Muutus geeni sees nukleotiidid (DNA molekulis on muutunud vaid mõned nukleotiidid) A, T, C, G Tekivad peamiselt replikatsiooni käigus sagedusega 1/miljoni nukleotiidi kohta. See toob kaasa vastava valgu aminohappelise koostise muutuse, mis omakorda väljendub fenotüübis. Tulemus võib olla väga suur või ka muutumatu. Kui üks või kaks nukleotiidi on kaduma läinud/juurde tulnud, muutub kogu aminohappeline järjestus valeks. Näiteid geenmutatsioonidest: · Varane raukumine- DNA helikaasi määravas geenis on viga. · Juuste täielik väljalangemine- A G · XY kromosoome kandev loode ei arene poisiks, kui testosterooni määravad geenis on viga. · Sirprakuline aneemia (ei haigestu malaariasse), sage Aafrika rahvastel. · Tsüstiline fibroos (kopsud limased, hingamisraskused) Eurooplastel levinuim geneetiline haigus (ei haigestu koolerasse, salmonelloosi) Leidub ka kasulikke geenmutatsioone:
~245 bp E. coli Initsiatsioon: Üheahelalisi DNA lõike nimetatakse matriitsideks Güraas on ensüüm (topoisomeraas), mis vabastab superspiralisatsiooni Initseeriv valk ja DNA helikaas seonduvad DNAga replikatsiooni kahvlis ja toimub lahtikeerdumine. Energia tuleb ATPst. ATP hüdrolüüs põhjustab konformatsioonilisi muutusi helikaasis DNA primaas seob järgnevalt helikaasi, nii et moodustub primosoom Primaas sünteesib väikese, 1012 RNA, millele DNA polümeraas hakkab sünteesima juurde nukleotiide Polümeraas III lisab nukleotiide 5' 3' suunas mõlemal ahelal, alustades RNA praimerist RNA praimer hiljem eemaldatakse ja asendatakse DNA polümeraasiga. Vahemik siilitakse ligaasi poolt. Üheahelaine DNA stabiliseeritakse kogu protsessi vältel spetsiaalse valguga Replikatsiooni mudel E. coli
kasutades ATP hüdrolüüsi energiat. * DNA replikatsiooni algatamine bakterite rakus Valgud, mis algatavad bakterites DNA replikatsiooni 2 replikatsiooni kahvlit lähtuvad originist ning liiguvad vastassuundades. Initsiaator proteiinid seostuvad spetsiifilisele DNA järjestusele replikatsiooni alguspunktis (replication origin) ja destabiliseerib kaksikheeliksi, moodustades struktuuri, mille puhul DNA on tugevalt pakitud ümber valgu. Kaks helikaasi seostuvad helicase-loading (DNA C valk) valkudega, mis inhibeerivad helikaasi, kuni see on täielikult replikatsiooni alguspunkti jõudnud. SSB valkude kaasabil helikaasid avavad DNA. Võimaldades seeläbi primaasil sisenda ja sünteesida initsiaalne praimer. Initsiaator proteiinid eemalduvad kui vasakpoolne replikatsiooni kahvel liigub neist läbi. * DNA replikatsiooni algatamine eukarüootide rakus See mehhanism kindlustab, et igat replikatsiooni alguspunkti aktiveeritakse ainult
Võib kaasa tuua vastava valgu aminohappelise koostise muutuse, mis omakorda väljendub fenotüübis. Tulemuses võib olla märkamatu või väga suur viga. Duchenne’i lihasdüstroofia- haigus, mille puhul lihased jäävad nõrgaks. Sünnitakse normaalselt, avaldub teismeeas. Hemofiilia- kaasasündinud verehüübimatus Huntingtoni tõbi- värisemine ja koordinatsioonihäired Mittetäielik luuteke- haigus, mis avaldub luumurdude ja -moondustega Progeeria- tekib hästi kiire vananemine. Helikaasi määravas geenis on viga. 4. Kromosoommutatsioonid. Millest tulenevad kromosoommutatsioonid? Näiteid haigustest. Vastus: Kromosoomi struktuuri muudatused, mida on võimalik mikroskoobiga näha. Inversioon- kromosoomi osa pöördub ümber Kassikisa sündroom- 5. kromosoomist osa puudu Martin Belli sündroom- deletsioon X kromosoomist Madal IQ, hüperaktiivne, keskendumishäired 5. Genoommutatsioonid? Millest tulenevad genoommutatsioonid? Näiteid haigustest.
mikroobe *Translatsioon - valgusüntees, mis toimub tsütoplasmas asuvates ribosoomides. *Transkriptsiooni käigus kopeeritakse DNAs paikneb informatsioon RNA-ahelasse. *Kui rakkudes läheb vaja mõnda valku, siis aktiveerub DNA-ahelas just selle valgu valmistamisjuhist sisaldav geen. *RNA-polümeraasi seondumist promootoriga ja sünteesi algust reguleerib suur hulk valke. *RNA-sünteesil pole vesiniksidemete katkestamisel helikaasi abi tarvis. *Sünteesitav RNA on komplementaarne ühega kahest DNA-ahelast. *Replikatsiooni eesmärk: päriliku info jõudmine kõikidesse rakkudesse ja selle edasiandmine järglastele. *Replikatsiooni sisu: DNA-ahela kopeerimine enne raku jagunemist. *Transkriptsiooni eesmärk: päriliku info avaldumine. *Transkriptsiooni sisu: RNA-ahela sünteesimine DNA-ahelas sisalduva info alusel. *Valgud on biomolekulid, mis koosnevad aminohapetest.
saaks lahku minna. DNA-polümeraas- ensüüm, mis sünteesib, kahe vana DNA-ahela järgi uued DNA-ahelad, lähtudes taas komplementaarsusprintsiibist. Tulemuseks on 2 uut DNA-kaksikheeliksit, millel mõlemal on 1 uus ja 1 vana ahel ja mõlemad nad sisaldavad sama geneetilist teavet. Promootor- geeni alguses olev DNA-järjestus. Terminaator- geeni lõpus olev DNa-järjestus. RNA-polümeraas- lõhub samuti vesiniksidemeid, kuid ta ei vaja selleks helikaasi abi. Kõik organismile vajalikud valgud moodustatakse rakkude sees aminohapetest. Inimese kehas 20 erinevat aminohapet, neist 8 on asendamatud ( peame ise toiduga saama, keha neid ei sünteesi). Peptiidside- kahe aminohappe vahele tekkinud ühendust. Valgud biomolekulid, koosnevad paljudest aminohapetest. Ribosoomid rakuorganellid, kus sünteesitakse valke. Translatsioon- valgusüntees, toimub tsütoplasmas olevatest ribosoomides, vajaliku info viib tsütoplasmasse mRNA
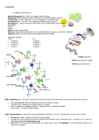
1. Helikaas katkestab vesiniksidemed ja DNA kaksikheelik avaneb. Süntees algab lahtikeerdunud DNA-ahela kohtadest 2. DNA-polümeraas- sünteesib mõlema vana ahela järgi uued DNA-ahelad. Tulemuseks on kaks uut DNA ahelat DNA transkriptsioon- DNA-ahela järgi sünteesitakse RNA-ahel. Toimub tsütoplasmas asuvas ribosoomides. 1. Promootor (geeni alguses oleva DNA-järjestusega) 2. Polümeraas peab katkestama vesiniksidemed DNA-ahelate vahel. (helikaasi vaja ei ole RNA puhul). Ensüümi möödumisel vesiniksidemed taastuvad. 3. RNA-polümeraas liigub piki DNAd edasi, kuni jõuab geeni lõppu. Terminaator on nukleotiidide järjestus, kus lõpeb RNA süntees. Geneetika 1.3 Pärilik info avaldub valkude sünteesi kaudu Valgud koosnevad aminohapetest Peptiidside- keemiline side, mis seob aminohapped (C-N) Translatsioon- valkude sünteesimine vastavalt mRNAs olevatele juhistele. Sellele eelneb mRNA süntees ehk
poole oleksid ja et vesiniksidemed tekkida saaksid.Antiparalleelsed s.t., üks ahel kulgeb suunas 5' 3' ja teine suunas 3' 5' , 10. Millises suunas toimub uute nukleiinhappe ahelate süntees? 5' -> 3' 11. Mis on replikatsioon, milliseid ensümaatilisi aktiivsusi on selle juures vaja? DNA molekuli "paljundamine" enne rakujagunemist. Ühest DNA molekulist saadakse 2, kusjuures mõlemas molekulis on üks ahel pärit vanast molekulist. Replikatsiooniks on vaja helikaasi, et lõhkuda ahelaid koos hoidvad vesiniksidemed. Polümeraasi, et lisada uusi nukleotiide. 12. Milline ensüüm teostab replikatsiooni? DNA-polümeraas 13. Kust saavad rakud energiat uute fosfodiestersidemete sünteesimiseks? Nukleotiidtrifosfaatidest, katkeb makroergiline side 14. Mida tähendab, et uute DNA ahelate süntees rakkudes on semikonservatiivne? Uues DNA kaksikheeliksis on üks ahel pärit vanast DNA molekulist ja teine on uus, sünteesitud ahel. 15
mis sisaldab enamuse GTF'dest. Arvatakse, et selline kompleks istub DNA'le ühe sündmusena. 9. Missugused TAFid on TBPga vahetus interaktsioonis? (TBP on eukarüootse transkriptsiooni faktori TFIID alaühik: TBP + 16 TAF'i = TFIID). Nime järgi võiks oletada, et kõik on. GTF'dest aga TFIIA. 10. Missugune TFII valk hüdrolüüsib ATPd ja mis protsessiga on tegu? TFIIH kahel alaühikul on helikaasi ja ATPaasi aktiivsus. Sellega aitab ta luua transkriptsioonimulli. TFIIH helikaasne aktiivsus saab oma energia ATP hüdrolüüsilt. Selle energia abil keerataksegi START-saidi piirkonnas DNA dupleksi lahti. (TFIIH ühe teise sub-kompleksi roll on fosforüleerida RNA polümeraasi suurima subühiku C-terminaalset domääni (CTD), mis on vajalik polümeraasi vabanemiseks PIC-ilt (pre-initsiatsiooni kompleks) ning transkriptsiooni üleminekul elongatsioonifaasi). 3 11
Viivisahelal võib jääda laadur kogu sünteesi ajaks kompleksiga kokku Eksonukleaas lõikab RNA praimeri välja ja DNA ligaas seob DNA ahela kokku 18. DNA replikatsiooni algatamine bakterite rakus Üks alguspunkt, helikaas liigub piki mahajäävat ahelat 1) Initsiaatorvalgud seostuvad spetsiifilisele DNA järjestusele replikatsiooni alguspunktis, ja destabiliseerib kaksikheeliksi, moodustades struktuuri, mille puhul DNA on tugevalt ümber valgu pakitud 2) Kaks helikaasi seostuvad helicase loading (DNA C valk) valkudega, mis inhibeerivad helikaasi, kuni too on replikatsiooni alguspunkti jõudnud. 3) SSB valkude kaasabil avavad helikaasid DNA võimaldades seeläbi praimaasil siseneda ja praimer sünteesida 4) Initsiaatorproteiinid eemalduvad, kui vasakpoolne replikatsiooni kahvel neist läbi liigub 19. DNA replikatsiooni algatamine eukarüootide rakus Mitu alguspunkti, Helikaas liigub piki liiderahelat
DNA ahelad. Replikatsioon lõppeb, kui mõlemalt DNA-ahelalt on sünteesitud uus DNA molekul. Selleks, et rakk saaks jaguneda, peab ta kõigepealt enda DNAd replitseerima. Seda protsessi alustatakse kindlatelt DNA lõikudelt, mida nimetatakse originideks. Originid sisaldavad DNA järjestusi, mille tunnevad ära replikatsiooni algatavad valgud. Need valgud seondavad omakorda erinevaid valke (nt helikaasi), et eraldada kahte DNA ahelat ning moodustada replikatsioonikahvleid. Initsiaatorvalkude algatusel keeratakse DNA ahelad lahti ning moodustub n-ö replikatsiooni-mull (DNAd sünteesitakse bidirektsionaalselt ehk mõlemas suunas). Originid on tavaliselt A-T rikkad (sisaldavad palju adeniini-tümiini aluspaare) ja aitavad sellega lahtikeerdumisele kaasa, sest A-T aluspaaridel on kaks vesiniksidet (mitte kolm, nagu C-G paaridel). Seega on A-T sidemeid lihtsam
nukleotiid ning replikatsioon saab jätkuda. Replikatsiooni protsess Originid Selleks, et rakk saaks jaguneda, peab ta kõigepealt enda DNAd replitseerima.[4] Seda protsessi alustatakse kindlatelt DNA lõikudelt, mida nimetatakse originideks. Originid sisaldavad DNA järjestusi, mille tunnevad ära replikatsiooni algatavad valgud (näiteks dnaA soolekepikesel ning ORC (Origin Recognition Complex) pärmis). Need valgud seondavad omakorda erinevaid valke (nt helikaasi), et eraldada kahte DNA ahelat ning moodustada replikatsioonikahvleid. Initsiaatorvalkude algatusel keeratakse DNA ahelad lahti ning moodustub n-ö replikatsiooni-mull (DNAd sünteesitakse bidirektsionaalselt ehk mõlemas suunas). Originid on tavaliselt A-T rikkad (sisaldavad palju adeniini- tümiini aluspaare) ja aitavad sellega lahtikeerdumisele kaasa, sest A-T aluspaaridel on kaks vesiniksidet(mitte kolm, nagu C-G paaridel). Seega on A-T sidemeid lihtsam lõhkuda, sest väikesema
pikendama moodustades lühikese 5'RNA-3'DNA tütarahela. Seejärel võtab polümeraas üle ja jätkab ahela pikendamist. Pol on seotud templeitahelaga seotuse tõttu Rfc-valguga (replication factor C), mis pöördudes muutub PCNA-ks (proliferating cell nuclear antigen trimeerne valk, mis ümbritseb tütarDNA-d). Tavaliselt on mehhanism kahesuunaline replikatsioonikahvlid liiguvad kahes suunas. Regulatsioon toimub kuuest MCM valgust koosneva helikaasi aktiivsuse kontrollimisega. Ensüümid: · Helikaas keerab DNA biheeliksi lahti · DNA güraas e. topoisomeraas kompenseerib lahtikeerdumist (aitab struktuuri hoida) · DNA-polümeraasid vastutavad polünukleotiidahelate sünteesi eest 5'->3' suunas o - 4 subühikut, polümeraasi tootlikus, pole 3' eksonukleaasi aktiivsust o - vigade parandus o - replitseeriv ensüüm mitokondrites o - peamine polümeraas
olenevalt raku tüübist ja diferentseerumisolekust, ning paiknevad üle tuuma. Transkriptsioonivabrikute diameeter on 45-100 nm. Transkriptsiooniga hõivatud RNA polümeraas II fraktsioon vabrikutes on seotud mingi alamstruktuuriga. Ehk vabrikud on immobiliseeritud mingi struktuuri külge. Vabrikud on valgurikkad – sisaldavad aktiivset või inaktiivset RNA polümeraasi, transkriptsioonifaktoreid, ribonukleoproteiine, ko- aktivaatoreid, RNA helikaasi ja splaissingu ja protsessingu ensüüme. Vabrik sisaldab ainult ühte tüüpi polümeraase. Ühes vabrikus võidakse transkribeerida samadelt või erinevatelt kromosoomidelt pärit geene. Teke – on vastuolulisi arvamusi selle kohta kas transkriptsiooni vabrikud pannakse kokku de novo vastusena transkriptsiooni nõudlusele või kas nad on stabiilsed struktuurid millede number raku tuumas püsib konstantsena.
lämmastikalauste ekspositsioon. Replikatsioon kahes suunas. Initsiatsioon: Üheahelalisi DNA lõike nimetatakse matriitsideks. Güraas on ensüüm (topoisomeraas), mis vabastab superspiralisatsiooni. Initseeriv valk ja DNA helikaas seonduvad DNAga replikatsiooni kahvlis ja toimub lahtikeerdumine. Energia tuleb ATPst. ATP hüdrolüüs põhjustab konformatsioonilisi muutusi helikaasis. DNA primaas seob järgnevalt helikaasi, nii et moodustub primosoom. Primaas sünteesib väikese, 10-12 RNA, millele DNA polümeraas hakkab sünteesima juurde nukleotiide. Polümeraas III lisab nukleotiide 5' 3' suunas mõlemal ahelal, alustades RNA praimerist. RNA praimer hiljem eemaldatakse ja asendatakse DNA polümeraasiga. Vahemik säilitakse ligaasi poolt. Üheahelaine DNA stabiliseeritakse kogu protsessi vältel spetsiaalse valguga. Kui replikatsioonikahvel kogu aeg avaneb, siis ühe DNA ahela süntees on pidev ehk toimub
UTR (untranslated regions) mRNA 5´ ja 3´otsas. REP Repetitive Extragenic palindromic sequences. Polü(A) järjestused mRNA 3´otsas - eukarüootides on polü A et stabiliseerida. Bakteritel vatsupidine funktsioon. 3´otsas polü(A)-järjestust sisaldavad RNA molekulid on PNPaasi ja RNaas II poolt paremini atakeeritavad. polü(A)-järjestus on nende eksonukleaaside kinnitumis-kohaks muidu vahetult 3´otsas sekundaarstruktuuri sisaldavatele mRNA-dele. Vajatakse helikaasi abi ja ribonukleaas kinnitub kehvemini, kui on sek struktuur, selliseid lagunatakse muidu aeglasemalt poly a annab toetsupinna. Polü(A) järjestusi sünteesivad polü(A) polümeraasid PAP I ja PAP II mRNA polüadenüleerimine on bakterirakus pidevalt toimuv protsess, mis aitab kaasa mRNA molekulide täielikule degradeerimisele. 9. Globaalne geeniregulatsioon bakterites ja selle uurimist võimaldavad meetodid.
Võimalik, et GATC saite aitab hemimetüleerituna hoida SeqA. Nimelt on näidatud, et SeqA-defektsetes mutantides toimub GATC saitide metüleerimine oriC piirkonnas varem kui metsiktüüpi bakterites. Replikatsiooni terminatsioon ja tütarmolekulide dekatenatsioon terC ja terB blokeerivad replikatsioonikahvli liikumist vasakult ning terD ja terA paremalt. Nende kahe regiooni vahe on 275 kb. ter järjestused seovad valku Tus, mis blokeerib replikatsioonikahvli liikumiseks vajaliku helikaasi töö. Rekombinatsioon dif saitidest Xer rekombinaasi toimel tagab tütarmolekulide teineteisest eraldumise, dekatenatsiooni. Vajalik on ka DNA güraasi ja topoisomeraasi IV juuresolek. Tütarkromosoomide jaotumine Septumi moodustumine raku pooldumiseks toimub siis, kui raku pikkus L (E. coli puhul on see 2,8 µm) on kahekordistunud (2L). Tütarkromosoomid liiguvad teineteisest kaugusele 1L. Üks võimalikke
Seostub ORI-ga järjestusspetsiifiliselt, harutab lahti ORI-kõrval oleva DNA, tekib replikatsiooni mull (silm). - Initsiaator kutsub kohale teisi valke (helikaase), mis seonduvad lahti tehtud DNA piirkonnaga ja replikatsioon läheb edasi. E. colil ORI – 3x13 piirkond + 4x9 piirkond. Dna – seondub 9-aluspaarilistele kordustele, tekib DNA topoloogia muutus, see kutsub esile ahelate lahtikeerdumise, et ahel ei katkeks, vajab topoisomeraasi. DnaB – DNA helikaas DnaC – DNA helikaasi laadija – suurendavad replikatsioonisilma Pärast DnaB ja DnaC seostumist toimub praimeri süntees. seejärel seostub DNA polümeraas III holoensüüm, sünteesitakse veidi DNA-d, lisandub libisev klamber DNA polümeraas III-le → täielik replikatsiooni mehhanism nüüd koos. See replikatsioon vajab ühe valgu sünteesi. DnaA lagundatakse. Järgmise replikatsiooni ringiks on vaja sünteesida uus DnaA valk (initsiaator), sest see on ühekordseks kasutamiseks.
1. rakk-rakk interaktsiooni loomine ehk MPF (mating-pair formation) 1. sugupiili teke, mis on vajalik rakk-rakk kontakti tekkeks ning DNA ülekandumiseks (tüüp 4 sekretsioonisüsteem ehk T4SS type 4 secretion system) 2. DNA ülekanne 1. relaksosoomi teke, üheahelalise katke tekkimine oriT järjestuses ning DNA lahti-hargnemine. Selleks on vajalik konjugatsioonispetsiifilise endonukleaasi ehk relaksaasi ekspressioon (mõnel juhul eraldi helikaasi ekspressioon) 2. DNA-relaksosoomi kompleksi esitlemine T4SS-le T4CP (T4SS coupling proteiin) abil ning DNA ülekandmine retsipienti 3. DNA süntees ja ligeerimine 4. (piili lagunemine ning) rakkude lahknemine. 100 Rakk-rakk kontakti loomiseks peab doonorrakk olema võimeline moodustama sugupiili. Sugupiilid võivad morfoloogiliselt erineda, neid on jagatud