ÖKOLOOGIA (LOOM .01.105) KORDAMISKÜSIMUSED, kevad 2011. a. 1. Ökoloogia aine, alajaotused; Teadus, mis käsitleb organismide ja keskkonna suhet. Kõikide sidemed kõikidega. Jaguneb: a) Ökofüsioloogia e molekulaarne ökoloogia b) Autökoloogia (isendi/organismi tasandil) c) Demökoloogia (populatsiooni tasandil) d) Sünökoloogia (eluskoosluse, populatsioonide tasandil) e) Süsteemökoloogia (ökosüsteemi tasandil, elus kooslus + eluta keskkond) f) Biosfäroloogia e biosfääri ökoloogia (globaalne ökosüsteem) 2. Ökoloogia põhimõisted isend (genet, kloon, ramet), populatsioon, kooslus, ökosüsteem, bioom; Isend- kindla genotüübiga organism
Ökoloogia on teaduslik õpetus organismide ja nende keskkonna vahelisest seostest ja mõjudest. Keskkonna alla kuulub nii biootiline kui ka abiootiline keskkond. Ökoloogiat võib defineeida ka kui organismide "kodu elu". Ökoloogia alajaotused: * molekulaarne ökoloogia (molekuli, organi ja isenfi tasandil) ; (ökofüsioloogia- uurib organismide kohanemisreaktsioone) * autökoloogia (isendi tasandil) * pop.ökoloogia e demökoloogia * kooslusökoloogia e sünökoloogia *geograafiline ökoloogia * biosfääriline ökoloogia 2. Ökoloogia põhimõisted isend (genet, kloon, ramet), populatsioon, kooslus, ökosüsteem, bioom (konspekt); Isend: unitaarne organism. Selline organism, kes ei moodusta mooduleid, mis oleksid kas suhteliselt või täiesti iseseisvad. Populatsioon: ühise genofondiga isendite kogum. Kooslus: koos eksisteerivad populatsioonid Ökosüsteem: hõlmab endas elukooslust ja selle abiootilist keskkonda Bioom: saransed ökosüsteemid üle maailma 3
ÖKOLOOGIA (LOOM .01.105) KORDAMISKÜSIMUSED, kevad 2013. a. 1. Ökoloogia aine, alajaotused; Ökoloogia on teadus organismide ja keskkonna vahelistest suhetest. Ökoloogia alajaotused on : · Ökofüsioloogia (organell, rakk, organ) · Autökoloogia (isend) organism ja keskkonna suhe isendi tasemel · Demökoloogia (populatsioon) · Sünökoloogia (kooslus) · Süsteemökoloogia (ökosüsteem, biosfäär) 2. Ökoloogia põhimõisted isend (genet, kloon, ramet), populatsioon, kooslus, ökosüsteem, bioom; · isend organism, mis ei moodusta iseseisvaid mooduleid o kloon ehk genet geneetiliselt identne moodulite kogum o ramet taime puhul võsu, risoomiga ühendatud, iseseisva juuresüsteemiga moodul · populatsioon ühise genofondiga isendite kogum · kooslus koos eksisteerivad populatsioonid · ökosüsteem elukooslus ja selle abiootiline kes
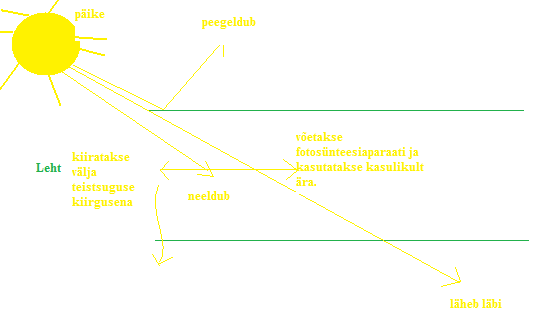
Ökosüsteem Esimesena kasutas seda sõna Tansley 1935. eluskooslus ja selle abiootiline keskkond. Nt kuumaveeallikas, lillepeenar vms. Bioom sarnaste ökosüsteemide kogum üle maailma. Nt troopiline vihmamets, tundra. Biosfäär globaalne ökosüsteem. Kõik teadaolevad ökosüsteemid on avatud süsteemid, st et nad vahetavad naabersüsteemidega nii ainet kui energiat, ja ei ole seetõttu eraldiseisvad üksused. Ökoloogia valdkonnad: · Molekulaarne ökoloogia meetodile viitav. · Ökofüsioloogia uurib füsioloogiliste protsesside kohanemist vastavalt keskkonnale. · Antökoloogia isendi suhted keskkonnaga. · Populatsiooniökoloogia, demökoloogia(demograafiline) · Sünökoloogia(koos, kaasa, kooslemise), kooslusökoloogia · Süsteemökoloogia · Geograafiline ökoloogia · Biosfäroloogia Ökoloogiline faktor(tegur) aine, energia või informatsiooni voog keskkonnast, mis
1. Aine, alajaotused (allpool) , areng. Ökoloogia - teadus, mis uurib elusa ja eluta looduse omavahelist suhet, ei keskendu ühele objektile, vaatleb tervikut. E. Haeckel 1869 ökoloogia on teadus organismide ja kk suhetest. E. Odum teadus looduse struktuurist ja funktsoonist. 2. Ökoloogia põhimõisted. Ökoloogia valdkonnad: 1) Organelli tase 2) Raku tase (ainurakse puhul isend) 3) Koe tase 4) Organi tase 5) Isendi tase autökoloogia, uurib abiootilisi kk faktoreid. 6) Populatsiooni tase demökoloogia e. populatsiooni ökoloogia. 7) Koosluse tase kooslusökoloogia e. sünökoloogia, uurib mitmeliigilisi pop. süsteeme.
1. Aine, alajaotused (allpool) , areng. Ökoloogia - teadus, mis uurib elusa ja eluta looduse omavahelist suhet, ei keskejdu ühele objektile, vaatleb tervikut. E. Haeckel 1869 - ökoloogia on teadus organismide ja kk suhetest. E. Odum - teadus looduse struktuurist ja funktsoonist. 2. Ökoloogia pôhimôisted. Ökoloogia valdkonnad: 1) Organelli tase- uurib olulisi eluavaldusi madalamal str tasemel 2) Raku tase (ainurakse puhul isend) 3) Koe tase 4) Organi tase- autökoloogia, org. Ja keskk. Suhete uurimine isendi tasandil 5) Isendi tase - autökoloogia, uurib abiootilisi kk faktoreid. 6) Populatsiooni tase - demökoloogia e. populatsiooni ökoloogia.
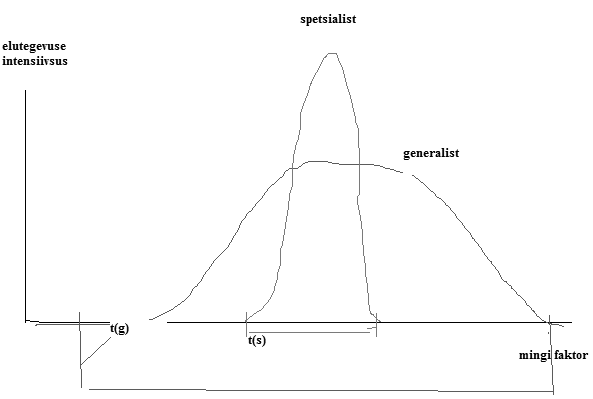
72. Kuidas avaldub produktsiooniefektiivsus? A. PE = In/Pa-1 B. PE = An/In C. PE = Pn/An-1 D. PE = Pn/An 73. Kas kiskja-saaklooma süsteemis: A. Kiskja populatsioonitiheduse madalseis vastab saaklooma populatsiooni suurimale tihedusele B. Kiskja ja saaklooma populatsioonitihedus võnguvad samas rütmis veerandperioodilise faasnihkega C. Saaklooma populatsioonitihedus on kiskja omast sõltumatu kuid fluktueerib samas rütmis 74. Termini ökoloogia võttis kasutusele: A. K. Zobel B. V. Masing C. Haeckel D. Hutchinson E. V. Gause 75. Seen võib olla: A. Loom B. Imetaja C. Nematood D. Parasiit E. Primaarne produtsent 76. Produktsiooni efektiivsus on suurem: A. Kõigusoojastel selgroogsetel? B. Püsisoojastel selgroogsetel? C. Nendel selgroogsetel, kelle energiakadu hingamisele on null? 77. Millise elemendi ringe puhul on atmosfäär väheoluliseks fondiks? A. N B. P C. C D. O 78
ÖKOLOOGIA KORDAMISKÜSIMUSTE VASTUSED EKSAMIKS 1. Ökoloogiateaduse uurimisobjektid - teadus interaktsioonid, mis määravad elusorganismide leviku ja arvukuse. 2. Ökoloogiliste tasemete hierarhia (alates kõrgemast tasemest) - Süsteemide ökoloogia (ecosystem), sünökoloogia (community), demökoloogia (population), autökoloogia (organism), ökofüsioloogia (organ system, organ, tissue, cell). 3. Populatsiooni mõiste - rühm ühe liigi isendeid, kes elavad koos samal ajal samas paigas.gh 4. Ökoloogilise teguri mõiste ja liigitus Selline aine-, energia-, ja infovoog keskkonnas, mis avaldab selles keskkonnas elavatele organismidele mõju.































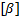
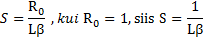
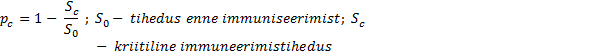
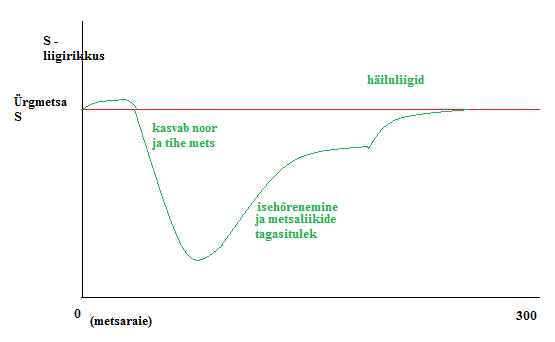
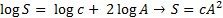
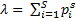

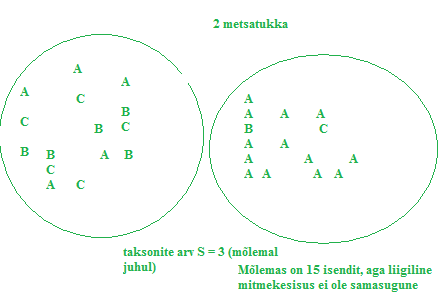
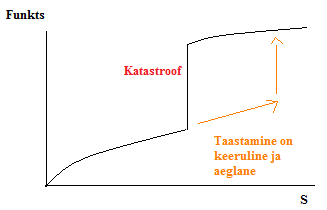
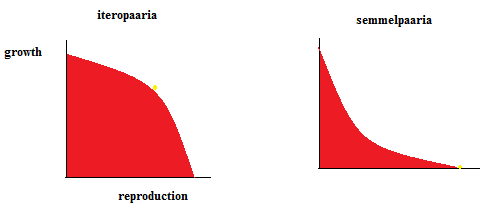
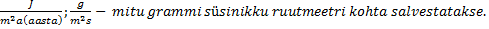
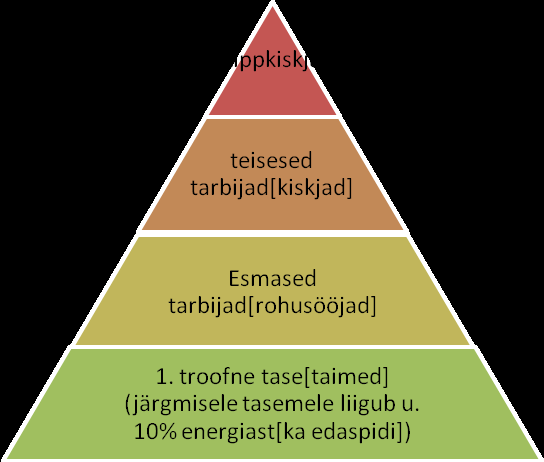
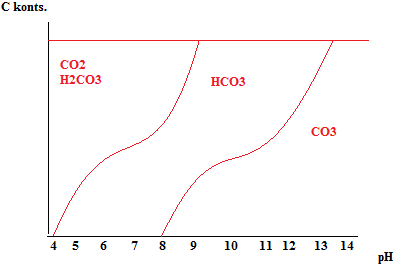


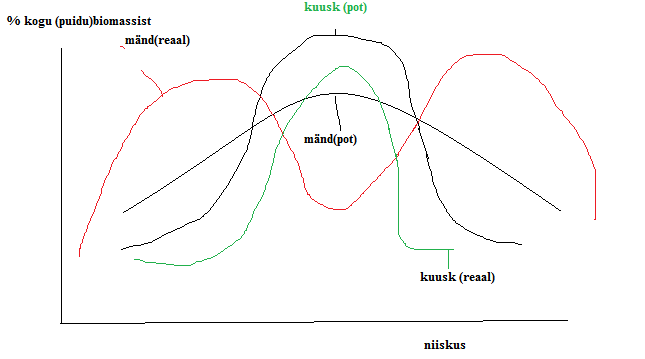
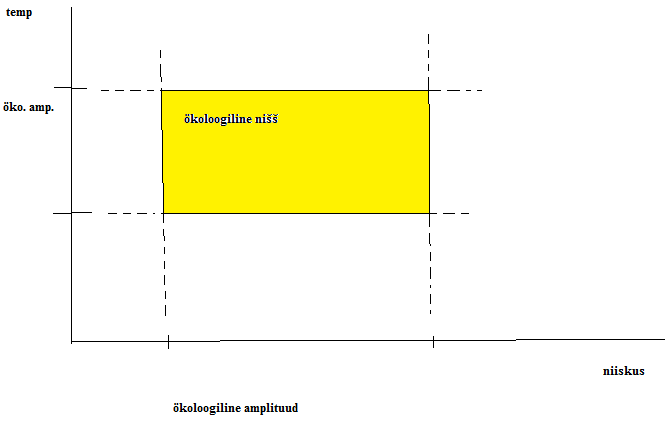
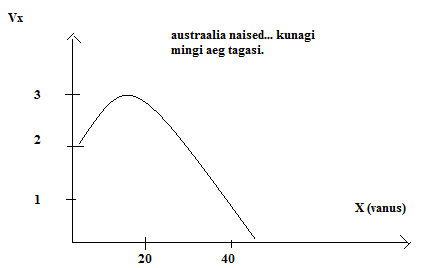
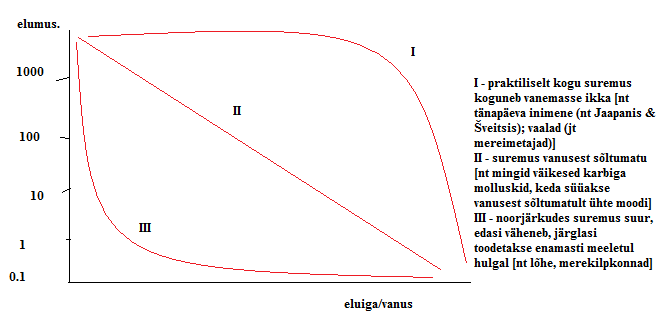
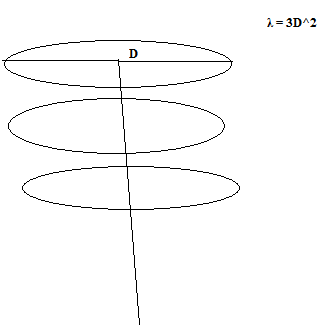
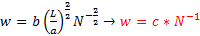
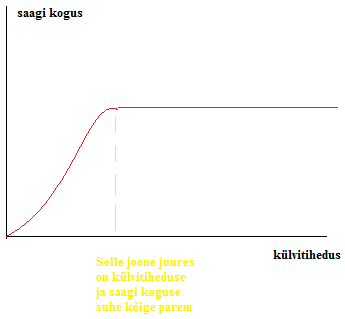
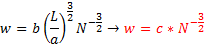
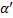
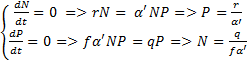
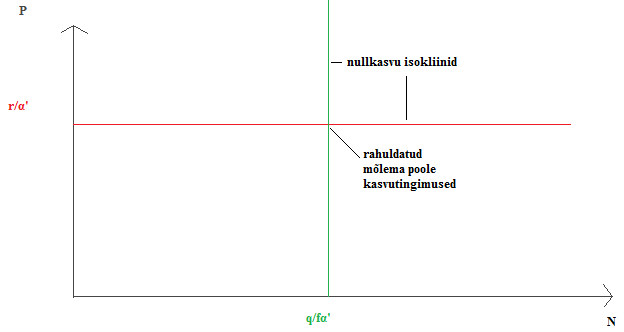
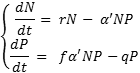
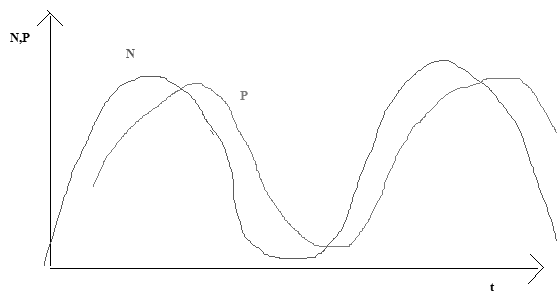




Kõik kommentaarid