloatsetaadiks, mis saab liituda järgmisesse tsüklisse kondenseerudes atsetüül-CoA-ga. Tsitraaditsükli reaktsioonid: I Atsetüül-CoA atsetüülgrupi ülekandega oksaloatsetaadile tekib ensüümi tsitraadi süntaas vahendusel tsitraat. Reaktsioon on pöördumatu. II Tsitraat isomeriseerub isotsitraadiks ensüümi akonitaas vahendusel. Reaktsioon on pöörduv. III Isotsitraadi osküdatiivse dekarboksüülimise tulemusel sünteesitakse -ketoglutaraat, eraldub CO 2 (väljub esimene C aatom) ja toodetakse NADH ensüümi isotsitraadi dehüdrogenaasi (tsitraaditsükli võtmeensüüm) vahendusel. Reaktsioon on pöördumatu.
Neid indiviide nimetatakse albiinodeks. Naha vähene pigmentatsioon on soodus nahavähi tekkeks, islmade pigmentatsioonihäired soodustavad valguspelguse (fotofoobia) teket. 19.Homotsüsteiin ja hüperhomotsüsteineemia Hcy on AH, mis tekib metioniinist. Kuna Met on asendamatu AH, konverteerib inimkeha umbes pool Hcy tagasi Met-ks. Hcy liigtaseme vältimiseks, ei tohi kudedes tekkida foolhappe, vitamiinide B12 ja B6 defitsiiti. Neid vajavad Met süntaas ja Hcy metabolism Cys-ks. Ravimid, mis mõjutavad vitamiin b& või foolhappe taset, võivad põhjustada Hcy taseme tõusu veres. Hüperhomotsüsteineemia on KVH ja neurodegeneratsiooni riskifaktor. Ilmneb tihti neerupuudulikkuse, suitsetamise, menopausi, vananemise puhul. Levinum on 5,10- metüleenTHF reduktaasi teatud defitsiit. Võib esineda ka teiste ensüümide defitsiiti. Tsüstatiooni süntaasi defektsus häirib Hcy konversiooni tsüstatiooniks, Hcy kuhjub ning
sellepärast tekib elektrokeemiline gradient membraani sise- ja väliskülje vahel. See avaldub selles, et välisküljel on suurem positiivne laeng (gradiendi elektriline komponent) ja suurem vesinikuioonide kontsentratsioon (gradiendi keemiline komponent). Mitokondri sisemembraanis on aga valke, mis suudavad neid prootoneid jälle gradiendi surve toimel sisse tuua ja selle prootonite voo energia arvelt toimubki ATP süntaasi abil ADP fosforüülimine. ATP süntaas koosneb kahest subühikust, üks neist (F0) paikneb sisemembraanis ja moodustab poori, mille kaudu prootonid saavad maatriksisse sisenega ning F1, mis katalüüsib ATP sünteesi. Praktiliselt saab hingamisahelat jagada neljaks kompleksiks. I kompleks võtab NADH + H+ elektronid ja annab need koensüüm Q-le. See on NADH dehüdrogenaas. Prootonid lasevad alga sisemembraani välisküljele ja coQ redutseerub coQH2. II kompleks (flavoproteiini hüdrogenaas) võtab
c) aeglustab vananemist (toitaineid liiguvad pritsitud piirkonda) d) soodustavad varte pikenemiskasvu ja rakkude venivuskasvu. e) vähendavad apikaalset domineerimist. Nimetage etüleeni biosünteesi raja lähteühendid ja peamised vaheproduktid Etüleeni biosünteesirada. Etüleeni sünteesitakse kõikides vananevates taimeosades, eriti palju aga valmivates viljades. Seda sünteesitakse metioniinist, millega peab seostuma ATP. Tekib S-adenosüülmetioniin. Oluliseks ensüümiks on ACC süntaas, mis saadud produktist sünteesib ACC, mis omakorda oksüdaaside toimel muudetakse etüleeniks. Pärssivaks kohaks võib olla ACC süntaasi geeniperekondade mitte ekspresseerumine. Nimetage etüleeni biosünteesi raja kaks peamist regulatsioonipunkti Milliste geenide ekspresseerumine etüleeni toimel aktiveerub ACC süntaasi, ACC oksüdaasi Loetlege etüleeni füsioloogilisi toimeid Etüleen kiirendab viljade valmimist ja lehtede vananemist ning nende maha langemist. Taime
Kõigepealt aktiivsaidi lüsiin deprotoneerub ja siis seostub CO2-ga. Karbamüülitud Rubisco seob magneesiumiooni ja muutub aktiivseks. Kuidas valgus põhjustab GAP dehüdrogenaasi aktiveerumist Calvini tsüklis PSI aktseptorpoolel ferredoksiin taandub, see taandab tioredoksiini ja viimane ensüümide tioolrühmad (S-S), muutes need sulfhüdrüül(SH) rühmadeks. S-S sidemete katkemine avab reaktsioonisaidi. Toimub siis eelnev valguse toimel ning nii atkiveruvad lisaks GAP DH-le ka ATP süntaas, Rubisco aktivaas,Fruktoosbisfosfataas,Ru5P ja Seduheptuloosbisfosfataas. Kuidas on defineeritud CO2 kompensatsioonipunkt CO2 kompensatsioonipunkt : selline CO2 konsentratsioon, mille juures CO2 assimilatsioonikiirus fotosünteesil on võrdne CO2 moodustumise kiirusega fotorespiratsioonil ja hingamisel. CO2 kompensatsioonipunkt on hästi madal C4 taimedel, mis näitab, et fotorespiratsioonil vabanev CO2 reassimileeritakse mesofüllirakkude poolt, enne kui see jõuab difundeerida lehest välja.
Glükogenolüüs on glükogeeni lõhustumine glükoosiks. Glükogenolüüs lühendab korraga glükogeeni paljusid ahelaid vabastamaks kiiresti rohkelt glükoosijääke (lõhustatakse 1,4 sidemed), hargnemispunktide elimineerimine, mil lõhustatakse 1,6 sidemed ning glükoos-1-P konversioon glükoos-6-P-ks. Glükogenees on glükogeeni biosüntees glükoosist tsütoplasmas, mis algab glükoosi aktiveerimisega ATP energia arvel Glc-6-P tekkega. Glükogeeni süntaas liidab glükoosijääke sidemega (1,4) tsütoplasmas asuvale ,,juuretisele", millega pikendatakse ahelaid ning sidemete (1,6) abil luuakse hargnemispunktid. Glükoneogenees on glükoosi biosüntees mittesahhariidsetest eelainetest (laktaat, püruvaat, glütserool, ossa aminohappeid). Peaaegu 90% toimub maksas ja u 10% neerukoores. Nt glükoneogenees on vältimatu ära hoidmaks hüpoglükeemiat. 32. Käärimised. Sahhariidide ainevahetuse eripära mäletsejalistel.
piisavalt. Kuigi suur hulk NADHd, mis tekib TCA tsükli reaktsioonide tulemusel, on põhimõtteliselt kasutatav taandavaks biosünteesiks, rakendatakse mitokondriaalse NADH keemiline potentsiaal kõige sagedamini energia saamise eesmärgil ehk ATP sünteesiks oksüdatiivse fosforüülimise protsessis. NADH oksüdeerimine ja ADP fosforüülimine ATPks on protsessid, milleks on vajalik mitokondri sisemembraaniga seotud elektronide ülekande ahel ja ATP süntaas. Nendes protsesside vahendavad mitokondri sisemembraani integraalsed valkkompleksid. NADH ja FADH2 loovutavad prootonid ja elektronid ära elektronide transportahelale. See ahel koosneb neljast valkkompleksist, mis koostöös liikuvate ehk mobiilsete vahendajatega kannavad elektronid üle hapnikule, redutseerides selle veeks. Samal ajal toimub prootonite pumpamine intermembraansesse ruumi, mille tulemusel tekib prootonite gradient mitokondri sisemembraanil
suhkur glükoos-1-fosfaat, mis on glükolüüsi substraadiks. Glükoos-1-fosfaat 3. UDP-glükoos (uridiindifosfaatglükoos) on glükoosijääkide doonor kasvavale glükogeeni polümeeri molekulile. Glükogeeni süntaasi reaktsioon moodustab (14) glükosiidsidemed. Esimene glükoos seotakse glükogeniini (valgu) türosiini OH rühmale. Glükogeeni süntaas kannab glükosüülühikud UDP-glükoosilt C-4 hüdroksüülrühmale glükogeeni mitteredutseerivas otsas. Amülo-(1,41,6)-trans-glükosülaas katalüüsib lineaarse ahela lõikamist ja liitmist C-6 OH-rühmale, millega moodustab uue haru. UDP-glükoos Glükogeeni fosforülaasi ja glükogeeni süntaasi regulatsioon toimub ka kovalentse modifitseerimise teel, mis on hormonaalse kontrolli all. 4
bakterist Tapab: 7 Liblikalised (lepidoptera) (liblikad, ööliblikad) Kahetiivalised (diptera) (kärbsed, sääsed) Mardikalised (coleoptera) (mardikad) Põllumajanduslik tähtsus: varreleedik, maisi juure uss ja puuvilla kuprauss Toksiini toime: seob putuka seedeelundkonna retseptoreid Tekitab poore plasma membraani Glyphosate (RoundupTM; Monsanto) blokeerib aminohapete sünteesi. Glüfosaat blokeerib ära ühe ensüümi (EPSP süntaas). Mõned EPSP süntaasid on resistentsed glüfosaadile Roundup-readyTM puuvillal ja sojal on ekstra EPSP ensüüm (loeng 7) Epigeneetika - Igasugune potentsiaalselt stabiilne ja pärandatav muutus geeni ekspressioonis, mis tekib ilma DNA järjestuse muutuseta. Tähendab informatsiooni DNA-s, mitte järjestuses, vaid DNA või kromatiini struktuuri muutusi. Praktikas epigeneetika kirjeldab fenomeni, kus geneetiliselt identsed rakud või organismid ekspresseerivad oma
mevalonaadist, millest tekib isopentenüülpürofosfaat (IPP). Praegu on teada, et selles rajas sünteesitakse seskviterpeenid (näit ABA), triterpeenid (steroidsed ühendid) ja polüterpeenid (kautsuk, gutta). Kloroplastides moodustub IPP GAP/püruvaatses rajas, mille olemasolu avastati suhteliselt hiljuti (1993). Püruvaat reageerib TPP-ga, moodustades kahesüsinikulise fragmendi, mis kondenseerub GAP-iga deoksü- ksüluloos-5-fosfaadiks (DOXP süntaas=transketolaas, dxs), mis redutseerub (+dehüdrateeritakse+fosforüülitakse) IPP-ks. Selles rajas sünteesitakse peamiselt mono-, di-(giberelliinid) ja tetraterpeenid (karotinoidid, fütool, plastokinoonid). Seega giberelliinide biosüntees toimub kolmes etapis. 1) proplastiidides C20 eellase ent-kaureeni süntees püruvaadist ja GAP-ist 2) ER-s ent-kaureeni oksüdeerumine ent-kaureenhappe tekkimisega Kahekümnenda
Fosfoproteiinid on näiteks piimavalk kaseiin, ka mitmed munades ja kalamarjas leiduvad valgud. Fosfoproteiinidele on iseloomulik suur asendamatute aminohapete sisaldus, seejuures inimesele sobivates proportsioonides, mis teeb neist kõrge toiteväärtusega valgud. Fosfoproteiinidena võib vaadelda ka paljusid ensüüme, mida fosforüülimise/defosforüülimise kaudu kas aktiveeritakse või inhibeeritakse. Niisugused ensüümid on näiteks glükogeeni süntaas ja glükogeeni fosforülaas, mis reguleerivad vastavalt glükogeeni sünteesi- ja degratsiooniprotsessi. Kromoproteiinide iseloomulikuks tunnuseks on see, et 13 Maris Kallus KKS 2010 prosteetiline rühm annab neile valkudele spetsiifilise värvuse. Kromoproteiinid on
Ja kasvatada sellest transgeenne taim. Bt-toksiini määrav geen on viidud taimesse. Võetakse cry geen bakterist, mis toodab putukspetsiifilist toksiini. See toksiin on putukatele kahjulik. Kuidas konstrueerida üht herbitsiididele tolerantset transgeenset taime? glüfosaat, glüfosinaat blokeerib aminohapete sünteesi. glüfosaat (herbitsiit) plokeerib ära ühe ensüümi mis vastutab aminohapete tootmise eest (EPSPS ensüüm ,EPSP süntaas). Et muuta taim herbitsiidi tolerantseks võiks talle siirdada, mõne teise organismi geene, mis kodeerivad sama ensüümi, aga vähe teisel kujul nii et herbitsiid sellega ei seondu. Võib ekspresseerida ka ensüümi, mis muudab herbitsiidi nii, et see ei seondu aminohapet sünteesiva ensüümiga. Mis on “Kuldne riis”? Üks varasemaid näiteid säärastest GMOdest on kuldne riis. Tavalise riisi sisse viiakse
b) Elektroni transpordisüsteem (ETS) (= elektroni ülekandeahel = hindamisahel) Elektronid, mida kannavad tsitraaditsüklis moodustunud taandatud koensüümid, läbivad valkude ja koensüümide ahelda (hingamisahela), et moodustada prootonigradient läbi mitokondri sisemembraani. c) Tsütokroomid Tsütokroom C on vesilahustuv elektronide kandja valk ja paikneb membraanidevahelises ruumis. Tsütokroomid on ühe-elektroni ülekande agendid. d) ATP süntaas Roteeruv molekulaarmootor, mille abil toimub ADP fosforüleerimine prootonigradiendi juuresolekul. F1-l on kolm omavahel interakteeruvat kuid eristatud aktiivtsentrit. Elektronide transpordist saadav energia moodustab prootonigradiendi, mis kutsub esile ATP süntaasi konformatsioonimuutused. Selle tulemuseks on substraadi (ADP) sidumine, ATP süntees ja produkti vabanemine. 2. Esitage mitokondri joonis (lõige) ja kirjeldage selle põhjal mitokondri ehitust.
2. Kui ensüüm katalüüsib rohekm kui ühte tüüpi reaktsiooni, tuleb nimetus koostada ainult ühest reaktsioonist. Sama kehtib ka alternatiivsete substraatide kohta. Kui väiksema spetsiifilisusega ensüüm, siis antakse substraadi nimi üldisemalt alkoholi dehüdrogenaas (pole spetiifiline ühele kindlale alkoholile). 3. kui soovitakse rõhutada ensüümi olulisust mingis sünteesiprotsessis, siis võib triviaalnimetustes kasutada nimetust süntaas (v.a. ligaaside puhul seal on süntetaas). Tselluloosi süntaas (oluline), süntaasi tuleb kasutada mõistuse piirides. EX! Ensüüminimede tuletamine. Antakse reaktsioon ette struktuuridena, tuleb reaktsiooni katalüüsiva ensüümi nimi kirjutada ja ka põhiklass. Reaktsiooni keemia järgi tuleb ära tunda põhireaktsioon tüüp (üks kuuest). EC.1 Oksüdoreduktaasid Katalüüsivad oksüdeerumis-redutseerumis reaktsioone (elektronide ülekanne substraatide vahel).
Kodeerib kokku 37 geeni, millest 13 on valkudele, peamiselt elektrontrasportahela valgud. Kodeeritavad valgud: mtETA, ATP sünteesi valgud, translatsioonivalgud. 16,5 kbp e 15k-17k aluspaari. 100-10.000 mitokondrit, igas oma DNA, kerge (C-rikas, 9geeni) ja raske ahel (G-rikas, 28 geeni). kodeerib 37 geeni, 13 valku, 22 tRNA'd, 2 rRNA. Elektrontransportahela valgud: NADH dehüdrogenaasid, koensüüm q - tsütokroom c reduktaas/tsütokroom b, tsütokroom c oksüdaas, ATP süntaas. 96. Kuidas mitokondrid liiguvad eellasrakkudelt tütarrakkudele? Mitokondrid poolduvad rakkudega sarnaselt, mõlemale tütarrakule umbes sama palju mitokondreid 97. Mis on tsütoplasmaatiline pärilikkus? Sugulisel paljunemisel pärinevad raku mitokondrid emasliinist, kuna viljastatud munarakul on vaid munarakult saadud tsütoplasma ja seega ainult munarakult saadud mitokondrid. Tsütoplasmaatilise pärilikkuse tõttu levivad mitmed mitokondriaalsete
• Toimub kloroplastides ja vaja on pigmenti nimega klorofüll c) Valgusstaadium ja pimestaadium e. Calvini tsükkel. Mis toimub? Kus toimub? • Valgusstaadiumis alguses pigmendi molekul ergastub ja sealt eraldub 1 elektron → elektron võetakse tagasi vee molekulilt → H₂O laguneb vesinikioonideks ja hapnikuks → vesinikioonide kontsentratsioon tülakoidi membraani sees ja väljas on erinev → vesinikioonid liiguvad läbi membraani → ATP-süntaas läheb käima ADP+fosfaatrühm → saadakse ATP • Toimub tülakoidi membraanis • Pimestaadiumis liigub klorofüllist eraldunud 1 elektron molekulile nimega NADP⁺ ja see redusteerub ja seob endaga vesinikioone → muutub NADPHks → ATP toimel hakatakse liitma CO₂ molekule + NADPH → saame C₆H₁₂O₆ d) Mis mõjutab fotosünteesi intensiivsust ja kas seda intensiivsust saab lõpmatuseni tõsta? • Mõjutab valgus ja temperatuur
keskkonda. Tavaliselt tekitavad bakterid PMF-i prootonite transportimisega tsütoplasmast välja. Kuna prootonid on positiivse laenguga, siis tekib kaks gradienti korraga keemiline gradient (ainet on ühel pool membraani rohkem kui teisel pool) ning elektriline gradient (membraani välisküljel on positiivne laeng, tsütoplasmapoolsel küljel negatiivne laeng). Selle tulemusena on soodustatud prootonite tagasi liikumine tsütoplasmasse, ehk ATP-süntaas kasutab ära prootonite passiivset liikumist mööda elektrilist ja keemilist gradienti. PMF-ga on tihedalt seotud kemiosmoos. Kemiosmoosi põhimõte on sarnane osmoosile, kus kahel pool membraani olev vesi liigub membraani sellele poolele, 34 kus on vähem vett. Kemiosmoosi korral liiguvad katioonid. Tavaliselt on selleks katiooniks metallid, millest omakorda levinuim on kaalium. Bakteris on kaalium
· Antimütsiin A takistab elektroni ülekannet tsütokroom b-lt tsütokroom c-le. Elektrontransportahela komponendid: 1. Flavoproteiinid, kannavad vesinikku 2. FeS valgud (mitteheemne raud) kannavad elektrone 3. Kinoonid, kannavad vesinikku 4. Tsütokroomid, sisaldavad heemi, kannavad elektrone Elektronide ja vesiniku ülekandjad vahelduvad membraanis. Moodustades linge ja see võimaldab prootonite suunatud liikumist läbi membraani välja. ATP süntaas on mikromootor, mis pöörleb, kui prootonid transporditakse läbi tema tsütoplasmasse. See pöörlemine toob kaasa ATP sünteesi. Bakteritel on ka selliseid ATP süntaase mis kasutavad põhiliselt Na + ioonide gradienti, kuid võivad ümber lülituda ka H+ või Li+ ioonide gradiendile. Ainete transport rakku. Difusioon, vahendatud difusioon, aktiivtransport oska neid lühidalt iseloomustada. Sümport, uniport, antiport. Poriinid välismembraanis, nende osa ainete liikumises rakku.
transportijad mitokondri maatriksist membraanidevahelisse ruumi. Elektroni seostumine/eraldumine muudab valguliste komplekside konformatsiooni rohkem/vähem afiinseks prootonite suhtes. Iga elektronpaari liikumisel hapnikule transporditakse 4 prootonit membraanide vahelisse ruumi. Milline valguline kompleks mitokondrites kasutab prootonite kontsentratsiooni erinevuses peituvat energiat ATP sünteesiks? ATP süntaas (F-tüüpi H-ATPaas). Koosneb kahest subühikust Fo ja F1. Kus see kompleks mitokondris paikneb? Millises mitokondri piirkonnas ATP sünteesitakse? Paikneb mitokondri sisemembraanis. Fo subühik paikneb membraanis, F1 membraani maatriksi poolsel küljel. Kuidas ATP transporditakse mitokondrist tsütosooli? ATP transporditakse ATP/ADP translokaasi abil välja Mitu prootonit kannab suktsinaat-CoQ reduktaas membraani ühelt küljelt teisele? Mitte ühtegi.
transportijad mitokondri maatriksist membraanidevahelisse ruumi. Elektroni seostumine/eraldumine muudab valguliste komplekside konformatsiooni rohkem/vähem afiinseks prootonite suhtes. Iga elektronpaari liikumisel hapnikule transporditakse 4 prootonit membraanide vahelisse ruumi. 9. Milline valguline kompleks mitokondrites kasutab prootonite kontsentratsiooni erinevuses peituvat energiat ATP sünteesiks? ATP süntaas (F-tüüpi H-ATPaas). Koosneb kahest subühikust Fo ja F1. 10. Kus see kompleks mitokondris paikneb? Millises mitokondri piirkonnas ATP sünteesitakse? Paikneb mitokondri sisemembraanis. Fo subühik paikneb membraanis, F1 membraani maatriksi poolsel küljel. 11. Kuidas ATP transporditakse mitokondrist tsütosooli? ATP transporditakse ATP/ADP translokaasi abil välja 12. Mitu prootonit kannab suktsinaat-CoQ reduktaas membraani ühelt küljelt teisele? Mitte ühtegi. 13
Klorofüllid on fotosünteesi põhipigmendid, mis neelavad nähtavat valgust. Fotofosforüülimine on ATP süntees valgusenergia arvel Fotoindutseeritud elektronide voog H2O-lt NADP+-le pumpab H+ läbi tülakoidi membraani stroomast luumenisse. Tülakoidi membraanis moodustub elektrokeemiline gradient. ATP sünteesi juhib tülakoidi luumenisse pumbatud H+ voolamine tagasi stroomasse ATP sünteesi eest kloroplastides vastutab ensüüm ATP süntaas, mis koosneb membraani läbivast prootonikanalist CF0 ja katalüütilisest perifeersest membraanivalgust CF1. Süsivesikute süntees Calvini tsüklis pimereaktsioonid. See on fotosünteetiline CO2 sidumise ja C taandamise tsükkel. Calvini (e. Calvin-Bensoni) tsüklis eristatakse 4 etappi: I etapp: CO2 sidumine aktseptorile, so pentoossuhkrule ribuloos-1,5- difosfaadile ning trioosi 3-fosfoglütseraadi teke
rasvkoes ja neerudes. Rasvhappe de novo süntees kujutab endast astetüül-CoA atsetüülijäägi korduvat lülitumist pikenevasse rasvhappe ahelasse. Rasvhappe de novo süntees toimub astmeliselt ja pole rasvhappeid lõhustava -oksüdatsiooni pöördprotsess. Süntees toimub tsütoplasmas. Süntees vajab redutseerivat energiat NADPH näol. Süntees vajab ATP, bikarbonaati ja mangaani. Sünteesiensüümide kompleks on multiensüümsüsteem mille üld nimi on rasvhappe süntaas. Süsteemi üheks komponendiks on atsüülkandja valk (ACP). Sünteesi iga ring pikendab ahelat 2 süsinikulise fragmendi võrra. Fragmengi doonoriks on melonüül-CoA ja vedavaks jõuks on CO2 vabanemine malonüül-CoA-lt. Sünteesi aktiveerib tsitraat ja inhibeerib rasvappe atsüül-CoA. Sünteesi soodustab kõrge insuliin. Rasvhappe de novo süntees on sisuliselt palmitaadi biosüntees. Palmitaadi baasil, pikendades ahelat ja tekitades kaksiksidemeid, toimub ka teiste rasvhapete süntees