said immuunsuse. Marutaudi võitmine teeb Pasteurist pühaku. 1888. aastal loodi Pasteuri instituut ja kuus aastat hiljem maetakse Pasteur tema enda loodud meditsiinipühamusse. R. Koch avastas vaktsiini Siberi katkule, tuberkuloosile ja koolerale. Bakterid on prokarüoodid ehk eeltuumsed. Bakteritel puudub tuum ja teised membraaniga ümbritsetud organellid. vibureid võib olla ka mitu Bakteril puudub tuum Rõngas DNA bakteri kromosoom genoom Bakteris peavad olema ka ribosoomid (valgusüntees). Gaasivakuoolid asuvad samuti bakteris, reguleerib kas bakter vajub põhja või tõuseb pinnale, toimib nagu kala ujupõis. Plasmiid väikerõngas DNA. Plasmiidis on geenid mis võimaldavad bakteril elada ekstreemsetes tingimustes. Ilma plastiidita (antibiootikumid, kõrge temperatuur, kõrge rõhk, happeline, aluseline keskkond). bakter on ilma plastiidita elujõuline Plasmiide võib olla ka mitu.
b) Mõnede bakterite kest on kaetud karvakestega, mis on analoogsed imetajate karvadega VÄÄR Mõnedel bakteritel ümbritseb rakukesta kaitsev limakest ehk kapsel. c) Bakterite patogeensus tuleneb nende poolt ümbritsevasse keskkonda eraldatavatest toksiinidest TÕENE 5. Mida sa tead limakapslist? Bakterid koosnevad limakapslist, see kaitseb rakku kuivamise eest, seob üksikud rakud kolooniaks ja hõlbustab liikumist 6. Mida tead bakteris leiduvast kromosoomist? Rakutuuma asendab tuumapiirkond, milles paikneb rõngjas kromosoom. Kõigil bakteritel on vaid üks kromosoom, mille geenide arv ulatub kuue tuhandeni. Enne bakteri jagunemist rõngaskromosoom kahekordistub. 7. Joonista bakteri skeem koos nimetustega. 8. Bakterid paljunevad: a) pungumisega, b) eostega, c) pooldumisega, d) mütseeli tükikeste abil, c) paljunemisrakukestega. 9. Paljunemise etapid: 1) Kromosoom kordistub
multiresitsentsusele. Siiski ei ole faagiresistentsuse tekkimine täielikult välistatud. Bakterid võivad muutuda faagiresistentseteks näiteks siis, kui neil tekivad mutatsioonid vastavates retseptorites, millega faag kinnitub bakteriraku pinnale. Võib ka juhtuda, et need retseptorid kaovad mutatsioonide toimel täielikult ära. Faagiresistentsus võib tekkida ka siis, kui bakter omandab restriktsiooni-modifikatsiooni süsteemi või kui bakteris tekib CRISPR järjestuste blokeerimisel adaptiivne immuunsus. Mõlemal juhul omandab bakter võime lagundada sisse tunginud faagi DNA-d (Nobrega et al., 2015). Siiski ei ole faagiresistentsus nii ulatuslik kui antibiootikumide resistentsus. Faagid üldiselt ei ületa liikide vahelisi piire ning kuna faagid mõjuvad spetsiifiliselt üksikutele bakteritüvedele, siis tekib faagiresistentsus vaid üksikul tüvel, mitte tervel liigil või sugukonnal (Carlton, 1999).
BAKTERID 1) Bakteri rakuosad/ehitus/osade ülesanded. - Bakteril ei ole tuuma. eeltuumse raku sisemuses puuduvad membraanidest koosnevad rakustruktuurid ja nendega ümbritsetud organellid, mille tõttu pole bakteritel tsütoplasmavõrgustikku, Golgi kompleksi, kloroplaste ega mitokondreid. Kapsel: Sisaldis: Tsütoplasma: ringleb bakteris vähem kui päristuumses rakus ja ainete kandumine ühest raku osast teisi toimub valdavalt difusiooni teel. Tuumapiirkond: seal paikneb rõnasjas kromosoom, mis koosneb ühest DNA molekulist. Ribosoomid: valgusüntees. Plasmiid: sisaldavad geene, mis on vajalikud bakteri kasvukeskkonna eripärast tulenevate ensüümide sünteesiks. Aitab lagundada ümbritsevas keskonnas leiduvaid orgaanilisi aineid. Vajalik toitumiseks. Viburid: nende abil liiguvad.
ristvaheseina sissekasvamise teel), mis algab sellest, et mikroob kasvab oma pärilikult määratud pikkuseni. Rakkude jagunemise alustamiseks on vajalik valk FtsZ. FtsZ (filamentous temperature sensitive) valk on eukarüootide tubuliiniga homoloogne valk, mis rakkude jagunemisel moodustab algul rõnga jagunemiskoha ümber ja see rõngas tõmbub hiljem kokku, aidates tütarrakkudel eralduda. Pooldumise eel toimub DNA replikatsioon, mille tulemusena moodustub bakteris kaks rõngaskromosoomi. Järgnevalt sünteesitakse raku keskossa rakumembraanid.. Protsess lõpeb raku keskele vaheseina tekkimisega, mille tulemusena moodustuvad kaks uut tütarrakku. Need võivad jääda üksteisega veel mõneks ajaks seotuks, moodustades erineva pikkusega ahelaid Koos tsütoplasma jagunemisega kaheks jaotuvad selles paiknevad rakuorganellid tütarrakkude vahel. Moodustunud tütarrakud on geneetiliselt samased lähterakuga
• Bakterite patogeensus tuleneb nende poolt ümbritsevasse keskkonda eraldatavatest toksiinidest - Bakterid, mis põhjustavad haigusi inimese organismi sattudes, nimetatakse patogeenseteks bakteriteks. Enamus neist moodustavad mürkaineid ehk toksiine. 5. Mida sa tead limakapslist? Limakapsel aitab mõningatel bakteritel neid kaitsta ning hõlbustab nende liikumist. 6. Mida tead bakteris leiduvast kromosoomist? Tsütoplasmas asub üks rõngaskromosoom, mis koosneb ühest DNA-molekulist 7. Joonista bakteri skeem koos nimetustega. 8. Bakterid paljunevad: a)eostega , b) pungumisega, c) paljunemisrakukestega, d) mütseeli tükikeste abil, c) .pooldumisega 9. Paljunemise etapid: • Bakterirakk suureneb, DNA replitseerub ning liigub raku poolustele • Rakukest ja –membraan sopistuvad ning moodustub rakuvahesein
Bioogia kt 2 Elusloodus jaguneb: üherakulised ja hulkraksed organismid.-üherakulisi on mitmeid kordi rohkem, aga enamus on väga väiksed ja palja silmaga seetõttu ei näe. Kõige pisem üherakuline on mükoplasma.(võib tekitada hingamisteede haigusi) Üherakulistel aine-, energia-, infovahetus ümbritseva keskkonnaga rakumembraani vahendusel. Bakteris on kerakujulised, pulkjad, niitjad, kruvikujulised. Eeltuumsed ehk prokarüoodid (bakterid- puudub membraaniga piiritletud tuum, vähem organelle ja membraanseid struktuure), päristuumsed ehk eukarüoodid (sisemus täidetud poolvedela tsütoplasmaga, milles on mitmesuguseid organelle). Eukarüoodid jaotatakse protistideks, taime-, seene- ja loomariigiks. Viirused ei ole eel- ega päristuumsed. Rakk on ümbritesetud rakumembraaniga
(fosfataasid-võtavad ära, kinaasid- panevad juurde). Geeni reg- kromosoomi üleminek valguks. Tsentromeer- hoiab koos kahte kromatiidi. Telomeerid- hoiavad kromosoomi otsi tugevana, et mitte lasta kromosoomil lüheneda ja pärilikkusel kaduda. Eukromatiin- lõdvalt pakitud , aktiivse transkrip. ala. Heterokromatiin- tugevalt pakitud, transkr. ei toimu. Operon- erinevaid valke kodeerivad alad. Speiser- geenide vaheline ala. Prokarüoodi geenis puuduvad.Operaator- negatiivne geenireg piirkond (bakteris). Repressor- DNAd siduv valk mis represeerib transkriptsiooni.cDNA- complementary DNA- küpsest mRNAst (RNA sõltuvast DNA polumeraasi abil ) sünteesitud DNA. Enhansereid vaja transkriptsiooniks. microRNAs (miRNA) üheahelalised RNA molekulid 21-23 nukleotiidi pikad. Reguleerivad geeniekspressiooni. miRNAd kodeerib geen mis on transkribeeritud DNAlt kuid ei ole transleeritud valguks ( mittekodeeriv DNA)Mediaatori funkts: (valgukompleks 50 erinevast valgust). Tähtis
Geenide regulatsiooni uurimine on molekulaarbioloogia peamine valdkond • Molekulaarbioloogia üks olulisemaid teemasid on – miks osa geene avalduvad ühtedes rakkudes/kudedes ja teised geenid teistes rakkudes/kudedes? • Kõige olulisem geeni avaldumise regulatsioon toimub transkriptsiooni tasemel. Geenide regulatsiooni tasemed Transkriptsiooni regulatsiooni uuringud said alguse E. coli bakteris Operonis on kolm geeni: Z, Y ja A. LacZ on b-galaktosidaas ehk ensüüm, mis lõikab laktoosi glükoosiks ja galaktoosiks. Lähedal asub LacI geen, mis kodeerib repressorvalku ja seondub Operaator regiooniga. Sellega blokeerib geenide ekspressiooni. Teisalt seondub hästi laktoosiga. Kui söötmes on laktoosi, siis vabaneb promootor ja LacZ geeni käivitub. Eukarüootse geeni regulatsioon
-) Fagotsütoos protess, mille abil rakk transpordib suuremaid aineosakesi. -) Karüoplasma poolvedel ,,täide" rakutuumas, nagu tsütoplasma rakus. -) Mütseel seeneniidistik. -) Homoloogiline kromosoom paaris kromosoomid (sarnase informatsiooni põhjal). -) Bakteritoksiin mürk, mida bakterid toodavad. * 2. Osa Kas väited on tõesed või valed? Paranda (kirjutan vaid õiged vastused) (2p) -) Ühest bakteris tekib üks spoor. -) Taimerakule iseloomulikud organellid on plastiidid ja vakuoolid. -) Ribosoomides toimub valgu süntees. -) Spooridel toimub ainevahetus maksimaalselt aeglaselt. -) Valguse puudumisel muutuvad kloroplastid leukoplastideks. -) Seened on heterotroofsed organismid. -) Tuumasisest plasmat nimetatakse karüoplasmaks. -) Ainetranspordiks läbi rakumembraani vajatakse täiendavat energiat, siis kui transport toimub aktiivselt (tahkete ainete transport).
Ligeerimissegu: 1 μl puhastatud PCRi produkti 1 μl pSTBlue-1 vektorit (9 ng/μl) 0,5 μl 10x puhvrit 1 μl T4 DNA ligaasi (sünteesib fosfodiester sidemeid) 1,5 μl MQ vett 48. 49.Transformeerimine 50. Eesmärgiks on peale ligeerimist saadud DNA molekuli viimine kompetentsetesse E. Coli bakterisse ja see on oluline selleks, et saaksime kiiresti ja odavalt ning väga väikese vigade hulgaga palju seda plasmiidi (paljundame seda bakteris). Bakterid on eelnevalt töödeldud divalentsete katioonidega (Ca2+, Mn2+), mis muudab rakuseina laengu positiivseks. Plasmiidse DNA lisamisel kinnitub see rakuseinale. Bakteri rakke kuumutatakse lühikest aega ning kuna E. Coli rakke töödeldakse külmades tingimustes, toimub temperatuuri muutus (heat shock) ning selle tulemusena tekivad rakuseina kahjustused, kust plasmiidne DNA pääseb rakku. 51
Mitmed bakterid võivad olla ohtlikud e patogeensed, sest põhjustavad tervisehäireid, kuna eritavad väliskeskkonda mürke bakteritoksiine. Bakteri rakutuuma asendab tuumapiirkond, milles paikneb rõngaskromosoom (koosneb ühest DNA molekulist ja tal pole vabu otsi). Kõikidel bakteritel on üks kromosoom. Enne bakteri jagunemist rõngaskromosoom kahekordistub. 7 Plasmiidid väiksemad DNA rõngad bakteris, mis on vajalikud spetsiifiliste ensüümide sünteesiks ellujäämisel. Plasmiidide arv pole püsiv Ained liiguvad bakteris difusiooni teel. Valgusüntees toimub ribosoomides Mõnes bakterite tsütoplasmas on gaasivakuoolid, mis aitavad bakteril veel oma kõrgust muuta. Bakterid paljunevad pooldumisega. Rakumembraan kestaga nöörib sisse ja moodustub kaks ühesuurust tütarrakku. Bakterid paljunevad suhteliselt kiiresti.
5 Vakuoolid Vakuool esineb enamasti taimerakkudes. Vakuool on põieke, mis on ümbritsetud membraaniga. Ta täidab rakus seedeülessandeid. Vakuool ilmub tsütoplasmasse ja täitub vedelikuga. Vakuool toob sisse väliskeskkonnast toitaineid, ning tema sisse tulevad jääkained, mida ta pärast eritab väliskeskkonda. Kui vakuool esineb bakteris, siis temas on gaas ja teda nimetatakse gaasivakuooliks. Parasiitidel puudub vakuool. Vakuooli ülessanded: 1) Ta lagundab ja on lüsosoom. 2) Temas säilitatakse aineid, seega on säilitusorganell. 3) Ta kontrollib raku siserõhku. 4) Tema läbi saab taimerakk ennast suurendada. Vakuoolis säilitatakse mittevajalikud ained, mida võib pärastpoole vaja minna. Näiteks: suhkrud
Restriktaase on erinevaid, igaühel oma “äratundmiskoht” DNA-l. Restritaasid lõikavad DNA sagely nii, et tekivad üheahelalised otsad - “kleepuvad otsad”. Selliste otstega DNA lõike on komplementaarsuse tõttu võimalik mugavalt liita. Kuidas kergesti aru saada, et soovitud geen on üle kandunud? - üle viidavale geenile on markergeen külge pandud (GFP-geen). Miks bakterirakus ei hakka inimese geen kohe tööle? Meis on intronid ja eksonid, kuid bakteris neid ei ole. Kui tahame bakterisse geeni via, siis peame selle mRNA alusel tegema DNA, seda protsessi, pöördtranskriptsiooni viib läbi PÖÖRDTRANSKRIPTAAS. Edasi saab cDNA juba liita bakteri DNA-ga ning ta hakkab tootma inimese organismis esinevat valku. Kui kasutame baktereid sel viisil mõne teise liigi geenide säilitamiseks, same genoomipanga. Mudelorganismid geneetikas: kiire põlvkondade vahetus, suhteliselt vähe geneetilist materjali,
geenitehnoloogilise avastuse eest (Paul Berg, Walter Gilbert ja Frederick Sanger). Patenteeritakse esimene organism - transgeenne "õli sööv" bakter. 1978: Inglismaal sünnib esimene katseklaasis vljastatud munarakust saadud embrüosiiratud laps. 1977: Saadakse inimese geeni esmakordne avaldumine bakterirakus. Järgmisel aastal saadakse esimest korda inimese valku (insuliin) transgeensetest bakteritest. 1976: Saavutatakse pärmseene geenide avaldumine bakteris Escherichia coli. Moodustatakse rahvusvaheline Rekombinantse DNA Järelvalve Komitee, kehtestatakse ametlikud reeglid rekombinantse DNA sisestamiseks mikroorganismidesse ja viirustesse. 1975: Luuakse hübridoomitehnoloogia monokloonsete antikehade saamiseks. 1973: Stanley Cohen ja Herbert Boyer sooritavad bakteril esimese eduka katse rekombinantse DNA-ga. 1972: Embrüosiirdamise moodsa tehnoloogilise väljatöötamine ja rakendamine põllumajandusloomadel.
efektile on näha see spetsiifiline koht ka valgusmikroskoobis nähtav.Ekspressioonikonstruktid olid plasmiidid GFP järjestuse ja erinevate signaalidega, mille tõttu ekspresseerus GFP erinevates rakupiirkondades. PEI polüetüleenimiin pakib DNA kokku ja interakteerub membraaniga ja difundeerub läbi membraani. 3. Millised elemendid peavad olema eukarüootses ekspressioonivektoris? Loomarakkudele mõeldud ekspressioonivektor (plasmiid) sisaldab: 1.komponente, mis võimaldavad plasmiidi bakteris paljundada (prokarüootset replikatsiooni alguspunkti, antibiootikumi resistentsusgeeni); 2.kloneerimisala (MCS) erinevate restriktaaside lõikekohad; 3.uuritavat järjestust (valkude korral tavaliselt cDNA kujul); 4.reportergeeni (GFP, lutsiferaas, lacZ jt.) või märgise (tag; FLAG, myc, V5, GFP) järjestust. 5.eukarüootset promootorit (CMV, SV40) ja polüadenüleerimisjärjestust (SV40); 6.selektsioonimarkeri ekspressioonikassetti. Praktiline töö III Immuunotsütokeemia 1
Mittevajalikud plasmiidid lagundatakse vastavate ensüümide poolt. Selle tulemusena muutub nii plasmiidide kui ka neis sisalduvate geenide arv. hõlbustab liikumist. Eeltuumse raku sisemuses puuduvad membraanidest koosnevad rakustruktuurid ja nendega ümbritsetud organellid. Seega ei ole bakteritel tsütoplasmavõrgustikku, Golgi kompleksi, kloroplaste, mitokondreid, tsentrosoom, tsütoskelett. Tsütoplasma ringleb bakteris vähem kui pärsituumses rakus ja ainete kandumine raku ühest osast teise toimub valdavalt difusiooni teel. Valgusüntees toimub ka bakterites ribosoomides. Ribosoomid on aga väiksemad ja sisaldavad vähem rRNA ja valgu molekule. Mõnede bakterite tsütoplasmas esinevad väikesemõõdulised gaasivakuoolid. Need on valgulise membraaniga ümbritsetud põiekesed. Gaasivakuoolid on iseloomulikumad vesikeskkonnas elutsevatele bakteritele. Need
ning valk seondub ka väiksema afiinsusega saitidesse. Dps seondub DNA-ga mittespetsiifiliselt, kaitstes DNA-d oksüdatiivsete kahjustuste eest. Paljud valgud degradeeritakse ja väheneb ka RNA stabiilsus. 3) Toimub valkude denaturatsioon ja proteolüüs. Selle kaitseks on indutseeritud HSP valgud ja valgud, mis parandavad kahjustusi aminohapetes (Pcm, MsrA). 4) Sünteesitakse uusi valke. Praeguseks on bakteris E. coli identifitseeritud ligi 100 statsionaarsele faasile spetsiifilist geeni. 5) Ribosoomide arv rakus väheneb. 70S ribosoome konverteeritakse 100S dimeerideks. Dimeriseerumisel osaleb 55 aminohappe suurune aluseline valk RMF (ribosome modulation factor). RMF-defektsed rakud elavad statsionaarset faasi halvasti üle. Dimeersed ribosoomid on resistentsemad proteaasidele ja nukleaasidele ning translatsiooniliselt inaktiivsed. Seni puuduvad andmed, kuidas toimub sellistel
Kasvajad tekivad juurtel, okstel ja tüvel. · Kasvajad takistavad vee- ja toitainete liikumist taimes. Taimed kääbustuvad ja on väheviljakad. · Algul on kasvaja pehme, soolatüükataoline, hiljem aga puitub. · Agrobacterium tumefaciensi poolt põhjustatud kasvaja õunapuu oksal · Agrobacteriumi kasutatakse taimede insenergeneetikas uute geenide sisseviimiseks taimegenoomi. Lisatavad geenid lülitatakse bakteris replitseeruvasse Ti plasmiidi. · Selle tehnoloogiaga on saadud näiteks herbitsiidide suhtes resistentseid taimi. · Herbitsiid "Roundup", mille toimeaine on glüfosaat, mis ihibeerib aromaatsete aminohapete sünteesi taimedes, nii kultuurtaimedes kui ka umbrohtudes. · Seda herbitsiidi kasutatakse nn musta maa tegemiseks enne külvi. · tumefaciensi ja mitmete teiste bakterite vastav ensüüm on aga glüfosaadile tundetu.
kõrgema efektiivsusega. Sageli on transkriptsiooni initsiatsiooniks vajalik ka spetsiifiliste TF-de olemasolu. Lisaks TF-dele toimub transkriptsiooni regulatsioon ka erinevate faktorite kaudu. 70 on vajalik bakterile eluliselt tähtsate geenide, nn. "house keeping" geenide avaldumiseks eksponentsiaalselt kasvavates 4 rakkudes. Seetõttu nimetatakse seda põhiliseks faktoriks. Bakteris Escherichia coli on põhiliseks faktoriks 70, Bacillus'es 43. Stressitingimustes (näit. temperatuuri tõus kasvukeskkonnas, toitainete nälg, oksüdatiivne stress) lülitatakse tööle geenid, mille promootoreid tunnevad ära alternatiivsed faktorid. faktorite tähistamisel kasutati algul indeksit, mis iseloomustas vastava valgu liikuvust SDS geelis. Paljud faktorid liikusid aga anomaalselt ning sellepärast hakati indeksina kasutama arvestuslikku molekulmassi
k. guide RNA). Giid-RNA-s asuvad mRNA-ga mittepaarduvates alades A nukleotiidid ja just nende vastu lisatakse mRNA-s uridiinmonofosfaadid. Mõjutab oluliselt geenide avaldumist trüpanosoomide ja taimede mitokondrites. 63. Transkriptsiooni ja translatsiooni toimumise aeg ja koht bakterites ja eukarüootides. Transkriptsioon ja valgusüntees (translatsioon) toimuvad bakterirakus korraga, kindel koht puudub, eukarüoodis aga eraldi etappidena ja erinevates kohtades. Bakteris : Iga mRNA molekul on samaaegselt transleeritav paljudel ribosoomidel polüribosoomil (polüsoomil). Valgusüntees toimub ribosoomidel, mis koosnevad 3-5 erinevast rRNA molekulist ja üle 50 erinevast ribosoomivalgust. Eukarüoodil: Transkriptsioon rakutuumas. Translatsioon tsütoplasmas. Bakterirakus ei ole ribosoomidel kindlat asukohta. 64. Ribosoomide ehitus prokarüootses ja eukarüootses rakus. Ribosoomid koosnevad väikesest ja suurest subühikust.
DNA güraasi on vaja E. coli DNA replikatsioonil vähendamaks replikatsioonikahvli ees tekkivat positiivset superspiralisatsiooni Pol III: põhiline DNA replikatsiooni läbiviija. Polümeraasi õlad sünteesivad erinevaid ahelaid: vasak õlg – juhtivat. Parem õlg – Mahajääva ahela Okazaki fragmente. Pol IV ja Pol V: DNA reparatsioonilised polümeraasid. vigaderohke DNA süntees, kui Pol III töö on blokeeritud DNA kahjustuse tõttu. Bakteris on põhiliseks DNA replikatsiooni läbiviivaks valguks DNA polümeraas Pol III. DNA replikatsiooni täpsuse eest vastutab Pol III subühik epsilon. Epsilonil on 3’- 5’ suunaline endonukleaasne aktiivsus, mis võimaldab valesti DNA ahelasse lülitatud nukleotiide kõrvaldada. 31 52. DNA replikatsioon juhtivalt ja mahajäävalt ahelalt. DNA ahelate komplementaarsuse tõttu on DNA ahelad vastassuunalised. Replikatsiooni alguspunktis
keskkonnasäästlikumat põllumajandust kui geenitehnoloogiline. Kardetud on ka Bt pikaajalise looduses akumuleerumise tagajärgi. Tegelikult on Bt-valk küllalt ebastabiilne ja tema kuhjumine mis tahes süsteemis ei ole reaalne. Samuti ei ole suudetud tõestada GM-taimede kahjulikku mõju mulla mikrofloorale, mida on eriti aktiivselt uuritud B-taimede puhul. Väide, nagu oleks taimedes toodetud Bt-toksiinil teistsugune aktiivsus kui bakteris sünteesitul, on kogu senise teadusteabe põhjal täiesti väär. Neidki hüpoteese on võimendatud Eesti ajakirjanduses. Keskkonnakaitsjate argumendid geenmuundamise vastu on selged. Looduses sellist asja ei esine. Ja säherdune geenidega manipuleerimine võib algatada pöördumatuid protsesse ökoloo- gilistes süsteemides. Ning vähendab ka liigilist mitmekesisust. Lisaks satuvad GM-firmade 3
1 toitumist ja hingamist; 2 paljunemist ja teatud keskkonna asustamist; olema resistentne välistele faktoritele tänu paindlikele geneetilistele mehhanismidele, s.o. ellu jääma äärmuslikes tingimustes. Bakterirakk koosneb tsütoplasmast ja membraanist. Bakterid paljunevad lihtsa pooldumise teel, mis algab sellest, et mikroob kasvab oma pärilikult määratud pikkuseni. Pooldumise eel toimub DNA replikatsioon, mille tulemusena moodustub bakteris kaks rõngaskromosoomi. Järgnevalt sünteesitakse raku keskossa rakumembraanid.. Protsess lõpeb raku keskele vaheseina tekkimisega, mille tulemusena moodustuvad kaks uut tütarrakku. Need võivad jääda üksteisega veel mõneks ajaks seotuks, moodustades erineva pikkusega ahelaid Koos tsütoplasma jagunemisega kaheks jaotuvad selles paiknevad rakuorganellid tütarrakkude vahel. Moodustunud tütarrakud on geneetiliselt samased lähterakuga. Pooldumisprotsessidega
tsütolüüs – rakk lõhkeb, sest palju vett tuleb rakku. Toimub hüpotooniliseks keskkonnas, kus vesi liigub rakku osmoosi teel (rakus konts. kõrge), membraan ei pea vastu. hemolüsiin – hemolüütiline toksiin, lüüsivad vereliblesid M. haemolytica – põhjustab veiste pneumooniat. FUR proteiin – ferric uptake regulator - rauasõltuv transkriptsiooni pidurdaja bakteritel (seondub promootorile) nn raudsõltuvad geenid. Sellega kontrollitakse raua sisaldust bakteris (kõrge kontsentratsioon võib bakterile ohtlik olla) NAD – nikotiinamiid adeniin dinukleotiid pertaktiin – välismembraani proteiin, mis aitab trahhea epiteelirakkudele kinnituda. Kontrolltöö 3 1. Er. rhusiopathiae üldiseloomustus ja virulentsusfaktorid Gram+, liikumatud, pulgad või filamendid, fakultatiivsed mikroaerofiilid, katalaas- neg, hea kasv vereagaril, põhjustab seal, kalkuni punataudi. Bakterit on
Viirused evolutsioneeruvad iseseisvalt (mutatsioonid, võtavad kaasa peremehe geene, võtavad kaasa teiste rakus olevate viiruste geene) Oblikatoorsed ained: 1)nuklehape DNA või RNA (pärilikkusaine). 2)Valk moodustab apsiidi (kaitseks - kaitsevalgud ja peremeesraku ära tundmiseks - retseptorvalgud). Suurus: alla valgusmikroskoobi nähtavuse piiri. Kuju: 1)Isodiameetriline (hulktahukas). 2)Pulkjas lühem, niitjas pikem. 3)Liitehitused (faagid) bakteris viljelevad viirused. Rühmad: 1) DNA. 2) RNA sünteesivad RNA põhjal DNA ja hakkavad seal paljunema. 3) (DNA+RNA Riketsiad bakterid) pole viirus, parasiitne, rakusisesed bakterparasiidid. Päritolu: 1) Eeleluline, ürgpuljongi esimeste elustruktuuride järglased. 2)Hulkuma läinud geenirühm viirused kasutavad enda arendamiseks peremeesraku geene. 3) Redutseerunud bakter. Viiruse leviku eeldus: Parasiit saab eksisteerida vaid piisavalt tihedas peremehe populatsioonis
(3) Ribosoomi A-saidis asuv aminoatsüül-tRNA liigub P-saiti ja enne seda P-saidis asunud tRNA, mis ei ole enam aminohappega seotud, liigub E-saiti. Ribosoom liigub EF-G toimel mRNA molekulil kolme nukleotiidi võrra edasi mRNA 3´-otsa suunas, tarbides GTP energiat ning A-sait jääb vabaks, seondumaks järgmise aminoatsüül-tRNA molekuliga 68. Kirjeldage translatsiooni terminatsiooniprotsessi. Translatsioon termineerub, kui terminatsioonifaktorid tunnevad A-saidis ära stop koodonid. Bakteris RF1 UAA ja UAG koodonid, RF2 UAA ja UGA. Eukarüootides on ainult eRF, mis tunneb ära kõiki 3 koodonit. A-saiti sisenenud RF muudab peptidüültransferaasi aktiivsust ja peptidüültransferaas lisab polüpeptiidahela viimase aminohappe karboksüülrühmale vee molekuli Vabaneb valmis polüpeptiidahel P-saidis asuvalt tRNA molekulilt ning vaba tRNA liigub ribosoomi E- saiti. Translatsiooni terminatsiooniprotsess lõpeb mRNA vabanemisega ribosoomilt ja ribosoomi
-bakterid (ja arhed) asustavad ka piirkondi, kus teiste organismide elu ei ole võimalik -bakterite olulisimaks rolliks on orgaanilise aine lagundamine -inimeses on sümbiootilisi baktereid (maos, soolestikus jne) ning teised bakterid põhjustavad haigusi -baktereid kasutatakse toiduainetööstuses, ravimitööstuses, biopuhastuses Bakteriraku ehitusest: 1. geeneetilise aine info: -bakteritel puudub rakutuum -bakteri DNA moodustab ühe rõngaskromosoomi -piirkond bakteris, kus DNA rõngaskromosoom paikneb, nimetatakse nukleoidiks -lisaks võivad bakterites olla ka plasmiidid, mille omadusteks, ülesanneteks on: 1. liiguvad kergelt rakust rakku ning omastavad võõraineid kergesti 2. omavad enüümide tootmiseks vajalikke geene 2. ribosoomid, mis - ei ole kinnitunud tsütoplasma võrgustikku - paiknevad rõngaskromosoomi lähedal -valgusüntees (ja seega ka bakteri elutegevus) on väga kiire 3. tsütoplasma (sarnane eukarüootsele rakule 4. rakumembraan 5
inserdi, aga valged sisaldasid õige plasmiidi. Valisin kõige suurema valge koloonia ja panin neid kasvama: Võtsin epsi, kus tegin augu (ilma hapnikuta bakretid suervad ära) Lisasin 1ml Amp-i sisalav vedelsööde Otsikuga katsusin valge kolooniat ja panin otsik epsi sisse Panin bakterid loksutavasse inkubaatorisse 16 tunniks kasvama 375 C juures 6.1 Praktikum – minipreparatsioon aluselise lüüsi meetodil Eesmärgiks on bakteris paljundatud plasmiidi välja puhastamine. Võtsin üleöö kasvanud bakterid ja fuugisin bakterid põhja Eemaldasin supernatanti Re-suspendeerisin pellet 0,2 ml-s E1 lahuses (50mM glükoos – teeb lahuse isotooniliseks, 25 mM Tris-HCl, pH 8,0, 10nM EDTA-Na2 – seob katioone, inhibeerides ensüüme(DNAaasid) ja destabiliseerib raku membraani, RnaasA 10 µl/ml – eemaldab bakteriaale RNA prepsist).
> kahjulike ainete lagundamiseks või vältimiseks > plasmiide ja neis asuvate geenide arv ei ole püsiva suurusega > mittevajalikud plasmiidid lagundatakse vastavate ensüümide poolt. Bakteriraku organellid ° Eeltuumse raku sisemuses puuduvad membraanidest koosnevad rakustuktuurid ja nendega ümbritsetud organellid. > puudub tsütoplasmavõrgustik, Golgi kompleks, kloroplastid, mitokondrid, tsentrosoom ja tsütoskelett. ° Tsütoplasma ringleb bakteris vähem kui päristuumses ° ainete kandumine toimub difusiooni teel peamiselt ° Bakteritel toimub valgusüntees ribosoomides (nagu ka eukarüootsetes) > mõõtmetelt väiksemad > vähem rRNA ja valgu molekule ° Mõnede bakterite tsütoplasmas gaasivakuoolid > valgulise membraaniga ümbritsetud põiekesed > iseloomulikumad vesikeskkonnas elavatele bakteritele Bakterite elutegevuse iseärasused
· P saidis toimub AH lisamine kasvavale polüpeptiidahelale · E saiti liigub tRNA, millelt AH on seotud polüpeptiidahelasse. Saitide põhiosad asuvad ribosoomi suures subühikus. mRNA asub väikeses subühikus. 66. Võrrelge translatsiooni initsiatsiooni bakterites ja eukarüootses rakus. Translatsiooni initsiatsioon sündmused, mis eelnevad peptiidsideme moodustumisele sünteesitava polüpeptiidi kahe esimese aminohappe vahel Bakteris: ribosoomid on jaotunud kaheks alaühikuks 30s ja 50s. Initsiaatorkoodon mRNA molekulis on AUG koodon. Ribosoomi 30s subühik ja mRNA moodustavad kompleksi IF3 juuresolekul. Metionüül-tRNA inaktiveerub initsiatsioonifaktoriga. Täielik initsiatsioonikompleks moodustub metionüül-tRNA ning mRNA/30s subühik/IF3 komplekside kombineerumisel IF1 ja GTP juuresolekul. Enne 50s subühiku lisandumist vabaneb IF3 50s subühiku liitumisel 30s subühikuga tarbitakse
GEENITEHNOLOOGIA Geenitehnoloogia seisneb: 1) DNA lõikude eraldamises, töötlemises in virto ja siirdamises. Kas sama või teise isendi gromosoomi siirdamine plasmiidi(on bakteris) või viirusesse. 2) Geenide suunatud väljalülitamistes ehk geeni knock out'is(lk 124). Geenitehnoloogiat rakendatakse transgeensete loomade ja taimede saamiseks.Geeniravis, ehk geeniteraapias. 3) Teostatakse sünnieelselt ja järgset diagnostikat DNA proovidega. 4) Tuvastatakse isadust, kurjategijat(ü.s. tuvastatakse inimest). Transgeensete organimside loomine põhineb restriktas tehnikal(see on bakterites olev ensüüm, mis lõikab DNA tükkideks). DNA ahelate lõigud on vastavad ehk
Arhede histoonid · Eukarüootidel on DNA pakitud ümber aluseliste konserveerunud järjestusega valkude - histoonide, moodustades nukleosoome (pildil nukleosoomidega DNA). Nukleosoomide vahele jäävad pakkumata DNA alad. · Histoonid on leitud eurüarhedel. Esmalt leiti need Thermoplasma acidophilumil ja näidati, et in vitro võivad nad seostuda DNAga, moodustades nukleosoomitaolisi struktuure. · Samas bakteris nähti elektronmikroskoobiga nukleosoomidega DNAd. · Ilmselt koosnevad arhede nukleosoomid histooni tetrameerist (eukarüootidel oktameerid), mille ümber on pakitud ca 80 bp DNAd. Arvatakse, et eukarüootide oktameerne histoonkompleks võis välja kujuneda arhede tetrameersest. Kompleks suurenes, kuna eukarüootidel on rohkem DNAd. Nendel arhedel, kellel histoone pole leitud, on kirjeldatud histoonitaolised valgud, mis ka seostuvad DNAga. Arhede rakuskelett
1. Transformatsioon - geneetilise informatsiooni ülekandumine ühest bakterirakust teise rakku isoleeritud DNA abil. Transformatsioon võib toimuda ka looduslikes tingimustes. Sel juhul kandub elusrakkudesse surnud rakkudest vabanenud DNA. Transkriptsioon ümberkirjutamine, DNA ühe ahela alusel komplementaarse RNA molekuli süntees. Translatsioon mRNA põhjal ribosoomides valguahela sünteesimine ehk lihtsamalt öeldes valgu süntees. (RNA alusel valgu süntees tsütoplasmas paiknevatel ribosoomidel.) Translatsiooniprotsess loob geneetilise koodi ehk vastavuse mRNAde nukleotiidahelate ja valkude polüpeptiidahelate vahel. 2. Initsiatsioon prokarüootidel: transkriptsioon algab sellega, et protsessi läbiviiv ensüüm RNA polümeraas kinnitub struktuurgeenide ees asuvas promootoris sisalduvatele transkriptsiooni algussignaalidele. RNA polümeraas ei vaja transkriptsiooni initsiatsiooniks praimerjärjestust nagu seda vajas DNA polümeraas. Promootori pi...
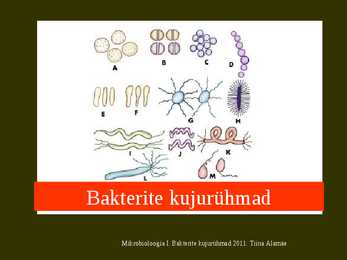
Planctomycese rosett. Fluorestseeruv geeniproov on Planctomyces'e rühma spetsiifiline Planctomycesel moodustub rakule valguline jätke ja raku vastaspoolusele moodustub pung. Mikrobioloogia I. Bakterite kujurühmad 2011. T Punguvad ja jätketega bakterid Verrucomicrobium spinosum on soolatüükataoliste jätketega bakter, keda on leitud veest ja mullast, aga ka inimese soolest. Eriti arvukalt neid mullas. Bakteris olemas muidu eukarüootidele omane tubuliini kodeeriv geen. Mikrobioloogia I. Bakterite kujurühmad 2011. Tiina Alamäe Punguvad ja jätketega bakterid Gallionella on levinuim rauabakter. Esineb rauarikastes allikates, drenaazhitorudes jne. Rakk eritab rauda oksüdeerides limasaba, millesse sadenevad
DNA replikatsioonikahvlite liikumine vastassuunas. DNA polümeraasid bakterites: E. coli DNA polümeraas I; replikatiivne DNA polümeraas III; DNA polümeraaside vigu korrigeeriv (proofreading) aktiivsus; vigaderohked DNA polümeraasid pol IV ja pol V bakterites. DNA replikatsiooni toimumine mõlemalt DNA ahelalt korraga: juhtiv ahel; mahajääv ahel; Okazaki fragmendid; praimosoom; replisoom. DNA replikatsiooni initsiatsioon oriC-lt bakteris E. coli. DNA ahelate lahtikeeramisel osalevad valgud. Veereva ratta replikatsiooni mudel. Eukarüoodi kromosoomi replikatsiooni eripärad. Replikon. Telomeeride pikkus ja vananemine. 12. Transkriptsioon ja RNA protsessing. Molekulaarbioloogia põhidogma. RNA tüübid: tRNA, rRNA, mRNA, snRNA. RNA süntees ja selles osalevad valgud; promootorid; transkriptsiooni regulaatorid.
Helicobacter pylori imetajate mao patogeen 1 667 867 bp 1576 geeni Mycoplasma genitalium imetajate patogeen 580 074 bp 480 geeni Kromosoomi struktuur ja pakitus sõltub bakteri kasvukiirusest. Kui E. coli kasvab kiiresti ning rakk sisaldab 4 8 kromosoomi, on kromosoomne DNA tugevasti pakitud ning DNA lokaalne super-spiraliseeritus varieerub tugevasti. Samas aeglaselt kasvavates rakkudes, kui bakteris on 1 2 kromo-soomi, pole kromosoom tugevalt pakitud ning on suhteliselt struktuuritult üle kogu tsütoplasma. Kromosoomi tihedus pole rakus ühtlane, vaid väheneb pooluste suunas. Kromosoom on tihedaim keskelt ning hõredaim otstest. DNA kondenseeritus muutub ka rakutsükli jooksul, ühe jagunemise ajal võib toimuda mitu kromosoomi kondenseerumist ja dekondenseerumist. Arvatakse, et replikatsiooni initsieerimisel on E. coli'l kromosoomi origin ja ter-piirkond raku keskel,
3. Bakterite kasv ja paljunemine Ühe bakteriraku võimet sünteesida oma raku komponente ja assimileerida energiat, mille tulemuseks on tema suurenemine (liigiomastes piirides) ja järgnev pooldumine, nimetatakse kasvuks. Kasvu laiem mõiste on mikroobide paljunemine. Bakterid paljunevad lihtsa pooldumise teel, mis algab sellest, et mikroob kasvab oma pärilikult määratud pikkuseni. Pooldumise eel toimub DNA replikatsioon, mille tulemusena moodustub bakteris kaks rõngaskromosoomi. Järgnevalt sünteesitakse raku keskossa rakumembraanid.. Protsess lõpeb raku keskele vaheseina tekkimisega, mille tulemusena moodustuvad kaks uut tütarrakku. Need võivad jääda üksteisega veel mõneks ajaks seotuks, moodustades erineva pikkusega ahelaid Koos tsütoplasma jagunemisega kaheks jaotuvad selles paiknevad rakuorganellid tütarrakkude vahel. Moodustunud tütarrakud on geneetiliselt samased lähterakuga
koodonini ning see tõttu algab translatsioon enamasti esimeselt AUG koodonilt. Prokarüootides on vaja mRNAs spetsiaalset RBS (ribosome binding site) järjestust AGGAGG, mis asub AUG koodonist 7 nukleotiidi kõrgemal. · Sarnasused üldjoontes on sama intsiatsiooniprotsess. :D 67. Kirjeldage translatsiooni elongatsiooniprotsessi. · Kolmeetapiline protsess üldjoontes sarnane bakteris ja eukarüoodis Aminoatsüül-tRNA seondub ribosoomi A-saiti ning paardub antikoodonjärjestuse kaudu parasjagu A-saidis asuva koodonjärjestusega mRNAs, olles assotsieerunud elongatsioonifaktoriga EF-Tu, mis on seotud GTP-ga. Moodustub peptiidside ribosoomi A-saidis oleva aminoatsüül-tRNA aminorühma ja ribosoomi P-saidis oleva tRNA-ga seotud kasvava polüpeptiidahela viimase AH karboksüülrühma vahele
experiments). Need katsed kinnitavad, et Okazaki fragmendid on DNA replikatsiooni vaheproduktiks. Juhtiva ja mahajääva DNA ahela süntees on väga täpselt koordineeritud protsess. Kogu replikatsiooniaparaati, mis liigub mööda DNA molekuli replikatsioonikahvlina, nimetatakse replisoomiks. Replisoom koosneb DNA polümeraas III holoensüümist, mille üks apoensüüm sünteesib juhtivat ahelat ja teine mahajäävat ahelat ning praimosoomist. DNA replikatsiooni initsiatsioon oriC-lt bakteris E. coli Algne initsiatsioonikompleks moodustub 20-30 DnaA monomeeri sidumisel. ATP juuresolekul toimub avatud kompleksi moodustumine, kusjuures DNA ahelate lahtikeerdumine algab kõige parempoolsemast 13-meerist. Seejärel moodustavad DnaB helikaas ja DnaC denatureerunud DNA-ga prepraimingkompleksi. SSB (üksikahelalise DNA-ga seonduv valk) ja güraasi (topoisomeraas II) juuresolekul jätkub DNA ahelate lahtikeerdumine mõlemas suunas. Järgneb praimeri süntees primaasi
valgusmikroskoobis analüüsitavad. Niisiis, võrreldes prokarüoodiga on eukarüoodi kromosoomide arv suurem, kromosoomid ise on palju suuremad ning nende ehitus keerulisem. Kromosoomi eri piirkonnad sisaldavad erinevaid DNA klasse ja täidavad kindlaid funktsioone. Eriti olulised kromosoomi kui terviku säilimisel on tsentromeeri ja telomeeri alad. 18. Milliseid rakke ümbritseb rakukest? Taimerakke + veel midagi?! 19. Mis on plasmiid? Bakteris asuvad ekstrakromosomaalsd DNA elemenid e. plasmiidid, DNA rõngasmolekulid. Aitavad halvas keskkonnas kohaneda?! 20. Rakutsükli etapid. Rakutsükkel koosneb mitoosist ja interfaasist. Mitoosis eristatakse karüokineesi ja tsütokineesi 21. Milleks on gameetide küpsemisel vaja meioosi? Gameetide keharakkudes on diploidne kromosoomistik, et sugulisel paljunemisel kromosoomide arv viljastumise tulemusena ei kahekordistuks, peab kromosoomide arv vähenema 2x. 22
Kõik raku sisemuses asuvad osad ujuvad poolvedelas plasmas. Suure osa taimerakust täidab vakuool, mis on täis rakumahla. Vakuool hoiab raku pallikesena pinges. Rakumahlas liiguvad kõik raku toitained. 39. Restriktaasid - ensüümid, mida toodavad bakterid enesekaitseks. Nad lõikavad DNA-d spetsiifilise nukleotiidse järjestuse järgi nii, et tekivad üheahelalised ehk kleepuvad otsad, mida on mugav komplementaarsuse põhjal liita. Bakteris tunnevad restriktaasid neile spetsiifilise koha 48 nukleotiidi järgi ära ja lõikavad võõra DNA sealt katki sellist mehhanismi nimetatakse restriktsiooniks. 40. DNA kloneerimise etapid. Kloonimise puhul võib eristada kaht meetodit: 1. Rakuvaba DNA kloonimine ehk in vitro DNA kloonimine ehk PCR. Polümeraasi ahelreaktsioon ehk PCR on meetod DNA või RNA järjestuse kordistamiseks. PCR- meetod võimaldab väga väikesest DNA lõigust luua miljoneid koopiaid. PCR
on tihe või on hulkrakne, haruldastel liikidel pole viiruseid, füüsikaliste välisjõudude toel nt. gripiviirus ja faagid, parasiitloomade abil, viirus paljuneb ka vaheperemehes nt. puukides entsefaliit ja sääskedes kollapalavik, taimedel mehhaniline levimine taimtoiduliste abil nt. lehetäid). * Viiruse leviku eeldus: Parasiit saab eksisteerida vaid piisavalt tihedas peremehe populatsioonis. Plasmiid viirusesarnane moodustis bakteris, omadustelt enamasti neutraalne, mõnedes keskkonnatingimustes suurendavad bakteri elujõudu, mõnedes vähendavad, on olulised bakterite evolutsioonis geenide ülekandega. BAKTERID Bakteri rakud pole membraanseid organelle, tuuma asendab tuumapiirkond, kus on rõngasjas kromosoom. rakukest, limakapsel, ribosoomid, viburid, plasmiidid (väiksemad DNA rõngad)-sisaldab geene, mis ensüüme sünteesivad. Paljunevad pooldumsiega. Võivad spoore moodustada!
1. Bakterirakus leiduvad polümeraasid: ! Replikatsiooni viivad läbi DNA polümeraasid (erinevad eri ahelatel). Replikatsioonijärgne parandamine: polümeraasid. Valepaardumiste eemaldamiseks: DNA polümeraas III – lisab nukleotiide sünteesitava ahela 3’ otsa; suudab eemaldada viimase DNA-le lisatud nukleotiidi (eksonukleaasne aktiivsus) st valesti lisatud nukleotiid eemaldatakse ning asendatakse õigega: DNA üks parandusmehhanisme bakteris. Pärast vea eemaldamist replikatsioon jätkub. DNA polümeraas I – aeglasem, aga tal on nii 3’ 5’ kui ka 5’ 3’ eksonukleaasne aktiivsus; Replikatsiooni käigus tunneb ära RNA praimeri ja asendab selle desoksüribonukleotiididega; Parandab vead, mida ei suutnud parandada DNA polümeraas III On ka DNA polümeraas V ja IV – neil puudub 3’-5’ eksonukleaasne aktiivsus, seetõttu teevad nad palju vigu 2
Enne seda tuleb RNA'st mingisugused jupid, lõigud välja lõigata (splaising?). Sellisel juhul muutub DNA kloonimine sisuliselt võimatuks, sest bakterites sellist väljalõikamise mehhanismi ei ole. Mida teha? Retroviirused sünteesivad ensüümi pöördtranskriptaas kasutab matriitsina RNA'd ja sünteesib sellele vastava DNA koopia. Seda retroviiruste ensüümi saab ära kasutada. Kui tahame kloonida rekombinantse valgu tootmiseks bakteris mingit sellist eukarüootset geeni, mida tavapäraselt splice'takse, siis ei alusta kloonimist sellest, et isoleerime geeni ja integreerime geeni mingisse plasmiidi vaid alustame sellest, et puhastame rakkudest vastava geeni pealt sünteesitud ja juba ära splicetud mRNA. Nüüd katseklaasis kasutades pöördtranskriptaasi sünteesime sellele mRNA'le ühe kaheahelalise DNA koopia. Seda koopiat kutsutakse sidiDNA'ks (?). Ja kõik jätkub endisel viisil. 05/11/09
Restriktaaside abiga võime saada erinevaid DNA lõike – siduvate ja tömpide otsadega. Siduvate otstega fragmendid on ühekordsete ahelatega ning neid saab taas omavahel liita. Restriktaase toodavad bakterid enesekaitseks, purustades erinevaid DNA ahelaid, jättes sellele „kleepuvad otsad“. Need jupid liidetakse uuesti komplementaarsuseprintsiibi kohaselt. Erinevate DNA lõikude liitmisel saame rekombinantse DNA. 39. Plasmiidid Plasmiidid – bakteris asuvad DNA väikesed rõngasmolekulid, millel on põhiliselt ainevahetuslik tähtsus. Plasmiidid sisaldavad geene, mis on vajalikud bakteri kasvukeskkonna eripärast tulenevate ensüümide sünteesiks. Need aitavad lagundada ümbritsevas keskkonnas leiduvaid orgaanilisi aineid. See on vajalik toitumiseks, aga ka elutegevuseks kahjulike ainete lagundamiseks või nende toime vältimiseks
bakteripopulatsiooni revertante (bakterirakke, kes on võimelised kolooniaid moodustama histidiini-vabas keskkonnas) 76. Põhilised DNA reparatsioonimehhanismid rakkudes. Valgusest sõltuv fotoreaktivatsioon Väljalõikereparatsioon (excision repair) Replikatsioonijärgne valepaardumisi kõrvaldav DNA “mismatch” reparatsioon MMR (mismatch repair) Rekombinatsiooniline reparatsioon 77. SOS vastus bakterites. Bakteris E. coli põhjustab SOS mutageneesi vigaderohke DNA polümeraas V, mis jätkab kahjustuse kohal peatunud DNA polümeraasi III asemel vigaderohket DNA sünteesi. See mehhanism käivitub ebasoodsates oludes. Vigaderohke DNA sünteesi tõttu toimub palju mutatsioone, millest osad võivad osutuda kasulikeks antud rasketes oludes. 78. Ristsiirde toimumise mehhanism Holliday mudeli põhjal.
viljakusega, antiensüümide tootmisega (sooleussid) Peremees tõrjeviisidega (linnud liivavannidega, suslikud pujude söömisega) On ka ruumi, pesa- ja transpordiparasiidid. Ektoparasiidid parasiit toitub või eluneb peremehe keha pinnal (peatäi, voodilutikas). Endoparasiididparasiit elutseb peremehe kehas (limuksolge, tuberkuloosibakter inimeses, bakteriofaag bakteris). o antibioos, organismide (pms.) mikroobide suhe, mille puhul üks liik mõjub teisele pärssivalt eritiste või laguainete vahendusel. Kokkusaamise korral Lahus elu korral A B A B - 0 0 0 (B eritab A-le ohtlikke aineid) Laiemas, vähem kasutatavas tähenduses on antibioos sama mis antagonism
On ka selliseid faage, mis on võimelised ründama tervet mikroobide perekonda või ka erinevatest perekondadest pärinevaid mikroobe. Bakteriofaagid jagunevad kahte suurde rühma: 1) Virulentsed faagid põhjustavad bakteriraku purunemist: 2) Mõõdukad ehk tempereeritud faagid need on algses genoomi soikeseisundis, ning hiljem aktiviseeruvad. Kui mõõdukas faag nakatab bakteriraku, siis esiteks võib bakter lüüsuda, teiseks lüsogeenne bakter. Lüsogeenses bakteris on faagi DNA bakteri genoomis, kuid ta pole aktiivne. Sellises vaikivas olekus genoomi kutsutakse profaagiks. Profaag on integreerunud genoomi ja asetub seal kindlas lookuses. Lüsogeenne bakterirakk ei nakatu teise bakteriofaagiga. Kuid UV-kiirgus, keemilised ained, võib toimuda profaagi genoomist väljalõikamine ja bakter võib selle tagajärjel lüüsuda. Bakteriofaagide levik Võib leida kõikjal, kus on mikroobe: vees, mullas, väljaheidetes, toiduainetes jne. On
On ka selliseid faage, mis on võimelised ründama tervet mikroobide perekonda või ka erinevatest perekondadest pärinevaid mikroobe. Bakteriofaagid jagunevad kahte suurde rühma: 1) Virulentsed faagid põhjustavad bakteriraku purunemist: 2) Mõõdukad ehk tempereeritud faagid need on algses genoomi soikeseisundis, ning hiljem aktiviseeruvad. Kui mõõdukas faag nakatab bakteriraku, siis esiteks võib bakter lüüsuda, teiseks lüsogeenne bakter. Lüsogeenses bakteris on faagi DNA bakteri genoomis, kuid ta pole aktiivne. Sellises vaikivas olekus genoomi kutsutakse profaagiks. Profaag on integreerunud genoomi ja asetub seal kindlas lookuses. Lüsogeenne bakterirakk ei nakatu teise bakteriofaagiga. Kuid UV-kiirgus, keemilised ained, võib toimuda profaagi genoomist väljalõikamine ja bakter võib selle tagajärjel lüüsuda. Bakteriofaagide levik Võib leida kõikjal, kus on mikroobe: vees, mullas, väljaheidetes, toiduainetes jne. On