b) Tõsta DNA sünteesiaega kolmelt minutilt nelja minutini c) Vähendada PCR-i tsüklite arvu 30-It 24-ni d) Tõsta praimerite liitumistemperatuuri 52°C-It 56°C-ni e) Tõsta DNA denaturatsiooni temperatuuri 94°C-It 96°C-ni 8. Milline järgnevatest väidetest on tõene tsirkulaarse kaheahelalise DNA genoomi osas, mille puhul on keemiliselt määratud, et see sisaldab 21 % adenini ? a) Genoomis on 21 % guaniini b) Guanini % ei saa niimoodi määrata c) Genoomis on 29 % guaniini d) Genoomis on 10.5 % guaniini e) Genoomis on 58 % guaniini 9. Rakulisel (in vivo) DNA kloonimisel on võimalik meid huvitava geenijärjestuse sisestamiseks plasmiidvektorisse kasutada erinevaid ensüüme. Milliseid ensüüme järgnevast valikust saaks rakendada: a) DNA topoisomeraas I b) DNaas I c) DNA polünukleotiid kinaas d) DNA ligaas e) T7 DNA polümeraas 10
2. renaturatsioon: kõrgemat järku struktuuride taastumine 8. DNA ehitus ja ülesanded. DNA - desoksüribonukleiinhape - mitokondrites, rakutuumas, kloroplastides Ehitus: kaks heeliksisse keerdunud ahelat, mis on kokku pandud nukleotiididest. Nukleotiidid on moodustunud desoksüriboosist, fosfaatrühmast ja lämmastikualusest. Nukleotiidid omakorda sisaldavad 4 lämmastikalust: adeniin, guaniin, tsütosiin, tümiin. (adeniini ja tümiini vahel on kaksikside, tsütosiini ja guaniini vahel kolmikside -- nukleotiidid on seotud vesiniksidemetega). DNA ülesanded: päriliku info säilitamine ja täpne edasiandmine 9. RNA ehitus ja ülesanded. RNA - ribonukleiinhape - mitokondris, rakutuumas, tsütoplasmas, ribosoomis, kloroplastis Ehitus: 1 ahel, kohati 2 ahelat, koosneb nukleotiididest, mis koosnevad samuti kolmest osast nagu DNA: lämmastikalusest, fosfaatrühmast ja riboosist. Samuti 4 lämmastikualust: adeniin,
kandjad e pärilikkusained Monomeer – polümeeri ehitusüksus; moodustab teiste omasugustega liitunult polümeerse molekuli Nukleotiid – nukleiinhappe ehitusüksus; koosneb suhkrust, fosfaatrühmast ja lämmastikalusest; suhkruks on RNA koostises riboos ja DNA-l desoksüriboos Komplementaarsusprintsiip – lämmastikaluste paardumise seaduspära; (nt ühe DNA-ahela adeniini vastas on alati teise ahela tümiin ja guaniini vastas tsütosiin) Kromosoom – terviklik DNA-molekul ja sellega seotud valgud Kromatiin – rakutuumas asuv pärilikkusaine koos selle pakkimises osalevate valkudega Tuumake – rakutuuma piirkond, kus sünteesitakse ribosoomi-RNAd(rRNA) ja moodustuvad ribosoomid Geen - DNA-molekuli lõik, mis kodeerib valku või määrab mingi RNA-molekuli sünteesi. Valku kodeerivate geenide järgi sünteesitakse mRNA. Geenid asuvad kromosoomides.
klorokiin on selle sünteetiline analoog. Toime põhineb malaariaparasiidi DNA transkriptsiooni blokeerimisel. Iseloomulik struktuur võimaldab interkaleeruda malaariaparasiidi DNA-ga. Aktinomütsiin D ja Adriamütsiin on antibiootikumid, mis sisaldavad vastavaid rühmi, millega töötavad interkalaarsete agentidena. Alküleerivad agendid Tugevad elektrofiilid. DNA-s on mitmeid nukleofiilsed gruppe, millest olulisim on guaniini 7. lämmastik. Agendid on väga reaktiivsed ning kasutatakse vähiravimitena. Ravimil peab olema kaks alküleerivat gruppi, mis reageerivad kahe lähestikku asetseva, kuid erinevates DNA ahelates paikneva G-ga, ristseovad kaks ahelat, blokeerides seega replikatsiooni ja transkriptsiooni. Samuti võib ravim siduda kaks guaniini samast ahelast, mistõttu takistab ka selline klammerdumine ligipääsu DNA ahelale. Alküleeritud guaniini
kõõlused , kaitse antikehad verehüümimisvalgud .valgud annavad energiat . NUKLEIINHAPPED DNA desoksüribonukleiinhape ,asub rakutuumas , kannab geenilist infot RNA Ribonukleiinhape , kopeerib ja salvestab DNA infot . Koosnevad mõlemad nukeotiididest . DNA ehitus · Sõltub nukeotiidide järjekorrast ja arvust . · On 4 erinevat nukeotiidi A sisaldab adeniini G- sisaldab guaniini C sisaldab tsütosiini T- sisaldab sümiini DNA molekul koosneb kahest ahelast , mis püsib koos üksteisele vastavuse põhimõtte alusel A -> T G -> C Selline ehitus muudab nad vastupidavaks ning tagab päriliku info esinemise vähemalt kahes koopias Nt : A C C T G G - A | | | | | | | T G G A C C T
sattunud. Ajapikku kaitsekiht kasvab , millega pakseneb ka pärlmutterkiht. Pärlikasvatuses ehk pärlifarmis kasvatatud pärlid on ainukesd ,,looduslikud pärlid", mida müüakse juveeliturgudel. Segadusse aetakse aga tänapäeval inimesi(kes soovivad endale osta loodusliku protsessi käigus arenenud pärlit) tehispärlite ehk pärliimitatsioonidega, mis on valminud tehastes kunstlikul teel. Pärliimitatsiionid on kaetud kalaluudest saadava guaniini ja laki seguga. Igastahes on tänapäeval pärlikasvatus kasulik mitmetel põhjustel: 1) pärlikasvatus päästis pärlikaubanduse väljasuremisest 2)pärlikasvatus päästis pärlikarpide ja austrite liigi väljasuremisest ja hävitamisest 3)pärlikasvatuses valmivad kvaliteetsemad ja looduslikumad pärlid, mis võimaldab ka inimestel endale muretseda ,,looduslikke pärleid" 4)pidurdab kunstlikke pärlite levikut ,,looduslike pärlite" olemasoluga, mis ei
Ta olevat ükskord külaliste silme all visanud muna- suuruse pärli äädikasse,kus see kiiresti lahustus.Siis joonud Kleopatra selle äädika ära. Ilmselt oli see maailma kalleim jook,sest mõnes lonksus mitte eriti kanges äädikas oli lahustunud 10 000 000 sestertsi ehk ligikaudu 60 000 rubla suurune varandus. Tänapäeval valmistatakse pärli imitatsioone klaasist ja tselluloidist.Nendest ainetest kuulikesed värvitakse üle kalasoomustest toodetava orgaanilise aine guaniini essentsiga ning eemalt on nad tõeliste pärlitega üsna sarnased. Pärlite " ravimine " s.t. tuhmunud pärlite läike taastamineon aktuaalne olnud üle aastatuhande.Väidetavalt pidi pärli tuhmunud helk kaduma kui seda soojendada lambarasvas või " ravida " piimaga.Samuti pärli iga pikendab nende kandmine. Pärlite kasutamine Pärle püütakse peamiselt nööpide valmistamiseks, s.o. pärlimutri pärast.Sajandeid
Aktivaatorid seonduvad DNA biheeliksi ''suurele vaole'' ja neil on teatud tunnusmotiivid. Helix-turn-helix;helix-loop-helix;zinc-finger;leucine zipper 6. Kuidas toimub ja milleks kasutatakse DNA metüleerimist imetaja genoomis? Loetle nähtusi, mis on seotud DNA metüleerimisega? Dna metüleerimine - tsütosiini viiendasse positsiooni lisatakse metüülrühm 5 metüültsütosiin. Protsessi viib läbi dna-polümeraas. Eelistatult metüleeritakse guaniini ees asuv tsütosiin cg. Dna metüleerimine pärsib geeni ekspressiooni. Metüleerimine on seotud geeni regulatsiooni, kromatiini struktuuri, X-inaktivatsiooni, epigeneetilise protsesside ja genoomse imprintingu nähtustega, imetaja genoomis ~3% tsütosiiinidest (-c-) metüleeritud 7. Mis on genoomne imprinting? Kus ja millal toimub imprintimine ja millega see nähtus on seotud loomulikult ning patoloogia korral? Nähtus, kus sama geen ekpresseerub erinevalt sõltuvalt
DNA tertsiaarstruktuur - nukleiinhappe ruumiline struktuur DNA kvaternaarstruktuur- kompleksid valkudega (kromatiin), kõrgemat järku nukleiinhapete organisatsioon Lämmastikalused paarduvad omavahel vesiniksidemete kaudu. Paarid moodustuvad puriinide ja pürimidiinide vahel komplementaarsusprintsiibi alusel. A=T(DNA) või U(RNA), G=C. A ja T kaks H-sidet, G ja C kolm. G:C piirkonnad on stabiilsemad. Chargraffi reeglid Adeniini hulk võrdub tümiini hulgaga, samamoodi guaniini hulk tsütosiiniga Adeniini ja guaniini hulk moodustab 50% kogu lämmastikaluste hulgast 4. RNA struktuurielemendid lämmastikalus (A, C, G või U) riboos, mille 2’ –OH rühm muudab RNA reaktiivsemaks kui DNA fosfaatrühm RNA võib moodustuda üheahelalise (nt hairpin) või kaheahelalise sekundaarstruktuuri või hoopis DNA-RNA dupleksi. Esineb ka tertsiaarstruktuuri. mRNA on kaetud 5’ otsast metüülguanosiinmütsikesega (cap), 3’ otsas on polü-A saba
ja RNA. Olenevalt sellest, kas nukleiinhapete koostisse kuulub riboos või desoksüriboos, jaotatakse neid ribonukleiinhapeteks (RNA) ja desoksüribonukleiinhapeteks (DNA). Nad erinevad üksteisest ka nende koostises leiduvate lämmastikaluste poolest. Lämmastikaluseid on kokku viis: puriinalused - adeniin (A) ja guaniin (G); pürimidiinalused - tümiin (T), tsütosiin (C) ja uratsiil (U). Lämmastikalustest esineb RNA molekulis adeniini (A), guaniini (G), tsütosiini (C) ja uratsiili (U), kuid puudub tümiin (T). 4.DNA ja RNA ehituse põhiprintsiibid. DNA paikneb rakutuumas kromosoomides. Erandiks on munarakud, kus osa DNA-st paiknebka tsütoplasmas. DNA sisaldab adeniini, guaniini, tsütosiini ja tümiini (puudub uratsiil). DNA on biheeliksi kujuline (kujutab endast kaksikspiraali, mis on pöördunud kellaosuti liikumise suunas) ning üksikute molekulide vahekaugus on konstantne (0,34 nm)
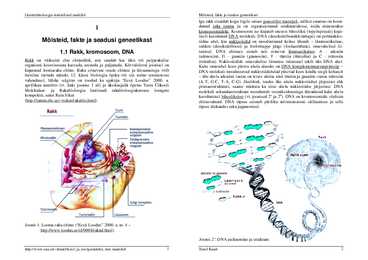
organismi koostisosana kasvada, areneda ja paljuneda. Kõrvaloleval joonisel on Kahe omavahel koos püsiva ahela aluseks on DNA komplementaarsusprintsiip kujutatud loomaraku ehitus. Raku erinevate osade ehituse ja ülesannetega võib DNA-molekuli moodustavad nukleotiidahelad püsivad koos kindla reegli kohaselt huviline tutvuda näiteks 12. klassi bioloogia õpiku või siis mõne teatmeteose ühe ahela adeniini vastas on teises ahelas alati tümiin ja guaniini vastas tsütosiin vahendusel, lühike selgitus on toodud ka ajakirja "Eesti Loodus" 2000. a. (A-T, G-C, T-A, C-G). Järelikult, teades ühe ahela nukleotiidset järjestust ehk aprillikuu numbris (vt. linki joonise 1 all) ja üksikasjalik õpetus Tartu Ülikooli primaarstruktuuri, saame määrata ka teise ahela nukleotiidse järjestuse. DNA
Nukleiinhapped DNA ja RNA, nukleotiidide polümeerid; päriliku info kandjad e pärilikkusained Monomeer polümeeri ehitusüksus; moodustab teiste omasugustega liitunult polümeerse molekuli Nukleotiid nukleiinhappe ehitusüksus; koosneb suhkrust, fosfaatrühmast ja lämmastikalusest; suhkruks on RNA koostises riboos ja DNA-l desoksüriboos Komplementaarsusprintsiip lämmastikaluste paardumise seaduspära;(nt ühe DNA-ahela adeniini vastas on alati teise ahela tümiin ja guaniini vastas tsütosiin) Kromosoom terviklik DNA-molekul ja sellega seotud valgud Kromatiin rakutuumas asuv pärilikkusaine koos selle pakkimises osalevate valkudega Tuumake rakutuuma piirkond, kus sünteesitakse ribosoomi-RNAd(rRNA) ja moodustuvad ribosoomid Genoom liigiomases ühekordses kromosoomikomplektis sisalduv geneetiline materjal Genotüüp organismi kõigi pärilike tegurite kogu ja koostoime Aluspaar kaks omavahel vesiniksidemetega seotud nukleotiidi, mis esinevad
teatav sümbol. Kõik kodeeritavad aminohapped on -aminohapped. Kodeeritavad aminohapped jagunevad asendamatuteks ja asendatavateks aminohapeteks. Asendamatud aminohapped. Evolutsiooni käigus on kõrgemad loomad,sealhulgas inimene,loobunud mõndade aminohapete valmistamisest ning hangivad need toiduga. Kokku on neid 8. asendamatuid aminohappeid sünteesivad taimed ja bakterid. Puriin-kahe aromaatse tuumaga orgaaniline alus, mh. nukleiinhapetes esineva adeniini ja guaniini struktuurne osa. Puriinaluste formaalne lähteühend, millest adeniin ja guaniin kuuluvad nukleiinhapetesse Pürimidiin-kuulub rakkude geneetilise materjali (DNA ja RNA). Hulka. Pürimidiinaluste formaalne lähteühend, millest tsütosiin, tümiin ja uratsiil kuuluvad nukleiinhapetesse. 14. Valkude klassifikatsioon: Lihtvalgud e proteiinid (koosnevad ainult aminohapetest) a) Fibrillaarsed valgud ehk skleroproteiinid- lahustumatud loomsed valgud tugi- ja
DNA-ligaas mahajääva ahela fragmentide ühendamine. Kuna uute polünukleotiidahelate süntees toimub 5' 3' suunas, siis üks ahel kasvab pidevalt juhtiv ahel (leading strand), teine aga lühikeste fragmentidena mahajääv ahel (lagging strand, uurija nime järge tuntakse neid kui Okazaki fragmente), mis seejärel ühendatakse. RIBONUKLEIINHAPPED (RNA): Sisaldavad -D-riboosi suhkru molekuli jäägina; Lämmastikalustest sisaldavad adeniini (A), guaniini (G), tsütosiini (C) uratsiili (U); Üheahelalised molekulid. Kuna enamasti N-alused pole paardunud, ei sisaldu komplementaarseid aluseid ka võrdsetes hulkades; Molekulmassid oluliselt väiksemad kui DNA molekulidel. Nukleotiidide arv vahemikus 75 ... mõni tuhat. RNA liigid: Ribosoomi RNA (rRNA). Seostub valkudega kompleksideks RIBOSOOMIDEKS. Ribosoomide molekulmassid on suurusjärgus 3x106; Informatsiooni- e matriits-RNA (mRNA). Kannab geneetilist informatsiooni DNA- lt ribosoomidele
Seega ei saa päris kindel olla, et aknet põhjustab Propionibacterium. Alati leitakse aknega haigete nahalt ka Staphylococcust. · P. acnes't on nahal v. palju, eriti palju seal, kus on rohkesti rasunäärmeid: laubal ja ninakülgedel. · Propionibaktereid on leitud ka tupest, suust ja väljaheidetest. · Naha propioonhapebakterid on v. nõudlikud söötme suhtes: vajavad peaaegu kõiki aminohappeid, paljusid vitamiine ja paljud ka guaniini ja adeniini. Perekond Micromonospora Morfoloogia: · Esineb vaid substraadimütseel. Koloonia on pinnalt kurruline, spoorid paiknevad üksikult mütseelil. Elupaik looduses: · neid leidub mullas ja vees. Kodudes õhu saastajad, elavad ka konditsioneerides. · Lagundavad biopolümeere (tselluloosi, ksülaani, kitiini). Biotehnoloogiline rakendus: · mitmed Micromonospora toodetud antibiootikumid on kasvajavastased
neid nukleotiitide alguseid ja lõppe RNAs nad oleksid CU (tsütosiin, uratsiil) ja AC (adeniin, tsütosiin). 39. RNA protsessimine. Protsess nimega ,,RNA editing" muudab transkriptis sisalduva info hulka, muutes aluspaaride järjestust asenduste teel. Algset staadiumi RNA protsessingus katalüüsib dimeerne capping ensüüm, mis seostub RNAPolII fosforüülitud CTDga. Viimases staadiumis kannavad erinevad ensüümsed alaühikud üle metüülrühmad S¬- adenosüülmetioniinilt guaniini asendis N7 olevale lämmastikule ning sünteesitud RNA 5'otsas paiknevate ribooside 2'oksügeenidele. Muudab CAA koodoni UAA poolt kodeeritud Stop koodoniks ning lõpeb poole lühema valgu apoB-48 sünteesiga. 40. Vaid puuviljakärbse emasisendid sünteesivad funktsionaalset SXL (sex lethal) valku, mis represseerib eksonite 2 ja 3 splaisingut sxl pre-mRNAs ja eksonite 1 ja 2 splaisingut tra (transformer) pre-mRNAs. TRA-TRA2 kompleksi sidumine dsx (double-sex) pre-mRNAle
1) luhike elutsukkel (viirustel ja bakteritel 20...30 min, mikroseentel 1...2 tundi). See voimaldab uurida geneetilist sidet vaga paljude polvkondade jooksul. 2) suur paljunemiskiirus 3) suguta sigimise korval esineb ka suguline sigimine 4) lihtne kasvatada. Bakteritest koige enam on uurimistoos kasutanud soolekepikest Escherichia coli ja tema faage. DNA ehitus DNA paikneb rakutuumas kromosoomides. Erandiks on munarakud, kus osa DNA-st paikneb ka tsutoplasmas. DNA sisaldab adeniini, guaniini, tsutosiini ja tumiini (puudub uratsiil). E. Chargraff toestas1950.a, et puriin-ja purimidiinaluste suhe DNA-s on alati vordne: A+G=T+C ning adeniini hulk vordub tumiiniga (A=T) ja guaniini hulk tsutosiiniga (G=C). 1950-ndatel aastatel uuriti DNA struktuuri ja mitmed teadlaste toogrupid joudsid jareldusele, et DNA on biheeliksi kujuline (kujutab endast kaksikspiraali, mis on poordunud kellaosuti liikumise suunas) ning uksikute molekulide vahekaugus on konstantne (0,34 nm). DNA
7. Transkriptsioonijärgne geeniekspressiooni kontroll 55. Ribolülitid ja nende roll geeniekspressiooni kontrollis Ribolülitid — lühikesed RNA järjestused, mille kuju muutub, kui nendele seostuvad väiksed molekulid (näit. metaboliidid). kui mõnda ainet on rakus palju, seostub ribolüliti sellega, ribolüliti konformatsioon muutub ning ta katkestab transkriptsiooni, mis toodaks antud metaboliiti juurde. paiknevad vastsünteesitud mRNA 5’-otsa läheduses. NÄITEKS: kui guaniini tase rakus on madal, siis elongeeriv RNA polümeraas transkribeerib puriini biosünteesi geene ja ensüüme, mis on vaja guaniini biosünteesiks. Kui guaniini on aga liiast, siis see seostub ribolülitile, põhjustades konformatsiooni muutuse, mis omakorda toob kaasa RNA plümeraasi termineerimise. 56. RNA splaissing, RNA alternatiivne splaissing, RNA alternatiivse splaissingu negatiivne ja positiivne kontroll RNA SPLAISSING
Ribosoomi sees tRNA- d liiguvad alguses A, siis P ja lõpuks E saiti. Ribosoomi subühikud liiguvad teineteise suhtes iga elongatsioonitsükli käigus. Elongatsioonil osalevad translatsiooni elongatsiooni faktorid, mille hulka kuuluvad 3 universaalset faktorit: EF-Tu (+ GTP) – vahendab aminoatsüül-tRNA 13 sisenemist ribosoomi vabasse A-saiti; EF-Ts – EF-Tu guaniini nukleotiidivahetusfaktor, katalüüsides GDP vabanemist EF-Tu-lt; EF-G (+ GTP) – katalüüsib tRNA ja mRNA translokatsiooni mööda ribosoomi pärast iga polüpeptiidi elongatsioonitsükli lõppu. Terminatsioon Terminatsiooni käigus peab vabanema valmissünteesitud peptiidahel ja ribosoomid tuleb vabastada kõigist tema ligandidest. Terminatsiooni- (ehk nonsense) ehk STOP-koodoneid on 3: • UAG - amber • UAA - ochre • UGA - opal
Eukarüootse pre-mRNA protsessimise jooksul toimub 3 peamist sündmust: (1) 5' capping 5' cap lisatakse vahetul pärast transkriptsiooni initsiatsiooni. Capping on iseloomulik vaid mRNAdele. Capping ensüüm seondub fosforüülitud RNAPolII CTD'ga. Ensüümi fosfotaasi aktiivsusega alaühik eemaldab ühe terminaalse fosfaatrühma. Seejärel lisatakse terminaalsetele fosfaatidele GTP, kaotades 2 protsessi käigus 2 GTP fosfaatrühma. Tulemuseks on 5'-5' trifosfaatne struktuur. Guaniini 7N asendis olevale lämmastikule kantakse üle metüülrühmad ning RNA 5' otsas paiknevate ribooside 2' oksügeenidele. (2) 3' lõikamine ja polüadenüülimine. Peaaegu kõik mRNAd sisaldavad järjestust AUAAA. Mis asub polü(A) sabast 10-35bp ülavoolu. Lõikamis- ja polüadenüülimis-spetsiifiline faktor (CPSF), mis koosneb neljast erinevast valgust, moodustab kõigepealt AAUAAA järjestusest ülavooli seonduva ebastabiilse kompleksi.
Võivad vahendada eksonite vahetust, kuna aktsepteerivad mobiilsete elementide insertsioone. Mõnikord väga pikad (>1Mb). Tekkinud koos eukarüootsuse tekkega, Grupp I ja II intronid esineb keeruline sekundaarstruktuur. On isesplaisuvad. Esinevad nii bakterites kui eukarüootides, esmajoones tRNA ja rRNA geenides. Võivad käituda mobiilsete elementidena. Grupp I ja II erinevad splaisingusignaalide poolest. Grupp II omab RT aktiivsust, grupp I vajab splaisinguks vaba guaniini, Arhebakterite intronid esinevad tRNA ja rRNA geenides. Konserveerunud struktuur ning self-splaising puuduvad. Esinevad valgulised splaisingufaktorid, puuduvad trans-toimivad RNA molekulid. Intronite päritolu: Intronid esimesena (intron first hypothesis) postuleerib, et intronid ja RNA splaising on jäänukid RNA maailmast ja eelnesid prokarüootsetele introniteta geenidele, mis kaotasid splaisingu võites efektiivse geeniekspressiooni, Intronid
Metülatsioon- lämmastikaluse aminorühmale DNA iseeneslik kahjustus. Depuriinimine ja desamiinimine hüdrolüüsi teel. Depuriinimine-lämmastikaluse eemaldumine suhkru küljest Desamiinimine- lämmastikaluse aminorühma asendamine hapnikuga Radiatsiooni ja kemikaalide poolt põhjustatud DNA kahjustused. Tümiini dimeeride ja O6-metüülguaniini teke ja parandamine. Kõrvutiasetesevate tümiinide vahele tekib side, sideme lagundab fotoreaktiivne ensüüm O6-metüülguaniini puhul tooimub guaniini alküleerimine(metüülgrupp lisatakse hapniku juurde). Metüültransferaas parandab vea. Aktiivtsentris tsüsteiin seotakse metüülrühmaga. Kaks põhilist DNA parandamise viisi (lämmastikaluste ja nukleotiidide välja lõikamine). Lämmastikaluse väljalõikamine- DNA glükosülaad eemaldab lämmastikaluse, AP endonukleaas ja fosfodiesteraas eemaldavad suhkru ja fosfaadi. DNA polümeraas lisab uue nukleotiidi ja ligaas kinnitab.
ja importiini vabanemine Ran importiin liiguvad läbi NPC tagasi tsütoplasmasse, kus toimub hüdrolüüs ja mõlemi komponendi vabanemine (11-37 lk.434) Ran valk annab energiat transpordiks, sest hüdrolüüsib GTP. Ran võib olla seotud GTP ja GDP-ga. Seda kontrollivad täiendavad valgud. GTPaasi aktiveeriv valk GAP (ingl GTPase activating protein) hüdrolüüsib GTP ja muudab Ran GTP Ran GDPks. Tuuma guaniini vahetusfaktor GEF (ingl guanine exchange factor) vahetab GDP GTP-ks. Seega tuumatranspordis osalevad mitmesugused GTP/GDP ainevahetusega seotud valgud. Nii nagu valgu ekspordi puhul, ka impordi puhul on oluline Ran GTP valgu (tagab GTPGDP) tsütoplasmaatiline lokalisatsioon ja RCC1 valgu (GDPGTP) lokalisats. tuumas. Kõigil retseptorvalkudel on tertsiaarstruktuuris piirkonnad nukleoporiinide ja Raniga seostumiseks. Erinevat tüüpi NLS ja NES (Nuclear export signal) jaoks on erinevad
T, C ja U puhul on tegemist pürimidiinidega, kus lämmastikuaatomid moodustavad ühe tsükli. 53 DNA kaksikahela ehitus 1953. a. kirjeldasid James Watson ja Francis Crick DNA ruumilise struktuuri. Mudeli loomisel lähtusid nad järgmisest informatsioonist: 1) Erwin Chargaff kolleegidega analüüsis DNA koostist. Segus, et tümiini kontsentratsioon vastas alati adeniini kontsentratsioonile ja tsütosiini kogus guaniini hulgale. Samuti näitasid nad, et pürimidiinide summaarne kontsentratsioon DNA molekulis vastas kõigil juhtudel puriinide summaarsele kontsentratsioonile. Samas oli molaarne suhe T + A/ C + G liigiti erinev. 2) Maurice Wilkins'i ja Rosalind Franklin'i poolt saadud DNA röntgenstruktuuri analüüsi tulemused. DNA kristallstruktuuri uuringud näitasid, et DNA on 2-ahelaline, kusjuures ümber molekuli telje kordub regulaarselt (iga 0,34 nm järel) teatav alamstruktuur.
A ja G on 2-tsüklilised lämmastikalused - puriinid. T, C ja U puhul on tegemist pürimidiinidega, kus lämmastikuaatomid moodustavad ühe tsükli. DNA kaksikahela ehitus 1953. a. kirjeldasid James Watson ja Francis Crick DNA ruumilise struktuuri. Mudeli loomisel lähtusid nad järgmisest informatsioonist: 1) Erwin Chargaff kolleegidega analüüsis DNA koostist. Segus, et tümiini kontsentratsioon vastas alati adeniini kontsentratsioonile ja tsütosiini kogus guaniini hulgale. Samuti näitasid nad, et pürimidiinide summaarne kontsentratsioon DNA molekulis vastas kõigil juhtudel puriinide summaarsele kontsentratsioonile. Samas oli molaarne suhe T + A/ C + G liigiti erinev. 2) Maurice Wilkins'i ja Rosalind Franklin'i poolt saadud DNA röntgenstruktuuri analüüsi tulemused. DNA kristallstruktuuri uuringud näitasid, et DNA on 2-ahelaline, kusjuures ümber molekuli telje kordub regulaarselt (iga 0,34 nm järel) teatav alamstruktuur.
tekkinud mutatsiooni avaldumise. 71. Mutatsioonisagedust mõjutavad tegurid. · DNA replikatsioonitäpsus · DNA reparatsiooni efektiivsus · Mutageensete faktorite olemasolu ja hulk keskkonnas: UV, röntgen, kemikaalid · Korduvjärjestused soodustavad DNA sünteesil indelite teket · Lämmastikaluste tautomeersed üleminekud võivad põhjustada asendusmutatsioone · Rakus metabolismi käigus tekkivad O2 radikaalid võivad oksüdeerida guaniini. Somaatilistes rakkudes tekkinud mutatsioonid ei pärandu edasi. Sugurakkude arengu alguses tekkinud läheb mitmele järglasele. Lõpuks saab vaid üks järglane mutatsiooni. 72. Spontaansed ja indutseeritud mutatsioonid. Mutatsioonitekke juhuslikkust tõendavad katsed. Spontaansed tekivad iseenesest, sagedus on madal Indutseeritud mutageenide toimel tekkinud mutatsioonid, sagedus mitu suurusjärku kõrgem. UV, kemikaalid
Eukarüootse pre-mRNA protsessimise jooksul toimub 3 peamist sündmust: (1) 5' capping 5' cap lisatakse vahetult pärast transkriptsiooni initsiatsiooni. Capping on iseloomulik vaid mRNAdele. Capping ensüüm seondub fosforüülitud RNAPolII CTD'ga. Ensüümi fosfotaasi aktiivsusega alaühik eemaldab ühe terminaalse fosfaatrühma. Seejärel lisatakse terminaalsetele fosfaatidele GTP, kaotades 2 protsessi käigus 2 GTP fosfaatrühma. Tulemuseks on 5'- 5' trifosfaatne struktuur. Guaniini 7N asendis olevale lämmastikule kantakse üle metüülrühmad ning RNA 5' otsas paiknevate ribooside 2' oksügeenidele. (2) 3' lõikamine ja polüadenüülimine. Peaaegu kõik mRNAd sisaldavad järjestust AUAAA. Lõikamis- ja polüadenüülimis-spetsiifiline faktor (CPSF), mis koosneb neljast erinevast valgust, moodustab ebastabiilse kompleksi. Seejärel lisanduvad kompleksile vähemalt 3 valku: lõikamist
Vatsabarjääri kahjustamise tõttu väheneb ka kaitse teiste patogeenide ja toksiinide 9 vastu lisaks mükotoksiinidele. Mükotoksiinide mõju vatsaseedele võib olla võrreldav vatsaatsidoosiga (Fink-Gremmels, 2008). 3.4.1. Aflatoksiinid Aflatoksiin läbib suures osas vatsa muutumatult ning ka metaboliseerumisel ei vähene selle toksilisus oluliselt, mistõttu toksiin on ohtlik ka mäletsejalistele. Sidudes guaniini toimib aflatoksiin DNA-d kahjustavalt ja põhjustab kas raku surma või selle muteerumise kasvajarakuks. On leitud kasvuiibe langust lihaveistel alates kogusest ratsioonis üle 0,7 g/kg, maksaturset kogusest üle 0,1 g/kg, sigimislangust piimalehmadel koguses 0,12 g/kg ning piimatoodangu lagust kuni 25% koguses üle 0,1 g/kg (Jouany1 ja Diaz, 2011). Osaliselt on kõik need ilmingud seotud aflatoksiini maksatoksilisusega, selle üheks omapäraseaks väljenduseks on fotosensibiseerumine pärast
71. Mutatsioonisagedust mõjutavad tegurid. · DNA replikatsiooni täpsus ning reparatsiooni efektiivsus · Mutageenide olemasolu ja hulk keskkonnas (UV, röntgenkiirgus, kemikaalid) · Korduvjärjestused (1 või mõni nukleotiid) soodustavad DNA sünteesil indelite (deletsioonide, duplikatsioonide) teket. · Lämmastikaluste tautomeersed üleminekud võivad põhjustada asendusmutatsioone. · Vabad hapniku radikaalid kahjustavad guaniini, mis seetõttu võib paarduda adeniiniga. 72. Spontaansed ja indutseeritud mutatsioonid. Mutatsioonitekke juhuslikkust tõendavad katsed. · Spontaansete mutatsioonide tekkesagedus on väga madal geeni kohta ~10 -4 10-7. · Indutseeritud mutatsioonid ehk mutatsioonid, mis on tekkinud mutageenide toimel tõstavad mutatsioonisagedust rakus võrreldes spontaanse mutatsioonisagedusega mitu suurusjärku.
uratsiili (U). A ja G on 2-tsüklilised lämmastikalused - puriinid. T, C ja U puhul on tegemist pürimidiinidega, kus lämmastikuaatomid moodustavad ühe tsükli. 3. DNA kaksikahela ehitus. 1953. a. kirjeldasid James Watson ja Francis Crick DNA ruumilise struktuuri. Mudeli loomisel lähtusid nad järgmisest informatsioonist: 1) Erwin Chargaff kolleegidega analüüsis DNA koostist. Segus, et tümiini kontsentratsioon vastas alati adeniini kontsentratsioonile ja tsütosiini kogus guaniini hulgale. Samuti näitasid nad, et pürimidiinide summaarne kontsentratsioon DNA molekulis vastas kõigil juhtudel puriinide summaarsele kontsentratsioonile. 2) Maurice Wilkins'i ja Rosalind Franklin'i poolt saadud DNA röntgenstruktuuri analüüsi tulemused. DNA kristallstruktuuri uuringud näitasid, et DNA on 2-ahelaline, kusjuures ümber molekuli telje kordub regulaarselt (iga 0,34 nm järel) teatav alamstruktuur.
(DNA). Nad erinevad üksteisest ka nende koostises leiduvate lämmastikaluste poolest. Lämmastikaluseid on kokku viis: puriinalused - adeniin (A) ja guaniin (G); pürimidiinalused - tümiin (T), tsütosiin (C) ja uratsiil (U). Lämmastikalustest esineb RNA molekulis adeniin (A), guaniin (G), tsütosiin (C) ja uratsiil (U), kuid puudub tümiin (T). DNA ehitus DNA paikneb rakutuumas kromosoomides. Erandiks on munarakud, kus osa DNA-st paikneb ka tsütoplasmas. DNA sisaldab adeniini, guaniini, tsütosiini ja tümiini (puudub uratsiil). E. Chargraff tõestas1950.a, et puriin- ja pürimidiinaluste suhe DNA-s on alati võrdne: A+G=T+C ning adeniini hulk võrdub tümiiniga (A=T) ja guaniini hulk tsütosiiniga (G=C). 1950. aastatel uurides DNA struktuuri jõudsid mitmed teadlaste töögrupid järeldusele, et DNA on biheeliksi kujuline (kujutab endast kaksikspiraali, mis on pöördunud kellaosuti liikumise suunas) ning üksikute molekulide vahekaugus on konstantne (0,34 nm). DNA
uratsiili. Uratsiil tekib tsütosiini deamineerimise tulemusena. Sellel ensüümil AP endonukleaasne aktiivsus puudub. 3-MeA DNA glükosülaasid I (Tag) ja II (AlkA) on erineva substraadispetsiifilisusega. AlkA on laiema substraadispetsiifilisusega, kõrvaldades nii 3-MeA, 3-MeG kui ka hüpoksantiini (hüpoksantiin tekib adeniini deamineerimisel). Tag ekspresseerub rakkudes konstitutiivselt, AlkA geeni transkriptsioon on aga samuti Ada valgu poolt reguleeritav. Oksüdeeritud guaniini GO reparatsioon Guaniini oksüdatsiooniproduktiks on 8-hüdroksüguaniin (GO). GO kahjustusi põhjustavad hapnikuradikaalid tekivad rakus ka tavaliste protsesside (elektronide transport, lipiidide peroksüdatsioon) tulemusena. Seetõttu on GO reparatsioon bakterirakus üks olulisemaid vigade ärahoidmise ja korrigeerimise süsteeme. GO-defektsetes tüvedes tõuseb mutatsioonisagedus võrreldavalt mutatsioonisageduse tõusuga, mis ilmneb replikatiivse DNA
(-NH2, =O ja tsükli 1. ja 3. positsiooni N või N-H). A=T ja A=U vahele moodustub kaks vesiniksidet, GC vahele kolm vesiniksidet. 9. Selgitage nn. Chargaff'i reegli (DNA molekulis A + G = T + C) struktuurset alust. Kas reegel kehtib ka RNA puhul? Põhjendus? Chargaff leidis, et DNA koostisesse kuuluvate lämmastikaluste vahekorras kehtivad alati kindlad reeglid: - puriin- ja pürimidiinaluste hulk on võrdne: A+G =T+C - adeniini hulk võrdub tümiiniga: A=T - guaniini hulk võrdub tsütosiiniga: G=C See reegel ei kehti RNA puhul, kuna RNA on üheahelaline. 10. Püüdke seletada a) miks kannab DNA happe nimetust sest DNA's nukleotiidid (need on polüprootsed happed) loovutavad prootoneid, et luua 3'-5'fosfodiestersidemeid, mis ühendavad nukleotiide nukleiinhapetes. b) miks hüdrolüüsub RNA leelises keskkonnas kergemini kui DNA. RNA hüdrolüüsub lahjas leelises keskkonnas, DNA on vastupidav leeliste suhtes, sest DNA'l puudub 2'-OH ja on
esinevad korduvjärjestustega regioonides. Tavaliselt patogeensed. · Kromosomaalsed vead numeraalsed ja struktuurilised, harvad ja patogeensed, tihti vähirakkudes. Sünonüümne mutatsioon aminohape ei muutu nukleotiidi asendudes; mittesünonüümne on tavalisem. · Tsütosiini deaminatsioon: C->U · Depurinatsioon viib deletsioonini · Alküleerivad ühendid rikuvad guaniini ära · Aluspaaride analoogia · T dimeerub UV toimel · metülatsiooni vead: adeniin -> hüpoksantiin · replikatsiooni, rekombinatsiooni vead Võib jagada veel funktsiooni kaotavad mutatsioonid (retesessiivsed nt geeni deletsioon, geenistr. muutus mRNA stabiilsuse muut, lugemisraami nihe, stoppkoodoni teke jne.) või funktsiooni muutvad (dominantsed üleekspressioon, retseptor pidevalt aktiveeritud, uus substraat, ioonkanalite funkts
geeni splaissingul. Tavaliselt fosforsidemeid üle kandes. DMD - 78 intronit, kana kollageen - vähemalt 50. Introneid lõigatakse välja: tRNA prekursoritest splaissingu endonukleaasiga + ligeerimine (fosfodiesteraasne, kinaasne, ligaasne aktiivsus) rRNA prekursoritest autokatalüütiliselt. pre-mRNA transkriptidest kaheetapiliselt splaissosoomide abil. 121. Autokatalüütiline splaising. Puudub trans; on ainult cis katalüütiline aktiivsus. Reaktsioon vajab guaniini vaba 3'-OH gruppi kui kofaktorit! Intron kõrvaldatakse kahe fosforsideme ülekandega. Väljalõigatud intron võib muuta ringikujuliseks teise fosforsidemete ülekandega. rRNA prekursorid. 122. Splaissosoomid. Splaissosoomides on 5 snRNPd: U1, U2, U4, U5, U6. SpRNAd - valk kompleksid. I etapis toimub 5' intron splaissingu saidis (tavaliselt GU; splice donor) katkemine, intramolekulaarsete fosfordiestersideme teke katkemispunkti G 5' süsinikku ja konserveerunud A 2?' süsinikku vahel
DNA molekul on kaksikspiraalse ehitusega st DNA sekundaarstruktuuri moodustavad kaks paralleelset ahelat, mis on omavahel vesiniksidemete abil ühendatud ja pöörduvad ümber ühise telje paremale. Seejuures on lämmastikalused paarunud nn komplementaarsusprintsiibist lähtudes: adeniin seostub kahe vesiniksideme abil tüminiiniga ja guaniin kolme vesiniksideme varal tsütosiiniga. Selletõttu ongi adeniini hulk elusrakus DNA-s alati võrdne tüminiini hulgaga, samuti tsütosiini hulk guaniini hulgaga. Seda tõsiasja, samuti seaduspärasust, et puriinaluste hulk rakus võrdub pürimidiinaluste hulgaga, nimetatakse Chargaffi reegliks. 5. DNA funktsioonid: Geneetilise info säilitamine muutumatul kujul ja edasiandmine tütarrakkudele raku jagunemisel, samuti valgusünteesiks vajaliku info ülekandmine RNA-le. 6. DNA replikatsioon: Replikatsioon on DNA kahekordistumine. Raku jagunemisele eelneva uue
dekodeerimises, geenide regulatsioonis ja ekspressioonis. ◦ mRNA: informatsiooni-RNA kannab DNA- lt valgusünteesiks vajaliku informatsiooni tuumast välja ribosoomi; ◦ tRNA: transport-RNA kannab aminohapped ribosoomi, kus geneetilise koodi alusel lisatakse need sünteesitavasse valguahelasse ◦ rRNA: ribosomaalne-RNA moodustab põhilise osa ribosoomidest ning viivad läbi valgusünteesi. Lämmastikalustest sisaldab adeniini (A), tsütosiini (C), guaniini (G) ja uratsiili (U). Erinevalt DNA-st on enamasti üheahelaline ja sisaldab riboosi. Lämmastikalused DNA ja RNA monomeeride nimetused: Lämmastikalus Monomeer Tähis DNA RNA Valem Adeniin Adenosiinfosfaat A X X C5H5N5 Guaniin Guanofosfaat G X X C5H5N5O Tsütosiin Tsütidiinfosfaat C X X C4H5N3O Tümiin Tümiinfosfaat T X - C5H6N2O2 Uratsiil Uridiinfosfaat U - X C4H4N2O2 Geneetiline kood
PKU võib avalduda erineval määral, sõltuvalt sellest, millise PAH alleeliga on tegemist. Mõnede mutatsioonide puhul jääb ensüüm osaliselt funktsionaalseks. PAH defektsust on võimalik ravida vastava dieediga, kus fenüülalaniini tase toidus on madal. Oluline on, et dieeti rakendataks imikutel kohe pärast sündimist. Lesh-Nyhan'i sündroom. Lesh-Nyhan'i sündroom (LNS) on samuti põhjustatud metabolismihäiretest ning avaldub juba imikutel. LNS on X-liiteline hüpoksantiini ja guaniini fosforibosüültransferaasi defektsusest tulenev geneetiline haigus. Kuna tegemist on retsessiivse alleeliga, on haiged tavaliselt poisslapsed. LNS-i põdevad haiged on vaimselt alaarenenud ning nende keha, eriti käed, teeb pidevaid aeglasi võnkuvaid liigutusi. Haiged kipuvad end ise kahjustama. See avaldub esmalt huulte ja suu seespoole närimises, hiljem asuvad nad ka oma käte kallale. Selleks, et LNS sündroomiga haigeid iseenda eest kaitsta, kõrvaldatakse neil hambad ning nad
ja ribosoomi- RNA .Olenevalt sellest, kas nukleiinhapete koostisse kuulub riboos või desoksüriboos, jaotatakse neid ribonukleiinhapeteks (RNA) ja desoksüribonukleiinhapeteks (DNA). Nad erinevad üksteisest ka nende koostises leiduvate lämmastikaluste poolest. Lämmastikaluseid on kokku viis: puriinalused - adeniin (A) ja guaniin (G); pürimidiinalused - tümiin (T), tsütosiin (C) ja uratsiil (U). Lämmastikalustest esineb RNA molekulis adeniini (A), guaniini (G), tsütosiini (C) ja uratsiili (U), kuid puudub tümiin (T). 40.TRIPLETT--kolm järjestikulist nukleotiidi DNA juhtahelas , KOODON --- kolm järjestikulist nukleotiidi mRNAs,mis kodeerivad ühte aminohapetest, ANTIKOODON---kolm mRNA koodonile komplementaarset nukleotiidi tRNAs. Geneetilise info koodiüksuseks on kolmest nukleotiidipaarist koosnev koodon(triplet).Esimesteks teadlasteks 1954 Gamow ja Crick avaldasid hüpoteese geneetilise koodi olemasolust
neil pole CTDd, siis capping on omane vaid mRNAdele. Capping ensüümi üks alaühikutest eemaldab _-fosfaadi sünteesitud RNA 5'otsast, s.o. RNAPolII kompleksist väljaulatuvast alast. Selle alaühiku teise domeeni aktiivsus lisab transkripti 5' otsa jäänud difosfaatrühmale GMP, nii et tekib unikaalne guanosiin 5'-5' trifosfaatne struktuur. Viimases staadiumis kannavad erinevad ensüümsed alaühikud üle metüülrühmad S-adenosüülmetioniinilt guaniini asendis N7 olevale lämmastikule ning sünteesitud RNA 5'otsas paiknevate ribooside 2'oksügeenidele. pre-mRNAde 3'lõikamine ja polüadenüülimine on omavahel tihedalt seotud protsessid Eukarüoodis on kõigil mRNAdel, v.a. histoonide mRNAd, 3' polü(A) saba. Primaarse transkripti 3' lõikamine eelneb polüadenüülimisele. Peaagu kõik mRNAd sisaldavad järjestust AUAAA, mis asub polü(A) sabast 10- 35bp ülavoolu
uratsiili. Uratsiil tekib tsütosiini deamineerimise tulemusena. Sellel ensüümil AP endonukleaasne aktiivsus puudub. 3-MeA DNA glükosülaasid I (Tag) ja II (AlkA) on erineva substraadispetsiifilisusega. AlkA on laiema substraadispetsiifilisusega, kõrvaldades nii 3-MeA, 3-MeG kui ka hüpoksantiini (hüpoksantiin tekib adeniini deamineerimisel). Tag ekspresseerub rakkudes konstitutiivselt, AlkA geeni transkriptsioon on aga samuti Ada valgu poolt reguleeritav. Oksüdeeritud guaniini GO reparatsioon Guaniini oksüdatsiooniproduktiks on 8-hüdroksüguaniin (GO). GO kahjustusi põhjustavad hapnikuradikaalid tekivad rakus ka tavaliste protsesside (elektronide transport, lipiidide peroksüdatsioon) tulemusena. Seetõttu on GO reparatsioon bakterirakus üks olulisemaid vigade ärahoidmise ja korrigeerimise süsteeme. GO-defektsetes tüvedes tõuseb mutatsioonisagedus võrreldavalt