Tallinna Ülikool
FOTOSÜNTEESI KEEMIA
Taimefüsioloogia referaat
Koostanud : Luise Tiks
Tallinn 2014
Sissejuhatus
Fotosüntees on protsess, mille abil mitmed bakteritüübid,
vetikad ning
vaskulaarsed taimed muundavad
valgusenergia orgaaniliste ainete
keemiliseks energiaks.
Fotosüntees hõlmab mitmeid füüsikalisi
ning keemilisi reaktsioone, mille käigus sünteesitakse
valgusenergia abil taandavaid agente (ferredoksiine ja
NADPH -d) ning
ATP-d. Saadud ühendeid kasutatakse lihtsate orgaaniliste ainete
sünteesiks süsihappegaasist ning lämmastik- ja sulfaatioonidest.
Sellised reaktsioonid on omased nii prokarüootidele kui
eukarüootidele ning on aluseks taime funktsioneerimisele.
Käesoleva töö eesmärgiks on anda ülevaade fotosünteesi
pimedusstaadiumis toimuvatest keemilistest reaktsioonidest, mis
võimaldavad organismidel keemiliste ühendite energiaks muundatud
päikeseenergiat kasutada süsiniku sidumiseks ning orgaaniliste
ainete sünteesiks. Lihtsustamise eesmärgil on töös lähemalt
kirjeldatud vaid C3 tüüpi fotosünteesi.
Töö põhineb David W. Lawleri
raamatul „Photosynthesis”
(
Oxford : BIOS, 2001).
ELEKTRONTRANSPORT FOTOSÜNTEESIS
Valgustsükli reaktsioonides toimub valgusenergia muundamine
keemiliseks energiaks. Valgusenergia mõjul toimub fotosünteesi
aktiivtsentri klorofülli-molekulide ergastamine, selle tulemusena
vabaneb elektron , mis transporditakse primaarsete aktseptoriteni, et
produtseerida tugevalt redutseeritud vaheühendeid. Saadud
vaheühendeid kasutatakse fotosünteesi pimedusstaadiumi
reaktsioonides süsihappegaasi taandamiseks.
Footoni jõudmine fotosüsteemi ning ergastumise liikumine fotosüsteem II ja I kannab energiat, mis on vajalik vee
oksüdatsiooniks, elektronide liikumiseks elektroni aktseptorile ning
prootonite liikumiseks tülakoidi luumenisse, et saaks toimuda ATP
süntees. Valgusenergia jõudmisel fotosüsteem II ( PSII )
reaktsioonitsentrisse, toimub klorofülli (P680) ergastumine, mille
tagajärjel vabaneb elektron. Selle tulemusena reaktsioonitsentri
P680 oksüdeerub ning tekib P680+. Tugeva oksüdeerijana eemaldab P680+ vett-lõhustavalt kompleksilt elektroni,
taastades nii neutraalse seisundi.
Vett-lõhustav kompleks on ensüüm, mis paikneb tülakoidmembraani
lumenaalsel küljel ning vastutab vee oksüdeerimise eest.
Ensüümkompleksiga on seotud Mn2+ ioonid , mille
oksüdeerumisel vabanev elektron liigub P680+-ile. Mn3+
ioonid omakorda osalevad väävli oksüdeerimisel. S toimib laengut
akumuleeriva kompleksina - vee oksüdeerimiseks peab eelpool kirjeldatud oksüdeerimiste ahel toimuma neli korda, kuni
produtseeritakse S4+. S4+ reageerib kahe
veemolekuliga:
S4+ + 2H2O → S + 4H+ + O2
Vabanenud 4H+ kasutatakse edaspidi ATP sünteesis.
Fotosüsteem I (PSI) ergastub samuti valgusenergia mõjul, seal
vabaneb elektron ning selle tulemusena toimub PSI-ga seotud
ferredoksiini redutseerimine . Redutseeritud ferredoksiin võtab
elektrondoonorina osa mitmetest bioloogilistest protsessidest,
näiteks NADP+ taandamine ferredoksiin NADP+
oksüdoreduktaasi mõjul edasise fotosünteesi käigus:
NADP+ + H+ + 2 ferredoksiin- →
NADPH + ferredoksiin
NADPH-d kasutatakse fotosünteesi pimedusstaadiumi reaktsioonides.
P680-lt vabanenud elektron liigub edasi läbi keerukate reaktsioonide
ning komplekside fotosüsteem I (PSI), taastades seal klorofülli
neutraalse seisundi.
ATP SÜNTEES
Adenosiin trifosfaadi süntees on põhiline energia salvestamise ning
edasi kandmise viis bioloogiliste protsessides. ATP võimaldab nii
energiat üle kanda kui ka fosforüleerimisprotsesse läbi viia. ATP
sünteesi mehhanism on sarnane nii bakterites , loomades kui taimedes.
Taimedes saadakse ATP sünteesiks vajalik energia fotosünteesi
valgusstaadiumist, kus kõrge redokspotentsiaaliga elektronide
liikumisel tülakoidis toimuvate reaktsioonide tulemusel vabanevad
vesinikioonid tekitavad prootongradiendi. Kõrge prootonite
kontsentratsioon kloroplasti luumenis põhjustab prootonite liikumise
läbi ATP süntaasi basaalse osa stroomasse, kus prootonite
kontsentratsioon on madal. Prootonite liikumine läbi ATP süntaasi
paneb ensüümi roteeruva kompleksi pöörlema ning varustab seeläbi
ATP sünteesiks vajaliku energiaga.
Reaktsiooni aktiveerimiseks peavad ADP ja fosfaatrühm liituma ATP
süntaasi „avatud” saidiga. Seejärel toimub rotatsioon, mille
tulemusena muutub aktiivsaidi struktuur ning ADP ja fosfaatrühm
moodustavad ATP, eraldub vesi. Edasisel rotatsioonil vabaneb ATP ning
aktiivsaidi struktuur taastub , mis võimaldab ADP ning fosfaatrühma
seondumist. Roteeruva kompleksi ühe täisringi jooksul
sünteesitakse 3 ATP molekuli ning selleks peab 12 H+
iooni liikuma läbi ATP süntaasi. ATP sünteesi kiirus on
reguleeritud valguse hulgaga , kuna suurem prootongradient võimaldab
intensiivsemat rotatsiooni. Reguleerivaks faktoriks on ka NADPH ja
ATP koguste suhe – NADPH liia korral suureneb ka ATP süntees, et
elektrone ning prootoneid maksimaalselt ära kasutada.
SÜSINIKU SIDUMINE NING SÜSINIKUÜHENDITE SÜNTEES
Süsiniku omastamine on väga täpselt reguleeritud hästi
integreeritud mehhanismide poolt, mis võimaldab fotosünteesi
aktiivsust pidevalt vajaduse ning muutuvate tingimustega vastavuses
hoida. Süsiniku assimilatsioon on tsükliline, autokatalüütiline
protsess, mida sageli nimetatakse ka Calvini tsükliks või PCR
tsükliks. CO2 assimilatsiooni mehhanismi, mis toimib PCR tsükli
kaudu, nimetatakse C3 fotosünteesiks, kuna esimene protsessi käigus
tekkiv stabiilne vaheprodukt on kolme-süsinikuline. Lisaks C3
fotosünteesile eristatakse veel C4 ning CAM fotosünteesi, mille
puhul eelnevad PCR tsüklile CO2 siduvad protsessid.
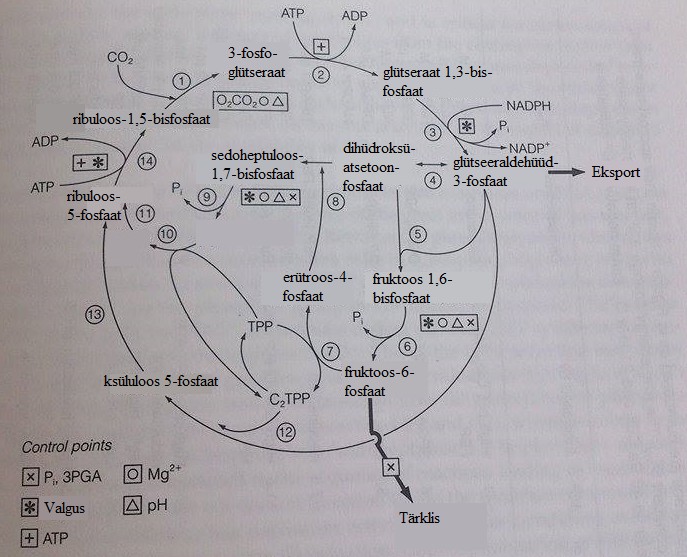
PCR tsükli reaktsioonid
PCR tsükkel on 13-astmeline karboksüleerimisprotsess (vt joonist
1), millest võtab osa 11 ensüümi. PCR tsükli alguseks loetakse
primaarset süsiniku ja ribuloos-1,5-bisfosfaadi (RuBP) sidumise reaktsiooni, mida katalüüsib RuBP karboksülaas- oksügenaas ( Rubisco ):
RuBP + CO2 → 2 3-PGA
Tavaolekus on Rubisco inaktiivne ning aktiivtsentrisse on tugevalt
seondunud RuBP molekul , mis takistab aktivatsiooniks vajalikku CO2
seondumist. Reaktsiooni käivitamiseks on vaja Rubisco aktivaasi, mis
eemaldab dekarbamüleeritud (inaktiivsest) aktiivtsentrist RuBP ning
võimaldab karbamüleerimist ning Mg2+ liitumist aktiivtsentrisse. Seeläbi formeerub katalüütiliselt aktiivne tsenter , kuhu liitub uuesti RuBP. Reaktsiooni käigus reageerib CO2
otseselt ning elektrofiilselt RuBP tautomeerse kompleksi C2-ga,
moodustades 6-süsinikulise vaheühendi
2-karboksü-3-ketoarabinitool-1,5-bisfosfaadi. Seejärel toimub vee
mõjul vaheühendi lagunemine kaheks 3-fosfoglütseerhappe molekuliks
– vesi loovutab hüdroksüülrühma C3 karbonüülrühmale,
moodustades deprotoneeritava gem-diooli, põhjustades C2-C3 sideme
lagunemist. Antud reaktsioon on PCR tsüklile ainomane ning seda
katalüüsiv ensüüm Rubisco esineb kõigis fotosünteesivates
organismides ning on tõenäoliselt üks levinumaid valke maal,
moodustades kuni 50% kõrgemate taimede lehtedes leiduvatest
lahustuvatest valkudest.
Teises PCR tsükli etapis fosforüleerib fosfoglütseraadi kinaas ATP
abil 3PGA 1,3-difosfoglütseerhappeks, mis on tänu kahele
happelisele anhüdriidsidemele aktiivsem. Seejärel asendab NADP
glütseeraldehüüd-3- fosfaadi dehüdrogenaas 1,3-difosfoglütseraadi
fosfaatrühma H+ vastu, moodustades glütseeraldehüüd-3- fosfaadi (GAP). Vesinikuallikana kasutatakse valges NADP+, kuid pimedas
kataboolse ATP sünteesi ajal NAD+. Tegemist on PCR tsükli ainsa
taandamisreaktsiooniga. GAP sünteesitakse trioos fosfoisomeraasi
toimel dihüdroatsetoonfosfaadiks (DHAP). 3-süsinikulistest GAP ja
DHAP molekulidest sünteesitakse aldolaaside mõjul 6-süsinikuline fruktoos -1,6-bisfosfaat (FBP) või 7-süsinikuline
sedoheptuloos-1,7-bisfosfaat (SBP). Fruktoos-1,6-bisfosfaas
defosforüleerimisel fruktoosi bisfosfataasiga saadakse
fruktoos-6- fosfaat (F6P). Osa fruktoos-6-fosfaadi molekule
kasutatakse tärklise sünteesiks, osa on vajalikud RuBP
taastamiseks.
RuBP taastamine saavutatakse 3-, 4-, 5- ja 6-süsinikuliste ühendite
vastastikuste üleminekute kaudu. Selleks eemaldab transketolaas
fruktoos-1,6-fosfaadilt ning sedoheptuloos-7-fosfaadilt
2-süsinikulised fragmendid , moodustades nii vastavalt
erütroos-4-fosfaadi (E4P) ning riboos-5-fosfaadi (R5P).
Erütroos-4-fosfaat reageerib omakorda DHAP-ga – tekib
sedoheptuloos-1,7-bisfosfaat, mis defosforüleeritakse bisfosfataasi
mõjul ja muundatakse transketolaasi mõjul riboos-5-fosfaadiks ning
seejärel ribuloos-5-fosfaadiks (Ru5P). Samal ajal moodustub
transketolaasi mõjul glükoaldehüüdist ja
glütseeraldehüüd-3-fosfaadist ksüluloos-5-fosfaat, mis
ribuloos-5-fosfaat 3-epimeraasi toimel Ru5P-ks konverteeritakse.
Calvini tsükli viimaseks sammuks loetakse RuBP taastamist Ru5P
fosforüleerimisel fosforibulokinaasi abil.
Joonis 1. PCR tsükkel
PCR tsükli produktide metabolism
Fotosünteesi üks põhilisemaid produkte on sahhariidid, mis on
edaspidiseks energiaallikaks nii taimele endale, kui ka
loomorganismidele. Kõige levinum oligosahhariid taimedes on sahharoos – varuaineid transporditakse taimeorganite vahel
peamiselt sahharoosi kujul. Sahharoos koosneb glükopüranoosist
ning fruktofuranoosist, mis on omavahel seotud 1-2 glükosiidsidemega.
Sahharoosi sünteesi esimene etapp on fruktoos-1,6-bisfosfaadist
hüdrolüüs:
Fru-1,6-BP + H2O > F6P + Pi
Seejärel katalüüsib sahharoosi fosfaatsüntaas Fru-6-P reaktsiooni
uridiindifosfaat-glükoosiga (UDPG). Selle tulemusena liigub
glükoosijääk UDPG-lt F6P-le:
UDPG + F6P > UDP + sahharoos-6-fosfaat + H+
Sahharoosi sünteesi viimases etapis toimub sahharoos-6-fosfaadi
defosforüleerimine sahharoosi fosfataasi poolt:
Sahharoos-6-fosfaat + H2O > sahharoos + Pi
Sahharoosi süntees on võimalik ka UDPG ning fruktoosi reaktsioonil,
sellisel juhul jääb defosforüülimisetapp vahelt ära. Taimedes on
säärasel reaktsioonil ebaoluline tähtsus.
Teine põhiline varuaine taimedes on tärklis, mille põhiline
funktsioon on energiavarude säilitamine taimes. Tärklis
sünteesitakse PCR- tsüklis toodetud fruktoos-6-fosfaadist, mis
heksoosfosfaatisomeraasi toimel glükoos -6-fosfaadiks
konverteeritakse. Seejärel muundatakse glükoos-6-fosfaat
fosfoglükomutaasi toimel glükoos-1-fosfaadiks, mis omakorda ADP
glükoosi pürofosforülaasi toimel ADP-ga liidetakse:
Glükoos-1-fosfaat + ATP → ADP-glükoos + PPi
ADP-glükoosi kasutatakse tärklise sünteesimisel, lisades tärklise
süntaasi abil ADP-glükoosile 1,4-sidemete kaudu glükoosimolekule.
Peale sahhariidide sünteesitakse kloroplastides ka organellile
vajalikke lipiide ning aminohappeid .
Lipiidid moodustavad kloroplasti kuivmassist umbes ühe kolmandiku.
Rasvhapete sünteesis kasutatakse PCR tsükli produkte, taandavaid
agente (nt ferrodoksiin ning NADPH) ja ATP-d ning peale kloroplasti
toimub sünteesi osa etappe ka tsütosoolis ning mitokondrites. PCR-i
produktidest sünteesitakse stroomas dihüdroksüatsetoonfosfaat, mis
liigub tsütosooli, ning sealt edasi mitokondrisse, kus sünteesitakse
atsetüül-CoA, mis liigub kloroplasti tagasi. Kloroplastis
konverteeritakse atsetüül-CoA-st malonüül-CoA ning seejärel
malonüül-ACP. Ühe atsetüül-ACP ning seitsme malonüül-ACP
liitmisel ning taandamisel NADPH abil saadakse palmitüül-ACP, mis
edasi hüdrolüüsitakse palmithappeks. Võimalik on ka palmitüül-ACP
edasine pikendamine teisteks rasvhapeteks ning rasvhapete edasine
modifitseerimine.
Kloroplastides leidub ensüüme enamiku valkudes leiduvate
aminohapete sünteesiks. Aminohapete sünteesis on väga oluline roll
lämmastiku assimilatsioonil. Taimed seovad lämmastikku peamiselt
NO3- kujul. Seejärel taandatakse nitraatioon
nitritiooniks ning ammoniaagiks:
NO3- + 2 e- + 2H+ → NO2-
+ H2O
NO2- + 6 e- + 7 H+ → NH3
+ 2 H2O
Seejärel seotakse NH3 2-oksoglutaraadiga või
glutamaadiga:
2-oksoglutaraat + NH3 + NAD(P)H + H+ → glutamaat +
NAD(P) + + H2O
Glutamaat + NH3 + ATP → ADP + Pi + H2O
Edasises aminohapete sünteesis kasutatakse peamiselt glutamaati ning
PCR tsükli produkte. Näiteks sünteesitakse 3PGA-st
fosfoenoolpüruvaat (PEP), seejärel konverteeritakse PEP
püruvaadiks, mille transamineerimisel glutamaadiga moodustub
alaniin.
Mõned aminohapped sünteesitakse osaliselt kloroplastis ning
osaliselt tsütoplasmas, kuid peamised prekursorid aminohapete
sünteesiks sünteesitakse kõik fotosünteesi või fotosünteesi
produktide modifitseerimise tulemusena kloroplastides.
KOKKUVÕTE
Fotosüntees on äärmiselt keerukas protsess, millest võtab osa
hulgaliselt ensüüme ning kofaktoreid. Kloroplastis on välja
kujunenud sünteesirajad enamiku vajalike sahhariidide,
kloroplastilipiidide ning aminohapete sünteesiks. Lisaks sellele
toodetakse sahhariide kõigi teiste rakuosade ning mitte
fotosünteesivate rakkude varustamiseks. Seetõttu on fotosüntees
äärmiselt kompleksne ning hästi reguleeritud protsesss.
Fotosünteesi lähteaineteks võib lugeda süsihappegaasi ja vee, produktideks peamisel fruktoos-6-fosfaadi ning lipiidide sünteesi
puhul ka dihüdroksüatsetoonfosfaadi. Protsessi erinevad etapid
kulgevad peamiselt tülakoidmembraanil, aga ka kloroplasti stroomas
ja luumenis. Erinevate etappide koostoime on tagatud seeläbi, et iga etappi koordineerib hulk ensüüme, mille hulgast olulisimaks võib
lugeda Rubisco.


























Kõik kommentaarid