AMINOHAPED
1. definitsioon
On karboksüülhapete
derivaadid , mis sissaldavad ühte
amiino - ja karbokspplrühma. Inimese kehas neid on 60.
inimkeha valgud ja
peptiidi koosnevad alfa-aminohpaetest.
2. Jaotus. 2.1 -
proteinogeensed kuuuvad valkude koostisse. Neid on 20 ja nendel on v'
hemalt üks kodoon inimese geneetilises
koodis . Neil on L-konfiguratsioon. Proteiinogeensete aminohapete derivaadid tekivad valgumolekulis juba olevate põhiaminohapete ensümaatilisel modifitseerimisel. (nt. Pro baasil tekib Hyp, Lys baasil Hyl.) Derivaate teke loob aluse valgu mingi spetsfunktsiooni täitmiseks. - Aproteingeensed ei kuulu valkude koostisee. Need on rakus vabalt või mittevalgulistes ühendites. Ornitiin, tsitrulliin, beeta-
alaniin , tauriin, homoseriin, betaiin. 2.2 -
Asendatavad inimkehas sünteesivad - Asendamatud inimkeha ei sünteesi neid ise ja peab nende vajamineva koguse saama toiduga ( Val, Ile, Leu, Thr, Trp, Lys, Met, Phe, His).
Asendamatuid AH-d vastavates
hulkades ja sobivates vahekordades sisaldavad näiteks muna, piim, juust ja liha.
Taimsed valgud (kaunviljad, pähklid, seemned) on sellised, kus puuduvad osad asendamatuid AH-d. Segatoidus
loomsed ja taimsed valgud täiendavad üksteist (soovitavalt 55%
loomseid ja 45% taimseid valke).
Sageli asendamatud AH
kirjutatakse asendatavate AH-ga paarides (Phe+Tyr; Met+Cys). S.t, et Tyr võib sünteesida
Phe baasil, kui Phe jääb üle. Kui Phe ei ole -> Tyr muutub asendamatuks.
Histidiin inimkeha ei süntees, normaalse söömisega sünteesivad His
seedekulgla mikroobid.
3. AH reaktsioonid 3.1 Dekarboksüülimine. CO2 ellimineerimine dekarboksülaasiga. Annab
biogeense amiini. (Glutamaat annab -aminobutüraadi (
GABA ); histidiin ->
histamiini ). Biogeensed
amiinid töötavad inimkehas
signaalmolekulina. 3.2 Desamiinimine. Aminorühma eraldamine molekulist ensüümi toimel. Annab vastaav ketohappe ja NH3.
3.3 Transamiinimine Aminorühma ülekandmine. Aminorühm
kogub glutamiinhappe molekulide vormis. Aminohappe alfa-aminorühm kantakse üle alfa- ketohappe alfa-süsinikule. Reaktsiooni käigus aminohappest tekib talle vastab ketohape, ketohappest ->
aminohape . Koensüümiks on püridoksaalfosfaat.
3.4 Peptiidsideme teke Ühe aminoghappe alfa-COOH interakteerub teise aminohappe alfa-NH2-ga, eraldyb veemolekul ja tekib
kovalentne peptiidside. Peptiidsideme süntees vajab energiat. Pep-side seob
aminohappejääke peptiidides ja valkudes.
PEPTIIDIDOn oligomeersed biomolekulid, nad koosnevad peptiidsidemega
seostunud aminohappejääkidest.
1.
Klassifikatsioon 1.1 Ahela pikkuse järgi oligopeptiidid 2-20 aminohappe jääki polüpeptiidid 21-50 jääke
kui on rohkem, sis on valk 1.2 Sünteesi koha järgi ribosomaalsed; mitteribosomaalsed; fragmenteeritud (pikk polüpeptiidne ahel inaktiivne. Proteaasiga tehakse lühemaks, mis siis omavad aktiivsust või ei oma.) 1.3 Ülesande järgi signaalmolekulid; antioksüdantid; antibiootikumid;
toksiinid .
Peptiidides esinevad minoorsed
haped või aproteinogeensed.
2. Biofunktsioonid antioksüdantsus pole toime suur, OH- radikaali püüdja puhverdusvõime eemaldavad laktaati, et atsidoosi ei tekeks kelateeriv toime antiglükosüülimine glõkoosi lisamine valgule, halb kui aktiivsele valgule glükoosi lisamisel muudetakse tema konformatsiooni ja funtsioon jääb täitmata (diabeet)
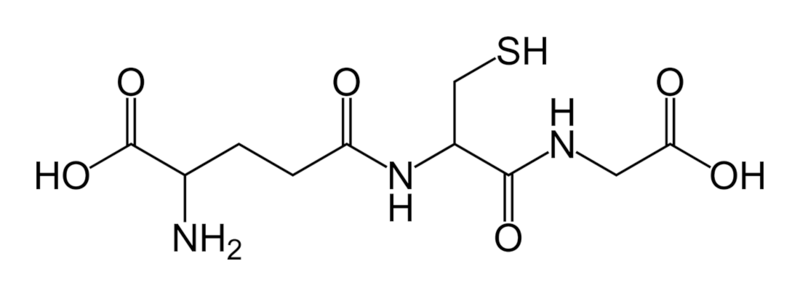
3. Glutatioon
glutamaat-tsüsteiin-glütsiin · sisaldab inimkeha keskset aminohapet (glutamaat ehk glutamiinhape, Glu) ·
ebatavaline -peptiidside (isopeptiidside) · reaktiivne tioolrühm (-SH) väävliaatomi elektronstruktuur on reaktiivsuse aluseks (redoksreaktisoon jt)
Glutatiooni biofunktsioonid
· antioksüdant
· stabiliseerib erütrotsüütide membraane
· kaitseb Hb denaturatsiooni eest
· kaitseb Hb oksüdeerimise eest (inhibeerib
metHb teket)
· mitmete ensüümide kofaktor
· reguleerib ensüümide tegevust (glutationüülimine)
· osaleb aminohapete
transpordis läbi membraani
· osaleb ksenobiootikumide detoksikatsiooniprotsessis
· taastab valkude tioolrühmi
· sünteesi põhikoht on maks
· sünteesi kiirust ja mahtu limiteerib tsüsteiini kättesaadavus/tase
· lisamanustamine on limiteeritud VALGUD
Valgud on
unikaalsed ja asendamatud
toitained , sest nad osalevad paljudes organismi füsioloogilistes protsessides
(ensüümid katalüüsivad reaktsioone, antikehad
kaitsevad organismi jne.). Valkude sünteesi
eelduseks on vabade
aminohapete olemasolu organismis. Kuna
aminohappeid peab olema nõutavates vahekordades ja piisavates hulkades
omandab õige toitumine erilise tähtsuse. Ööpäevas lammutab inimorganism umbes 400g kehavalke ja samapalju ka
sünteesitakse, et säiliks tasakaal. Kõige kiiremini uuenevad soole limaskesta valgud, samuti maksa, pankrease, neerude
ja vereplasma valgud. Aeglaselt asenduvad lihaste ja naha valgud.
Valgud on ühendid, mida iseloomustab: · Kõrgmolekulaarsus; · Koosnevad peptiidsidemega seotud aminohappejääkidest; · Spetsiifilisus (eri organismi valgud on erinevad) · Ahela
lineaarsus ; · Bioloogiline aktiivsus; · N sisaldus.
Klassifikatsioon koostise alusel: Lihtvalgud koosnevad ainult AH-jääkidest. Mida ühekülgsem on Ah koostis, seda spetsiifilisemat funktsiooni see valk täidab.
Liitvalgud koosnevad valgulisest ja mittevalgulisest osast. 1. valk + glükoos = glükoproteiin membraanides
retseptorvalgud , viirusvastane interferoon. 2. valk + nukleiinhape = nukleoproteiin kromosoomides ja ribosoomides. 3. valk +
pigment = kromoproteiin
heem ja klorofüll. 4. valk + fosfor = fosfoproteiin piimavalk
kaseiin . 5. valk +
lipiid = lipoproteiin biomembraanides ja verest lipiidide transportija. 6. valk +
metall = metalloproteiin liiteensüümid ja
transferiin (Fe transportiv valk).
1. Valkude ehitusprinsiibid: 1) Primaar- e esmane struktuur AH suhteline hulk ja järjestus polüpeptiidahelas, mis on geneetiliselt määratletud. On aluseks kõikide kõrgemat järku struktuuride moodustamisele. Siduvaks sidemeks on peptiidside, teised sidemed esinevad ebakorrapäraselt. Selles võib peituda geneetiline viga.
2) Sekundaarstruktuur esineb kahel kujul: · polüpeptiidahela kokkukeerdumisel spiraaliks. Stabiliseerivateks sidemeteks on molekulisisesed H-sidemed; · -struktuur esineb kahe variandina: a) üks polüpeptiidahel voltub mitmekordselt üksteise kohal kokku. Stabiliseerivateks sidemeteks on molekulisisesed H-sidemed; b) mitu polüpeptiidahelat voltuvad paralleelselt ja nad asuvad kohakuti. Stabiliseerivateks sidemeteks on molekulisisesed H-sidemed;
Sekundaarne struktuur esineb juustes, küüntes, sulgedes, ämblikuvõrgus, siidiniidis jne. Enamasti valgud ei ole 100% või -struktuuris, vaid üks neist vormidest esineb ülekaaluliselt.
3)
Tertsiaalstruktuur tekib polüpeptiidahela spetaiaalsel ruumilisel kokkupakkimisel kas: · Kerakujuliseks gloobuliks (
sagedasem ) · Niitjaks, ellipsoidseks fibrilliks. Stabiliseerivateks sidemeteks on: a) molekulisisesed H-sidemed; b)
ioonilised sidemed (erinevalt laetud radikaalide vastasmõju tekkinud) c) hüdrofoobsed sidemed (hüdrofoobsete AH radikaalide vastasmõju) d) S-S tüüpi sidemed, mis tekivad 2 tioolrühma vastasmõjul.
Tertsiaarne struktuur esineb: osa ensüüme,
histoonvalgud , albumiinid,
globuliinid ,
fibrinogeen , sidekoelised valgud. Kõik aminovalgud omavad 3o struktuuri.
4) Kvaternaarstuktuur mitmest polüpeptiidahelast tekkiv valgu
molekul , mis on struktuurselt ja funktsionaalselt terviklik. Ehitusüksusteks on: · Subühikud, s.o struktuurid, mis iseseisvalt bioloogilist aktiivsust ei oma; · Protomeeridest subühikud, millel on teatav iseseisev katalüütiline aktiivsus.
Annab uusi funktsioone valgule (peenregulatsioon).
Kui polümeerne valk on geneetilise veaga -> tervikkuna struktuur on vale, vaid molekulid vüivad osaliselt
funktsioneerida.
2.
Denaturatsioon ja
renaturatsioon Valkude denaturatsioon valkude kõrgemat järku struktuuride lagunemine (kuumutamisel, kiirgus,
vibratsioon ,
tugev alus/hape). Valk ei täita oma fuktsioone.
Valkude renaturatsioon kõrgemat järku struktuuride
taastumine . On võimalik, kui denatureeriv faktor ei olnud
tugev , toimis vähe aega, kui
primaarstruktuur on elav.
Valkude hüdrolüüs peptiidsidemete lõhkumine ehk primaarstruktuuri lagunemine (hapetes keetmisel). Inimese
maos toimub pepsiini abil HCl keskkonnas.
3. Individuaalsed valgud
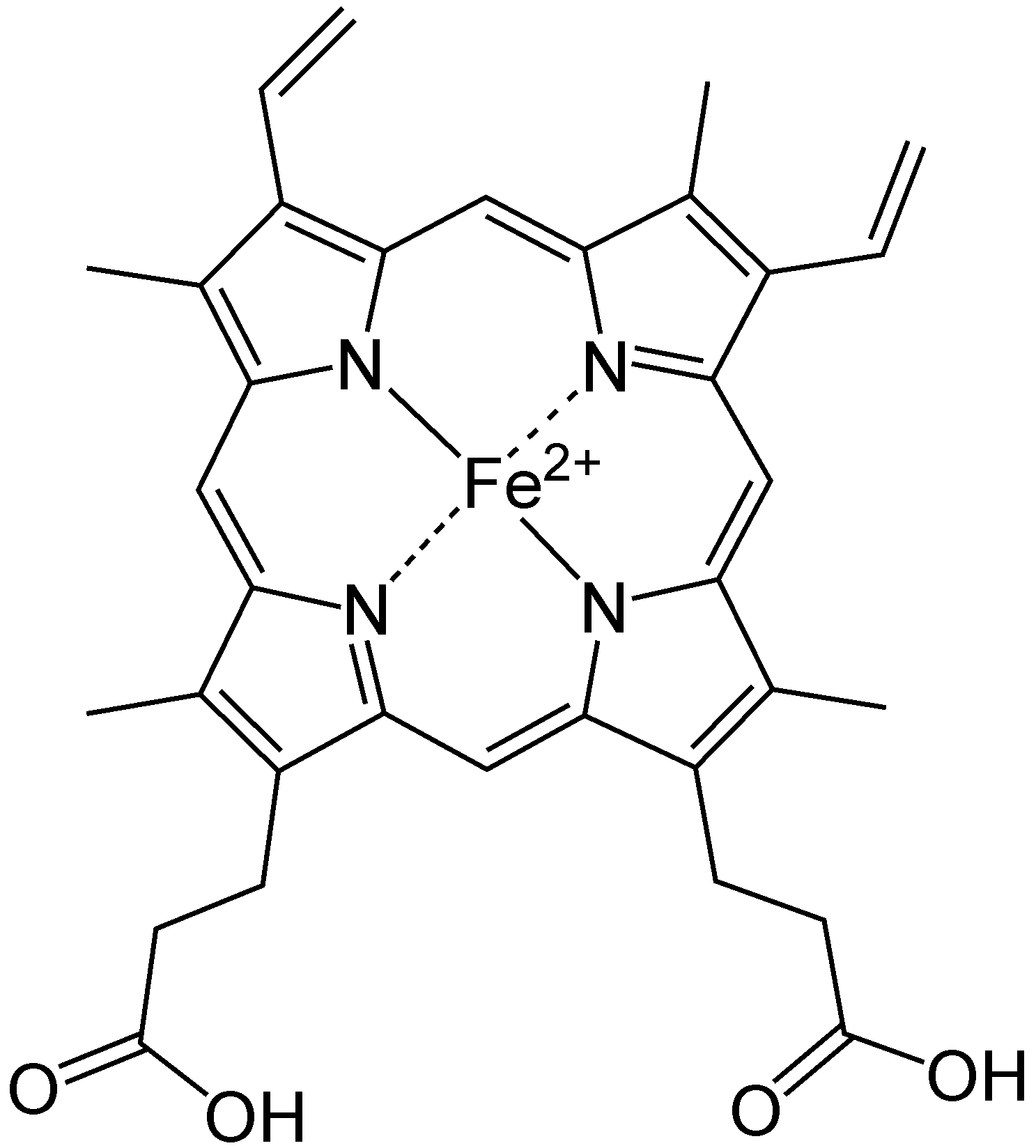
hemoglobiin ja müoglobiin.
Heem tähtsain komponent on Raud.
Ioniseeritud raud annab koordinatiivseid sidemeid. Üks
nendest tekib O2 vaba elektroonpaari osalusel ja sel viisil
seobki Hb transporditava molekulaarse O2.
3.1 Müoglobiin
sisaldub lihastes.
Omab tertsiaalse struktuuri (1 polüpeptiidahel).
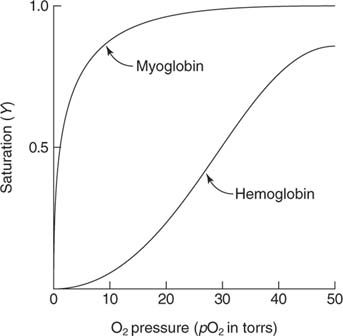
Mb omab 1 heemi grupp, 1 heem soib 1 hapnikumolekuli. Kuna
sidumise sõltuvus O2 partsiaalrõhust on
hüperboolne, suudab Mb intensiivselt töötavas
lihases madala osarõhu juures (5 mmHg) vabastada O2, et
lihasrakkude mitokondrite tsütokroomid toodaks ATP. Mb küllastub hapnikuga madala partsiaalrõhu juures.
Mb on ehituse tõttu O2 spetsiaalsalvestaja ja ajutine
depoo lihaskoe suure hapnikutarbe jaoks koomavate pingutuste
korral.
Mb ei saa töttada O2 transpordijana kopsudest kudedesse, sest 2+ mmHg tasemel ta hapniku ei loovuta. 3.2 Hemoglobiin
Sisaldub veres. Erütrotsüütides.
Kvaternaarne struktuur 2 alfa ja 2 beeta polüpeptiidi.
Koosneb globiinist ja 4 heemist.
Funktsioon O2 sidumine kopsukapillaarides ja transport kudedesse.
O2 sidumisel toimub SU kooperatiivsus, see tagab selle, et kopsukapillaaris O2 partsiaalrõhu üsna
kitsas vahemikus
toimuvast muutusest
piisab , et tagada Hb O2-ga küllastamine efektiivsus ja kiirus. Tänu kooperatiivsusele tagab vaid
4-5 kordne hapniku partsiaalrõhu langus koekapillassris O2 piisava vabanemise.
Kopsudes, kus hapniku partsiaalrõhk on kõrge,
seondub hemoglobiin hapnikuga:
Hb + 4O2 -> Hb(O 2) 4
Lihasrakkudes, kus hapniku partsiaalrõhk on madal, vabaneb hapnik hemoglobiiniga kompleksist
Hb(O 2) 4 -> Hb + 4O2
2,3-bisfosfoglütseraat. On Hb allosteeriline regulaator. BPG toodab erütrotsüütides anaeroobse glükolüüsi
kõrvaltee. BPG seostub desoksüHb-ga, kergendab O2 vabastamist -> vähendab Hb afiinsust hapnikule.
Hb Klassifikatsioon.
HbF loote Hb, ehitus 22, HbA täiskasvanute Hb, 22. HbF seob hapniku afiinsemalt, ei teki hüpoksiat.
Hb -desoksühemoglobiin
HbO2 oksühemoglobiin, arterites
HbCO karboksühemoglobiin (vingugaas + Hb). CO seostumine blokeerib O2 seostumise madalate
kontsentratsioonide juures.
MetHb methemoglobiin. Oksüdeeritud mittefunktsionaalne vorm (ei seo ega transporteeri O2), heemis on 3-
valentne raud. MetHb suure koguse korral
sureb org-m hapnikunälga. Kogust võivad tekitada sulfoonamiidid,
anilinvärvid, nitroühendid,
nitritid , endogeensed oksüdantid. MetHb teket takistavad vit C, glükoos.
4. Sidekoe valgud.
Kollageen ja
elastiin 4.1 Kollageenid annavad sidekoedele tugevust.
Struktuur.
primaarstruktuur antihelikaalsete aminohapete järjestus. (Gly, Pro, Hyp, Ala, Hyl). Iga kolmas jääk on
Gly ja polüpeptiidahelad koosnevad valdavalt korduvblokist (Gly-X-Y)n, kus X Pro ja Y Hyp /Hyl.
Jäägid on põhiAH-te derivaadid,
tekkivad põhiAH-te ensümaatilisel reaktsioonidel. (Pro-Hyp; Lys-Hyl)
Ensüümid vajavad: Cu - osaleb
kollageeni ja
elastiini formeerumisel koostöös
vitamiin C-ga, vit C defitsiidiga
kukuvad hambad välja, sest ei teki
sidekoes tugevat tuge kollageen O2 - Puudus võib tekkida
aneemia või vereringluse probleemide korral (
tromb ), ateroskleroos,
kui
koed ei saa piisavalt hapnikku. Siis ei teki kollageeni õige
strktuur alfa-
keto -glutaraat - Defitsiit võib tekkida, kui prevaleerib protolüüs ehk valgu lammutamine. See
võib esineda põletiku korral. Süntees jääb maha, tekib palju ammoniaaki, seda tuleb korjata ja detoksikeerida,
mistõttu jääb seda väheks.
Sekundaarstruktuur atüüpiline alfa-heeliks. Pole vesiniksidemeid, teda stabiliseerib Pro ja Hyp tuumade
repulsioon.
Tertsiaalstruktuur tropokollageen. (3 alfa-ahela paremkeerduvus). On väga tugev ja praktiliselt ei veni,
kuna naaberahelaid on seostunud H-, ioonsete-, hüdrofoobsete- ja kovalentsete ristsidemetega Lys jääkide vahel.
Kvaternaar mikrofibrill. Kllageensed SU-id on seovad kollageensed ristsidemed.
Kovalentne ristside tagab kollageeni tugevust. · Kollageeni poolväärtusperiood on keskmiselt aasta, ei ole kiiresti metaboliseeriv. Mida vanem on inimene, seda rohkem elab kollageeni molekul, seda rohkem
ristsidemeid tekib -> kollageen muutub tugevamaks aga ka hapremaks -> venitus väheneb.
· Glükosüülimine - süsvuvesiku liitumine valkude molekuli. Muutuvad kollageeni konformatsioon ja funktsioonid. Liigglükolüüsil areneb diabeet, ilmnevad sidekoe nõrkuse sümptomid.
4.2
Elastiinid annab suurte arterite seinte, kopsude, elastsete ligamentide, naha mitmesuunaline
venivus /kokkutõmbumine.
· Primaarstruktuuris ei ole korduvjärjestust. Gly rikkad osad vahlduvad Ala ja Lys lühilõikedega. · Nad ei sisalda Hyl ja süsivesikuid · On vaid üks geneetiline tüüp
Ehitus.
4 polüpeptiidseid ahelaid. Enmasti sisaldab Gly, Ala, Pro, Lys ja Hyp.
Elastiinid sünteesitakse tropoelastiini monomeeridena, milles Pro hõdroksülaas tekitab Hyp. Tropoelastiin on
ebakorrapäraselt keerdunud struktuur, mis võimaldab venimist. Tropoelastiini rakust tekitab Lys oksüdaas
lüsiinijääkidest tsüklilisi desmosiinseid ristsidemeid. Võrkjas desmosiin tagab elastiinide suure elastsuse ning
venitavuse.
Elastiini struktuuri
tekkes võtavad osa samad
faktorid , nagu kollageeni kohta.
5. Lihaskontraktsiooni valgud · Lihased 40...50% inimese kehamassist. · Lihased muundavad ATP ja kreatiinfosfaadi keemilise energia mehhaaniliseks energiaks (tööks) ja soojuseks. · Selles tegevuses on
kesksed lihaste põhivalgud
aktiin ja müosiin. · See tähendab, et kontraktiilsed valgud tagavad skeletilihaste, silelihaste ning südamelihaste töö.
5.1 Aktiin · peened heterogeensed · globulaarsed SU polümeriseeruvad kaksikhelikaalseks F-aktiiniks ( Mg, ATP) · F-aktiin ühineb fibrillaarse tropomüosiini (T M) ja globulaarse troponiiniga (Tn) aktiinfilamendiks · TM tugevdab filamenti · regulatoorne Tn muudab aktiinfilamendi kaltsiumtundlikuks. Tn-C (kaltsiumsiduv SU), Tn-I (inhibeeriv, aktiiniga seostuv SU) ja Tn-T (tropomüosiiniga seostuv SU)
Vajab Mg ja ATP-d. Moodustatakse pikk
filament ehk F-aktiin. 2 polüpeptiidahelat (2 F-aktiini) moodustavad
aktiinifilamendi.
5.2 Müosiin
Müosiinmolekuli 6 alfa-hlikaalset polüpeptiidset raskahelat keerduvad ümber üksteise moodustades
müosiinimolekuli saba. Müosiinipeakestel on ATPaasne aktiivsus (seovad ja lõhustavad ATP) ning aktiini ja
regulatoorsete kergahelate sidumisalad.
Hing lubab müosiinipeakestel saba suhtes teatud nurga ulatuses painduda, mis on lihaskontraktsiooniks hädavajalik. ATP roll: · ATP on kontraktsiooni vahetu
energiaallikas : tema hüdrolüütilise lõhustumise energiamuut kasutub müosiinipeakeste liikumiseks (keemiline energia konverteerub mehhaaniliseks) · ATP hüdrolüütilise lõhustumise energiamuudu arvel eemaldab Ca-
pump sarkoplasmast kaltsiumiioone (vastu kontsentratsioonigradienti)
6. Vereplasma valgud
Vereplasma proteinogramm elektroforeesi abil eraldatud valgud.
Jaotus: · albumiinid · globuliinid ( 1, 2, , )
Vereseerumis:
Prealbumiin
Albumiin 1-globuliinid (nt. 1-antiproteaas, 1-happeline glükoproteiin, -lipoproteiin)
2-globuliinid (nt. 2-makroglobuliin, haptoglobuliin, tseruloplasmiin, CRP)
-globuliinid (nt. transferriin, C3-komplementfaktor, -lipoproteiin)
-globuliinid (nt. IgG, IgA, IgM, IgD, IgE)
( vereplasmas oleks ka fibrinogeen)
Veres on albumiine rohkem. Väga kiiresti metaboliseeruvad valgud. Verevalkude järgi saab leida, mis organite
funktsioonid on nõrgenenud või häirunud.
Valgu koefitsient albumiinide ja globuliinide suhe (1,5-2,5)
Valgufraktsioonide väärtused seerumis
Prealbumiin: 0,1-0,42 g/l
Albumiin: 34-50 g/l
Funktsioonid: · kaitsefunktsioon (immuuboglobuliinid) · ensümaatiline funktsioon · (nõrk)
puhver , pH aäilitamine · proteaaside inhibeerimne (hoiab aktiivsust tasakaalus, väldib nende liigset valkude lõhustamist) · antioksüdatiivne funktsioon · transport- albumiinid.
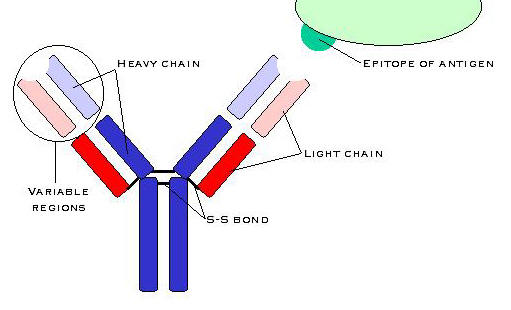
-globuliine struktuur.
Molekulil on kaks kergahelat (L-ahel) ja kaks raskahelat (H-ahel). Molekuli stabiliseerivad ahelatevahelised
disulfiidsillad ja hüdrofoobsed sidemed.
Ahelatel on varieeruvad ja konstantsed regioonid. Fab on antigeeni sidumise
regioon, Fc interakteerub immuunsüsteemi teise konplemendiga. ,,Hing" lubab raskahelatel igas suunas käänduda.
Di-, tri- ja pentameersed vormid koosnevad 2,3 ja 5-st monomeerist ja seondavast ahelast.




























Kõik kommentaarid