Kullman, B. (2002). Horisontaalsest geeniülekandest seeneriigis. In: I. Puura & T. Teder (toim), Võrkude teooria.
Schola Biotheoretica 28: 21-33. Tartu, Sulemees. 88 lk. ( ISBN 9989-9278-6-9)
Horisontaalsest geeniülekandest seeneriigis
Bellis Kullman
Intratsellulaarse sümbiogeneesi käigus on enamus protomitokondri geene transformeerunud peremees –tuuma
horisontaalse geeniülekande (HGT) käigus. Võrreldes erinevate seente mitokondriaalseid genoome (mtGenoom),
on jälgitav kodeerivate järjestuste asendumine mittekodeerivatega kuni nende kadumiseni. Pärmirakus ei muutu
mtGenoom ja tuumagenoom üksteisest sõltumatult. Võimalik, et HGT mitokondri ja tuuma vahel on siiani
käimasolev protsess.
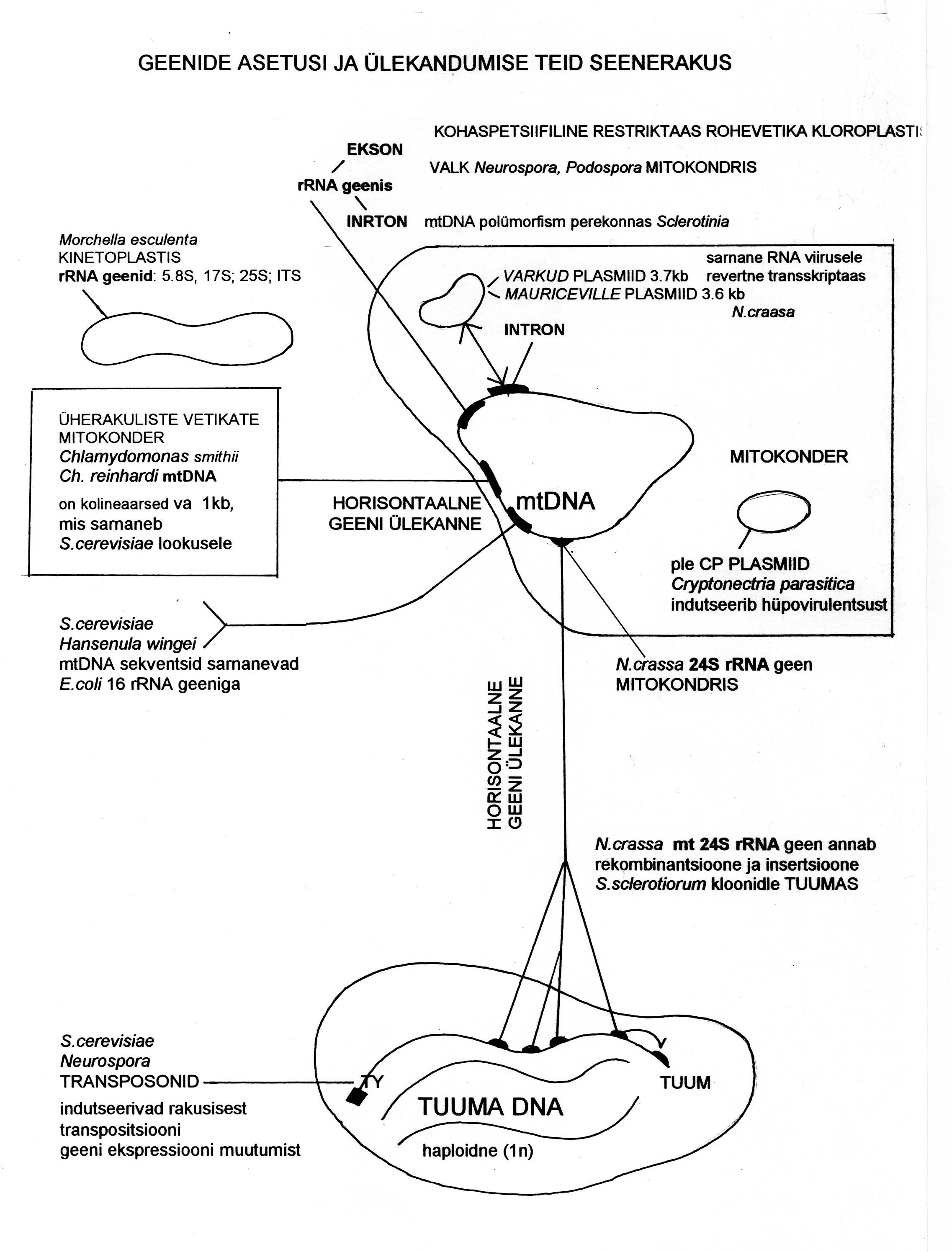
Seenerakus esineb palju viirusi ja plasmiide, mis võivad osaleda geenide ülekandel erinevate
organismide vahel. Plasmiid võib olla mtGenoomi koosseisus või iseseisvalt paljuneda ja omada kindlaid
funktsioone (raku kasvu pidurdamine , vananemine, plasmiidis kodeeritud mürkide abil konkurentide
surmamine). Neid geene antakse edasi tsütoplasmaga seenehüüfide kokkukasvamisel (heteroplasmas). On
kirjeldatud HGT bakterilt seenele, seenelt seenele (eri liikide või sama liigi eri isendite mtGenoomide vahel,
seeneperekondade või ühe perekonna liikide tuumagenoomide vahel) ja seenelt taimele (sümbiontse või
parasiitsete seene mtGenoomist taime mtGenoomi). HGT on leitud rDNA I grupi intronite osas ja mitmete valke
kodeerivate järjestuste osas.
Nii pärmidel kui hüüfe moodustavatel seentel on leitud mitootiline geenisiire, mis on sisuliselt sama,
mis bakterite konjugatsioonil toimuv HGT. Kui haploidne tuum siseneb teise hüüfi, võib ta liikuda mitootiliselt
jagunedes, säilitades oma intaktsuse. Selliselt tekkivad dikarüootsete hüüfide tuumad võivad olla küllaltki
erinevad ( aneuploidsus , kromosoomi pikkuse polümorfism ). Tuumade liitumisel (konjugatsioonil? või/ja
kopulatsioonil) võib toimuda geenisiire sarnaselt bakteritega (HGT) või ka nii nagu tõelises meioosis
(genealoogiline, vertikaalne geenisiire - VGT). Seente tuumade jagunemisviiside ebastabiilsus ja mitmekesisus
võib olla põhjuseks, miks nii mõnelgi juhul pärilikkuse ‘võrku’ on ‘puust’ üpriski raske eristada. Kas ka
mitokondril on olnud roll homoloogiliste kromosoomide evolutsioonis , jääb mõistatuseks.
Viimasel ajal on kogunenud üha enam andmeid horisontaalse geeniülekande (HGT – horizontal gene transfer )
kohta seeneriigis. Nii leivapärmseene kui ka teiste seente mitokondriaalne genoom (mtGenoom) on tugevalt
rekombinantne. Geneetilisi muutusi ja rekombinatsioone on kindlaks tehtud nii kunstlikul ristamisel ( Fincham ,
1979), kui ka looduslikes populatsioonides ( Saville , 1998). On isegi leitud, et klonaalselt paljunevatel seentel on
raku tuumagenoomi muutumisele kaasnenud mtGenoomi rekombinantne (retikulaarne) muutumine ( Anderson ,
2001). Teadaolevalt paljunevad mitokondrid inimesel jt. imetajatel klonaalselt ning nende järgi on võimalik
määrata päritolu ‘mitokondriaalsest Evast ’. On esitatud küll ka vastupidiseid seisukohi (Awadalla, 1999), mis
aga on osutunud ekslikeks andmete tõlgenduseks ( Kivisild & Villems, 2000).
Arvatakse, et HGT ja endosümbioos on mänginud rolli nii valkude kui DNA mosaiiksuse tekkel ( Nara
et al., 2000). Tõenäoliselt on HGT toimunud bakterite eellaste ja eukarüootide eellaste vahel väga varasest
organismide arengust alates mõlemas suunas ning võib toimuda mõnedes organismides tänapäevalgi. On
andmeid, et on toimunud HGT bakterite eellastelt seente eellastele (Brinkmann et al., 2001). Varakult pärast
Archaea ja eukarüootide divergeerumist võis aga toimuda vastupidine paljude geenide ülekanne eukarüootidelt
bakteritele (Brown et al., 1999). Hilisemast ajast on teada HGT bakteritelt seentele ( eukarüoodid ). Näiteks
pärineb seene Orpinomyces joyonii tselluloosi lagundamisel tähtsat rolli omav geen (celA) bakterilt
Fibrobacter succinogenes. See võimaldab seenel elada uues toitekeskkonnas, herbivoorsete imetajate vatsas
( Garcia -Vallve et al., 2000).
HGT prokarüootidelt eukarüootidele avastati 1990 aastal, kui võrreldi isopenitsilliin-N süntetaasi
(IPNS) geenide ja 5S rRNA geenide vastavaid evolutsioonilisi distantse. Analüüsid näitasid veenvalt , et IPNS
geen oli kahel korral horisontaalselt üle kantud bakteritelt hüüfe moodustavatele kottseentele (Panalva et al.,
1990).
Kuigi on teda, et DNA transport läbi membraanide ja rakkude vahel toimub, ei teata selle toimumise
mehhanismi. Prokarüootidel toimub HGT peamiselt bakteriaalse konjugatsiooni käigus. Bakteritel on leitud
paljunemise ajal konjugatsiooni protsessis toimiv integraalne DNAd siduv valk TrwB (Escherichia coli).
Arvatakse, et sellel valgul võib olla roll geeni ülekandmisel bakterilt taimele ning bakterilt seenele (Comis- Ruth
et al., 2001).
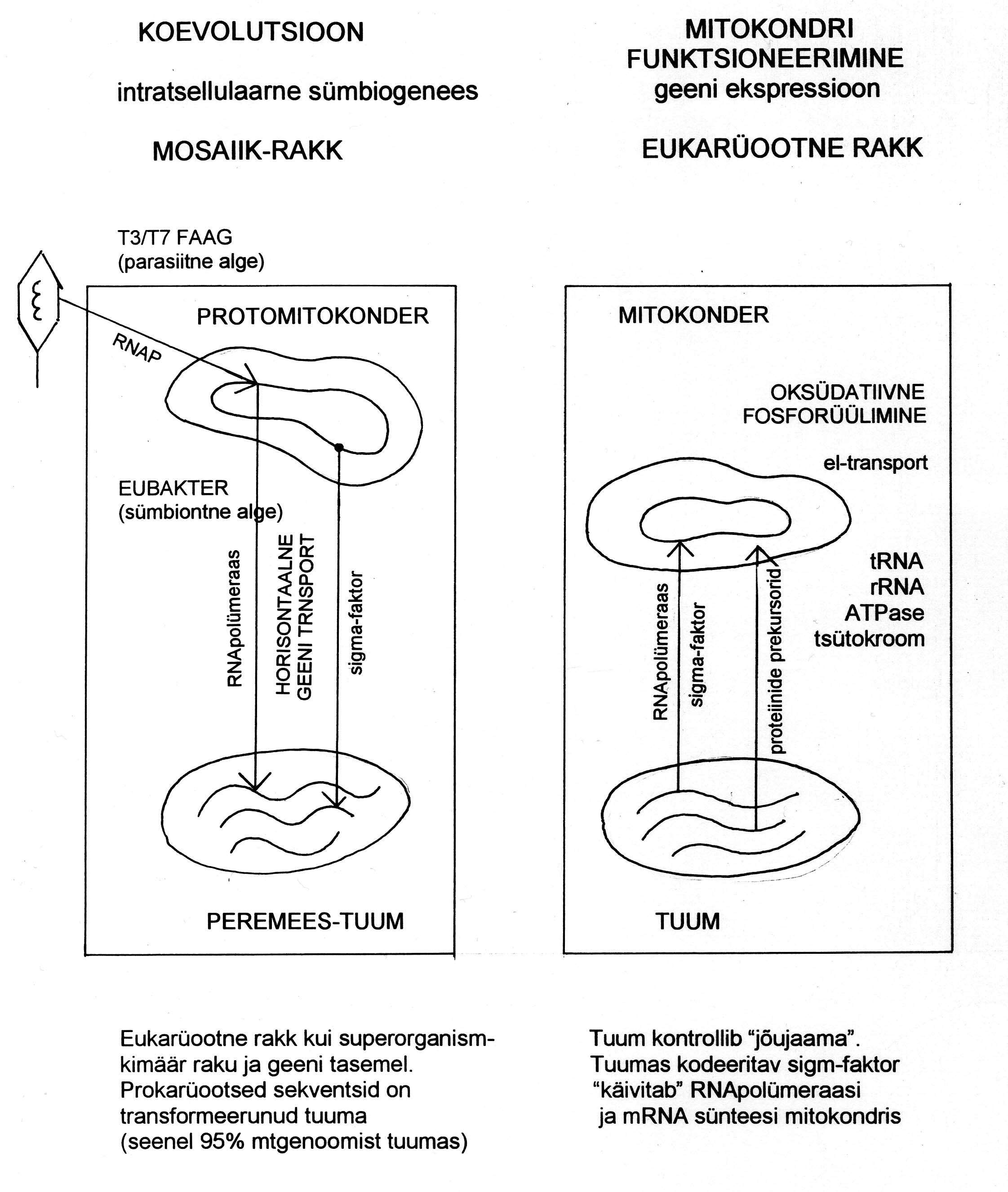
Joonis 1. Tuuma ja mitokondri koevolutsioon (vasakul). Eukarüootses rakus reguleerib tuum energia tootmist
mitokondris (paremal).
On andmeid ka eükarüootide omavahelise HGT kohta. Nii on leitud HGT seenelt taimele
parasiitsete (Kim et al., 2000) ja sümbiontsete seente puhul (Voughn et al., 1995), kes elavad tihedas kontaktis
oma peremeestaimedega. Tegelikult on tegemist tsütoplasmaatilise pärilikkuse muutumisega. Nimelt on HGT
toimunud seene mitokondrist taime mitokondrisse. Mitokonder on rõngasja kromosoomiga rakuorganell.
Mitokondri mtGenoom on haploidne, seetõttu kasutatakse tema kohta ka terminit haplotüüp. (Käesolevas töös
seda ei kasutata kuna hüüfe moodustavate seente tuumad on samuti valdavalt haploidsed). Mitokondril arvatakse
olevat eubakteriaalne päritolu (joonis 1). Kui oletada, et bakteriaalse päritoluga mitokondrid on säilitanud
bakteritele omaseid paljunemisviise, siis võib neil HGT aset leida konjugatsiooni käigus heteroplasmas
(piirkondades, kus peremeestaime ja seene tsütoplasmad kokku puutuvad). Õistaimel Peperoma polybotrya on
mitokondri coxI geenis ekson ja intron , milledel on selgelt erinev evolutsiooniline päritolu. Eksoni DNA
järjestus on lähedane teiste õistaimede sama geeni vastavatele järjestusele, intronit aga teistel soontaimedel pole
leitud. Selle introni järjestus sarnaneb rohkem seente kui sammaltaime, helviku Marchantia või rohevetika
Prototheca mitokondri vastavale geenijärjestusele. Arvatakse, et coxI introni doonoriks võis olla mingi
sümbiontne mükoriisaseen (Voughn, et al., 1995).
Viimase aastakümne jooksul on neid uusi meetodeid rakendades tehtud jahmatavaid avastusi, mille hulka kuulub ka seente liikidevahelise hübridisatsiooni võime (Schardl & Craven, 2003). Seeneriigis valitsevad haploidid, vahel küll dikarüootsel viisil. Kui inimesel saavad ema munarakk ja isa seemnerakk kokku, siis seeneeosed, munaraku ja seemneraku ekvivalendid, idanevad eraldi hüüfideks. Tavaliselt peetakse seeneks selle viljakeha, tegelikult on seen ise aga maa all ja ajab seal oma niidistikku laiali. Nõiaringis kasvavad seened on tegelikult kõik üks ja sama seen, selle erinevad viljakehad. Nagu õunapuu ja õunad. Teineteise äratundmiseks võivad kaks ühetuumsete rakkudega hüüfi kasutada feromoone. Seejärel võib toimuda midagi sellist, mida me oleme harjunud seksiks pidama. Kui hüüfide tuumadel on erinev nn paarumisfaktor, siis moodustatakse nende vahele torujas kanal, mille
Põhjustavad eranditult kottseened. Aeglane protsess. Võib toimuda ekstreemsetes tingimustes: külmas, kuivas, liigniiskes. Kahjustab vanu puitesemeid. Puidukaitsevahendid ei aita. Majaseente levikut ja arengut soodustavad tingimused. Kindel temperatuur ja niiskus. Majaseened toituvad puidus ja puidutoodetes leiduvates ainetest: tselluloosist, hemitselluloosist ja ligniinist (olenevalt liigist). 4. Mükoriisa Mükoriisaks nimetatakse seente ja taimejuurte kooselulisi vorme, kus seen ja taim on vastastikku kasulikes suhetes. Seenjuur on peamine organ, mille abil taimed hangivad toitaineid ja vett. Ehitus: Ainete vahetuseks seene ja taime vahel tekivad taimejuure- ja seenerakkude ühisstruktuurid. Tüübid: Arbuskulaarne mükoriisa (AM) - vanim mükoriisa tüüp. Seda moodustavad kõik ikkesseened. Taimedest esineb see põhiliselt rohttaimedel. Iiseloomulikuks tunnuseks on põõsasjalt harunenud seenehüüfide esinemine taimejuure rakkudes. Need tekivad juure
Genoomika kursuse kordamispunktid 1. Mis on ja mida uurib genoomika? Genoomika - teadus genoomide ehitusest. Genoomika uurib põhjusi, miks konkreetne DNA järjestus on evolutsioonis välja valitud (säilunud). Genoomika ülesandeks on mitte ainult teada konkreetse geeni ja selle produkti funktsiooni organismis, vaid ka kõikide geenide, nende produktide, funktsioonide ja regulatsiooni seoseid, mis viivad organismi tekkeni. GENes and chromosOMEs (kromosoomide täielik kogu koos neis sisalduvate geenidega). Genoomika on geneetika edasiarendus tegeledes: Genoomikaartide ja ülesehitusega, DNA sekveneerimisprobleemidega, Andmete säilitamise ja töötlemisega (bioinformaatika), Geenide identifitseerimisega, Funktsionaalse analüüsiga (funktsionaalne genoomika), Genoomide evolutsiooniga, Farmakogeneetiliste probleemidega jne. 3. Genoomika suundumused ja probleemid. Post-genoomika e. modulaarne bioloogia: Genoomide tasemel - Molekulaarne fülogenees, võrdlev genoomi
Mida tähendab fingerprintimine. See on mikrosatelliitse kordus DNA määramine. Mikrosatelliidid on kiiresti muteeruvad mittekodeerivad DNA lõigud, mis koosnevad tandeemselt korduvatest nukleiididest. Kuna nad muteeruvad suhteliselt kiiresti, siis populatsioonid erinevad korduste arvu poolest. Esmalt koguti üle Aafrika eri elevantide fecest, eraldati sealt DNA ja määrati referents proovid. Seejärel eraldati spetsiaalse meetodiga salakaubana konfiskeeritud elevandiluust DNA ja võrreldi mikrosatellitide korduseid referntsiga. Genotüüp = organismi geenide (alleelide) kogum Fenotüüp = organismi tunnuste kogum Monohübriidne = kahe erineva homosügoodi ristamine (erinevad tunnused) Vastastikune ristamine (retsiprookne) = tunnused vahetatakse ristamiseks erinevatel sugupooltel (kui tulemus ei muutu, siis tunnus ei ole seotud sooga) Dihübriidne ristamine = ristatakse kahe tunnuse suhtes erinevaid homosügoote F1 x F1 ristamine: Mendel avastas, et tunnus, mis ei avaldunud I põlvko
Handbook of Meat Processing Handbook of Meat Processing Fidel Toldrá EDITOR A John Wiley & Sons, Inc., Publication Edition first published 2010 © 2010 Blackwell Publishing Blackwell Publishing was acquired by John Wiley & Sons in February 2007. Blackwell’s publishing program has been merged with Wiley’s global Scientific, Technical, and Medical business to form Wiley-Blackwell. Editorial Office 2121 State Avenue, Ames, Iowa 50014-8300, USA For details of our global editorial offices, for customer services, and for information about how to apply for permission to reuse the copyright material in this book, please see our website at www.wiley.com/ wiley-blackwell. Authorization to photocopy items for internal or personal use, or the internal or personal use of specific clients, is granted by Blackwell Publishing, provided that the base fee is paid directly to the Copyright Clearance Center, 222 Rosewood Drive, Danvers, MA 01923. F
Geenitehnoloogia kordamisküsimused 1.Suhkrute lühiiseloomustus Suhkrud (süsivesikud)- orgaanilised ühendid, mille koostisesse kuuluvad süsinik (C), vesinik (H) ja hapnik (O). Suhkruid jagatakse 3 rühma: 1)Monosahhariidid (lihtsuhkrud) (üks tsükkel)- kõige lihtsamad süsivesikud, mis koosnevad 3-6 süsinikuaatomist. Tähtsamad neist on: 1. 5-süsinikuga e pentoosid · riboos (C5H10O5)- kuulub RNA (nukleotiidi) koostisesse. · desoksüriboos (C5H10O4)- kuulub DNA (nukleotiidi) koostisesse. 2. 6-süsinikuga (heksoosid) i. glükoos (viinamarjasuhkur) (C6H12O6)- tähtis energiallikas. Taimedes moodustub glükoos fotosünteesi käigus ja tihti talletatakse ,see tärklisena. Loomad saavad glükoosi toiduga nt tärklise lõhustamisel seedeelundkonnas. ii. Fruktoos (puuviljasuhkur )(C6H12O6)- puuviljades ja mees esinev monosahhariid. Seda
Geneetika 2 kordamisküsimused Lisaks tekstile ja õpikule vaadake kindlasti ka materjali slaididelt. 1. Võrrelge lüütilq aaise ja mõõduka bakteriofaagi paljunemistsüklit VIRULENTSED FAAGID – põhjustavad peremeesraku surma MÕÕDUKAD FAAGID – võivad püsida rakus ilma seda hävitamata o Lüütiline ja lüsogeenne fvgvb89htsükkel. Lüsogeenne tsükkel võib keskkonnatingimuste muutudes üle minna lüütiliseks tsükliks Lüütiline: kinnitub peremeesrakule antiretseptori vahendusel; genoom sisestatakse rakku, tehakse palju DNA/RNA koopiaid, viiruspartiklid pannakse kokku, rakk lüüsitakse Lüsogeenne: kinnitub peremeesrakule antiretseptori vahendusel; genoom sisestatakse rakku, genoom integreerub peremehe genoomi ja kandub kromosoomi koostisosana tütarrakkudesse. Keskkonnatingimuste muutudes võib lüsogeenne faag minna üle lüütilisse tsüklisse, mille käigus sünteesitakse viiruse partikleid ning pannakse need kokku. Lõpuks rakk lü
1. PPT 1. Geeniused ja geenid geen (ingl. Gene): Spetsiifilise bioloogilise funktsiooniga geneetiline determinant. Pärilikkuse ühik, mis asub kromosoomi kindlas punktis (lookuses). DNA segment, mis kodeerib mingit kindlat RNA-d ja mRNA kaudu kindlat polüpeptiidi ning mida saab eksperimentaalselt eristada cis- trans- või komplementatsioonitestiga. Geenius: harukordselt andekas inimene, suurvaim. Geenius on see, kes on suutnud oma päriliku potentsiaali ideaalselt hästi realiseerida. 2. Autismi geneetiline alus autism (ingl. Autism) Endassesulgumus, lapsepõlves ilmnev psüühikahäire, esineb ka täiskasvanuil. Põhjuseid otsitakse geenidest, sünniprotsessist, loote- kui ka beebieast. Milles ollakse kindlad on see, et antud häiretel on bioloogiline alus ning et lastevanemate kasvatusmeetodid ei põhjusta lapsel seda häiret. Erinevate uuringute järgi ühemunakaksikutel 60-
























Kõik kommentaarid