Kompleksid kas moodustuvad või ei moodustu. 5. Missugused TAFid on TBPga vahetus interaktsioonis? TBPga seonduvad TFIIB, TFIIA TBP is a subunit of the eukaryotic transcription factor TFIID. 6. Missugune TFII valk hüdrolüüsib ATPd ja mis protsessiga on tegu? ATPd hüdrolüüsib helikaasse aktiivsusega TFIIH, mis kasutab ATP hüdrolüüsi energiat selleks, et DNA dupleks startsaidis lahti kerida. 7. Võrdle transkriptsiooni initsitatsiooni protsesse prokarüootidel ja eukarüoo-tidel. Prokarüoodid toimuvad tsütoplasmas. *RNA polümeraasi holoensüümi (2 , ß, ß', ) seondumine DNA promootorpiirkonda *RNA polümeraasi toimel lokaalne DNA-ahelate lahtikeeramine ja matriitsahela teke *Fosfodiestersidemete esmane teke tekkivas RNA ahelas, pärast 8 9 lülitust vabaneb - faktor ja põhiensüüm jätkab (2 , ß, ß') elongatsiooni - faktor tunneb ära -35 või -10 (promootori ülesvoolu, 5' poolne järjestus) konsensus-järjestust TTGACA
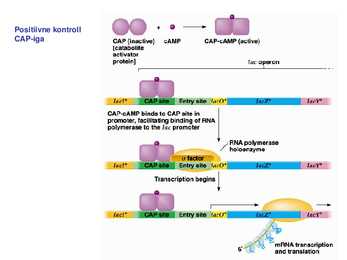
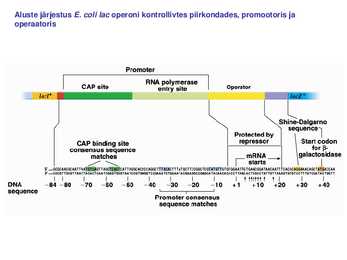
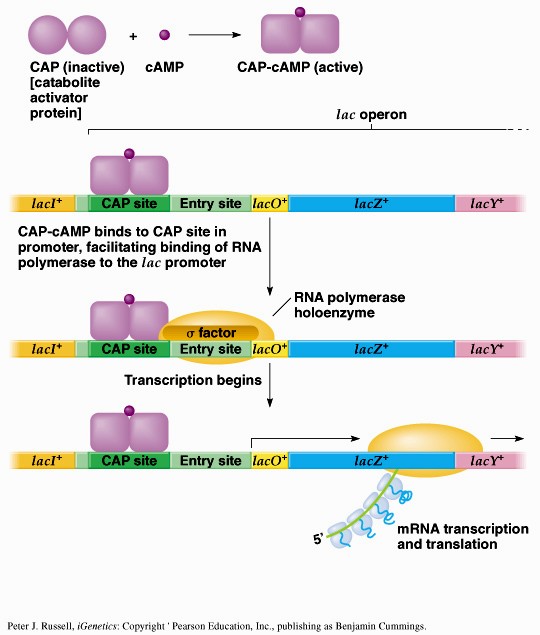
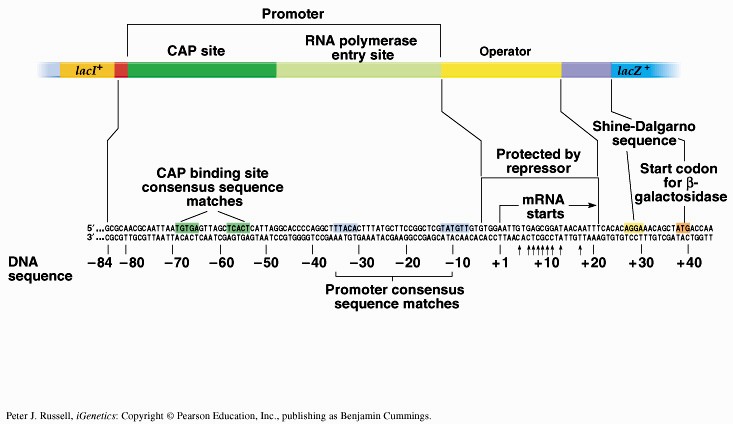
Sageli on transkriptsiooni initsiatsiooniks vajalik ka spetsiifiliste TF-de olemasolu. Kui transkriptsiooni kontrolliv järjestus -35 on vaevu äratuntav on vajlikud transkriptsiooni aktivaatorid. Miks ei ole konsensus igalpool? vaja geeniregulliks. Aktiveeritavatel promootoritel on -35 heksameer konsensusjärjestusest TTGACA märkimisväärselt erinev konsensusjärjestusest ja sel juhul soodustab aktivaator polümeraasi seondumist promootorile. Lisaks TF-dele toimub transkriptsiooni regulatsioon ka erinevate faktorite kaudu. RNA polümeraasi subühikud on eraldivõttes inaktiivsed. Ainult ß`on võimeline DNA-ga mittespetsiifiliselt seonduma. Seetõttu seondub ka RNA polümeraasi apoensüüm DNA-ga mittespetsiifiliselt. faktori lisandumisel toimub RNA polümeraasi spetsiifiline seondumine promootoralale ligikaudu 1000 korda kõrgema efektiivsusega. Holoensüümi koosseisu kuulub ka faktor. faktor on vajalik RNA polümeraasi spetsiifiliseks
füsioloogia). Organitest moodustuvad ELUNDKONNAD e. ORGANSÜSTEEMID (füsioloogia) . Elundkond on samuti elu organiseerituse üks tase. Organismist eraldatuna ei ole ühelgi koel, organ(süsteemil) elu tunnuseid. Seetõttu peetakse rakust järgmiseks oluliseks tasemeks organismi taset. - ORGANISMI tase – organism talitlus sõltub tema elundite(kondanse) koostööst, regulatsiooniga tagatakse sisekeskkonna stabiilsus, neuraalne regulatsioon, humoraalne regulatsioon. Organismid koos omakorda moodustavad POPULATSIOONIDES taseme – ühel asustusalal elavad sama liiki organismid. - LIIGI tase – üks peamisi. Uurib, mis on ühe konkreetse liigi eripära ÖKÖSÜSTEEMI TASE – organismid + keskkond (st ümbritsev elus- ja eluta lodus – ökoloogia). Ühisel territooriumil omavahel toitumissuhetes olevad organismid – Kogu ELU KÕRGEIM TASE
ORGANEID (anatoomia, füsioloogia). Organitest moodustuvad ELUNDKONNAD e. ORGANSÜSTEEMID (füsioloogia) . Elundkond on samuti elu organiseerituse üks tase. Organismist eraldatuna ei ole ühelgi koel, organ(süsteemil) elu tunnuseid. Seetõttu peetakse rakust järgmiseks oluliseks tasemeks organismi taset. - ORGANISMI tase organism talitlus sõltub tema elundite(kondanse) koostööst, regulatsiooniga tagatakse sisekeskkonna stabiilsus, neuraalne regulatsioon, humoraalne regulatsioon. Organismid koos omakorda moodustavad POPULATSIOONIDES taseme ühel asustusalal elavad sama liiki organismid. - LIIGI tase üks peamisi. Uurib, mis on ühe konkreetse liigi eripära ÖKÖSÜSTEEMI TASE organismid + keskkond (st ümbritsev elus- ja eluta lodus ökoloogia). Ühisel territooriumil omavahel toitumissuhetes olevad organismid Kogu ELU KÕRGEIM TASE BIOSFÄÄR, hõlmab kogu Maad ümbritsevat elu sisaldavat kihti.
viiruse assambleerimisel o Geneetilise materjali segunemine võib toimuda siis, kui 2 sarnast viirust nakatavad sama rakku Rekombinantsed girpiviirused ja nendega seotud ohud: Linnugripp (H5N1): o Kandub lindudelt inimestele o Kardeti viirustevahelist rekombineerumist – uus tüüp, mille vastu puudub täielikult immuunsus o Hispaania gripp 1918 – pandeemia; viirus suutis välja lülitada RIG1 geeni, mis põhjustab immuunsüsteemi ülereageerimise H1N1 tüvi – seaviirus, „uut tüüpi“ viirus: o RNA segmendid on segunenud o Segunenud segmendid on pärit erinevatelt gripiviirustelt (kahelt seaviiruselt, inimese viiruselt ja linnuviiruselt) o Hakkas levima Mehhikos o Kandub inimeselt inimesele o Kardeti pandeemiat 3. Selgitage retroviiruse paljunemist HIV näitel. HIV-ga nakatunute ravi Retroviirused:
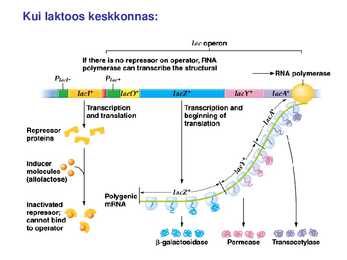
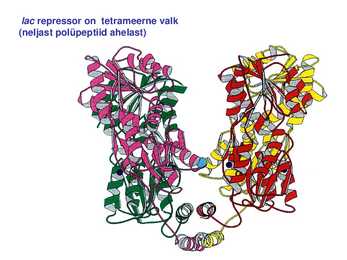
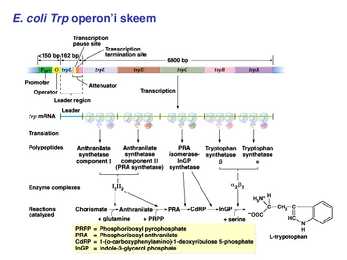
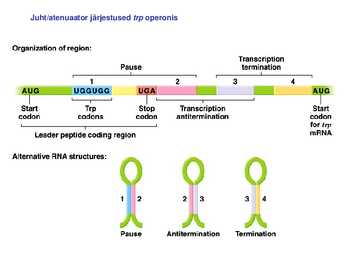
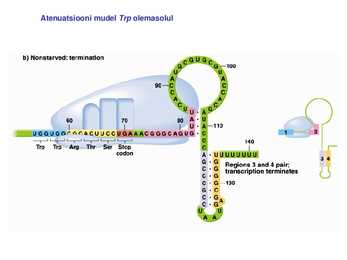
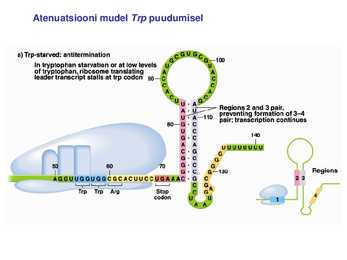
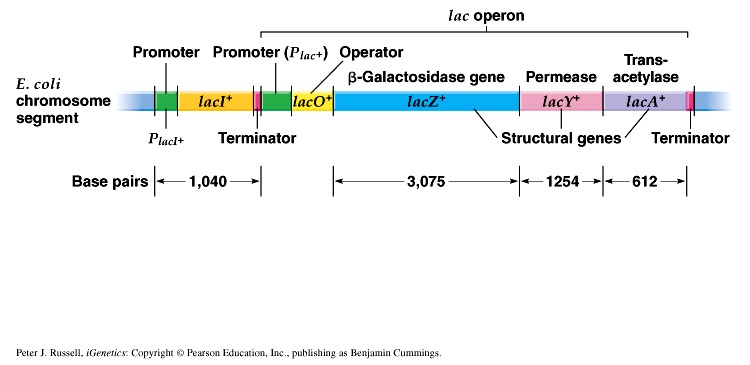
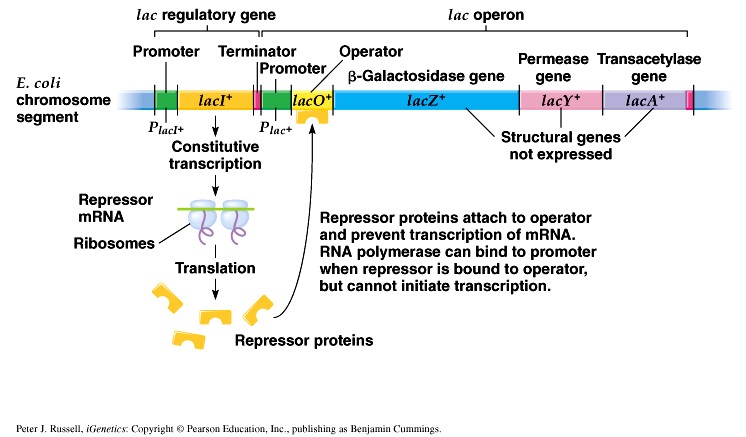
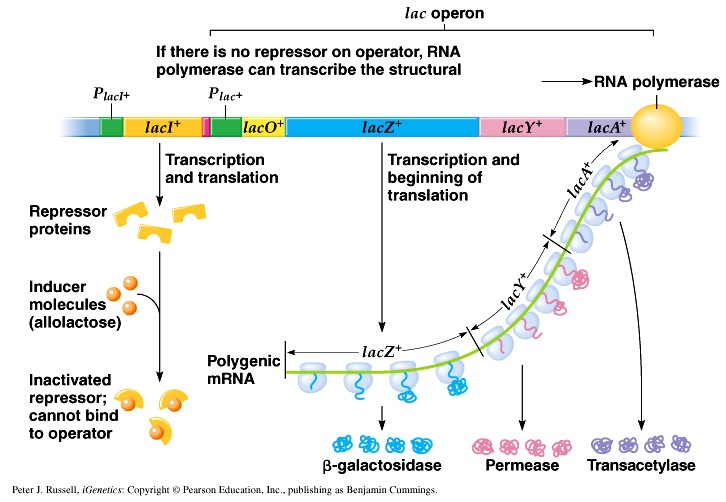
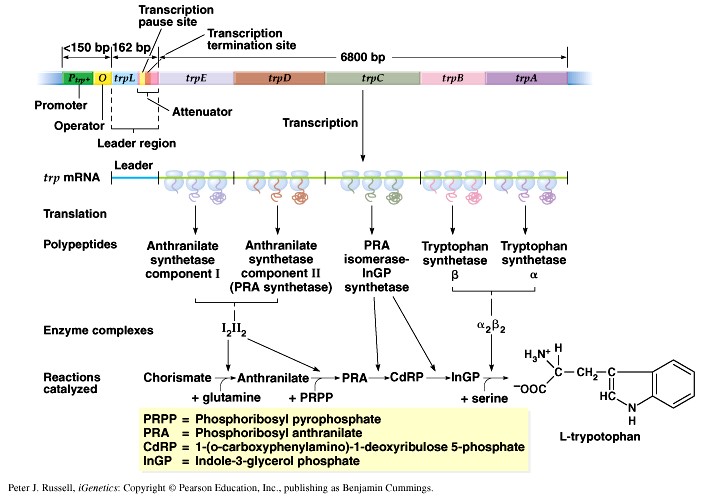
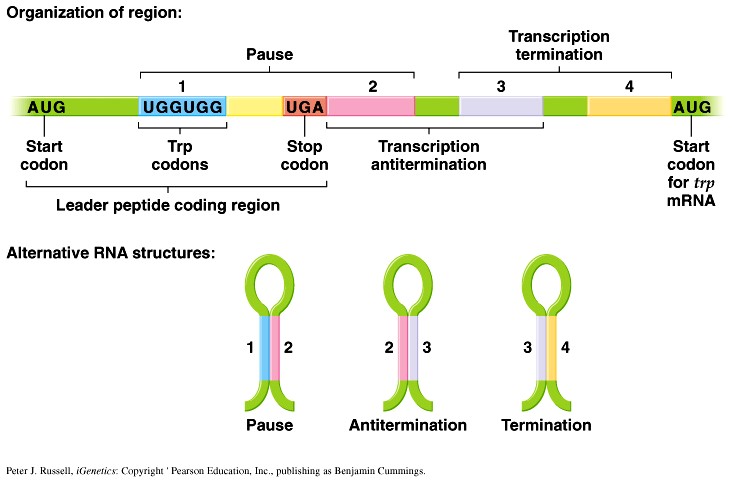
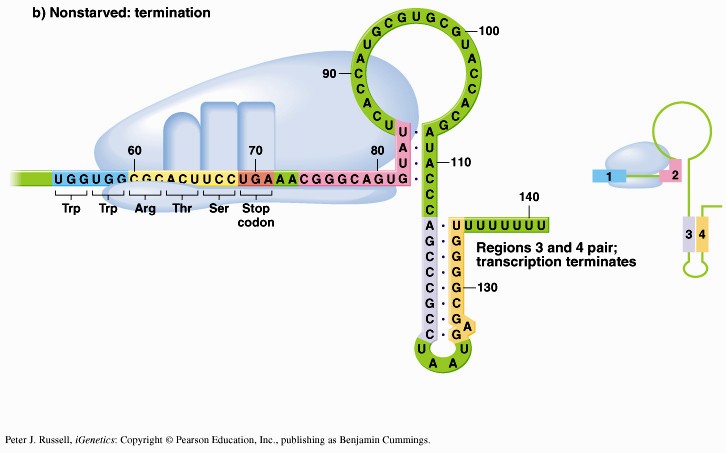
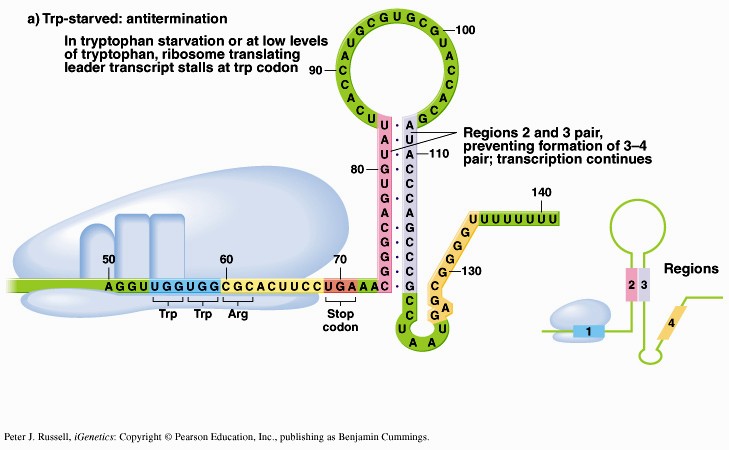
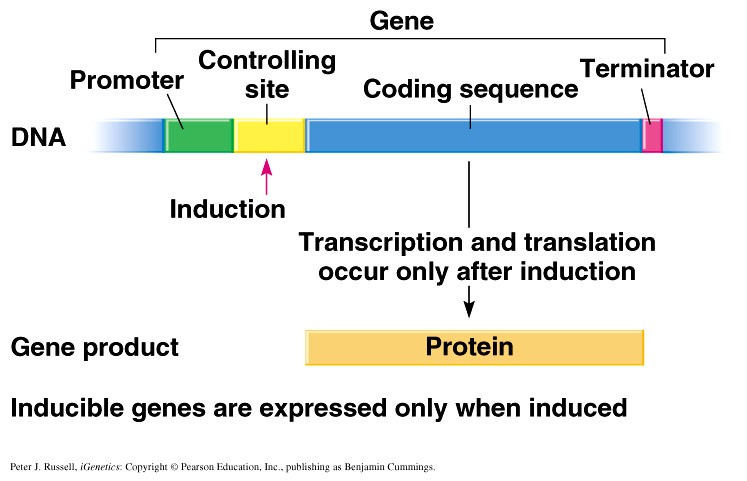
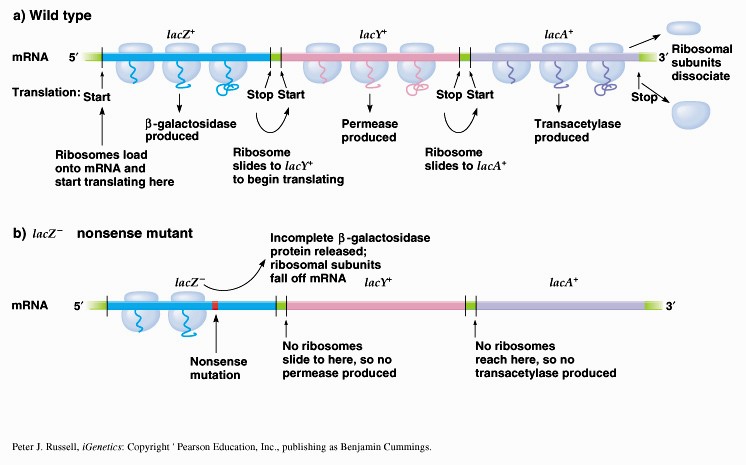
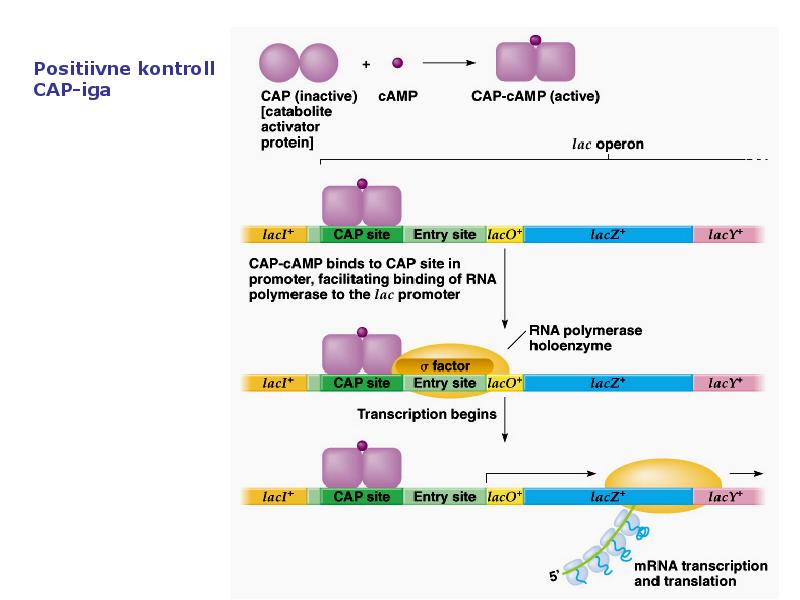
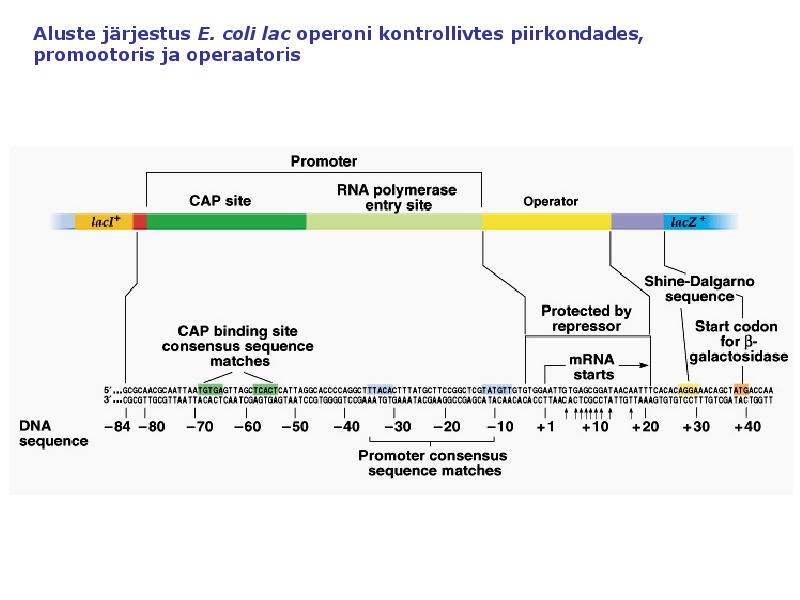
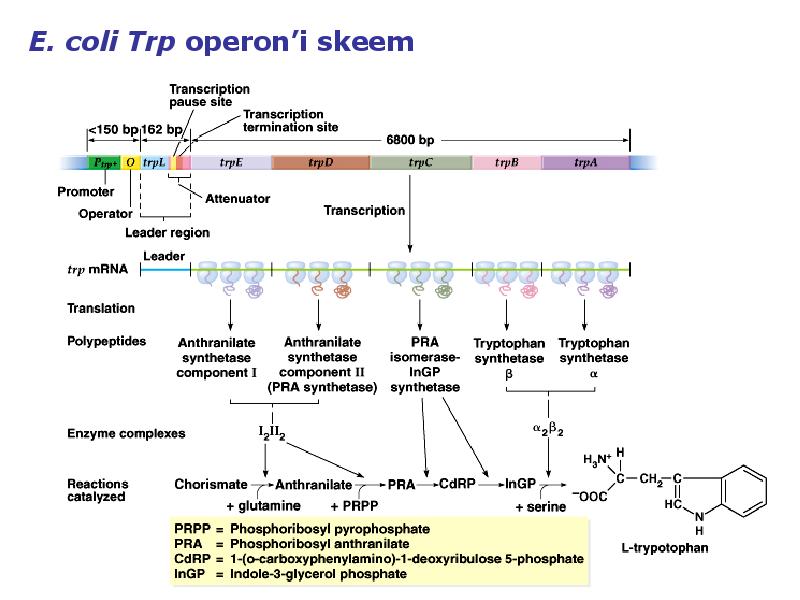
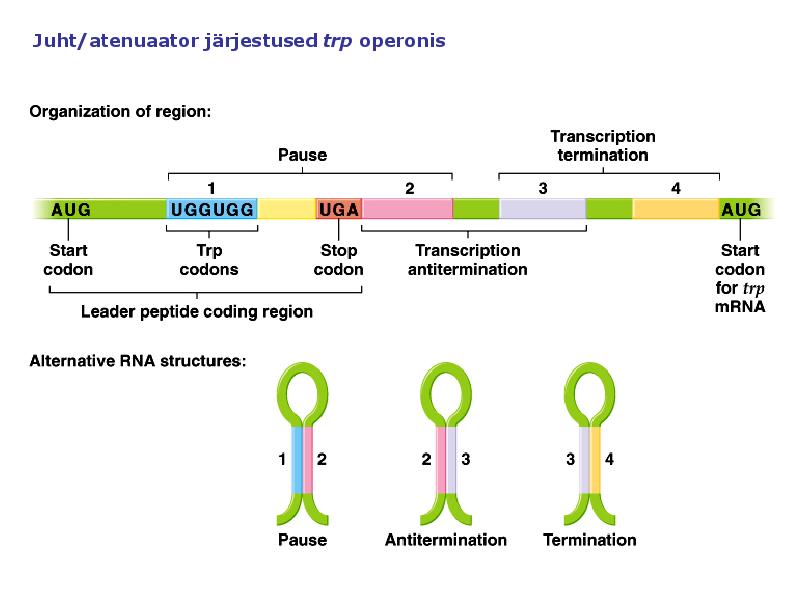
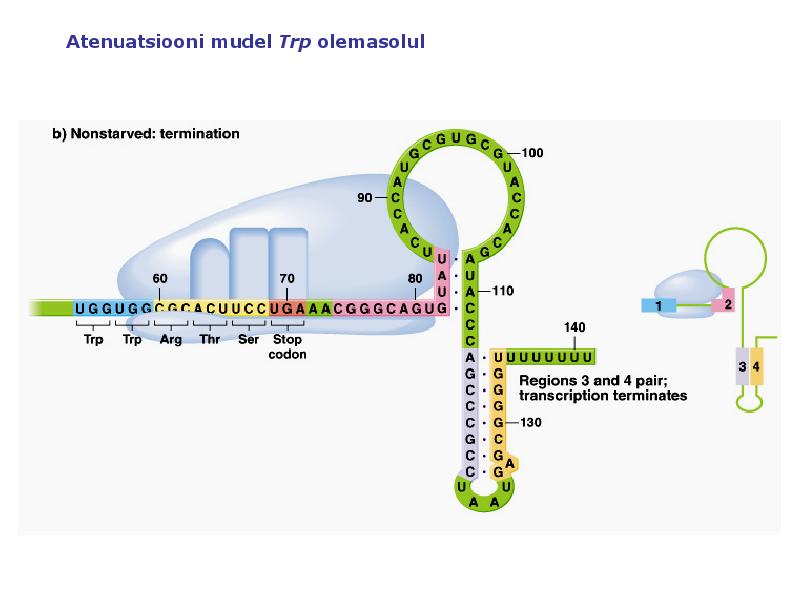
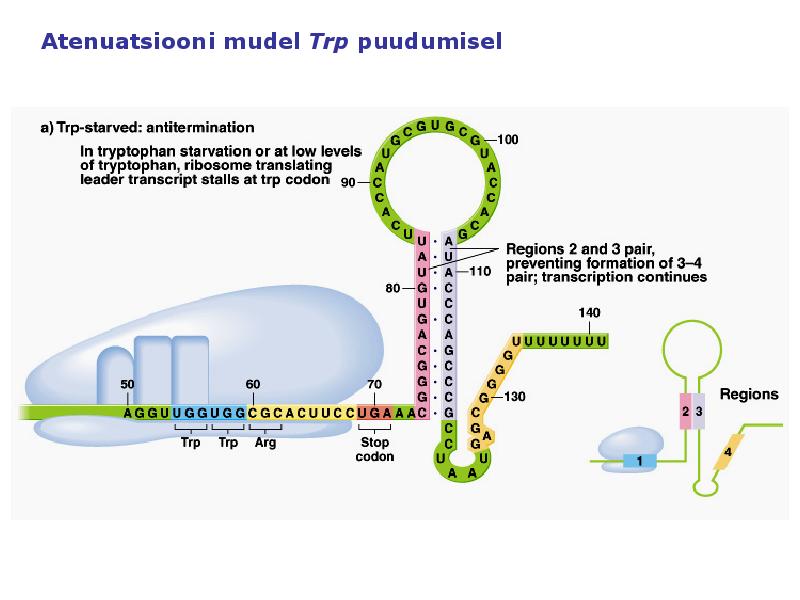
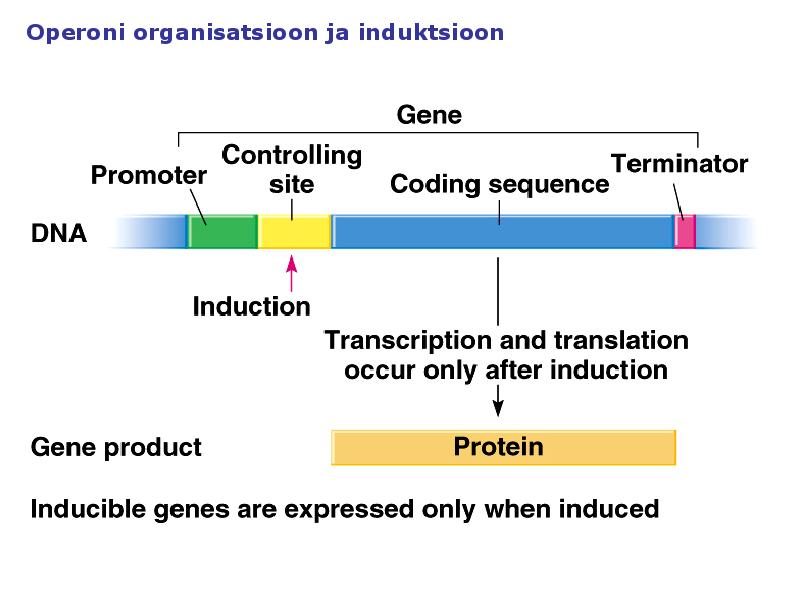
1. Sissejuhatus Metaboolne ja geneetiline regulatsioon bakterites Bakterirakkude efektiivseks kasvuks on vaja, et kõiki raku põhilisi ehitusblokke ja nendeks vajalikke makromolekule produtseeritaks õiges vahekorras. Selleks, et sünteesi lõpp-produktide kontsentratsioon rakus liiga kõrgele ei tõuseks, on rakus välja kujunenud kaks kontrollmehhanismi: 1. Ensüümiaktiivsuse tagasisidestuslik inhibitsioon (feedback inhibition) metaboolne regulatsioon 2. Ensüümi sünteesi repressioon geneetiline regulatsioon Tagasisidestusliku inhibitsiooni tulemusena inhibeeritakse rakus juba olemasoleva ensüümi aktiivsus reaktsiooni lõpp-produkti poolt. Inhibitsiooni võib esile kutsuda ka teatav metabolismiraja vaheprodukt. Geneetilise repressiooni korral inhibeerib tavaliselt lõpp-produkt metabolismiraja esimese ensüümi sünteesi vastava geeni avaldumise pärssimise kaudu. Metaboolne regulatsioon tagasisidestusliku inhibitsiooni kaudu
olekus. Substraadi kontsentratsiooni tõustes muutub tasakaal T-olekust Rüleku suunas. 7. Kirjeldage transkriptsiooni alustamist prokarüootsetes ja eukarüootsetes rakkudes. Prokarüoodid Eukarüoodid Transkptsioon algab kui RNA- Eukarüootidel on 3 tüüpi RNA-polümeraase: polümeraasi σ-faktor seondub geeni • RNA-pol. I (tRNA, rRNA, väikesed RNAd) promootoriga ja lōpeb geeni • RNA-pol. II (mRNA) terminaatorjärjes-tuse juures. • RNA-pol. III (tRNA, rRNA, väikesed RNAd) Promootorjärjestus on asümmeetriline ja määrab RNA sünteesi suuna. Prokarüootidel on RNA polümeraasi II transkriptsiooni üks RNA-polümeraas, mis sünteesib alustamiseks on vaja alati kōiki RNA tüüpe
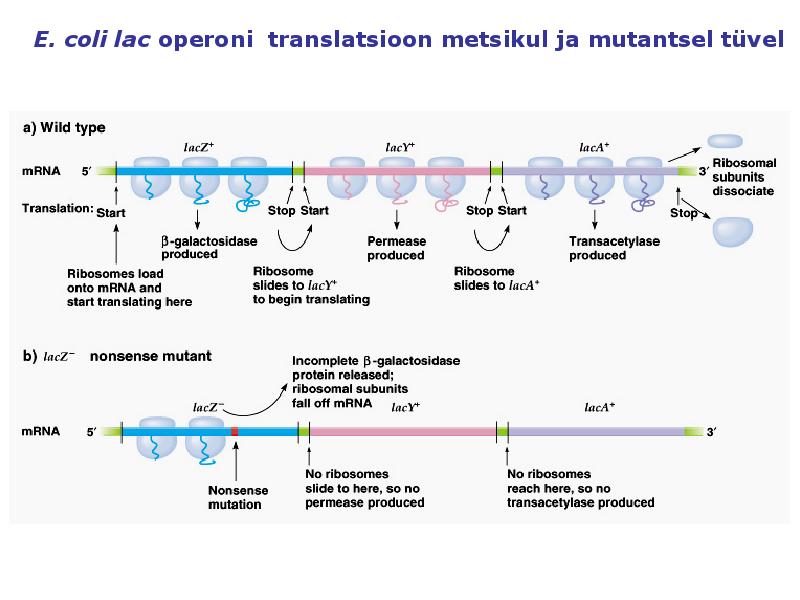
1 MOLEKULAAR- JA RAKUBIOLOOGIA | YTM0011 II KONTROLLTÖÖ KORDAMISKÜSIMUSED | MIHKEL HEINMAA TTÜ YAGB31 | 08/10/10 TRANSKRIPTSIOON JA GEENI REGULATSIOON 1. Missuguseid geneetilise infovahetuse protsesse tähistavad a) transformatsioon, 2) transkriptsioon 3) translatsioon. Transformatsioon on geneetilise info ülekandumine ühest bakterirakust teise rakust isoleeritud DNA abil. Transkriptsioon ehk RNA süntees on DNA ühe ahela (matriitsahel) alusel komplementaarse RNA ahela süntees.







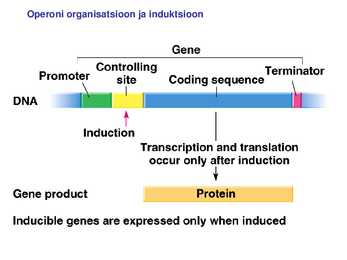



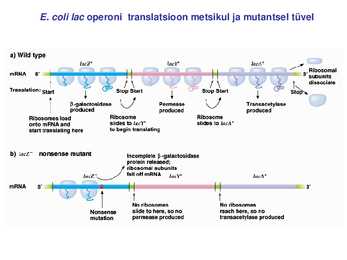




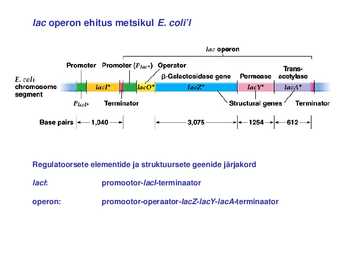
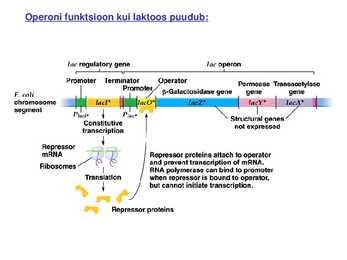

























































Kõik kommentaarid