Valkude ruumiline struktuur 1. Valkude struktuuri määravad
faktorid 2. Valkude sekundaarstruktuur
3. Valkude
tertsiaarstruktuur 4. Valkude
kvaternaarstruktuur 5. Valkude struktuuri näiteid: RibonukleaasA, Müoglobiin,
Hemoglobiin , Insuliin Valkude struktuur?
·
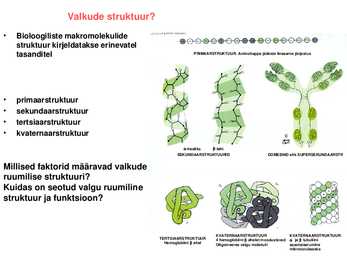
Bioloogiliste makromolekulide struktuur kirjeldatakse erinevatel tasanditel
PRIMAARSTRUKTUUR . Aminohappe jääkide lineaarne järjestus
· primaarstruktuur
· sekundaarstruktuur
· tertsiaarstruktuur
· kvaternaarstruktuur
heeliks leht SEKUNDAARSTRUKTUURID
DOMEENID ehk SUPERSEKUNDAARSTR
Millised faktorid määravad valkude ruumilise struktuuri?
Kuidas on seotud valgu ruumiline struktuur ja funktsioon?
KVATERNAARSTRUKTUUR KVATERNAARSTRUKTUUR TERTSIAARSTRUKTUUR 4
hemoglobiini ahelat moodustavad ja tubuliini Hemoglobiini ahel Oligomeerse valgu molekuli assotsieerumine mikrotorukeseks Valkude ruumiline struktuur
regulaarne või irregulaarne?
Primaarstruktuurid tohutult varieeruvad 1934
pepsiini kristallidelt
difraktsioon 1958 müoglobiini struktuur (Kendrew) 1965 lüsotsüüm
raeguseks on teada ca 10000 valgu struktuur
"
Perhaps the most remarkable
features of the molecule are its
complexity and its
lack of symmetry. The arrangement
seems to be
almost totally lacking in the kind of regularities which one instinctively anticipates, and it is more complicated
than has been predicted by any theory."
John Kendrew kommenteerides esimest globulaarse valgu kristallstruktuuri,
Cambridge University, 1958 Valkude struktuurse organisatsiooni
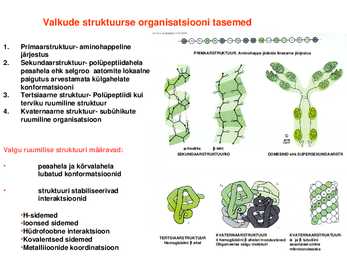
tasemed1. Primaarstruktuur aminohappeline järjestus PRIMAARSTRUKTUUR. Aminohappe jääkide lineaarne järjestus
2. Sekundaarstruktuur polüpeptiidahela peaahela ehk
selgroo aatomite
lokaalne paigutus arvestamata külgahelate konformatsiooni
3.
Tertsiaarne struktuur Polüpeptiidi kui terviku ruumiline struktuur
4. Kvaternaarne struktuur subühikute ruumiline
organisatsioonValgu ruumilise struktuuri määravad:
heeliks leht SEKUNDAARSTRUKTUURID DOMEENID ehk SUPERSEKUNDAARSTR
· peaahela ja kõrvalahela lubatud konformatsioonid
· struktuuri stabiliseerivad
interaktsioonid·Hsidemed ·Ioonsed sidemed ·Hüdrofoobne
interaktsioon KVATERNAARSTRUKTUUR KVATERNAARSTRUKTUUR ·Kovalentsed sidemed TERTSIAARSTRUKTUUR Hemoglobiini ahel 4 hemoglobiini ahelat moodustavad ja tubuliini Oligomeerse valgu molekuli assotsieerumine ·Metalliioonide koordinatsioon mikrotorukeseks Peptiidrühma ruumiline struktuur I
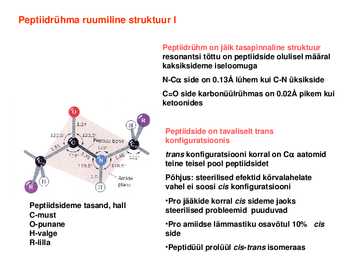
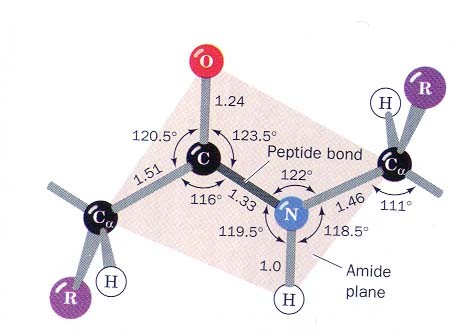
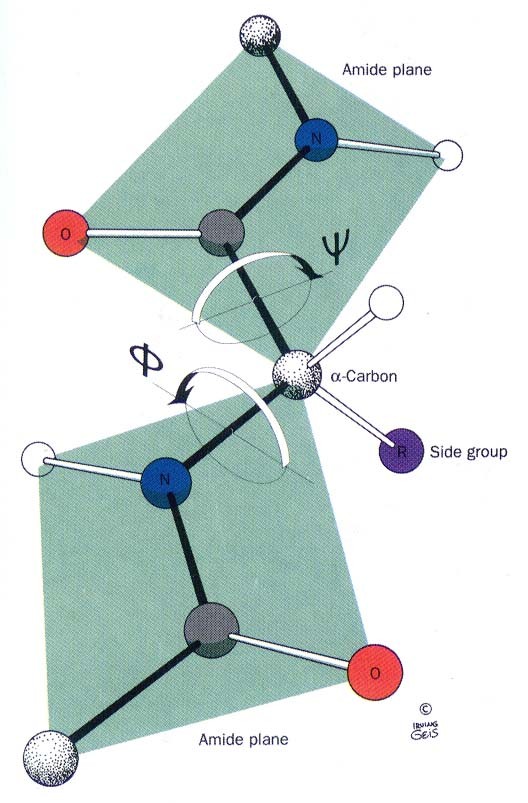
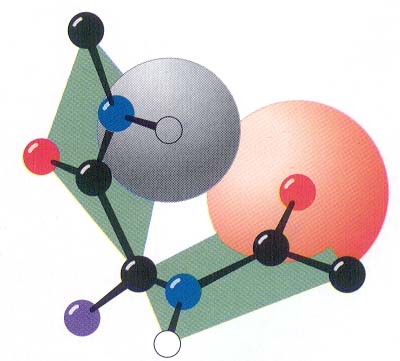
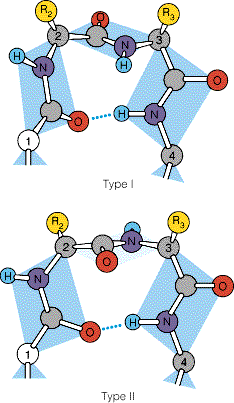
Peptiidrühm on jäik tasapinnaline struktuur resonantsi tõttu on peptiidside olulisel määral kaksiksideme iseloomuga NC side on 0.13Å lühem kui CN üksikside C=O side karbonüülrühmas on 0.02Å pikem kui ketoonides
Peptiidside on tavaliselt trans konfiguratsioonis trans konfiguratsiooni korral on C
aatomid teine teisel pool
peptiidsidet Põhjus: steerilised efektid kõrvalahelate vahel ei soosi cis konfiguratsiooni ·Pro jääkide korral cis sideme jaoks Peptiidsideme tasand, hall steerilised probleemid puuduvad Cmust Opunane ·Pro amiidse lämmastiku osavõtul 10% cis Hvalge side Rlilla ·Peptidüül prolüül cistrans
isomeraas Regulaarsed sekundaarstruktuuri elemendid valgus
Linus Paulig koos kaastöötajatega
1950ndadRegulaarsed polüpeptiidahela konformatsioonid peavad rahuldama teatud
kindlaid nõudeid:
1. Polüpeptiidis esinevad sidemete pikkused ja nurgad peavad võimalikult vähe kõrvale
kalduma peptiidide puhul katseliselt määratutest 2. Kaks aatomit ei tohi paikneda teineteisele lähemal kui lubavad vastavad van der Waalsi
raadiused 3. Peptiidside peab olema planaarne ja trans konfiguratsioonis 4. Polüpeptiidahela amiidrühma
prootonid ja karbonüülsed hapnikud peavad olema kaasatud vesiniksidemete moodustamisse Peptiidahela
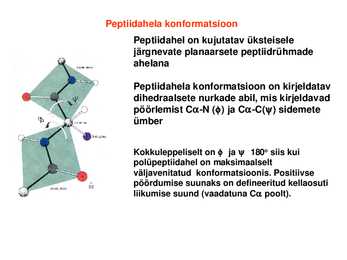
konformatsioon Peptiidahel on
kujutatav üksteisele järgnevate planaarsete peptiidrühmade ahelana
Peptiidahela konformatsioon on kirjeldatav dihedraalsete nurkade abil, mis
kirjeldavad pöörlemist CN () ja CC() sidemete ümber
Kokkuleppeliselt on ja 180o siis kui polüpeptiidahel on maksimaalselt väljavenitatud konformatsioonis. Positiivse pöördumise suunaks on defineeritud
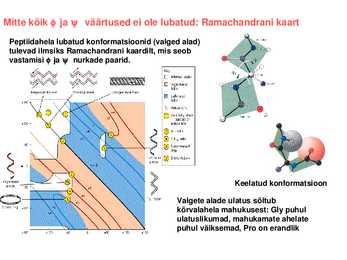
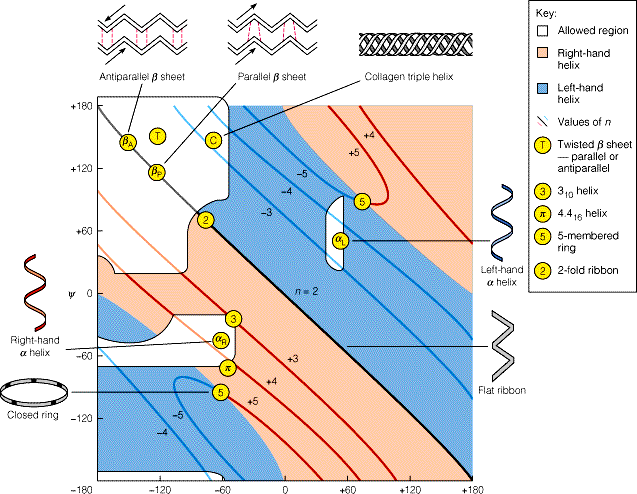
kellaosuti liikumise suund (vaadatuna C poolt). Mitte kõik ja väärtused ei ole lubatud: Ramachandrani kaart Peptiidahela lubatud konformatsioonid (valged alad) tulevad ilmsiks Ramachandrani kaardilt, mis seob vastamisi ja nurkade paarid.
Keelatud konformatsioon
Valgete alade ulatus sõltub kõrvalahela mahukusest: Gly puhul ulatuslikumad, mahukamate
ahelate puhul väiksemad, Pro on
erandlik Regulaarsete (korduvate) ja väärtustega sekundaarstruktuurid
2 olulist regulaarset sekundaarstruktuuri, kus
vesiniksidemed peaahela elementide vahel stabiliseerivad molekuli on:
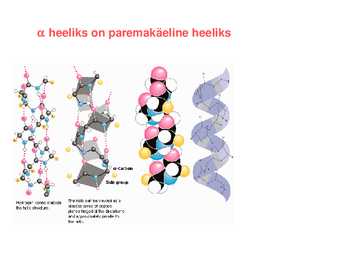
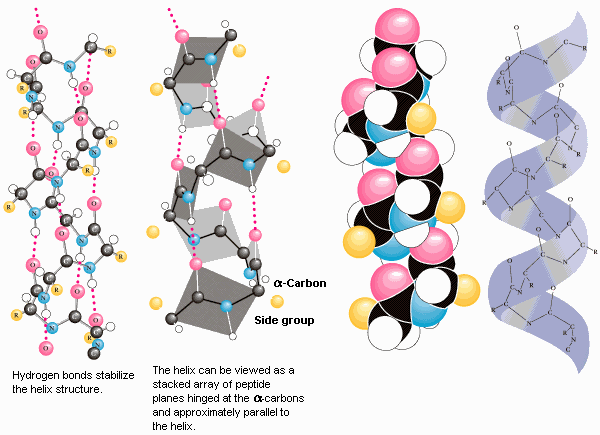
1. heeliks 2. struktuur heeliks: 1951 Paulig
· Ainuke heeliks, mis vastab piiranguteta Ramachandrani kaardile ja millel on energeetiliselt kasulikud regulaarsed H sidemed
· 3.6 aminohapet tiiru peale
heeliksi tõus 1,5Å · Keskmine heeliski pikkus 12 aminohappe jääki, 18Å · Peptiidrühma karbonüül moodustab H sideme n+4 peptiirühma NH rühmaga, iga amiidi
vesinik ja karbonüüli hapnik moodustavad sideme · Heeliksi sisemuses aatomid tihedalt pakitud · Mitu heeliksit võivad valgus omakorda kokku keerdunud olla heeliks on paremakäeline heeliks Regulaarsete (korduvate) ja väärtustega sekundaarstruktuurid II
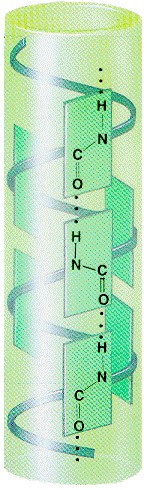
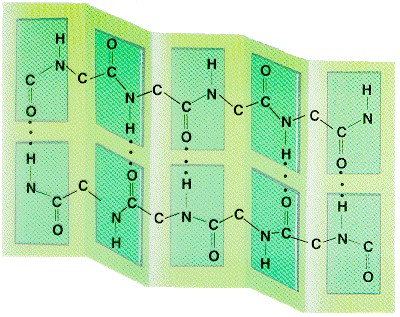
struktuur: Paulig ja Corey 1951 volditud leht
H sidemed ei moodusta sama ahela vaid külgneva ahela rühmitustega. Stabiliseerivad H sidemed tekivad primaarstruktuuris üksteisest kaugel asuvate aminohappe jääkide vahel. 1. Antiparalleelne leht külgnevate ahelate N C suunalisus
vastupidine 2. Paralleelne leht külgnevate ahelate suunalisus sama lehed koosnevad 2..12
ahelast , iga ahel keskmiselt 6 aminohappe jääki Paralleelsed lehed alla 5 ahela haruldased, nende stabiilsus väiksem. Sageli esinevad paralleelne ja antiparalleelne struktuur
segamini . Lehed tihti pöördes Ahelate omavahel ühendamise topoloogia võib olla kompleksne · lehe struktuuris ei ole karbonüülid orienteeritud ühes suunas nagu heeliksi korral. · heeliksis on ühe aminohappe jäägi kohta 0.15 nm heeliksi
pikkuses , lehe struktuuris 0.320.34 nm ·Mõlemad regulaarsed
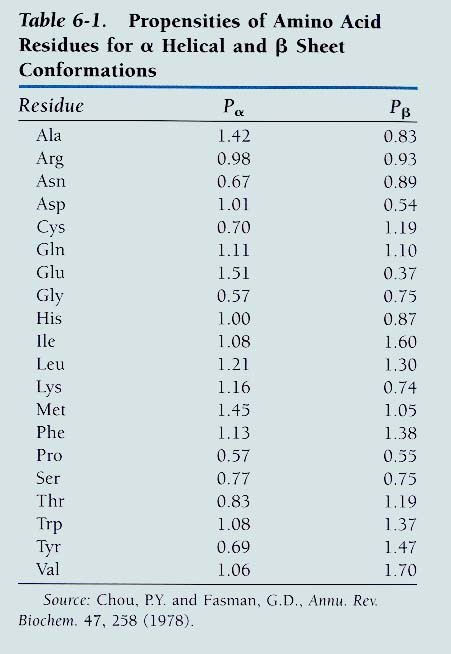
struktuurid on sageli bipolaarsed. Solvendi poole eksponeeritud osas paiknevad polaarsed ja laetud aminohappe jäägid Kas sekundaarstruktuur on ennustatav primaarstruktuurist lähtudes?
Chou ja Fasman 1976
·Homoloogia alusel
modelleerimine ·Valkude kokkupakkumise modelleerimine ·Irregulaarsused standardsetes struktuurides ·
Helix capping (Asn, Gln)
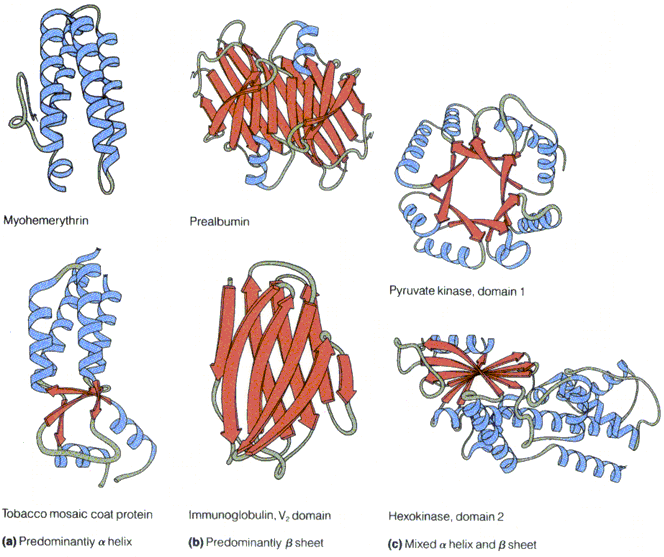
Valgud jaotatakse ajalooliselt globulaarseteks ja fiibervalkudeks
Esialgne jaotus
morfoloogia alusel
Fiibervalgud
domineerib 1 sekundaarstruktuur
Fiibervalgud on tüüpiliselt vees lahustumatud kasutusel sidekudedes
Näited:
Kollageen ,
keratiin , siidi fibroiin
Globulaarsed valgud vees lahustuvad. Enamus valke rakkudes. Heterogeense sekundaarstruktuuriga. Oluline tertsiaarstruktuur
tuleneb ebareeglipärastest kõrvalahelate interaktsioonidest (interaktsioonid solvendiga, hüdrofoobne
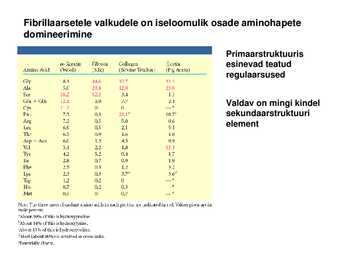
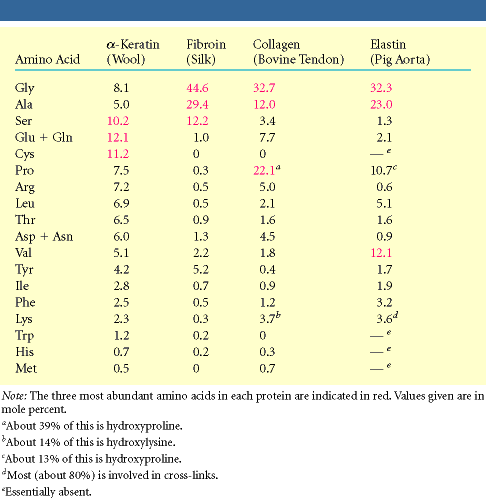
interaktsioon , ioonsed interaktsioonid, vesinikside ja kovalentsed sidemed) Fibrillaarsetele valkudele on iseloomulik osade aminohapete domineerimine
Primaarstruktuuris esinevad teatud regulaarsused
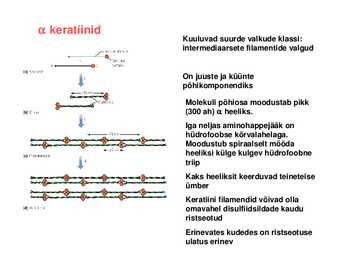
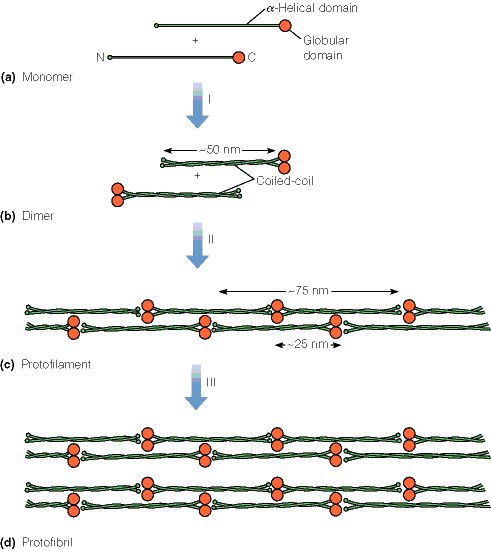
Valdav on mingi kindel sekundaarstruktuuri element keratiinid Kuuluvad suurde valkude klassi: intermediaarsete filamentide valgud
On juuste ja küünte põhikomponendiks
Molekuli põhiosa moodustab pikk (300 ah) heeliks. Iga neljas aminohappejääk on hüdrofoobse kõrvalahelaga. Moodustub spiraalselt mööda heeliksi külge kulgev hüdrofoobne
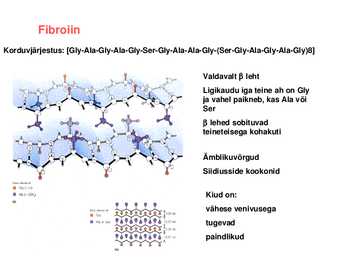
triip Kaks heeliksit keerduvad teineteise ümber Keratiini filamendid võivad olla omavahel disulfiidsildade kaudu ristseotud Erinevates kudedes on ristseotuse ulatus erinev Fibroiin
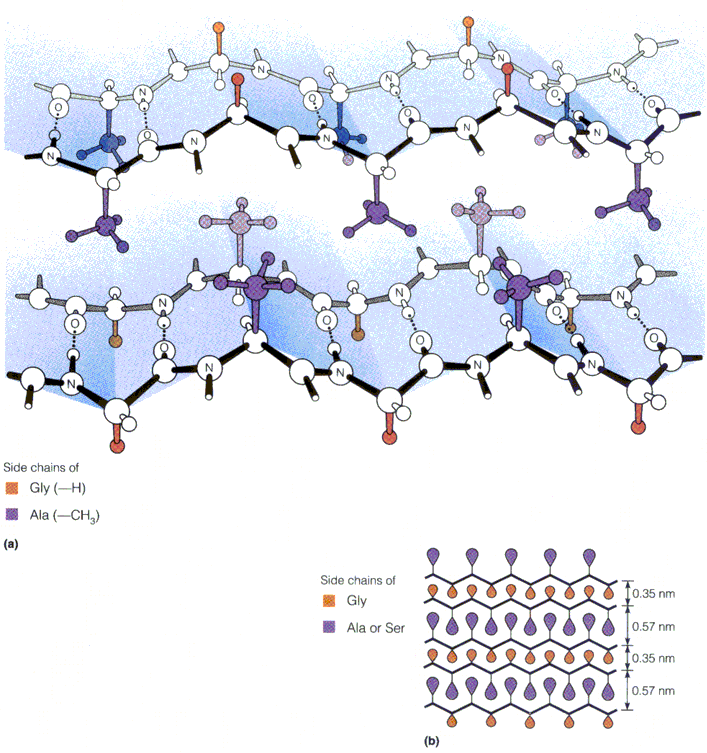
Korduvjärjestus: [GlyAlaGlyAlaGlySerGlyAlaAlaGly(SerGlyAlaGlyAlaGly)8]
Valdavalt leht Ligikaudu iga teine ah on Gly ja vahel paikneb, kas Ala või Ser lehed sobituvad teineteisega kohakuti
Ämblikuvõrgud Siidiusside kookonid
Kiud on: vähese venivusega tugevad paindlikud
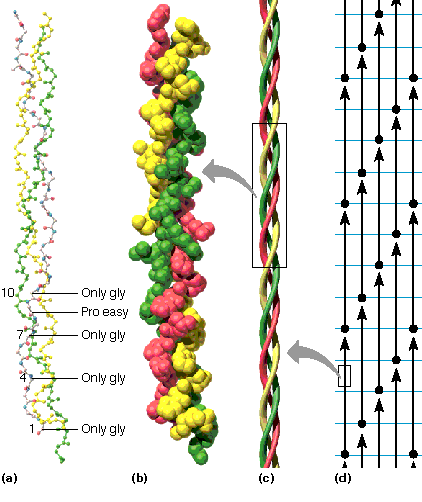
Kollageeni kolmikheeliks imetajal on 1/3 valgust kollageen
·Heeliks (ca 1000 ah) on vasakukäeline ja rohkem välja venitatud kui heeliks ·Kolm heeliksit keerduvad paremakäeliselt moodustades kolmikheeliksi · Primaarstruktuur koosneb kordustest Gly XaaYaa ·Xaa on enamasti Pro ja Yaa hüdroksüproliin
Moodustab luudes maatriksi kuhu ladestuvad mineraalained Kõõluste põhikomponent Oluline naha struktuuri tagamisel
Kollageen hoiab loomi koos
Vitamiin C ehk askorbiinhappe peamine funktsioon on osalemine kollageeni proliini ja lüsiini jääkide hüdroksüleerimises.
Defitsiidi korral areneb välja
skorbuut Globulaarsed valgud: ruumilise struktuuri ja funktsiooni
mitmekesisusValkude ruumilise struktuuri määramine
1. Röntgenkristallograafia 2. Tuumamagnetresonants spektroskoopia ehk NMR
1. 2.
Vajalikud valgu
kristallid NMR töötab valgu lahusega
Tüüpiliselt saavutatav 23.5Å Limiteeritud ca 30 kD suurusete lahutuvus valkudega
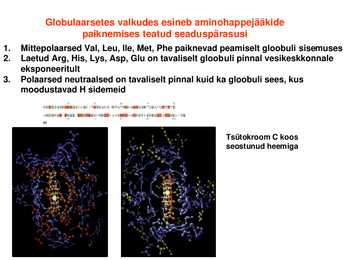
Kristallstruktuur kattub küllaltki hästi struktuuriga lahuses Globulaarsetes valkudes esineb aminohappejääkide paiknemises teatud seaduspärasusi
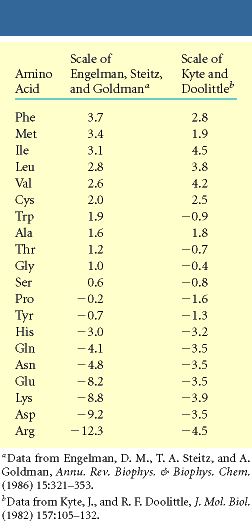
1. Mittepolaarsed Val, Leu, Ile, Met, Phe paiknevad peamiselt gloobuli sisemuses
2. Laetud Arg, His, Lys, Asp, Glu on tavaliselt gloobuli pinnal vesikeskkonnale eksponeeritult
3. Polaarsed neutraalsed on tavaliselt pinnal kuid ka gloobuli sees, kus moodustavad H
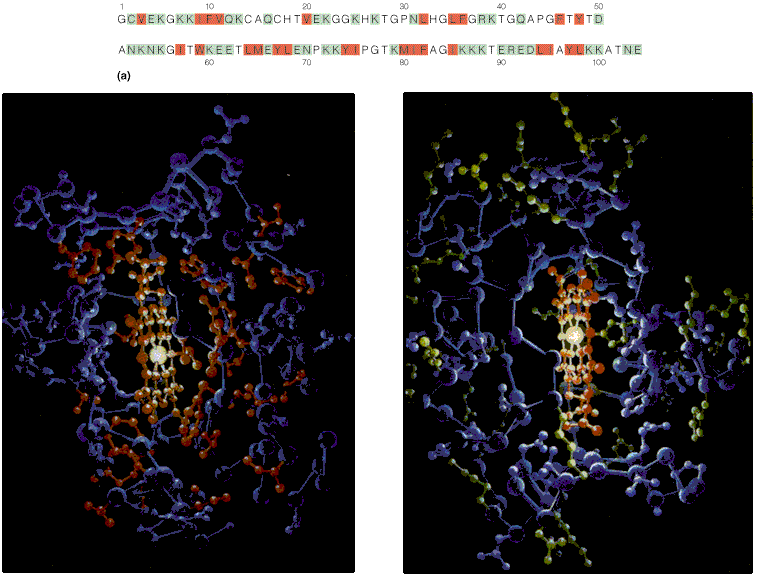
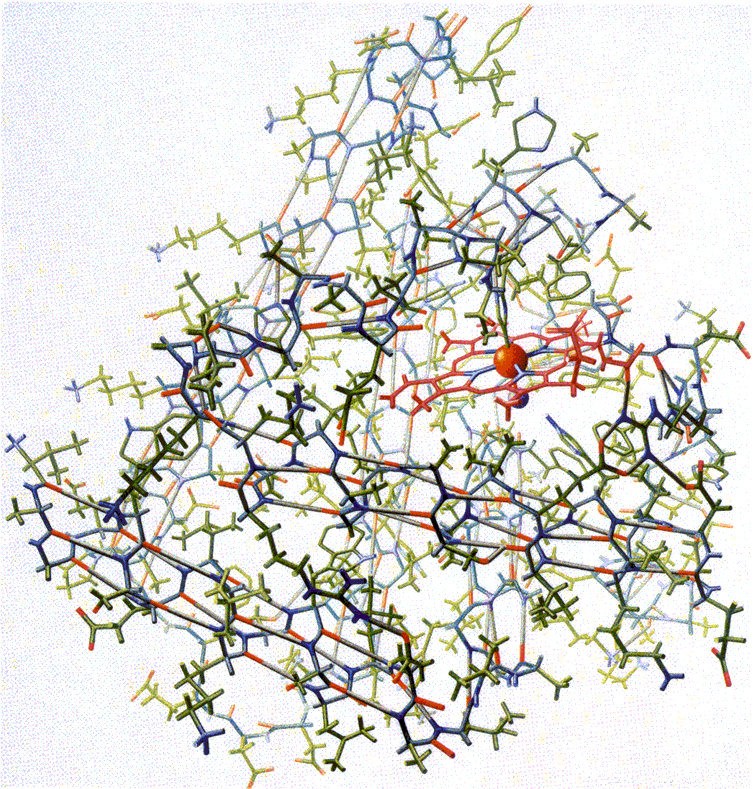
sidemeidTsütokroom C koos
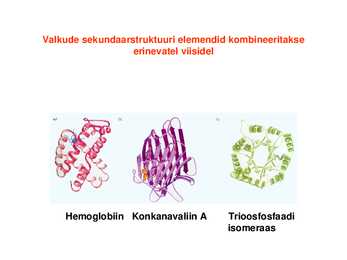
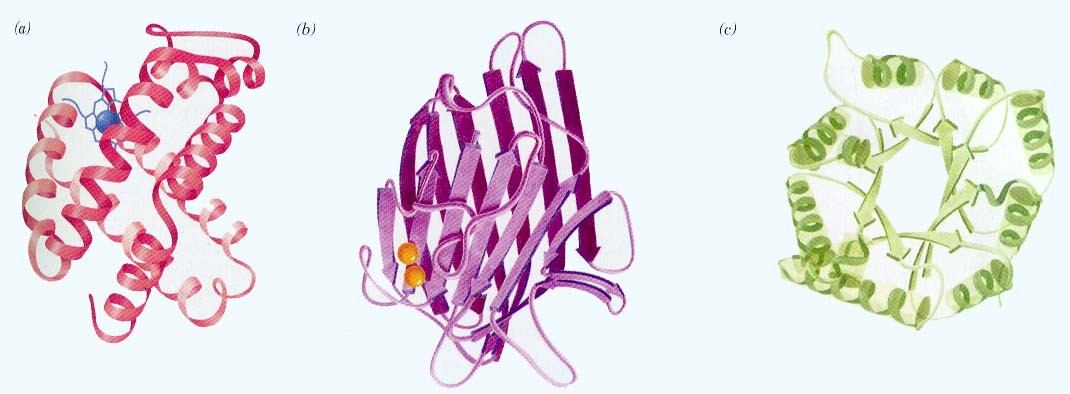
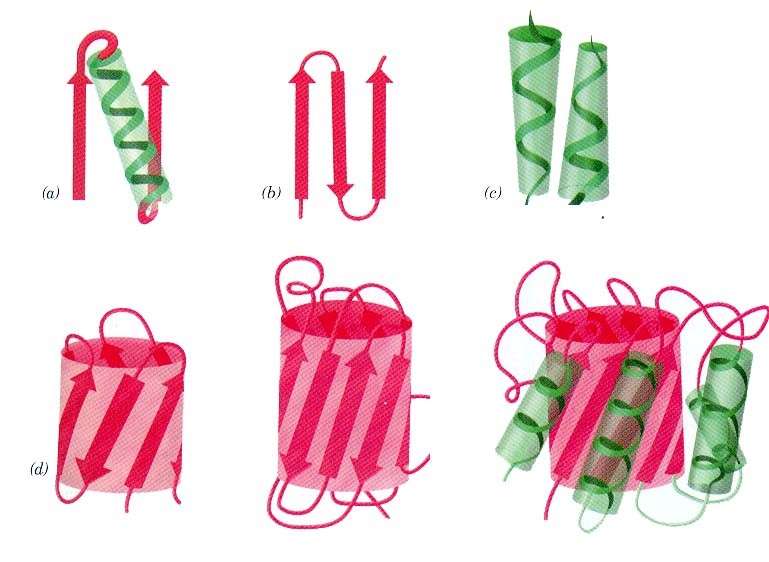
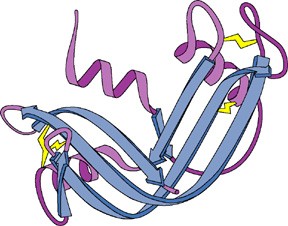
seostunud heemiga Valkude sekundaarstruktuuri elemendid kombineeritakse erinevatel viisidel
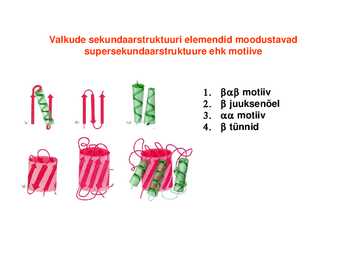
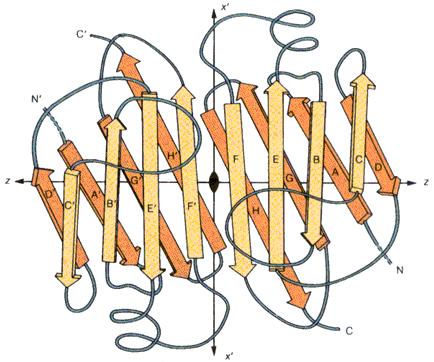
Hemoglobiin Konkanavaliin A Trioosfosfaadi isomeraas Valkude sekundaarstruktuuri elemendid moodustavad supersekundaarstruktuure ehk
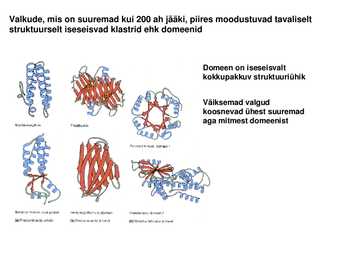
motiive1. motiiv 2. juuksenõel 3. motiiv 4. tünnid Valkude, mis on suuremad kui 200 ah jääki, piires moodustuvad tavaliselt struktuurselt iseseisvad klastrid ehk domeenid
Domeen on iseseisvalt kokkupakkuv struktuuriühik
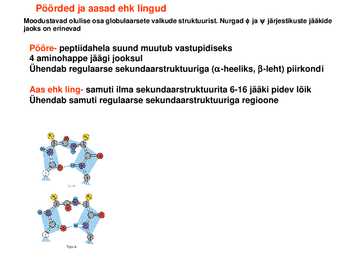
Väiksemad valgud koosnevad ühest suuremad aga mitmest domeenist Pöörded ja aasad ehk lingud
Moodustavad olulise osa globulaarsete valkude struktuurist. Nurgad ja järjestikuste jääkide jaoks on erinevad
Pööre peptiidahela suund muutub vastupidiseks 4 aminohappe jäägi jooksul Ühendab regulaarse sekundaarstruktuuriga (heeliks, leht) piirkondi
Aas ehk
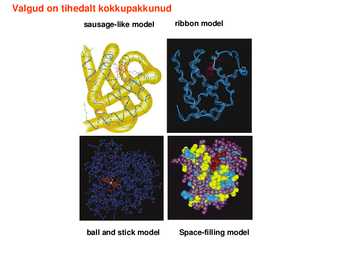
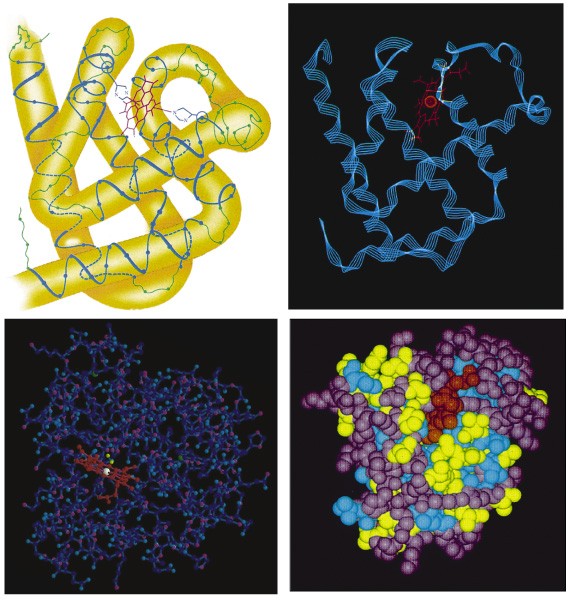
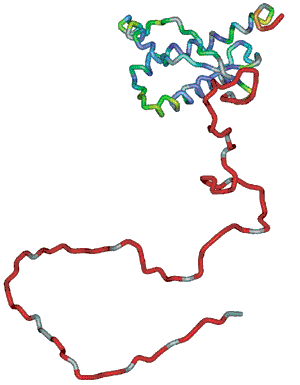
ling samuti ilma sekundaarstruktuurita 616 jääki pidev lõik Ühendab samuti regulaarse sekundaarstruktuuriga regioone Valgud on tihedalt kokkupakkunud sausagelike model ribbon model
ball and
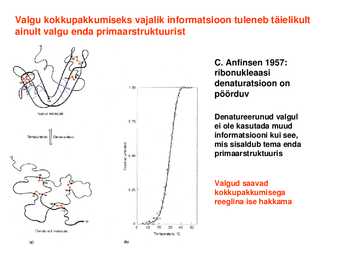
stick model Spacefilling model Valgu kokkupakkumiseks vajalik informatsioon tuleneb täielikult ainult valgu enda primaarstruktuurist
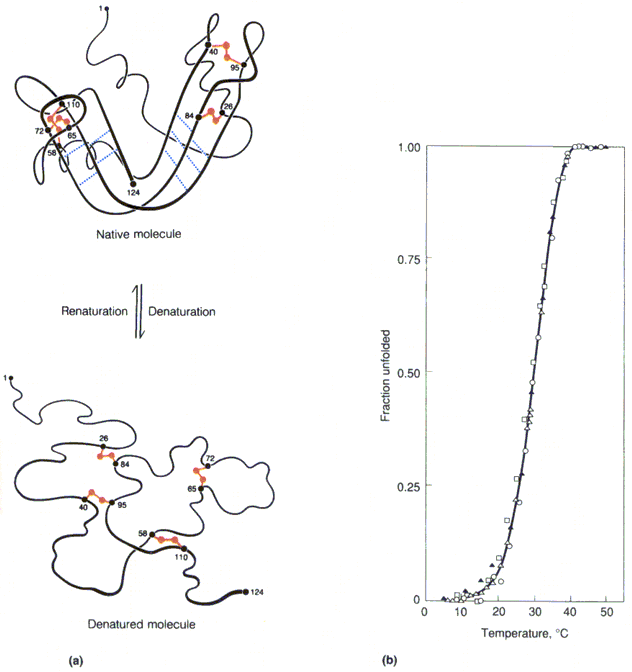
C. Anfinsen 1957: ribonukleaasi
denaturatsioon on pöörduv
Denatureerunud
valgul ei ole kasutada muud informatsiooni kui see, mis sisaldub tema enda primaarstruktuuris
Valgud saavad kokkupakkumisega reeglina ise hakkama Valkude kokkupakkumine ja stabiilsus
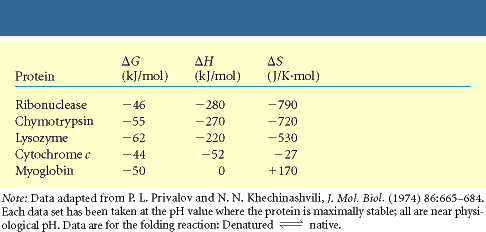
Natiivsed valgud on sageli väga kergelt denatureeritavad: 100 aminohappe jäägi kohta 40 kJ/mol (see on ca kahe H sideme energia)!
Valgu struktuuri stabiliseerivad faktorid: 1. Hüdrofoobne efekt 2. Nõrgad interaktsioon:
elektrostaatiline , Van der Waals, H side 3. Kovalentsed sidemed: SS
sillad tsüsteiini jääkide vahel 4. Stabiliseerivad metallioonid (Zn
finger )
Valgu struktuuri destabiliseerib konformatsiooniline
entroopia Erinevate faktorite osakaal valkude kokkupakkumises sõltub konkreetsest valgust
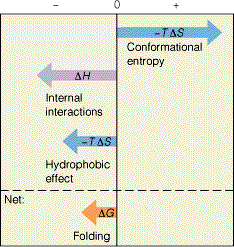
G = H T S
TS jaguneb: konformatsiooniline entroopia (ebasoodne) hüdrofoobne efekt (soodne)
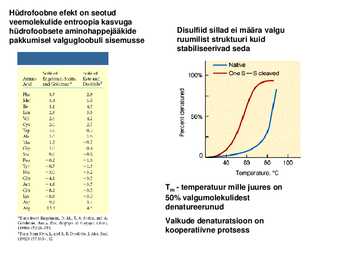
Soodne H jaguneb: soolasillad molekulisisesed H sidemed van der Waalsi interaktsioonid disulfiidsillad Hüdrofoobne efekt on seotud veemolekulide entroopia kasvuga hüdrofoobsete aminohappejääkide Disulfiid sillad ei määra valgu pakkumisel valgugloobuli sisemusse ruumilist struktuuri kuid stabiliseerivad seda
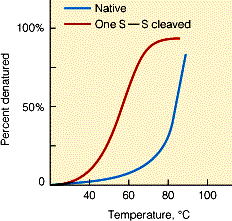
Tm temperatuur mille juures on 50% valgumolekulidest denatureerunud Valkude denaturatsioon on
kooperatiivne protsess Valkude denaturatsioon ja
renaturatsioonDenatureerivad faktorid 1. Temperatuur.
Termiline denaturatsioon on kooperatiivne. Muna
keetmine 2. pH HCl maos 3. Detergendid 4. Kaotroopsed ühendid (Guanidiinium ioon,
karbamiid )
Väiksemad valgud denatureeruvad reeglina pöörduvalt
Suuremad valgud kalduvad denatureerudes agregeeruma ja selline denaturatsioon on pöördumatu Valkude kokkupakkumise
kineetika Levinthali
paradoks : väikese valgu (ribonukleaas, 124 ah) puhul on võimalik ligikaudu 1050 erinevat konformatsiooni. Kui valk kulutaks iga konformatsiooni läbiproovimiseks
1013 sekundit, siis kuluks kõikide proovimiseks 1030 aastat. Tegelikult omandab ribonukleaas
funktsionaalse ruumilise struktuuri vähem kui 1 minuti jooksul.
Valkude kokkupakkumist uuritakse jälgides eelstatsionaarse faasi kineetikat. Nõuab reaktsiooni jälgimist esimese mõne millisekundi jooksul.
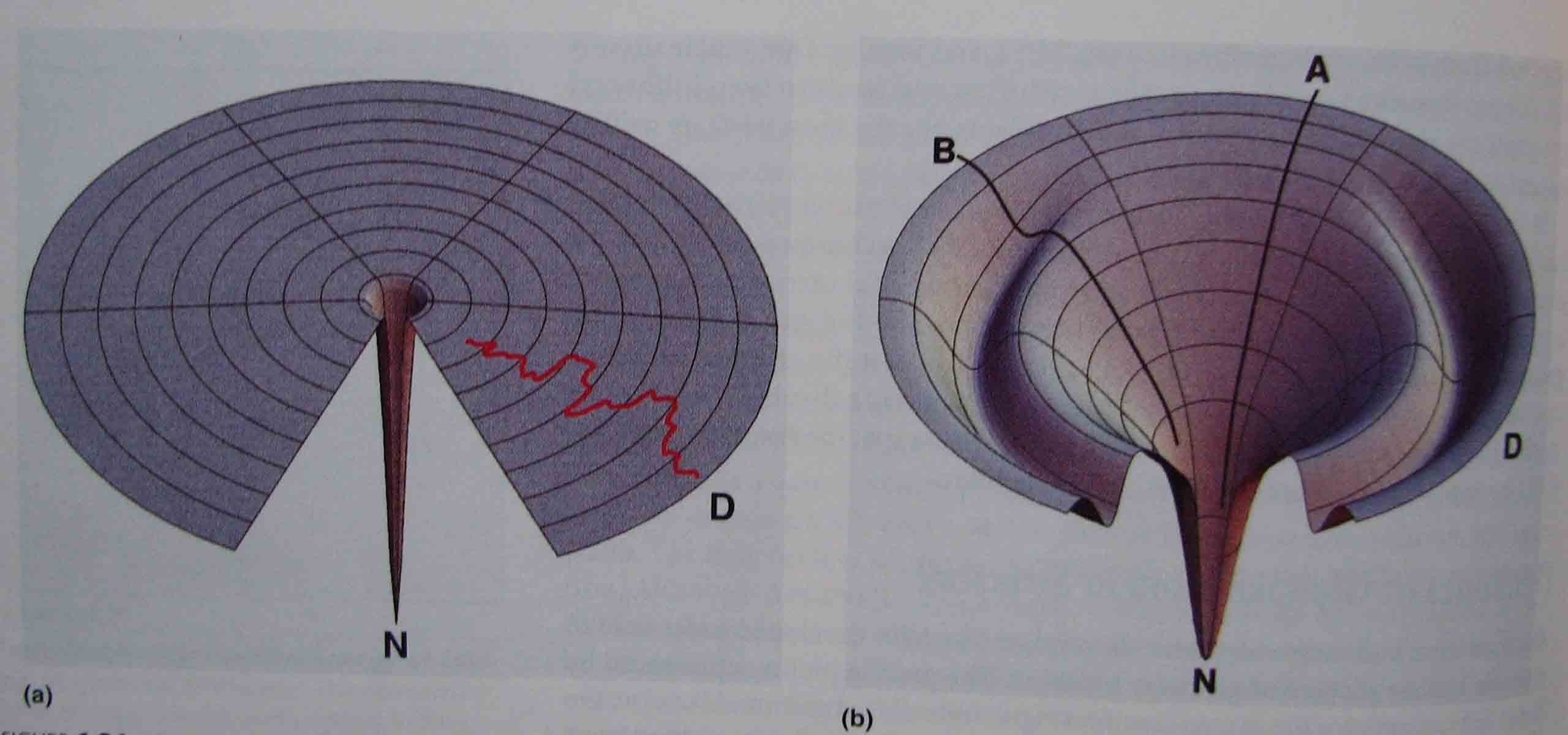
Levinthali golfiväljaku kujuline Lehtrikujuline energiapind
energiapind Valkude kokkupakkumine on kooperatiivne protsess
1. Lokaalsete sekundaarstruktuuride
formeerumine 5 ms
2. Tertsiaarstruktuuri moodustumine algab, subdomeenide teke , hüdrofoobne kollaps 1s
3. Domeenide lõplik moodustumine, H sidemed 5s
4. Multidomeense struktuuri moodustumine
Kokkupakkumine võib olla pidurdatud metastabiilsete vaheühendite tekke kaudu. Kokkupakkumist assisteerivad ensüümid: 1. Peptidüülprolüül isomeraas 2. Disulfiidi isomeraas 3. Saperonid (E.coli GroELGroES) Hemoglobiin ja müoglobiin
Hemoglobiin hapniku transportimine veres. Punaste vereliblede valk
Müoglobiin lihasrakkude valk
Mõlemad valgud sisaldavad prosteetilise rühmana heemi
Müoglobiin monomeerne valk, 1 heem
Hemoglobiin tetrameerne valk, 4 heemi
Kopsudes, kus hapniku partsiaalrõhk on kõrge,
seondub hemoglobiin hapnikuga: Hb + 4O2 Hb(O2)4
Lihasrakkudes, kus hapniku partsiaalrõhk on madal, vabaneb hapnik hemoglobiiniga kompleksist
Hb(O2)4 Hb + 4O2 Valkudes esinevad molekulaarsed liikumised
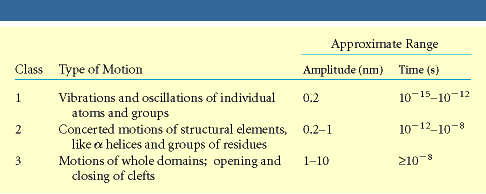
Valgud ei ole jäigad struktuurid
Valkudes võib rääkida kolme tüüpi molekulaarsetest liikumistest, mis erinevad liikumise
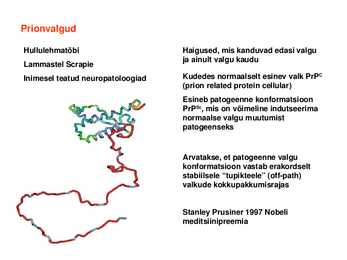
ulatuse ja kiiruse poolest Prionvalgud
Hullulehmatõbi Haigused, mis
kanduvad edasi valgu ja ainult valgu kaudu
Lammastel Scrapie
Inimesel teatud neuropatoloogiad Kudedes normaalselt esinev valk PrPC (
prion related
protein cellular) Esineb patogeenne konformatsioon PrPSc, mis on võimeline indutseerima normaalse valgu muutumist patogeenseks
Arvatakse, et patogeenne valgu konformatsioon vastab erakordselt stabiilsele "tupikteele" (offpath) valkude kokkupakkumisrajas
Stanley Prusiner 1997 Nobeli meditsiinipreemia Valkude kvaternaarstruktuur
Moodustub individuaalsete polüpeptiidahelate subühikute omavahelisel assotsiatsioonil
Subühikute assotsiatsioon enamast mittekovalentne
Monomeerne, oligomeerne, multimeerne valk
Dimeerne, trimeerne, tertameerne jne.
Kõige laiemalt on levinud dimeerid
Identsed või sarnased subühikud homo:
Homodimeer, homotrimeer jne.
Erinevad subühikud hetero:
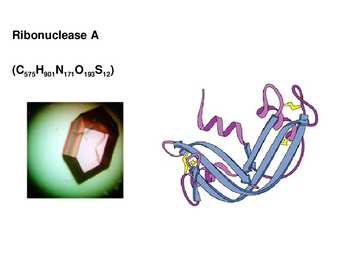
Heterodimeer, heterotrimeer jne. Ribonuclease A (C575H901N171O193S12)





































Kõik kommentaarid